The aim of this study was to obtain a better under-standing of the palynomorph flora, the age and thedepositional environment of the Neill Klinter Group inJameson Land, East Greenland (Fig. 1). Although thesedimentology, fossil faunas, ichnofaunas, lithostrati-graphy and sequence stratigraphy of this successionhave been studied in detail (Rosenkrantz 1934; Sykes1974; Dam 1990a, b, 1991; Dam & Surlyk 1995, 1998),
there are few published papers on the palynology of theNeill Klinter Group. The group has recently been dividedinto four formations and nine members (Fig. 2) and adetailed sequence stratigraphic correlation betweenEast Greenland and Norway has been established (Dam& Surlyk 1995, 1998). Lithostratigraphic units of thegroup that are precisely dated by macrofossils are re-stricted to the Rævekløft Formation and the Lepidopteriselv,
Palynostratigraphy and palaeoenvironments of theRævekløft, Gule Horn and Ostreaelv Formations(Lower–Middle Jurassic), Neill Klinter Group, JamesonLand, East Greenland
Eva B. Koppelhus and Gregers Dam
The Neill Klinter Group of Jameson Land, East Greenland contains rich and diverse palynomorphassemblages. Spores, pollen and freshwater algae dominate most of the samples, but dinofla-gellate cysts and acritarchs also form important components. The ages suggested by the paly-nomorphs from the Rævekløft, Gule Horn and Ostreaelv Formations span the period from theEarly Pliensbachian to the early Aalenian. The number of palynomorphs identified totals 136,including 83 miospore and 53 microplankton species; they are grouped into seven palynologi-cal assemblage zones.
In general, there is good agreement between the palynological and sedimentological data, andthe palynological data has refined the understanding of the depositional palaeoenvironments ofthe Neill Klinter Group. In some cases, the boundaries of the palynological assemblage zonesare congruent with major sequence stratigraphic surfaces and the palynological data thus sup-port the sequence stratigraphic interpretation. In other cases, however, regional correlation indi-cates that the zone boundaries cross important sequence stratigraphic surfaces, such as sequenceboundaries; such behaviour is thought to reflect the facies-dependent nature of certain of thepalynological assemblage zones. The pattern of palynological events in East Greenland has alsobeen recognised on the mid-Norwegian shelf.
Keywords: East Greenland, Jameson Land Basin, Lower–Middle Jurassic, Early Pliensbachian – early Aalenian,
Nathorst Fjeld and Skævdal Members of the OstreaelvFormation. The Neill Klinter Group was sampled inten-sively for palynological analysis at a number of differentlocalities in Jameson Land (Fig. 1). This paper onlyincludes data from the lowermost three formations of theNeill Klinter Group (Rævekløft, Gule Horn and Ostrea-elv Formations). The uppermost formation, the SortehatFormation, is treated in an accompanying paper (Koppel-hus & Hansen 2003, this volume). Seven palynomorphassemblage zones have been established from the mostcomplete section, at Albuen (Figs 3, 4). Data from otherlocalities have been correlated with this section.
Geological settingThe Upper Palaeozoic – Mesozoic Jameson Land Basinis located in the present-day land areas of JamesonLand and Scoresby Land, at the southern end of the EastGreenland rift system (Fig. 1; Surlyk 1978). This systemis part of a larger rift complex separating Greenland fromNorway before the opening of the North Atlantic Ocean(Ziegler 1988). The Jameson Land Basin is bounded tothe east and west by major N–S-trending faults, and tothe north by a NW–SE cross-fault in Kong Oscar Fjord(Surlyk 1977a, 1978, 1990a). The southern boundary is
25 km
Illoqqortoormiut
24°W
72°N
71°N
N
Neill Klinter GroupStudied localities
Normal fault
?
?
Scoresby Sund
?
JamesonLand
ScoresbyLand
KongOscar Fjord
Liaselv
Rhætelv
Lepidopteriselv
Ranunkeldal
EnhjørningenDal
Horsedal
MoskusoksekløftAstartekløft
GoniomyakløftAlbuen
Rævekløft
Harris Fjeld
Nathorst Fjeld
ElisBjerg
DusénBjerg
Sortehat
Primulaelv
QupaulakajikSkævdal
Vardekløft
Tancrediakløft
?
22°W
LiverpoolLand
Greenland
Fig. 1. Map of the Jameson Landregion showing the outcrop of theNeill Klinter Group, the location ofsections forming the basis of thisstudy and additional localitiesmentioned in the text.
unknown, but the basin probably extended south ofScoresby Sund, an area that is now covered byPalaeogene plateau basalts. The basin was initiated inthe Devonian due to extensional collapse of theover-thickened crust of the Caledonian mountain belt.The Devonian phase was probably associated withstrike-slip or oblique-slip deformation resulting in thedevelopment of NW–SE-trending transverse faults inthe north-eastern part of the basin. During LateCarboniferous – Early Permian times, the oblique-slipregime changed to a more orthogonal extensionalregime, resulting in the development of basin marginhalf-grabens (Surlyk et al. 1984, 1986; Surlyk 1990a;Larsen & Marcussen 1992). The period of extensionaltectonics was followed by a long period of subsidencelasting from the Late Permian to the Cretaceous, inter-rupted by minor episodes of rifting and faulting (Surlyk1977a, b, 1990a; Clemmensen 1980a; Surlyk et al. 1981,1986; Larsen & Marcussen 1992).
Triassic – Early Jurassic sedimentation in the JamesonLand Basin was, in addition to tectonic subsidence, also
influenced by climate, drainage patterns and eustasy.During the Triassic – earliest Jurassic, a major lacustrinecomplex was situated in the Jameson Land Basin. Thelacustrine deposits record a long-term change from awarm arid to a more temperate humid climate (Clem-mensen 1978a, b, 1979, 1980a, b; Bromley & Asgaard1979; Dam & Surlyk 1992, 1993). This long-term climaticchange was mainly governed by a gradual northwardsdrift of the Laurasian continent and was accompaniedby a long-term eustatic sea-level rise during the EarlyJurassic. In the Pliensbachian, the lacustrine complexwas transformed into a shallow marine embaymentmarking the first fully-marine inundation of the basinsince Late Permian – Early Triassic times (Surlyk 1990b).
The sandstones and mudstones of the Neill KlinterGroup were deposited in a wide, shallow tide- andstorm-influenced marine embayment, during a periodof relative tectonic quiescence. The facies pattern wascontrolled mainly by relative sea-level fluctuations, sed-iment influx and basinal currents (Sykes 1974; Dam &Surlyk 1995, 1998).
Sortehat
Ostreaelv
Gule Horn
Elis Bjerg
Skævdal
HarrisFjeld
NathorstFjeld
Lepidopteriselv
Trefjord Bjerg
MemberFormation
Albuen
Astartekløft Horsedal
Rævekløft
Rhaetian
Hettangian
Sinemurian
Pliensbachian
Toarcian
Low
erM
iddl
e
Jura
ssic
Tria
s.
Upp
er
Aalenian
Bajocian
Bathonian
Callovian
Oxfordian
Kimmeridgian
Volgian
GroupChronostratigraphy Formation
Sortehat
Ostreaelv
Gule Horn
Rævekløft
Kap Stewart
Jam
eson
Lan
d
Super-group
Scoresby Sund
HallBredning
Vardekløft
Neill Klinter
Raukelv
Hareelv
Olympen
Fossil-bjerget
Pelion
Rhæ
telvPrimulaelv
Innakajik
Fig. 2. Jurassic lithostratigraphy of Jameson Landshowing the detailed subdivision of the Neill KlinterGroup formalised by Dam & Surlyk (1998). Modifiedfrom Surlyk (2003, this volume, fig. 5).
Stratigraphy The Pliensbachian – lower Aalenian succession nowreferred to the Neill Klinter Group was initially describedby Rosenkrantz (1929), but was first formally estab-lished as a formation by Surlyk et al. (1973). The NeillKlinter Formation (sensu Surlyk et al. 1973) has subse-quently been raised to group status (Dam & Surlyk1998) and the Rævekløft, Gule Horn and OstreaelvMembers of Surlyk et al. (1973) have been elevated toformation status. Moreover, the former Sortehat Memberof the Vardekløft Formation (Surlyk et al. 1973) hasbeen promoted to formation and transferred to the NeillKlinter Group (Dam & Surlyk 1998). These authorsdivided the Gule Horn Formation into two new mem-bers and the Ostreaelv Formation into seven new mem-bers (Fig. 2).
The Neill Klinter Group is exposed in Jameson Landand Scoresby Land, and in a small fault-bounded out-lier in the southern part of Liverpool Land (Fig. 1); it is300–450 m thick. The boundary between the Kap StewartGroup and the succeeding Neill Klinter Group is an ero-sional unconformity along the south-eastern basin mar-gin, representing a major hiatus corresponding to theSinemurian Stage (Harris 1931; Surlyk 1991; Dam &Surlyk 1995, 1998). The unconformity passes basin-wards into a conformity and the contact between thelacustrine mudstones of the Kap Stewart Group andthe shallow marine sandstones of the Neill Klinter Groupis gradational (Fig. 5; Surlyk 1991; Dam & Surlyk 1995,1998). The upper boundary of the Neill Klinter Groupis placed at a sharp unconformity between the mud-stones of the Sortehat Formation and the sandstones ofthe Vardekløft Group (Surlyk et al. 1973; Surlyk 1990a;Engkilde 1994; Koppelhus & Hansen 2003, this vol-ume). The Neill Klinter Group and most of its con-stituent formations and members show an overall sheetgeometry, although the thicknesses of the units aregreatest in the basin centre and thin towards the mar-gins (Dam & Surlyk 1995, 1998).
A rich marine fauna is present in the lower part ofthe Neill Klinter Group (Rosenkrantz 1934). It occursin the Rævekløft Formation and is restricted to certainlevels separated by largely unfossiliferous intervals.Rosenkrantz (1934) identified a lower division, with adiverse fauna (150 species) dominated by bivalves, gas-tropods, cephalopods, echinoids and crinoids, and anupper division yielding a relatively sparse fauna (c. 20molluscan species). Ammonites of the genus Uptoniaoccur in the lower division, suggesting that these bedsbelong to the Early Pliensbachian Jamesoni Zone
(Rosenkrantz 1934). In the upper division, Rosenkrantz(1934) found two ammonites, Beaniceras sp. andLytoceras fimbriatum. Rosenkrantz (1934) referred thisdivision to the Ibex Zone, although the bed also yieldedan ammonite that appears to be Aegoceras aff. capri-cornus of the maculatum group, indicative of theDavoei Zone (Callomon 1961; Surlyk et al. 1973). Allthe belemnites recovered from the Rævekløft Formationby Rosenkrantz were apparently collected from theJamesoni Zone interval. They indicate that the JamesoniZone as adopted by Rosenkrantz (1934) includes theEarly Pliensbachian Jamesoni Zone to at least the IbexZone and possibly the early Davoie Zone (Doyle 1991).
The marine macrofossils of the Lepidopteriselv,Nathorst Fjeld, Skævdal and Trefjord Bjerg Members ofthe Ostreaelv Formation and the Sortehat Formationare bivalves, brachiopods, crinoids, belemnites, am-monites, and vertebrates (Rosenkrantz 1934). Ammonitescollected on the top of Elis Bjerg from strata belongingto the Lepidopteriselv Member include Dactyliocerassemicelatum (Simpson) sensu Howarth 1992 (probablyincluding D. groenlandicum Rosenkrantz 1934) andHildaites sp. aff. murleyi (Moxon). Dactylioceras semi-celatum belongs to the Early Toarcian TenuicostatumZone, Semicelatum Subzone (J.H. Callomon, personalcommunication 1993). Hildaites sp. is an early form, rem-iniscent of Protogrammoceras. Dactylioceras sp. hasalso been collected at Nathorst Fjeld in the Nathorst FjeldMember and in the lower part of the Skævdal Member,also suggesting an Early Toarcian Tenuicostatum Zoneage (C. Bjerrum and J.H. Callomon, personal commu-nications 1996). Dactylioceras sp. and Hildaites sp.,suggestive of the Early Toarcian, have also been col-lected from the Lepidopteriselv Member in Horsedal,although they were both loose specimens. Phydoleocerassp. has been collected on Nathorst Fjeld in the TrefjordBjerg Member, just beneath the boundary of the SortehatFormation (C. Bjerrum, personal communication 1996).
The Lepidopteriselv Member has been correlated onsequence stratigraphic grounds with the Nathorst FjeldMember (Dam & Surlyk 1995, 1998). On Nathorst Fjeld,Rosenkrantz (1934) collected a specimen of the belem-nite Parapassolotheuthis polita at an altitude of 494 m,and ‘Parabrachybelus’ subaduncatus at 509 m. Thelower level probably belongs to the Nathorst FjeldMember, and the upper level to the overlying SkævdalMember. The two species have restricted ranges andare not known to be widespread in Europe. Para-passolotheuthis polita has only been recorded from theEarly Toarcian latest Falciferum Zone or earliest BifronsZone (Commune Subzone) in Britain, while ‘Para-
brachybelus’ subaduncatus, which so far has only beenrecorded from mainland Europe, has a range probablyrestricted to the latest Toarcian Levesquei Zone (Doyle1991). The ammonite Dactylioceras semicelatum (Simp-son) has been collected at the base of the SkævdalMember at Nathorst Fjeld (C. Bjerrum, personal com-munication 1996) and in the Lepidopteriselv Memberon top of Elis Bjerg indicating an Early ToarcianTenuicostatum Zone, Semicelatum Subzone age (J.H.Callomon, personal communication 1993). Based onthese data, the Nathorst Fjeld and LepidopteriselvMembers and the lower part of the Skævdal Memberinclude strata with an Early Toarcian TenuicostatumZone to latest Falciferum Zone or earliest Bifrons Zoneage. Belemnites suggest that the Skævdal Member mayalso include strata with a latest Toarcian Levesquei Zoneage (Doyle 1991; Dam & Surlyk 1998), suggesting eitherthat the Skævdal Member has a very long age range,that the belemnites cannot be used stratigraphically orthat the D. semicelatum at the base of the SkævdalMember is reworked.
Dam & Surlyk (1995, 1998) interpreted the Neill KlinterGroup within a sequence stratigraphic framework andattempted a sequence stratigraphic correlation with thecoeval Tilje, Ror, Ile and Not Formations on the mid-Norwegian shelf. This comparison demonstrated that theLower Jurassic in both regions consists of six sequencesand it appears feasible to directly correlate systems tractson a scale of a few tens of metres between East Greenlandand the mid-Norwegian shelf (see Fig. 17).
Previous palynological workPrevious reports on the palynology of the Neill KlinterGroup have been published by Lund & Pedersen (1985)and Underhill & Partington (1994). The former authorsstudied the Neill Klinter Group together with the over-lying Vardekløft Group (sensu Surlyk 2003, this vol-ume, fig. 5) and the lower part of the Hareelv Formation.Based on material collected from Vardekløft, in thesouth-eastern part of the basin (Fig. 1), Lund & Pedersen(1985) proposed four assemblage zones for the entiresuccession based on the miospore assemblages. Dino-flagellate cysts were used to improve the age correla-tion of the spore-pollen assemblages. The threelowermost assemblage zones A, B and C of Lund &Pedersen (1985) cover the Rævekløft, Gule Horn andOstreaelv Formations of the Neill Klinter Group.Assemblage Zone A is divided into subassemblages A1and A2; the age of the zone was suggested to be Late
Pliensbachian because of the presence of the sporeKraeuselisporites reissingeri and the dinoflagellate cystNannoceratopsis triceras. Assemblage Zone B is char-acterised by abundant Spheripollenites subgranulatusand Luehndea spinosa, and an Early Toarcian age wasproposed. Assemblage Zone C is characterised by theincoming of the pollen Callialasporites dampieri, thespores Sestrosporites pseudoalveolatus and Staplini-sporites caminus and the dinoflagellate cyst Parvocystacontracta (now Susadinium scrofoides); these specieswere considered to indicate a Late Toarcian age for thelowermost Assemblage Zone C (subassemblage (C)).
Underhill & Partington (1994) discussed the devel-opment of the Lower Jurassic in East Greenland in con-nection with a sequence stratigraphic study of the NorthSea. They included 3 sections from Jameson Land,Section 1 from Liaselv, Section 2 from Vardekløft andSection 3 from the Harris Fjeld/Primulaelv area (Fig. 1).Sections 1 and 2 cover the uppermost few metres ofthe Ostreaelv Formation and all of the Sortehat Formationand Vardekløft Group. Section 3 covers 170 m of theNeill Klinter Group. Underhill & Partington (1994)analysed 48 samples and recognised 11 events. Theysuggested a Late Pliensbachian – earliest Toarcian agefor the Gule Horn Formation and a Toarcian age forthe Ostreaelv Formation.
Materials and methodsMost of the samples used in this study are from the sec-tion at Albuen, in the fifth ravine north of Skævdal (Figs1, 3). Intervals that proved inaccessible in this ravinewere sampled at Astartekløft (see Figs 1, 9). Sampleswere also obtained from Rævekløft, Tancrediakløft, Qupa-ulakajik, Albuen, Goniomyakløft, Astartekløft, Moskus-oksekløft, Harris Fjeld, Primulaelv, Lepidopteriselv,Liaselv, Horsedal and Ranunkeldal (Fig. 1). A numberof samples collected by Claus Heinberg and Tove Birke-lund in 1974 from Lepidopteriselv were also includedin the study.
The samples were processed for their palynologicalcontent using the techniques adopted at the formerGeological Survey of Greenland, as described byNøhr-Hansen (1993). Over 210 samples were analysedfor their palynological content by means of a trans-mission light microscope. Two hundred specimens werecounted in each sample and all species were registeredin the range chart programme SIS and on the videodatabase at the Geological Survey of Denmark andGreenland, where the slides are stored.
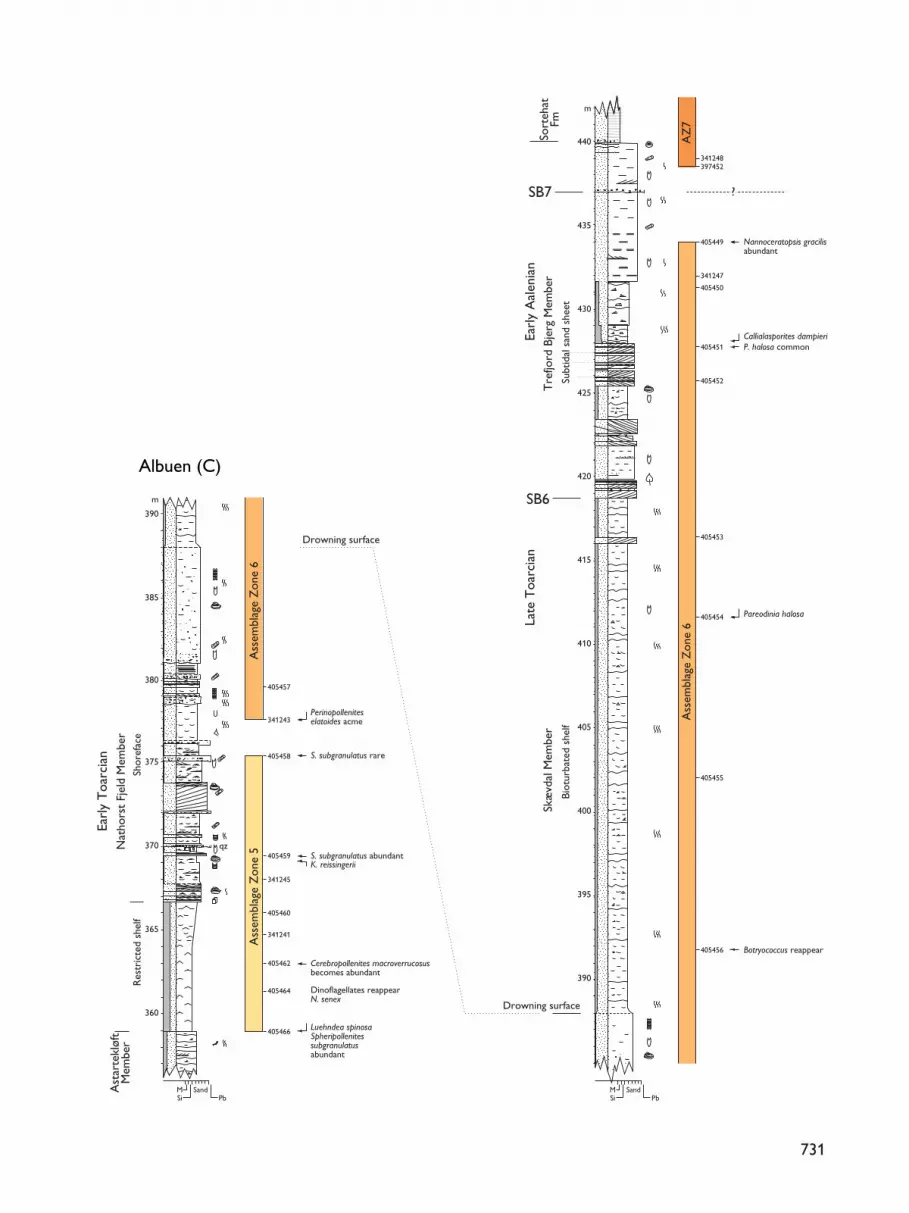
Fig. 3A–C. Sedimentological logs through the Gule Horn (A, B) andOstreaelv (B, C) Formations, Neill Klinter Group, at Albuen (forlocation, see Fig. 1); the Gule Horn Formation comprises the Elis Bjergand Albuen Members, the Ostreaelv Formation is composed of theAstartekløft, Nathorst Fjeld, Skævdal and Trefjord Bjerg Members.Sequence boundaries (SB3–7), palynomorph assemblage zones (AZ)and sample numbers are indicated. The legend accompanying thisfigure is also applicable to Figs 5, 7, 9 and 15.
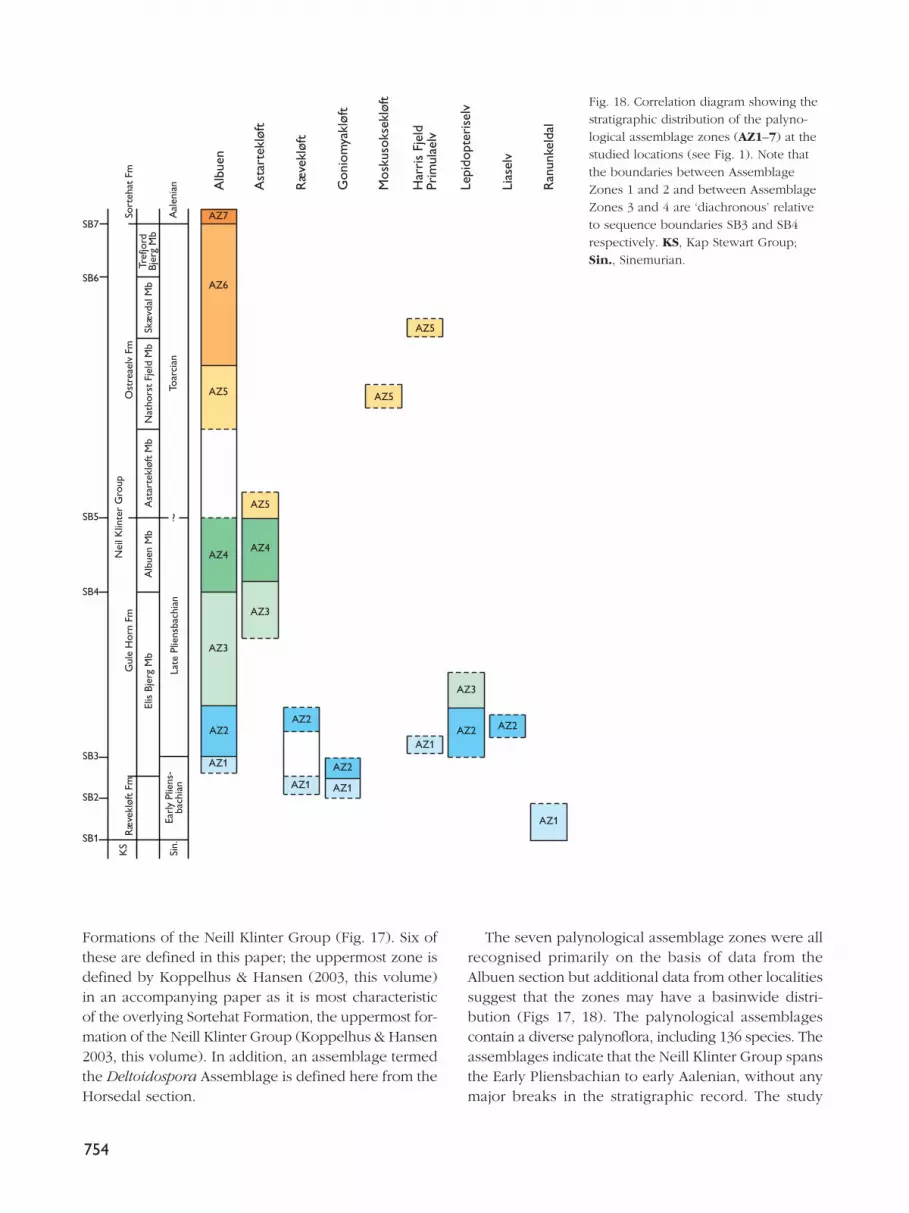
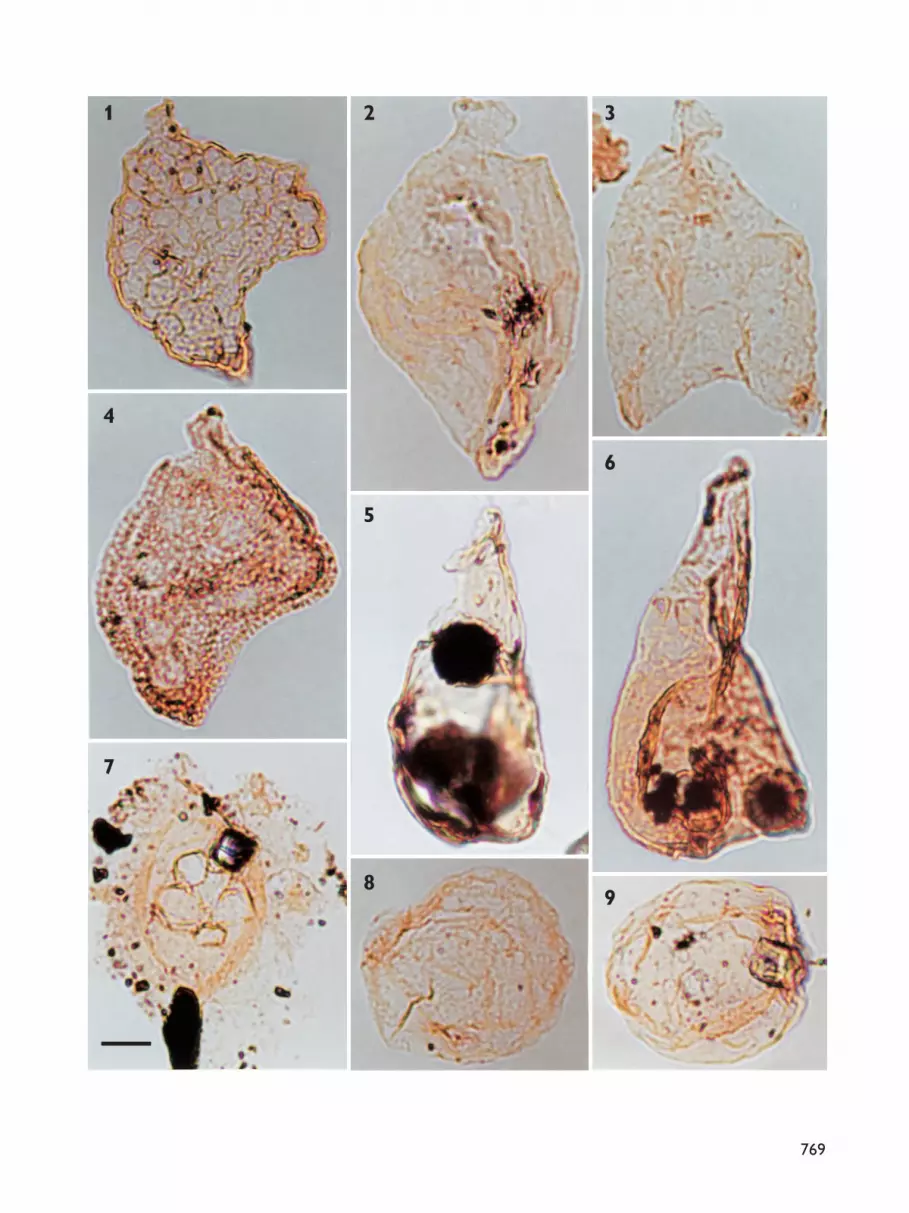
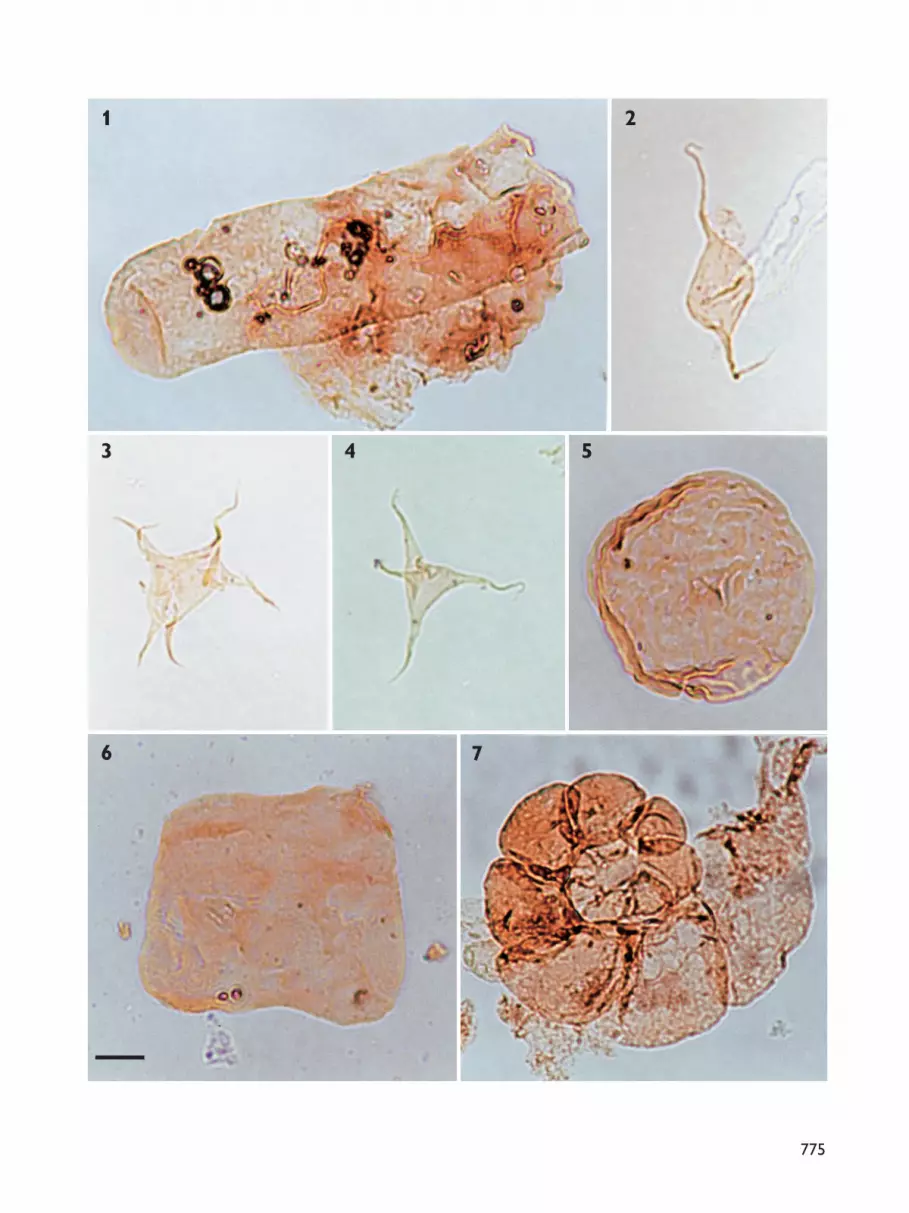
Palynological zones: definitionNine assemblage zones have been recognised in theNeill Klinter Group. The zones are based on the com-position of the entire assemblage of spores, pollen,dinoflagellate cysts, acritarchs and freshwater algae. Theassemblage zones are numbered 1–9 and named afterthe species which dominate the assemblages. AssemblageZones 1–6 are defined and described in detail in thispaper, together with a brief description of AssemblageZone 7. Assemblage Zones 7–9 are formally defined inthe accompanying paper on the Sortehat Formation(Koppelhus & Hansen 2003, this volume). The recordedtaxa are listed in full in Appendix 1 and the importantspecies are illustrated in Plates 1–7.
The most complete section was sampled at Albuen(Figs 1, 3); approximately 70 samples were analysed forpalynomorphs from this section (Fig. 4, facing page744). These samples yielded rich though not very well-preserved palynomorph assemblages (Plates 1–7), withthe exception of samples taken close to the Palaeogenesills and dykes that penetrate the sedimentary succes-sion. These samples were either barren or the paly-nomorphs present were so dark as to be indeterminate.From the productive samples, approximately 150 speciesof spores, pollen, dinoflagellate cysts, acritarchs andfreshwater algae were identified (Appendix 1). Theintensively sampled Albuen section forms the referencesection for Assemblage Zones 1–6 defined in this paper;Assemblage Zones 7–9 are defined from the boreholeat Sortehat (Fig. 1; Koppelhus & Hansen 2003, this vol-ume). In addition to the Albuen reference section,important data for the definition of the assemblagezones were obtained from Ranunkeldal, Goniomyakløftand Astartekløft (Fig. 1).
In addition to the formal assemblage zones definedhere, a distinctive palynological assemblage was iden-tified in samples from the Horsedal Member of theOstreaelv Formation at Horsedal. This assemblage isdefined as the Deltoidospora Assemblage (see belowunder the Horsedal locality).
Assemblage Zone 1:Cerebropollenites thiergartii – Pinuspollenitesminimus – Botryococcus
new assemblage zone
Occurrence. Albuen 211–222.5 mGoniomyakløft 226.5 m (single sample)
Primulaelv 310 m (single sample) Rævekløft 118–122 mRanunkeldal 306–352.5 m
This assemblage zone was recorded from the RævekløftFormation at Rævekløft and Goniomyakløft and fromthe Elis Bjerg Member (Gule Horn Formation) at Albuen,Primulaelv and Ranunkeldal. Relative to the sequencestratigraphic scheme of Dam & Surlyk (1995, 1998), theassemblage occurs within sequences SQ1 and SQ2 (seeFig. 17).
Reference section. Albuen, 211 m (sample 405401) –222.5 m (sample 405404; Figs 1–4).
Additional section. Ranunkeldal, 306 m (sample 341171)– 352.5 m (sample 341173; Figs 5, 6).
Base. The base is not seen in the reference section atAlbuen (Figs 3A, 4). At Ranunkeldal, the base of thezone is placed at sample 341171, immediately abovethe base of the Neill Klinter Group (Figs 5, 6); samplesfrom the underlying Kap Stewart Group are consideredto represent a different assemblage but further work isrequired to precisely define the base of AssemblageZone 1.
Top. The top of the zone is defined by the last sampleshowing this assemblage (sample 405404 at 222.5 m atAlbuen) beneath the first appearance of Nannoceratopsissenex and N. sp. (Figs 3A, 4).
Characteristics. Terrestrial palynomorphs (spores andpollen) dominate together with Botryococcus sp. Themost common spores are Deltoidospora sp. andBaculatisporites sp., and the pollen is dominated byCerebropollenites thiergartii and Pinuspollenites min-imus and other bisaccates. Other characteristic speciesare Cerebropollenites macroverrucosus, Chasmatosporiteshians and C. major. No dinoflagellate cysts have beendetermined with certainty, but a few acritarchs were iden-tified. The freshwater alga Botryococcus sp. occurs inabundance.
Suggested age. An Early Pliensbachian age is proposedbased on the presence of abundant Cerebropollenitesthiergartii.
Palaeoenvironment. The assemblage records a strongterrestrial signal, most components being indicative offreshwater to brackish conditions. Botryococcus, for
732
733
MPbSi
SandM
PbSiSand
285
290
295
341168
341167
341169
300
305
310
315
320
325
m
341171
341170
341172
330
335
340
345
350
355
360
m
341173
Ope
n la
cust
rine
Stor
m-d
omin
ated
offs
hore
tra
nsiti
on
Kap
Ste
war
t G
roup
Wav
e- a
nd s
torm
-dom
inat
ed s
hore
face
Elis
Bje
rg M
embe
r (G
ule
Hor
n Fm
)
Elis
Bje
rg M
embe
r
Ass
embl
age
Zon
e 1
Ass
embl
age
not
defin
ed
Ass
embl
age
Zon
e 1
Tid
al c
hann
elRanunkeldal
SB1
qz
Fig. 5. Sedimentological log through theuppermost part of the Kap StewartGroup and the Elis Bjerg Member (GuleHorn Formation) of the Neill KlinterGroup in Ranunkeldal (for location, seeFig. 1). Sequence boundary (SB1),palynomorph Assemblage Zone 1 andsample numbers are indicated. Forlegend, see Fig. 3; arrows denote grain-size trends.
example, is a planktonic green alga that occurs incolonies. It is known to adapt to different aquatic envi-ronments (fresh to brackish water), has been recordedfrom tropical to subarctic regions and has a stratigraphicrange from the Precambrian to the present day(Guy-Ohlson 1992). When large numbers of Botry-ococcus are recorded, it indicates that the depositionalenvironment was strongly influenced by fresh or brack-ish waters (Guy-Ohlson 1992).
Remarks. Assemblage Zone 1 is equivalent to As-semblage Zone A of Lund & Pedersen (1985), and prob-ably to the lower part of section 3 of Underhill &Partington (1994). The assemblage is similar but notidentical to that described from the ChasmatosporitesZone (miospore) and the Mendicodinium reticulatumZone (dinoflagellate) from the successions of Bornholmand the Øresund area (Koppelhus & Nielsen 1994;Koppelhus & Batten 1996).
This assemblage zone is characteristic of the Elis BjergMember of the Gule Horn Formation where it typicallyoccurs within sequence SQ3 of Dam & Surlyk (1995,1998), for example at Albuen (Figs 3A, 4), Lepidopteris-elv, Liaselv and Rævekløft. At Goniomyakløft, however,Assemblage Zone 2 was identified in the uppermostRævekløft Formation and the lowermost Elis BjergMember (Figs 7, 8), within sequence SQ2 of Dam &Surlyk (1995, 1998).
Reference section. Albuen, 229 m (sample 405405) –241 m (sample 405417; Figs 3A, 4).
Additional section. Goniomyakløft, 229.8 m (sample405469) – 235 m (sample 405471; Figs 7, 8).
Base. The base of the zone is defined by the sampleshowing the first appearance of Nannoceratopsis senexand N. sp.
Top. The top of the zone is defined by the last sampleshowing this palynomorph assemblage, above whichNannoceratopsis disappears together with most otherdinoflagellate cysts.
735
240
245
m
MSi Pb
Sand
225
230
235 405471
405470
405469
405468
405467
Drowning surface
Goniomyakløft
Stor
m-d
omin
ated
sand
y sh
oal
Subt
idal
san
d sh
eet
Res
tric
ted
shel
fU
pper
sho
refa
ce
Ræ
vekl
øft
Form
atio
nEl
is B
jerg
Mem
ber
Ass
embl
age
Zon
e 2
Gul
e H
orn
Form
atio
n
SB3
AZ1
Fig. 7. Sedimentological log through the uppermost part of theRævekløft Formation and the Elis Bjerg Member (Gule HornFormation) at Goniomyakløft (for location, see Fig. 1). Sequenceboundary (SB3), palynomorph assemblage zones (AZ) and sam-ple numbers are indicated. For legend, see Fig. 3. Note that theexact location of sample 405467 is uncertain due to imprecisefield records; although here placed immediately beneath the non-exposed interval (226.5–227.8 m), it is possible that it derives fromimmediately above this interval. For legend, see Fig. 3.
736
Gon
iom
yakl
øft
Lower Pliensbachian
Rævekløft Formation
Lower Jurassic
235.
0023
4.50
232.
40
229.
80
226.
50
4054
7140
5470
4054
69
4054
68
4054
671Kekryphalospora distincta
2Retitriletes semimuris
3Retitriletes austroclavatoides
4Stereisporites stereoides
5Lycopodiacidites rugulatus
6Baculatisporites sp.
7Deltoidospora spp.
8Densoisporites velatus
9Staplinisporites caminus
10Retitriletes clavatoides
11Striatella seebergensis
12Leptolepidites sp.
13Densoisporites scanicus
14Retitriletes sp.
15Foraminisporis jurassicus
16Tigrisporites microrugulatus
17Striatella scanica
18Rogalskaisporites cicatricosus
19Neoraistrickia gristhorpensis
20Megaspore spp.
21Striatella jurassica
22Todisporites major
23Cibotiumsporites jurienensis
24Neoraistrickia sp.
25Corollina torosus
26Quadraeculina anellaeformis
27Chasmatosporites apertus
28Perinopollenites elatoides
29Bisaccate spp.
30Cerebropollenites thiergartii
31Chasmatosporites hians
32Cerebropollenites macroverrucosus
33Chasmatosporites minor
34Pinuspollenites minimus
35Chasmatosporites major
36Corollina sp.
37Callialasporites sp.
38Dapcodinium sp.
39Mancodinium semitabulatum
40Dinocyst spp.
41Nannoceratopsis sp.
42Crassosphaera spp.
43Acritarch spp.
44Tasmanites sp.
45Botryococcus spp.
46Foraminiferal linings
47Foraminifera spp.
?
?
? ?
?
?
??
Alp
habe
tical
spe
cies
list
43A
crita
rch
spp.
6Bacu
latisp
orites
sp.
29Bi
sacc
ate
spp.
45Bot
ryoc
occu
s sp
p.37
Calli
ala
spor
ites
sp.
32Cer
ebro
pol
lenites
macr
over
ruco
sus
30Cer
ebro
pol
lenites
thie
rgart
ii27
Chasm
ato
spor
ites
aper
tus
31Chasm
ato
spor
ites
hia
ns
35Chasm
ato
spor
ites
majo
r33
Chasm
ato
spor
ites
min
or23
Cib
otiu
msp
orites
jurien
ensis
36Cor
ollin
a s
p.25
Cor
ollin
a t
oros
us
42Cra
ssos
phaer
a s
p.38
Dapco
din
ium
sp.
7D
elto
idos
por
a s
pp.
8D
enso
ispor
ites
vel
atu
s13
Den
soispor
ites
sca
nic
us
40D
inoc
yst
spp.
46Fo
ram
inife
ral l
inin
gs47
Fora
min
ifera
spp
.15
Fora
min
ispor
is jura
ssic
us
1Kek
ryphalo
spor
a d
istinct
a12
Lepto
lepid
ites
sp.
5Ly
copod
iaci
dites
rugu
latu
s39
Manco
din
ium
sem
itabula
tum
20M
egas
pore
spp
.41
Nannoc
erato
psis
sp.
19N
eora
istr
ickia
gris t
hor
pen
sis
24N
eora
istr
ickia
sp.
28Pe
rinop
olle
nites
ela
toid
es34
Pinusp
olle
nites
min
imus
26Q
uadra
eculin
a a
nel
laef
orm
is3
Ret
itrile
tes
aust
rocl
ava
toid
es10
Ret
itrile
tes
clava
toid
es2
Ret
itrile
tes
sem
imuris
14Ret
itrile
tes
sp.
18Rog
alskaispor
ites
cic
atr
icos
us
9Sta
plin
ispor
ites
cam
inus
4Ste
reispor
ites
ste
reoi
des
21Str
iate
lla jura
ssic
a17
Str
iate
lla s
canic
a11
Str
iate
lla s
eeber
gensis
44Ta
smanites
sp.
16Tig
risp
orites
mic
roru
gula
tus
22To
dispor
ites
majo
r
System
Stage
Palynological Assembl. Zones
Lithostratigraphy Elis BjergMb
UpperPliensbach.
(m)
Sample height
Sample number
225
235
215
205
12
Fig.
8. Pal
ynom
orp
h d
istrib
utio
n c
har
t fo
r th
e upper
most
par
t of th
e Ræ
vekl
øft F
orm
atio
n a
nd the
Elis
Bje
rg M
ember
(G
ule
Horn
Form
atio
n)
at G
onio
mya
kløft (
for lo
catio
n, se
e Fi
g. 1
).Fo
r le
gend, se
e Fi
g. 4
.
Characteristics. Terrestrial palynomorphs dominate theassemblage together with Botryococcus sp., as inAssemblage Zone 1; the spore Kekryphalospora dis-tincta appears for the first time. The difference betweenthis assemblage and that of Assemblage Zone 1 is theappearance of dinoflagellate cysts, including Manco-dinium semitabulatum, Nannoceratopsis senex, N. gra-cilis, N. plegas, N. triangulata and Parvocysta barbataand the presence of more acritarchs, including Limbicystabjaerkei.
Suggested age. A Late Pliensbachian age is suggestedbased on the first appearance of the spore Kekry-phalospora distincta in sample 405411 (236.50 m) in theAlbuen section (Fig. 3A); this species is known to havea range from Late Pliensbachian to Early Bajocian (Fenton& Riding 1987).
Palaeoenvironment. Although influenced by brackish tomarine waters, there is still a strong terrestrial signal.
Remarks. The spore Kekryphalospora distincta is alsoknown from assemblages of Pliensbachian age fromthe Danish area (Anholt borehole, Øresund borehole15 and the Korsodde section on Bornholm; Seidenkrantzet al. 1993; Koppelhus & Nielsen 1994; Koppelhus &Batten 1996). The appearance of Nannoceratopsis tri-angulata is particularly noteworthy as this species haspreviously only been recorded from NW Germanywhere it occurs in the uppermost Toarcian (Prauss1987). The appearance of Limbicysta bjaerkei andParvocysta barbata is unexpected at this level, as theyare known to have their first appearance in the BifronsZone (late Early Toarcian) in the North Sea and Svalbardand on the mid-Norwegian shelf (Bjærke 1980a; Riding& Thomas 1992; I. Throndsen, personal communication1996). However, these anomalous occurrences werealso recognised at this level in the Neill Klinter Groupby Underhill & Partington (1994); L. bjaerkei wasrecorded in their section 3 between 360 m and 370 m.Two explanations are possible: either P. barbata andL. bjaerkei have a longer range than recently reportedor the sediments are younger than expected.
In Assemblage Zone 2, acritarchs are more commonthan in the underlying zone and in the succeedingzones; this pattern is known from the Lower Jurassicin England and Wales (Wall 1965).
Assemblage Zone 3:Chasmatosporites – Cerebropollenites thiergartii– Botryococcus
new assemblage zone
Occurrence. Albuen 246–284.7 mAstartekløft 266–328 mLepidopteriselv 674–700 m
Assemblage Zone 3 is confined to the uppermost partof the Elis Bjerg Member (Gule Horn Formation) in theAlbuen and Lepidopteriselv sections; relative to thesequence stratigraphic scheme of Dam & Surlyk (1995,1998), the assemblage occurs within the upper levelsof sequence SQ3, below sequence boundary SB4 inthese sections (Fig. 3A). At Astartekløft, however, theassemblage spans the boundary between the Elis BjergMember and the succeeding Albuen Member, thus strad-dling the sequence boundary (SB4) between sequencesSQ3 and SQ4 (Fig. 10). It should be noted, however,that recognition of assemblage Zone 3 is based on onlytwo widely spaced samples at the Astartekløft locality(Fig. 10).
Reference section. Albuen, 246 m (sample 405418) –284.7 m (sample 405427; Figs 3A, 4).
Additional section. Astartekløft, 266 m (sample 346614)– 328 m (sample 346627; Fig. 10)
Base. The base is placed at the first sample in whichdinoflagellate cysts are absent or rare, succeeding sam-ples of Assemblage Zone 2 characterised by a numberof dinoflagellate species.
Top. The top of the assemblage is placed at the last sam-ple showing the assemblage described below; abovethis level, the palynomorph assemblage is dominatedby bisaccate pollen.
Characteristics. This zone is also dominated by terres-trial material. A number of spores have their first appear-ance, such as Striatella jurassica, Kraeuselisporitesreissingeri, Taurocusporites verrucatus and Densoi-sporites velatus. The pollen species and Botryococcussp. are very consistent; only few dinoflagellate cystsand acritarchs were recorded.
Suggested age. A Late Pliensbachian age is proposedbased on the absence of marker species indicative of
a younger age. The miospore assemblage is a contin-uation of Assemblage Zone 2.
Palaeoenvironment. The palynology displays an over-whelmingly terrestrial signal; there is very little evi-dence of marine influence.
Remarks. Cerebropollenites thiergartii is commonthrough much of the zone but becomes rare towardsthe top of the zone and in succeeding zones.
Assemblage Zone 4: Bisaccates
new assemblage zone
Occurrence. Albuen 287–297.8 mAstartekløft 339–340 m
This assemblage zone is restricted to the Albuen Member(Gule Horn Formation) at both Albuen and Astartekløft(Figs 3A, 3B, 4, 9, 10); it thus falls within sequence SQ4of Dam & Surlyk (1995, 1998).
Reference section. Albuen, 287 m (sample 405428) –297.8 m (sample 405434; Figs 3A, 3B, 4).
Additional section. Astartekløft, 339 m (sample 405472)– 340 m (sample 405473; Figs 9, 10).
Base. The base is defined by the first sample domi-nated overwhelmingly by bisaccate pollen, to the exclu-sion of most other palynomorphs.
Top. The top of the zone is defined by the last samplecomposed predominantly of bisaccate pollen; the suc-ceeding sample, defining the base of the overlyingAssemblage Zone 5, is characterised by the first appear-ance of Spheripollenites subgranulatus.
Characteristics. This assemblage zone is characterised byvery poor preservation of the few palynomorphs presentand by the absence of marine palynomorphs. Bisaccatepollen are common in most of the samples. The spores,pollen and Botryococcus sp. that were abundant in theAssemblage Zones 1–3 are absent in this zone.
Suggested age. A Late Pliensbachian age is assigned tothis zone, as for Assemblage Zone 3 (see above).
Palaeoenvironment. Taken at face value, the palyno-morph data suggest that the sediments of the AlbuenMember were deposited in a more distal position rel-ative to the source than that suggested by the previousassemblage zones. Amongst all palynomorphs, bisac-cate pollen are known to be found farthest away fromthe source, because of their ability to be transportedby air. However, bisaccate pollen are also known to havea thick wall and therefore may be preferentially pre-served. Thus, although lack of marine palynomorphscould be interpreted in terms of a non-marine envi-ronment, it could also have resulted from selectivedestruction of the more thin-walled marine paly-nomorphs during intrusion of Palaeogene igneous sillsand dykes in the Albuen Member.
Remarks. This interval was not recognised in previousstudies by Lund & Pedersen (1985) and Underhill &Partington (1994).
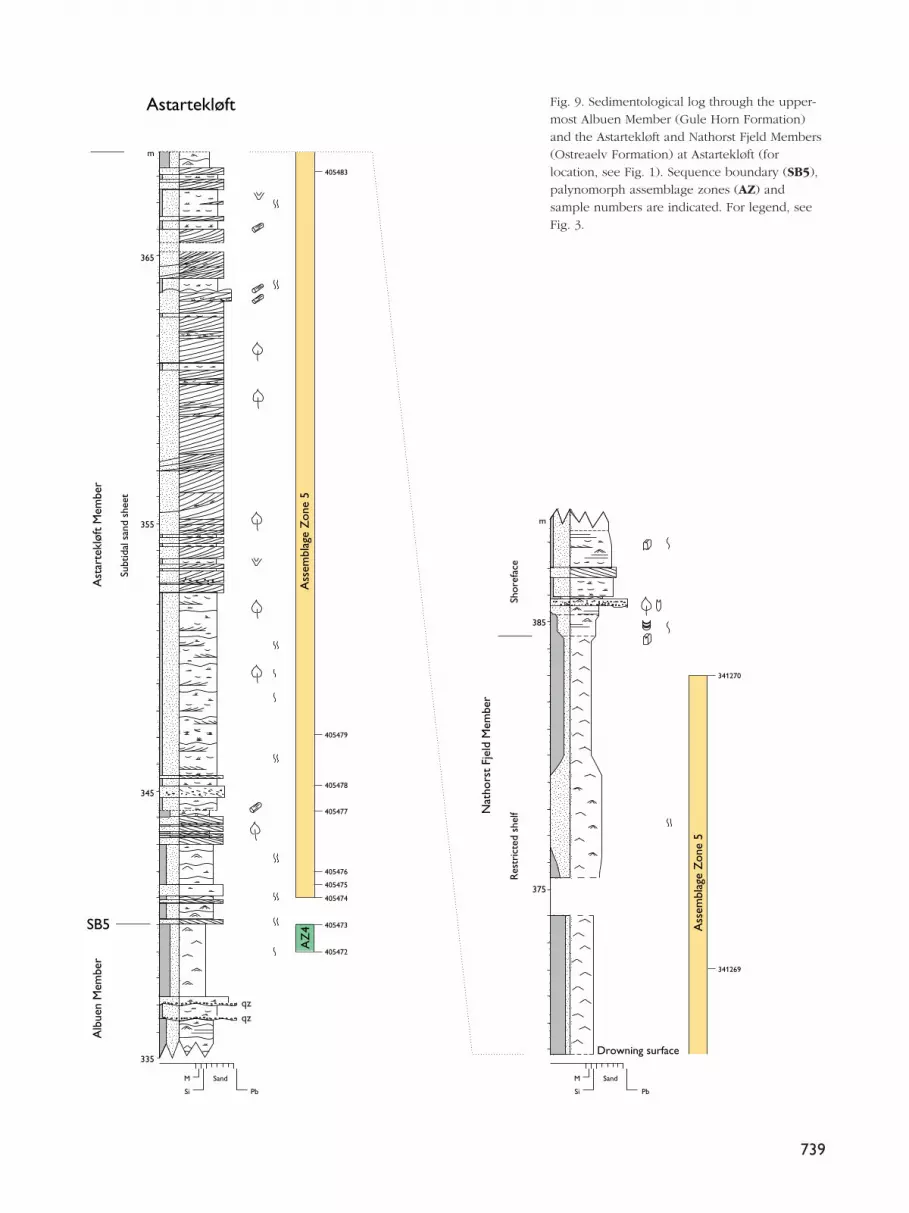
Assemblage Zone 5:Spheripollenites subgranulatus – Cerebropollenitesmacroverrucosus – Luehndea spinosa
new assemblage zone
Occurrence. Albuen 359–375.5 mAstartekløft 341–383 mMoskusoksekløft 359 m (single sample)Primulaelv 451 m (single sample)
In the Albuen reference section, Assemblage Zone 5 wasonly recorded from the lower Nathorst Fjeld Memberof the Ostreaelv Formation; it should be noted that theunderlying Astartekløft Member (also Ostreaelv For-mation) was not sampled in this section so the potentialdownwards range of the assemblage is poorly con-strained (Figs 3B, 3C, 4). At Astartekløft itself, however,Assemblage Zone 5 extends from the lowermostAstartekløft Member up into the Nathorst Fjeld Member(Figs 9, 10). Relative to the sequence stratigraphicscheme of Dam & Surlyk (1995, 1998), AssemblageZone 5 occurs within sequence SQ5, beginning imme-diately above the sequence boundary (SB5) in theAstartekløft section (Fig. 9) and extending up to some10 m beneath the flooding surface at Albuen (Fig. 3C).
Reference section. Albuen, 359 m (sample 405466) –375.5 m (sample 405458; Figs 3B, 3C, 4).
738
739
M
Si Pb
Sand
335
345
355
365
m
375
Drowning surface
SB5
M
Si Pb
Sand
385
m
qzqz
Res
tric
ted
shel
fSh
oref
ace
AstartekløftSu
btid
al s
and
shee
t
Nat
hors
t Fj
eld
Mem
ber
Ast
arte
kløf
t M
embe
r
Ass
embl
age
Zon
e 5
AZ
4 Ass
embl
age
Zon
e 5
Alb
uen
Mem
ber
341270
341269
405472
405473
405474
405475
405476
405477
405478
405483
405479
Fig. 9. Sedimentological log through the upper-most Albuen Member (Gule Horn Formation)and the Astartekløft and Nathorst Fjeld Members(Ostreaelv Formation) at Astartekløft (forlocation, see Fig. 1). Sequence boundary (SB5),palynomorph assemblage zones (AZ) andsample numbers are indicated. For legend, seeFig. 3.
740
Ast
arte
kløft
Lower Jurassic
Toarcian
Ostreaelv FormationAstartekløft MemberNathorst Fjeld Member
Additional section. Astartekløft, 341 m (sample 405474)– 383 m (sample 341270; Figs 9, 10).
Base. The base of the assemblage is defined by the firstappearance, in abundance, of Spheripollenites sub-granulatus; at Albuen, this coincides with the firstappearance of Luehndea spinosa, although this specieswas not recorded in the Astartekløft section.
Top. This is defined by the uppermost sample showingthe palynomorph assemblage characteristic of the zone(i.e. sample 405458 at Albuen). The succeeding sampleshows an acme of Perinopollenites elatoides, defining thebase of Assemblage Zone 6. The top of the assemblageis not seen in the Astartekløft section (Fig. 9).
Characteristics. In Assemblage Zone 5, the pollenSpheripollenites subgranulatus appears in abundancefor the first time; this species was only recorded in onesample below this level, in sample 346627 at 328 m inthe Astartekløft section (Figs 9, 10). The assemblage isalso marked by the reappearance of abundant Nanno-ceratopsis senex and N. gracilis. The pollen Cerebro-pollenites macroverrucosus and to a lesser degreeCorollina torosus are more common than in the under-lying assemblage. Spherical bodies of uncertain affin-ity are also abundant (on the distribution chart they areregistered under micromiscellanea).
Suggested age. An Early–Late Toarcian age is proposedbased on the presence and acme of Spheripollenitessubgranulatus which is known to be abundant inToarcian strata in the Danish Basin and the North Sea(Dybkjær 1991; Batten et al. 1994; Koppelhus & Nielsen1994; Koppelhus & Batten 1996). Palaeoenvironment. This assemblage contains elementsindicative of both brackish and marine conditions.
Remarks. In Germany, the North Sea area and the DanishBasin, the dinoflagellate cyst Luehndea spinosa is knownto appear within the Margaritatus, Spinatus andTenuicostatum Zones which span the Late Pliensbachianto Early Toarcian (Morgenroth 1970; Riding & Thomas1992; Poulsen 1996). The abundant spherical bodies ofunknown affinity are also known from Svalbard, theNorth Sea and the Baltic Sea in upper Pliensbachianand Toarcian strata (Bjærke 1980b; Dybkjær 1991;Koppelhus & Nielsen 1994); on the mid-Norway shelf,these forms are known to occur in Lower Toarcian strata(I. Throndsen, personal communication 1996).
This zone correlates with Assemblage Zone B ofLund & Pedersen (1985). It has not been possible torecognise this assemblage in the data presented byUnderhill & Partington (1994). Comparison with theMicrocysta erugata taxa range-zone of Smelror & Below(1992) has been attempted, but the two zones havevery few species in common.
Assemblage Zone 6: Perinopollenites elatoides
new assemblage zone
Occurrence. Albuen 377.5–434 mEnhjørningen Dal 414–424.67 mSortehat (core) 12.65–26.28 m
In the Albuen section, Assemblage Zone 6 extends fromthe upper Nathorst Fjeld Member through the SkævdalMember and much of the Trefjord Bjerg Member (allOstreaelv Formation; Figs 3C, 4). Relative to the sequencestratigraphic scheme of Dam & Surlyk (1995, 1998), theassemblage occurs within sequences SQ5 and SQ6,spanning the sequence boundary SB6 (Figs 3C, 4). Datafor the Enhjørningen Dal and Sortehat sections is pre-sented in the companion paper by Koppelhus & Hansen(2003, this volume). In the Sortehat core, AssemblageZone 6 is confined to the upper Ostreaelv Formation(Trefjord Bjerg Member); the top of the assemblage liesimmediately beneath the boundary between theOstreaelv and Sortehat Formations. At EnhjørningenDal, however, Assemblage Zone 6 spans the boundarybetween these two formations, extending some 6 m upinto the Sortehat Formation (Koppelhus & Hansen 2003,this volume).
Reference section. Albuen, 377.5 m (sample 341243) –434 m (sample 405449; Figs 3C, 4).
Base. The base of the zone is placed at the sample inwhich Perinopollenites elatoides reaches its acme; it isaccompanied by abundant Chasmatosporites hians andC. major.
Top. The upper boundary of Assemblage Zone 6 isplaced at the last sample showing the palynomorphassemblage described here; above this level, Botry-ococcus sp. becomes very dominant, defining the baseof Assemblage Zone 7.
741
Characteristics. The zone is characterised particularlyby the acme of Perinopollenites elatoides. The sporesStaplinisporites caminus and Sestrosporites pseudoalveo-latus have their first appearance within the zone togetherwith the dinoflagellate cysts Phallocysta eumekes,Wallodinium laganum, Scriniocassis sp. and Dissilio-dinium sp. The dinoflagellate cysts Nannoceratopsisgracilis and N. senex continue to be common in mostof the samples. Botryococcus sp. re-appears at 392 m(sample 405456; Fig. 3C) and continues to be commonto the top of this zone.
Suggested age. A Late Toarcian – early Aalenian age issuggested based on the first appearances of the sporesStaplinisporites caminus and Sestrosporites pseudoalveo-latus together with the dinoflagellate cysts Phallocystaeumekes, Wallodinium laganum, Scriniocassis sp. andDissiliodinium sp. The occurrence of abundant Perino-pollenites elatoides is a feature of Aalenian sedimentsin the Danish Basin and on Bornholm (Dybkjær 1991;Koppelhus & Nielsen 1994).
Palaeoenvironment. The palynomorphs indicate thatthe palaeoenvironment was influenced by both fresh,brackish and marine waters, suggesting a nearshoreenvironment.
Remarks. The two species Chasmatosporites hians andC. major are very abundant in the lowermost samplein this interval, higher up they become rare. In offshoremid-Norway, C. hians has a maximum appearance afterthe Spheripollenites acme (I. Throndsen, personal com-munication 1996). The pollen Callialasporites dampieriis known to appear in the uppermost Toarcian andlowermost Aalenian in north-west Scotland (Riding etal. 1991). This zone correlates with Assemblage ZoneC in Lund & Pedersen (1985) and the assemblage foundin sample B2/57 at approximately 460 m in Primulaelvby Underhill & Partington (1994). The dinoflagellatecyst Wallodinium laganum appears for the first timein this zone and is only known from the Late ToarcianLevesquei Zone in northern Germany and England(Feist-Burkhardt & Monteil 1994). The ammoniteDactylioceras sp. occurs at the base of the SkævdalMember at Nathorst Fjeld, suggesting an Early ToarcianTenuicostatum Zone age for the lower part of the assem-blage zone.
Assemblage Zone 7: Botryococcus
This assemblage is formally defined in the companionpaper by Koppelhus & Hansen (2003, this volume); asummary is given here.
Occurrence. Albuen 438.5–443.5 mEnhjørningen Dal 424.86–445 mPelion 550–567 mSortehat (core) 27.82–36.36 m
At Albuen, this assemblage is represented in the upper-most few metres of the Trefjord Member (OstreaelvFormation) and extends up into the Sortehat Formation(Figs 3C, 4); additional data for the uppermost TrefjordMember and the overlying Sortehat Formation in theAlbuen section are given in Koppelhus & Hansen (2003,this volume). In the cored section from Sortehat, theassemblage occurs in the lower levels of the SortehatFormation, the base of the zone being immediatelyabove the base of the formation (Koppelhus & Hansen2003, this volume). At Enhjørningen Dal, AssemblageZone 7 is also restricted to the lower Sortehat Formationalthough here the base is some 6 m above the lowerboundary of the Sortehat Formation. Detailed discus-sion and the full dataset are given in Koppelhus &Hansen (2003, this volume). Assemblage Zone 7 occurswithin the lower levels of sequence SQ7 of Dam &Surlyk (1995, 1998).
Reference section. Sortehat (core), 27.82 m (sample303143-73) – 36.36 m (sample 303143-62).
Additional sections. See Koppelhus & Hansen (2003, thisvolume).
Base. The base of the assemblage is placed at the firstsample in which Botryococcus sp. overwhelmingly dom-inates the assemblage. In the Albuen section (Figs 3C,4), this event coincides with the first co-occurrence ofCallialasporites dampieri (pollen) and Mendicodiniumgroenlandicum (dinoflagellate cyst) although in othersections (e.g. Enhjørningen Dal, Sortehat; Koppelhus &Hansen 2003, this volume) these species first occurtogether some metres below the Botryococcus sp. influx.
Top. The upper boundary is defined by the uppermostsample showing the Botryococcus-dominated assem-blage. Above this level, Botryococcus sp. disappear andNannoceratopsis gracilis and N. senex become abundantonce more.
742
Characteristics. The assemblage is characterised by theoverwhelming dominance of Botryococcus sp. and thescarcity of dinoflagellates.
Suggested age. An Aalenian age is proposed based onthe abundance of Callialasporites dampieri. Pollen fromthe genus Callialasporites are known to appear first insediments of Late Toarcian and Aalenian age in Swedenand the Danish area (Guy-Ohlson 1988; Koppelhus &Nielsen 1994).
Palaeoenvironment. The fresh and brackish water algaBotryococcus is known from recent environments toproduce blooms at certain times of the year. The coloniesfloat at the water surface under calm conditions and sub-sequently sink when the water is disturbed. When theydie, they float within the surface waters and can betransported by wind far from the area where they wereproduced. Palaeoenvironmental interpretation basedsolely on the presence of Botryococcus is therefore dan-gerous; the degree to which the Botryococcus in thissuccession is allochthonous is unknown. Further dis-cussion of the environmental implications of this assem-blage is given by Koppelhus & Hansen (2003, thisvolume).
Additional palynological resultsAs noted earlier, definition of the six assemblage zonesdescribed here is based primarily on the section atAlbuen, the palynostratigraphy of which is thus pre-sented in detail above. In addition to this reference sec-tion, however, a series of other sections were includedin the study, some of which yield important supple-mentary data for the definition of the assemblage zones(see above). The palynostratigraphic results from theseadditional localities, spread widely in the Jameson LandBasin (Fig. 1), are described below, broadly from southto north.
Rævekløft At Rævekløft, nine samples were collected, six fromthe Rævekløft Formation (405435–405440) and threefrom the Elis Bjerg Member (Gule Horn Formation,405441–405443; Fig. 11). The samples collected fromthe Rævekløft Formation are separated by a gap ofc. 100 m from those collected in the Elis Bjerg Memberand the boundary between the two units was not
exposed. All the samples yielded abundant poorly pre-served palynomorphs. Terrestrial material dominatestogether with the freshwater alga Botryococcus sp.Bisaccate pollen are the most abundant palynomorphsin all the samples. A few acritarchs and questionabledinoflagellate cysts were found. Based on the presenceof the spores Deltoidospora and Baculatisporites, thepollen Cerebropollenites thiergartii and Pinuspollenitesminimus and the lack of dinoflagellate cysts, the sam-ples 405435–405440 (?upper Rævekløft Formation) areassigned to Assemblage Zone 1. In the samples405441–405443 (Elis Bjerg Member), the dinoflagellatecysts Nannoceratopsis senex and Mancodinium semi-tabulatum appear for the first time, indicating that theassemblage belongs to Assemblage Zone 2.
Suggested age. An Early Pliensbachian age is suggestedfor Assemblage Zone 1, based on the presence of Cere-bropollenites thiergartii together with Pinuspollenitesminimus; a Late Pliensbachian age is proposed forAssemblage Zone 2 based on the occurrences of Nanno-ceratopsis senex and Mancodinium semitabulatum.
Tancrediakløft Only one sample (341229; Rævekløft Formation) wasstudied from this locality. The sample contained onlyblack material which was not identifiable.
Qupaulakajik Only one sample (341254; Albuen Member, Gule HornFormation) was studied from this locality. The paly-nomorphs were black and indeterminate.
GoniomyakløftFive samples were studied (Figs 7, 8). The sample 405467,from the uppermost part of Rævekløft Formation, yieldedan assemblage rich in poorly preserved palynomorphs,dominated by terrestrial material. However, several spec-imens of the dinoflagellate cyst genus Dapcodiniumwere found together with a single specimen of Tas-manites. The former are similar to Dapcodinium priscum,but not identical to specimens of this species describedfrom Northwest Europe. The assemblage is suggestedto belong to Assemblage Zone 1.
743
744
Lower Jurassic
Upper Pliensbachian
Gule Horn Formation
12
Elis Bjerg Member
Lower Pliensbachian
Rævekløft Fm
210.
0020
9.00
208.
00
122.
0012
1.00
120.
0011
9.00
118.
5011
8.00
4054
4340
5442
4054
41
4054
4040
5439
4054
3840
5437
4054
3640
5435
1Rogalskaisporites cicatricosus
2Conbaculatisporites mesozoicus
3Lycopodiacidites rugulatus
4Deltoidospora spp.
5Retitriletes clavatoides
6Retitriletes semimuris
7Retitriletes sp.
8Baculatisporites sp.
9Tigrisporites microrugulatus
10Foraminisporis jurassicus
11Kraeuselisporites reissingeri
12Tripartina variabilis
13Stereisporites stereoides
14Densoisporites scanicus
15Retitriletes austroclavatoides
16Todisporites minor
17Kekryphalospora distincta
18Cibotiumsporites jurienensis
19Striatella parva
20Todisporites major
21Striatella jurassica
22Chasmatosporites hians
23Cerebropollenites thiergartii
24Vesicaspora fuscus
25Quadraeculina anellaeformis
26Perinopollenites elatoides
27Pinuspollenites minimus
28Bisaccate spp.
29Cerebropollenites macroverrucosus
30Chasmatosporites major
31Chasmatosporites apertus
32Corollina torosus
33Monosulcites punctatus
34Taeniasporites rhaeticus
35Nannoceratopsis senex
36Acritarch spp.
37Veryhachium sp.
38Leiofusa jurassica
39Leiosphaeridia spp.
40Botryococcus spp.
41Tasmanites sp.
R
Alp
habe
tical
spe
cies
list
36A
crita
rch
spp.
8Bacu
latisp
orites
sp.
28Bi
sacc
ate
spp.
40Bot
ryoc
occu
s sp
p.29
Cer
ebro
pol
lenites
macr
over
ruco
sus
23Cer
ebro
pol
lenites
thie
rgart
ii31
Chasm
ato
spor
ites
aper
tus
22Chasm
ato
spor
ites
hia
ns
30Chasm
ato
spor
ites
majo
r18
Cib
otiu
msp
orites
jurien
ensis
2Con
bacu
latisp
orites
mes
ozoi
cus
32Cor
ollin
a t
oros
us
4D
elto
idos
por
a s
pp.
14D
enso
ispor
ites
sca
nic
us
10Fo
ram
inispor
is jura
ssic
us
17Kek
ryphalo
spor
a d
istinct
a11
Kra
euse
lispor
ites
rei
ssin
geri
38Le
iofu
sa jura
ssic
a39
Leio
sphaer
idia
spp
.3
Lyco
pod
iaci
dites
rugu
latu
s33
Mon
osulc
ites
punct
atu
s35
Nannoc
erato
psis
senex
26Pe
rinop
olle
nites
ela
toid
es27
Pinusp
olle
nites
min
imus
25Q
uadra
eculin
a a
nel
laef
orm
is15
Ret
itrile
tes
aust
rocl
ava
toid
es5
Ret
itrile
tes
clava
toid
es6
Ret
itrile
tes
sem
imuris
7Ret
itrile
tes
sp.
1Rog
alskaispor
ites
cic
atr
icos
us
13Ste
reispor
ites
ste
reoi
des
21Str
iate
lla jura
ssic
a19
Str
iate
lla p
arv
a34
Taen
iasp
orites
rhaet
icus
41Ta
smanites
sp.
9Tig
risp
orites
mic
roru
gula
tus
20To
dispor
ites
majo
r16
Todispor
ites
min
or12
Trip
art
ina v
ariabili
s37
Very
hach
ium
sp.
24Ve
sica
spor
a fusc
us
Ræ
vekl
øft
200
205
210
125
120
115
System
Stage
Palynological Assembl. Zones
Lithostratigraphy
(m)
Sample height
Sample number
Inte
rval
not
sam
pled
Fig.
11.
Pal
ynom
orp
h d
istrib
utio
n c
har
t fo
r th
e Ræ
vekl
øft F
orm
atio
n a
nd G
ule
Horn
Form
atio
n (
Elis
Bje
rg M
ember
) at
Ræ
vekl
øft (
for
loca
tion, se
e Fi
g. 1
). F
or
lege
nd, se
e Fi
g. 4
.
Sample 405468 was collected just beneath the bound-ary between the Rævekløft Formation and the suc-ceeding Elis Bjerg Member (Gule Horn Formation). Thissample yielded a similar palynomorph assemblage tothat described above but included the first appearanceof the dinoflagellate cyst Mancodinium semitabula-tum. It is suggestive of the somewhat younger As-semblage Zone 2. The samples 405469–405471 are fromthe Elis Bjerg Member. They lack recognisable dinofla-gellate cysts although acritarchs and some question-able dinoflagellate cysts were found together withCrassosphaera sp., foraminiferal inner linings and abun-dant Botryococcus sp. This assemblage is also suggestedto belong to Assemblage Zone 2.
Suggested age. An ?Early–Late Pliensbachian age is pro-posed, based on the appearance of Mancodinium semi-tabulatum.
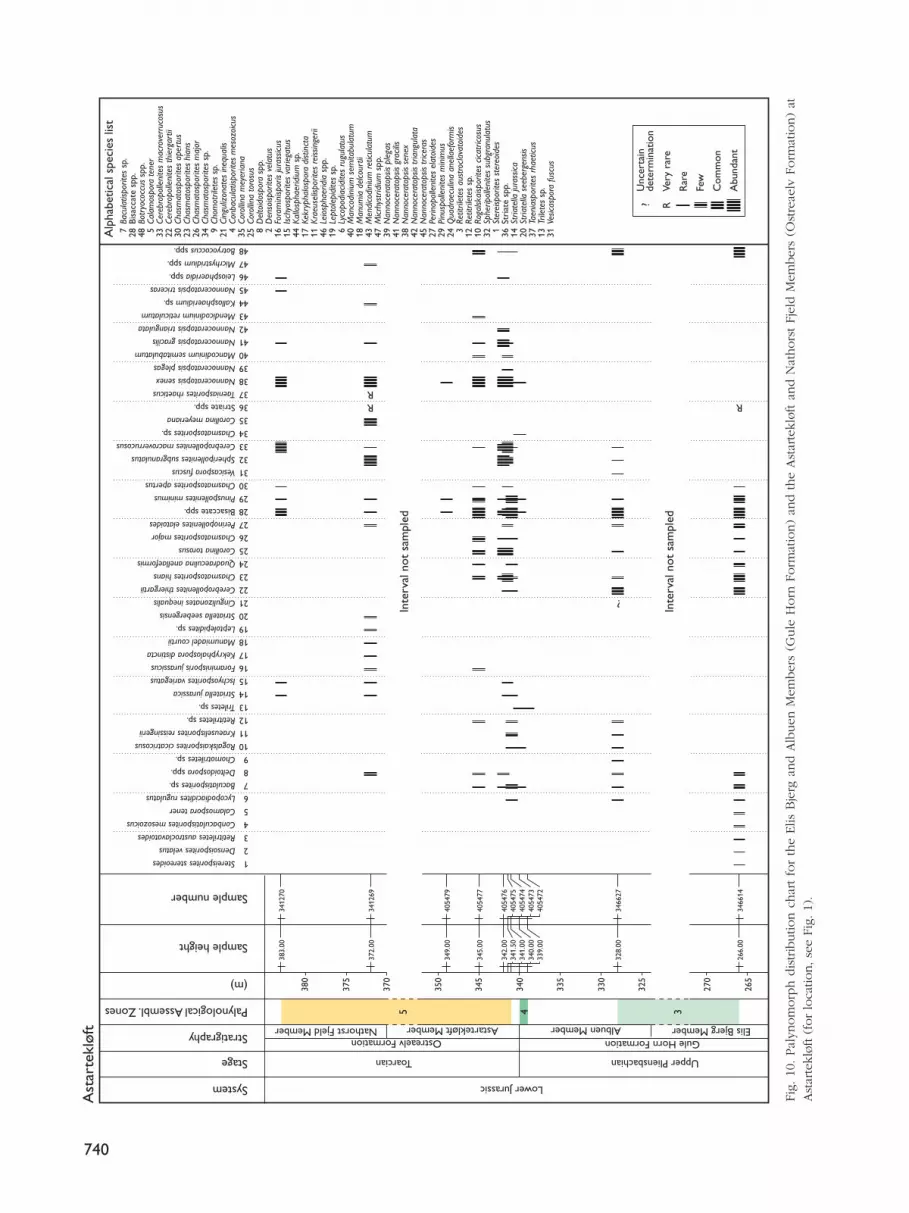
AstartekløftFourteen samples from the northern and southern sideof Astartekløft were investigated palynologically (Figs9, 10; note that the barren samples are not shown onFig. 10). The lowermost samples, 346614 at 266 m and346627 at 328 m, are from the lower Elis Bjerg Member(Gule Horn Formation) and the lower Albuen Member(Gule Horn Formation), respectively (Fig. 10). Thesesamples are dominated by Botryococcus sp. and bisac-cate pollen, together with common Cerebropollenitesthiergartii and Chasmatosporites hians, and are referredto Assemblage Zone 3.
The two samples 405472 and 405473 from the upperAlbuen Member (Figs 9, 10) contain abundant bisac-cate pollen; this and the lack of other palynomorphsindicate Assemblage Zone 4.
Samples 405474 and 405475 are from the lowermostbeds of the Astartekløft Member; they record the firstappearance of Spheripollenites subgranulatus, togetherwith abundant Cerebropollenites macroverrucosus andthe re-appearance of the dinoflagellate cyst Nanno-ceratopsis senex. Sample 405477, also from the Astarte-kløft Member, lacks Spheripollenites subgranulatus butNannoceratopsis senex is common. In sample 405483from the Astartekløft Member, the organic material isblack and indeterminate and thus this sample does notappear on Figure 10. In sample 341269, from the NathorstFjeld Member, Spheripollenites subgranulatus is abun-dant, and in the uppermost sample (341270), also fromthe Nathorst Fjeld Member, Spheripollenites subgranu-
latus is absent but Nannoceratopsis senex is abundanttogether with Cerebropollenites macroverrucosus. It issuggested that the assemblages recorded between sam-ple 405474, at the base of the Astartekløft Member, andthe uppermost sample 341270, in the Nathorst FjeldMember, are compatible with Assemblage Zone 5 asdefined from the Albuen section.
Suggested age. The lowermost part of the succession isreferred to the Upper Pliensbachian based on the abun-dance of Cerebropollenites thiergartii and Chasmato-sporites hians. An Early Toarcian age for the uppermostpart is based on the first appearance and abundanceof both Spheripollenites subgranulatus and Cerebro-pollenites macroverrucosus.
MoskusoksekløftOnly one sample (341260; Nathorst Fjeld Member,Ostreaelv Formation) was investigated from this local-ity; it yielded the dinoflagellate cysts Nannoceratopsissenex and N. triangulata and the pollen Spheripollenitessubgranulatus and Cerebropollenites macroverrucosus(Fig. 12A). This assemblage is referred to AssemblageZone 5.
Suggested age. An Early Toarcian age is proposed basedon the presence of Spheripollenites subgranulatus andCerebropollenites macroverrucosus together with Nanno-ceratopsis senex and N. triangulata.
Harris Fjeld One sample (346741) was studied from this locality,from the lower part of Elis Bjerg Member (Fig. 12B).The palynological assemblage is dominated by poorlypreserved bisaccate pollen and the assemblage wasdeemed too poor to determine to which assemblagezone it belongs.
Primulaelv Three samples (346746, 346745, 346753) were analysedfrom this locality (Fig. 12C), the first two from the ElisBjerg Member and the third from the lowermost mud-stones of the Skævdal Member. The lowermost sampleis tentatively referred to Assemblage Zone 1, based ona very poor assemblage of bisaccates, Pinuspollenites
745
746
Syst
emLo
wer
Jura
ssic
Stag
eTo
arci
an
Paly
nolo
gica
l Ass
embl
. Zon
es
Lith
ostr
atig
raph
yN
atho
rst
Fjel
d M
b
Ost
reae
lv F
orm
atio
n
Sam
ple
heig
ht
359.00Sa
mpl
e nu
mbe
r341260
1D
elto
idos
por
a sp
p.
2St
riat
ella
jura
ssic
a
3Is
chyo
spor
ites
varieg
atus
4Bac
ulat
ispor
ites
sp.
5Sp
heripol
leni
tes
subg
ranu
latu
s
6Cer
ebro
pol
leni
tes
mac
rove
rruc
osus
7Cor
ollin
a to
rosu
s
8Bi
sacc
ate
spp.
9Cor
ollin
a m
eyer
iana
10Cha
smat
ospor
ites
maj
or
11Cer
ebro
pol
leni
tes
thie
rgar
tii
12N
anno
cera
topsis
sene
x
13N
anno
cera
topsis
tria
ngul
ata
14Cym
atio
spha
era
sp.
15Fo
ram
inife
ra s
pp.
16Bot
ryoc
occu
s sp
p.
Alphabetical species list4 Baculatisporites sp.8 Bisaccate spp.
Fig. 12. Palynomorph distribution charts (for locations, see Fig. 1). A, Ostreaelv Formation (Nathorst Fjeld Member) at Moskusoksekløft.B, Gule Horn Formation (Elis Bjerg Member) at Harris Fjeld. C, Gule Horn Formation (Elis Bjerg Member) and Ostreaelv Formation(Nathorst Fjeld (Nath. F.) and Skævdal (Skæv.) Members) at Primulaelv.
minimus and common Botryococcus sp. The secondsample (346745) was barren and thus does not appearon the distribution chart (Fig. 12C). The uppermostsample is referred to Assemblage Zone 5 on the basisof abundant Spheripollenites subgranulatus, Cerebro-pollenites macroverrucosus and Nannoceratopsis senex.
Suggested age. A Late Pliensbachian age is tentativelysuggested for the lowermost sample (346746) based ona very poor assemblage in which only bisaccates andPinuspollenites minimus are common. An Early Toarcianage is suggested for the uppermost sample (346753)based on abundant Spheripollenites subgranulatus,Cerebropollenites macroverrucosus and Nannoceratopsissenex.
Lepidopteriselv Ten samples (139137–139146) were studied from thissection and all of them are rich in palynomorphs (Fig.13, following page 744). These samples were collected
by Claus Heinberg and Tove Birkelund in 1974, and theywere thus not assigned to the recently-defined mem-bers. However, comparing their field notes with oursedimentological logs, it has been possible to assign thesamples to the Elis Bjerg Member.
The samples 139237–139240 are rich in the sporesDeltoidospora and Baculatisporites, the pollen Pinus-pollenites minimus and Cerebropollenites thiergartii,bisaccate pollen and Botryococcus sp. In sample 139141,the dinoflagellate cyst Nannoceratopsis senex is veryabundant and Botryococcus sp. is rare. Samples139142–139146 are again rich in spores and pollen andBotryococcus sp. whereas dinoflagellate cysts are rare.
The palynological results allow us to suggest thatthe lowermost five samples (139137–139141) belongto Assemblage Zone 2. This is based on the commonoccurrence of Cerebropollenites thiergartii and Nanno-ceratopsis senex. The next five samples (139142–139146)are referred to Assemblage Zone 3 based on the pres-ence of Cerebropollenites thiergartii, Pinuspollenitesminimus and bisaccate pollen and the fact that dinofla-gellate cysts are rare.
747
LiaselvSy
stem
Low
er Ju
rass
ic
Stag
eU
pper
Plie
nsba
chia
n
Paly
nolo
gica
l Ass
embl
. Zon
es
Lith
ostr
atig
raph
yG
ule
Hor
n Fo
rmat
ion
Elis
Bje
rg M
embe
r
(m)
325
315
305
295
Sam
ple
heig
ht325.00
299.00
293.00
Sam
ple
num
ber
346665
346662
346667
1Bacu
latisp
orites
sp.
2D
elto
idos
por
a s
pp.
3Ly
copod
iaci
dites
rugu
latu
s
4Ret
itrile
tes
sem
imuris
5N
eora
istr
ickia
sp.
6Str
iate
lla s
eeber
gensis
7D
enso
ispor
ites
sca
nic
us
8Ret
itrile
tes
sp.
9Bi
sacc
ate
spp.
10Pi
nusp
olle
nites
min
imus
11Pe
rinop
olle
nites
ela
toid
es
12Cer
ebro
pol
lenites
thie
rgart
ii
13Chasm
ato
spor
ites
hia
ns
14Cer
ebro
pol
lenites
macr
over
ruco
sus
15Q
uadra
eculin
a a
nel
laef
orm
is
16Chasm
ato
spor
ites
majo
r
17Ve
sica
spor
a fusc
us
18Cor
ollin
a t
oros
us
19N
annoc
erato
psis
senex
20M
endic
odin
ium
sp.
21M
anco
din
ium
sem
itabula
tum
22D
inoc
yst
spp.
23A
crita
rch
spp.
24Bot
ryoc
occu
s sp
p.
?
Alphabetical species list23 Acritarch spp.1 Baculatisporites sp.9 Bisaccate spp.
Fig. 14. Palynomorph distribution chart for the Gule Horn Formation (Elis Bjerg Member) at Liaselv (for location, see Fig. 1).
Suggested age. A Late Pliensbachian age is proposed forthe samples 139137–139146 based on the presence ofCerebropollenites thiergartii, Pinuspollenites minimus,Nannoceratopsis species and Mancodinium semitabu-latum.
LiaselvThree samples (346662, 346665, 346667) were analysedfrom the Elis Bjerg Member (Fig. 14); in general, preser-vation of the palynomorphs is very poor. All three sam-ples are dominated by bisaccate pollen and Botryococcussp. and it is suggested that they belong to AssemblageZone 2.
Suggested age. A Late Pliensbachian age has been sug-gested because of the abundance of Pinuspollenitesminimus and bisaccates.
Horsedal and the Deltoidospora Assemblage Four samples (346696, 346700, 346701, 346703) wereanalysed from the Horsedal Member at Horsedal (Figs15, 16). Sample 346696 was barren. Sample 346700,from a coal bed, yielded an assemblage composed pre-dominantly of laevigate spores (pteridophyte spores)of the genus Deltoidospora. Such an assemblage has notbeen recorded before in samples from the Neill KlinterGroup at Albuen or at any other locality in Jameson Landand Scoresby Land. The assemblage totally lacksmicroplankton. Sample 346701 yielded a more diverseassemblage, but is dominated by bisaccate pollen andBotryococcus sp. The uppermost sample, 346703, yieldedonly bisaccate pollen.
It has not, based on the present material, been pos-sible to place this assemblage within any of the assem-blage zones defined above from the Albuen succession.The assemblage is thus defined as a new assemblage,named the Deltoidospora Assemblage, which is presentlyonly recognised at Horsedal in the Horsedal Memberof the Ostreaelv Formation.
Suggested age. This assemblage is not age specific; itcould occur within any stage of the Jurassic.
Palaeoenvironment. This assemblage is indicative ofan enclosed swamp area (lagoon, pond, small lake)with a dense vegetation of ferns.
Ranunkeldal Seven samples were analysed from this section (Figs 5,6). Samples 341167–341170 were sampled in the upper-most part of the Kap Stewart Group and samples
748
M
Si Pb
Sand
815
820
346700
825
346703
346701
830
m
Horsedal
Wav
e- a
nd s
torm
-dom
inat
ed la
goon
Del
toid
ospor
a A
ssem
blag
eO
stre
aelv
For
mat
ion
Hor
seda
l Mem
ber
Fig. 15. Sedimentological log through part of the Horsedal Member(Ostreaelv Formation) in Horsedal (for location, see Fig. 1).Sample numbers are indicated; arrows denote grain-size trends.For legend, see Fig. 3.
341171–341173 are from the Elis Bjerg Member of theGule Horn Formation. Preservation of the palynomorphsfrom all these samples is very poor. However, the paly-nomorph assemblages are dominated by the laevigatespore Deltoidospora sp., bisaccate pollen, Chasmato-sporites hians, Cerebropollenites thiergartii and Quadrae-culina anellaeformis. Botryococcus sp. is present in allsamples, but is only abundant in the uppermost sam-ple. In sample 341168, a dinoflagellate cyst has beenfound; it is similar to Mendicodinium reticulatum, butshows some anomalous features. The occurrence ofthis dinoflagellate cyst suggests that the environmentwas influenced by brackish waters, at least for a shorttime. A single Nannoceratopsis gracilis cyst was observedin sample 341171 from 306 m. In this sample, aTasmanites was found together with a poorly preservedforaminiferal inner-lining. In the uppermost sample,several Leiofusa jurassica were recorded.
The palynomorph assemblages from the Kap StewartGroup in Ranunkeldal are suggested to belong to aseparate assemblage zone. The samples from the ElisBjerg Member are referred to Assemblage Zone 1.
Suggested age. A ?Late Sinemurian age is suggested forthe Kap Stewart Group samples because of the occur-rence of the dinoflagellate comparable to Mendico-dinium reticulatum. This species has been found onBornholm, Denmark in sediments of latest Sinemurianand earliest Pliensbachian age (Batten et al. 1994;Koppelhus & Nielsen 1994). A Pliensbachian age is sug-
gested for the Elis Bjerg Member samples because ofthe presence of Nannoceratopsis gracilis.
Palaeoenvironment. A non-marine, freshwater envi-ronment is indicated for the Kap Stewart Group sam-ples, although the presence of a dinoflagellate cyst insample 341168 suggests the influence of brackish water,albeit only temporarily. The samples from the Elis BjergMember indicate marine influence.
Depositional environments and assemblage zonesThe palynological results presented above provide anadditional dataset with which to constrain palaeoenvi-ronmental and sequence stratigraphic interpretations.In the following section, the individual palynologicalassemblage zones, together with the DeltoidosporaAssemblage, are discussed in relation to the sedimen-tological and stratigraphic data.
Assemblage Zone 1:Cerebropollenites thiergartii – Pinuspollenitesminimus – Botryococcus
This assemblage zone is characteristic of the sedimen-tary succession referred to sequence SQ2 of Dam &Surlyk (1995, 1998) at Qupaulakajik, Albuen and
749
HorsedalSy
stem
Stag
e
Lith
ostr
atig
raph
yO
stre
aelv
For
mat
ion
Hor
seda
l Mem
ber
Paly
nolo
gica
l Ass
embl
age
? ?
Del
toid
ospor
a
(m)
825
815
Sam
ple
heig
ht828.00
823.00
813.00
Sam
ple
num
ber
346703
346701
346700
1D
elto
idos
por
a s
pp.
2Bacu
latisp
orites
sp.
3Cala
mos
por
a t
ener
4Ret
itrile
tes
sp.
5N
eora
istr
ickia
sp.
6Ret
itrile
tes
aust
rocl
ava
toid
es
7Ly
copod
iaci
dites
rugu
latu
s
8D
enso
ispor
ites
sca
nic
us
9Chasm
ato
spor
ites
majo
r
10Pe
rinop
olle
nites
ela
toid
es
11Cer
ebro
pol
lenites
thie
rgart
ii
12Spher
ipol
lenites
psila
tus
13Cer
ebro
pol
lenites
macr
over
ruco
sus
14Chasm
ato
spor
ites
hia
ns
15Pi
nusp
olle
nites
min
imus
16Bi
sacc
ate
spp.
17Ve
sica
spor
a fusc
us
18Q
uadra
eculin
a a
nel
laef
orm
is
19Pa
reod
inia
halo
sa
20M
endic
odin
ium
sp.
21N
annoc
erato
psis
sp.
22Bot
ryoc
occu
s sp
p.
?
Alphabetical Species List2 Baculatisporites sp.
16 Bisaccate spp.22 Botryococcus spp.3 Calamospora tener
Fig. 16. Palynomorph distribution chart for the Horsedal Member (Ostreaelv Formation) in Horsedal (for location, see Fig. 1).
Goniomyakløft, which consists of the upper part of theRævekløft Formation and the lowermost part of the ElisBjerg Member (Fig. 17). It is also present in the lowerpart of sequence SQ3 at Primulaelv, however, and inthe lowermost part of the Elis Bjerg Member in Ranunkel-dal, just above the Kap Stewart Formation. The base ofthe assemblage zone thus coincides with the base ofsequence SQ1 (i.e. SB1) and the zone extends up intothe lowermost part of sequence SQ3 of Dam & Surlyk(1995, 1998). At the studied locations, the upper partof the Rævekløft Formation consists of cross-bedded,fossiliferous medium- to very coarse-grained sandstones(Fig. 7), interpreted to represent fields of dunes orshoreface ridges on the shoreface (Dam & Surlyk 1995,1998). In the Elis Bjerg Member, the assemblage is pre-sent in subtidal sand sheet, shoreface and offshore tran-sition deposits (Figs 3A, 5). The Rævekløft Formationis capped by an important drowning surface that definesthe base of the Elis Bjerg Member (Figs 7, 17; Dam &Surlyk 1995, 1998).
The palynological assemblage is uniform, being dom-inated by bisaccate pollen and the freshwater to brack-ish alga Botryococcus sp. Spores include commonDeltoidospora and Baculatisporites sp., and locallyLycopodiacidites rugulatus. Among the pollen, Pinus-pollenites minimus and Cerebropollenites thiergartii arecommon. Acritarchs are rare and dinoflagellate cysts areabsent with the exception of one Nannoceratopsis gra-cilis cyst and the undetermined dinoflagellate cyst (cf.Mendicodinium reticulatum) from the Ranunkeldalsection. The assemblage probably reflects a vegetationwith few fern species and several gymnosperm species.These grew close to a fresh or brackish water envi-ronment, where the Botryococcus algae lived. Whenseen in the light of the strong marine indicators pro-vided by the macrofossils, sedimentary structures andichnology, it is suggested that this palynomorph assem-blage is dominantly allochthonous, having been trans-ported from a terrestrial to a shallow marine environ-ment. Similar palynomorph assemblages are knownfrom other areas, for example in the uppermostSinemurian and lowermost Pliensbachian of Bornholmin the Baltic Sea (Koppelhus & Nielsen 1994).
Assemblage Zone 2:Nannoceratopsis–Botryococcus
The assemblage zone is characteristic of the middlepart of the Elis Bjerg Member at Albuen, Lepidopteriselvand Liaselv in subtidal sand sheet and storm-dominated
shoreface deposits, but also occurs in similar depositsin the lower part of the member at Goniomyakløft andthe upper part of the member at Qupaulakajik (Figs 2,17). The assemblage zone is most characteristic of thelower part of sequence SQ3 of Dam & Surlyk (1995,1998), but is also locally present in the uppermost partof sequence SQ2 (Fig. 17). The top is placed below thetransition from subtidal sand sheet deposits to tidalchannel deposits of the Elis Bjerg Member (Fig. 3A).
The palynomorph assemblage is dominated by thesame spores, pollen and Botryococcus sp. that charac-terise Assemblage Zone 1, but the incoming of Nanno-ceratopsis senex, N. gracilis, Parvocysta barbata andMancodinium semitabulatum together with Limbicystabjaerkei and a few more acritarchs indicates an increasein marine influence. In most of the samples, there arebetween three and seven different dinoflagellate cystspecies and between one and five acritarch species.
The marine interpretation of this palynomorph assem-blage zone is in agreement with the sedimentologicaland ichnological data, which also indicate a shallowmarine environment (Dam & Surlyk 1995, 1998).
Assemblage Zone 3:Chasmatosporites – Cerebropollenites thiergartii– Botryococcus
The assemblage is characterised by rare dinoflagellatecysts such as Mendicodinium reticulatum and Nanno-ceratopsis spp. A few acritarchs are present, togetherwith abundant pollen and Botryococcus. The assemblageis present in the upper part of the Elis Bjerg Member,but may extend into the lowermost part of the AlbuenMember (Fig. 17). Assemblage Zone 3 is dominantly pre-sent in stacked tidal channel and wave- and storm-dom-inated shoreface deposits (Fig. 3A; Dam & Surlyk 1995,1998), whereas the sample from the Albuen Memberwas from heterolithic lower shoreface deposits. Thebase of the assemblage zone is placed just below thetransition from subtidal sand sheet deposits to tidalchannel deposits. At Albuen, the top of the zone occursjust beneath the boundary between tidal channeldeposits of the Elis Bjerg Member and storm-dominatedoffshore transition deposits of the Albuen Member; thisboundary has been interpreted as a coalesced sequenceboundary and transgressive surface (Dam & Surlyk1995, 1998). At Astartekløft, however, the lowermost sam-ple in the Albuen Member is also referred to AssemblageZone 3 (Fig. 10).
750
751
050
100
150
km
50 m
HST
HST
HST
HST
HST
TST
TST
SQ6
SQ5
SQ7
SQ4
SQ3
SQ2
SQ1
Tre
fjord
Bje
rg
Qupaulakajik/Rævekløft
Albuen
Goniomyakløft
Astartekløft
Moskusoksekløft
Harris Fjeld (N)
Nathorst Fjeld
Dusén BjergT
idal
cha
nnel
s an
dsu
btid
al s
hoal
s
Rhætelv
Horsedal
Halten Terrace,Norway
Lepidopteriselv/Liaselv
Primulaelv
Harris Fjeld (S)
Ast
arte
kløf
t/H
orse
dal
Skæ
vdal
Mb
Fm
Sortehat Ostreaelv Gule Horn
Alb
uen
Elis
Bje
rg
Ræ
vekl
øft
TST
TST
TST
HST
TST
TSTLST
Nat
hors
t Fj
eld/
Har
ris
Fjel
d/Le
pido
pter
isel
v
Sout
hN
orth
Shor
efac
e
Ter
min
al lo
be
Lago
on
Offs
hore
tra
nsiti
on
Faci
es a
ssoc
iatio
n bo
unda
ry
Sequ
ence
bou
ndar
y
Maj
or fl
oodi
ng s
urfa
ce
Not Ile Ror
Tilj
e
Åre
Ass
embl
age
Zon
e 4
Ass
embl
age
Zon
e 3
Ass
embl
age
Zon
e 2
Ass
embl
age
Zon
e 1
Del
toid
ospor
a A
ssem
blag
e
Ass
embl
age
Zon
e 7
Ass
embl
age
Zon
e 6
Ass
embl
age
Zon
e 5
Fig.
17.
North–s
outh
corr
elat
ion p
anel
of th
e Lo
wer
– lo
wer
Mid
dle
Jura
ssic
Nei
ll K
linte
r G
roup o
f Ja
mes
on L
and, E
ast G
reen
land s
how
ing
the
dis
trib
utio
n o
f th
e pal
ynolo
gica
l ass
em-
bla
ge z
ones
rep
orted
her
e re
lativ
e to
the
mai
n s
equen
ce s
trat
igra
phic
ele
men
ts a
nd th
e dep
osi
tional
envi
ronm
ents
. The
corr
elat
ive
form
atio
ns
of th
e H
alte
n T
erra
ce, o
ffsh
ore
Norw
ay,
are
show
n o
n the
righ
t-han
d s
ide
of th
e figu
re. N
ote
that
pal
ynolo
gica
l dat
a ar
e sc
arce
north o
f Ast
arte
kløft. The
dotted
ver
tical
lin
es indic
ate
the
loca
litie
s on w
hic
h the
sequen
cest
ratig
raphic
inte
rpre
tatio
n o
f D
am &
Surlyk
(19
98)
is b
ased
. Ass
embla
ge Z
ones
8 a
nd 9
in the
Sorteh
at F
orm
atio
n a
re n
ot in
dic
ated
; th
e re
ader
is
refe
rred
to K
oppel
hus
& H
anse
n(2
003,
this
volu
me)
. Fi
gure
modifie
d fro
m D
am &
Surlyk
(19
98). S
Q, se
quen
ce;
LST, lo
wst
and s
yste
ms
trac
t; T
ST, tran
sgre
ssiv
e sy
stem
s trac
t; H
ST, hig
hst
and s
yste
ms
trac
t.
The shift from Assemblage Zone 2 to AssemblageZone 3 records a change from a marine setting to anenvironment with a strongly terrestrial character, albeitwith evidence of periodic marine influence. This flo-ral/faunal change is compatible with the sedimento-logical record (Dam & Surlyk 1995, 1998), whichindicates a change from subtidal sand sheet depositionin a shallow marine environment to a tidal channelenvironment, where a larger degree of terrestrial influ-ence is to be expected.
Assemblage Zone 4: BisaccatesThis palynomorph assemblage is very uniform, beingdominated overwhelmingly by bisaccate pollen. Thecombined data from the Albuen and Astartekløft sec-tions suggest that the assemblage characterises muchof the Albuen Member of the Gule Horn Formation. AtAlbuen, the base of the assemblage zone occurs imme-diately above the coalesced sequence boundary andtransgressive surface that separates the Elis Bjerg andAlbuen Members (Fig. 3B; SB4 of Dam & Surlyk 1995,1998); at Astartekløft, in contrast, Assemblage Zone 3straddles this sequence boundary, Assemblage Zone 4being recognised only in the uppermost levels of theAlbuen Member (Fig. 10).
The upper levels of the Albuen Member proved inac-cessible at Albuen and were not sampled; the top ofthe zone is placed at the uppermost sample, some 12 mbelow the top of the member. At Astartekløft, however,detailed sampling across the boundary between theAlbuen and Astartekløft Members demonstrated thatthe boundary between Assemblage Zones 4 and 5 coin-cides closely with this surface which is interpreted asa sequence boundary (Figs 9, 10, 17; SB5 of Dam &Surlyk 1995, 1998).
The Albuen Member is heterolithic, being composedof alternating mudstones and well-sorted fine-grainedsandstones deposited in a storm-dominated lowershoreface environment (Dam & Surlyk 1995, 1998). Afew coarse-grained pebbly sheets, moulded into largesymmetrical ripples, and massive sandy mudstonesdeposited from debris flows are commonly interbed-ded with the heterolithic deposits.
It is well-known that bisaccate pollen is commonlyconcentrated in distal marine settings, beyond the reachof other terrestrial palynomorphs. On this basis, thisassemblage could therefore be interpreted to reflect anoffshore marine environment, beyond the depositionalrange of other land-derived elements. The sedimento-
logical data, however, testify to a lower shoreface envi-ronment (Dam & Surlyk 1998), and other land-derivedelements should therefore be present. The absence ofmarine palynomorphs is also difficult to explain. Asdiscussed earlier, such thin-walled forms may have beenselectively destroyed by thermal effects caused by theintrusion of volcanics into the sediments. Alternatively,the lack of marine palynomorphs could reflect partial iso-lation of the embayment resulting in the developmentof a fresh to brackish water environment; this could alsoexplain the lack of tidal indicators in this member.
Assemblage Zone 5:Spheripollenites subgranulatus – Cerebropollenitesmacroverrucosus – Luehndea spinosa
The base of Assemblage Zone 5 in the Albuen sectionis characterised by the sudden incoming of Spheri-pollenites subgranulatus and the reappearance of dinofla-gellate cysts together with a more diverse pollen floraand the freshwater alga Botryococcus sp. In sample405466 at 259 m in the Albuen section (Figs 3B, 4A),Luehndea spinosa appears for the first time together withcommon spherical dinocysts that are of unknown affin-ity, but have been recorded from Spitsbergen, the DanishSubbasin and Bornholm, Denmark (Bjærke 1980a;Dybkjær 1991; Koppelhus & Nielsen 1994).
At Astartekløft, the lower zone boundary is placedjust above the sequence boundary between the Albuenand Astartekløft Members (Figs 9, 17; SB5 of Dam &Surlyk 1995); the upwards extent of the zone is poorlyconstrained at this locality. At Albuen, the upper bound-ary of the zone is placed approximately 10 m belowthe drowning surface that separates the heavily bio-turbated shoreface sandstones of the Nathorst FjeldMember from bioturbated shelf deposits of the SkævdalMember (Fig. 3C). At Primulaelv, a single sample showsthat this assemblage is also present just above the drown-ing surface (Fig. 17). Along Neill Klinter, the AstartekløftMember includes three facies associations, tidal chan-nel, subtidal sand sheet and storm-dominated sandyshoal associations (Fig. 3B). The tidal channel and sub-tidal sand sheet deposits are similar to those of the ElisBjerg Member, discussed above under AssemblageZones 2 and 3. The storm-dominated sandy shoaldeposits form a laterally continuous succession, com-posed of well-sorted fine- to medium-grained sand-stone beds (Dam & Surlyk 1995, 1998). The NathorstFjeld Member forms a single coarsening-upwardssuccession consisting of alternating silty mudstones and
752
thin laminae of very fine- to fine-grained sandstones,grading upwards into fine- to coarse-grained sand-stones. The sandstones are cross-bedded, wave ripplecross-laminated, hummocky cross-stratified and bio-turbated. The coarsening-upwards succession reflectsan increase in energy with time and is interpreted torecord progressive shallowing from an offshore transi-tion setting to a shoreface environment.
The Spheripollenites subgranulatus – Cerebropollenitesmacroverrucosus – Luehndea spinosa assemblage com-prises a mixture of spores, pollen, a few dinoflagellatecyst species and acritarchs and the freshwater algaBotryococcus. The assemblage zone indicates a brack-ish to marine environment with a large input of terres-trial material. This is in accordance with the sedi-mentological data indicating various environments in amarginal shallow marine setting.
Assemblage Zone 6: Perinopollenites elatoides
Assemblage Zone 6, which is typical of the Skævdal andthe Trefjord Bjerg Members (Fig. 17) is characterised bythe abundance of the pollen Perinopollenites elatoidesand the absence or scarcity of Spheripollenites sub-granulatus. Overall, the palynomorph assemblage isdominated by different pollen species but dinoflagel-late cysts are also significant, including Dissiliodiniumsp., Phallocysta eumekes, Pareodinia halosa, andKallospharidium sp.; acritarchs are also present.
At Albuen, in the south-eastern part of the basin, thelower boundary is placed some 10 m below the drown-ing surface that defines the top of the Nathorst FjeldMember (Figs 3C, 17). The upper boundary of theassemblage zone at Albuen is placed at 434 m in theupper Trefjord Bjerg Member. Succeeding samples, justbelow the boundary between the Trefjord Bjerg Memberand the Sortehat Formation, are referred to AssemblageZone 7, which is characteristic of the lower SortehatFormation (Figs 3C, 17). At 437 m, between these twosampled levels, is an erosional surface that is drapedby well-rounded quartzite pebbles up to 3 cm across;this surface is defined as a sequence boundary (SB7 ofDam & Surlyk 1998).
The Skævdal Member consists of bioturbated muddysandstones and deposition probably took place in alow-energy shelf environment (Dam & Surlyk 1995,1998). Primary physical structures only occur locallyand include wave ripple cross-lamination, cross-lami-nation and cross-bedding. Stratigraphic variations inthe mud content suggests that the heavily bioturbated
muddy sandstones were originally deposited as het-eroliths (Dam & Surlyk 1995, 1998).
The Skævdal Member is truncated by a prominentbasinwide erosional unconformity, in places draped bya lag conglomerate. The unconformity marks a basin-wide seawards shift in facies and is interpreted as asequence boundary (SB6 of Dam & Surlyk 1995, 1998).At Albuen, the sequence boundary is overlain by sub-tidal cross-bedded sandstones of the Trefjord BjergMember deposited in an extensive subtidal dune field.
The palynomorph Assemblage Zone 6 is indicativeof deposition in a marine environment with a largeinput of terrigenous material. This is in accordance withthe sedimentological data that indicate a shallow marineenvironment (Dam & Surlyk 1995, 1998).
Assemblage Zone 7: Botryococcus
This assemblage is characterised by abundant Botry-ococcus. In a few samples near the lower boundary ofthe zone, both Botryococcus and dinoflagellate cystsoccur in abundance, but the latter become rare upwardswithin the zone; the top of the zone is marked by there-appearance of dinoflagellate cysts and the disap-pearance of Botryococcus. The depositional environmentof this assemblage zone is discussed in detail in an accom-panying paper (Koppelhus & Hansen 2003, this volume).
Deltoidospora Assemblage This assemblage is restricted to the Horsedal Memberin the northern part of the basin (Figs 15–17). Thismember is made up of minor coarsening-upwards suc-cessions, 1–6 m thick, deposited in wave-dominatedbeaches or delta systems that prograded into an exten-sive lagoonal environment (Dam & Surlyk 1995, 1998).
The palynomorph assemblage is overwhelminglydominated by laevigate spores (pteridophyte spores) ofthe genus Deltoidospora and is suggestive of an enclosedswamp area (lagoon, pond, small lake) with a densevegetation of ferns. This is in close agreement with thedepositional environment suggested by sedimentaryfacies analysis (Dam & Surlyk 1995, 1998).
Discussion and conclusionsSeven palynological assemblage zones have been recog-nised in the Rævekløft, Gule Horn and Ostreaelv
753
Formations of the Neill Klinter Group (Fig. 17). Six ofthese are defined in this paper; the uppermost zone isdefined by Koppelhus & Hansen (2003, this volume)in an accompanying paper as it is most characteristicof the overlying Sortehat Formation, the uppermost for-mation of the Neill Klinter Group (Koppelhus & Hansen2003, this volume). In addition, an assemblage termedthe Deltoidospora Assemblage is defined here from theHorsedal section.
The seven palynological assemblage zones were allrecognised primarily on the basis of data from theAlbuen section but additional data from other localitiessuggest that the zones may have a basinwide distri-bution (Figs 17, 18). The palynological assemblagescontain a diverse palynoflora, including 136 species. Theassemblages indicate that the Neill Klinter Group spansthe Early Pliensbachian to early Aalenian, without anymajor breaks in the stratigraphic record. The study
754
AZ7
AZ6
AZ5
AZ4
AZ5
AZ3
AZ3
AZ2
AZ1
SB7
SB6
SB5
SB4
SB3
SB2
SB1
Aal
enia
nTo
arci
anLa
te P
liens
bach
ian
Sort
ehat
Fm
Ost
reae
lv F
mN
eil K
linte
r G
roup
Gul
e H
orn
FmR
æve
kløf
t Fm
KS
Sin.
Earl
y Pl
iens
-ba
chia
n
Skæ
vdal
Mb
Nat
hors
t Fj
eld
Mb
Ast
arte
kløf
t M
bEl
is B
jerg
Mb
Alb
uen
Mb
Tref
jord
Bjer
g M
b
?
AZ2
AZ5
AZ3
AZ2 AZ2
AZ1 AZ1
AZ2
AZ4
AZ1
AZ5
Alb
uen
Ast
arte
kløf
t
Ræ
vekl
øft
Gon
iom
yakl
øft
Lepi
dopt
eris
elv
Lias
elv
Ran
unke
ldal
Har
ris
Fjel
dPr
imul
aelv
Mos
kuso
ksek
løft
AZ1
Fig. 18. Correlation diagram showing thestratigraphic distribution of the palyno-logical assemblage zones (AZ1–7) at thestudied locations (see Fig. 1). Note thatthe boundaries between AssemblageZones 1 and 2 and between AssemblageZones 3 and 4 are ‘diachronous’ relativeto sequence boundaries SB3 and SB4respectively. KS, Kap Stewart Group;Sin., Sinemurian.
755
shows that the palynomorph flora of the Neill KlinterGroup is strongly influenced by the amount of organicmatter derived from land plants and freshwater envi-ronments, yet the brackish and marine microplanktonplay a very important role in the interpretation of theenvironment and in establishing a palynostratigraphyfor the group.
Comparing the sequence stratigraphic frameworkdeveloped by Dam & Surlyk (1995, 1998) with the dis-
tribution of the palynological assemblages, it is clear thatsome of the major sequence stratigraphic and litho-logical boundaries are reflected by changes in the assem-blages. Assemblage Zones 1–3 are characteristic ofsequences SQ1–3. Figures 17 and 18 illustrate that theboundary between Assemblages 1 and 2 is diachro-nous on a regional scale with respect to the importantsequence stratigraphic surfaces (e.g. SB3). Given thatthe sole difference between these two zones is the pres-
Sphaeromorph clusters acmeChasmatosporites sp.N. gracilis/senex acmeSpheripollenites acmeAbundant–common L. spinosa
Luehndea spinosa FADC. thiergartii present but becomesrare in younger sediments
Mancodinium semitabulatum
N. senex/gracilis FADAbundant–common Botryococcus spp.
Common acritarchs
Fig. 19. Diagram showing major palyno-events in the Neill Klinter Groupcompared to the Early Jurassic palyno-events recorded from the Halten Terrace,mid-Norwegian shelf (I. Throndsen,personal communication 1996). FAD, firstappearance datum.
756
ence/absence of marine dinoflagellates, such diachrone-ity is not surprising. It can be attributed to variations interrestrial input and the dominance of the freshwaterplume laterally along the basin margin.
The transition from Assemblage Zone 2 to AssemblageZone 3 is marked by a decrease in dinoflagellate cystswhich coincides with a gradual overall change from adominance of subtidal sand sheet deposits to a domi-nance of tidal channel deposits in the upper part of theElis Bjerg Member (Figs 3A, 17; Dam & Surlyk 1998).The strengthening of the terrestrial signal recorded bythe change in the palynological assemblage is thus inaccordance with the sedimentological record.
At Albuen, sequence boundary SB4 separatesAssemblage Zones 3 and 4; at Astartekløft, however, asample from a few metres above the sequence bound-ary is referred to Assemblage Zone 3. This may be dueto reworking of the uppermost sediments of the Elis BjergMember in the underlying sequence. Palynological datafrom the Astartekløft section suggest that SB5 separatesAssemblage Zones 4 and 5, whereas the transition fromAssemblage Zone 5 to 6 appears to be diachronous(Fig. 17).
Sequence boundary SB7 separates Assemblage Zones6 and 7 (Fig. 17). With respect to the sequence strati-graphic interpretation given by Dam & Surlyk (1995),an important conclusion of this study is that AssemblageZone 7, characteristic of the lower part of the overly-ing Sortehat Formation (Koppelhus & Hansen 2003,this volume), also occurs in the uppermost few metresof the Trefjord Bjerg Member at Albuen, above a later-ally persistent erosional surface with a conglomerate lag(Figs 3C, 17). At other localities, a thin conglomeratelayer separates the Trefjord Bjerg Member and theSortehat Formation. Dam & Surlyk (1995) interpretedthe Trefjord Bjerg Member – Sortehat Formation bound-ary as a coalesced sequence boundary and transgres-sive surface. However, the palynological data suggestthat at Albuen the sequence boundary should be placedbeneath this boundary, at the conglomerate-draped ero-sional surface, and thus that the uppermost sandstones(c. 3 m thick) of the Trefjord Bjerg Member representa thin lowstand or transgressive package (Figs 3C, 17).Moreover, the palynological data suggest that the inferredmajor flooding surfaces in the sequence stratigraphicanalysis (i.e. base Albuen Member and base SortehatFormation), may in fact record partial isolation of theembayment from the seaway between Norway andGreenland causing a freshwater to brackish environmentto develop. This would also explain the lack of tidalindicators in the Albuen Member. If this is the case, the
Albuen Member does not represent transgressive andhighstand deposits in a sequence stratigraphic sense,but rather a period of physical isolation of the basin fromthe sea.
The palynostratigraphy has proven to be an impor-tant tool in confirming the high-resolution sequencestratigraphic correlation between East Greenland andthe Halten Terrace of the mid-Norwegian shelf pro-posed by Dam & Surlyk (1995). This detailed palyno-logical study should make it possible to correlate to otherlocalities in the North Atlantic and to the mid-Norwegianarea and the northern part of the North Sea. The pat-tern of events seen in the East Greenland material ismirrored by data from the Halten Terrace. Thus, accord-ing to I. Throndsen (personal communication 1996), thePliensbachian is characterised by common to abundantBotryococcus; this is followed by the incoming of thedinoflagellate cysts Nannoceratopsis gracilis, N. senexand Mancodinium semitabulatum together with com-mon to abundant Cerebropollenites thiergartii pollen andabundant bisaccate pollen in the Upper Pliensbachian(Fig. 19). The dinoflagellate cyst Luehndea spinosaoccurs in the uppermost Pliensbachian and lowermostToarcian together with Nannoceratopsis gracilis and N.senex followed by an acme of Spheripollenites sub-granulatus together with the sphaeromorph clusters. Inthe Upper Toarcian, the dinoflagellate cyst Parvocystacomplex appears together with abundant Perinopolleniteselatoides pollen and the first Callialasporites dampieriappear together with the dinoflagellate cyst Wallodiniumin the lowermost Aalenian.
AcknowledgementsThe work carried out in connection with this projectby E.B.K. was supported by a grant from the EFP-93Projects 1313/93-0010 and 0017. The work of G.D. hasbeen supported by BP Exploration Operating CompanyLimited, London, and the Carlsberg Foundation. Themanuscript was read by Karen Dybkjær, James B. Riding,Jon R. Ineson and Finn Surlyk who offered many help-ful suggestions. In particular, we thank Ingar Throndsenwho placed unpublished data from the Halten Terraceat our disposal, Henrik Nøhr-Hansen for patient assis-tance with the range charts and Jon R. Ineson for care-ful editing of the manuscript.
Triassic to Middle Jurassic palynofacies and palynomiscel-lanea in the Danish Basin and Fennoscandian Border Zone.Cahiers de Micropaléontologie 9, 21–45.
Bjærke, T. 1980a: Mesozoic palynology of Svalbard V. Dino-flagellates from the Agardhfjellet Member (Middle and UpperJurassic) in Spitsbergen. Norsk Polarinstitutt Skrifter 172,145–167.
Bjærke, T. 1980b: Mesozoic palynology of Svalbard IV. Toarciandinoflagellates from Spitsbergen. Palynology 4, 57–77.
Bromley, R.G. & Asgaard, U. 1979: Triassic freshwater ichno-coenoses from Carlsberg Fjord, East Greenland. Palaeo-geography, Palaeoclimatology, Palaeoecology 28, 39–80.
Callomon, J.H. 1961: The Jurassic system in East Greenland. In:Raasch, G.O. (ed.): Geology of the Arctic 1, 258–268. Toronto:University of Toronto Press.
Clemmensen, L.B. 1978a: Alternating aeolian, sabkha and shal-low-lake deposits from the Middle Triassic Gipsdalen Formation,Scoresby Land, East Greenland. Palaeogeography, Palaeo-climatology, Palaeoecology 24, 111–135.
Clemmensen, L.B. 1978b: Lacustrine facies and stromatolites fromthe Middle Triassic of East Greenland. Journal of SedimentaryPetrology 48, 1111–1128.
Clemmensen, L.B. 1979: Triassic lacustrine red-beds and palaeo-climate: the ‘Buntsandstein’ of Helgoland and the Malmros KlintMember of East Greenland. Geologische Rundschau 68,748–774.
Clemmensen, L.B. 1980a: Triassic rift sedimentation and palaeo-geography of central East Greenland. Bulletin GrønlandsGeologiske Undersøgelse 136, 72 pp.
Clemmensen, L.B. 1980b: Triassic lithostratigraphy of East Green-land between Scoresby Sund and Kejser Franz Josephs Fjord.Bulletin Grønlands Geologiske Undersøgelse 139, 56 pp.
Dam, G. 1990a: Taxonomy of trace fossils from the shallow marineLower Jurassic Neill Klinter Formation, East Greenland. Bulletinof the Geological Society of Denmark 38, 119–144.
Dam, G. 1990b: Palaeoenvironmental significance of trace fos-sils from the shallow marine Lower Jurassic Neill KlinterFormation, East Greenland. Palaeogeography, Palaeoclima-tology, Palaeoecology 79, 221–248.
Dam, G. 1991: A sedimentological analysis of the continental andshallow marine Upper Triassic to Lower Jurassic successionin Jameson Land, East Greenland, 243 pp. Unpublished Ph.D.thesis, University of Copenhagen, Denmark.
Dam, G. & Surlyk, F. 1992: Forced regressions in a large wave-and storm-dominated anoxic lake, Rhaetian–Sinemurian KapStewart Formation, East Greenland. Geology 20, 749–752.
Dam, G. & Surlyk, F. 1993: Cyclic sedimentation in a large wave-and storm-dominated anoxic lake; Kap Stewart Formation(Rhaetian–Sinemurian), Jameson Land, East Greenland. In:Posamentier, H.W. et al. (eds): Sequence stratigraphy andfacies associations. International Association of Sedimentolo-gists Special Publication 18, 419–448.
Dam, G. & Surlyk, F. 1995: Sequence stratigraphic correlation ofLower Jurassic shallow marine and paralic successions across
the Greenland–Norway seaway. In: Steel, R.J. et al. (eds):Sequence stratigraphy on the Northwest European margin.Norwegian Petroleum Society (NPF) Special Publication 5,483–509.
Dam, G. & Surlyk, F. 1998: Stratigraphy of the Neill Klinter Group;a Lower – lower Middle Jurassic tidal embayment succession,Jameson Land, East Greenland. Geology of Greenland SurveyBulletin 175, 80 pp.
Doyle, P. 1991: Belemnites from the Lower Jurassic of East Green-land and their biostratigraphical and biogeographical signi-ficance. Bulletin of the Geological Society of Denmark 39,123–141.
Dybkjær, K. 1991: Palynological zonation and palynofacies inves-tigation of the Fjerritslev Formation (Lower Jurassic – basalMiddle Jurassic) in the Danish Subbasin. Danmarks GeologiskeUndersøgelse Serie A 30, 150 pp.
Engkilde, M. 1994: The Middle Jurassic Vardekløft Formation,East Greenland: depositional environments and sequencestratigraphy of shallow marine sandstones deposited in a low-gradient epeiric seaway, 207 pp. Unpublished Ph.D. thesis,University of Copenhagen, Denmark.
Feist-Burkhardt, S. & Monteil, E. 1994: Wallodinium cylindricumand ‘Wallodinium elongatum’: stratigraphic problem and tax-onomical solution. Cahiers de Micropaléontologie 9, 5–15.
Fenton, J. & Riding, J.B. 1987: Kekryphalospora distincta gen. etsp. nov., a trilete spore from the Lower and Middle Jurassicof North-West Europe. Pollen et Spores 19, 427–434.
Guy-Ohlson, D. 1988: Toarcian palynostratigraphical correlationswithin and between different biogeographical provinces. In:Rocha, R.B. & Soares, A.F. (eds): 2nd International Symposiumon Jurassic Stratigraphy (Lisbon 1987) 1, 807–820. Lisbon:Universidade Nova de Lisboa.
Guy-Ohlson, D. 1992: Botryococcus as an aid in the interpreta-tion of palaeoenvironment and depositional processes. Reviewof Palaeobotany and Palynology 71, 1–15.
Harris, T.M. 1931: Rhaetic floras. Biological Reviews of theCambridge Philosophical Society 6, 133–162.
Koppelhus, E.B. & Batten, D.J. 1996: Applications of a paly-nomorph zonation to a series of short borehole sections,Lower to Middle Jurassic, Øresund, Denmark. In: Jansonius,J. & McGregor, D.C. (eds): Palynology: principles and appli-cations. American Association of Stratigraphic PalynologistsFoundation 2, 779–793.
Koppelhus, E.B. & Hansen, C.F. 2003: Palynostratigraphy andpalaeoenvironment of the Middle Jurassic Sortehat Formation(Neill Klinter Group), Jameson Land, East Greenland. In:Ineson, J.R. & Surlyk, F. (eds): The Jurassic of Denmark andGreenland. Geological Survey of Denmark and GreenlandBulletin 1, 777–811 (this volume).
Koppelhus, E.B. & Nielsen, L.H. 1994: Palynostratigraphy andpalaeoenvironments of the Lower to Middle Jurassic BagåFormation of Bornholm, Denmark. Palynology 18, 139–194.
Larsen, H.C. & Marcussen, C. 1992: Sill-intrusion, flood basaltemplacement and deep crustal structure of the Scoresby Sundregion, East Greenland. In: Storey, B.C., Alabaster, T. &Pankhurst, R.J. (eds): Magmatism and the causes of conti-nental break-up. Geological Society Special Publication
757
(London) 68, 365–386.Lund, J.J. & Pedersen, K.R. 1985: Palynology of the marine Jurassic
formations in the Vardekløft ravine, Jameson Land, EastGreenland. Bulletin of the Geological Society of Denmark 33,371–400.
Morgenroth, P. 1970: Dinoflagellate cysts from the Lias Delta ofLühnde/Germany. Neues Jahrbuch für Geologie und Palä-ontologie Abhandlungen 136, 345–359.
Nøhr-Hansen, H. 1993: Dinoflagellate cyst stratigraphy of theBarremian to Albian, Lower Cretaceous, North-East Greenland.Bulletin Grønlands Geologiske Undersøgelse 166, 171 pp.
Poulsen, N.E. 1996: Dinoflagellate cysts from marine Jurassicdeposits of Denmark and Poland. American Association ofStratigraphic Palynologists Contributions Series 31, 227 pp.
Prauss, M. 1987: Nannoceratopsis triangulata n.sp. – eine neueDinozysten-Spezies aus dem Obertoarcium von NW-Deutsch-land. Neues Jahrbuch für Geologie und Paläontologie Abhand-lungen 176, 129–136.
Riding, J.B. & Thomas, J.E. 1992: Dinoflagellate cysts of theJurassic System. In: Powell, A.J. (ed.): A stratigraphic index ofdinoflagellate cysts, 7–97. British Micropalaeontological SocietyPublication Series. London: Chapman & Hall.
Riding, J.B., Walton, W. & Shaw, D. 1991: Toarcian to Bathonian(Jurassic) palynology of the Inner Hebrides, Northwest Scotland.Palynology 15, 115–179.
Rosenkrantz, A. 1929: Preliminary account of the geology of theScoresby Sound District. Meddelelser om Grønland 73(2),135–154.
Rosenkrantz, A. 1934: The Lower Jurassic rocks of East Greenland,Part I. Meddelelser om Grønland 110(1), 150 pp.
Seidenkrantz, M.-S., Koppelhus, E.B. & Ravn-Sørensen, H. 1993:Biostratigraphy and palaeoenvironmental analysis of a Lowerto Middle Jurassic succession on Anholt, Denmark. Journal ofMicropalaeontology 12, 201–218.
Smelror, M. & Below, R. 1992: Dinoflagellate biostratigraphy ofthe Toarcian to Lower Oxfordian (Jurassic) of the Barents Searegion. Norwegian Petroleum Society (NPF) Special Publication2, 495–513.
Surlyk, F. 1977a: Mesozoic faulting in East Greenland. In: Frost,R.T.C. & Dikkers, A.J. (eds): Fault tectonics in NW Europe.Geologie en Mijnbouw 56, 311–327.
Surlyk, F. 1977b: Stratigraphy, tectonics and palaeogeography ofthe Jurassic sediments of the areas north of Kong Oscars Fjord,East Greenland. Bulletin Grønlands Geologiske Undersøgelse123, 56 pp.
Surlyk, F. 1978: Jurassic basin evolution of East Greenland. Nature274, 130–133.
Surlyk, F. 1990a: Timing, style and sedimentary evolution of LatePalaeozoic – Mesozoic extensional basins of East Greenland.In: Hardman, R.F.P. & Brooks, J. (eds): Tectonic events respon-
sible for Britain’s oil and gas reserves. Geological SocietySpecial Publications (London) 55, 107–125.
Surlyk, F. 1990b: A Jurassic sea-level curve for East Greenland.Palaeogeography, Palaeoclimatology, Palaeoecology 78, 71–85.
Surlyk, F. 1991: Sequence stratigraphy of the Jurassic – lowermostCretaceous of East Greenland. American Association ofPetroleum Geologists Bulletin 75, 1468–1488.
Surlyk, F. 2003: The Jurassic of East Greenland: a sedimentaryrecord of thermal subsidence, onset and culmination of rift-ing. In: Ineson, J.R. & Surlyk, F. (eds): The Jurassic of Denmarkand Greenland. Geological Survey of Denmark and GreenlandBulletin 1, 659–722 (this volume).
Surlyk, F., Callomon, J.H., Bromley, R.G. & Birkelund, T. 1973:Stratigraphy of the Jurassic – Lower Cretaceous sediments ofJameson Land and Scoresby Land, East Greenland. BulletinGrønlands Geologiske Undersøgelse 105, 76 pp. (also Med-delelser om Grønland 193(5)).
Surlyk, F., Clemmensen, L.B. & Larsen, H.C. 1981: Post-Paleozoicevolution of the East Greenland continental margin. In: Kerr,J.W., Fergusson, A.J. & Machan, L.C. (eds): Geology of the NorthAtlantic borderlands. Canadian Society of Petroleum GeologistsMemoir 7, 611–645.
Surlyk, F., Piasecki, S., Rolle, F., Thomsen, E. & Wrang, P. 1984:The Permian basin of East Greenland. In: Spencer, A. et al.(eds): Petroleum Geology of the North European Margin,303–315. London: Graham & Trotman for the NorwegianPetroleum Society (NPF).
Surlyk, F., Hurst, J.M., Piasecki, S., Rolle, F., Scholle, P.A., Stemmerik,L. & Thomsen, E. 1986: The Permian of the western marginof the Greenland Sea – a future exploration target. In: Halbouty,M.T. (ed.): Future petroleum provinces of the world. AmericanAssociation of Petroleum Geologists Memoir 40, 629–659.
Sykes, R.M. 1974: Sedimentological studies in southern JamesonLand, East Greenland. II. Offshore–estuarine regressivesequences in the Neill Klinter Formation (Pliensbachian–Toarcian). Bulletin of the Geological Society of Denmark 23,213–224.
Underhill, J.R. & Partington, M.A. 1994: Use of genetic sequencestratigraphy in defining and determining a regional tectoniccontrol on the ‘Mid-Cimmerian Unconformity’ – implicationsfor North Sea basin development and the global sea-levelchart. In: Weimer, P. & Posamentier, H.W. (eds): Siliciclasticsequence stratigraphy. Recent developments and applications.American Association of Petroleum Geologists Memoir 58,449–484.
Wall, D. 1965: Microplankton, pollen, and spores from the LowerJurassic in Britain. Micropaleontology 11, 151–190.
Ziegler, P.A. 1988: Evolution of the Arctic – North Atlantic andthe western Tethys. American Association of PetroleumGeologists Memoir 43, 198 pp.
758
Manuscript received 21 February 1997; revision accepted 1 June 2000.
Palynomorphs from the Neill Klinter Group at the Albuen section. The scale bar is 10 microns. For each of theillustrated specimens (Plates 1–7), the England Finder Reference (EFR) is given.
Fig. 4A. Terrestrial palynomorph distribution chart for the Gule Horn and Ostreaelv Formations at Albuen (for location, see Fig. 1). M, Middle Jurassic; A, Aalenian; As, Astartekløft Member.
Fig. 4B. Marine palynomorph distribution chart for the Gule Horn and Ostreaelv Formations at Albuen (for location, see Fig. 1). M, Middle Jurassic; A, Aalenian; As, Astartekløft Member.