Structural organization of the ‘‘zipper line’’ in Drosophila species with giant spermatozoa Romano Dallai * , David Mercati, Fabiola Giusti University of Siena, Department of Evolutionary Biology, Via A. Moro, 2 I-53100 Siena, Italy Received 15 June 2007; received in revised form 5 September 2007; accepted 7 September 2007 Available online 14 September 2007 Abstract The ‘‘zipper line’’ of Drosophila melanogaster and of Drosophila species characterized by giant spermatozoa (D. hydei, D. kanekoi and D. bifurca) was studied by electron microscopy using conventional thin-sections, lectin labeling and freeze-fracture replicas. In cross sec- tions the membrane specializations are located either at the level of the short cistern close to the large mitochondrial derivative where a small tuft of glycocalyx is visible or, in species characterized by long spermatozoa, along a cistern beneath the plasma membrane. In correspondence of such cistern, the plasma membrane exhibits a thick and extended glycocalyx. At this level, as well as at the short tuft of D. melanogaster, a-mannose residues were detected. The ‘‘zipper’’ of D. melanogaster consists of rows of intramembrane particles lon- gitudinally disposed along the sperm tail and associated with the external face of the plasma membrane. On the protoplasmatic face a narrow ribbon of transversal grooves is visible. Freeze-fracture replicas have revealed, in the region characterized by extended glycocalyx, the presence of a large ribbon of intramembrane particles disposed in parallel transversal rows, associated with the protoplasmatic mem- brane face. On the complementary external face a ribbon of parallel transversal grooves was observed. It is suggested that membrane specializations are mechanical devices to protect spermatozoa from torsion and bending in the seminal vesicles and then in the female storage organ. Ó 2007 Published by Elsevier Inc. Keywords: Drosophila sperm ultrastructure; Freeze-fracture replicas; Flagellar axoneme; Membrane specializations 1. Introduction The plasma membrane of insect sperm cells is very often specialized in different regions along the cell; these may rep- resent specific sites for gamete interaction at fertilization (Pasini et al., 1999; Perotti et al., 2001), or sites where sper- matozoa stick together in temporary aggregations of two or more (spermatodesms) to increase the number of male germ cells transferred to the female at mating (Jamieson et al., 1999). These membrane specializations might also have some mechanical role. Among the several membrane specializations described so far, the most common is the ‘‘zipper line’’, which appears in freeze-fracturing preparations as longitudinal double rows of staggered intramembrane particles (IMPs) along one of the axonemal doublets. The ‘‘zipper’’ was described in the principal piece of the mammalian sperm tail (Friend and Fawcett, 1974; Friend, 1984) where it may be involved in connecting the plasma membrane to underlying structures, thus preventing the passive fluttering of the membrane during sperm movement. A similar fea- ture was observed by Baccetti et al. (1971) in the medfly Ceratitis capitata, in a study on the fine structure of the insect plasma membrane. Dallai and Afzelius (1982) also found, by a freeze-fracture study of several heteropteran insect spermatozoa, a ‘‘zipper line’’ formed by a few rows of longitudinal intramembrane particles located, along the sperm tail, in one of the grooves between the axonemal complex and the mitochondrial derivative. The particles were seen mainly associated with the P-face of the plasma 1047-8477/$ - see front matter Ó 2007 Published by Elsevier Inc. doi:10.1016/j.jsb.2007.09.004 * Corresponding author. Fax: +39 0577 234476. E-mail address: [email protected](R. Dallai). www.elsevier.com/locate/yjsbi Available online at www.sciencedirect.com Journal of Structural Biology 161 (2008) 43–54 Journal of Structural Biology
Transcript
Available online at www.sciencedirect.comJournal of
www.elsevier.com/locate/yjsbi
Journal of Structural Biology 161 (2008) 43–54
StructuralBiology
Structural organization of the ‘‘zipper line’’ in Drosophila specieswith giant spermatozoa
Romano Dallai *, David Mercati, Fabiola Giusti
University of Siena, Department of Evolutionary Biology, Via A. Moro, 2 I-53100 Siena, Italy
Received 15 June 2007; received in revised form 5 September 2007; accepted 7 September 2007Available online 14 September 2007
Abstract
The ‘‘zipper line’’ of Drosophila melanogaster and of Drosophila species characterized by giant spermatozoa (D. hydei, D. kanekoi andD. bifurca) was studied by electron microscopy using conventional thin-sections, lectin labeling and freeze-fracture replicas. In cross sec-tions the membrane specializations are located either at the level of the short cistern close to the large mitochondrial derivative where asmall tuft of glycocalyx is visible or, in species characterized by long spermatozoa, along a cistern beneath the plasma membrane. Incorrespondence of such cistern, the plasma membrane exhibits a thick and extended glycocalyx. At this level, as well as at the short tuftof D. melanogaster, a-mannose residues were detected. The ‘‘zipper’’ of D. melanogaster consists of rows of intramembrane particles lon-gitudinally disposed along the sperm tail and associated with the external face of the plasma membrane. On the protoplasmatic face anarrow ribbon of transversal grooves is visible. Freeze-fracture replicas have revealed, in the region characterized by extended glycocalyx,the presence of a large ribbon of intramembrane particles disposed in parallel transversal rows, associated with the protoplasmatic mem-brane face. On the complementary external face a ribbon of parallel transversal grooves was observed. It is suggested that membranespecializations are mechanical devices to protect spermatozoa from torsion and bending in the seminal vesicles and then in the femalestorage organ.� 2007 Published by Elsevier Inc.
The plasma membrane of insect sperm cells is very oftenspecialized in different regions along the cell; these may rep-resent specific sites for gamete interaction at fertilization(Pasini et al., 1999; Perotti et al., 2001), or sites where sper-matozoa stick together in temporary aggregations of twoor more (spermatodesms) to increase the number of malegerm cells transferred to the female at mating (Jamiesonet al., 1999). These membrane specializations might alsohave some mechanical role.
Among the several membrane specializations describedso far, the most common is the ‘‘zipper line’’, which
1047-8477/$ - see front matter � 2007 Published by Elsevier Inc.
appears in freeze-fracturing preparations as longitudinaldouble rows of staggered intramembrane particles (IMPs)along one of the axonemal doublets. The ‘‘zipper’’ wasdescribed in the principal piece of the mammalian spermtail (Friend and Fawcett, 1974; Friend, 1984) where itmay be involved in connecting the plasma membrane tounderlying structures, thus preventing the passive flutteringof the membrane during sperm movement. A similar fea-ture was observed by Baccetti et al. (1971) in the medflyCeratitis capitata, in a study on the fine structure of theinsect plasma membrane. Dallai and Afzelius (1982) alsofound, by a freeze-fracture study of several heteropteraninsect spermatozoa, a ‘‘zipper line’’ formed by a few rowsof longitudinal intramembrane particles located, alongthe sperm tail, in one of the grooves between the axonemalcomplex and the mitochondrial derivative. The particleswere seen mainly associated with the P-face of the plasma
44 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
membrane. Bao and de Souza (1992) observed a similarstructure in freeze-fracture replicas of the mosquito spermplasma membrane.
The ‘‘zipper line’’ was recognized in thin sections byTokuyasu (1974) in a study on the dynamics of spermato-genesis of Drosophila melanogaster. He observed the‘‘appendicle stripe’’, a small tuft on the outside of thesperm cell membrane, located along the line of associationbetween the remnants of the periaxonemal sheath and theplasma membrane at the side of the major mitochondrialderivative. Similarly, on the outer leaflet of the plasmamembrane of the heteropteran spermatozoa, a few shortrodlets were seen to extend outside (Dallai and Afzelius,1982). Perotti and Riva (1988) and Perotti and Pasini(1995) detected, in D. melanogaster, specific glycoconju-gates at the ‘‘appendicle stripe’’ and suggested the presenceat this point of a-mannose residues.
In the present study, using correlated thin sections,freeze-fracture replicas and the lectin-binding technique,we investigate the possible function of the ‘‘zipper line’’in different Drosophila species characterized by longspermatozoa.
Animal species with internal fertilization have developedsperm cells whose length generally is greater than thosepracticing external fertilization (Baccetti and Afzelius,1976). Insects, however, exhibit an extreme variability ofthis character: from spermatozoa of only a few microme-ters such as those of some termites (1.5–1.7 lm) (Jamiesonet al., 1999), Protura (1–2 lm) (Dallai et al., 1990) or cad-disflies (4.5–10 lm) (Dallai and Afzelius, 1995) to those ofD. bifurca which measure many times the length of themale producing them.
Among the well-studied Drosophila, spermatozoa rangefrom 10 lm of D. azteca (Joly, 1987) to D. bifurca whichwith its 58.290 lm is the longest insect spermatozoa discov-ered so far (Joly et al., 1995; Pitnick et al., 1995b). The rea-son for the acquisition of long spermatozoa in this group ofspecies is still matter of study and various hypotheses havebeen offered to explain how selection pressure is responsi-ble for sperm evolution and for increasing sperm lengthin Drosophila (Pitnick et al., 1995b). The existence of giantsperm would seem to contradict the well known view of themale sexual strategy (Parker, 1982), as their productionrepresents a great cost energetically, reduces the numberof sperms and also delays male sexual maturation (Pitnicket al., 1995a). It has been suggested that long spermatozoacould be more efficient in terms of sperm velocity and deliv-ery of resources to the egg. Neither of these suggestions arevalid in the case of Drosophila, considering the short dis-tance sperm cells must travel in the female genital tractto reach eggs (Pitnick and Markow, 1994) and that theentire long spermatozoa does not enter into the egg (Pit-nick et al., 1995b).
It is worthy of mention that even though the length ofthe Drosophila sperm axoneme can vary, reaching up to6 cm in D. bifurca, its fine structure remains unchangedfrom the 9 + 9 + 2 axonemal pattern with accessory
tubules with 13 protofilaments and a compact intertubularmaterial with an electron-transparent row of dots along theouter edge. This could be explained bearing in mind thatthe axonemal pattern present in Brachycera had reachedthe maximum possible efficiency for sperm motility andthat it could not allow any further modifications in its axo-nemal components without losing its functionality (Dallaiet al., 2007). This claim is supported by the considerationthat a highly specialized b-tubulin is needed to constructthe Drosophila sperm tail axoneme (Nielsen and Raff,2002) and the brachyceran axoneme is a stable rather thana changeable feature (Nielsen et al., 2006).
We ask whether the increased sperm length and the con-sequent difficulty of the plasma membrane to maintain itsintegrity during sperm bending and sperm movement, iscounterbalanced by specializations which reinforce thesperm plasma membrane, particularly at the sides involvedin mechanical stress. The study also defines, in greaterdetail, these particular sperm tail membrane specializationsin D. melanogaster.
2. Materials and methods
2.1. Study animals
Populations of the following Drosophila species werereared at 25 �C on a standard medium:
• D. bifurca (sperm length 58.29 mm) (Pitnick et al.,1995b)
• D. kanekoi (sperm length 24.29 mm) (Pitnick et al.,1999)
• D. hydei (sperm length 23.32 mm) (Pitnick et al., 1999)• D. melanogaster (sperm length 1.91 mm) (Pitnick et al.,
1999)
Stocks of D. hydei came from the Tucson DrosophilaStock Center, USA; those of D. bifurca and D. kanekoi
were kindly donated by Prof. Scott Pitnick (Syracuse Uni-versity) and the D. melanogaster stock was kindly donatedby Prof. Giuliano Callaini (Siena University).
Several virgin males of each species were isolated fromthe rest of the population after being anesthetized withCO2. The flies were kept at the same temperature untilsexual maturity was reached, that is around 20 days forD. bifurca, 15 days for D. hydei and D. kanekoi and 7 daysfor D. melanogaster.
2.2. Interference contrast and fluorescence microscopy
Testis and seminal vesicles of D. bifurca and D. hydei
mature males were dissected in 0.1 M phosphate bufferedsaline (PBS) pH 7.2. Samples were fixed for 10 min inmethanol at �20 �C, washed in PBS and stained for5 min with 1 lg/ml of the DNA-specific dye Hoechst33258 in PBS. Samples were then washed in PBS andmounted in 90% glycerol containing 2.5% n-propylgallate.
R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54 45
Interference contrast and fluorescence observations weredone with a Leica DMRB light microscope equipped withUV light and UV filters.
2.3. Scanning electron microscopy
The isolated seminal vesicles were split open to adhereon a poly-L-lysine pre-treated glass coverslips fixed with2.5% glutaraldehyde in phosphate buffer (PB) 0.1 M pH7.2 for 1 h at 4 �C and post-fixed in 1% OsO4 in PB. Sam-ples were then dehydrated in ethanol, dried in a BalzersCPD 030 critical point dryer and coated with gold in a Bal-zers MED 010 sputtering device. Specimens were examinedin a Philips XL20 scanning electron microscope operatingat 20 kV.
2.4. Transmission electron microscopy
Testis, deferent ducts and seminal vesicles of males of D.
bifurca, D. kanekoi, D. hydei, and D. melanogaster were dis-sected in PB, containing 1.8% sucrose. Samples were fixedin 2.5% glutaraldehyde in PB to which 1% tannic acid wasadded (thus omitting the osmium fixation step) and blockstained with 1% uranyl acetate (Dallai and Afzelius,1990). The material was then washed in PB, dehydratedin ethanol, and embedded in Epon–Araldite mixture.Ultrathin sections obtained with Reichert Ultracut II Eultramicrotome were routinely stained and observed witha Philips CM 10 transmission electron microscope operat-ing at 80 kV. Semithin sections, of about 1 lm, werestained for 5 min with 1.5% toluidine blue and observedwith a Leica DMRB light microscope.
2.5. Freeze fracture
After isolation from adult males by dissection in PB,seminal vesicles were cryoprotected in 10%, 20%, and30% glycerol in PB for varying periods of time, then frozenby plunging into propane cooled in liquid nitrogen. Thematerial was then mounted in a Balzer’s BAF 301 freeze-fracturing device and fractured at �100 �C to �115 �C at2 · 10�6 Torr. The preparations were shadowed with plat-inum–carbon and backed with carbon. The tissue wasremoved with sodium hypochlorite and the replicas, rinsedin water, were mounted on copper grids for examinationwith a Philips CM 10 TEM at 80 kV.
2.6. Lectin labeling
Free spermatozoa of each species were dissected in PBSpH 7.4. Unfixed sperm cells were incubated for 1 h at roomtemperature in a solution of 10–20 lg/ml 10 nm gold-con-jugated lectins: Concanavalia ensiformis (Con A) andGalanthus nivalis (GNA) agglutinins (EY LaboratoriesInc.) were used for our experiments. The samples wererinsed several times in PBS then fixed for 2 h at 4 �C in2.5% glutaraldehyde in PBS. The material was then rinsed
in buffer and post-fixed in 1% OsO4 in PBS for 1 h at 4 �C.After dehydration in a graded series of ethanol, the mate-rial was embedded in an Epon–Araldite mixture, sectioned,routinely stained and observed with a Philips CM 10 trans-mission electron microscope at 80 kV.
Controls with the labeled lectins, each one in presence of0.3 M a-D-mannose, were performed.
2.7. Sperm unwinding
Observations were done on deferent ducts and seminalvesicles of D. hydei and D. bifurca opened in PB, using aLeica DMLB light microscope equipped with phase con-trast optics and recorded by a Sony DXC-930P 3 CCDcamera and a digital video recorder Sony DVCAM DSR-30P. Video images were captured using the Pinnacle Studio8.8 software (Pinnacle System, GmbH, Braunschweig, Ger-many) and edited using Adobe Photoshop.
3. Results
3.1. Drosophila melanogaster
The species has sperm 1.91 mm long (Pitnick et al.,1999). The sperm tail ultrastructure has been described ingreat detail in several papers dealing with axonemal orga-nization (Perotti, 1969; Stanley et al., 1972; Tokuyasu,1974; Dallai and Afzelius, 1990). The sperm tail consistsof 9 + 9 + 2 axoneme; the accessory tubules have 13 proto-filaments separated by intertubular material which shows,on the external side, few electron-transparent dots in lineararray (Fig. 1a). On the opposite side of the axoneme twomitochondrial derivatives of different size are present. Theyhave a central portion of crystallized material. The majormitochondrial derivative is found on the right side if theaxoneme is clockwise-oriented. The sperm plasma mem-brane, which shows the ‘‘fruit-fly model’’ (Baccetti et al.,1971), exhibits on the outer side facing the extremity ofthe residual periaxonemal sheath, a tuft of material whichprojects from the thin glycocalyx (Fig. 1a). This structureis the ‘‘appendicle stripe’’ described by Tokuyasu (1974).It is found at the right side of the plasma membrane, notfar from the major mitochondrial derivative, when the axo-neme is clockwise-oriented, thus with the observer lookingfrom the centriolar region.
Perotti and Riva (1988) previously reported that the‘‘appendicle’’ contains membrane-bound carbohydrates.We have confirmed these using lectins conjugated with col-loidal gold particles. In correspondence with the appendiclethe specific lectins used, Con A (Fig. 1b) and GNA(Fig. 1c), gave a positive result and gold particles wereassociated with the tuft of glycocalyx. We thus infer thepresence of a-mannose residues at this position.
Replicas of mature spermatozoa taken from seminalvesicles and prepared with the freeze-fracture techniqueexhibit a membrane specialization at the side of the plasmamembrane corresponding to the major mitochondrial
Fig. 1. Drosophila melanogaster. (a) Cross section through the sperm tail. The axoneme consists of a 9 + 2 which is surrounded by nine accessory microtubuleseach with 13 protofilaments in the tubular wall. Conspicuous intertubular material showing linear electron transparent dots on the outer profile is present(arrows). There are two partially crystallized mitochondrial derivatives (md) of different size. A little dense material beside the large mitochondrial derivative isvisible (asterisk); this is the remnant of the periaxonemal cistern present in the spermatid. On the outer leaflet of the plasma membrane a short thick glycocalyxis evident (arrowhead). Scale bar 0.1 lm. (b and c) Micrographs after treatment with ConA (b) and GNA (c) lectins conjugated with 10 nm colloidal gold.There are polysaccharide residues at the thick glycocalyx on the plasma membrane (arrowheads). Scale bars 0.1 lm. (d–f) Freeze-fracture replicas of theplasma membrane of the sperm tail. On the P-membrane face (PF) a longitudinal ribbon of grooves in parallel transversal rows is visible (arrows); on one sideof the ribbon a longitudinal row of small IMPs is present (arrowheads). On the complementary E-membrane face (EF) a continuous longitudinal row of IMPs(white arrowheads) and a parallel row of discontinuous IMPs (white arrows) are also visible. ax, axoneme; md, mitochondrial derivative. Scale bars 0.5 lm.
46 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54 47
derivative. On the protoplasmatic membrane face (P-face)a narrow ribbon of short grooves aligned in parallel trans-versal rows are visible (Fig. 1d and e). The ribbon runsalong the longitudinal axis of the sperm tail. On one sideof the ribbon a row of 8 nm IMPs with particles often fusedin short rods is evident. The complementary external mem-brane face (E-face) is decorated by a longitudinal row ofIMPs, 10-nm thick, often flanked by a parallel discontinu-ous row (Fig. 1d–f).
3.2. Drosophila hydei
This species is characterized by long spermatozoa(23.32 mm) (Pitnick et al., 1999) with sperm nuclei of about
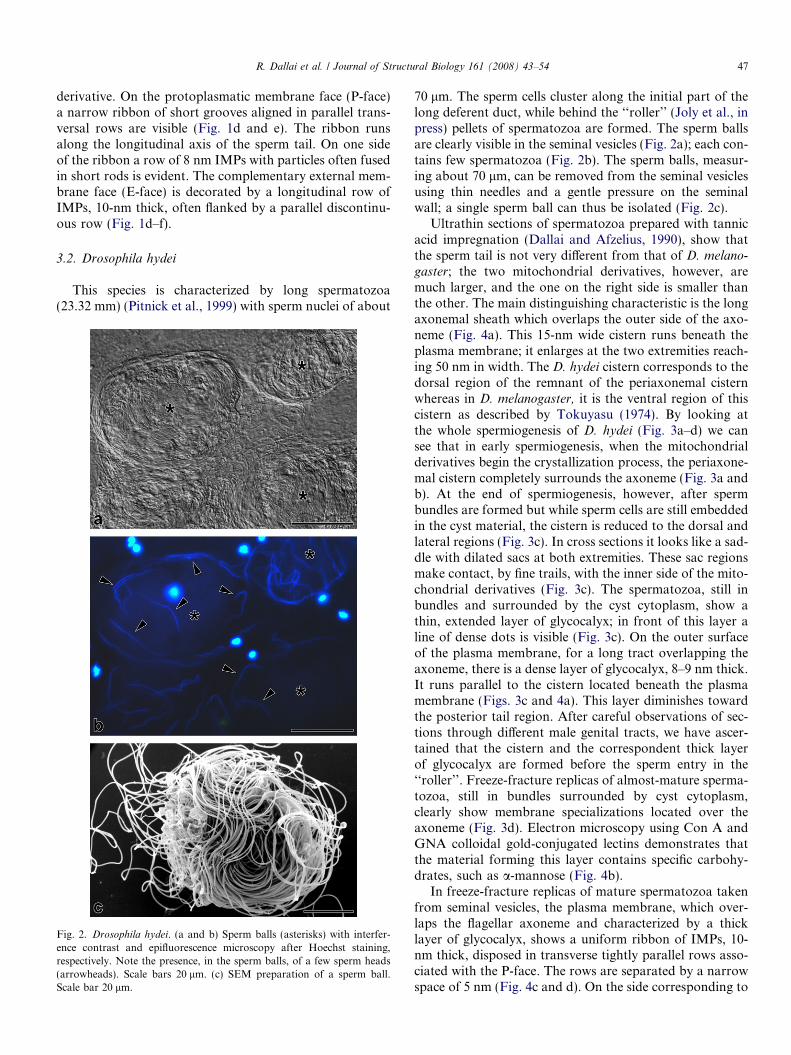
Fig. 2. Drosophila hydei. (a and b) Sperm balls (asterisks) with interfer-ence contrast and epifluorescence microscopy after Hoechst staining,respectively. Note the presence, in the sperm balls, of a few sperm heads(arrowheads). Scale bars 20 lm. (c) SEM preparation of a sperm ball.Scale bar 20 lm.
70 lm. The sperm cells cluster along the initial part of thelong deferent duct, while behind the ‘‘roller’’ (Joly et al., inpress) pellets of spermatozoa are formed. The sperm ballsare clearly visible in the seminal vesicles (Fig. 2a); each con-tains few spermatozoa (Fig. 2b). The sperm balls, measur-ing about 70 lm, can be removed from the seminal vesiclesusing thin needles and a gentle pressure on the seminalwall; a single sperm ball can thus be isolated (Fig. 2c).
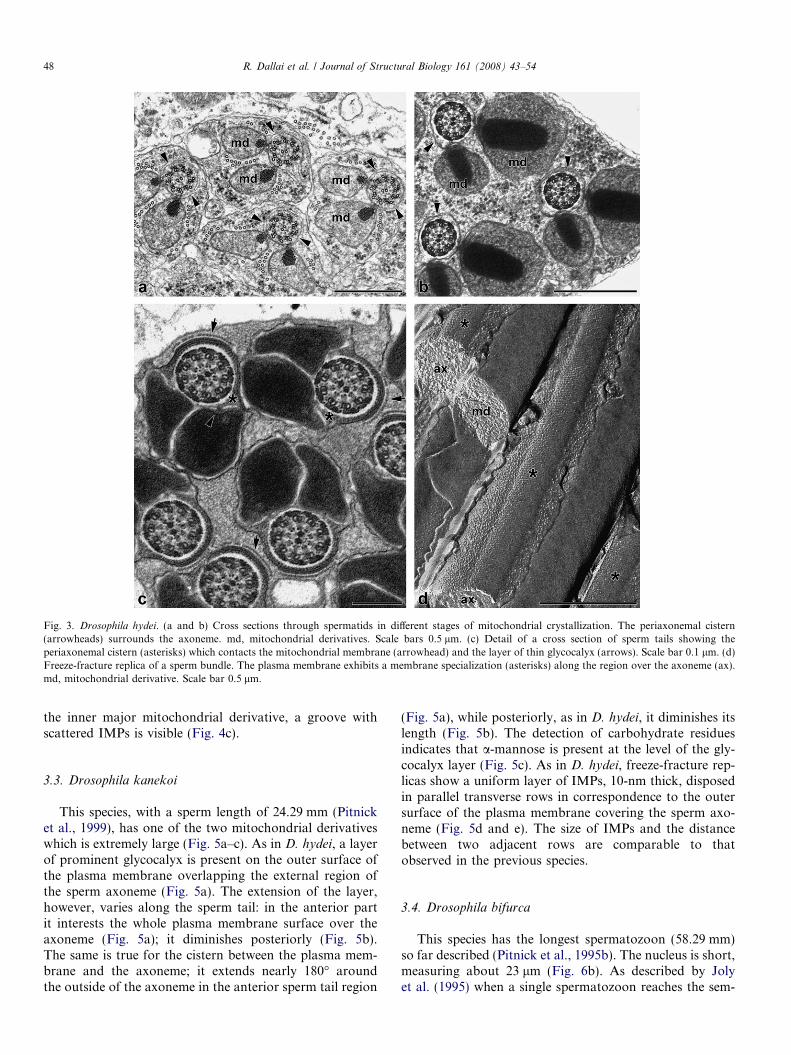
Ultrathin sections of spermatozoa prepared with tannicacid impregnation (Dallai and Afzelius, 1990), show thatthe sperm tail is not very different from that of D. melano-
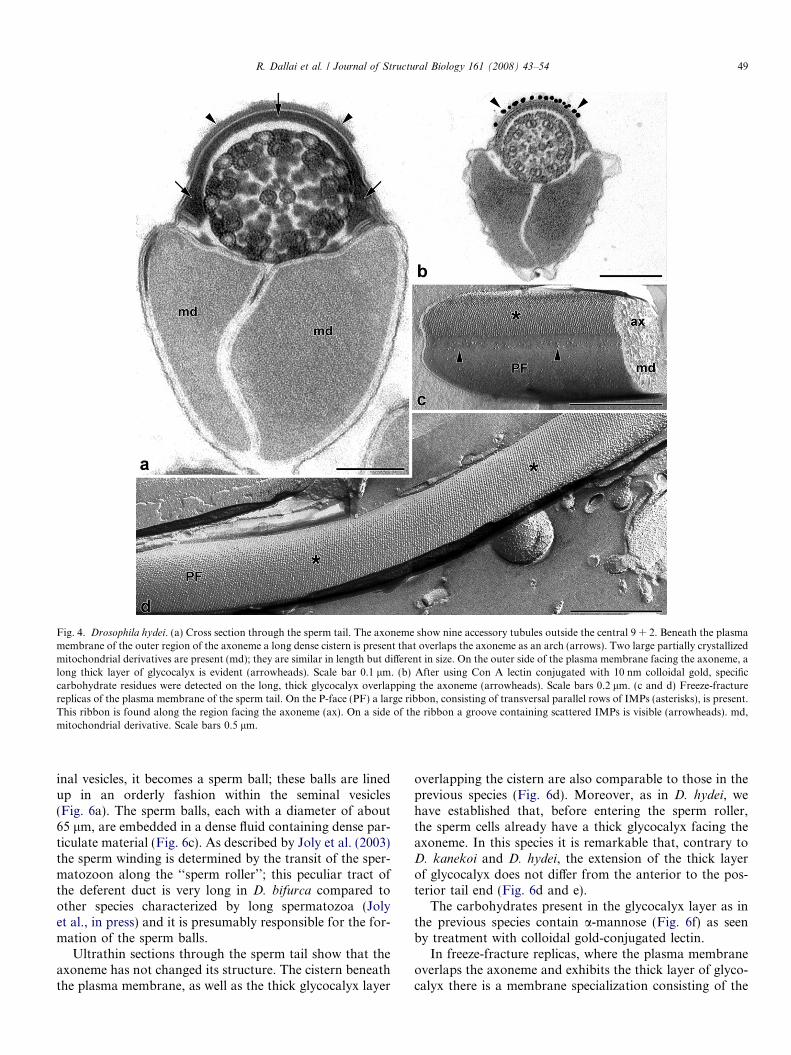
gaster; the two mitochondrial derivatives, however, aremuch larger, and the one on the right side is smaller thanthe other. The main distinguishing characteristic is the longaxonemal sheath which overlaps the outer side of the axo-neme (Fig. 4a). This 15-nm wide cistern runs beneath theplasma membrane; it enlarges at the two extremities reach-ing 50 nm in width. The D. hydei cistern corresponds to thedorsal region of the remnant of the periaxonemal cisternwhereas in D. melanogaster, it is the ventral region of thiscistern as described by Tokuyasu (1974). By looking atthe whole spermiogenesis of D. hydei (Fig. 3a–d) we cansee that in early spermiogenesis, when the mitochondrialderivatives begin the crystallization process, the periaxone-mal cistern completely surrounds the axoneme (Fig. 3a andb). At the end of spermiogenesis, however, after spermbundles are formed but while sperm cells are still embeddedin the cyst material, the cistern is reduced to the dorsal andlateral regions (Fig. 3c). In cross sections it looks like a sad-dle with dilated sacs at both extremities. These sac regionsmake contact, by fine trails, with the inner side of the mito-chondrial derivatives (Fig. 3c). The spermatozoa, still inbundles and surrounded by the cyst cytoplasm, show athin, extended layer of glycocalyx; in front of this layer aline of dense dots is visible (Fig. 3c). On the outer surfaceof the plasma membrane, for a long tract overlapping theaxoneme, there is a dense layer of glycocalyx, 8–9 nm thick.It runs parallel to the cistern located beneath the plasmamembrane (Figs. 3c and 4a). This layer diminishes towardthe posterior tail region. After careful observations of sec-tions through different male genital tracts, we have ascer-tained that the cistern and the correspondent thick layerof glycocalyx are formed before the sperm entry in the‘‘roller’’. Freeze-fracture replicas of almost-mature sperma-tozoa, still in bundles surrounded by cyst cytoplasm,clearly show membrane specializations located over theaxoneme (Fig. 3d). Electron microscopy using Con A andGNA colloidal gold-conjugated lectins demonstrates thatthe material forming this layer contains specific carbohy-drates, such as a-mannose (Fig. 4b).
In freeze-fracture replicas of mature spermatozoa takenfrom seminal vesicles, the plasma membrane, which over-laps the flagellar axoneme and characterized by a thicklayer of glycocalyx, shows a uniform ribbon of IMPs, 10-nm thick, disposed in transverse tightly parallel rows asso-ciated with the P-face. The rows are separated by a narrowspace of 5 nm (Fig. 4c and d). On the side corresponding to
Fig. 3. Drosophila hydei. (a and b) Cross sections through spermatids in different stages of mitochondrial crystallization. The periaxonemal cistern(arrowheads) surrounds the axoneme. md, mitochondrial derivatives. Scale bars 0.5 lm. (c) Detail of a cross section of sperm tails showing theperiaxonemal cistern (asterisks) which contacts the mitochondrial membrane (arrowhead) and the layer of thin glycocalyx (arrows). Scale bar 0.1 lm. (d)Freeze-fracture replica of a sperm bundle. The plasma membrane exhibits a membrane specialization (asterisks) along the region over the axoneme (ax).md, mitochondrial derivative. Scale bar 0.5 lm.
48 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
the inner major mitochondrial derivative, a groove withscattered IMPs is visible (Fig. 4c).
3.3. Drosophila kanekoi
This species, with a sperm length of 24.29 mm (Pitnicket al., 1999), has one of the two mitochondrial derivativeswhich is extremely large (Fig. 5a–c). As in D. hydei, a layerof prominent glycocalyx is present on the outer surface ofthe plasma membrane overlapping the external region ofthe sperm axoneme (Fig. 5a). The extension of the layer,however, varies along the sperm tail: in the anterior partit interests the whole plasma membrane surface over theaxoneme (Fig. 5a); it diminishes posteriorly (Fig. 5b).The same is true for the cistern between the plasma mem-brane and the axoneme; it extends nearly 180� aroundthe outside of the axoneme in the anterior sperm tail region
(Fig. 5a), while posteriorly, as in D. hydei, it diminishes itslength (Fig. 5b). The detection of carbohydrate residuesindicates that a-mannose is present at the level of the gly-cocalyx layer (Fig. 5c). As in D. hydei, freeze-fracture rep-licas show a uniform layer of IMPs, 10-nm thick, disposedin parallel transverse rows in correspondence to the outersurface of the plasma membrane covering the sperm axo-neme (Fig. 5d and e). The size of IMPs and the distancebetween two adjacent rows are comparable to thatobserved in the previous species.
3.4. Drosophila bifurca
This species has the longest spermatozoon (58.29 mm)so far described (Pitnick et al., 1995b). The nucleus is short,measuring about 23 lm (Fig. 6b). As described by Jolyet al. (1995) when a single spermatozoon reaches the sem-
Fig. 4. Drosophila hydei. (a) Cross section through the sperm tail. The axoneme show nine accessory tubules outside the central 9 + 2. Beneath the plasmamembrane of the outer region of the axoneme a long dense cistern is present that overlaps the axoneme as an arch (arrows). Two large partially crystallizedmitochondrial derivatives are present (md); they are similar in length but different in size. On the outer side of the plasma membrane facing the axoneme, along thick layer of glycocalyx is evident (arrowheads). Scale bar 0.1 lm. (b) After using Con A lectin conjugated with 10 nm colloidal gold, specificcarbohydrate residues were detected on the long, thick glycocalyx overlapping the axoneme (arrowheads). Scale bars 0.2 lm. (c and d) Freeze-fracturereplicas of the plasma membrane of the sperm tail. On the P-face (PF) a large ribbon, consisting of transversal parallel rows of IMPs (asterisks), is present.This ribbon is found along the region facing the axoneme (ax). On a side of the ribbon a groove containing scattered IMPs is visible (arrowheads). md,mitochondrial derivative. Scale bars 0.5 lm.
R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54 49
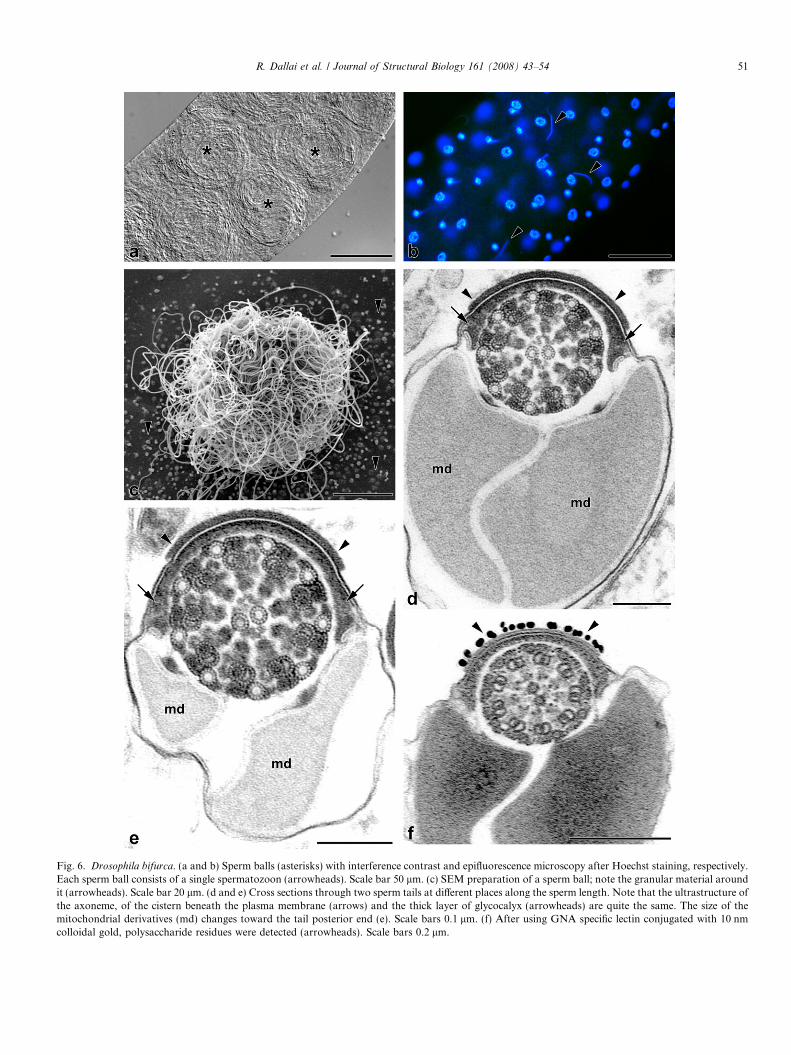
inal vesicles, it becomes a sperm ball; these balls are linedup in an orderly fashion within the seminal vesicles(Fig. 6a). The sperm balls, each with a diameter of about65 lm, are embedded in a dense fluid containing dense par-ticulate material (Fig. 6c). As described by Joly et al. (2003)the sperm winding is determined by the transit of the sper-matozoon along the ‘‘sperm roller’’; this peculiar tract ofthe deferent duct is very long in D. bifurca compared toother species characterized by long spermatozoa (Jolyet al., in press) and it is presumably responsible for the for-mation of the sperm balls.
Ultrathin sections through the sperm tail show that theaxoneme has not changed its structure. The cistern beneaththe plasma membrane, as well as the thick glycocalyx layer
overlapping the cistern are also comparable to those in theprevious species (Fig. 6d). Moreover, as in D. hydei, wehave established that, before entering the sperm roller,the sperm cells already have a thick glycocalyx facing theaxoneme. In this species it is remarkable that, contrary toD. kanekoi and D. hydei, the extension of the thick layerof glycocalyx does not differ from the anterior to the pos-terior tail end (Fig. 6d and e).
The carbohydrates present in the glycocalyx layer as inthe previous species contain a-mannose (Fig. 6f) as seenby treatment with colloidal gold-conjugated lectin.
In freeze-fracture replicas, where the plasma membraneoverlaps the axoneme and exhibits the thick layer of glyco-calyx there is a membrane specialization consisting of the
Fig. 5. Drosophila kanekoi. (a) Cross section through the anterior region of the sperm tail. The axoneme is protected by a long dense cistern (arrow) andon the corresponding plasma membrane a thick layer of glycocalyx is visible (arrowheads). Note also the great different size of the two mitochondrialderivatives (md). Scale bar 0.2 lm. (b) Cross section through the posterior region of the sperm tail. At this level the dense cistern (arrow) is reduced and thecorresponding thick layer of glycocalyx is less extended (arrowheads). md, mitochondrial derivatives. Scale bar 0.1 lm. (c) After using Con A lectinconjugated with 10 nm colloidal gold, specific polysaccharide residues at the level of the thick glycocalyx were detected (arrowheads). Scale bars 0.5 lm. (dand e) Freeze-fracture replicas of sperm tails showing membrane specialization at the region of plasma membrane overlapping the axoneme (ax). On the P-membrane face (PF) a large ribbon of IMPs disposed in parallel rows is visible (asterisks). md, mitochondrial derivative. Scale bars 0.2 lm.
50 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
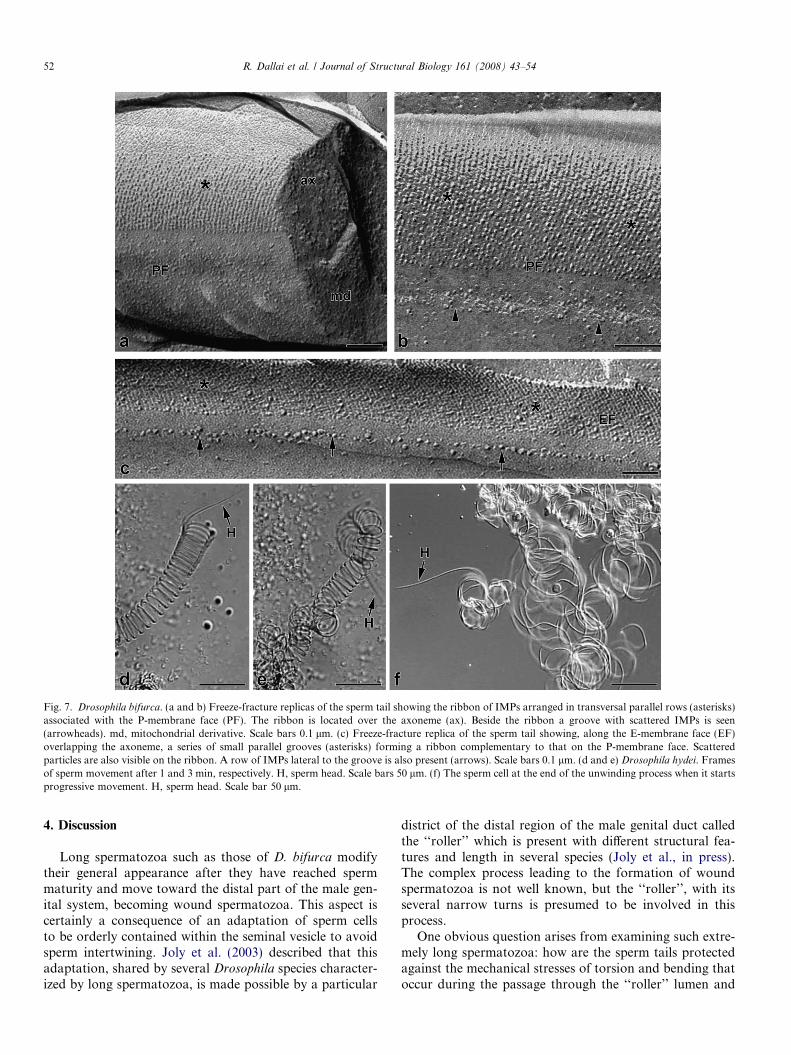
similar array of IMPs described in the previous species.These IMPs of about 10 nm are associated with the P-faceand are arranged in parallel rows (Fig. 7a and b). On oneside of the ribbon a narrow groove with few small intra-membrane particles is visible (Fig. 7b). The complementaryE-face shows a series of parallel thin grooves with scatteredparticles forming a ribbon. A longitudinal discontinuousrow of particles, 10-nm thick, is also visible on one sideof this ribbon (Fig. 7c).
3.5. Sperm unwinding
When the seminal vesicles of D. hydei are opened in PB,the sperm balls exit into the medium. At the beginning, the
sperm slowly unwinds, giving rise to a long spring formedby a tight series of spiral rings, of about 27 lm in diameter,which rotate around the longitudinal axis of the spring(Fig. 7d and e). Progressively, the spring elongates, the dis-tance between adjacent rings increases, and it begins to loseits perfect helical appearance; when the whole spermato-zoon has disentangled from the mass of spermatozoaaround it, the free sperm beats with increasing frequencyand the long sperm tail moves forward following a veryirregular path characterized by bends of large amplitude(Fig. 7f). It is of interest to mention that the diameter ofabout 27 lm of the spiral rings has also been determinedin the other species with long spermatozoa examined in thispaper.
Fig. 6. Drosophila bifurca. (a and b) Sperm balls (asterisks) with interference contrast and epifluorescence microscopy after Hoechst staining, respectively.Each sperm ball consists of a single spermatozoon (arrowheads). Scale bar 50 lm. (c) SEM preparation of a sperm ball; note the granular material aroundit (arrowheads). Scale bar 20 lm. (d and e) Cross sections through two sperm tails at different places along the sperm length. Note that the ultrastructure ofthe axoneme, of the cistern beneath the plasma membrane (arrows) and the thick layer of glycocalyx (arrowheads) are quite the same. The size of themitochondrial derivatives (md) changes toward the tail posterior end (e). Scale bars 0.1 lm. (f) After using GNA specific lectin conjugated with 10 nmcolloidal gold, polysaccharide residues were detected (arrowheads). Scale bars 0.2 lm.
R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54 51
Fig. 7. Drosophila bifurca. (a and b) Freeze-fracture replicas of the sperm tail showing the ribbon of IMPs arranged in transversal parallel rows (asterisks)associated with the P-membrane face (PF). The ribbon is located over the axoneme (ax). Beside the ribbon a groove with scattered IMPs is seen(arrowheads). md, mitochondrial derivative. Scale bars 0.1 lm. (c) Freeze-fracture replica of the sperm tail showing, along the E-membrane face (EF)overlapping the axoneme, a series of small parallel grooves (asterisks) forming a ribbon complementary to that on the P-membrane face. Scatteredparticles are also visible on the ribbon. A row of IMPs lateral to the groove is also present (arrows). Scale bars 0.1 lm. (d and e) Drosophila hydei. Framesof sperm movement after 1 and 3 min, respectively. H, sperm head. Scale bars 50 lm. (f) The sperm cell at the end of the unwinding process when it startsprogressive movement. H, sperm head. Scale bar 50 lm.
52 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
4. Discussion
Long spermatozoa such as those of D. bifurca modifytheir general appearance after they have reached spermmaturity and move toward the distal part of the male gen-ital system, becoming wound spermatozoa. This aspect iscertainly a consequence of an adaptation of sperm cellsto be orderly contained within the seminal vesicle to avoidsperm intertwining. Joly et al. (2003) described that thisadaptation, shared by several Drosophila species character-ized by long spermatozoa, is made possible by a particular
district of the distal region of the male genital duct calledthe ‘‘roller’’ which is present with different structural fea-tures and length in several species (Joly et al., in press).The complex process leading to the formation of woundspermatozoa is not well known, but the ‘‘roller’’, with itsseveral narrow turns is presumed to be involved in thisprocess.
One obvious question arises from examining such extre-mely long spermatozoa: how are the sperm tails protectedagainst the mechanical stresses of torsion and bending thatoccur during the passage through the ‘‘roller’’ lumen and
R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54 53
later during flagellar movement? Neesen et al. (1999) sug-gested that flagellar proteins stabilize the sperm tails. Topreserve the sperm tail integrity the plasma membranemust be reinforced in those points where it is liable to bedamaged. These points likely correspond to the beatingplane as defined by Afzelius (1959) and by Folliot andMaillet (1970), represented by the plane orthogonal to thatpassing through the two central tubules. According toTokuyasu (1974), however, the axoneme has no firm con-nection with the axonemal sheath or the cell membrane;this leads to an unstable position of the beating plane. Acertain rotational freedom has been suggested in relationto the rest of the tail even though the variations of the axo-nemal position are very restricted (Tokuyasu, 1974).
In the present study, we have demonstrated that there isa great difference between the sperm tail of D. melanogas-
ter, and those of the three species characterized by long(D. hydei and D. kanekoi) and extremely long (D. bifurca)spermatozoa. The latter species have a long cistern of densematerial beneath the plasma membrane at the region over-lapping the flagellar axoneme. At the same level, on theouter surface of the plasma membrane, a prominent andextended glycocalyx is evident. Instead, D. melanogaster
has only a short cistern and a simple tuft on the outer sur-face of the plasma membrane. The cisterns also derive fromdifferent remnants of the periaxonemal cisterns present inthe spermatids: in D. melanogaster it represents the ventralregion of the periaxonemal cistern, whilst in the specieswith long spermatozoa, it is the dorsal part of the periax-onemal cistern. The extended glycocalyx, however, seemsto be analogous to the small tuft (the ‘‘appendicle’’)described by Tokuyasu (1974) in D. melanogaster. At thelevel of this structure specific glycoproteins were detectedby Perotti and Riva (1988) using lectin conjugated with fer-ritin. We have confirmed, in all four species, the presence ofglycoproteins that could be involved in the reinforcementof the plasma membrane at this point. In particularly, aswe have obtained an identical labeling pattern from boththe lectins used, we can assume the presence at this levelof a-mannose terminal residues. Our findings are consistentwith recent studies indicating that the majority of the Dro-
sophila N-glycans contain a-mannose-terminated oligosac-charides. The intramembrane particles evidenced in freeze-fracture replicas, which give rise to membrane specializa-tions, are likely transmembrane integral proteins that couldrepresent specific points for the anchorage of the glyco-proteic chains forming the glycocalyx. Alternatively, itcould be possible that intramembrane particles are the pro-teic component of the glycocalyx.
We have also demonstrated by freeze-fracture replicasthat at the level of the ‘‘appendicle’’ of D. melanogaster
two different features are recognizable: longitudinal rowsof staggered IMPs, formerly described by Baccetti et al.(1971), which are associated with the E-face of the plasmamembrane and a complementary narrow longitudinal rib-bon of smaller grooves on the P-face of the membrane witha simple row of small IMPs nearby. Thus, the ‘‘zipper line’’
membrane specialization is more complex than was origi-nally described. Also of interest are the different fracturingresults obtained for the membrane specializations in thefour species. While in D. melanogaster the rows of IMPsare associated with the E-face of the membrane, in the spe-cies with long spermatozoa the IMPs are associated withthe P-face. This could be due to the different size and posi-tion of the proteins within the membrane to which the gly-cocalyx is bound, or to the different location of themembrane specializations which, in the species with longthick glycocalyx, being over the axoneme could be moreeasily fractured.
The membrane specializations and the thick glycocalyxare formed when sperm cells are still clustered in spermcyst. The presence of a simple tuft in D. melanogaster
would indicate that the structure is a condition both forsperm bending in the seminal vesicles where they formsperm bundles (Tokuyasu, 1974) as well as for sperm motil-ity in the small female seminal receptacle. The extendedsperm membrane specializations observed in the region fac-ing the axoneme of species characterized by long spermato-zoa are presumed to be mechanical devices to protect thespermatozoa from torsions and bending during the passagealong the roller where they become wound. In this contestit is of interest to mention that in the sperm ball the singlesperm forms several coils; a section through the sperm ballshows that the prominent glycocalyx of a coil, might inter-act with the plasma membrane of the overlapping spermcoil at the surface opposite to the glycocalyx. Thus, the gly-coconjugates might interact with complementary moleculesof the sperm plasma membrane itself, locking the tail intothe ball. Finally, the membrane specialization may alsohelp the sperm to be stored within the female seminalreceptacle (Joly, personal communication) which is verylong in Drosophila bifurca (Pitnick et al., 1999).
Finally, we would like to consider the series of ringswhich form the sperm ‘‘spring’’ and the process of spermunwinding. When the spermatozoa are released in the med-ium they first relax the spring so that the distance betweenthe rings increases; at the end of the process the spermato-zoa have straightened and begin to move forward. Wehypothesize that this particular sequence of events is theconsequence of the sperm passage through the sperm rolleras well as of the presence of the thick glycocalyx over theflagellar axoneme. It is likely that the winding which occursas the sperm pass through the sperm roller stores mechan-ical energy which is then released when the spermatozoabegin unrolling. Thus, the membrane specializations aresomewhat involved in establishing the type of sperm motil-ity as well as in preserving the membrane from flutteringduring flagellar beating. It would be of interest to verifythis hypothesis with future research.
Acknowledgments
We thank Prof. S. Pitnick (University of Syracuse, USA)and Prof. G. Callaini (University of Siena, Italy) for pro-
54 R. Dallai et al. / Journal of Structural Biology 161 (2008) 43–54
viding some of the Drosophila species studied in this paper.We want to thank Prof. L. Robbins (University of Siena,Italy) for helping us in rearing the animals. We are also in-debted to Dr. D. Joly (CNRS Gif sur Yvette Cedex,France) for the critical reading of the manuscript. Grantfrom MUR to R.D. is also gratefully acknowledged. Wewish to thank an anonymous reviewer for his useful com-ments which have improved the paper.
References
Afzelius, B.A, 1959. Electron microscopy of the sperm tail. J. Biophys.Biochem. Cytol. 5, 269–278.
Baccetti, B., Afzelius, B.A., 1976. The Biology of the Sperm Cell. KargerPress, Basel, 254 p.
Baccetti, B., Bigliardi, E., Rosati, F., 1971. The spermatozoon ofArthropoda. XIII. The cell surface. J. Ultrastruct. Res. 35, 582–605.
Bao, S.N., de Souza, W., 1992. Distribution of intramembranous particlesand filipin-sterol complexes in the spermatid and spermatozoon ofCulex quinquefasciatus (Culicidae). Biol. Cell 75, 37–44.
Dallai, R., Afzelius, B.A., 1982. On zipper-lines or particle arrays withinthe plasma membrane of hemipteran spermatozoa (Heteroptera,Insecta). J. Ultrastruct. Res. 80, 197–205.
Dallai, R., Afzelius, B.A., 1990. Microtubular diversity in insect sperma-tozoa: results obtained with a new fixative. J. Struct. Biol. 103, 164–179.
Dallai, R., Afzelius, B.A., 1995. Sperm structure of Trichoptera II. Theaflagellated spermatozoa of Hydroptila, Orthotrichia and Stactobia
(Hydroptilidae). Int. J. Insect Morphol. Embryol. 24, 161–170.Dallai, R., Xue, L., Yin, W.Y., 1990. Aflagellated spermatozoa of
Huhentomon and Acerella (Protura, Apterygota). Int. J. Insect Mor-phol. Embryol. 19, 211–217.
Dallai, R., Lombardo, B.M., Mercati, D., Vanin, S., Lupetti, P., 2007.Sperm structure of Limoniidae and their phylogenetic relationshipwith Tipulidae (Diptera, Nematocera). Arthropod Struct. Dev..doi:10.1016/j.asd.2007.05.00.
Folliot, R., Maillet, P.L., 1970. Ultrastructure de la spermiogenese et duspermatozoıde de divers insectes Homopteres. In: Baccetti, B. (Ed.),Comparative Spermatology. Academic Press, New York, pp. 289–300.
Friend, D.S., 1984. Membrane organization and differentiation in theguinea-pig spermatozoon. In: Van Blerkom, J., Motta, P.M. (Eds.),Ultrastructure of Reproduction. Martinus Nijhoff, Boston, pp. 75–85.
Friend, D.S., Fawcett, D.W., 1974. Membrane differentiation in freeze-fractured mammalian sperm. J. Cell Biol. 63, 641–664.
Jamieson, B.G.M., Dallai, R., Afzelius, B.A., 1999. InsectsTheir Sperma-tozoa and Phylogeny. Scientific Publishers, New Hampshire, USA,555p.
Joly, D., 1987. Between species divergence of cyst length distributions inthe Drosophila melanogaster species complex. Jpn. J. Genet. 62, 257–263.
Joly, D., Bressac, C., Lachaise, D., 1995. Disentangling giant sperm.Nature 377, 202.
Joly, D., Bressac, C., Jaillard, D., Lachaise, D., Lemullois, M., 2003. Thesperm roller: a modified testicular duct linked to giant sperm transportwithin the male reproductive tract. J. Struct. Biol. 142, 348–355.
Joly, D., Luck, N., Dejonghe, B., in press. Adaptations to long sperm inDrosophila: correlated development of the sperm roller and spermpackaging. J. Exp. Zool. (Mol. Dev. Evol) 308B, doi:10.1002/jez.b.21167.
Neesen, J., Heinlein, U.A.O., Glatzer, K.H., Bunemann, H., 1999.Proteins with tandemly arranged repeats of a highly charged 16-amino-acid motif encoded by the Dhmst101 gene family are structuralcomponents of the outer sheath of the extremely elongated sperm tailsof Drosophila hydei. Dev. Growth Differ. 41, 93–99.
Nielsen, M.G, Raff, E.C., 2002. The best of all worlds or the best possibleworld? Developmental constraint in the evolution of beta tubulin andthe sperm tail axoneme. Evol. Dev. 4, 303–315.
Nielsen, M.G., Caserta, J.M., Kidd, S.J., Phillips, C.M., 2006. Functionalconstraint underlies 60 million years stasis of Dipteran testis-specific b-tubulin. Evol. Dev. 8, 23–29.
Parker, G.A., 1982. Why are there so many tiny sperm? Spermcompetition and the maintenance of two sexes. J. Theor. Biol. 96,281–294.
Pasini, M.E., Cattaneo, F., Pinto, M.R., De Santis, R., Perotti, M.E., 1999.Plasma membrane association and preliminary characterization ofDrosophila sperm surface glycosidases. Mol. Reprod. Dev. 52, 166–173.
Perotti, M.E., 1969. Ultrastructure of the mature sperm of Drosophila
melanogaster Meig. J. Submicrosc. Cytol. 1, 171–196.Perotti, M.E., Riva, A., 1988. Concanavalin A binding sites on the surface
of Drosophila melanogaster sperm: a fluorescent and ultrastructuralstudy. J. Ultrastruct. Res. 100, 173–182.
Perotti, M.E., Pasini, M., 1995. Glycoconjugates of the surface of thespermatozoa of Drosophila melanogaster: a qualitative and quantita-tive study. J. Exp. Zool. 271, 311–318.
Perotti, M.E., Cattaneo, F., Pasini, M.E., Verni, F., Hackstein, J.H., 2001.Male sterile mutant casanova gives clues to mechanisms of sperm-egginteractions in Drosophila melanogaster. Mol. Reprod. Dev. 60, 248–259.
Pitnick, S., Markow, T.A., 1994. Male gametic strategies: sperm size,testes size and the allocation of ejaculates among successive mates bythe sperm-limited fly Drosophila pachea and its relatives. Am. Nat. 143,785–819.
Pitnick, S., Markow, T.A., Spicer, G.S., 1995a. Delayed male maturity is acost of producing large sperm in Drosophila. Proc. Nat. Acad. Sci.USA 92, 10614–10618.
Pitnick, S., Spicer, G.S., Markow, T.A., 1995b. How long is a giant sperm?Nature 375, 109.
Pitnick, S., Markow, T.A., Spicer, G.S., 1999. Evolution of multiple kindsof female sperm storage organs in Drosophila. Evolution 53, 1804–1822.
Stanley, H.P., Bowman, J.T., Romrell, L.J., Reed, S.C., Wilkinson, R.F.,1972. Fine structure of normal spermatid differentiation in Drosophila
melanogaster. J. Ultrastruct. Res. 41, 433–466.Tokuyasu, K.T., 1974. Dynamics of spermiogenesis in Drosophila mela-