onmental degradation associated with the end-Guadalupian, end-Permian, andend-Triassic extinctions has been inferred from numerous geochemical and sedimentological studies, but thelong-term impacts of this extinction-associated stress on the evolutionary trajectories of marineinvertebrates have not been explored. An examination of the diversity, extinction, paleoenvironmentalrange, and geographical distribution of marine stenolaemate bryozoans during the Permian to Jurassicinterval provides striking new evidence of the taxonomic and ecological influence of these mid-Phanerozoicextinctions on one of the most abundant components of the Paleozoic Fauna. Elevated bryozoan extinctionrates during the Late Permian and Late Triassic were coupled with major changes in their habitats. Bryozoansgradually disappeared from deep-water offshore settings during the Late Permian and from nearshore andoffshore settings during the Late Triassic. Re-colonization of these environments in the wake of each crisiswas delayed but coupled with increases in global generic diversity. The taxonomic effects of the end-Guadalupian extinction were milder than previously described, even though ecologically bryozoans werebecoming restricted to nearshore settings. The end-Permian mass extinction remained the largest forbryozoans, drastically reducing global and assemblage generic diversity and triggering a permanent changein their paleoenvironmental preferences from nearshore to mid-shelf settings. The 285 Myr dominance ofstenolaemate bryozoans ended during the Late Triassic when all but one order (Cyclostomata) becameextinct, initiating a taxonomic switch between stenolaemate and gymnolaemate bryozoans. Moreover,spatio-temporal variations in the paleoenvironmental history of bryozoans imply that Late Permian and LateTriassic marine environmental instability resulted largely from some stressful deep-water phenomenon.High extinction rates in nearshore environments in the Late Permian provide a link between marine andterrestrial/atmosphere extinction-related perturbations.
The transition between Paleozoic and Mesozoic communities ischaracterized by three devastating crises (Raup and Sepkoski,1982). Theend-Guadalupian (Middle Permian) and end-Permian extinctions aretwo independent events that took place within 8 million years (Myr) ofeach other in the Late Permian (e.g., Stanley and Yang, 1994; Gradsteinet al., 2004; Mundil et al., 2004). Together, they contributed to the mostsevere drop in biodiversity of the Phanerozoic and a major shift in thetaxonomic and ecological structure of marine communities (Gould andCalloway, 1980; Fraiser and Bottjer, 2005; Erwin, 2006; Clapham andBottjer, 2007a; Fraiser and Bottjer, 2007b). Fifty-twomillionyears later atthe end of the Triassic, many of the Paleozoic survivors who hadstruggled to re-diversify in the wake of the Late Permian events becameextinct. These three extinctions, spanning about 61 Myr, were accom-
1 213 740 8801.
l rights reserved.
panied by at least two periods of sustained environmental instability.Late Permian environmental degradation likely started at the end of theMiddle Permian and persisted through the Early Triassic (e.g., Isozaki,1997; Grice et al., 2005; Huey and Ward, 2005; Algeo et al., 2007). LateTriassic environmental stress inferred fromocean-atmospheremodelingbased on elevated pCO2 (McElwain et al., 1999; Huynh and Poulsen,2005) was likely initiated during the Rhaetian (Ciarapica, 2007).
The end-Guadalupian and end-Permian extinctions preferentiallyterminated sessile members of the Paleozoic Fauna (i.e., bryozoans,brachiopods, and crinoids) (Knoll et al., 1996, 2007). While all groupssuffered considerable losses at the end of the Permian, members of theModern Fauna (i.e., bivalves, gastropods) fared better in the immediateaftermath. Knoll et al. (2007) reviewed the effects of environmentalperturbations on both faunas and concluded that extinction selectivityin the Late Permian was linked to environmental stress, specificallyhypercapnia. Hypercapnia, or CO2-poisoning, is one of several proposedmechanisms for the end-Permian extinction; others include variousoceanographic, climatic, and extraterrestrial processes (Erwin, 2006).Regardless of the trigger, Late Permian–Early Triassic environmental
Fig. 1. Changes in the generic diversity of members of the Paleozoic and Modern Faunas across the end-Guadalupian and end-Permian extinctions. Modified from Knoll et al. (1996).Gray area represents the duration of extinction-related environmental stress based on sedimentological and geochemical evidence (Isozaki, 1997; Woods et al., 1999; Payne et al.,2004; Pruss et al., 2004).
147C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
stress had a more lasting effect on members of the Paleozoic Fauna.Secular changes in biodiversity during the Permian–Triassic (P/T)interval indicate that the re-diversification of sessile organisms duringthe Early Triassic was slow and that pre-extinction (Middle Permian)diversity levels were not yet re-established as of the Late Triassic (Fig.1).
In this study, we examined the transition of bryozoans through thePermian to Jurassic interval in an attempt to constrain the effects ofmassextinction-related environmental stress on their evolutionary history.Stenolaemate bryozoans, diverse and abundant members of thePaleozoic Fauna, nearly became extinct at the end of the Triassic (onegroup, Cyclostomata, survived) andwere replaced as thedominantpost-Jurassic group by the Modern Fauna gymnolaemate bryozoans (Fig. 2)(Sepkoski, 1981; Taylor and Larwood, 1990). Analyses of bryozoandiversity changes, paleoenvironmental transitions, and paleogeographicdistribution have the potential to reveal why these mid-Phanerozoicextinctions had amuchmore significant impact on somegroups (i.e., thePaleozoic Fauna) than others.
2. Mid-Phanerozoic extinctions: sources of environmental stress
2.1. End-Guadalupian extinction
The end-Guadalupian extinction is an independent Late Permianevent with species extinction rates comparable to those of the end-
Fig. 2. Schematic illustrating the range of stenolaemate and gymnolaemate (Gymno) bryo
Permian (Stanley and Yang, 1994). Although still a matter of contention,volcanism (Emeishan igneous province), global sea level fall, and a lateMiddle Permian high bio-productivity period (the “Kamura” event) areall potential kill mechanisms (Hallam and Wignall, 1999; Zhou et al.,2002; Isozaki et al., 2007). However, negative carbon isotopic excursionsacross the extinction boundary and a gradual trend towards decreasingvalues during the early Late Permian suggest that the causalmechanismwas gradual, which precludes catastrophic processes (i.e., extraterres-trial impact, gas hydrate release) and implies that extinction-relatedenvironmental instability was likely protracted through the Middle–Late Permianboundary (see Isozaki et al., 2007, Fig. 4). In fact, lithologicalstudies have shown that the development of deep-water anoxiaassociated with the P/T superanoxia event was initiated at the end ofthe Middle Permian (Isozaki, 1997) and is the likely source of thesustained Late Permian environmental degradation.
2.2. End-Permian extinction
The end-Permian mass extinction is arguably the largest suchevent, marked by the disappearance of about 80% of marine speciesand 49% and 63% of marine and terrestrial families, respectively (Raupand Sepkoski, 1982; Stanley and Yang, 1994; Benton, 1995). Hypothe-tical causal mechanisms for the end-Permian extinction include arange of gradual and catastrophic processes: widespread oceanic and
zoans through the Permian to Jurassic interval. Data from Powers and Pachut (2008).
148 C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
atmospheric anoxia (Wignall and Twitchett, 1996; Isozaki, 1997; Hueyand Ward, 2005), hypercapnia (Knoll et al., 1996), euxinia (H2Spoisoning) (Nielsen and Shen, 2004; Grice et al., 2005; Kump et al.,2005), massive volcanism and global warming (Renne et al., 1995;Kamo et al., 2003), methane oxidation (Krull and Retallack, 2000;Ryskin, 2003), and a meteorite impact (Becker et al., 2001). Thefundamental kill mechanism remains unclear and is likely acombination of several of the aforementioned processes (Ward,2006). Whereas evidence for a catastrophic extraterrestrial scenariois not compelling (Isozaki, 2001; Farley et al., 2005), a wealth ofgeochemical, sedimentological and paleontological data indicate thatthe end-Permian crisis was the result of a prolonged interval ofenvironmental degradation that lasted through the Early Triassic(Isozaki, 1997;Woods et al., 1999; Payne et al., 2004; Pruss et al., 2004,2006; Clapham and Bottjer, 2007b).
2.3. End-Triassic extinction
Whereas 80% of marine invertebrate species became extinctduring the end-Triassic biotic event (Sepkoski, 1996), marinevertebrates were not significantly affected (Benton, 1991; Hallamand Wignall, 1997). Postulated extinction mechanisms include CO2
and SO2 emissions associated with flood basalt volcanism of theCentral Atlantic Magmatic Province, gas hydrate release fromseafloor sediments, and a meteorite impact (Pálfy et al., 2001;Beerling and Berner, 2002; Cohen and Coe, 2002; Hesselbo et al.,2002; Olsen et al., 2002). Evidence of anoxic or euxinic conditions inthe late Triassic is not conclusive (e.g., Tanner et al., 2004), althoughanoxic conditions have been reported from several Late Triassicsections in Europe and British Columbia (Sephton et al., 2002; Wardet al., 2004; Ciarapica, 2007; van de Schootbrugge et al., 2007). LateTriassic marine anoxia is also invoked from ocean-atmospheremodeling that suggests enhanced oceanic stratification anddecreased levels of oxygen in the water column (Huynh and Poulsen,2005). However, negative carbon isotopic excursions from severalsections around the world provide the most compelling evidence forenvironmental instability during the Triassic–Jurassic (T/J) boundaryinterval (McElwain et al., 1999; Pálfy et al., 2001; Ward et al., 2001;Hesselbo et al., 2002).
3. Methods
We created two global data sets using bryozoan marine fossilassemblages in order to quantify the diversity, extinction, anddistribution patterns of bryozoans through the Permian to Jurassicinterval. Data were culled primarily from published literature,taxonomic monographs, and the Paleobiology Database (www.paleodb.org) (for list of references used in this study see Powers andBottjer, 2007).
In the first compilation, we recorded the presence of marinestenolaemate bryozoan genera at the stage level to calculate newextinction percentage rates for the end-Guadalupian, end-Permian,and end-Triassic events. Previously calculated rates for these extinc-tions were outdated in light of many recently published taxonomicstudies and improvements in P/T and T/J biostratigraphy thatenhanced the time-stratigraphic resolution of many bryozoanassemblages previously assigned to older broad time units (i.e.,Kazanian, Dzhufian).
The second compilation recorded the generic composition, geo-graphic and stratigraphic location, and depositional environment of396 marine bryozoan assemblages from the Early Permian throughthe Early Jurassic. Depositional settings, categorized into seven broadenvironmental groups along an onshore–offshore gradient, wereidentified using physical processes, mainly water energy, as reflectedin the type of sediments and its grain size, fossil preservation, andsedimentary features (for detailed methodology and paleoenviron-
mental criteria see Sepkoski and Miller, 1985; Sepkoski, 1987; Bottjeret al., 1988; Bottjer and Jablonski, 1988; Powers and Bottjer, 2007).Environmental categories are defined in Powers and Bottjer (2007)and include nearshore environments above fairweather wave base(including high-energy settings such as shoals and bars), inner shelfand lagoon environments above normal stormwave base, middle shelfenvironments between normal and maximum stormwave base, reefsand bioherms, outer shelf settings immediately landward of the shelfedge, deep-water slope mud mounds, and slope and basin settingsbeyond the shelf edge.
Generic richness data for bryozoan faunal assemblages assigned toan environmental categorywere plotted against time to create a time–environment (T–E) diagram. A T–E diagram consists of the onshore–offshore gradient described above on the horizontal axis and time onthe vertical axis. Time is divided into 20 Permian to Jurassic timeintervals representing a different stage based on the InternationalStratigraphic Chart (Gradstein et al., 2004). The resulting matrixcontains 140 T–E bins. Fossil assemblages were assigned to a time binusing conodont, fusulinid, or brachiopod biostratigraphic data. Weused a taphonomic control group, the rhynchonelliform Brachiopoda,to determine potential preservational bias in instances where nobryozoan genera were reported for a particular T–E bin. Rhynchonelli-form brachiopods and bryozoans are commonly found together inmarine communities and as lophotrochozoans, they share a similarlophophorate filter-feeding life habit and skeletal mineralogy. Thepresence of rhynchonelliform brachiopods then implies that theabsence of bryozoans from those settings is a real phenomenon, not ataphonomic artifact. Unequal sampling among environmental binsand differential preservation of bryozoan faunal assemblages causedby facies variation were accounted for by selecting and plotting themost generically diverse bryozoan assemblage for each environmentalcategory on the T–E diagram. Similar T–E diagrams were constructedfor each Permian–Early Jurassic marine bryozoan order to furtherdetermine the degree to which environmental factors controlled theonshore–offshore distribution of marine bryozoans.
Finally, the paleogeographic distribution of bryozoan faunalassemblages combined with paleoenvironmental data was plottedon paleogeographic reconstructions (Scotese, 2001) to examinegeographical trends across the end-Guadalupian, end-Permian, andend-Triassic extinction intervals.
4. Results
4.1. Bryozoan generic diversity and extinction rates
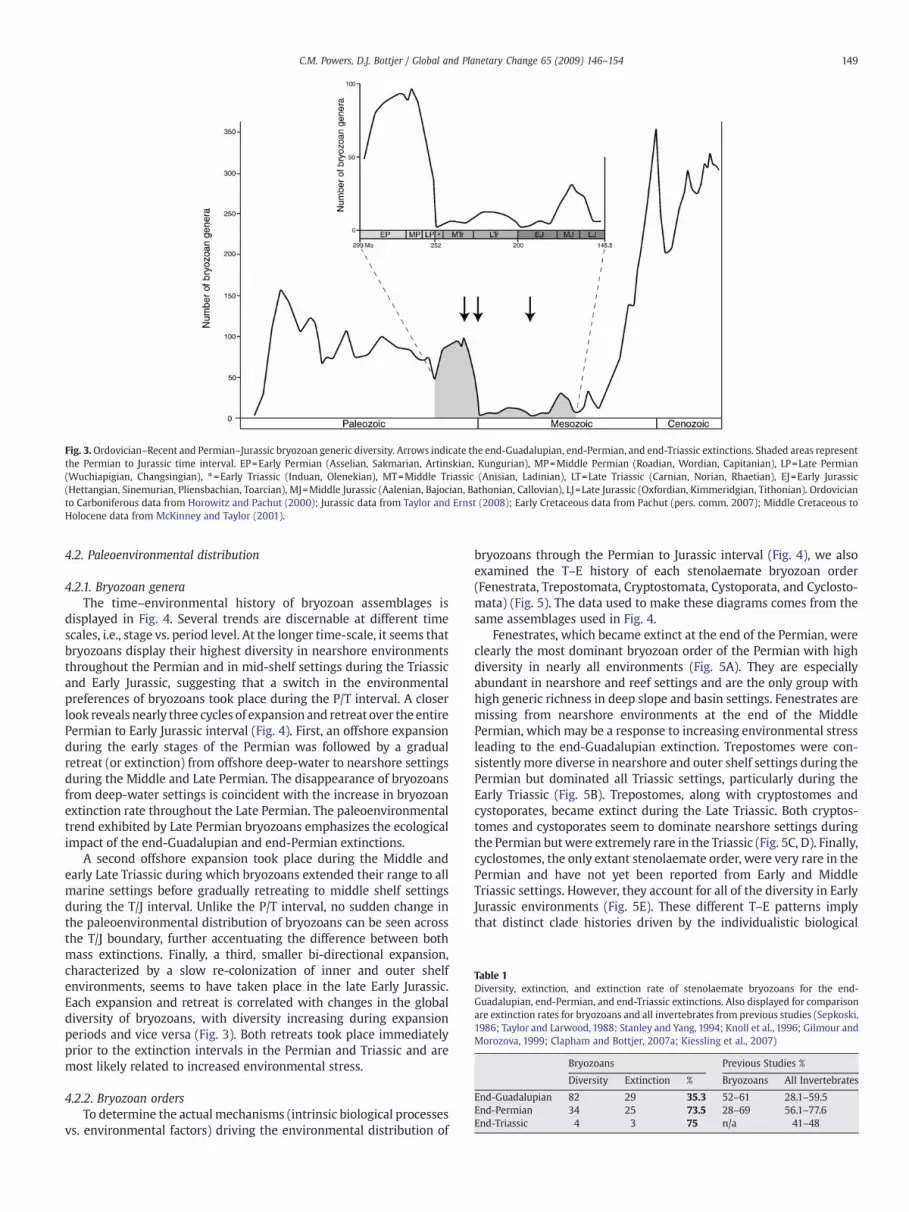
The Permian to Jurassic generic diversity of stenolaemate bryozo-ans is characterized by an abrupt extinction in the Late Permian andlow diversity levels throughout the Triassic and Jurassic (Fig. 3). Pre-extinction diversity levels of marine bryozoan genera were not re-established until themid-Cretaceous, ~150Myr later (Fig. 3). New LatePermian extinction rates, calculated using our genus-level compila-tion but excluding stratigraphic singletons, differ from previouslypublished rates for bryozoans, but fall within the intensity range for allmarine invertebrates (Table 1). Bryozoan extinction intensity steadilyincreased throughout the Late Permian, with extinction of 35.3% (29/82) at the end of the Guadalupian and 73.5% (25/34) for the end-Permianmass extinction. The end-Triassicmass extinction seemed themost severe, with 75% (3/4) of bryozoan genera becoming extinct.However, the standing diversity of bryozoan genera in the latestTriassic had been severely diminished by a relatively high extinction atthe Norian/Rhaetian boundary (60%) (Powers and Pachut, 2008) andwas much lower than at the end of the Permian (Table 1), thereforeenhancing the magnitude of the end-Triassic extinction. The increas-ing rates of extinction throughout the Late Permian and Late Triassicmay be symptomatic of protracted extinction-related environmentalstress.
Table 1Diversity, extinction, and extinction rate of stenolaemate bryozoans for the end-Guadalupian, end-Permian, and end-Triassic extinctions. Also displayed for comparisonare extinction rates for bryozoans and all invertebrates from previous studies (Sepkoski,1986; Taylor and Larwood, 1988; Stanley and Yang, 1994; Knoll et al., 1996; Gilmour andMorozova, 1999; Clapham and Bottjer, 2007a; Kiessling et al., 2007)
Bryozoans Previous Studies %
Diversity Extinction % Bryozoans All Invertebrates
Fig. 3.Ordovician–Recent and Permian–Jurassic bryozoan generic diversity. Arrows indicate the end-Guadalupian, end-Permian, and end-Triassic extinctions. Shaded areas representthe Permian to Jurassic time interval. EP=Early Permian (Asselian, Sakmarian, Artinskian, Kungurian), MP=Middle Permian (Roadian, Wordian, Capitanian), LP=Late Permian(Wuchiapigian, Changsingian), ⁎=Early Triassic (Induan, Olenekian), MT=Middle Triassic (Anisian, Ladinian), LT=Late Triassic (Carnian, Norian, Rhaetian), EJ=Early Jurassic(Hettangian, Sinemurian, Pliensbachian, Toarcian), MJ=Middle Jurassic (Aalenian, Bajocian, Bathonian, Callovian), LJ=Late Jurassic (Oxfordian, Kimmeridgian, Tithonian). Ordovicianto Carboniferous data from Horowitz and Pachut (2000); Jurassic data from Taylor and Ernst (2008); Early Cretaceous data from Pachut (pers. comm. 2007); Middle Cretaceous toHolocene data from McKinney and Taylor (2001).
149C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
4.2. Paleoenvironmental distribution
4.2.1. Bryozoan generaThe time–environmental history of bryozoan assemblages is
displayed in Fig. 4. Several trends are discernable at different timescales, i.e., stage vs. period level. At the longer time-scale, it seems thatbryozoans display their highest diversity in nearshore environmentsthroughout the Permian and in mid-shelf settings during the Triassicand Early Jurassic, suggesting that a switch in the environmentalpreferences of bryozoans took place during the P/T interval. A closerlook reveals nearly three cycles of expansion and retreat over the entirePermian to Early Jurassic interval (Fig. 4). First, an offshore expansionduring the early stages of the Permian was followed by a gradualretreat (or extinction) from offshore deep-water to nearshore settingsduring the Middle and Late Permian. The disappearance of bryozoansfrom deep-water settings is coincident with the increase in bryozoanextinction rate throughout the Late Permian. The paleoenvironmentaltrend exhibited by Late Permian bryozoans emphasizes the ecologicalimpact of the end-Guadalupian and end-Permian extinctions.
A second offshore expansion took place during the Middle andearly Late Triassic during which bryozoans extended their range to allmarine settings before gradually retreating to middle shelf settingsduring the T/J interval. Unlike the P/T interval, no sudden change inthe paleoenvironmental distribution of bryozoans can be seen acrossthe T/J boundary, further accentuating the difference between bothmass extinctions. Finally, a third, smaller bi-directional expansion,characterized by a slow re-colonization of inner and outer shelfenvironments, seems to have taken place in the late Early Jurassic.Each expansion and retreat is correlated with changes in the globaldiversity of bryozoans, with diversity increasing during expansionperiods and vice versa (Fig. 3). Both retreats took place immediatelyprior to the extinction intervals in the Permian and Triassic and aremost likely related to increased environmental stress.
4.2.2. Bryozoan ordersTo determine the actual mechanisms (intrinsic biological processes
vs. environmental factors) driving the environmental distribution of
bryozoans through the Permian to Jurassic interval (Fig. 4), we alsoexamined the T–E history of each stenolaemate bryozoan order(Fenestrata, Trepostomata, Cryptostomata, Cystoporata, and Cyclosto-mata) (Fig. 5). The data used to make these diagrams comes from thesame assemblages used in Fig. 4.
Fenestrates, which became extinct at the end of the Permian, wereclearly the most dominant bryozoan order of the Permian with highdiversity in nearly all environments (Fig. 5A). They are especiallyabundant in nearshore and reef settings and are the only group withhigh generic richness in deep slope and basin settings. Fenestrates aremissing from nearshore environments at the end of the MiddlePermian, which may be a response to increasing environmental stressleading to the end-Guadalupian extinction. Trepostomes were con-sistently more diverse in nearshore and outer shelf settings during thePermian but dominated all Triassic settings, particularly during theEarly Triassic (Fig. 5B). Trepostomes, along with cryptostomes andcystoporates, became extinct during the Late Triassic. Both cryptos-tomes and cystoporates seem to dominate nearshore settings duringthe Permian but were extremely rare in the Triassic (Fig. 5C, D). Finally,cyclostomes, the only extant stenolaemate order, were very rare in thePermian and have not yet been reported from Early and MiddleTriassic settings. However, they account for all of the diversity in EarlyJurassic environments (Fig. 5E). These different T–E patterns implythat distinct clade histories driven by the individualistic biological
Fig. 4. Contoured maximum generic diversity of Permian–Early Jurassic marine stenolaemate bryozoan assemblages. Each dot represents a data point, either for a bryozoanassemblage or for the absence of bryozoans validated by the taphonomic control group. Contouring was done separately for each time period using cubic interpolation. E=Early,M=Middle, L=Late, NS=Nearshore, IS & L=Inner Shelf and Lagoon, MS=Middle Shelf, R=Reefs, OS=Outer Shelf, SM=Slope Mounds, S & DB=Slope and Deep Basin.
150 C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
tendencies of each stenolaemate order underlie the paleoenviron-mental trends of stenolaemate bryozoans as a group.
4.3. Paleogeography
The paleogeographical distribution of all known marine bryozoanassemblages was plotted across each extinction interval (Fig. 6). Thedistribution of bryozoans across the end-Guadalupian and end-Triassic extinctions remained similar across each extinction interval.However, a striking switch in assemblage distribution took placeacross the P/T boundary. Bryozoan assemblages that were widespreadduring the Late Permian became restricted to high latitude settingsalong northwest Pangea in the eastern Panthalassa. These patternsfurther emphasize the important ecologic and taxonomic implicationsof the end-Permian mass extinction.
5. Evolutionary and ecological impact of long-termenvironmental stress
Bryozoan diversity and distribution through the Permian toJurassic interval clearly indicate that prolonged Late Permian andLate Triassic environmental stress in marine environments had aconsiderable impact on the evolutionary history of stenolaematebryozoans. Extinction rates during the Late Permian and Late Triassic
were elevated above background rates and Late Permian originationswere suppressed. The declines in diversity were coupled with gradualchanges in the environmental range of bryozoan genera, suggestingthat the effect of extinction-related stress varied among marinesettings and between mass extinctions. However, the different T–Ehistories of each stenolaemate order prior to the Late Permian andLate Triassic imply that during times of reduced stress, intrinsicbiological processes largely drove the paleoenvironmental distribu-tion of bryozoans.
The end-Guadalupian extinction was not as severe as previouslydescribed for bryozoans (Gilmour and Morozova, 1999), but insteadcan be considered the first in a series of events that ended the 285Myrdominance of marine stenolaemate bryozoans. The lack of geogra-phical trends in the distribution of Capitanian and Wuchiapingianassemblages (Fig. 6) indicates that the extinctionwas global, as shownby Gilmour and Morozova (1999) in their analysis of Late Permianbryozoan faunal provinces.
The end-Permian extinction remains the largest, eliminating allbut nine stenolaemate genera belonging to five families. ThesePaleozoic holdovers, most of them trepostomes, dominated theTriassic recovery, both in terms of diversity and environmental extent(Fig. 5B). In addition to the considerable loss in biodiversity, theenvironmental distribution of bryozoans through the P/T intervalprovides striking new evidence of the influence of extinction-related
Fig. 5. Contoured maximum generic diversity of Permian–Early Jurassic marine bryozoan orders. A=Fenestrata, B=Trepostomata, C=Cryptostomata, D=Cystoporata,E=Cyclostomata. See Fig. 4 for explanation of acronyms.
151C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
Fig. 6. Paleogeographic distribution of all marine stenolaemate bryozoan assemblages across the end-Guadalupian, end-Permian, and end-Triassic extinction intervals. Base mapsmodified from Scotese (2002).
152 C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
stress on the marine realm. Late Permian bryozoan assemblagegeneric richness declined significantly starting at the end-Guadalu-pian boundary and bryozoans did not occupy deep offshore settings.These patterns imply that environmental degradation resulted fromthe gradual encroachment of some deep-water phenomenon onto theshelves. As discussed in Section 2, sources of Late Permian environ-mental stress are numerous but most notably include the develop-ment of deep-water anoxia initiated at the end of the Middle Permian(Isozaki, 1997).
A second interesting trend to emerge from the spatial distribution ofbryozoans is the apparent long-term switch in their environmentalpreferences coincidentwith the end-Permianmass extinction. Thepost-Jurassic T–E history of cheilostome bryozoans (see Bottjer and Jablonski,1988, Fig. 2) and preliminary data on Middle and Late Jurassiccyclostomes indicate that assemblage generic richness has indeedremained higher in middle and outer shelf environments since thebeginning of the Mesozoic. Contemporaneous to this environmentalswitch is a change in the global latitudinal distribution of bryozoan-richdeposits from a Paleozoic tropical to amodern non-tropical distribution(Taylor and Allison, 1998). Both of these long-term changes inenvironmental preferences correspond to the taxonomic turnoverbetween stenolaemate and gymnolaemate bryozoans that graduallycame about during the Late Permian to Jurassic interval.
Geographical trends in bryozoan assemblages across the P/Tboundary reveal a third, albeit short-lived, change in environmentalpreferences. The restricted occurrence of Early Triassic bryozoansoutside of Tethys compared to their otherwise cosmopolitan distribu-tion within Tethys in the Late Permian suggests that boreal settingsmay have served as refugia for bryozoans. In fact, geochemical,sedimentological, and paleontological evidences indicate that envir-onmental instability in high latitude settings was short-lived and the
recovery of marine ecosystems there occurred much more rapidlythan in Early Triassic equatorial environments (Wignall et al., 1998;Pruss and Bottjer, 2004).
Extinction intensity for the end-Triassic extinction seems largerthan for the end-Permian. However, the standing diversity ofbryozoans in the Triassic was extremely low compared to the Permian,especially in the latest Triassic. Extinction was already elevatedfollowing the relative Carnian high in bryozoan diversity (Powers andPachut, 2008) and at least one of the remaining four stenolaemateorders became extinct before the end of the Triassic (Fig. 2). Two otherstenolaemate orders became extinct during the Norian stage, bringingan end to almost all Paleozoic holdovers and the stenolaemate285 Myr history of dominance. While Late Triassic environmentalstress is not causally related to the prolonged Late Permian–EarlyTriassic instability, both episodes contributed to the decline indiversity and assemblage generic richness of bryozoans that beganwith the end-Guadalupian extinction, and continued to suppressoriginations throughout the Triassic and the early Jurassic (Powersand Pachut, 2008; Taylor and Ernst, 2008).
6. Late Permian terrestrial–marine link
As reviewed in a previous section of this paper, the gradual andprotracted nature of Late Permian–Early Triassic and Late Triassicenvironmental degradation was the result of oceanographic and atmo-spheric processes (see Isozaki, 1997; McElwain et al., 1999; Grice et al.,2005; Huey andWard, 2005), although contention over the exact natureof the relationship between events in the marine and terrestrial realmstill exists. Environmental trends in bryozoanextinction rates prior to theend-Guadalupian and end-Permian mass extinction provide furtherevidence for the link between terrestrial and marine perturbations. For
Table 2Diversity, extinction, and extinction rates of stenolaemate bryozoans acrossenvironments for the end-Guadalupian and end-Permian extinctions
153C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
both events, extinction intensitywashighest in nearshore environments.A global end-Permian extinction of rooted plants, which contributed to asignificant increase in terrestrial erosion and sediment influx into theoceans, and a biocalcification crisis resulting from elevated atmosphericCO2mayhave caused increased instability in coastal settings (Ward et al.,2000; Sephton et al., 2005; Fraiser and Bottjer, 2007a). While higherextinction rates in nearshore environments are expected during back-ground times of extinction due to changing and unpredictable condi-tions, extinction intensity generally increases offshore during times ofmass extinction (Sepkoski, 1987). If reefs are excluded because theirplacement along the shelf can vary significantly, extinction intensityfrom the inner shelf to the outer shelf does increase (Table 2), thushighlighting the anomalous extinction intensity in nearshore settings.
7. Conclusions
Diversity and spatio-temporal analyses of Permian to Jurassicmarine stenolaemate bryozoans reveal that long-term Late Permianand Late Triassic environmental instability had a major impact on theevolutionary trajectory of bryozoans, facilitating a taxonomic switchbetween stenolaemate and gymnolaemate bryozoans and a long-termecological change in their environmental and geographical prefer-ences. The importance of this protracted extinction-related stress onmarine settings and organisms reinforces the notion of an oceano-graphic process as the trigger of both the Late Permian and LateTriassic extinctions. Integrating global diversity studies of fossilmarine organisms with their long-term paleoenvironmental trendsallows us to better constrain and understand the effects of large-scaleenvironmental perturbations on marine settings.
Acknowledgements
We thank Paul Taylor and Joseph Pachut for sharing their Jurassicand Cretaceous bryozoan diversity data, respectively. MatthewClapham provided invaluable help with Permian biostratigraphicdata. This research was supported by a USC WiSE grant to D.J.B. andgrants from the Geological Society of America, The PaleontologicalSociety, the American Museum of Natural History, the Yale PeabodyMuseum, and the USC Department of Earth Sciences to C.M.P.
References
Algeo, T.J., Hannigan, R., Rowe, H., Brookfield, M., Baud, A., Krystyn, L., Ellwood, B.B.,2007. Sequencing events across the Permian–Triassic boundary, Guryul Ravine(Kashmir, India). Palaeogeography, Palaeoclimatology, Palaeoecology 252,328–346.
Becker, L., Poreda, R.J., Hunt, A.G., Bunch, T.E., Rampino, M.R., 2001. Impact event at thePermian–Triassic boundary: evidence from extraterrestrial noble gases in full-erenes. Science 291, 1530–1533.
Beerling, D., Berner, R.A., 2002. Biogeochemical constraints on the Triassic–Jurassicboundary carbon cycle event. Global Biogeochemical Cycles 16, 1036. doi:10.1029/2001GB001637.
Benton, M.J., 1991. What really happened in the Late Triassic? Historical Biology 5,263–278.
Benton, M.J., 1995. Diversification and extinction in the history of life. Science 268,52–58.
Bottjer, D.J., Jablonski, D., 1988. Paleoenvironmental patterns in the evolution of post-Paleozoic benthic marine invertebrates. Palaios 3, 540–560.
Bottjer, D.J., Droser, M.L., Jablonski, D., 1988. Palaeoenvironmental trends in the historyof trace fossils. Nature 333, 252–255.
Ciarapica, G., 2007. Regional and global changes around the Triassic–Jurassic boundaryreflected in the Late Norian–Hettangian history of the Apennine basins.Palaeogeography, Palaeoclimatology, Palaeoecology 244, 34–51.
Clapham, M.E., Bottjer, D.J., 2007a. Permian marine paleoecology and its implicationsfor large-scale decoupling of brachiopod and bivalve abundance and diversityduring the Lopingian (Late Permian). Palaeogeography, Palaeoclimatology, Palaeoe-cology 249, 283–301.
Clapham, M.E., Bottjer, D.J., 2007b. Prolonged Permian–Triassic ecological crisisrecorded by molluscan dominance in Late Permian offshore assemblages.Proceeding of the National Academy of Sciences 104, 12971–12975.
Cohen, A.S., Coe, A.L., 2002. New geochemical evidence for the onset of volcanism in theCentral Atlantic magmatic province and environmental change at the Triassic–Jurassic boundary. Geology 30, 267–270.
Erwin, D.H., 2006. Extinction: How Life on Earth Nearly Ended 250 Million Years Ago.Princeton University Press, Princeton, New Jersey.
Farley, K.A., Ward, P., Garrison, G., Mukhopadhyay, S., 2005. Absence of extraterrestrial3He in Permian–Triassic age sedimentary rocks. Earth and Planetary Science Letters240, 265–275.
Fraiser, M.L., Bottjer, D.J., 2005. Restructuring in benthic level-bottom shallow marinecommunities due to prolonged environmental stress following the end-Permianmass extinction. Comptes Rendus Palevol 4, 515–523.
Fraiser, M.L., Bottjer, D.J., 2007a. Elevated atmospheric CO2 and the delayed bioticrecovery from the end-Permian mass extinction. Palaeogeography, Palaeoclimatol-ogy, Palaeoecology 252, 164–175.
Fraiser, M.L., Bottjer, D.J., 2007b. When bivalves took over the world. Paleobiology 33,397–413.
Gilmour, E.H., Morozova, I.P., 1999. Biogeography of the Late Permian bryozoans.Paleontological Journal 33, 36–51.
Gould, S.J., Calloway, C.B., 1980. Clams and brachiopods — ships that pass in the night.Paleobiology 6, 383–396.
Gradstein, F.M., Ogg, J.G., Smith, A.G., Bleeker,W., Lourens, L.J., 2004. A newgeologic timescale with special reference to Precambrian and Neogene. Episodes 27, 83–100.
Grice, K., Cao, C., Love, G.D., Böttcher, M.E., Twitchett, R.J., Grosjean, E., Summons, R.E.,Turgeon, S.C., Dunning, W., Jin, Y., 2005. Photic zone euxinia during the Permian–Triassic superanoxic event. Science 307, 706–709.
Hallam, A., Wignall, P.B., 1997. Mass Extinctions and their Aftermath. Oxford UniversityPress, New York.
Hallam, A., Wignall, P.B., 1999. Mass extinctions and sea-level changes. Earth-ScienceReviews 48, 217–250.
Hesselbo, S.P., Robinson, S.A., Surlyk, F., Piasecki, S., 2002. Terrestrial and marineextinction at the Triassic–Jurassic boundary synchronized with major carbon-cycleperturbation: a link to initiation of massive volcanism? Geology 30, 251–254.
Horowitz, A.S., Pachut, J.F., 2000. The fossil record of bryozoan species diversity. In:Herrera-Cubilla, A., Jackson, J.B.C. (Eds.), Proceedings of the 11th InternationalBryozoology Association Conference. Smithsonian Tropical Research Institute,Balboa, R.P. , pp. 245–248.
Huey, R.B., Ward, P.D., 2005. Hypoxia, global warming, and terrestrial Late Permianextinctions. Science 308, 398–401.
Huynh, T.T., Poulsen, C.J., 2005. Rising atmospheric CO2 as a possible trigger for the end-Triassic mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology 217,223–242.
Isozaki, Y., 1997. Permo–Triassic boundary superanoxia and stratified superocean:records from lost deep sea. Science 276, 235–238.
Isozaki, Y., 2001. An extraterrestrial impact at the Permian–Triassic boundary? Science293, 2343a.
Isozaki, Y., Kawahata, H., Ota, A., 2007. A unique carbon isotope record across theGuadalupian–Lopingian (Middle–Upper Permian) boundary in mid-oceanic paleo-atoll carbonates: The high-productivity “Kamura event” and its collapse inPanthalassa. Global and Planetary Change 55, 21–38.
Kamo, S.L., Czamanske, G.K., Amelin, Y., Fedorenko, V.A., Davis, D.W., Trofimov, V.R.,2003. Rapid eruption of Siberian flood-volcanic rocks and evidence for coincidencewith the Permian–Triassic boundary and mass extinction at 251 Ma. Earth andPlanetary Science Letters 214, 75–91.
Kiessling, W., Aberhan, M., Brenneis, B., Wagner, P.J., 2007. Extinction trajectories ofbenthic organisms across the Triassic–Jurassic boundary. Palaeogeography, Palaeo-climatology, Palaeoecology 244, 201–222.
Knoll, A.H., Bambach, R.K., Canfield, D.E., Grotzinger, J.P., 1996. Comparative Earthhistory and Late Permian mass extinction. Science 273, 452–457.
Krull, E.S., Retallack, G.J., 2000. δ13C depth profiles from paleosols across the Permian–Triassic boundary: evidence for methane release. Geological Society of AmericaBulletin 112, 1459–1472.
Kump, L.R., Pavlov, A., Arthur, M.A., 2005. Massive release of hydrogen sulfide to thesurface ocean and atmosphere during intervals of oceanic anoxia. Geology 33,397–400.
McElwain, J.C., Beerling, D.J., Woodward, F.I., 1999. Fossil plants and global warming atthe Triassic–Jurassic boundary. Science 285, 1386–1390.
McKinney, F.K., Taylor, P.D., 2001. Bryozoan generic extinction and originations duringthe last one hundred million years. Palaeontologica Electronica 4, 1–26.
Mundil, R., Ludwig, K.R., Metcalfe, I., Renne, P.R., 2004. Age and timing of the Permianmass extinctions: U/Pb dating of closed-system zircons. Science 305, 1760–1763.
154 C.M. Powers, D.J. Bottjer / Global and Planetary Change 65 (2009) 146–154
Nielsen, J.K., Shen, Y., 2004. Evidence for sulfidic deep water during the Late Permian inthe East Greenland Basin. Geology 32, 1037–1040.
Olsen, P.E., Kent, D.V., Sues, H.D., Koeberl, C., Huber, H., Montanari, A., Rainforth, E.C.,Fowell, S.J., Szajna, M.J., Hartline, B.W., 2002. Ascent of dinosaurs linked to aniridium anomaly at the Triassic–Jurassic boundary. Science 296, 1305–1307.
Pálfy, J., Demény, A., Haas, J., Hetényi, M., Orchard, M.J., Veto, I., 2001. Carbon isotopeanomaly and other geochemical changes at the Triassic–Jurassic boundary from amarine section in Hungary. Geology 29, 1047–1050.
Payne, J.L., Lehrmann, D.J., Wei, J., Orchard, M.J., Schrag, D.P., Knoll, A.H., 2004. Largeperturbations of the carbon cycle from the end-Permian extinction. Science 305,506–509.
Powers, C.M., Bottjer, D.J., 2007. Bryozoan paleoecology indicates mid-Phanerozoicextinctions were the product of long-term environmental stress. Geology 35,995–998.
Powers, C.M., Pachut, J.F., 2008. Diversity and distribution of Triassic bryozoans in theaftermath of the end-Permian mass extinction. Journal of Paleontology 82,393–402.
Pruss, S.B., Bottjer, D.J., 2004. Early Triassic trace fossils of the western United States andtheir implications for prolonged environmental stress from the end-Permian massextinction. Palaios 19, 551–564.
Pruss, S.B., Bottjer, D.J., Corsetti, F.A., Baud, A., 2006. A global marine sedimentaryresponse to the end-Permian mass extinction: examples from southern Turkey andthe western United States. Earth-Science Reviews 78, 193–206.
Pruss, S.B., Fraiser, M., Bottjer, D.J., 2004. The proliferation of Early Triassic wrinklestructures: implications for environmental stress following the end-Permian massextinction. Geology 32, 461–464.
Raup, D.M., Sepkoski, J.J., 1982. Mass extinctions in themarine fossil record. Science 215,1501–1503.
Renne, P.R., Zichao, Z., Richards, M.A., Black, M.T., Basu, A.R., 1995. Synchrony and causalrelations between Permian–Triassic boundary crises and Siberian flood volcanism.Science 269, 1413–1416.
Ryskin, G., 2003. Methane-driven oceanic eruptions and mass extinctions. Geology 31,741–744.
Scotese, C.R., 2002. http://www.scotese.com. (PALEOMAP website).Sephton, M.A., Amor, K., Franchi, I.A., Wignall, P.B., Newton, R., Zonneveld, J.-P., 2002.
Carbon and nitrogen isotope disturbances and an end-Norian (Late Triassic)extinction event. Geology 30, 1119–1122.
Sephton, M.A., Looy, C.V., Brinkhuis, H., Wignall, P.B., de Leeuw, J.W., Visscher, H., 2005.Catastrophic soil erosion during the end-Permian biotic crisis. Geology 33,941–944.
Sepkoski, J.J., 1981. A factor analytic description of the Phanerozoic marine fossil record.Paleobiology 7, 36–53.
Sepkoski, J.J., 1986. Phanerozoic overview of mass extinctions. In: Raup, D.M., Jablonski, D.(Eds.), Patterns and Processes in the History of Life. Springer-Verlag, Berlin, pp. 277–295.
Sepkoski, J.J., 1987. Environmental trends in extinction during the Paleozoic. Science235, 64–66.
Sepkoski, J.J., 1996. Patterns of Phanerozoic extinction: a perspective from globaldatabases. In: Walliser, O.H. (Ed.), Global Events and Event Stratigraphy in thePhanerozoic. Springer-Verlag, Berlin, pp. 35–51.
Sepkoski, J.J., Miller, A.I., 1985. Evolutionary faunas and the distribution of Paleozoicmarine communities in space and time. In: Valentine, J.W. (Ed.), PhanerozoicDiversity Patterns: Profiles in Macroevolution. Princeton University Press, Prince-ton, pp. 153–190.
Stanley, S.M., Yang, X., 1994. A double mass extinction at the end of the Paleozoic era.Science 266, 1340–1344.
Tanner, L.H., Lucas, S.G., Chapman, M.G., 2004. Assessing the record and causes of LateTriassic extinctions. Earth-Science Reviews 65, 103–139.
Taylor, P.D., Larwood, G.P., 1988. Mass extinctions and the pattern of bryozoan evolution.In: Larwood, G.P. (Ed.), Extinction and Survival in the Fossil Record. Clarendon Press,Oxford, pp. 99–119.
Taylor, P.D., Larwood,G.P.,1990.Majorevolutionary radiations in theBryozoa. In: Taylor, P.D.,Larwood, G.P. (Eds.), Major Evolutionary Radiations. Clarendon, Oxford, pp. 209–233.
Taylor, P.D., Allison, P.A., 1998. Bryozoan carbonates through time and space. Geology26, 459–462.
Taylor, P.D., Ernst, A., 2008. Bryozoans in transition: the depauperate and patchy Jurassicbiota. Palaeogeography, Palaeoclimatology, Palaeoecology 263, 9–23.
van de Schootbrugge, B., Tremolada, F., Rosenthal, Y., Bailey, T.R., Feist-Burkhardt, S.,Brinkhuis, H., Pross, J., Kent, D.V., Falkowski, P.G., 2007. End-Triassic calcificationcrisis and blooms of organic-walled ‘disaster species. Palaeogeography, Palaeocli-matology, Palaeoecology 244, 126–141.
Ward, P.D., 2006. Impact from the Deep. Scientific American 395, 65–71.Ward, P.D., Garrison, G.H., Haggart, J.W., Kring, D.A., Beattie, M.J., 2004. Isotopic
evidence bearing on Late Triassic extinction events, Queen Charlotte Islands, BritishColumbia, and implications for the duration and cause of the Triassic/Jurassic massextinction. Earth and Planetary Science Letters 224, 589–600.
Ward, P.D., Haggart, J.W., Carter, E.S., Wilbur, D., Tipper, H.W., Evans, T., 2001. Suddenproductivity collapse associated with the Triassic–Jurassic boundary mass extinc-tion. Science 292, 1148–1151.
Ward, P.D., Montgomery, D.R., Smith, R., 2000. Altered river morphology in South Africarelated to the Permian–Triassic extinction. Science 289, 1740–1743.
Wignall, P.B., Twitchett, R.J., 1996. Oceanic anoxia and the End Permian mass extinction.Science 272, 1155–1158.
Wignall, P.B., Morante, R., Newton, R., 1998. The Permo–Triassic transition inSpitsbergen: δ13Corg chemostratigraphy, Fe and S geochemistry; facies, fauna andtrace fossils. Geological Magazine 135, 47–62.
Woods, A.D., Bottjer, D.J., Mutti, M., Morrison, J., 1999. Lower Triassic large sea-floorcarbonate cements: their origin and a mechanism for the prolonged biotic recoveryfrom the end-Permian mass extinction. Geology 27, 645–648.
Zhou, M., Malpas, J., Song, X., Robinson, P.T., Sun, M., Kennedy, A.K., Lesher, C.M., Keays,R.R., 2002. A temporal link between the Emeishan large igneous province (SWChina) and the end-Guadalupian mass extinction. Earth and Planetary ScienceLetters 196, 113–122.