213
| Date post: | 24-Mar-2023 |
| Category: |
Documents |
| Upload: | khangminh22 |
| View: | 0 times |
| Download: | 0 times |


Palaeobiogeography and BiodiversityChange: the Ordovician and
Mesozoic-Cenozoic Radiations

Geological Society Special PublicationsSociety Book Editors
A. J. FLEET (CHIEF EDITOR)P. DOYLE
F. J. GREGORYJ. S. GRIFFITHSA. J. HARTLEY
R. E. HOLDSWORTHA. C. MORTONN. S. ROBINSM. S. STOKERJ. P. TURNER
Reviewing procedures
The Society makes every effort to ensure that the scientific and production quality of its books matches that ofits journals. Since 1997, all book proposals have been refereed by specialist reviewers as well as by the Society'sBooks Editorial Committee. If the referees identify weaknesses in the proposal, these must be addressedbefore the proposal is accepted.
Once the book is accepted, the Society has a team of Book Editors (listed above) who ensure that the vol-ume editors follow strict guidelines on refereeing and quality control. We insist that individual papers can onlybe accepted after satisfactory review by two independent referees. The questions on the review forms aresimilar to those for Journal of the Geological Society. The referees forms and comments must be available tothe Society's Book Editors on request.
Although many of the books result from meetings, the editors are expected to commission papers that werenot presented at the meeting to ensure that the book provides a balanced coverage of the subject. Being accept-ed for presentation at the meeting does not guarantee inclusion in the book.
Geological Society Special Publications are included in the ISI Science Citation Index, but they do not havean impact factor, the latter being applicable only to journals.
More information about submitting a proposal and producing a Special Publication can be found on theSociety's web site: www.geolsoc.org.uk.
It is recommended that reference to all or part of this book should be made in one of the follow-ing ways:
CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change:the Ordovician and Mesozoic-Cenozoic Radiations. Geological Society, London, SpecialPublications, 194.
COPE, J. C. W. 2002. Diversification and biogeography of bivalves during the Ordovician Period.In: CRAME, J. A. & OWEN, A. W. (eds) Palaeobiogeography and Biodiversity Change: theOrdovician and Mesozoic-Cenozoic Radiations. Geological Society, London, SpecialPublications, 194, 35-52.

GEOLOGICAL SOCIETY SPECIAL PUBLICATIONS NO. 194
Palaeobiogeography and BiodiversityChange: the Ordovician and
Mesozoic-Cenozoic Radiations
E D I T E D B Y
J. A. CRAMEBritish Antarctic Survey, Cambridge, UK
and
A. W. OWENUniversity of Glasgow, UK
2002Published by
The Geological SocietyLondon

THE GEOLOGICAL SOCIETY
The Geological Society of London (GSL) was founded in 1807. It is the oldest national geological society in theworld and the largest in Europe. It was incorporated under Royal Charter in 1825 and is Registered Charity210161.
The Society is the UK national learned and professional society for geology with a worldwide Fellowship(FGS) of 9000. The Society has the power to confer Chartered status on suitably qualified Fellows, and about2000 of the Fellowship carry the title (CGeol). Chartered Geologists may also obtain the equivalent Europeantitle, European Geologist (EurGeol). One fifth of the Society's fellowship resides outside the UK. To find outmore about the Society, log on to www.geolsoc.org.uk.
The Geological Society Publishing House (Bath. UK) produces the Society's international journals andbooks, and acts as European distributor for selected publications of the American Association of PetroleumGeologists (AAPG), the American Geological Institute (AGI), the Indonesian Petroleum Association (IPA).the Geological Society of America (GSA), the Society for Sedimentary Geology (SEPM) and the Geologists'Association (GA). Joint marketing agreements ensure that GSL Fellows may purchase these societies* publi-cations at a discount. The Society's online bookshop (accessible from www.geolsoc.org.uk) offers secure bookpurchasing with your credit or debit card.
To find out about joining the Society and benefiting from substantial discounts on publications of GSL andother societies worldwide, consult www.geolsoc.org.uk. or contact the Fellowship Department at: TheGeological Society, Burlington House, Piccadilly, London W1J 0BG: Tel. +44 (0)20 7434 9944; Fax +44 (0)207439 8975; Email: [email protected].
For information about the Society's meetings, consult Events on www.geolsoc.org.uk. To find out more aboutthe Society's Corporate Affiliates Scheme, write to [email protected].
Published by The Geological Society from:The Geological Society Publishing HouseUnit 7, Brassmill Enterprise CentreBrassmill LaneBath BA1 3JN,UK
(Orders: Tel. +44 (0)1225 445046Fax +44 (0)1225 442836)
Online bookshop: http://bookshop.geolsoc.org.uk
The publishers make no representation, express orimplied, with regard to the accuracy of theinformation contained in this book and cannotaccept any legal responsibility for any errors oromissions that may be made.
© The Geological Society of London 2002. All rightsreserved. No reproduction, copy or transmission ofthis publication may be made without writtenpermission. No paragraph of this publication may bereproduced, copied or transmitted save with theprovisions of the Copyright Licensing Agency, 90Tottenham Court Road, London W1P 9HE. Usersregistered with the Copyright Clearance Center, 27Congress Street, Salem, MA 01970, USA: the item-fee code for this publication is 0305-8719/02/$15.00.
British Library Cataloguing in Publication DataA catalogue record for this book is available fromthe British Library.
ISBN 1-86239-106-8
DistributorsUSA
AAPG BookstorePO Box 979TulsaOK 74101-0979USA
Orders: Tel. +1 918 584-2555Fax + 1 918 560-2652E-mail [email protected]
IndiaAffiliated East-West Press PVT LtdG-l/16 Ansari Road, Daryaganj.New Delhi 110 002India
Orders: Tel. +91 11 327-9113Fax + 91 11 326-0538E-mail [email protected]
JapanKanda Book Trading Co.Cityhouse Tama 204Tsurumaki 1-3-10Tama-shiTokyo 206-0034Japan
Orders: Tel. +81 (0)423 57-7650Fax +81 (0)423 57-7651
Typeset by Type Study, Scarborough, UKPrinted by Hobbs the Printers, UK.

Contents
Preface
OWEN, A. W. & CRAME, I A. Palaeobiogeography and the Ordovician andMesozoic-Cenozoic biotic radiations
BASSETT, M. G., POPOV, L. E. & HOLMER, L. E. Brachiopods: Cambrian-Tremadocprecursors to Ordovician radiation events
HARPER, D. A. T. & MAC NIOCAILL, C. Early Ordovician rhynchonelliformeanbrachiopod biodiversity: comparing some platforms, margins and intra-oceanicsites around the lapetus Ocean
COPE, J. C. W. Diversification and biogeography of bivalves during the OrdovicianPeriod
TURVEY, S. T. Phylogeny of the Reedocalymeninae (Trilobita): implications forEarly Ordovician biogeography of Gondwana
SMITH, M. P., DONOGHUE, P. C. I & SANSOM, I. J. The spatial and temporaldiversification of Early Palaeozoic vertebrates
ARMSTRONG, H. A. & OWEN, A. W. Euconodont diversity changes in a cooling and closinglapetus Ocean
BOTTING, J. P The role of pyroclastic volcanism in Ordovician diversification
HART, M. B., OXFORD, M. J. & HUDSON, W. The early evolution and palaeobiogeographyof Mesozoic planktonic foraminifera
ABERHAN, M. Opening of the Hispanic Corridor and Early Jurassic bivalvebiodiversity
CANTRILL, D. J. & POOLE, L Cretaceous patterns of floristic change in theAntarctic Peninsula
CRAME, J. A. & ROSEN, B. R. Cenozoic palaeogeography and the rise of modernbiodiversity patterns
MARKWICK, P.J. & LUPIA, R. Palaeontological databases for palaeobiogeography,palaeoecology and biodiversity: a question of scale
MARKWICK, P.J. Integrating the present and past records of climate, biodiversityand biogeography: implications for palaeoecology and palaeoclimatology
Index
vi
1
13
25
35
53
69
85
99
115
127
141
153
169
179
201

Preface
Biodiversity change is now one of the most important topics of investigation for biogeographer andpalaeobiogeographer alike. Demonstrably, great shifts in the numbers of plant and animal taxa areoccurring on certain parts of the Earth's surface at the present day, and much effort is currentlybeing expended to determine why this should be so. Climate change in particular is held by manyto be the most likely cause of dramatic range shifts and local extinctions.
With their longer time perspective, palaeontologists are continually surveying the fossil recordfor signs of global biodiversity change in the past. Much attention has rightly been paid to studyingthe spectacular mass extinctions during the Phanerozoic, but what happens in between them? Manypalaeobiologists now believe that there may have been two pronounced intervals when life radiated(i.e. diversified) spectacularly: the Ordovician Period, and the mid-Mesozoic-Cenozoic eras. Theseepisodes mark the steepest sustained rises on the 'curve of Life through time'; the interveningSilurian-Jurassic interval is the much flatter plateau, punctuated by mass extinctions and theirrecovery intervals.
Both the scale of these spectacular diversity increases and their probable causes are currentlytopics of intense debate. To the geologist, there is an intriguing link here between dispersed conti-nents (as opposed to the presence of supercontinents), changing climates (both intervals endedwith widespread glaciations), and the proliferation of life. We now know that the accumulation ofLife on Earth is extremely complex; it is not just a matter of packing more taxa into any one habi-tat, but also of packing more habitats within a province, more provinces within a region, and moreregions within the biosphere. Were the Ordovican and mid-Mesozoic-Cenozoic two intervalsof time when there was a fundamental reorganization of biodiversity on a hierarchy of biogeo-graphical scales?
Further study of these two great evolutionary radiations by a mixed group of geologists, palaeon-tologists and biogeographers was the theme of the 2001 Lyell Meeting convened on behalf of theJoint Committee for Palaeontology representing the Palaeontological Association, GeologicalSociety of London and British Micropalaeontological Society. Entitled 'Palaeobiogeography andBiodiversity Change', it was held at Burlington House on 21 February 2001 and received generousfinancial support from the Geological Society and Palaeontological Association. Some 75 scientistsattended and contributed to a lively debate on the fundamental causes of major evolutionaryradiations. With half of the programme devoted to the Ordovician radiation, and half to theMesozoic-Cenozoic one, it was possible to compare and contrast these two great evolutionaryevents.
The results of this meeting are presented in this volume as a series of individual papers. Theserepresent by no means the last word on this important topic, but serve instead as an introductionto some of the key issues involved. In particular it is hoped that, collectively, they demonstrate howlong periods of time and plate tectonic movements can have a fundamental influence on the gener-ation and maintenance of major biodiversity patterns.
We are indebted to many colleagues for their help in refereeing the papers in the volume, includ-ing: H. A. Armstrong, R. Askin, C. Babin, M. J. Benton, M. Caron, A. Clarke, E. N. K. Clarkson,S. Damborenea, W. T. Dean, A. Hallam, R. Hill, P. Janvier, S. Lidgard, C. Little, R. A. Livermore,A. I. Miller, T. McCormick, J. A. Rasmussen, M. Romano, A. W. A. Rushton, D. J. Siveter, M. P.Smith, S. P. Tunnicliff, J. Whittaker and P. B. Wignall.
Alistair Crame and Alan Owen

Palaeobiogeography and the Ordovician and Mesozoic-Cenozoicbiotic radiations
A. W. OWEN1 & J. A. CRAME2
1 Division of Earth Sciences, University of Glasgow, Gregory Building, Lilybank Gardens,Glasgow G12 8QQ, UK (e-mail: [email protected])
2British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 0ET, UK(e-mail: A. [email protected])
Abstract: Curves of taxonomic diversity through geological time consistently show majorevolutionary radiations during the Ordovician Period and from the mid-Mesozoic to thepresent day. Both intervals were characterized by marked biotic provincialism, reflectingepisodes of major continental break-up and global tectonism, and their later historiesfeatured steep global climatic gradients. The Ordovician radiation can be recognized at awide spectrum of taxonomic levels from species to class, and the biogeographical patternsassociated with the radiation of individual clades reflect a complex combination of platedistribution, tectonic activity, sedimentary environment, sea-level rise and, ultimately,glaciation. The true scale of the mid-Mesozoic-Cenozoic biotic radiation is currently a topicof intense debate but there is no doubt that it affected plants and animals in both the marineand terrestrial realms. The role of land bridges and ocean gateways in controlling the for-mation of biodiversity patterns has been a persistent theme in Mesozoic-Cenozoic bio-geography, and a complex set of Neogene tectonic events probably aided the developmentof both latitudinal and longitudinal provinces during the Cenozoic. The present volumehighlights some of the successes across a spectrum of approaches to unravelling theOrdovician and Mesozoic-Cenozoic radiations within the context of palaeobiogeography.
' ... geologic history is the pacemaker of bio-logic diversity' (Schopf 1979, p.454)
Post-Cambrian curves of taxonomic diversitythrough geological time consistently show amajor evolutionary radiation during the Ordo-vician Period, and a second, larger one from themid-Mesozoic to the present day. Between thesesteep upward slopes the Silurian-Jurassic inter-val usually appears as a distinct plateau punctu-ated by mass extinctions and their recoveryintervals (e.g. Sepkoski 1981, 1993, 1997; Niklaset al. 1985; Padian & Clemens 1985; Benton1995, 2000, 2001; Conway Morris 1998).Although this general pattern seems to berobust, it is becoming increasingly clear that itmay include biases introduced by both thequality of the fossil record and the samplingprocedures employed. For example, Miller &Foote (1996) have shown how the use of morerigorous taxonomic and diversity metrics canalter the perception of the Ordovician radiation,and Alroy (2000; Alroy et al. 2001), using a tech-nique that standardizes sampling to absolutetime intervals, has cast doubt on the scale of themid-Cretaceous-Recent one. Potential biaseson a variety of spatial and temporal scales havebeen further highlighted by Smith (2000; Smithet al. 2001) and McCormick & Owen (2001).
Nevertheless, the general trends are stillprominent across a wide range of faunal andfloral groups (Benton 1999), and have been thefocus of intense scientific debate. The Ordo-vician radiation reflects the rise of Sepkoski's(1981) Palaeozoic Evolutionary Fauna and themid-Mesozoic to Recent one, at least in themarine realm, the full development of theModern Evolutionary Fauna. Sheehan (2001b)has recently summarized the gross ecologicalchanges associated with the rise of these Evol-utionary Faunas. The mid-Mesozoic to Recentwas also the time of spectacular radiation ofmajor terrestrial groups such as the floweringplants (angiosperms), birds and mammals(Hallam 1994). Why have there been these twogreat pulses in the diversification of life? Arethere any obvious similarities or differencesbetween them that may provide clues as to whattriggers biological diversification on a globalscale? As both these intervals were character-ized by marked biotic provincialism, reflectingepisodes of major continental break-up andglobal tectonism, it has become particularlyimportant to ask whether there may be a keylink between palaeogeographical configurationand biotic radiations. This is especially so as thelatter parts of both intervals featured steepglobal climatic gradients.
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194, 1-11.0305-8719/02/$15.00 © The Geological Society of London 2002.

2 A.W. OWEN & J. A CRAME
In comparing the Vendian-Ordovician andMesozoic marine diversifications, Erwin et al.(1987) highlighted the much lower number oftaxonomically high level (order and above)originations in the later event. They attributedthis to differences in the occupation of 'adaptivespace' with the early Phanerozoic radiationreflecting the greater opportunity for theappearance of the novel morphologies used todiagnose higher level taxa. In the presentvolume, the accent is very much on diversifica-tion at lower taxonomic levels, and the LateCenozoic-Recent interval in particular ischaracterized by the proliferation of species-richclades (Crame 2001).
It is also clear that to make headway in thestudy of taxonomic diversity patterns, on eithertemporal or spatial scales, we have to be con-sistent in what is being measured and there issome confusion in terminology within the litera-ture. Much of the rigorous definition of diversitymeasurement has been in relation to terrestrialenvironments. There are two basic categoriesof measurement: inventory diversity (sensuWhittaker 1977), that records the numbers oftaxa per unit area (and may be weighted to takeaccount of proportional abundances), and dif-ferentiation diversity that provides a measure ofdifference (or similarity) between levels ofinventory diversity. Alpha (or within-habitat)diversity is the most common form of inventorydiversity and records the number of taxa perarea of homogenous habitat and so reflectsspecies packing within a community. Beta (orbetween-habitat) diversity is the category ofdifferentiation diversity that measures the vari-ation in taxonomic composition between areasof alpha diversity (Magurran 1988). Whittaker(1977) used the terms 'gamma diversity' toreflect the number of taxa in an island ordistinctive landscape and 'epsilon diversity' forthe inventory diversity of a large biogeographicregion. Using that terminological scheme, theterm 'delta diversity' is used for the variationbetween areas of gamma diversity within an areaof epsilon diversity (Magurran 1988). However,in palaeontological analyses of marine faunas,many workers (e.g. Sepkoski 1988 and refer-ences therein), have adopted a simpler schemewhereby gamma diversity is viewed as a measureof differentiation diversity at a larger spatialscale than beta, measuring taxonomic differenti-ation between geographical regions and thus areflection of provinciality or endemicity.
Miller (e.g. 1997a, b, 1998, 2000) has con-sistently emphasized that it is essential to dissectthe global diversity curves in order for themto be understood. If the patterns can be
comprehended, then the processes that drivethem can be addressed. Even understanding thepartitioning of diversity change through theareal scales of its measurement is a significantchallenge. Thus for the Ordovician, for example,the alpha diversities of a major clade mayremain constant, in contrast to the global diver-sity change (e.g. Westrop & Adrain 1998; Adrainet al. 2000), increasing alpha diversities may beset against decreasing beta trends within apalaeogeographical region (e.g. Miller & Mao1998) and measured increases in alpha and betadiversities may be insufficient to account for thescale of global biodiversity increase (Sepkoski1988). To what extent therefore do gamma anddelta diversity levels (i.e. provinciality) hold thekey to understanding global diversity trendsthrough time?
Biogeography and biodiversity changeConnections between biodiversity change andprovinciality are well demonstrated in theliterature (see Jablonski et al. 1985 for review).Boucot (1975, see also 1983) suggested thatprovincialism may be an important factorunderlying diversity change in the Silurian andDevonian. Valentine (1973) and Schopf (1979)established a strong link between changes inendemism and the dramatic rise in taxa pro-duced by the mid-Mesozoic-Cenozoic radiation.Valentine et al. (1978) attributed high marinespecies diversity in the Cenozoic to the markedrise in provinciality and simulated Phanerozoicmarine diversity in terms of changing provincialpatterns. They suggested as much as a five-foldincrease in provinciality since the Late Palaeo-zoic, although this figure has been disputed(Bambach 1990). More recently, Smith (1988,fig. 8; see also Brenchley & Harper 1998, fig.8.13) combined the schematic representation ofthe major continental plates and global marinefamilial diversity curve of Valentine & Moores(1970, 1972) to highlight the correspondencebetween diversity peaks and plate dispersal.Such a compilation would now be equally appro-priate using Sepkoski's (1997) familial or genus-level curves.
Miller (1997c) drew attention to work byJablonski (1987) on Cretaceous molluscs andother studies that show that geographical rangeswere strongly correlated with stratigraphicaldurations, but these ranges were establishedvery early in the histories of individual species.Miller showed a general increase in thelongevity of Ordovician genera as well as anoverall range expansion and suggested that byanalogy with the Cretaceous mollusc data, the

PALAEOBIOGEOGRAPHY AND BIOTIC RADIATIONS 3
Ordovician genera may have become morespecies-rich. Geographical range is also import-ant in terms of differential survival duringextinction events. Geographically widespreadclades have a much higher probability of sur-vival, as shown for example by Westrop (1991)for the latest Cambrian, Owen & Robertson(1995) and Sheehan et al. (1996) for the end-Ordovician and Jablonski (1986) for theend-Cretaceous extinction event.
Ordovician biogeography and biodiversityIn the introductory paper to the volume arisingfrom the 1982 International Ordovician Sym-posium, Jaanusson (1984) highlighted threereasons why he considered the Ordovician to be'so special': (i) extensive development of epi-continental seas, (ii) marked changes in thecomposition of skeleton-bearing faunas conse-quent on the appearance and diversification ofmajor taxonornic groups, and (iii) pronouncedbiogeographical differentiation. The second ofthese has become the focus of considerableinternational attention in recent years, partiallyformalized under IGCP Project 410, The GreatOrdovician Biodiversification Event' (seeWebby et al. 1999). It is pertinent to assess theinfluence of the first and especially the third ofJaanusson's 'special' attributes of the Ordovi-cian to that diversification.
The Ordovician radiation can be recognizedat a wide spectrum of taxonomic levels fromspecies to class. The major component of thediversification took place in clades that typifiedthe Palaeozoic Fauna (Sepkoski 1981, 1995)both in groups that had a Cambrian history suchas rhynchonelliformean ('articulate') brachio-pods (e.g. Bassett et al. 2002) and vertebrates(e.g. Smith et al. 2002), and in groups that firstappeared during the Period (e.g. bryozoans andsome major echinoderm clades; Sepkoski 1995).Some elements of the Modern Fauna such asbivalve molluscs (e.g. Cope 2002) also radiatedat this time as did clades within groups thatcharacterize the Cambrian Fauna, in particulargroups of 'inarticulated' brachiopods (Sepkoski1995) and a substantial set of trilobite familiesconstituting the so-called 'Whiterock Fauna'(Adrain et al. 1998).
Sepkoski (1988) argued that changes in alphaand beta diversities accounted for only abouthalf of the increase in global genus-level diver-sity during the Ordovician and that much of theremaining increase was probably not attribu-table to changes reflecting provinciality orendemicity. Instead, he attributed much of theremaining diversity increase to 'hidden sources
of beta diversity' such as the appearance and/orexpansion of communities that were low inoverall extent but rich in species, such as organicreefs, hardground communities, bryozoan thick-ets and crinoid gardens. This view was endorsedby Zhuravlev (2001) in a thoughtful comparisonof the Cambrian and Ordovician radiations.Such communities reflect changes at a fairly highlevel in the ecological architecture of marine life(see Droser et al. 1997, 2000) but the lack ofcorrelation between significant biodiversitychange and palaeobiogeography during theOrdovician is surprising. Sepkoski consideredthat there was no change in the number ofprovinces from the Cambrian into the earlyOrdovician and that the degree of provincialitydecreased during late Ordovician. However, hisassessment did not take account of majorpalaeogeographical changes such as the frag-mentation of the Gondwanan margin (Dalziel1997; van Staal et al. 1998; Cocks 2001;McNamara et al. 2001, Bassett et al. 2002).Coupled with this, the plate tectonic dynamismof the Ordovician may have been crucial ingenerating hitherto unsuspected provinciality.
Miller (1997b) compared genus-level diversitycurves from six palaeocontinents using both rawand rarefied (see Miller & Foote 1996) data. Theextreme ends of the spectrum of palaeocontinen-tal variation in diversity patterns were those ofLaurentia and South China which both lay at lowlatitudes. Moreover, in terms of the partitioningof overall diversity change, Miller & Mao (1998)showed that whilst mean alpha diversitiesshowed an overall slight increase from theTremadoc to the Ashgill in both regions, therewas a fall in mean beta diversity and this wasmuch more marked in South China than inLaurentia. In itself, the fall in beta diversityshown in both regions set against rising alphadiversity is surprising, and was provisionallyexplained as reflecting the increasing environ-mental and geographical range of many generathroughout the Ordovician (see also Miller1997c), possibly along with an increase in thespecies richness of these genera. This wouldreduce the beta diversity through time asmeasured at genus level as individual generabecame more widespread causing a decrease inthe distinction between assemblages at thistaxonomic level. Miller & Mao (1998, p. 306) alsoargued that the increase in beta diversity recog-nized by Sepkoski (1988), largely on the basis ofLaurentian data, represented a net change fromthe Cambrian to the Ordovician and was not indirect contradiction of the decreasing trend thatthey recorded through the Ordovician.
In a clever reversal of Sepkoski's (1988)

4 A.W. OWEN & J. A CRAME
question about the partitioning of diversityduring the Ordovician biodiversification,Brenchley et al. (2001) recently addressed theissue of where the diversity was lost during thetwo phases of extinction at the end of thatPeriod, They argued that the considerablereduction in alpha diversity had a significantimpact on both beta diversity and provinciality,Given the speed of the extinctions, it was themajor loss of endemic taxa that significantlyreduced the number of provinces, rather thanplate movements,
Emerging patterns
The biogeographical patterns associated withthe radiation of individual clades during theEarly Palaeozoic are only just starting to cometo light and reflect a complex combination ofplate distribution, tectonic activity, sedimentaryenvironment, sea-level rise and, ultimately,climatic gradient. Smith et al. (2002) argue thatthe latest common ancestor of all 'ostracoderms'and jawed vertebrates was Laurentian but there-after the early Palaeozoic diversification of thevertebrates reflected complex biogeographicalpatterns of dispersal, vicariance and tectonicconvergence. In contrast, an 'out of Gondwana'radiation for many invertebrate groups is arecurring theme. However, the picture iscomplex as that continent extended from thesouth pole to north of the equator (Cocks 2001;Dalziel 1997) and encompassed both silici-clastic- and (at lower latitudes) carbonate-dominated shelf environments. Thus whileBassett et al. (2002) have shown that the mid-Cambrian precursors of the typical PalaeozoicFauna rhynchonelliformean ('articulate')brachiopod communities first appeared inshallow-water carbonate settings in north andeast Gondwana, the early Ordovician radiationof the bivalve molluscs took place in the shallow-water siliciclastic shelves of Gondwana (Babin1995; Cope 2002) at all latitudes. Only late in theOrdovician did bivalves become established onthe low latitude carbonate shelves of Laurentiaand Baltica. Preliminary analysis suggests thatthe rise to dominance of the Whiterock Fauna oftrilobite families (Adrain et al. 1998) took placea little earlier on the fairly high latitude silici-clastic margins of Gondwana (Owen &McCormick in press) than on low latitudeLaurentia. A cladistic analysis by Turvey (2002)of one of the groups belonging to the WhiterockFauna, the Reedocalymeninae (one of thearchetypical Gondwanan groups), points to aseries of biogeographical events affecting largeareas of that plate during the early Ordovician.
The nature of the depositional environmentclearly played a significant role in the diversifi-cation of clades during the Ordovician. Miller's(1997b) preliminary analysis of diversity curvesfrom six palaeocontinents identified differencesin the scale and timing of changes both in termsof total diversity and that of individual majorclades representing all three EvolutionaryFaunas. These differences were linked to theamount and timing of siliciclastic input, includ-ing the effects of uplift caused by orogeny andvolcanism (see also Miller & Mao 1995, 1998),and therefore emphasize the importance of platetectonic activity to the promotion of biodiversitychange. Miller (1997b) suggested that the spreadof siliciclastic sediment after the TaconicOrogeny triggered the flourishing of bivalves inLaurentia following their earlier radiation insiliciclastic sedimentary environments inGondwana,
The Taconic Orogeny and the resultantincrease in weathering of siliceous rocks in thelate Ordovician may also have been responsiblefor the profound change in climate leading to theHirnantian glaciation (Kump et al. 1999). Thisvery short-lived glaciation during what wasessentially a greenhouse world was the proximalcause of the complex series of events that pro-duced the end-Ordovician mass extinctions (seereviews in Brenchley et al. 2001; Sheehan 2001a).The climatic decline (see Armstrong & Coe1997; Kump et al. 1999) that led up to it may alsohave had some effect on biodiversity. Cope(2002) recognized an increase in bivalveendemicity as climatic gradients steepenedduring the late Ordovician, An analysis ofconodont biofacies by Armstrong & Owen(2002) suggests that diversity fell in lowlatitude Laurentia coincident with the onset ofocean cooling in the early Ashgill, whereas onAvalonia the decline took place a little laterwhen that microcontinent drifted northwardsinto tropical latitudes.
Plate tectonic activity clearly played a signifi-cant role in controlling biodiversity changeduring the Ordovician. On a local scale, but withpossible wider consequence, Botting (2002) hasprovided a possible mechanism for diversifica-tion as a result of repeated volcanic activitywhereby ash falls produce overturn of thestratified watermass. This overturn could causeblooms in the plankton and benthos and thegeneration of localized, temporary benthicIslands' within the otherwise dysaerobic shelfwhich were the sites of high speciation rates. Ona much larger scale, Bassett et al. (2002) linkthe early and mid-Ordovician spread anddiversification of the rhynchonelliformean

PALAEOBIOGEOGRAPHY AND BIOTIC RADIATIONS 5
brachiopods typical of the Palaeozoic Evol-utionary Fauna to a combination of global sea-level rise and fragmentation of the Gondwananmargin. Harper & MacNiocaill (2002) reach asimilar conclusion and also demonstrate theimportance of intra-oceanic sites in the develop-ment, divergence and survival of taxa.
Mesozoic-Cenozoic biogeography andbiodiversityThe true scale of the mid-Mesozoic-Cenozoicbiotic radiation is currently a topic of intensedebate. Whereas a new compilation of genericrather than familial data suggests that it may nothave been as marked as Sepkoski (1981, 1993)originally indicated (Alroy et al. 2001), adetailed taxonomic investigation of just onesmall part of the Late Cenozoic tropics suggeststhat it was considerably greater (Jackson &Johnson 2000, 2001). One thing that is beyonddispute about this radiation is that it affectedplants and animals in both the marine andterrestrial realms. The spectacular radiation ofthe angiosperms, mammals and certain reptilegroups (such as the snakes) on land is matchedby that of the planktonic foraminifera, neo-gastropods, heteroconch bivalves, cheilostomebryozoans, decapod crustaceans and teleost fishin shallow seas (Hallam 1994). This co-radiationof such disparate taxa is often taken as strongevidence for a real evolutionary phenomenon(Signor 1990; Benton 1999). This was also thetime when some of the largest (in terms ofspecies richness) clades evolved. For example,some of the massive eudicot angiosperm cladeshave between 20 000 and 25 000 extant taxa, andboth the neogastropod subfamily Coninae andthe heteroconch bivalve family Veneridae haveapproximately 500 living species each (Crame2001).
Some geographical constraints on theradiation
It now seems almost certain that the extantplanktonic formainifera evolved from benthonicancestors in Tethyan (i.e. tropical/subtropical)facies in the Early Jurassic (Toarcian) (Hart etal. 2002). A biostratigraphic analysis hasindicated some expansion within the NorthAtlantic-European region in both theBajoican-Bathonian and Aptian, but it was notuntil the latest Albian that a near-global distri-bution was achieved. There is a strong linkhere between phases of diversification asrevealed by stratigraphic analysis, continental
fragmentation, and the expansion of shelf seas.Interestingly enough, it would appear thatlatitudinal gradients in taxonomic diversity hadbeen established by the Campanian-Maastricht-ian (Huber 1988), and these have persistedthrough to the present day (Be 1977).
The role of land bridges and ocean gatewaysin controlling the formation of biodiversitypatterns has been a persistent theme in Meso-zoic-Cenozoic biogeography (e.g. Hallam1981). Have they, perhaps, created bottlenecksin the distribution of biotas that are largeenough to be discernible in the fossil record?This forms the central theme in the papers byAberhan (2002) and Cantrill & Poole (2002).and is discussed in a more general way by Crame& Rosen (2002).
In his study of Early Jurassic bivalve biodi-versity, Aberhan (2002) has been particularlyconcerned with the role of the Hispanic Corri-dor, a putative narrow seaway across centralPangaea linking the eastern Pacific with westernTethys. Was there a preferential extinction ofendemic South American taxa across thePliensbachian-Toarcian boundary due to asudden influx of cosmopolitan Tethyan forms,or was a renewed rise in late Toarcian-Aalenianspecies diversity in NW Europe attributable toan influx from Andean South America in theother direction? Using a comprehensivespecies-level database, Aberhan is able todemonstrate that, in fact, in both regions earlyPliensbachian to Aalenian immigration ratesthrough the corridor remained low. In compari-son, the within-region origination of newspecies played a much more important role inthe recovery of post-extinction faunas. Therelative importance of immigration versus insitu radiation in the generation of large-scalebiodiversity patterns throughout the fossilrecord is still a controversial issue (e.g. Vermeij1993).
By the time of radiation of crown groupangiosperms (flowering plants) in the EarlyCretaceous, the major southern hemispherecontinents had largely separated from oneanother and Antarctica was rapidly becomingisolated. It has long been thought that, byAptian-Albian times, the only major terrestrialconnection between western Gondwana (SouthAmerica and Africa) and eastern Gondwana(India, Antarctica and Australasia) was thevolcanic archipelago of the Antarctic Peninsularegion. However, in a new analysis of southern,high latitude Cretaceous floras, Cantrill & Poole(2002) cast some doubt on this theory. Whereasthe earliest known angiosperms from theAntarctic Peninsula are Early Albian in age,

6 A.W. OWEN & J. A CRAME
there are angiospermous pollen grains fromAustralia that are Early Aptian. Quite what thealternative immigration route may have been isuncertain, but Cantrill & Poole point to theproven existence of Turonian terrestrial sedi-ments on the Kerguelen Plateau (ShipboardScientific Party 2000) and Late Cretaceousdinosaur finds that link Madagascar, SouthAmerica and India (Sampson et al. 1998). It isclear that the earliest Antarctic angiospermsoccupied areas of disturbance and were under-storey colonizers. Their major pulse of radiationin the Turonian was coincident with the acme ofCretaceous global warmth.
The relative importance of tectonics andclimate
In a wide-ranging review, Crame & Rosen(2002) take as their starting point the majorextant biodiversity patterns, and then workback through time. They address the reason forthere being discrete tropical high-diversity fociin both the marine and terrestrial realms, withsteep latitudinal diversity gradients leadingaway from them. Whatever the ultimate causeof tropical high diversity per se, it is clear that acritical sequence of Early Miocene (c. 20 Ma)tectonic events fractured a once homogeneoustropical biota into several distinct centres;the Indo-West Pacific (IWP) and theAtlantic-Caribbean-East Pacific (ACEP) arethe most important of these in the marinerealm. Together with the physical isolation ofAntarctica, these same plate tectonic move-ments contributed significantly to the globalreorganization of major ocean current systemsand Cenozoic cooling. This in turn led to amarked increase in provincialism and bioticdifferentiation on a regional scale. Even so,something else may have been involved in thedevelopment of the steepest diversity gradientsduring the Late Cenozoic. Rates of originationmay have been driven by an increase inMilankovitch cyclicity over the last 10-15 Maand there may be some crucial links herebetween the geographical range of an organismand its propensity to allopatric speciation.However, the issue is a complex one and is yetto be fully resolved. The more frequent natureof climatic change in high latitude regions mightmean that ecological isolates are less likely topersist or, alternatively, the more constantenvironment of the tropics could mean thatranges can be more easily split by slighttemperature shifts (Chown & Gaston 2000;Dynesius & Jansson 2000).
The importance of databases in biodiversitystudies
With the widespread availability of relationaldatabase and GIS software, it is now much easierto build a comprehensive biodiversity databaseon a desktop computer. However, in a methodo-logical review of just how such a database shouldbe constructed, Markwick & Lupia (2002)emphasize the need to take into account varia-tions in scale, inconsistency in the data, andpotential errors (inaccuracy). Scale is a particu-larly critical issue, for, whereas 'large-scale' to abiologist refers to a large area or duration, a'large-scale map' is usually one of great detailbut small area. As this can lead to confusionwhen using a GIS for biological or palaeobio-logical purposes, Markwick & Lupia advocatethe adoption of two terms from landscapeecology. The first of these is 'grain', which is theminimum resolution or scale of an observation,and the second is 'extent', which is the totalamount of space or time observed. Thus, a large-scale map is fine-grained but of limited extent. Itis the heteorogeneities in scale (grain) in thefossil record that are potentially the majorcauses of problems in the construction ofpalaeontological databases.
Markwick (2002) goes on to show how a com-prehensive database of Recent North American,European, South African and Australian non-avian tetrapods can be used to tackle somefundamental problems in the study of latitudinaldiversity gradients. Pooling taxa from all theseregions it can be shown that there is a strong,monotonic relationship with both absolutelatitude and mean annual temperature (MAT);the highest diversities are clearly associated withthe highest MATs and lowest latitudes. Never-theless, it is clear that there are strong differencesin the form of the latitudinal gradients betweenthe ectotherms (reptiles and amphibians) andendotherms (mammals). Whereas the formershow comparatively simple, steep gradients,reflecting their dependence on the immediateenvironment, the latter show a more complexpattern, based on an indirect dependence onthe environment (through various feedingstrategies). Markwick concludes that patterns ofterrestrial species diversity are not simply a func-tion of the amount of available energy, but howthat energy is procured. As the proportion ofectotherms to endotherms shows a strong linearrelationship to temperature, it can be used toretrodict MATs in the fossil record. An exampleis given for the Middle Eocene Messel fauna ofGermany that shows close agreement with otherpalaeotemperature estimates.

PALAEOBIOGEOGRAPHY AND BIOTIC RADIATIONS 7
Synopsis
The Ordovician Period witnessed the rise ofPalaeozoic Evolutionary Fauna, comprisingelements such as the rhynchonelliformeanbrachiopods, bryozoans, echinoderms, primitivevertebrates and many families of trilobites. TheMesozoic-Cenozoic radiation includes the riseof such groups as the flowering plants, birds,mammals, marine molluscs, teleost fish anddecapod crustaceans. Full substantiation ofthese patterns remains a major task and willinvolve a massive coordinated effort in taxo-nomic palaeontology (Kerr 2001; Jackson &Johnson 2001).
If these two steep increases in taxonomicdiversity are not artefacts of the fossil record, achallenge of equal dimensions is to establishwhat has been driving them. One of the simplestexplanations to consider is that they representtwo intervals in which there were fundamentalincreases in within-habitat species richness (i.e.alpha diversity). In a seminal study of Phanero-zoic marine communities, Bambach (1977)suggested that there may well have been peri-odic expansion of available ecospace throughthe development of new or increased resourcesupplies (see also Bambach 1993). Rosenzweig(1995, p. 306), for example, has suggested thatthe Ordovician was the first time in Earth historythat muddy bottoms were extensively colonizedby marine organisms. Precisely why this mayhave been so is uncertain, but dissolved oxygenlevels may have increased until they reachedsome sort of threshold and allowed widespreadcolonization. Similarly, Bambach (1977, 1993)linked the late Mesozoic-Cenozoic rise ofboth terrestrial and marine organisms to theblossoming of the angiosperms. Here was anabundant source of both new food and habitatspace for a wide variety of organisms. Inaddition, Bambach (1977) pointed to both thelate Ordovician and Cenozoic glaciations as apossible source of oceanic nutrient recycling.Changes in thermohaline circulation broughtabout by climate change could have led to sub-stantial oceanic mixing, which in turn affectedthe food resources of shelf seas. In bothinstances though, the global diversity increasewas well underway and in the case of the Ordo-vician may have already reached its plateau bythe time such processes began.
In contrast to attempts to seek a limitednumber of global controls on biodiversitychange, Miller (e.g. 2000) has argued that it ismore realistic to try to understand the regionalpatterns and the processes likely to have driventhem. None the less, some form of periodic
increase in resource supply could lead to a rapidexpansion in the numbers of taxa within somehabitats and regions. Once an adaptive thresh-old has been breached it may be possible to packmore taxa into a locality or region within acomparatively short space of time. Patterns ofalpha and gamma diversity could be expected toincrease periodically rather than continuously.The concept of non-hierarchical, non-additivelevels of ecological change introduced by Droseret al (1997, see also 2000) promises to be a usefulway of addressing step changes in biodiversityand/or the utilization of ecospace, the two notnecessarily changing in tandem.
However, it is clear that something other thana sheer rise in numbers must be involved. Thereis abundant evidence that differentiation ofbiotas has occurred and on a variety of geo-graphical scales. For example, in their review ofthe Cenozoic diversification event, Crame &Rosen (2002) indicate that a complex set ofNeogene tectonic events probably aided thedevelopment of both latitudinal and longitudinalprovinces. There are indications to suggest thatbetween-habitat diversity (i.e. beta diversity)increases in tandem with alpha diversity but aswell as this, between-community and between-province diversity must be increasing too. Simi-larly Harper & MacNiocaill (2002) argue that thebreak-up of the Gondwanan margin promoted arise in between-province diversity amongst therhynchonelliformean brachiopods and thisbreak-up, together with sea-level rise that pro-moted migration over the continental shelves,also produced a rise in alpha diversities as estab-lished communities were augmented by immi-grants. Subsequent to this, beta diversities rose ascommunities became increasingly developed indeeper water environments. This pattern ofnearshore innovation and offshore expansion ofcommunities at the expense of the existing com-munity types during the Ordovician was docu-mented by Jablonski et al. (1983; see also Bassettet al. 2002). However, Westrop & Adrain (1998)and Adrain et al. (2000) have also demonstratedthat for the trilobites, alpha diversities remainedremarkably constant across the shelf throughoutthe Period and so, whilst their diversity relativeto the rapidly diversifying clades might havedeclined, they were not simply being displacedby them and 'pushed' into deeper water refuges.Jablonski & Bottjer (1991) also showed that theorigins of post-Palaeozoic benthonic orders werelargely in nearshore environments, but Jacobs &Lindberg (1998) have argued that this onlyapplied prior to the Turonian after which off-shore bottom waters became more widely oxicand amenable to originations. That change also

8 A.W. OWEN & J. A CRAME
marked the demise of dysoxic faunas, theexpansion of deep burrowers into offshoreenvironments and the offshore expansion ofbryozoan diversity.
Coupled with any purely tectonic definition ofprovinces, it is clear that biotic differentiationwas assisted by the steepened latitudinal tem-perature gradients that became establishedtowards the end of each of these intervals. Thestudies by Cope (2002), Armstrong & Owen(2002), Harper & MacNiocall (2002), Crame &Rosen (2002), Cantrill & Poole (2002) andMarkwick (2002) all emphasize that climatechange has assisted range shifts into higher orlower latitudes. The differentiation of biotasalong environmental gradients undoubtedlyenhanced the process of global biodiversifi-cation during both the Ordovician and Meso-zoic-Cenozoic. In the case of the latter, there arestill areas of the Earth's surface, such as the veryhighest and lowest latitudes, about which verylittle is known, which must bias both theobserved patterns and their interpretations.Tropical systems are particularly poorly knownfor many intervals in time, and one can onlyguess at how much 'hidden diversity' is lockedup within them (Sepkoski 1988; Jackson &Johnson 2001).
Clearly, there is still a great deal to learn aboutthe precise mechanisms underpinning the majorbiotic radiations and it is unlikely that they willbe attributable to a single, simple cause. Thepalaeobiologist is uniquely placed to study theinteraction of physical and biological forces thathave shaped the major patterns of life on Earthand understanding the palaeobiogeographicalcontext is crucial. It encompasses a multitude ofcontrols on the distribution and ecology oforganisms, from the location of continents,islands and oceans to the global climate. Import-antly, especially from a geological perspective, itrepresents a series of very dynamic systems. Thepresent volume reflects some of the wide spec-trum of approaches to studying biodiversitychange on a variety of spatial and temporalscales, from taxonomy to the analysis of largedatabases, and emphasizes their links to thecomponents of these systems in the two mostsustained episodes of overall biotic radiation inthe Phanerozoic.
We thank the referees, E. Clarkson and P. Wignall fortheir helpful comments. A.W.O. acknowledgessupport from NERC Grant GR3/11834 and this paperis a contribution to IGCP 410 'The Great OrdovicianBiodiversification Event'. J.A.C. is supported by BASCore Project, 'Antarctic marine biodiversity: ahistorical perspective'.
References
ABERHAN, M. 2002. Opening of the Hispanic Corridorand Early Jurassic bivalve biodiversity. In: CRAME.J. A. & OWEN. A. W. (eds) Palaeobiogeography andBiodiversity Change: the Ordovician and Meso-zoic-Cenozoic Radiations. Geological Society.London, Special Publications, 194. 127-139.
ADRAIN, J. M.. FORTEY, R. A. & WESTROR S. R. 1998.Post-Cambrian trilobite diversity and evol-utionary faunas. Science, 280. 1922-1925.
ADRAIN, J. M.. WESTROR S. R.. CHATTERTON. B. D. E. &RAMSKOLD. L. 2000. Silurian trilobite alphadiversity and the end-Ordovician mass extinction.Paleobiology, 26, 625–464.
ALROY, J. 2000. Successive approximations of diversitycurves: ten more years in the library. Geohgy. 28.1023-1026.
ALROY, J., MARSHALL. C. R., BAMBACH. R. K.. ET AL.2001. Effects of sampling standardization on esti-mates of Phanerozoic marine diversification. Pro-ceedings of the National Academy of Science of theUSA, 98. 6261-6266.
ARMSTRONG. H. A. & COE. A. L. 1997. Deep-seasediments record the geophysiology of the lateOrdovician glaciation. Journal of the GeologicalSociety, London. 154, 929-943.
ARMSTRONG, H. A. & OWEN. A. W. 2002. Euconodontdiversity changes in a cooling and closing IapetusOcean. In: CRAME. J. A. & OWEN, A. W. (eds)Palaeobiogeography and Biodiversity Change: theOrdovician and Mesozoic-Cenozoic Radiations.Geological Society. London, Special Publications,194, 85-98.
BABIN. C. 1995. The initial Ordovician mollusc radia-tions on the western Gondwanan shelves. In:COOPER, J. D., DROSER, M. L. & FINNEY. S. C. (eds)Ordovician Odyssey: Short Papers for the SeventhInternational Symposium on the OrdovicianSystem. SEPM Pacific Section. Fullerton Cali-fornia, 491-498.
BAMBACH, R. K. 1977. Species richness and marinebenthis habitats through the Phanerozoic. Paleo-biology, 3, 152-167.
BAMBACH, R. K. 1990. Late Palaeozoic provinciality inthe marine realm. In: MCKERROW. W. S. &SCOTESE, C. R. (eds) Palaeozoic Palaeogeographyand Biogoegraphy. Geological Society. London,Memoir, 12, 307-323.
BAMBACH, R. K. 1993. Seafood through time: changesin biomass, energetics and productivity in themarine ecosystem. Paleobiology, 19, 372-397.
BASSETT, M. G, POPOV, L. E. & HOLMER. L. E. 2002.Brachiopods: Cambrian-Tremadoc precursors toOrdovician radiation events. In: CRAME. J. A. &OWEN, A. W. (eds) Palaeobiogeography and Bio-diversity Change: the Ordovician and Meso-zoic-Cenozoic Radiations. Geological Society.London, Special Publications. 194. 13-23.
BE, A. W. H. 1977. An ecological, zoogeographic andtaxonomic review of recent planktonicforaminifera. In: RAMSAY, A. T. S. (ed.) OceanicMicropalaeontology Volume 1. Academic Press.London, 1-88.

PALAEOBIOGEOGRAPHY AND BIOTIC RADIATIONS 9
BENTON, M. J. 1995. Diversification and extinction inthe history of life. Science, 268, 52-58.
BENTON, M. J. 1999. The history of life: large data-bases in palaeontology. In: HARPER, D. A. T. (ed.)Numerical Palaeobiology, Wiley, Chichester,249-283.
BENTON, M. J. 2000. Vertebrate Palaeontology. Black-well Science, Oxford.
BENTON, M. J. 2001. Biodiversity on land and in thesea. Geological Journal, 36, 211-230.
BOTTING, J. P. 2002. The role of pyroclastic voicanismin Ordovician diversification. In: CRAME, J. A. &OWEN, A. W. (eds) Palaeobiogeography andBiodiversity Change: the Ordovician and Meso-zoic-Cenozoic Radiations. Geological Society,London, Special Publications, 194, 99-113.
BOUCOT, A. J. 1975. Evolution and Extinction RateControls. Elsevier, Amsterdam.
BOUCOT, A. J. 1983. Does evolution take place in anecological vacuum? II. Journal of Paleontology,57, 1-30.
BRENCHLEY, P. J. & HARPER, D. A. T. 1998. Palaeo-ecology: Ecosystems, Environments and Evol-ution, Chapman & Hall, London.
BRENCHLEY, P. J., MARSHALL, J. D. & UNDERWOOD,C. J. 2001. Do all mass extinctions representan ecological crisis? Geological Journal, 36,329-340.
CANTRILL, D. J. & POOLE, I.2002. Cretaceous patternsof floristic change in the Antarctic Peninsula. In:CRAME, J. A. & OWEN, A. W. (eds) Palaeobiogeog-raphy and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. GeologicalSociety, London, Special Publications, 194,141-152.
CHOWN, S. L. & GASTON, K. J. 2000. Areas, cradlesand museums: the latitudinal gradient in speciesrichness. Trends in Ecology & Evolution, 15,311-315.
COCKS, L. R. M. 2001. Ordovician and Silurian globalgeography. Journal of the Geological Society,London, 158, 197-210.
CONWAY MORRIS, S. 1998. The evolution of diversity inancient ecosystems: a review. PhilosophicalTransactions of the Royal Society of London, B,353, 327-345.
COPE, J. C. W. 2002. Diversification and biogeographyof bivalves during the Ordovician Period. In:CRAME,J. A. & OWEN, A. W. (eds) Palaeobiogeog-raphy and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. GeologicalSociety, London, Special Publications, 194, 35-52.
CRAME, J. A. 2001. Taxonomic diversity gradientsthrough geological time. Diversity and Distri-butions, 7, 175-189.
CRAME, J. A. & ROSEN, B. R. 2002. Cenozoic palaeo-geography and the rise of modern biodiversitypatterns. In: CRAME, J. A. & OWEN, A. W. (eds)Palaeobiogeography and Biodiversity Change:the Ordovician and Mesozoic-Cenozoic Radia-tions. Geological Society, London, Special Publi-cations, 194, 153-168.
DALZIEL, I. W. D. 1997. Neoproterozoic-Paleozoicgeography and tectonics: review, hypothesis,
environmental speculation. Geological Society ofAmerica Bulletin, 109, 16-42.
DROSER, M. L., BOTtjER, D. J. & SHEEHAN, P. M. 1997.Evaluating the ecological architecture of majorevents in the Phanerozoic history of marine inver-tebrate life. Geology, 25, 167-170.
DROSER, M. L., BOTTJER, D. X, SHEEHAN, P. M. &McGEE, J. R. JR. 2000. Decoupling of taxonomicand ecological severity of Phanerozoic marinemass extinctions. Geology, 28, 675-678.
DYNESIUS, M. & JANSSON, R. 2000. Evolutionary con-sequences of changes in species' geographicaldistributions driven by Milankovitch climateoscillations. Proceedings of the National Academyof Sciences USA, 97, 9115-9120.
ERWIN,D. H., VALENTINE, J.W.& SEPKOSKI,J.J. 1987. Acomparative study of diversification events: theearly Paleozoic versus the Mesozoic. Evolution.41, 1177-1186.
HALLAM, A. 1981. Relative importance of platemovements, eustasy and climate in controllingmajor biogeographical changes since the EarlyMesozoic. In: NELSON, G. & ROSEN, D. E. (eds)Vicariance Biogeography. A Critique. ColumbiaUniversity Press, New York, 303-330.
HALLAM, A. 1994. An Outline of Phanerozoic Bio-geography. Oxford University Press, Oxford.
HARPER, D. A. T. & MAC NIOCAILL, C. 2002.Early Ordovician rhynchonelliformean brachio-pod biodiversity: comparing some platforms,margins and intra-oceanic sites around thelapetus Ocean. In: OWEN, A. W. & CRAME, J. A.(eds) Palaeobiogeography and BiodiversityChange: the Ordovician and Mesozoic-CenozoicRadiations. Geological Society, London, SpecialPublications, 194, 25-34.
HART, M. B., OXFORD, M. J. & HUDSON, W. 2002. Theearly evolution and palaeobiogeography of Meso-zoic planktonic foraminifera. In: OWEN, A. W. &CRAME, J. A. (eds) Palaeobiogeography andBiodiversity Change: the Ordovician and Meso-zoic-Cenozoic Radiations. Geological Society.London, Special Publications, 194, 115-125.
HUBER, B. T. 1988. Upper Campanian - Paleoceneforaminifera from the James Ross Island region,Antarctic Peninsula. In: FELDMANN, R. M. &WOODBURNE, M. O. (eds) Geology and Paleon-tology of Seymour Island, Antarctic Peninsula.Geological Society of America, Memoir, 169,163-252.
JAANUSSON, V. 1984. What is so special about theOrdovician? In: BRUTON, D. L. (ed.) Aspects of theOrdovician System. Palaeontological Contri-butions from the University of Oslo, 295, Univer-sitetsforlaget, Oslo, 1-3.
JABLONSKI, D. 1986. Background and mass extinctions:the alternation of macroevolutionary regimes.Science, 231, 129-133.
JABLONSKI, D. 1987. Heritability at the species level:analysis of geographic ranges of Cretaceous mol-lusks. Science, 238, 360-363.
JABLONSKI, D. & BOTTJER, D. J. 1991. Environmentalpatterns in the origins of higher taxa: the post-Paleozoic marine record. Science, 252, 1831-1833.

10 A.W. OWEN & J. A CRAME
JABLONSKI, D., SEPOSKI, J. J., BOTTJER, D. J. &SHEEHAN. P. M. 1983. Onshore-offshore patternsin the evolution of Phanerozoic shelf communi-ties. Science, 222, 1123-1125.
JABLONSKI, D., FLESSA, K. W. & VALENTINE, J. W.1985. Biogeography and paleobiology. Paleobiol-ogy, 11, 75-90.
JACKSON, J. B. C. & JOHNSON, K. G. 2000. Life in thelast few million years. In: ERWIN, D. H. & WING. S.L. (eds) Deep Time: Paleobiology's Perspective.Paleobiology, Supplement, 26(4). 221-235.
JACKSON, J. B. C & JOHNSON. K. G. 2001. Measuringpast biodiversity. Science, 293, 2402-2404.
JACOBS. D. K. & LINDBERG, D. R. 1998. Oxygen andevolutionary patterns in the sea: onshore/offshoretrends and recent recruitment of deep sea faunas.Proceedings of the National Academy of Scienceof the USA, 95, 9396-9401.
KERR, R. A. 2001. Putting limits on the diversity of life.Science, 292, 1481.
KUMP, L. R., ARTHUR, M. A., PATZKOWSKY, M. E.,GIBBS, M. T., PINKUS, D. S. & SHEEHAN, P. M. 1999.A weathering hypothesis for glaciation at highatmospheric pCO2 during the Late Ordovician.Palaeogeography, Palaeoclimatologv, Palaeo-ecology, 152, 173-187.
MCCORMICK, T. & OWEN, A. W. 2001. Assessing trilo-bite biodiversity change in the Ordovician of theBritish Isles. Geological Journal, 36, 279-290.
MCNAMARA, A. K., MAC NIOCAILL, C, VAN DERPLUIJM, B. A. & VAN DER Voo, R. 2001. WestAfrican proximity of the Avalon terrane in thelatest Precambrian. Geological Society ofAmerica Bulletin, 113, 1161-1170.
MAGURRAN, A. E. 1988. Ecological Diversity and itsMeasurement. Croom Helm, London.
MARKWICK, P. J. 2002. Integrating the present andpast records of climate, biodiversity and bio-geography: implications for palaeoecology andpalaeoclimatology. In: CRAME, J. A & OWEN, A.W. (eds) Palaeobiogeography and BiodiversityChange: the Ordovician and Mesozoic-CenozoicRadiations. Geological Society, London, SpecialPublications, 194, 179-199.
MARKWICK, P. J. & LUPIA, R. 2002. Palaeontologicaldatabases for palaeobiogeography, palaeo-ecology and biodiversity: a question of scale. In:CRAME, J. A. & OWEN, A. W. (eds) Palaeobiogeog-raphy and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. GeologicalSociety, London, Special Publications, 194,169-178.
MILLAR, A. I. 1997a. Dissecting global diversitypatterns. Annual Review of Ecology and System-atics, 28, 85-104.
MILLAR, A. I. 1997b. Comparative diversificationdynamics among palaeocontinents during theOrdovician radiation. Geobios, 20, 397-406.
MILLAR, A. I. 1997c. A new look at age and area:the geographical and environmental expansion ofgenera during the Ordovician. Paleobiology, 23,410-419.
MILLAR, A. I. 1998. Biotic transitions in globalmarine diversity. Science, 281, 1157-1160.
MILLAR. A. I. 2000. Conversations about Phanerozoicglobal diversity. In: ERWIN. D. H. & WING. S. L.(eds) Deep Time: Paleobiology's Perspective.Paleobiology, Supplement. 26(4) 53-73.
MILLAR. A. I. & FOOTE. M. 1996. Calibrating theOrdovician radiation of marine life: implicationsfor Phanerozoic diversity trends. Paleobiology.22, 304-309.
MILLAR, A. I. & MAO, SHUGUANG. 1995. Associationof orogenic activity with the Ordovician radiationof marine life. Geology, 23. 305-308.
MILLAR. A. I. & MAO, SHUGUANG, 1998. Scales ofdiversification and the Ordovician radiation. In:MCKINNEY. M. L. & DRAKE. J. A. (eds) Bio-diversity Dynamics: Turnover of Populations,Taxa and Communities. Columbia UniversityPress, New York. 288-310.
NIKLAS. K. J., TIFFNEY, B. H. & KNOLL. A. H. 1985.Patterns in vascular land plant diversification: ananalysis at the species level. In: VALENTINE. J.W. (ed.) Phanerozoic Diversity Patterns: Profilesin Macroevolution. Princetown University Press.Princetown. 97-128.
OWEN. A.W. & MCCORMICK. T. (in press). Ordoviciantrilobite biodiversity change in the Anglo-Welshsector of Avalonia - a comparison of regional andglobal patterns. Special Papers in Palaeontology.
OWEN, A. W. & ROBERTSON. D. B. R. 1995. Ecologicalchanges during the end-Ordovician extinction.Modern Geology, 20, 21-39.
PADIAN, K. & CLEMENS, W. A. 1985. Terrestrial verte-brate diversity: episodes and insights. In: VALEN-TINE, J. W. (ed.) Phanerozoic Diversity Patterns.Profiles in Macroevolution. Princetown Uni-versity Press. Princetown. 41-96.
ROSENZWEIG, M. L. 1995. Species Diversity in Spaceand Time. Cambridge University Press. Cam-bridge.
SAMPSON, S. D.. WITMER, L. M.. FORSTER. C. A..KRAUSE. D. W., O ' C O N N O R , P. M.. DODSON, P. &RAVOAVY, F. 1998. Predatory dinosaur remainsfrom Madagascar: implications for theCretaceous biogeography of Gondwana. Science,280, 1048-1051.
SCHOPF, T. J. M. 1979. The role of biogeographicalprovinces in regulating marine faunal diversitythrough geologic time. In: GRAY, J. & BorcoT. A.J. (eds) Historical Biogeography Plate Tectonics,and the Changing Environment. Oregon StateUniversity Press. Corvallis, 449-457.
SEPKOSKI. J. J. 1981. A factor analytical description ofthe Phanerozoic marine fossil record. Paleobiol-ogy, 7, 36-53.
SEPKOSKI. J. J. 1988. Alpha, beta, or gamma: wheredoes all the diversity go? Paleobiology, 14.221-234.
SEPKOSKI, J. J. 1993. Ten years in the library: newdata confirm paleontological patterns. Paleobiol-ogy, 19. 43-51.
SEPKOSKI. J. J. 1995. The Ordovician radiations:diversification and extinction shown by globalgenus-level taxonomic data In: COOPER. J. D..DROSER, M. L. & FINNFY. S. C. (eds) Ordovi-cian Odyssey: Short Papers for the Seventh

PALAEOBIOGEOGRAPHY AND BIOTIC RADIATIONS 11
International Symposium on the OrdovicianSystem. SEPM Pacific Section, Fullerton, Cali-fornia, 393-396.
SEPKOSKI, J. J. 1997. Biodiversity: past, present, andfuture. Journal of Paleontology, 71, 533-539.
SHEEHAN, P. M. 2001a. The late Ordovician massextinction. Annual Review of Earth and PlanetarySciences, 29, 331-364.
SHEEHAN, P. M. 2001b. History of marine biodiversity.Geological Journal, 36, 231-249.
SHEEHAN, P. M., COOROUGH, P. J. & FASTOVSKY, D. E.1996. Biotic selectivity during the K/T and LateOrdovician extinction events. In: RYDER, G.,FASTOVSKY, D. E. & GARTNER, S. (eds) TheCretaceous-Tertiary Event and other Catastro-phies in Earth History. Geological Society ofAmerica, Special Papers, 307, 477-489.
SHIPBOARD SCIENTIFIC PARTY, 2000. Leg 183 summary:Kerguelen Plateau - Broken Ridge - A largeigneous province. In: COFFIN, M. F., FREY,F. A., WALLACE, P. J. et al. (eds) ProceedingsOcean Drilling Program, Initial Reports, 183,1-101.
SIGNOR, P. W. 1990. The geologic history of diversity.Annual Reviews of Ecology and Systematics, 21,509-539.
SMITH, A. B. 2000. Large-scale heterogeneity of thefossil record: implications for Phanerozoic bio-diversity studies. Philosophical Transactions ofthe Royal Society of London, B, 356, 351-367.
SMITH, A. B., GALE, A. S. & MONKS, N. E. A. 2001. Sea-level change and rock-record bias in theCretaceous: a problem for extinction and bio-diversity studies. Paleobiology, 27, 241-253.
SMITH, M. P., DONOGHUE, P. C. J. & SANSOM, I. J.2002. The spatial and temporal diversificationof Early Palaeozoic vertebrates. In: CRAME,J. A. & OWEN, A. W. (eds) Palaeobio-geography and Biodiversity Change: the Ordovi-cian and Mesozoic-Cenozoic Radiations.Geological Society, London, Special Publications,194, 69-83.
SMITH, P. L. 1988. Palaeobiogeography and platetectonics. Geoscience Canada, 15, 261-279.
TURVEY, S. T. 2002. Phylogeny of the Reedocaly-meninae (Trilobita): implications for EarlyOrdovician biogeography of Gondwana. In:
CRAME, J. A. & OWEN, A. W. (eds) Palaeo-biogeography and Biodiversity Change:the Ordovician and Mesozoic-CenozoicRadiations. Geological Society, London, SpecialPublications, 194, 53-68.
VALENTINE, J. W. 1973. Evolutionary Paleoecology ofthe Marine Biosphere. Prentice-Hall, EnglewoodCliffs, NJ.
VALENTINE, J. W. & MOORES, E. M. 1970. Plate tec-tonic regulation of biodiversity and sea level: amodel. Nature, 228, 657-659.
VALENTINE, J. W. & MOORES, E. M. 1972. Globaltectonics and the fossil record. Journal ofGeology, 80, 167-184.
VALENTINE, J. W., FOIN, T. C. & PEART, D. 1978. Aprovincial model of Phanerozoic marine diversity.Paleobiology, 4, 55-66.
VAN STAAL, C. R., DEWEY, J. F, MAC NIOCAILL, C.& MCKERROW, W. S. 1998. The Cambrian-Siluriantectonic evolution of the northern Appalachiansand British Caledonides: history of complex,west and southwest Pacific-type segment oflapetus. In: BLUNDELL, D. J. & SCOTT, A. C. (eds)Lyell: the Past is the Key to the Present. GeologicalSociety of London, Special Publications, 143,199-242.
VERMEIJ, G. J. 1993. The biological history of aseaway. Science, 260, 1603-1604.
WEBBY, B. D., PARIS, F. & DROSER, M. L. 1999. IGCPProject No. 410: The great Ordovician biodiversi-fication event: implications for global correlationand resources. Acta Universitatis Carolinae-Geo-logica 43: xi-xiii.
WESTROP, S. 1991. Intercontinental variation in massextinction patterns: influence of biogeographicstructure. Paleobiology, 17, 363-368.
WESTROP, S. & ADRAIN, J. 1998. Trilobite alpha diver-sity and the reorganization of Ordovician benthicmarine communities. Paleobiology, 24, 1-16.
WHITTAKER, R. H. 1977. Evolution of species diversityin land communities. Evolutionary Biology, 10,1-67.
ZHURAVLEV, A. Y. 2001. Biotic diversity and structureduring the Neoproterozoic-Ordovician tran-sition. In: ZHURAVLEV, A. Y. & RIDING, R. (eds)The Ecology of the Cambrian Radiation. Colum-bia University Press, New York, 173-199.

This page intentionally left blank

Brachiopods: Camhrian-Treniadoc precursors to Ordovicianradiation events
MICHAEL G. BASSETT 1, LEONID E. POPOV1 & LARS E. HOLMER2
1Department of Geology, National Museum of Wales, Cathays Park, CardiffCF10 3NP, UK (e-mail: [email protected])
2Institute of Earth Sciences, Department of Historical Geology and Palaeontology,University of Uppsala Norbyvagen 22, S-75236, Uppsala, Sweden
(e-mail: [email protected])
Abstract: Brachiopod-dominated palaeoeommunities incorporating a structure typical offaunal groups within the Palaeozoic Evolutionary Fauna were already present in North andEast Gondwana and associated terranes as early as the mid-Cambrian, confined exclusivelyto shallow marine, inshore environments. The late Cambrian and Tremadoc record of thesefaunas is incomplete, because of pronounced global sea-level lowstand and subsequentbreak-up and destruction of the Cambrian Gondwanan margin. It is likely, however, thatthose groups later forming the core of the Palaeozoic Evolutionary Fauna evolvedoriginally in shallow-water environments of low-latitude peri-Gondwana, and dispersedwidely when favourable ecological conditions developed. Conspicuous sea-level risethrough the early to mid-Arenig provided newly available habitats in the expanding epeiricseas, where the new faunas evolved and diversified by the mid-Ordovician, when rapid driftseparated the early Palaeozoic continents. Relatively short-lived precursor and transitionalbrachiopod assemblages can be identified on most of the main palaeocontinents prior tothe Ordovician radiation of the Palaeozoic Evolutionary Fauna.
The Ordovician evolutionary radiation ofmarine metazoans was the second significantbiodiversification event in Phanerozoic biotichistory, following the origin and explosiveradiation of skeletonized faunas at the beginningof the Cambrian. Ordovician events resulted notonly in a significant global increase of taxonomicdiversity, but also in substantial changes ofbenthic community structure on marine shelves,where the dominant trilobite-lingulate brachio-pod associations of the Cambrian EvolutionaryFauna were replaced mainly by more advancedand structured benthic assemblages of thePalaeozoic Evolutionary Fauna, dominated byfilter-feeders and especially by rhynchonelli-formean brachiopods, bryozoans and pelmato-zoan echinoderms (Sepkoski 1981, 1995). Therewas substantial overlap between these twobiotopes in the earlier Ordovician, but gener-ally on some major Lower Palaeozoic plates,including Laurentia, Baltica and Siberia, fullydeveloped communities of the PalaeozoicEvolutionary Fauna have little in common withtransitional faunas in which direct descendantsof local Cambrian lineages predominate, orwhere transformation of the assemblagesoccurred almost abruptly so that newly emerg-ing benthic faunas had little evident linkage withtheir Cambrian and early Ordovician predeces-sors (Sokolov 1982; Sepkoski & Sheehan 1983;
Popov 1993; Patzkowsky 1995).Rhynchonelliformean brachiopod assem-
blages that we identify and name below astransitional to the Palaeozoic EvolutionaryFauna (e.g. Clarkella Fauna and various syn-trophinidine and Tritoechia-Protambonitesassociations) require particular discussion anddefinition. They already formed trophicstructures (dominance of suspension-feeders)and tiering (presence of at least two levels)characteristic of the Palaeozoic Fauna, but thetaxonomic composition of brachiopod generaand families retains distinct links to the ancestralCambrian faunas, whereas their relationshipwith succeeding brachiopod faunas is lessevident. These transitional assemblages alsolack ostracodes and bryozoans, which areusually important components of Ordovicianbiotopes.
The nature of faunal patterns and replace-ment was especially significant on the shallowshelves of Gondwana and its closely associatedmarginal terranes, where the development ofdistinctive trophic and taxonomic communitystructures was rooted deep in the Cambrian.This suggests that increased faunal exchangebetween palaeoplates at the beginning of theOrdovician could have been an important factorin triggering subsequent changes in communitystructure more widely across marine shelves,
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194, 13-23.0305-8719/02/$15.00 © The Geological Society of London 2002.
14 M. G. BASSETT, L. E. POPOV & L. E. HOLMER
resulting in the significant increases in bio-diversity that characterize the OrdovicianPeriod.
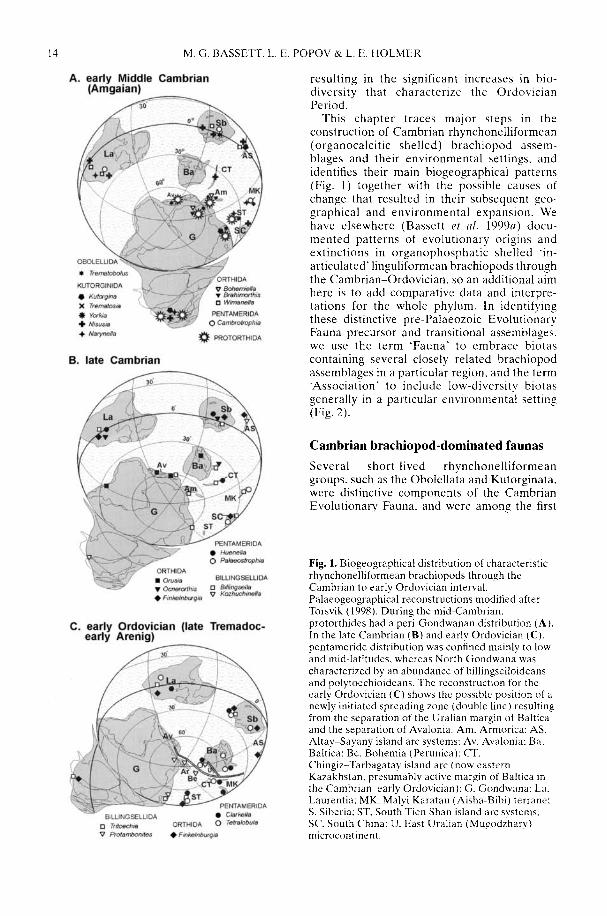
This chapter traces major steps in theconstruction of Cambrian rhynchonelliformean(organocalcitic shelled) brachiopod assem-blages and their environmental settings, andidentifies their main biogeographical patterns(Fig. 1) together with the possible causes ofchange that resulted in their subsequent geo-graphical and environmental expansion. Wehave elsewhere (Bassett et al. 1999a) docu-mented patterns of evolutionary origins andextinctions in organophosphatic shelled 'in-articulated' linguliformean brachiopods throughthe Cambrian-Ordovician, so an additional aimhere is to add comparative data and interpre-tations for the whole phylum. In identifyingthese distinctive pre-Palaeozoic EvolutionaryFauna precursor and transitional assemblages,we use the term 'Fauna' to embrace biotascontaining several closely related brachiopodassemblages in a particular region, and the term'Association' to include low-diversity biotasgenerally in a particular environmental setting(Fig. 2).
Cambrian brachiopod-dominated faunasSeveral short-lived rhynchonelliformeangroups, such as the Obolellata and Kutorginata,were distinctive components of the CambrianEvolutionary Fauna, and were among the first
Fig. 1. Biogeographical distribution of characteristicrhynchonelliformean brachiopods through theCambrian to early Ordovician interval.Palaeogeographical reconstructions modified afterTorsvik (1998). During the mid-Cambrian,protorthides had a peri-Gondwanan distribution (A).In the late Cambrian (B) and early Ordovician (C).pentameride distribution was confined mainly to lowand mid-latitudes, whereas North Gondwana wascharacterized by an abundance of hillingselloideansand polytoechioideans. The reconstruction for theearly Ordovician (C) shows the possible position of anewly initiated spreading zone (double line) resultingfrom the separation of the Uralian margin of Balticaand the separation of Avalonia. Am, Armorica: AS.Altay-Sayany island arc systems: Av. Avalonia: Ba.Baltica: Be, Bohemia (Perunica); CT.Chingiz-Tarbagatay island arc (now easternKazakhstan, presumably active margin of Baltica inthe Cambrian-early Ordovician): G. Gondwana: La.Laurentia: MK, Malyi Karatau (Aisha-Bibi) terrane:S. Siberia: ST, South Tien Shan island arc systems:SC. South China: U, East Uralian (Mugodzhary)microcontinent.
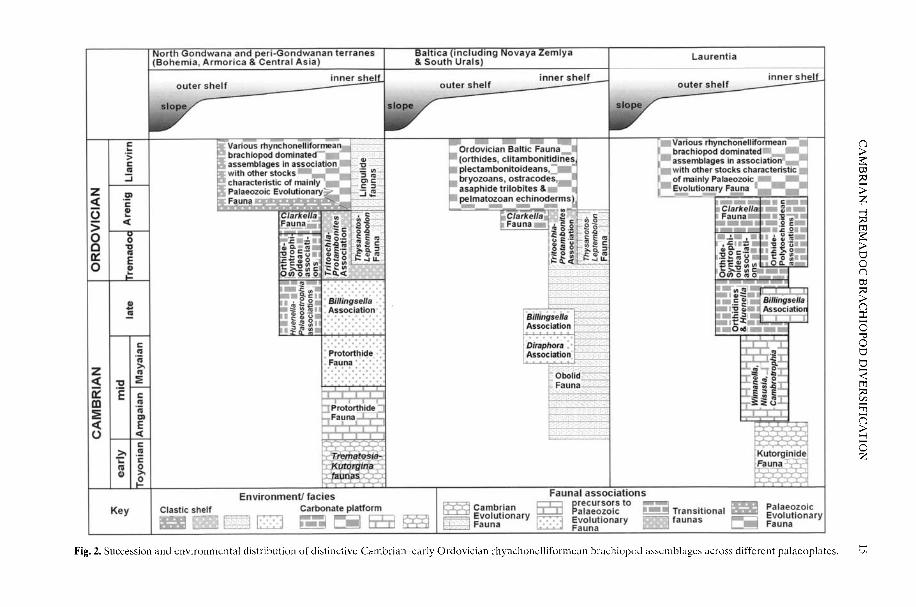
Fig. 2. Succession and environmental distribution of distinctive Cambrian-early Ordovician rhynchonclliformean brachiopod assemblages across different palaeoplates.

16 M. G. BASSETT. L. E. POPOV & L. E. HOLMER
animals to form distinctive communities withessentially the same trophic and environmentalcharacteristics as brachiopod-dominated com-munities of the Palaeozoic Evolutionary Fauna.The kutorginides Kutorgina and Trematosiaand the obolellide Trematobolus formed theoldest known rhynchonelliformean brachiopod-dominated assemblages (Figs 1, 2). These assem-blages were almost invariably of low taxonomicdiversity, generally comprising or dominated bya single species, living in low-energy, shallowshelf environments affected by storm events, onlow-latitude carbonate platforms usually in oradjacent to areas of hypersaline carbonateaccumulation. Two illustrative examples are themarginal part of the Turukhansk-Irkutsk faciesbelt on the Siberian Plate, characterized bydeposition of evaporites (Pelman 1992), and theBurj Formation on the eastern Dead Sea coast ofJordan together with the closely adjacent andcoeval Nimra Formation of the southern Negevdesert, Israel (Cooper 1976; and our unpub-lished studies), deposited on northern peri-Gondwana. These successions also contain theoldest known brachiopod coquinoid concentra-tions, usually in storm beds, which in some casesare of remarkable abundance and extent(Powell 1989). With the extinction of obolellidesand kutorginides at the end of the earlyCambrian, rhynchonelliformean brachiopod-dominated assemblages and shell bed accumu-lations declined during the mid-Cambrian inSiberia and Laurentia, but they remained acharacteristic feature of shallow shelf environ-ments across Gondwana and peri-Gondwananregions, as discussed below.
By mid-Cambrian times, benthic palaeo-communities with a structure and dominanttaxonomic composition typical of the PalaeozoicEvolutionary Fauna occurred widely acrossGondwana (Middle East, Australia) and neigh-bouring areas, where they were confined exclus-ively to shallow, inshore marine environments(e.g. Cooper 1976; Roberts & Jell 1990). Twocharacteristic assemblages can be recognized.
The first is named here as the ProtorthideFauna (Fig. 2), comprising a medium-diversityassociation of archaic orthidines and protor-thides and often including some obolellide,kutorginide and linguliformean taxa (Fig. 1A).We interpret protorthides as a stem group topentamerides and clitambonitoideans (unpub-lished studies; see also Williams et al. 1996). Thisfauna embraced a wide variety of mid-Cambrianbrachiopod associations containing character-istic protorthide components (e.g. Protorthis inNew Brunswuck; Jamesella in Bohemia, Spainand North Africa; Glyptoria and Psiloria in
Israel and Kyrgizstan; Arctochedra in Kyrgizstanand Australia, etc.). Nearly all known pro-torthide occurrences (Fig. 1A) except Kyrgizs-tan and Alaska were parts of Gondwanathrough the Cambrian (Dalziel 1997; Torsvik &Rehnstrom 2001). Lower to Middle Cambriandeposits of South Tien Shan in Kyrgizstanprobably originated in an adjacent peri-Gondwanan island arc (Fig. 1A; Holmer et al.2000). The occurrence of the protorthide Arc-tochedra in Alaska (Cooper 1936) does notseemingly indicate expansion of the group toLaurentia, but in our view was probably relatedto an exotic terrane of uncertain origin.
The second assemblage is defined here asthe Billingsella Association (Fig. 2). It is a low-diversity association dominated by Billingsellaitself or by related billingsellid genera (e.g.Cymbithyris), and shows patterns characteristicof opportunistic life strategies (Alexander1977), e.g. limited areal distribution, highdensity clustering in thin, widespread iso-chronous horizons, overwhelming numericaldominance in the assemblage and remarkableabundance in atypical facies, mostly relatedto nearshore depositional environments.Billingsellids often formed inshore coquinoidaccumulations. The most spectacular examplesknown to us are extensive shell beds in theUpper Cambrian Kyjandy Formation of north-central Kazakhstan (Nikitin 1956) and in theMiddle to Upper Cambrian Derenjal Formationof east-central Iran (Fig. 3a). We interpretbillingsellides as the ancestral stock of theStrophomenata including strophomenides.orthotetides and polytoechioideans (see alsoWilliams et al. 1996). The Diraphora Associationis a generally monospecific assemblage whichimmediately precedes the Billingsella Associ-ation in regions such as Novaya Zemlya (Fig. 2;Popov 1984); the ecological setting of both theseassociations appears to be very similar.
The Protorthide Fauna was confined exclus-ively to the mid-Cambrian of peri-Gondwana.whereas the Billingsella Association becamealmost cosmopolitan in the late Cambrian (Fig.1). The rhynchonell iformean taxa in bothassemblages can be traced phylogenetically intoOrdovician descendants, and both the Protor-thide Fauna and Billingsella Association prob-ably represent nuclei that gave rise to stockscharacteristic of the Palaeozoic EvolutionaryFauna.
In the Tremadoc-early Arenig, theBillingsella Association sensu stricto wasreplaced across the shallow clastic shelvesof North Gondwana (Fig. 2; Iran. NorthAfrica, Armorica) by low-diversity assemblages

CAMBRIAN-TREMADOC BRACHIOPOD DIVERSIFICATION 17
Fig. 3. Examples of late Cambrian-early Ordovician brachiopods that occur typically as coquinoidaccumulations. (a) Storm bed formed by the accumulation of Billingsella sp., NMW 2001.45G.1, UpperCambrian, Derenjal Formation, Shirgesht, Tabas Region, East Central Iran. (b) Shell bed comprising thesyntrophioidean Clarkella supina Nikitin, NMW 98.67G.95, Lower Ordovician, Koagash Formation, KoagashRiver, South Urals, Kazakhstan. (c, d) Billingsella sp., NMW 2001.45G.2, interior and exterior of ventral valve,horizon and locality as in (a). (e, f) Protambonites sp. NMW 2001.45G.3, Lower Ordovician, ShirgeshtFormation, Shirgesht, Tabas Region, East Central Iran. (g) Bedding surface with accumulation of Huenellatexana (Walcott), USNM 52494, Upper Cambrian, Packsaddle Mountain, Llano County, Texas, USA. (h, k)Characteristic protorthide taxa from coquina accumulations in the Middle Cambrian, Amgaian, ArpatektyrMountains, South Kyrgizstan: (h, k) NMW 98.69G.22, dorsal valve interior and NMW 98.69G.24 ventral valveof Arctohedra pyramidalis Aksarina; (i) CNIGR Museum 22/12761, dorsal valve exterior of Glyptoriagulchensis Popov and Tikhonov. NMW, National Museum of Wales, Cardiff; USNM, United States NationalMuseum, Washington DC; CNIGR, Central Geological Research and Exploration Museum, St Petersburg.

18 M. G. BASSETT, L. E. POPOV & L. E. HOLMER
dominated by the polytoechioideans Protam-bonites (Fig. 3e-f) or Tritoechia (e.g. Serre de losCabos area of NW Spain, Shirgesht Formationof central Iran; Villas et al. 1995; Bassett et al.1999b). These assemblages also spread tothe Uralian margin of Baltica near theTremadoc-Arenig boundary (Popov et al. 2001).Polytoechioideans were probably deriveddirectly from the Billingsella Association(Popov et al. 2001) and demonstrate similarenvironmental requirements (Figs 1C, 2). InIran and the South Urals they co-occur with aThysanotos-Leptembolon Association of lin-guliformean brachiopods (Fig. 2; Popov &Holmer 1994, 1995; Bassett et al. 1999b).
Another distinctive group of late Cambrian-Tremadoc rhynchonelliformean assemblageswas formed by syntrophioideans. These earlypentamerides were most typical of low-latitudecarbonate platforms on microcontinents andisland arcs associated with Gondwana (e.g. Southand North China, Central Asia, etc.) and Lau-rentia (Figs 1B, 2). The core of these assemblageswas generally formed by a single taxon, usuallyHuenella (Fig. 3g) or Palaeostrophia. Associatedbrachiopods were mostly orthides (e.g. Apheoor-this, Eoorthis, etc.) and less abundantBillingsella. The co-occurrence of these faunaswith pelmatozoan echinoderms such aseocrinoids is fairly typical, suggesting a relativelycomplex trophic structure involving at least twotiering levels. By the end of the Tremadoc (lowerParoistodus proteus Biozone), the syntro-phioidean-dominated faunas were transformedto medium-diversity associations typified by theco-occurrence of the pentamerides Clarkella,Diaphelasma, Glyptotrophia, Tetralobula, etc.Here we identify this as the Clarkella Fauna(Figs 1C, 2, 3b). Brachiopods from the LowerOrdovician Olenty Formation of north-centralKazakhstan described by Nikitin (1956) includea good example of this newly emerging fauna.The Clarkella Fauna is unknown in WestGondwana, which by then had drifted to a highlatitude (Fig. 1C), but it is characteristic of lowlatitude Laurentia and of the numerous terranesnow incorporated into the complex tectoniccollage of Central Asia and Kazakhstan, and isalso recorded in South China and the Uralianmargin of Baltica, located at that time in temper-ate latitudes. In all these regions, assemblages ofthe Clarkella Fauna occur in shallow-watercarbonate depositional environments; they alsoincorporate descendants of local late Cambrianlineages (e.g. Finkelnburgia, Apheoorthis, Eoor-this, etc.) and polytoechioideans, but showlittle in common with succeeding Ordovicianbrachiopod associations.
Laurentia, Baltica and Siberia: generalpatterns of faunal replacement in the earlyOrdovicianThe general characters of faunal replacementprior to the early Ordovician radiation are welldocumented and analysed only for Laurentia(Sepkoski & Sheehan 1983; Droser & Sheehan1997), where the transition from benthiccommunity types of the Cambrian EvolutionaryFauna to those characteristic of the PalaeozoicEvolutionary Fauna occurred in the Ibexian-early Whiterockian. Notable diversification ofechinoderms (e.g. Sprinkle 1995) in the Ibexianrepresents the earliest indication of the Ordo-vician radiation in Laurentia (Guensberg &Sprinkle 1992), whereas the diversity ofrhynchonelliformean brachiopods remainedrelatively low, represented mostly by familiestransitional from the Cambrian (Patzkowsky1995). The abundance of polytoechiides andsyntrophiidines diagnostic of the ClarkellaFauna is also characteristic. Typical Ordovicianrhynchonelliformeans (e.g. camerelloideans andplectambonitoideans) together with bryozoansbecame increasingly abundant during theWhiterockian (Wilson et al. 1992), which was theinterval when benthic assemblages with astructure and composition characteristic of thePalaeozoic Evolutionary Fauna became fullyformed and widespread across Laurentia(Patzkowsky 1995; Sepkoski & Sheehan 1983;Droser & Sheehan 1997).
Available data from Baltica demonstrate thata benthic structure typical of the PalaeozoicEvolutionary Fauna arose there as early as theBillingenian (mid-Arenig, Prioniodus elegansBiozone), including the earliest representativesof plectambonitoideans, camerelloideans,endopunctate orthides and clitambonitidinesamong brachiopods, plus bryozoans, ostracodes,pelmatozoan echinoderms and asaphide trilo-bites (Fig. 2; Popov 1993; Pushkin & Popov1999). This newly emergent fauna had noobvious origins in the low-diversity obolid-dominated assemblages of shallow shelfenvironments or in the predominantly dysoxicouter shelf olenid trilobite faunas characteristicof Baltica in the late Cambrian-Tremadoc. Thesharp nature of faunal replacement in Balto-scandia suggests regional extinction and sub-sequent fairly rapid immigration of new taxa,concomitant with significant environmentalchanges including the onset of continuouscarbonate sedimentation and the developmentof numerous hardground surfaces (Dronov et al.1996).
In Siberia, early Ordovician (Tremadoc to

CAMBRIAN-TREMADOC BRACHIOPOD DIVERSIFICATION 19
early Arenig) benthic assemblages retainedmajor characters of the Cambrian EvolutionaryFauna, with abundant trilobites and with low-diversity orthide brachiopod assemblages rep-resented by families transitional from the lateCambrian, accompanied by syntrophioideans(Sokolov 1982). The transitional intervalthrough the late Arenig-Llanvirn was character-ized by the presence of an essentially endemiclow-diversity fauna of trilobites and rhyncho-nelliformeans, the problematical molluscsAngarella and Moyeronia, gastropods, locallyabundant ostracodes, and the rare tabulate coralCryptolichenaria (Kanygin et al. 1984). Otherdistinctive components of the PalaeozoicEvolutionary Fauna, such as rhynchonelli-formean brachiopods of the orders Stropho-menida, Triplesiida and Rhynchonellida,bryozoans and crinoids were introduced intoSiberia only during the late Llanvirn-Llandeilo,where their sudden appearance suggests a majorimmigration from Laurentia (Rozman 1977).
Brachiopod radiation andpalaeogeographyWithin the newly emergent Ordovician faunas,rhynchonelliformean brachiopods were amongthe most important components, which insignificant part defined the structure of benthiccommunities. Characters of faunal turnoverand replacement in the early Ordovician ofLaurentia, Baltica and Siberia - the three majorearly Palaeozoic plates outside Gondwana -suggest the importance of large-scale immi-gration of new brachiopod taxa, such asstrophomenides, endopunctate orthides, clitam-bonitidines, triplesiides, and possibly camerel-loideans, often of uncertain geographical andevolutionary origin, which comprise a significantcomponent of the new faunas. Present-dayrhynchonelliformeans and many bryozoanshave a very low dispersal potential because ofthe very short free-swimming duration of theirlecithotrophic larvae (Hyman 1959). Recentphylogenetic studies suggest that lecithotrophiclarvae evolved early in ancestors of craniifor-mean and rhynchonelliformean brachiopods(Williams et al. 1996), and data from Palaeozoicrhynchonelliformean brachiopod biogeographyalso point to a relatively low ability for migrationacross oceans (Boucot 1975; Williams 1973,1976). Their rapid expansion across the shelvesof all major palaeoplates in the early to mid-Ordovician may therefore reflect a uniquecombination of environmental, geographicaland tectonic factors that allowed them to crossoceanic divides separating major continents and
then to settle, replace or integrate successfullyinto previously aboriginal benthic assemblages.
Following the early Cambrian radiation,when the trilobite and lingulate brachiopod-dominated Cambrian Evolutionary Fauna wasestablished and diversified, the mid-Cambrianto Tremadoc interval was a time of relativeevolutionary and environmental conservatism.By comparison with the succeeding Ordovician,this interval was characterized by pronouncedglobal sea-level lowstand with insignificant sea-level fluctuations, as indicated recently by theanalysis of almost continuous shallow marinedeposition in the East Baltic (Artyushkov et al.2000), accompanied by minimal volcanicactivity, while the expansion of lapetus led toincreasing separation of Baltica from Laurentia.By contrast, the Arenig-Llanvirn was a periodof remarkable tectonic activity, which includedthe break-up of the Gondwanan margin, result-ing in particular in the origin of Avalonia as aseparate microplate (Torsvik 1998) and separ-ation of the East Uralian microcontinent fromBaltica (Sengor & Natal'in 1996). Onset of arcvolcanism in the Tremadoc of Eastern Avalonia(e.g. Kokelaar et al. 1984) and the late Cambrianto early Tremadoc rapid subsidence of the Welshbasin (Prigmore et al. 1997) possibly suggestinitial development of rifting of Avalonia fromGondwana at about that time. Prigmore et al.(1997) supposed that the wider separation ofAvalonia from Gondwana possibly coincidedwith a second phase of subsidence in theArenig-early Llanvirn, although only by thelate Llanvirn had evident biogeographicaldifferences developed between Avalonian andGondwanan shallow-shelf benthic faunas(Cocks 1999). Available data for the Uralianmargin of Baltica are less focused, but theysuggest initiation of late Cambrian to earlyOrdovician rifting of the Uralian margin ofBaltica (Zonenshain et al. 1984), and possiblythe detachment of island arc systems east ofBaltica related to backarc spreading (Sengorand Natal'in 1996).
The possible position of Baltica in relation toNorth Gondwana (Fig. 1C) suggests that thischain of events may have resulted in the for-mation of a new spreading zone comparablewith that of the present-day North Atlanticoceanic ridge, followed by a tectonically inducedsea-level rise and a change of direction in therotation of Baltica, leading to initiation ofthe closure of lapetus. Volcanism commencedin Avalonia by at least the late Tremadoc(Kokelaar et al. 1984), and the presence of anactive volcanic arc in the proximity of theCaledonian margin of Baltica at about that time

20 M. G. BASSETT. L. E. POPOV & L. E. HOLMER
became evident (Sturt & Roberts 1991). Evi-dence of increased early Ordovician island arcvolcanism in the orogenic belts of Central andEast Asia (Nikitin et al. 1991; Apollonov 2000)also suggests that the relatively narrow oceanicdivide between the eastern margin of Balticaand equatorial Gondwana was occupied bychains of volcanic island arcs and microconti-nents acting possibly as faunal 'bridges' betweenthese two continents.
There is little doubt that the OrdovicianPeriod mainly documents an interval of substan-tial sea-level rise, with a maximum in theCaradoc when most palaeoplates were flooded(Barnes et al. 1996). Superimposed second-ordereustatic cycles corresponding approximately tothe Tremadoc, Arenig and Llanvirn also can berecognized (Fortey 1984). The first episode ofsea-level rise, corresponding to the transgressivephase of the Black Mountain eustatic event(mid-Tremadoc) of Miller (1984), is recorded onnearly all early Palaeozoic continents. This andthe second transgressive phase in the Arenigreduced the influx of siliciclastic sediments intothe large epeiric basins and provided good con-ditions for carbonate deposition, both in low(Laurentia, Siberia, North China) and temper-ate (Baltica, South China) latitudes. Areas ofshallow marine carbonate deposition withnumerous hardgrounds provided environmentsin which the earliest rhynchonelliformeanbrachiopod-dominated associations evolved.
Little is known of the characteristics of faunalreplacement during the late Cambrian to earlyOrdovician of Gondwana and its associatedterranes. In part this is a result of the markedperiod of sea-level lowstand through the lateCambrian to early Tremadoc, and the subse-quent destruction of the Cambrian Gondwananmargin through the Palaeozoic, when areas ofCambrian shallow marine deposition wereseparated and later obscured within tectoniccollages of Western Europe, the Middle East,and Central and Southeast Asia. By the begin-ning of the Ordovician, the North African andMiddle Eastern sectors of Gondwana driftedinto high latitudes and brachiopod assemblageslinked to areas of carbonate deposition mainlydisappeared (Fig. 1C). However, the lateCambrian Billingsella Association and succeed-ing early Ordovician polytoechioidean associ-ations remained characteristic of some regions,for example in Iran and Armorica (Fig. 2). It islikely that in the late Cambrian-Tremadoc,benthic assemblages transitional to the Palaeo-zoic Evolutionary Fauna persisted mainly infaunas of peri-Gondwanan terranes, such as theSouth and North China plates, or on island arcs
and microplates incorporated into the orogenicbelts of Central Asia, where the Clarkella Faunabecame widespread in the late Tremadoc-earlyArenig (Nikitin 1956; Holmer et al. 2001) andalso colonized the Uralian margin of Baltica(Bondarev 1968) before that plate began itsrapid northwesterly drift away from Gondwana.The early Arenig brachiopod-dominated faunaof the Billingenian Regional Stage (mid-Arenig) in the East Baltic contains all the mainfeatures of the Palaeozoic Evolutionary Fauna,including such characteristic groups as bryo-zoans and ostracodes (Popov 1993; Pushkin &Popov 1999). Assemblages of similar characterappeared in Laurentia somewhat later, in themid-Whiterockian (Wilson et al. 1992). Similarfaunas in Siberia, which was possibly the mostisolated continent of that time, appeared only inthe late Llanvirn-Llandeilo. The abrupt natureof the faunal replacement in Baltica and Siberia,together with the abundance of new groups ofhigh taxonomic rank and of taxa unrooted inindigenous lineages, suggests that the newlyappearing faunas were mature assemblages thathad evolved and immigrated from elsewhere.By contrast, faunal replacement in Laurentiawas passive and non-competitive in nature(Westrop & Adrain 1998), and newly appearingtaxa of rhynchonell iformean brachiopodsand bryozoans were incorporated into thetransformed benthic assemblages together withlocally evolved faunas of trilobites andechinoderms.
Synopsis
The pre-Arenig nuclei of benthic communitiesdominated by suspension-feeders with a struc-ture and composition characteristic of thePalaeozoic Evolutionary Fauna most probablyevolved on the Cambrian shallow carbonateshelves of equatorial Gondwana and peri-Gondwanan terranes. Dispersion of thesefaunas outside Gondwana is evident from thelate Cambrian-Tremadoc, when syntrophioi-dean, Billingsella and polytoechiid associationsreached Laurentia, Siberia and the Uralianmargin of Baltica, but wider significant trans-formation of benthic shelf assemblages wasdelayed until the Arenig-Llanvirn and thenoccurred diachronously across the major palaeo-continents. Substantial sea-level rise from theTremadoc to Caradoc, coupled with extensivecarbonate deposition and the development ofassociated hardgrounds in the shallow epeiricseas of low and temperate latitudes, establishedincreased ecospace for colonization by the newlyemerging benthic assemblages of the Palaeozoic

CAMBRIAN-TREMADOC BRACHIOPOD DIVERSIFICATION 21
Evolutionary Fauna. These faunas included newstocks of rhynchonelliformean brachiopods, andpossibly bryozoans (Hu & Spjeldnaes 1991),which were preadapted to these environments.Thus the well known onshore-offshore patternin ecological expansion of rhynchonelliformeanbrachiopod assemblages in the Ordovician(Sepkoski 1991) reflects the fact that areas ofshallow carbonate deposition were the primaryenvironments in which these brachiopodsflourished in the Cambrian, and which they thenoccupied first in their geographical expansion inthe early Ordovician.
As noted by Sepkoski & Sheehan (1983),'there was no immediately obvious physicaltrigger for such a great burst of evolutionaryactivity' to drive the Ordovician biodiversi-fication. However, significant intensification ofplate tectonic activity at the beginning of theOrdovician, including rifting of the Gondwananmargin and the Uralian margin of Baltica,resulted in the origin of extensive new spreadingzones and substantial increase of island arcvolcanism and eustatic sea-level rise, whichprovided a physical background for the remark-able transformations of marine biodiversity.
Reconstructions of Baltica and Laurentiaduring the Tremadoc-early Arenig place themin the relative proximity of Gondwana (Dalziel1997; Torsvik 1998; Torsvik & Rehnstrom 2001).Benthic faunal groups with low dispersal poten-tial thus had enhanced opportunities to cross theintervening oceans; at the same time, newlyemergent volcanic arcs formed 'island hoppingbridges' between the major continents, tofurther develop the pathways for such migra-tions. Conspicuous sea-level rise at the begin-ning of the Ordovician led to the development ofvast new shelves and platforms in the expandingepeiric seas, which the new faunas invaded, thenevolved and diversified by mid-Ordovician timeswhen rapid drift began the separation of theearly Palaeozoic continents.
M.G.B. and L.E.P. acknowledge support from theRoyal Society of London and the National Museum ofWales for work on early Palaeozoic brachiopods.L.E.H. completed part of his contribution in Cardiffunder a European Science Exchange Programmefunded by the Royal Society and Royal SwedishAcademy, awarded jointly with M.G.B. L.E.H. andL.E.P. acknowledge the receipt of grants from theSwedish Natural Science Research Council (NFR).This paper is a contribution to IGCP Project 410. Weare grateful to A. W. A. Rushton for his mosthelpful comments that have improved the manuscript,and to A. W. Owen for his scientific and editorialcomments.
References
ALEXANDER, R. R. 1977. Growth, morphology andecology of Paleozoic and Mezosoic opportunisticspecies of brachiopods from Idaho-Utah. Journalof Paleontology, 51, 1133-1149.
APOLLONOV, M. 2000. Geodinmicheskaya evolyutsiyaKazakhstana v rannem paleozoe (s pozitsyi klas-sicheskoi tektoniki plit) [Geodynamic evolutionof Kazakhstan in the early Palaeozoic (fromclassic plate tectonic positions)]. In: BESPAEV, H.A. (ed.) Geodinamika i minerageniya Kaza-khstana, 1, VUC Publishing House, Almaty,46-63 (in Russian).
ARTYUSHKOV, E.V., LINDSTROM, M. & POPOV, L. E.2000. Relative sea-level changes in Baltoscandiain the Cambrian and early Ordovician: the pre-dominance of tectonic factors and the absence oflarge scale eustatic fluctuations. Tectonophyisics,320, 375-407.
BARNES, C.R., FORTEY, R. A. & WILLIAMS, S. H. 1996.The pattern of global bio-events during theOrdovician Period. In: WALLISER, O. T. (ed.)Global Events and Event Stratigraphy in thePhanerozoic. Springer, Berlin-Heidelberg,139-172.
BASSETT, M. G., POPOV, L. E. & HOLMER, L. E. 1999a.Organophosphatic brachiopods: patterns of bio-diversification and extinction in the early Palaeo-zoic. Geobios, 32, 145-163.
BASSETT, M. G., DASTANPOUR, M. & POPOV, L. E.1999b. New data on Ordovician faunas andstratigraphy of the Kerman and Tabas regions,east-central Iran. Acta Universitatis Carolinae,Geologica, 43, 483–486.
BONDAREV, V. I. 1968. Stratigrafiya i kharakternyebrakhiopody ordovikskikh otlozhenii yuga NovoiZemli, ostrova Vaigach i severnogo Pai-Khoia[Stratigraphy and characteristic brachiopods ofthe Ordovician deposits of southern NovayaZemlya, Isle of Vaigach and northern Pai-Khoi].Trudy Nauchno- Issledovatel'skogo InstitutaGeotogii Arktiki (NIIGA), 157. 3-144 (inRussian).
BOUCOT, A. 1975. Evolution and Extinction RateControls. Developments in Paleontology andStratigraphy, Elsevier, Amsterdam.
COCKS, L. R. M. 1999. The Early Palaeozoic geogra-phy of Europe. Journal of the Geological Society,London, 157, 1-10.
COOPER, G. A. 1936. New Cambrian brachiopods fromAlaska. Journal of Paleontology, 10, 210-214.
COOPER, G. A. 1976. Lower Cambrian brachiopodsfrom the Rift Valley (Israel and Jordan). Journalof Paleontology, 50, 269-289.
DALZIEL, I. W. D. 1997. Neoproterozoic-Paleozoicgeography and tectonics: Review, hypothesis,environmental speculation. Geological Society ofAmerica Bulletin, 109, 16-42.
DRONOV, A. Ju., SAVITSKY, Ju. V., FEDOROV, P. V. &TSYGANOVA, E. A. 1996. Detailed lithostratigra-phy of the Ordovician lower Volkhovian lime-stone along the eastern part of the Baltic-LadogaGlint, northwestern Russia. GFF, 118, 19-24.

22 M. G. BASSETT, L. E. POPOV & L. E. HOLMER
DROSER, M. L. & SHEEHAN, P. M. 1997. Palaeoecologyof the Ordovician radiation: resolution of large-scale patterns with individual clade histories,palaeogeography and environments. Geobios, 20.221-229.
FORTEY, R. A. 1984. Global earlier Ordoviciantransgressions and regressions and their bio-logical implications. In: BRUTON, D. L. (ed.)Aspects of the Ordovician System. Universityof Oslo Palaeontological Contributions, 295,37-50.
GUENSBERG, T. E. & SPRINKLE, J. 1992. Rise ofechinoderms in the Paleozoic fauna: Significanceof paleoenvironmental controls. Geology, 20,407–410.
HOLMER, L. E., POPOV, L. E. & BASSETT, M. G. 2000.Early Ordovician organophosphatic brachiopodswith Baltoscandian affinities from the AlayRange, southern Kyrgyzstan. GFF, 122, 367-375.
HOLMER, L. E., POPOV, L. E., KONEVA, S. P. &BASSETT, M. G. 2001. Cambrian - early Ordo-vician brachiopods from Malyi Karatau, thewestern Balkhash Region, and northern TienShan, Central Asia. Special Papers in Palaeontol-ogy, 65, 1-180.
Hu, ZHAO-XUN & SPJELDNAES, N. 1991. Early Ordovi-cian bryozoans from China. Bulletin de la Societedes Sciences Naturelle Ouest France, Hors Serie 1,179-189.
HYMAN, L. H. 1959. The Invertebrates 5. SmallerCoelomate Groups. McGraw Hill, New York.
KANYGIN, A. V., MOSKALENKO, T. A., OBUT, A. M. &SENNIKOV, N. V. 1984. Ordovik [Ordovician]. In:Fanerozoi Sibiri. 1. Vend, paleozoi. Trudy Insti-tuta geologii i geifiziki Sibirskogo otdelenivaAkademii Nauk SSSR, 595, 60-88 (in Russian).
KOKELAAR, B. P., HOWELLS, M. F., BEVINS, R. E.,ROACH, R. A. & DUNKLEY, P. N. 1984. TheOrdovician marginal basin of Wales. In: KOKE-LAAR, B. P. & HOWELLS, M. F. (eds) MarginalBasin Geology: Volcanic and Associated Sedi-mentary and Tectonic Processes in Modern andAncient Marginal Basins. Geological Society,London, Special Publications, 16, 245-269.
MILLER, J. F. 1984. Cambrian and earliest Ordovicianconodont evolution, biofacies and provincialism.In: CLARK, D. L. (ed.) Conodont Biofacies andProvincialism. Geological Society of America,Special Papers, 196, 43-68.
NIKITIN, I. F. 1956. Brakhiopody kembriya i nizhnegoordovika severo-vostoka Tsentral'nogo Kaza-khstana [Cambrian and Lower Ordovicianbrachiopods of north-eastern Central Kaza-khstan]. Nauka, Alma-Ata (in Russian).
NIKITIN, I. F., FRID, N. M. & ZVONTSOV, V. S. 1991.Paleogeography and main features of volcanicityin the Ordovician of Kazakhstan and North Tien-Shan. Geological Survey of Canada, Paper 90-9,259-270.
PATZKOWSKY, M. E. 1995. Ecologic aspects of theOrdovician radiation of articulate brachiopods.In: COOPER, J. D., DROSER, M. L. & FINNEY, S. C.(eds) Ordovician Odyssey: Short Papers forthe Seventh International Symposium on the
Ordovician System. Pacific Section, Society forSedimentary Geology (SEPM). Fullerton. Cali-fornia, 413-414.
PELMAN, Yu. L. 1992. Biostratigraficheskoyeraschleneniye kembriya Severnoi Azii pobrakhiopodam. 1. Sibirskaya Platforma [Bio-stratigraphical subdivision of the Cambrian ofnorth Asia based on brachiopods. 1. SiberianPlatform.] In: REPINA. L. N. & ROZANOV.A. Yu. (eds) Drevneishie brakhiopody terri-torii Severnoi Evrazii. Rossiiskaya AkademiyaNauk, Paleontologicheskii Institut. Sibirskoyeotdeleniye, Obyedinennyi institut geologii.geofiziki i mineralogii. Novosibirsk. 10-21 (inRussian).
POPOV, L. E. 1984. Kembriiskie zamkovyebrakhiopody iz yuzhnogo berega guby Gribovoi.Yiuzhnyi ostrov [Cambrian articulate brachio-pods from the southern coast of the GribovajaHarbor, Southern Island]. In: BONDAREV. V. I.(ed.) Novaya Zemlia na Rannikh EtapakhGeologicheskogo Razvitiya. Sevmorgeologiya.Leningrad, 27-34 (in Russian).
POPOV, L. E. 1993. Dinamika raznoobraziya bentosnoifauny na granitsre kembriya i ordovika naterritorii Vostochno-Evropeiskoi platformy i eeobramleniya [Dynamics of faunal biodiversityacross the Cambrian-Ordovician boundary on theterritory of the East European Plate and adjacentareas]. Stratigrafiya i korrelyatsyia. 1, 33-38 (inRussian).
POPOV, L. E. & HOLMER. L. E. 1994. Cam-brian-Ordovician lingulate brachiopods fromScandinavia, Kazakhstan, and South UralMountains. Fossils and Strata, 35, 1-156.
POPOV, L. E. & HOLMER. L. E. 1995. Distribution ofbrachiopods across the Cambrian-Ordovicianboundary on the East European Plate andadjacent areas. In: COOPER. J. D., DROSER. M. L.& FINNEY, S. C. (eds) Ordovician Odyssey: ShortPapers for the Seventh International Symposiumon the Ordovician System. Pacific Section. Societyfor Sedimentary Geology (SEPM). Fullerton.California, 117-120.
POPOV, L. E., V I N N , O. & N I K I T I N A . O. I. 2001.Brachiopods of the redefined family Tritoechi-idae from the Ordovician of Kazakhstan andSouth Urals. Geobios, 32. 131-155.
POWELL, J. H. 1989. Stratigraphy and sedimentation ofPhanerozoic rocks in Central and South Jordan.Part A. Ram and Khreim Groups. I: 50000Mapping Series Geological Bulletin. 11. 1-72.Amman. Jordan.
PRIGMORE, J. K., ButLER, A. J. & WOODCOCK. N. H.1997. Rifting during separation of EasternAvalonia from Gondwana: Evidence fromsubsidence analysis. Geology. 25. 203-206.
PUSHKIN, V. I. & POPOV, L. E. 1999. Early Ordovicianbryozoans from north-western Russia. Palaeon-tology. 42, 171-189.
ROBERTS, J. & JELL. P. A. 1990. Early Middle Cam-brian (Ordian) brachiopods of the Coonigan For-mation, western New South Wales. Alcheringa.14, 257-309.

CAMBRIAN-TREMADOC BRACHIOPOD DIVERSIFICATION 23
ROZMAN, Kh. S. 1977. Biostratigrafiya i zoogeo-grafiyaverkhnego ordovika Severnoi Azii i SevernoiAmeriki (po brakhiopodam) [Biostratigraphyand zoogeography of the Upper Ordovician ofNorth Asia and North America (based onbrachiopods)]. Trudy Geologicheskogo institutaAkademii Nauk SSSR, 305, 1-171 (in Russian).
SENGOR, A. M. C. & NATAL'IN, B. A. 1996. Paleotec-tonics of Asia: fragments of a synthesis. In: YIN,A. & HARRISON, M. (eds) The Tectonic Evolutionof Asia. Cambridge University Press, Cambridge,486-640.
SEPKOSKI, J. J. Jr. 1981. A factor analysis description ofthe Phanerozoic marine fossil record. Paleobiol-ogy. 7, 36-53.
SEPKOSKI, J. J. Jr. 1991. A model of onshore-offshorechange in faunal diversity. Paleobiology, 17,157-176.
SEPKOSKI, J. J. Jr. 1995. The Ordovician radiations:diversification and extinction shown by globalgenus level taxonomic data. In: COOPER, J. D.,DROSER, M. L. & FINNEY, S. C. (eds) OrdovicianOdyssey: Short Papers for the Seventh Inter-national Symposium on the Ordovician System.Pacific Section, Society for Sedimentary Geology(SEPM), Fullerton, California, 493-496.
SEPKOSKI, J. J. Jr & SHEEHAN, P. M. 1983. Diversifica-tion, faunal change and community replacementduring the Ordovician radiations. In: TEVESZ, M.J. S. & MCCALL, L. L. (eds) Biotic Interpretationsin Recent and Fossil Communities. Plenum Press,New York, 673-717.
SOKOLOV, B. S. (ed.) 1982. Ordovik Sibirskoi platformy(opornyi razrez na reke Kulyumbe) [Ordovicianof Siberian Platform (key section on theKulyumbe River)]. Trudy Instituta geologii igeifiziki Sibirskogo otdeleniya Akademii NaukSSSR, 506, 60-88 (in Russian).
SPRINKLE, J. 1995. Do eocrinoids belong to the Cam-brian or to the Paleozoic evolutionary fauna? In:COOPER, J. D., DROSER, M. L. & FINNEY, S. C.(eds) Ordovician Odyssey: Short Papers for theSeventh International Symposium on the Ordovi-cian System. Pacific Section, Society for Sedi-mentary Geology (SEPM), Fullerton, California,397–400.
STURT, B. A. & ROBERTS, D. 1991. Tectonostrati-graphic relationships and obduction histories ofScandinavian ophiolitic terranes. In: PETERS, T. et
al. (eds) Ophiolite Genesis and Evolution of theOceanic Lithosphere. Ministry of Petroleum andMinerals, Sultanate of Oman; Kluver, Amster-dam, 745-769.
TORSVIK, T. H. 1998. Palaeozoic palaeogeography: ANorth Atlantic viewpoint. GFF, 120, 109-118.
TORSVIK, T. H. & REHNSTROM, E. F. 2001. Cambrianpalaeomagnetic data from Baltica: implicationsfor true polar wander and Cambrian palaeo-geography. Journal of the Geological Society,London, 158, 321-329.
VILLAS, E., ARBIZU, M., BERNARDEZ, E., MENDEZ-BEDIA, I. & ARAMBURU, C. 1995. Protambonitcsprimigenus (Brachiopoda, Clitambonitidina) y ellimite Cambrico-Ordovicico en la Serre de losCabos (Zona Asturoccidental-leonesa, NO deEspana). Espanola de Paleontologia, 10(2),140-150.
WESTROP, S. R. & ADRAIN, J. M. 1998. Trilobite alphadiversity and the reorganization of Ordovicianbenthic marine communities. Paleobiology, 24,1-16.
WILLIAMS, A. 1973. Distribution of brachiopod assem-blages in relation to Ordovician paleogeography.In: HUGHES, N. F. (ed.) Organisms and Conti-nents Through Time. Special Papers in Palaeon-tology, 12, 241-269 (also published as SystematicsAssociation Publication, 9).
WILLIAMS, A. 1976. Plate tectonics and biofacies evol-ution as factors in Ordovician correlation. In:BASSETT, M. G. (ed.) The Ordovician System:Proceedings of a Palaeontological AssociationSymposium, Birmingham, September 1974. Uni-versity of Wales Press and National Museum ofWales, Cardiff, 29-66.
WILLIAMS, A., CARLSON, S. J., BRUNTON, C. H. C.,HOLMER, L. E. & POPOV, L. E. 1996. A supra-ordinal classification of the Brachiopoda. Philo-sophical Transactions of the Royal Society,Biological Sciences, 35, 1171-1193.
WILSON, G. B., PALMER, T. J., GUENSBERG, T. E.,FINTON, C. D. & KAUFMAN, L. E. 1992. Thedevelopment of an early Ordovician hardgroundcommunity in response to rapid sea-floor calciteprecipitation. Lethaia, 25, 19-34.
ZONENSHAIN, L. P., KORINEVSKY, V. G., KlJZMIN, M. I.,PECHERSKY, D. M., KHAIN, V. V. & MATVEENKOV,V. V. 1984. Plate tectonic model of the South Uralsdevelopment. Tectonophysics, 109, 95-135.

This page intentionally left blank

Early Ordovician rhynchonelliformean brachiopod biodiversity:comparing some platforms, margins and intra-oceanic sites around
the Iapetus Ocean
DAVID A.T. HARPER1 & CONALL MAC NIOCAILL2
1 Geological Museum, Oster Voldgade 5-7, DK-1350 Copenhagen, Denmark(e-mail:[email protected]
2Department of Earth Sciences, University of Oxford, Parks Road, Oxford, UK(e-mail:[email protected]
Abstract: During the Arenig-Llanvirn interval a series of radiations across all the majorclades, established the Brachiopoda as a major component of the Palaeozoic benthos. Radi-ations on Baltica and Laurentia during the Arenig formed the basis for two distinct bio-geographical provinces with contrasting articulated brachiopod faunas. These platformprovinces were supplemented by the marginal Celtic and Toquima-Head provinces; thelatter included both marginal and intra-ocean island sites. These marginal and intra-oceanicsites may have served as both 'cradles and museums' alternately providing sources for radi-ations on the platforms and refugia for otherwise relict taxa. Such terranes also partitionedoceanic circulation patterns within the Iapetus Ocean and provided mosaics of rapidlychanging, nearshore unstable environments. In contrast to later Ordovician brachiopodfaunas, many early Ordovician genera are reported from only one or two sites in the Iapetusregion; narrow geographical ranges are characteristic of many taxa. The strong biogeo-graphic differentiation at the generic level is less marked at the familial and higher levelssuggesting a series of late Cambrian and early Ordovician migrations prior to the moreregional development of the Arenig-Llanvirn brachiopod biofacies across the Iapetusterranes. However, the origination of many brachiopod taxa apparently occurred outsidethe lapetus region suggesting that the initial stages of the Ordovician radiations here werefirst spiked by a series of immigrations.
The Ordovician brachiopod faunas of theGreater Iapetus Region include arguably thebest-documented assemblages of the period.The faunas are biostratigraphically and taxo-nomically well constrained while their palaeo-environmental and palaeogeographical settingshave been significantly refined during the last 30years. Recent reviews have emphasized theresolution of current palaeogeographicalmodels (e.g. Cocks 2000, 2001) and the qualityof faunal data available for the early Ordovician(e.g. Harper et al. 1996). Harper & Sandy (2001)have placed Ordovician provinciality within thecontext of the changing biogeographicalpatterns of the entire Palaeozoic. During theearly Ordovician, brachiopod provinciality wasmarked in comparison with subsequent Silurianfaunas (Boucot 1990); comparative provincial-ity was, however, re-established during themid-Devonian (Boucot 1993, 1999). Thisenhanced biogeographical differentation wasassociated with high diversities which togetherprovided opportunities to test palaeogeo-graphical models for the mid-Devonian(McKerrow et al. 2000), similar to those pro-vided by early Ordovician faunas. But although
data for Lower Ordovician faunas are charac-teristically dispersed and uneven, current infor-mation is sufficient to frame and test variousmodels for the early Ordovician radiationwithin the Iapetus region.
Ordovician radiation
The Ordovician radiation is now firmly recog-nized as one of the most significant events inmetazoan history (Droser & Sheehan 1997). Thedetailed anatomy of the radiation is currently anarea of intense investigation. New Bambachianmegaguilds (Droser et al. 1997), associated withdeep burrowing and complex tiering, arefeatures of the radiation together with the estab-lishment of new community types as well as theappearance of stromatoporoid reef structures(Bottjer et al. 2001). In addition, the capacity ofeach Bambachian megaguild was expanded totheir Palaeozoic levels during a relatively shortperiod of time.
Initial detailed research, mainly in the GreatBasin of Laurentia (see Droser & Sheehan 1997for summary), has indicated the importance ofthe faunal turnover at the base of the Whiterock
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194, 25-34.0305-8719/02/$15.00 © The Geological Society of London 2002.
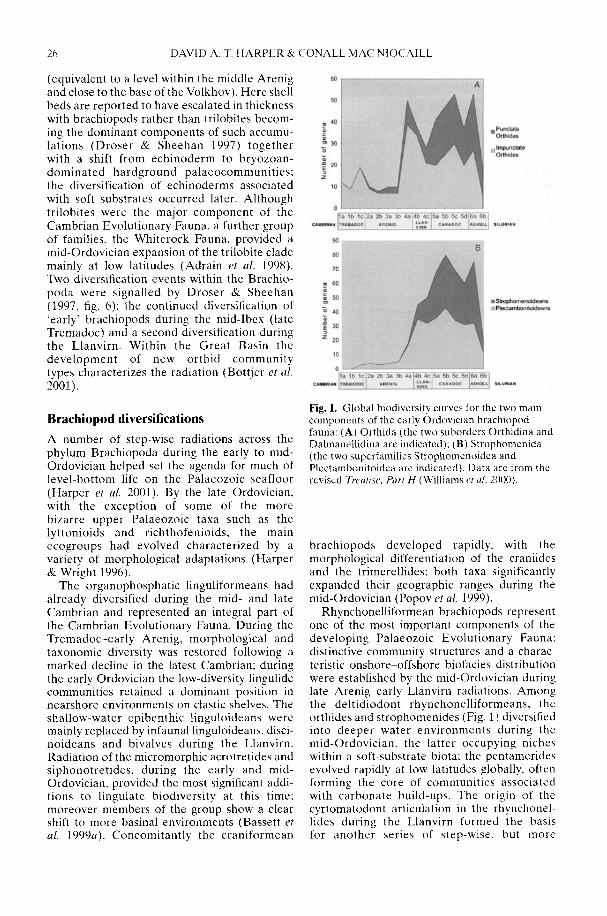
26 DAVID A. T. HARPER & CONALL MAC NIOCAILL
(equivalent to a level within the middle Arenigand close to the base of the Volkhov). Here shellbeds are reported to have escalated in thicknesswith brachiopods rather than trilobites becom-ing the dominant components of such accumu-lations (Droser & Sheehan 1997) togetherwith a shift from echinoderm to bryozoan-dominated hardground palaeocommunities;the diversification of echinoderms associatedwith soft substrates occurred later. Althoughtrilobites were the major component of theCambrian Evolutionary Fauna, a further groupof families, the Whiterock Fauna, provided amid-Ordovician expansion of the trilobite clademainly at low latitudes (Adrain et al. 1998).Two diversification events within the Brachio-poda were signalled by Droser & Sheehan(1997, fig. 6): the continued diversification of'early' brachiopods during the mid-Ibex (lateTremadoc) and a second diversification duringthe Llanvirn. Within the Great Basin thedevelopment of new orthid communitytypes characterizes the radiation (Bottjer et al.2001).
Brachiopod diversificationsA number of step-wise radiations across thephylum Brachiopoda during the early to mid-Ordovician helped set the agenda for much oflevel-bottom life on the Palaeozoic seafloor(Harper et al. 2001). By the late Ordovician,with the exception of some of the morebizarre upper Palaeozoic taxa such as thelyttonioids and richthofenioids, the mainecogroups had evolved characterized by avariety of morphological adaptations (Harper& Wright 1996).
The organophosphatic linguliformeans hadalready diversified during the mid- and lateCambrian and represented an integral part ofthe Cambrian Evolutionary Fauna. During theTremadoc-early Arenig, morphological andtaxonomic diversity was restored following amarked decline in the latest Cambrian; duringthe early Ordovician the low-diversity lingulidecommunities retained a dominant position innearshore environments on clastic shelves. Theshallow-water epibenthic linguloideans weremainly replaced by infaunal linguloideans, disci-noideans and bivalves during the Llanvirn.Radiation of the micromorphic acrotretides andsiphonotretides, during the early and mid-Ordovician, provided the most significant addi-tions to lingulate biodiversity at this time;moreover members of the group show a clearshift to more basinal environments (Bassett etal. 1999a). Concomitantly the craniformean
Fig. 1. Global biodiversity curves for the two maincomponents of the early Ordovician brachiopodfauna: (A) Orthida (the two suborders Orthidina andDalmanellidina are indicated): (B) Strophomenida(the two superfamilies Strophomenoidea andPlectambonitoidea are indicated). Data are from therevised Treatise, Part H (Williams el al. 2000).
brachiopods developed rapidly, with themorphological differentiation of the craniidesand the trimerellides; both taxa significantlyexpanded their geographic ranges during themid-Ordovician (Popov et al. 1999).
Rhynchonelliformean brachiopods representone of the most important components of thedeveloping Palaeozoic Evolutionary Fauna:distinctive community structures and a charac-teristic onshore-offshore biofacies distributionwere established by the mid-Ordovician duringlate Arenig-early Llanvirn radiations. Amongthe deltidiodont rhynchonelliformeans, theorthides and strophomenides (Fig. 1) diversifiedinto deeper water environments during themid-Ordovician, the latter occupying nicheswithin a soft-substrate biota; the pentameridesevolved rapidly at low latitudes globally, oftenforming the core of communities associatedwith carbonate build-ups. The origin of thecyrtomatodont articulation in the rhynchonel-lides during the Llanvirn formed the basisfor another series of step-wise, but more

EARLY ORDOVICIAN BRACHIOPOD BIODIVERSITY 27
restricted, radiations during the mid- to lateOrdovician (Bassett et al. 1999b); ecologicalcompetition and displacement both within thecyrtomatodonts and between the deltidiodontswas marked. Among the spire-bearing brachio-pods only the atrypides diversifed during theCaradoc in the tropics, whereas the athyrididesand the spiriferides were not numerically signifi-cant components of the Ordovician radiations.The pentamerides and spire-bearers were toform the basis of subsequent radiations duringthe early Silurian, commonly associated withcarbonate environments (Rong & Harper 2000;Harper & Rong 2001).
During the early Ordovician the contrastingescalation of the pedunculate orthides and therecumbent strophomenides was the mostmarked amongst the rhynchonelliformeans.Global datasets suggest initial (late Arenig-early Llanvirn) radiations were dominated bythe orthides whereas the strophomenides firstdiversified during the early Llanvirn but mostmarkedly during the Caradoc (Harper et al.19990).
The 'Greater Iapetus Region'The Iapetus Ocean was bordered by a number ofplatform provinces and punctuated by a varietyof marginal and oceanic terranes. This sector ofthe Earth's crust has provided a critical field areafor the understanding of the dynamics andevolution of Early Palaeozoic brachiopodfaunas. Terrane models for most parts of theregion are relatively far advanced (Harper 1998)and the extent of most continents and theirmargins are now well defined. The margins of,for example, Avalonia (Cocks et al. 1997) andBaltica (Cocks & Fortey 1998) have beendescribed in detail.
Perspectives on the development of theIapetus Ocean system have changed markedlyduring the past three decades. The innovativeorthogonal model for oceanic opening andclosure (Wilson 1966), which has formed thebasis for the Wilson Cycle, indicated the separ-ation of North American and Europeanbrachiopod faunas during the Early Palaeozoic.Statistical analyses of a range of brachiopodfaunas mainly associated with the Iapetus region(Williams 1969, 1973) suggested a number ofprovinces existed within the Greater Iapetusregion. Nevertheless, as closure of the IapetusOcean progressed sequentially less mobileorganisms were able to cross a narrowingseaway (McKerrow & Cocks 1976); for example,many brachiopod genera were present on bothsides of the ocean by the Caradoc whereas
similar brachiopod species were present in bothEurope and North America during the Ashgill(McKerrow & Cocks 1976). During the early tomid-1980s a series of more complex models wasdeveloped for the ocean emphasizing, forexample, the role of decreasing endemicity withdepth (Cocks & Fortey 1982) and the partici-pation of a more complex system of continents,including Laurentia, Avalonia and Baltica,within the cycle (Cocks & Fortey 1982; Fortey &Cocks 1986). Each continent, and to a lesserextent its margins, contained a distinctive earlyOrdovician brachiopod fauna defining aprovince and suggesting oceanic separationfrom coeval continental areas; distinctionsbecome less clear during and following the earlyCaradoc gracilis transgression.
Neuman, in a series of papers (Neuman 1972,1984; Neuman & Harper 1992), added greaterrealism to these palaeogeographical reconstruc-tions with the addition of islands and archipela-gos into otherwise barren oceanic tracts, basedon data initially from the Appalachians. Theimportance of marginal and oceanic sites wasconfirmed and developed, on the basis of datafrom Scandinavia, by Bruton & Harper (1981,1985). More focused statistical analyses of thebrachiopod faunas from around and within thelapetus Ocean (Neuman & Harper 1992; Harperet al. 1996) confirmed the status of the platformprovinces but also flagged groups of marginaland ocean biotas associated with islands com-posed of both basement and volcanic material.For example, the Toquima-Table Head faunasdeveloped circumferal and seaward of Laurentiaat low latitudes whereas the Celtic faunasformed a high-latitude belt marginal to Gond-wana (Harper et al. 1996). These models havebeen tested by seriation, using simulated anneal-ing (Ryan et al. 1999). This algorithm clearlyrecognized clusters of associated faunas but wasless successful in clarifying the relationshipsbetween the groupings. Possibly many of theearly Ordovician high-latitude faunas weremore isolated from coeval faunas elsewherehaving fewer widespread taxa and moreendemics than those faunas from low latitudes.That situation contrasts with the pattern for thelatest Ordovician, when a clearer latitudinalgradient across more cosmopolitan faunas isapparent (Ryan et al. 1999).
Palaeogeographical templates for the earlyOrdovician of the Iapetus region have increas-ingly emphasized the role of marginal andoceanic tracts now entrained as a variety ofterranes within the Appalachian-Caledonianmountain belt. The biological importance ofthese sites has been specified in a number of

28 DAVID A. T. HARPER & CONALL MAC NIOCAILL
Fig. 2. Palaeogeographical reconstructions for (A) early Arenig and (B) the mid-Llanvrin. The early Arenig tomid-Llanvirn interval highlights the development of a series of peri-Gondwanan terranes, many forming thebasis for the Celtic Province. Laurentia has been positioned according to the palaeomagnetic compilation ofMac Niocaill & Smethurst (1994); Baltica and Gondwana after Torsvik et al. (1996): Siberia after Smethurst etal. (1998); Avalonia has been palaeolatitudinally positioned after Mac Niocaill (2000). The locations of intra-lapetan arcs are based on Mac Niocaill et al. (1997) and van Staal et al. (1998). Abbreviations used are asfollows: Hol. Holonda; DA, Dashwoods block; Precord, Precordillera; Gand, Gander terrane: Carol. Carolinaterrane; ATA, Armorican Terrane Assemblage, the individual elements being as follows: Ib. Iberia: M.Meguma; Arm. Armorica; Boh, Bohemia [= Perunica]: Am. Alpine Massifs.
contributions (Neuman 1972, 1976; Bruton &Harper 1981, 1985; Harper et al. 1996); roles ascentres for speciation, refugia during regres-sion and stepping stones along migrationalroutes have been proposed. A number ofislands clearly changed their faunal affinitiesduring cross-latitudinal movement (Owen et al.1992).
The addition of realistic ocean-circulationpatterns (Christiansen & Stouge 1999) hasenhanced understanding of the mechanics of thelapetus oceanic system. Predicted warm- andcool-water gyres may have also provided apartition between the low-latitude Laurentianand Toquima-Table Head faunas and thoseat high latitudes associated with Avalonia,Gondwana and the Celtic group of faunas. Thepresence of island chains within the IapetusOcean may have helped develop the interfacebetween the two current systems.
Platforms, margins and intra-oceanic sites
The early Ordovician Iapetus region contains aspectrum of continental and ocean terranes in avariety of tectonic settings (Fig. 2). The largerplatform areas of Laurentia, Baltica and
Gondwana are supplemented by marginal sitessuch as the Oslo region and Western Newfound-land and microcontinents such as Avalonia andPerunica. Although there are abundant brachio-pod data from many of these sites there are fewregional databases. Preliminary databases, atthe generic level, have been established here asan initial test for a number of hypotheses regard-ing the early Ordovician brachiopod radiation.The East Baltic (Harper & Hints 2001).Morocco (Havlicek 1971), Welsh Basin(Lockley 1983) and the Prague Basin (Havlicek1976, 1998) have been used as proxies for theBaltic, Gondwanan, Avalonian and Bohemianbiogeographic units. Nevertheless such areas,for example Baltica, contain a range of facies.and initial studies (e.g. Jaanusson 1973, 1976;Harper 1986) have emphasized that the variousfacies belts reacted in different ways to changinglatitude and waves of immigrants.
Critical to the early Ordovician radiations isthe development of marginal and intra-oceanicterranes and their roles in the development,dispersal and survival of taxa, the partition ofoceanic circulation patterns within the IapetusOcean and the provision of mosaics of rapidlychanging, nearshore unstable environments.

EARLY ORDOVICIAN BRACHIOPOD BIODIVERSITY 29
Harper et al. (1996) identified nearly 20 earlyOrdovician intra-oceanic and marginal siteswithin the Iapetus region. A range of multi-variate analyses confirmed the coherence of theToquima-Table Head (low latitude) and Celtic(high latitude) groups of faunas. A numberof authors have emphasized the biologicalimportance of these sites (Neuman 1972,1984; Bruton & Harper 1981, 1985; Fortey 1984;Harper 1992; Harper et al. 1996). These marginaland intra-oceanic sites may have served as both'cradles' and 'museums' alternately providingsources for radiations on the platforms andrefugia for otherwise relict taxa.
The early Ordovician rhynchonelliformeanbrachiopod fauna is dominated by billingsellides(including the clitambonitidines) and orthides.Analysis of the distribution of these taxaacross the Celtic and Toquima-Table Head belts,targeting the marginal and intra-oceanic sites,suggests that 32% (N = 45) of the brachiopods atthese sites were endemic to either a single local-ity or to a group of adjacent localities (Fig. 3).Moreover 30% of the taxa represent the firstoccurrence of a genus and 25% the last. Only13% of the occurrences represent within-rangespikes. The available data tend to confirm therole of these sites as the locus for endemics andpossible roles as cradles and refugia within theIapetus Ocean system.
Early to mid-Ordovician development ofthe Iapetus brachiopod faunaThe initial break-up of the northern margins ofGondwana and the arcs around Laurentiacreated a diversity of terranes (Fig. 2). Thisdisparate array of terranes formed an idealenvironment for the development of gammadiversity (inter-province) across the IapetusOcean. The isolation of these terranes wasenhanced by the late Arenig regression, whenthere was a concentration of island biotas(Fortey 1984). Many islands and archipelagosmay have been emergent, with narrow shelfareas with relatively small populations. Themajority of brachiopod faunas inhabitedshallow-water, nearshore environments, prob-ably no deeper than Benthic Assemblage zones3-4 (Cocks 1996). By comparison with thenon-articulates, dispersion of the rhynchonelli-formeans was relatively limited; the latterprobably possessed relatively short, nektoben-thic larval stages restricting the rapid spread ofmany articulated groups (Harper & Sandy2001).
Subsequent transgressions promoted migra-tion and the subsequent colonization of the shelf
Fig. 3. Proportions of endemics, first, last and mid-range occurrences of billingsellide and orthide generaacross the marginal Celtic and Table Head faunas.Data are modified from Harper et al. (1996).
areas of the platform provinces (Figs 4 and 5).These mechanisms provided for the escalationof alpha diversity (within-community) as estab-lished regional palaeocommunities were supple-mented by immigrants. These diversificationswere most marked in shallow-water palaeo-communities, for example in the inner,nearshore facies of Avalonia and the Balticprovince (Harper & Hints 2001). This diversityspike is most obvious along the margins ofBaltica, for example in the Oslo Region (Harper1986). The faunas were dominated by largerpedunculate orthidine brachiopods. In additionto sea-level changes the approach of terranes,particularly from the peri-Gondwanan region,may have provided additional taxa to participatein these radiations (Homier et al 2000). Thisprocess of integration has also been suggestedfor the development of the Celtic terranes(Bruton & Harper 1985).
The most marked radiations, however,occurred during the Caradoc (Figs 4 and 5). Thisdiversity spike is clear in global databases butis also obvious in the regional datasets forAvalonia, Baltica, Gondwana and Perunica.These later diversifications may have beenassociated with beta diversity changes as avariety of new palaeocommunities inhabiteddeep-water environments with a greaterdominance of the recumbent strophomenides(Harper et al. 1999b; Rong et al. 1999).
Comparison of early and late Ordovicianbrachiopod biofaciesIn contrast to later Ordovician brachiopodfaunas, a large number of early Ordoviciangenera are reported from only one or two sitesin the Iapetus region; narrow geographicalranges are characteristic of many early Ordo-vician taxa (Fig. 6). The distribution patterns ofthese taxa show a positive skewness, suggesting

30 DAVID A. T. HARPER & CONALL MAC NIOCAILL
Fig. 4. Diversity profiles (unstandardized numbers of genera) through (A) the shallow-water North Estonianconfacies belt and (B) the deeper-water Livonian Tongue (equivalent to the Central Baltoscandian confaciesbelt of Jaanusson 1976). Sea-level stands are calibrated in terms of Benthic Assemblage zones. Data aremodified from Harper & Hints (2001).

EARLY ORDOVICIAN BRACHIOPOD BIODIVERSITY 31
Fig. 5. Diversity profiles (unstandardized numbers of genera) from Avalonia (based on sources in Lockley(1983) and some additional data), part of the Gondwanan margin (based on Havlicek 1971) and Perunica(based on sources in Havlicek 1998).
that the majority of taxa are in fact reportedfrom only one or two sites. This pattern contrastswith that for the late Ordovician where a lessskewed distribution indicates fewer endemictaxa and more widespread genera. Seriation ofthese sites has emphasized these contrastingspatial patterns (Ryan et al. 1999). EarlyOrdovician faunas were successfully arrangedinto cohesive sets representing the main provin-cial groupings; the links between these groupswere less clear. Faunal gradients are muchclearer during the late Ordovician when a seriesof overlapping provinces can be recognized fromthe high-latitude Bani province, through theKosov province to the low-latitude Edgewoodprovince (Rong & Harper 1988; Owen et al.1991; Ryan et al. 1999).
Conclusions
This study emphasizes the role of marginal andoceanic sites in the Ordovician radiation. Intra-and marginal Iapetus Ocean terranes acted as
both cradles and museums for brachiopod taxaduring the early Ordovician (late Arenig to earlyLlanvirn) phases of the diversification. Terranefragmentation, particularly at high latitudes, wasessential for the generation of an initial gammadiversity. Diversification, however, was morepronounced when individual platform com-munities first expanded in size with the develop-ment of alpha diversity, during the Llanvirn.These diversifications may have helped propelthe offshore movement of communities duringthe Caradoc as beta diversity intensified.
During the early Ordovician many brachio-pod families were relatively widespread: bio-geographical signals provided by genericdistributions. This suggests that the origins ofmany Ordovician brachiopod families may betracked back into the Cambrian outside thelapetus region. The circum-Iapetus platformsand margins provided opportunities but not theorigins for a mid-Arenig radiation. The earlyOrdovician dispersion of the continents andassociated outboard terranes provided the

32 DAVID A. T. HARPER & CONALL MAC NIOCAILL
Fig. 6. Comparison of some of the distributionalproperties of early and late Ordovician brachiopodfaunas. Data are modified from sources in Harper etal. (1996) and Owen et al. (1991).
conditions for geographic isolation andspeciation. Migration of taxa onto the platformsduring the early Llanvirn generated the firstincreases in within-community diversity thatmay have driven the development of new,particularly deeper-water, community typesespecially during the Caradoc,
D.A.T.H. thanks the Danish Natural Science Council(SNF) for financial support. This paper is a contri-bution to IGCP 410 The Great Ordovician Biodiver-sification Event'. Helpful comments from T.McCormick, A. Owen and an anonymous reviewerimproved the manuscript.
ReferencesADRAIN, J. M.. FORTEY, R. A. & WESTROP, S. R. 1998.
Post-Cambrian trilobite diversity and evol-utionary faunas. Science, 280, 1922-1925.
BASSETT, M. G., POPOV, L. E. & HOLMER, L. E. 1999a.Organophosphatic brachiopods: Patterns of bio-diversification and extinction in the Early Palaeo-zoic. Geobios, 32, 145-163.
BASSETT, M. G, POPOV, L. E. & SOKIRIN, E. V. 1999b.Patterns of diversification in Ordovician cyr-tomatodont rhynchonellate brachiopods. ActaUniversitatis Carolinae, Geologica. 43, 329-332.
BOTTJER. D. J., DROSER, M. L., SHEEHAN, P. M.& MCGHEE JR., G. R. 2001. The ecological
architecture of major events in the Phanerozoichistory of marine invertebrate life. In: A lLMON.W. D. & BOTtjER. D. J. (eds) EvolutionaryPaleoecology. Columbia University Press. NewYork, 35-61".
Borcor, A. J. 1990. Silurian biogeography. In: McK-ERROW. W. S. & SCOTESE. C R. (eds) PalaeozoicPalaeogeography and Biogeography. GeologicalSociety. London. Memoir, 12. 191-196.
BOUCOT, A. J. 1993. Paleozoic paleogeography andbiogeography. Revista Espahola de Paleontologia,No Extraordinario. 15-20.
BoucOT. A. J. 1999. Southern African Phanerozoicmarine invertebrates: biogeography. pateoecol-ogy. climatology and comments on adjacentregions. Journal of African Earth Sciences. 28,129-143.
BRuTON, D. L. & HARPER, D. A.T. 1981. Brachiopodsand trilobites of the early Ordovician serpentineOtta Conglomerate, south central Norway. NorskGeologisk Tidsskrift. 61.151-181.
BRITON, D. L. & HARPER, D. A. T. 1985. EarlyOrdovician (Arenig-Llanvirn) faunas fromoceanic islands in the Appalachian-Caledonianorogen. In: GEE, D. G. & STuRT. B. A. (eds)The Caledonian Orogen - Scandinavia andRelated Areas. John Wiley & Sons. Chichester.359-368.
CHRISTIANSEN, J. L. & STOUGE. S. 1999. Using palaeo-oceanical modelling in reconstructing EarlyOrdovician palaeogeography. Acta UniversitatisCarolinae, Geologica, 43. 515-518.
COCKS, L. R. M. 1996. Lower Palaeozoic brachio-pod communities. In: COPPER. P. & JIN Jsuo(eds) Brachiopods. A. A. Balkema. Rotterdam.69-71.
COCKS, L. R. M. 2000. The Early Palaeozoic geographyof Europe. Journal of the Geological Society,London. 157, 1-10.
COCKS. L. R. M. 2001. Ordovician and Silurian globalgeography. Journal of the Geological Society,London. 158, 197-210.
COCKS, L. R. M. & FORtEY. R. A. 1982. Faunal evidencefor oceanic separation in the Palaeozoic ofBritain. Journal of the Geological Society,London, 139, 465-478.
COCKS. L. R. M. & FORTEY, R. A. 1998. The LowerPalaeozoic margins of Baltica. GFF. 120. 173-179.
COCKS, L. R. M., MCKERROW. W. S. & VAN STAAL. C. R.1997. The margins of Avalonia. Geological Maga-zine. 134. 627-636.
DROSER, M. L. & SHEEHAN, P. M. 1997. Palaeoecologyof the Ordovician radiation: resolution of large-scale patterns within individual clade histories,palaeogeography and environments. Geobios. 20.221-229.
DROSER, M. L., BOTtjER, D. J. & SHEEHAN. P. M. 1997.Evaluating the ecological architecture of majorevents in the Phanerozoic history of marine inver-tebrate life. Geology. 25. 167-170.
FORTEY, R. A. 1984. Global earlier Ordovician trans-gressions and regressions and their biologicalimplications. In: BRUTON. D. L. (ed.) Aspectsof the Ordovician System. Palaeontological

EARLY ORDOVICIAN BRACHIOPOD BIODIVERSITY 33
Contributions from the University of Oslo, 295,37-50.
FORTEY, R. A. & COCKS, L. R. M. 1986. Marginal faunalbelts and their stratigraphical implications.Journal of the Geological Society, London, 143,151-160.
HARPER, D. A.T. 1986. Distributional trends within theOrdovieian brachiopod faunas of the Oslo Region,south Norway. In: RACHEBOEUF, P. R. & EMIG, C.C. (eds) Les Brachiopodes fossils et actuels. Bios-tratigraphie du Paleozoique, 4, 465–475.
HARPER, D. A. T. 1992. Ordovieian provincial signalsfrom Appalachian-Caledonian terranes. TerraNova, 4, 204-209.
HARPER, D. A. T. 1998. Interpreting erogenic belts:principles and examples. In: DOYLE, P. &BENNETT, M. R. (eds) Unlocking the Strati-graphical Record: Advances in Modern Stratigra-phy. John Wiley & Sons, Chichester, 491-524.
HARPER, D. A. T. & HINTS, L. 2001. Distribution anddiversity of Ordovieian articulated brachiopods inthe East Baltic. In: BRUNTON, C. H. C, COCKS, L.R. M. & LONG, S. (eds) Brachiopods Past andPresent. Taylor & Francis, New York & London,315-326.
HARPER, D. A. T. & RONG JiA-Yu 2001. Palaeozoicbrachiopod extinctions, survival and recovery:patterns within the rhynchonelliformeans. Geo-logical Journal, 36, 317-328.
HARPER, D. A. T. & SANDY, M. R. 2001. Paleozoicbrachiopod biogeography. In: CARLSON, S. J. &SANDY, M. R. (eds) Brachiopods Ancient andModern. A Tribute to G. Arthur Cooper. Paleon-tological Society Papers, 7, 207-222
HARPER, D. A.T. & WRIGHT, A. D. 1996. Brachiopods.In: HARPER, D. A. T. & OWEN, A. W. (eds) Fossilsof the Upper Ordovieian. PalaeontologicalAssociation, London, 63-94.
HARPER, D. A.T, MAC NIOCAILL, C. & WILLIAMS, S. H.1996. The palaeogeography of early OrdovieianIapetus terranes: an integration of faunal andpalaeomagnetic constraints. Palaeogeography,Palaeoclimatology, Palaeoecology, 121, 297-312.
HARPER, D. A. T., RONG JiA-Yu & SHEEHAN, P. M.1999a. Ordovieian diversity patterns in earlyrhynchonelliform (protorthide, orthide andstrophomenide) brachiopods. Acta UniversitatisCarolinae, Geologica, 43, 325-327.
HARPER, D. A.T., RONG JIA-Yu AND ZHAN R E N - B I N1999b. Late Ordovieian development of deep-water brachiopod faunas. Acta UniversitatisCarolinae, Geologica, 43, 351-353.
HARPER, D. A. T, BASSETT, M. G., COCKS, L. R. M.,COPPER, P., ET AL. 2001. Ordovieian brachiopoddiversity. Paleobios, 21 (2nd Supplement), 5-6.
HAVLICEK, V. 1971. Brachiopodes de L'Ordovicien duMaroc. Notes et Memoires du Service Geologique,230, 1-135.
HAVLICEK, V. 1976. Evolution of Ordovieian brachio-pod communities in the Mediterranean province.In: BASSETT, M.G. (ed.) The Ordovician System.University of Wales Press and the Palaeontologi-cal Association, 349-358.
HAVIJCEK, V. 1998. Ordovieian. In: CHULPAC, I.,
HAVLICEK, V. K R I Z , J., KUKAL, Z. & STORCH, P.(eds) Palaeozoic of the Barrandian (Cam-brian-Devonian). Czech Geological Survey,Prague, 41-79.
HOLMER, L. E., POPOV, L. E. & BASSETT, M. G. 2000.Early Ordovician organophosphatic brachiopodswith Baltoscandian affinities from the AltayRange, southern Krygyzstan, GFF, 122, 367-375.
JAANUSSON, V. 1973. Ordovician articulate brachio-pods. In: HALLAM, A. (ed.) Atlas of Palaeobio-geography, Elsevier, Amsterdam, 19-26.
JAANUSSON, V. 1976. Faunal dynamics in the MiddleOrdovician (Viruan) of Baltoscandia. In:BASSETT, M. G. (ed.) The Ordovician System. Uni-versity of Wales Press and the PalaeontologicalAssociation, 301-326.
LOCKLEY, M. G. 1983. A review of the brachiopoddominated palaeocommunities from the typeOrdovician. Palaeontology, 26, 111-145.
MCKERROW, W. S. & COCKS, L. R. M. 1976. Progressivefaunal migration across the Iapetus Ocean.Nature, 263, 304-306.
MCKERROW, W. S., MAC NIOCAILL, C., AHLBERG, P. E.,CLAYTON, G., CLEAL, C. J. & EAGAR, R. M. C 2000.The Late Palaeozoic relations between Gond-wana and Laurussia. In: FRANKE, W., HAAK, V.,ONCKEN, O. & TANNER, D. (eds) Orogenic Pro-cesses: Quantification and Modelling in theVariscan Belt. Geological Society, London,Special Publications, 179, 9-20.
MAC NIOCAILL, C. 2000. A new Silurian palaeolatitudefor eastern Avalonia and evidence for crustalrotations in the Avalonian margin of southwest-ern Ireland. Geophysical Journal International,141,661-671.
MAC NIOCAILL, C. & SMETHURST, M. A. 1994. Palaeo-zoic palaeogeography of Laurentia and itsmargins: a reassessment of palaeomagnetic data.Geophysical Journal International, 116, 715-725.
MAC NIOCAILL, C., VAN DER PLUIJM, B. A. & VAN DERVoo, R. 1997. Ordovician paleogeography and theevolution of the Iapetus Ocean. Geology, 25,159-162.
NEUMAN, R. B. 1972. Brachiopods of early Ordovicianvolcanic islands. International Geological Con-gress, Montreal, Section, 7, 297-302.
NEUMAN, R. B. 1976. Early Ordovician (Late Arenig)brachiopods from Virgin Arm, New World Island,Newfoundland. Bulletin of the Geological Surveyof Canada, 261, 11-61.
NEUMAN, R. B. 1984. Geology and paleobiology ofislands in the Ordovician Iapetus Ocean. Geo-logical Society of America, Bulletin, 95, 1188-1201.
NEUMAN, R. B. & HARPER, D. A. T. 1992. Paleogeo-graphic significance of Arenig-LlanvirnToquima-Table Head and Celtic assemblages. In:WEBBY, B. D. & LAURIE, J. R. (eds) GlobalPerspectives in Ordovician Geology. Balkema,Rotterdam, 241-254.
OWEN, A. W., HARPER, D. A. T. & RONG JIA-Yu 1991.Hirnantian trilobites and brachiopods in spaceand time. In: BARNES, C. & WILLIAMS, S.H. (eds.)Advances in Ordovician Geology. GeologicalSurvey of Canada, Paper 90-9, 179-190.

34 DAVID A. T. HARPER & CONALL MAC NIOCAILL
OWEN, A. W., HARPER, D. A. T. & ROMANO, M. 1992.The Ordovician biogeography of theGrangegeeth terrane and the position of theIapetus suture in eastern Ireland. Journal of theGeological Society, London, 149, 3-6.
POPOV, L.E., BASSETT, M.G., HOLMER, L.E. & GORJAN-SKY, V. Yu. 1999. Ordovician patterns of diversifi-cation in craniiformean brachiopods. ActaUniversitatis Carolinae, Geologica, 43, 321-324.
RONG JIA-Yu & HARPER, D. A. T. 1988. A global syn-thesis of the latest Ordovician Hirnantianbrachiopod fauna. Transactions of the RoyalSociety of Edinburgh: Earth Sciences, 79, 383–402.
RONG JIA-Yu & HARPER, D. A. T. 2000. Brachiopodsurvival and recovery from the latest Ordovicianmass extinctions. Geological Journal, 34, 321-348.
RONG JIA-Yu, ZHAN REN-BIN & HARPER, D. A. T.1999. Late Ordovician (Caradoc-Ashgill)brachiopod faunas with Foliomena based on datafrom China. Palaios, 14, 412-431.
RYAN, P. D., RYAN, M. & HARPER, D. A. T. 1999. Seri-ation: a new approach. In: HARPER, D.A.T. (ed.)Numerical Palaeobiology. John Wiley & Sons,Chichester, 433-449.
SMETHURST, M. A., KHRAMOV, A. & TORSVIK, T. H.1998. Palaeomagnetic constraints on the Neopro-terozoic-Palaeozoic drift history of the SiberianPlatform: Rodinia to Pangea. Earth ScienceReviews, 43, 1-24.
TORSVIK,T. H., SMETHURST, M. A., MEERT, J. G., ET AL.1996. Continental break-up and collision in the
Neoproterozoic and Palaeozoic: A tale of Balticaand Laurentia. Earth Science Reviews, 40.229-258.
VAN STAAL, C. R., D E w e Y . J. F., MAC NIOCAILL. C. &MCKERROW. W. S. 1998. The tectonic evolutionof the Northern Appalachians and BritishCaledonides: history of a complex, west andsouthwest Pacific-type segment of Iapetus. In:B l u n D E L L . D.J. & Scon. A.C. (eds) Lyell:the Past is the Key to the Present. GeologicalSociety. London, Special Publications. 143.199-242.
WILLIAMS. A. 1969. Ordovician faunal provinces withreference to brachiopod distribution. In: WOOD.A. (ed.) The Pre-Cambrian and Lower PalaeozoicRocks of Wales. University of Wales Press.Cardiff. 117-154.
WILLIAMS, A. 1973. Distribution of brachiopod assem-blages in relation to Ordovician palaeogeogra-phy. In: HUGHES, N. F. (ed.) Organisms andContinents through Time. Special Papers inPalaeontology. 12, 241-269.
WILLIAMS, A., BRUNTON,C. H. C. & CARLSON. S. J. (eds)2000. Treatise on Invertebrate Paleontology, PartH, Brachiopoda revised. Volume 2: Linguli-fonnea, Craniiformea, and Rhynchonelliformea(part). Geological Society of America and Uni-versity of Kansas Press. Boulder. Colorado andLawrence, Kansas.
WILSON, J.T. 1966. Did the Atlantic close then reopen?Nature, 211. 676-681.

Diversification and biogeography of bivalves during the OrdovicianPeriod
JOHN C. W. COPEDepartment of Earth Sciences, Cardiff University, PO Box 914, Cardiff CF10 3YE, UK
(e-mail: [email protected])
Abstract: Bivalves have a wide distribution in the Lower and to a lesser extent MiddleCambrian rocks, but they have not yet been certainly identified in the Upper Cambrian.Recent discoveries have significantly increased our knowledge of Lower Ordovicianbivalve faunas and their explosive radiation from the Early Ordovician apparentlycoincides with the evolution of the feeding gill. Early Ordovician faunas were confined tothe siliciclastic facies of Gondwanan shelf seas; most genera were clearly latitudinallyconstrained, but others apparently migrated over wide latitudes. By the Mid-Ordovician,bivalves had begun to escape the confines of Gondwana and marked latitudinal differencesin the composition of the faunas became apparent, with pteriomorphians showing clearpreference for low latitudes, whilst heteroconchs clearly preferred median to high latitudes;surprisingly, nuculoids were both most diverse in terms of species and most abundant as apercentage of individuals within the bivalve populations at low latitudes. It was in the LateOrdovician that bivalves colonized the low-latitude carbonate platforms of Laurentia andBaltica, leading to a second major diversification particularly within the pteriomorphianbivalves, which developed semi-infaunal and epifaunal habits; they became the dominantlow-latitude bivalve group. The latest-Ordovician eustatic regression which exposed thelow-latitude carbonate platforms resulted in a major reduction in the epifaunal and semi-infaunal bivalves involving extinction of many genera, including the only Ordovician boringbivalves.
Thirty years ago Pojeta (1971) produced hisclassic work on Ordovician bivalves that illus-trated something of the early diversity of thislong-neglected fossil group. At the time of thispublication, some revisionary work had beendone on Late Ordovician faunas from NorthAmerica (e.g. Pojeta 1962, 1966), but save theworks on French faunas of the Montagne Noire(Thoral 1935) and Armorica (Babin 1966) littlework had been done in the first 70 years of thetwentieth century on Early Ordovician bivalvefaunas. Phylogenetic links were necessarilysketchy (Pojeta 1971) and Cambrian bivalveswere of doubtful interpretation. Pojeta's (1971)reservations about Lamellodonta, the earliestbivalve accepted by the Treatise (Cox et al.1969-1971), were confirmed when the genus wasshown to be based upon a distorted brachiopod(Havlicek & Kriz 1978). Pojeta later (1973,1975) went on to demonstrate the bivalve affini-ties of the widely dispersed Lower CambrianFordilla, whilst Jell (1980) described anotherEarly Cambrian genus Pojetaia from Australia,subsequently found in Bornholm, China andMorocco and found to extend into the Mid-Cambrian. Other Mid-Cambrian forms havebeen described from New Zealand (McKinnon1982), Bornholm (Berg-Madsen 1987; Hinz-Schallreuter 1995, 2000) and Morocco (Geyer &
Streng 1998). The lack of bivalves from theUpper Cambrian remains an embarrassinghiatus in the study of early bivalve evolution.Berg-Madsen (1987) recorded a Tuarangia froma Polish erratic that contained conodonts sheregarded as Upper Cambrian, but Hinz-Schall-reuter (2000) pointed out that tricuspidate west -ergaardodinids also occur in the MiddleCambrian. Pojeta (1980) figured a rather fea-tureless shell from the Upper Cambrian ofMaryland, but this single specimen has nouniquely bivalve characteristics. Pojeta (2000)has recently reviewed the Cambrian history ofthe Bivalvia.
Lower Ordovician bivalves are much morediverse, much larger in size and more commonas fossils. Cope (1995, 1997b) correlated thesefactors with the evolution of the feeding gill inthe latest Cambrian or earliest Ordovician.Bivalves remain, however, very rare fossils inLower Ordovician rocks. Thus from theTremadoc, bivalves are known from four areas:Salta Province, Argentina (Harrington 1938);the Montagne Noire (Thoral 1935; Babin1982a); central Australia (Pojeta & Gilbert-Tomlinson 1977; originally described asprobably of Arenig age but later shown to belate Tremadoc by Shergold et al. 1991) andthe Moroccan Anti-Atlas (pers. obs. 2001).
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194, 25-52.0305-8719/02/$15.00 © The Geological Society of London 2002.

36 JOHN C. W. COPE
Altogether some nine or ten species are knownso far from Tremadoc rocks. From the lower partof the Arenig Series, bivalves are known fromSalta Province, Argentina (Harrington 1938);the Montagne Noire (Babin 1982a); Afghanis-tan (Desparmet et al. 1971); originally reportedas Tremadoc but more probably of Arenig age;Babin & Gutierrez-Marco 1991); the MoroccanAnti-Atlas (Babin & Destombes 1992); RamseyIsland, South Wales (Hicks 1873; Carter 1971);and most recently from Llangynog, South Wales(Cope 1996b), the latter locality yielding nofewer than 20 species belonging to 18 genera,representing well over half the species knownhitherto from rocks of this age. After the Arenig,bivalves become commoner and Babin &Gutierrez-Marco (1991) were able to report onMiddle Ordovician bivalves collected from nofewer than 87 localities from Spain alone.
All the Early Ordovician bivalves known arefrom Gondwana and it now seems clearthat they were thus geographically restricted(Avalonia being so close to the Gondwananmargins in Arenig times that, for the purposes ofpalaeobiogeography, it can still be considered aspart of Gondwana). It is assumed that, althoughthey were able to migrate freely around theGondwanan margins, the width of the oceanicareas around Gondwana precluded bivalvelarvae from crossing to other continental shelvesbefore the Mid-Ordovician.
The major radiations of the Early Ordovicianresulted in the appearance of most bivalvegroups by the end of that time. Cope (2000)proposed a revised classification of the Bivalviarecognizing that the division into the subclassesbased on gill grade, Protobranchia Pelseneer,1889 and Autolamellibranchiata Grobben, 1894,long used by zoologists, could now be applied tothe fossil record. The classification of theBivalvia used in this paper is essentially that ofCope (2000) shown in Figure 1. The only differ-ence between that figure and Cope (2000, fig. 2)is that the Nucinelloidea are not shown, as theydo not appear until well after the Ordovician.Continental reconstructions for the Ordovicianare shown in Figure 2.
Protobranchia
Nuculoida
Amongst the protobranchs the Nuculoida arewell represented in the Early Ordovician andrange from low-latitude areas like Australia, tohigh-latitude areas such as the Montagne Noireand the Moroccan Anti-Atlas. The simplestforms, with undifferentiated taxodont dentition,
Fig. 1. Classification of the Bivalvia adopted herein,showing the phylogenetic links between the majorbivalve groups. Modified after Cope (2000).
belong to the family Praenuculidae McAlester,1969 (in Cox et al. 1969-1971). The earliestpraenuculids include forms like Paulinea andPemarnia from the Early Arenig of South Wales(Cope 1996b); the latter also occurs in the LateArenig of the Welsh Borderland (Cope 1999).Some other praenuculids also seem to be ofrestricted geographical range, such as FideraPojeta & Gilbert-Tomlinson, 1977, from theMiddle to Upper Ordovician of Tasmania.However, Praenucula Pfab, 1934 itself occurswidely in the Middle Ordovician of Europe fromBohemia (e.g. P. expansa Pfab, 1934) westwardsto Portugal (e.g. P. riheiro (Sharpe, 1853)); andhas also been recorded from the Lower, Middleand Upper Ordovician of the Moroccan Anti-Atlas (Babin & Destombes 1992). Praenuculaand the praenuculid Palaeoconcha wererecorded from the upper Middle or lower UpperOrdovician of Argentina by Sanchez (1990).Dzik (1994) figured a minute juvenile nuculoidfrom the Lower Llanvirn of Baltica that appearsto be a Praenucula, as indicated by Babin (2000);this is notable in another context, as it is one ofthe earliest bivalves to have reached Baltica.Praenucula also occurs in the Upper Ordovicianof Laurentia and P.filistriata (Ulrich, 1894) fromOhio was figured under the generic name
ORDOVICIAN BIVALVE BIOGEOGRAPHY 37
Fig. 2. Continental reconstructions for the Ordovician. (a) Early Ordovician. Note near-polar position of theMontagne Noire and the lower-latitude position of Avalonia which is still on the Gondwanan margin. Contrastwith the lower latitudes of Argentina and Australia, (b) Mid-Ordovieian. Note lower-latitude position ofAvalonia, whilst Armorica stays at polar latitudes, (c) Late Ordovician. Note positions of Baltica andLaurentia, both areas of carbonate platforms.Key to abbreviations: Ar, Arrnorica; Av, Avalonia; B, Baltica; Bh, Bohemia; G, Gondwana; K, Kazakhstania;La, Laurentia; S, Siberia; SC, South China; U, East Uralia. Reconstructions principally based on those ofTorsvik (1998) with modifications by L. E. Popov.
Deceptrix by Pojeta (1978 p1. 2, figs 1-2), butCope (1997b) showed that the latter genus was acardiolariid and not a praenueulid (see below).Some of the species figured previously asDeceptrix, in addition to some of those figuredby Pojeta (1971, 1978), including those figuredby Tunnicliff (1982) and Cope (1996a), are
now recognized as praenuculids and notcardiolariids. For those species Cope (1997b)proposed the praenueulid genus Homilodontawith Arca subtruncata Portlock, 1843 as typespecies; a full description and designation of alectotype of this species was made by Tunnicliff(1982, pp. 60-61, p1. 9, figs 1-7, 9-11)

38 JOHN C. W. COPE
Similodonta Soot-Ryen, 1964, is a character-istic praenuculid genus of the Late Ordovicianof Laurentia and Baltica. It is characterized byan abrupt 90° angle in the middle of the hinge-plate, although Soot-Ryen (1964), in her originaldiagnosis, claimed that the angle was near 80°.The genus is well exemplified by the type speciesS. similis (Ulrich, 1892) from the RichmondGroup of Minnesota. Other Laurentian speciesinclude S. recurva (Ulrich, 1892) and S. costata(Branson, 1909) together with the Ashgillspecies S. magna (Lamont, 1946) and S. collina(Reed, 1946) refigured by Cope (1996a) bothfrom Scotland, and a probable (but unidentified)species figured from Northern Ireland byTunnicliff (1982). An as yet un-named newgenus, clearly related to Similodonta, wasfigured by Pojeta (1978, p1. 2, figs 9-12). LateOrdovician species of Similodonta are alsoknown from Baltica, and include S. spjeldnaesi(Soot-Ryen & Soot-Ryen, 1960) from the UpperCaradoc of the Oslo region; the same specieswas also recorded from the Ashgill of theOslo region by Toni (1975). It could thus beconsidered that Similodonta was a genuscharacteristic of low-latitude areas (presumablywith warm water). The origins of Similodontaare, however, probably Avalonian. Cope (1999)has figured the earliest known species, S. ceryx,from the Aurelucian Stage of the LowerCaradoc of mid-Wales and suggested that it mayhave evolved from the early Llanvirn praenu-culid genus Arcodonta Cope, 1999, putting theorigin of the genus in Avalonia. Similodonta alsooccurs in the Ashgill Series of North Wales(collections of the British Geological Survey).Another possible Gondwanan Similodonta wasfigured by Steinmann & Hoek (1912) from theUpper(?) Ordovician of Bolivia according toBabin (1993), but the text seems to make it clearthat these specimens are distorted lingulidbrachiopods. From the Precordillera ofArgentina Sanchez (1999a) has figured whatappear very clearly to be genera closely relatedto Similodonta. These include Villicumia andTrigonoconcha, the latter with a hinge-plateangle close to 65°; these clearly represent part ofa local Caradoc bivalve diversification that wasdescribed by Sanchez (1999b); both thesenew genera could have been derived fromSimilodonta.
The Llanvirn Tironucula Morris & Fortey,1976, from the Laurentian margin on Svalbard,was possibly the first bivalve to reach Laurentia.Its juvenile dentition is similar to that of manypraenuculids, but it develops blade-like teethontogenetically. Babin (1982a) proposed thefamily Tironuculidae to accommodate this
genus and the clearly related Ekaterodonta,originally described from the Lower Arenig ofthe Montagne Noire (Babin 1982a). This lattergenus is now also known from the LowerLlanvirn of Spain (Babin & Gutierrez-Marco1991) and from the Upper Llanvirn of Bolivia(Babin & Branisa 1987) and, unlike Tironucula,was a high-latitude genus. Another tironuculid,Natasia, has been described from the Lower andMiddle Arenig of NW Argentina by Sanchez(1996) and was subsequently assigned to aseparate subfamily within the Tironuculidae(Sanchez 1997a).
The apparently wide geographical ranges ofsome other nuculoid forms may in some casessimply be due to inappropriate generic assigna-tions. Thus the genus Ctenodonta. that has beenwidely reported from the low-latitude UpperOrdovician rocks, is based on the species C.nasuta (Hall, 1847) from the Upper Ordovicianof the United States. However, many otherspecies have been assigned to this genus in thepast and, as remarked on by Pojeta (1971, p. 15),all that this generic name effectively denotes isthat a nuculoid is being described. There do notappear to be any definitive records of this genusfrom the Early Ordovician although poorlypreserved material from the Tremadoc ofArgentina (Palaeoneilo iruyensis Harrington,1938) was tentatively assigned to Ctenodonta byPojeta & Gilbert-Tomlinson (1977), and Babin& Destombes (1992) figured two forms theycompared to Ctenodonta from the MiddleArenig of Morocco. Definitive records ofCtenodonta are from the Middle Ordovician oflow-latitude Gondwana (Australia; Pojeta &Gilbert-Tomlinson 1977) and the UpperOrdovician of Laurentia (Pojeta 1971) andSiberia (Krasilova 1976, 1979). Other Ordo-vician genera related to Ctenodonta includeTancrediopsis, the earliest known of whichis from the Llanvirn of Mid-Wales (Cope1999); the somewhat younger type species (T.contracta) was originally described fromthe Alumette Islands, Ontario, Canada, bySalter (1859). Alococoncha Pojeta & Gilbert-Tomlinson, 1977 is only known from the MiddleOrdovician of the Amadeus Basin, Australia.Siberoctenia Krasilova, 1976, from the UpperOrdovician of Siberia, has a large number ofsmall teeth.
Pojeta (1988, pp. 210-211) decided that themonotypic superfamily Ctenodontoidea and thefamily Ctenodontidae were poorly founded andof uncertain diagnosis and suggested that, forthe time being, the few genera included inthe family could be incorporated into theNuculanoid family Malletiidae. Nuculanoids are

ORDOVICIAN BIVALVE BIOGEOGRAPHY 39
nuculoids that have a posteriorly elongate shelland the family Malletiidae includes all theOrdovician nuculanoids. Excluding formsrelated to Ctenodonta, the earliest malletiidsappear to be from the Gondwanan MiddleOrdovician and include the genus Cadomia deTromelin, 1877 (de Tromelin & Lebesconte1877), first reported from Armorica, but morerecently reported from the Middle Ordovicianof Argentina (Sanchez 1986). The genusMyoplusia Neumayr, 1884, shows multiplemuscle scars, and other malletiids include theendemic Australian genera Johnmartinia,Lophoconcha, Sthenodonta and Zeehania(Pojeta & Gilbert-Tomlinson 1977). The generaNuculites and Palaeoneilo are very widelyreported from the Upper Ordovician (andyounger rocks) and appear cosmopolitan, but,as commented upon by Pojeta & Gilbert-Tomlinson (1977, p. 16), both genera are in needof revision and probably include several distinctgeneric level taxa.
Solemyoida
The other protobranch group to appear is theSolemyoida, known first from the early Arenigof South Wales (Cope 1996b), although aspointed out by Cope (1997b) the MontagneNoire specimen figured as a Cymatonotal byBabin (1982a, p1. 11, fig. 17) could possiblybelong here too. The genus Ovatoconcha Cope,1996b possesses the strongly anisomyarian ante-riorly elongate shell and radial ornamentationtypical of later solemyoids; it also apparentlyhad a thick periostracal fringe that projectedbeyond the calcified margin of the valves, as inRecent solemyoids (Cope 2000). Pojeta (1988)discussed the origin of the solemyoids andderived them from ctenodontid nuculoids; theearliest form he identified with the solemyoidswas the Baltic species Dystactella aedilis(Eichwald, 1856) from the early Late Ordo-vician of Estonia. Cope (2000) showed how thepresence of ligamental nymphs in Ctenodontaand the Solemyoida, claimed as a synapomorphyto the two groups by Waller (1990, 1998), wasindependently derived, as Mid-Ordovicianctenodontids apparently lack the nymphspossessed by Late Ordovician ctenodontids. Thenymphs of Ctenodonta were probably necessi-tated in the Late Ordovician as the umbonesmigrated rearwards, thus shortening the liga-ment and making stronger insertion essential(Cope 2000). This therefore confirmed Cope'searlier (19976) view that Pojeta's claimedintermediate forms between solemyoids andctenodontids must be persistent intermediate
stocks, as fully evolved solemyoids were presentat least as early as the Early Arenig. No pre-Arenig bivalves are known that could indicatethe origins of Ovatoconcha.
Autolamellibranchia
Cardiolarioidea
Cope (1995) reported that the palaeotaxodontgenus Cardiolaria had a dentition that was sosimilar in some respects to that of the hetero-conch Glyptarca that they were likely to bephylogenetically related. He suggested that theasymmetrical hinge in Cardiolaria could havebeen evolved concomitantly with the evolutionof the filibranch gill and that the hinge wasdesigned to permit far wider valve opening thanthat permitted by the nuculoid hinge. Cope later(19976) proposed the family Cardiolariidae forCardiolaria and related genera, but still retainedthem within the Palaeotaxodonta. Subsequentlyas further cardiolariid taxa have been recog-nized, the Cardiolarioidea was proposed as asuperfamily (Cope 2000) and it was establishedthat it was then possible to classify Ordovicianbivalve groups on the basis of their possessingeither a protobranch or a more advanced gilltype, thus reflecting the principal divisions ofmany zoological classifications of the bivalves. Itwas clear at this point that the subclass Palaeo-taxodonta Korobkov, 1954 was a paraphyleticcombination of protobranch nuculoids andautolamellibranch cardiolarioids, and should beabandoned.
The earliest cardiolarioids known hitherto arefrom the Arenig, but if the ideas of Cope (1995,19976) on the evolution of the filibranch gill arecorrect, they should predate other autolamelli-branchs. Cardiolaria itself is best known fromthe Middle Ordovician, where it is widespread,particularly across European Gondwana, fromPortugal to Bohemia and into Morocco. It isknown earlier, from the Upper Arenig ofArmorica (Babin 1966) and also occurs in theArenig of Guadarranque, Extramadura, Spain(fide Babin 1993). As earlier Ordovician faunasare still known from so few localities, its lack ofearlier occurrence is not seen as a majorproblem. Cardiolaria is also known from theearly Caradoc of the Central Andean Basin ofNW Argentina and Bolivia (Sanchez 19996).
One cardiolarioid has an extraordinarily widelatitudinal range. Cope (2000) proposed thefamily Eritropidae to include the distinctivecardiolarioid Eritropis. Eritropis has a pro-nounced posterior carina and was first describedfrom the Middle Ordovician (Llanvirn Series) of

40 JOHN C. W. COPE
the Amadeus Basin, Australia, by Pojeta &Gilbert-Tomlinson (1977). The same genusoccurs in the Llanvirn Series of mid-Wales(Cope 1999) showing that it was able to migratefor large distances around the Gondwananmargins to Avalonia, apparently quite indepen-dently of water temperature. Eritropis may alsohave reached Bolivia as Arca gracilis Hoek, 1912(in Steinmann & Hoek 1912, pp. 248-249, p1. 8,fig. 17) appears remarkably similar in externalmorphology to Eritropis, but regrettably thedentition is unknown. The only other interpre-tation of this latter fossil is that it is a glyptarcoidbivalve related to either Glyptarca or Hemipri-onodonta. Despite the wide latitudinal toleranceof Eritropis, the other eritropid genusdescribed by Pojeta & Gilbert-Tomlinson(1977), Inaequidens, appears restricted to itstype area.
Trigonioida
The Trigonioida are a distinctive autolamelli-branch group that can be distinguished from theearly Ordovician; they may be separated fromthe heteroconchs by their type of dentition anda short parivincular ligament with strongnymphae and deep ligament grooves (Johnston1996; Johnston & Zhang 1998); this type ofligament is unlike that of the actinodonts, agroup of heteroconchs to which the trigonioidshave often been related (e.g. Pojeta 1978; Babin1993; Waller 1998). Its earliest representativeappears to be Noradonta from the Early Arenigof the Montagne Noire (Babin 1982a); this genuswas first described from the Nora Formation(earliest Mid-Ordovician) of the GeorginaBasin, Australia, by Pojeta & Gilbert-Tomlinson(1977) and this is again an example of a genusthat was able to migrate from the very highlatitudes of the Montagne Noire to the lowlatitude of Australia. The dentition ofNoradonta has affinities with that of the cardio-larioids, suggesting derivation from them (Cope2000). Noradonta is a genus which forms asuitable ancestor for Tromelinodonta (Babin1982b) from the Late Arenig of Armorica. Inturn, Tromelinodonta, by loss of posterior teeth,could produce the typical form of Lyrodesmathat has a radiating subumbonal sheaf of crenu-late teeth. The earliest Lyrodesma recordedhitherto is from the murchisoni Zone of theLlanvirn Series of mid-Wales (Cope 1999).Slightly younger ones are known from Spain(Gutierrez-Marco & Babin 1999) and there aremany species known from the Late Ordo-vician; these include forms from high-latitudeGondwana such as the Armorican L. secure de
Tromelin & Lebesconte, 1876. or the Lyrodesmadescribed from Bolivia by Sanchez & Suarez-Soruco (1996). Undescribed Avalonian formsoccur in both the Caradoc and Ashgill of Walesand NW England (collections of the BritishGeological Survey). Low-latitude Laurentianforms include such species as L. majus (Ulrich,1879) or the Baltican lyrodesmatid figured byNeuman et al. (1997). Unlike contemporarybivalves which favoured silty muds, lyrodes-matids seem characteristic of the unstablehabitats represented by high-energy sands(Cope 1999) and it is noteworthy that this issimilar to the habitat of Mesozoic trigoniids(Stanley 1977). Post-Ordovician lyrodesmatidswere first reported from the Llandovery Seriesof the Laurentian Silurian by Harrison &Harrison (1975), but have subsequently beenfound to occur as late as the Middle Devonian(Pojeta & Zhang 1986). The ventrally flaringcrenulate teeth and dorsal musculature withpedal retractors at the hinge-plate extremitiesprovide good evidence of a phylogenetic linkbetween Lyrodesma and the trigonioids. UpperPalaeozoic trigonioids lack the dental crenula-tion but have the same musculature as both thelyrodesmatids and Mesozoic trigonioids (Newell& Boyd 1975) and the juvenile dentition ofLyrodesma was found to be very similar to theschizodid dentition, suggesting paedomorphicretention of this feature in the Upper Palaeozoicschizodids (Harrison & Harrison 1975). Withinthe Late Ordovician the lyrodesmatids gaverise to the genus Pseudarca de Tromelin &Lebesconte, 1875. For over a century this genuswas of doubtful affinities and was included in theNuculanoidea by Cox et al. (1969), althoughBabin (1966) had suggested it was probably alyrodesmatid. Later, Babin (1987) described therediscovered type material, and, noting itspeculiar dentition with a diverging sheath ofshort teeth, concluded that it was a lyrodes-matid. Tunnicliff (1987) described material fromthe Caradoc rocks of North Wales that includeda species of Pseudarca, previously unknown inBritain, and independently came to the con-clusion that its affinities were lyrodesmatid.Brachilyrodesma Pojeta & Gilbert-Tomlinson,1977 is a Mid- to Late Ordovician lyrodesmatidfrom the Toko Group of the Georgina Basin,Australia, unknown from elsewhere.
Heteroconchia
The Heteroconchia Hertwig, 1895 is a combi-nation of the subclasses PalaeoheterodontaNewell, 1965 and Heterodonta Neumayr, 1884.but as used here excludes the Trigonioida which

ORDOVICIAN BIVALVE BIOGEOGRAPHY 41
were included in the Palaeoheterodonta by Coxetal (1969).
The actinodontid heteroconchs include formswith a sheaf of ventrally radiating teeth. The firstsuch form to be described was Actinodontacuneata Phillips, 1848, from the Middle Llan-dovery (Aeronian) of South Wales. Pojeta(1971) wrongly recorded this species as from theOrdovician and a number of other authors havereported the genus from the Ordovician. Forexample Barrois (1891) figured several bivalvesfrom the Gres Armoricain that he attributed toActinodonta, and Babin (1966) also attributedsome Armorican forms to the genus. Thelectotype of A. cuneata was designated byStubblefield (1938) and a latex cast of this speci-men was figured by Pojeta (1978). The teethbranch out subumbonally and several of theposterior teeth are elongated. It has nowbecome clear that Actinodonta is confined to theSilurian and that Ordovician forms ascribed tothat genus will need new names. Within theLower Ordovician, the genus CarminodontaCope, 1996b approaches Actinodonta the closestin dentition; the former is only known from theLower Arenig of the Llangynog Inlier, SouthWales. Other actinodontids with this type ofdentition include the Mid-Ordovician genusCopidens Pojeta & Gilbert-Tomlinson, 1977from the Georgina Basin of Australia; thisspecies was also recorded from South China byGuo (1988) as a new genus and species,Zadimerodia fastigata, but the two forms areclearly conspecific (Fang & Cope, unpublishedwork). Ananterodonta Babin & Gutierrez-Marco, 1985 from the Lower Llanvirn of Spain,is a further actinodont of this type; this is stillknown only from a single specimen. The earliestCycloconcha was recorded from the MiddleOrdovician of Argentina by Sanchez (1986) butis subsequently known from Laurentia. LateOrdovician species of Cycloconcha show thetypical actinodont subumbonal multiplicity ofteeth; such a form is the C. mediocardinalisMiller, 1874, figured by Pojeta & Runnegar(1985, fig. 13A).
Heteroconchs with reduced dentition aremore widely dispersed. The family RedoniidaeBabin, 1966 comprises heteroconchs with muchreduced subumbonal teeth and lacking anteriordentition. Redonia itself has a wide distribution,being recorded from the Lower Arenig of theMontagne Noire (Babin 1982a) and Morocco(Babin & Destombes 1992). The genus was alsorecorded from the lowest Middle Arenig ofArgentina (Sanchez & Babin 1994; Sanchez1997b). It occurs in the Upper Arenig of theWelsh Borderland (Cope 1999) and Turkey
(Dean & Monod 1970) and in the Llanvirn ofMorocco (Babin & Destombes 1992) andBohemia (Barrande 1881). Other redoniidsinclude the Avalonian genus Moridunia which isabundant in the Lower Arenig of South Wales(Cope 1996b) and probably occurs too in thepoorly preserved Ramsey Island fauna (Hicks1873), whence it was recorded as an Actinodonta(Carter 1971). In Spain another local genus isfound in the Llanvirn, Dulcineia Babin &Gutierrez Marco, 1991, that is characterized bycrenulate teeth. Redoniids appear to be charac-teristic of high to median latitudes as no memberof the family has yet been found in any of the lowlatitude areas.
Another group with very much reduceddentition, frequently related in the past to theheteroconchs, is the family Modiomorphidae.Pojeta (1971, p. 20) remarked upon the vagueconcept of this taxon and the fact that it prob-ably included more than one family level group.Recent work has confirmed this view (Fang &Morris 1997), and it now seems that there aretwo separate bivalve groups involved, neither ofwhich should be assigned to the heteroconchs.These are discussed more fully below.
The genus Babinka Barrande, 1881 is one ofthe most perplexing of Ordovician bivalves asfar as its affinities are concerned. Its multiplemuscle scars suggested that it was closely relatedto monoplacophorans to McAlester (1965, 1966)who decided that it was an early lucinoid.Starobogatov (1971) disagreed with McAlester'sorientation of Babinka and thus its suggestedaffinities, but Pojeta (1978) agreed with theorientation and systematic placing. Otherworkers, however (e.g. Babin 1982a; Cope19976), have preferred to regard it as a hetero-conch. More recently, Taylor & Glover (2000)have described the anterior respiratory pouch inthe lucinoids; this is apparently absent inBabinka and Taylor & Glover (2000, p. 221)concluded that McAlester (1965) had exagger-ated the lucinoid characters of the genus. Thestratigraphical and biogeographical range ofBabinka is also interesting. The earliestexamples are from the Tremadoc of theMontagne Noire (Babin 1982a), but the genus isknown from the Lower Arenig of Morocco(Babin & Destombes 1982), from the UpperArenig of South Wales (Cope 19976), the LowerLlanvirn of the Welsh Borderland (Cope 1999)and Sweden (Soot-Ryen 1969) and the Llanvirnof Bohemia (Barrande 1881). The genus is thuswidely dispersed over the high to mid-latitudesof Gondwana and Avalonia and was one of thefirst bivalves to reach Baltica.
Babinka is far from the only Ordovician

42 JOHN C. W. COPE
heteroconch genus to display a multiplicity ofmuscle scars. Such features are common in othergenera of heteroconchs, including CycloconchaMiller, 1874, Celtoconcha Cope, 1996b andCoxiconcha Babin, 1966. Pojeta & Runnegar(1985) suggested that the multiple accessorymuscle scars in both Babinka and Coxiconchaindicated derivation from Cycloconcha. Thissuggestion seems untenable in view of thefact that the former two genera have a well-established Early Ordovician history, whereasthe latter does not appear until the Mid-Ordovician. Coxiconcha is widely dispersed,being known earliest from the Early Arenig ofthe Montagne Noire (Babin 19820), but in theMid-Ordovician is known in France, Spain andPortugal (Babin 1977), Morocco (Babin &Destombes 1992), and Bohemia (Kriz 1995),reaching Bolivia by the Late Llanvirn (Babin &Branisa 1987).
Another group assigned to the heteroconchsis the Superfamily Glyptarcoidea Cope, 1996b.Cope (1996b) pointed out that these differ fromother heteroconchs in that their teeth radiatedorsally, rather than ventrally as in other hete-roconchs, and in their cladistic analysis ofbivalves Carter et al. (2000) concluded thatGlyptarca was a pteriomorph. The type speciesof Glyptarca, G. primaeva Hicks, 1873,described from the Lower Arenig of RamseyIsland, South Wales, although abundant in thetype locality, is poorly preserved and little of thedentition is known apart from a small sheaf ofanterior teeth and a blade-like posterior tooth;no subumbonal dentition is visible. As a result ofthis view of its dentition it was thought to have asubumbonal lacuna in the dentition. The genuswas thus viewed as an ideal ancestor for thecyrtodontids, which have curved dorsally flaringanterior teeth, a subumbonal dental lacuna andone or more straight posterior teeth (e.g. Morris1978). Cope (19966) figured a new species, G.serrata, from the Llangynog Inlier, some 60 kmeast of Ramsey Island, but at the same horizon,which showed perfectly preserved dentition,demonstrating that the genus had in reality acomplex subumbonal overlap of the anteriorand posterior teeth and thus there was no directlink to the cyrtodontids. Cope (1995, 19976)showed that this dentition bears strong simi-larities to that of cardiolarioids and these mightwell have been ancestral to the glyptarcoids. Theglyptarcoids include the genera Glyptarca andCeltoconcha, both known from the LowerArenig of Wales (Cope 19966), the former alsooccurring in the Llanvirn of Mid-Wales (Cope1999) where it occurs together with anotherglyptarcoid, Camnantia Cope, 1999. The only
other glyptarcoid known hitherto was widelyreported as Glyptarca, but following Cope's(19966) redefinition of the genus, is now knownto belong to a separate genus, HemiprionodontaCope, 19966 (type species ?Dolabra lusitanicaSharpe, 1853). This genus is known widely fromthe Middle Ordovician of Spain (Babin &Gutierrez-Marco 1991) and Armorica (Babin1966) and has also been reported from theMiddle Ordovician of Saudi Arabia (Fortey &Morris 1982). Hemiprionodonta is known fromboth the Late Llanvirn (Llandeilian) and theCaradoc of Morocco (Babin & Destombes1992).
PteriomorphiaGlyptarca has dentition closely related to that ofthe genus Catamarcaia Sanchez & Babin, 1993from the upper part of the Middle Arenig ofArgentina; this latter genus is the earliest formthat has a duplivincular ligament. Sanchez &Babin (1993) believed that it was an earlypteriomorph lacking the central edentulous areaon the hinge-plate and Cope (1997a) decidedthat it was the earliest arcoid sensu stricto that hethen included in the Neotaxodonta. Morerecently Cope (2000) accepted that the Neo-taxodonta could be included within the Pterio-morpha, as they shared many features includingthe multiple ligamental insertions, but heretained the Order Arcoida in a restricted senseand recognized the Cyrtodontoida as a separateorder. As Catamarcaia had a unique mix ofcharacters, the monotypic family Catamar-caidae was proposed for it (Cope 2000). Thedeficient early fossil record of the OrderArcoida (sensu Cope 1997a) needs examination;after Catamarcaia in the Middle Arenig, noarcoids are known from the remainder of theOrdovician. The next record is that ofAlytodonta Cope, 19976 from the LowerLlandovery of Girvan, Scotland, on the Lau-rentian margin, and then the Wenlock generaTrecanolia and Uskardita (Ratter & Cope 1998)to add to the previously known Freja Liljedahl.1984. Judging from the occurrence of these latergenera, it appears possible that arcoids may haveinhabited some of the nearest inshore environ-ments and this may explain their absence thusfar from the Mid- and Late Ordovician records,which must be put down to collection failure.
In their cladistic review of early bivalve phy-logeny Carter et al. (2000) preferred to derivethe glyptarcoids from Catamarcaia and assignedthem to the Pteriomorphia. Clearly Glyptarcalies close to the point of divergence of theheteroconchs and pteriomorphians and thus

ORDOVICIAN BIVALVE BIOGEOGRAPHY 43
they show affinities to both groups and it couldbe that their placement is correct within thelatter clade; there are many early arcoids andcyrtodonts that apparently do not have multipleligamental insertions, including the earliestcyrtodontids known from the Tremadoc ofAustralia (Cyrtodonta and Pharcidoconcha;Pojeta & Gilbert-Tomlinson 1977), and from theLower Arenig of Wales (Cyrtodonta andCyrtodontula; Cope 1996b), together with thestrongly ribbed genus Falcatodonta (Cope1996b).
Cyrtodonta itself ranges from the TremadocSeries of Australia through to the Silurian. In theEarly Ordovician, cyrtodontids seem to havebeen little constrained by latitude; they appearfirst in the Tremadoc of Australia (Pojeta &Gilbert-Tomlinson 1977) and they also occur inthe Lower Arenig of Avalonia (Cope 1996b).They have been recorded with a query from thehigher latitudes of the Mid-Ordovician of bothSpain (Babin & Gutierrez-Marco 1991) andMorocco (Babin & Destombes 1992). In the lasttwo cases no dentition is known and the genericassignment was made on the basis of shell shape.Cyrtodontula was also recorded from the MiddleOrdovician of Argentina by Sanchez (1990).However, it is on the Late Ordovician low-latitude carbonate platforms that cyrtodontsbecame most diverse (Cope & Babin 1999) andby then clearly showing a preference for lowlatitudes. In Laurentia Cyrtodonta occurstogether with such genera as Cyrtodontula,Ortonella and Vanuxemia. The earliest Laurent-ian occurrence of cyrtodontids appears to be theshallow water St Peter's Sandstone fauna ofMinnesota first described by Sardeson (1896)from the late Chazyan ( = Early Caradoc). Thisfauna includes two species each of Cyrtodontaand Vanuxemia (Sardeson 1939a, b). The lattergenus is also found on the Late Ordoviciancarbonate platform of Baltica, whence Isberg(1934) also described the genera Thorslundiaand Warburgia. Soot-Ryen & Soot-Ryen (1960)also record Cyrtodontula from the Oslo area.Cyrtodonta and Vanuxemia were recordedfrom Kazakhstania by Khalfin (1958) whilstCyrtodonta, Cyrtodontula, Plethocardia,Ortonella and Vanuxemia were recorded fromSiberia by Krasilova (1970, 1979)
Accompanying the cyrtodontids are a varietyof genera of ambonychiids; these are pterioidbivalves that have their umbones at the anteriorend of the shell and commonly have a byssalsinus. They are strongly anisomyarian or mono-myarian in the adult stage and the posterioradductor is often displaced towards the centre ofthe valves. Such clearly epifaunal forms are,
unsurprisingly, common in the low-latitudecarbonate platforms. However, the earliestambonychiid recorded hitherto is from Ava-lonia, from the Middle Arenig of South Wales(Cope 19966), where it occurs in a sandstone;this is an undetermined species of Cleionychia,which is also the earliest ambonychiid genus tooccur in the Upper Ordovician of NorthAmerica (Pojeta 1966). Middle Ordovicianambonychiids are known from Australia whencePojeta & Gilbert-Tomlinson (1977) describedspecies of Glyptonychia, Leconychia andPteronychia. Pojeta (1966) illustrated somethingof the large variety of North American ambony-chiids and figured Late Ordovician species ofAllonychia, Ambonychiopsis, Anomalodonta,Ambonychia, Cleionychia, Eridonychia, Mary-onychia, Opisthoptera and Psilonychia; to thesemay be added Claudeonychia Pojeta, 1997.Some of these genera are also known from theBaltic Upper Ordovician carbonate platforms(Isberg 1934) where the fauna also includesgenera such as Anomalocoelia, Paramytilarcaand Praeanomalodonta. The ambonychiidfaunas from around the continents of Kaza-khstania and Siberia are not as varied as those ofLaurentia and Baltica but include species ofAmbonychia and Cleionychia (Khalfin 1958;Krasilova 1979).
Pterineids are also particularly characteristicof the Upper Ordovician, but the earliest is aPalaeopteria from the Lower Arenig of SouthWales (Cope 19966) only known from a singleright valve. The next oldest genus is Gond-wanan, in this case from the Amadeus Basin ofAustralia, the genus Denticelox Pojeta &Gilbert-Tomlinson, 1977, which, unlike laterpterineids is biconvex. Carotidens Foerste, 1910and Palaeopteria occur in the Upper Ordovicianof the United States and Canada; both thesegenera have asymmetrical valves, with a flatterright than left valve.
The last group of pteriomorphian bivalvesto evolve in the Ordovician was the limids.Tunnicliff (1987) described the genus Myo-dakryotus from the Caradoc of North Wales.This genus has characteristics in common withboth cyrtodontids and later limids, suggestingthat the group was derived from the cyrtodon-tids. Pojeta & Runnegar (1985, fig. 17) figured alimid species from the Late Ordovician ofCanada and the United States, which theyassigned with a query to the genus ProlobellaUlrich, 1894, regarded by Cox et al. (1969) as apalaeoheterodont. Ordovician limids lack theinternal ligament of more recent forms (Pojeta& Runnegar 1985).
A further group of the pteriomorphians is the

44 JOHN C. W. COPE
superfamily Modiolopsoidea Fischer, 1887, for-merly often classified with the heteroconchs, butshown by Carter & Seed (1998) to include sometaxa that display multiply inserted non-parivin-cular ligaments; they may thus be related to thepteriomorphs. In the Treatise (Cox el al. 1969)the modiolopsoids are included in the orderModiomorphoida and the superfamily Modio-morphoidea. Fang & Morris (1997) revised thetaxonomy of this problematic superfamily andshowed that it included two distinct groups ofbivalves: the modiomorphoids, typified bythe Devonian genus Modiomorpha, had well-developed ligamental nymphs that allowedthem to be distinguished from the Ordovicianmodiolopsoids, a group with which they hadfrequently been confused, but which lacked theligamental nymphs. Fang & Morris (1997) alsoshowed that the family Permophoridae Van dePoel, 1959 was a subjective junior synonym ofthe family Modiomorphidae Miller, 1877 andrecommended that order Modiomorphoida beabandoned and that the superfamily Modiomor-phoidea Miller, 1877, as used in the Treatise(Cox et al. 1969), should be replaced by theModiolopsoidea Fischer, 1887, in which theyrecognized the families Modiolopsidae Fischer,1887, Colpomyidae Pojeta & Gilbert-Tomlinson,1977 and the Modiolodontidae Fang & Morris,1997. Carter & Seed (1998) showed that somemodiolopsoids had multiply inserted non-parivincular ligaments and suggested that theiraffinities thus lay with the pteriomorphs (seeabove). The modiomorphoids, on the otherhand, possessed a periostracum with calcifiedspicules and Carter & Aller (1975) suggestedthat this feature, widely found in the anom-alodesmatans, indicated that their affinities laywith that group.
Modiolopsids are recorded as far back asthe Early Tremadoc of Argentina, whenceHarrington (1938) figured a form he describedas Cosmogoniophorina tenuicostata sp. nov.Harrington figured three syntypes, two of which(Harrington 1938, p1. 3, figs 1, 5) show an almostcarinate shell and one (Harrington 1938, fig. 1)also shows clear radial ornamentation on thepost-umbilical shoulder. There appears to belittle quarrel that these are modiolopsids, butI find myself in agreement with Pojeta(1971, p. 17) that Harrington's other syntype(Harrington 1938, p1. 3, fig. 4) appears to showtaxodont teeth, and a less marked post-umbonalshoulder and apparently lacks radial ornamenta-tion, suggesting this specimen is a nuculoid. Toclarify matters I designate the specimen figuredby Harrington (1938, p1. 4, fig. 1) as lectotype ofthe species Goniophorina (Cosmogoniophorina)
tenuicostata (Harrington), 1938. The samespecies has been recently recorded from the earlyMiddle Arenig of Argentina by Sanchez (1997b)who noted that as Harrington did not describethe dentition, the generic and subgenericassignments are uncertain.
The next oldest species are two species ofModiolopsis from the Early Arenig of SouthWales (Cope 19966) where the modiolopsidsCosmogoniophorina and Parallelodus alsooccur. Middle Ordovician modiolopsids arerecorded possibly from Spain (Babin &Gutierrez-Marco 1991, pp. 123-124), from theMiddle and Upper Ordovician of Argentina(Sanchez 1990, 1999b) and the Upper Ordo-vician of Laurentia (Pojeta 1971), Kazakhstania(Khalfin 1958) and Siberia (Krasilova 1970.1979). The genus Corallidomns appears in theUpper Ordovician; this genus is noteworthybecause it is the first boring bivalve (as firstreported by Whitfield 1893) and lived in cryptsexcavated into colonial corals or bryozoans;the same mode of life was followed bySemicorallidomus Isberg, 1934.
Colpomyids normally have rather amorphousbulbous-shaped teeth, referred to as articulatingdevices by Pojeta and Gilbert-Tomlinson (1977p. 27), that are not mounted on a hinge-plate.The earliest forms are from the Late Tremadocof the Amadeus Basin of Australia, Colpantvxand Xestoconcha (Pojeta & Gilbert-Tomlinson1977); the latter genus also occurs in the EarlyArenig of South Wales (Cope 1996b).Colpomyids are known from the UpperOrdovician of Laurentia (Pojeta 1978). Siberia(Krasilova 1979) and western Gondwana(Sanchez 1999b) and survived into the Silurian.
The Modiolodontidae are modiolopsoids thathave a small number of cardinal teeth in eachvalve. They are known from the Middle andUpper Ordovician and younger rocks. One ofthe earliest is the Modioliodon recorded fromthe Llanvirn of Avalonia by Cope (1999).
Anomalodesmata
The earliest generally accepted anomalodes-matan is Arenigomya from the Early Arenig ofSouth Wales (Cope 19966). Arenigomya showsalready several of the principal characters of thegroup, including a finely pustulate shell orna-ment and an edentulous hinge that sometimescarries subumbonal articulating devices (seeCope 19966, p1. 7, figs 12-14; text-fig. 7). Thesecharacters are evident in the genera Cuneamyaand Rhytimya (the latter also possesses a pos-terior gape), both of which are known from theLate Ordovician of the United States, Canada

ORDOVICIAN BIVALVE BIOGEOGRAPHY 45
and Siberia; Cuneamya is also recorded fromKazakhstania (Khalfin 1958). Cuneamyaappears earliest, however, in the MiddleOrdovician of Argentina (Sanchez 1990).
Another Ordovician group may also beanomalodesmatans: this is the elongate orthono-tians which were elevated to subclass level byPojeta (1978). Of the Ordovician genera Pojeta(1978) included in the Orthonotia, Psiloconchawas later transferred to the Solemyoida (Pojeta1988) and the elongate Cymatonota, Soleno-morpha and Prothyris would now be regardedby most workers as anomalodesmatans. Brev-iorthonota Krasilova, 1979, from the UpperOrdovician of Siberia, also belongs here. Theearliest possible Cymatonota was recorded fromthe Early Arenig of the Montagne Noire byBabin (1982a) but as discussed above, there is apossibility that this specimen may be a solemyid.The orthonotids were included in the Anom-alodesmata by Cox et al. (1969).
Waller (1990) suggested that the ligamentalnymphs and outer prismatic shell layer of themodiomorphoids suggested that they werelinked to the anomalodesmatans and this sup-ported Carter & Aller's (1975) suggestion thatthe calcified spicules of the modiomorphoidperiostracum indicated that their affinities laywith the anomalodesmatans. Fang & Morris(1997) showed that the modiomorphoideans asdefined by Cox et al. (1969) included both theDevonian modiomorphoids and the Ordo-vician-Silurian modiolopsoids, a group withwhich they had frequently been confused, butwhich lacked the ligamental nymphs. The latterare now regarded as pteriomorphians and arediscussed above.
Summary and conclusionsAlthough fairly widely dispersed during theEarly and Mid-Cambrian, it is suspected thatduring the Late Cambrian bivalves may havebecome restricted to Gondwana, but they are sofar unknown with certainty from that Epoch.
In the Tremadoc Epoch of the OrdovicianPeriod bivalves are known from Gondwananhigh latitudes (Morocco, Montagne Noire),median latitudes (Argentina) and low latitudes(Australia). The most diverse fauna so fardiscovered is that of the Amadeus Basin inAustralia (Pojeta & Gilbert-Tomlinson 1977)which already contains pteriomorphians that arenot known with certainty from higher latitudesin rocks of that age. The Early Ordovician initialdiversification of the bivalves was restricted toGondwana (including Avalonia at this time) andby the Mid-Arenig all the principal clades of
bivalves had evolved. Some forms weredecidedly high-latitude in distribution, whilstothers were just as clearly confined to lowlatitudes; most areas had some endemic genera.Comparison between the high latitude of theMontagne Noire and the lower latitude ofAvalonia for the Early Arenig (Fig. 3a) shows afar greater diversity in Avalonian latitudes.Avalonia has at least 18 genera belonging to allthe major bivalve groups, with great diversity ofpteriomorphians and to a lesser extent hetero-conchs, whereas nuculoids and heteroconchs areequally important in the less diverse faunas ofhigher latitudes. Even though Avalonian wasnot far north of the Gondwanan margin in EarlyArenig times, the difference in pteriomorphiandiversity is pronounced. It is a pity that there areno known low-latitude faunas of this age withwhich to make further comparisons. It thusseems that climatic gradients played some partin controlling bivalve diversity in the EarlyOrdovician, but the fact that a few genera occurat both high and median latitudes suggests thatsuch gradients may not have been particularlystrong at this time. In some cases the gradualmigration of a species across Gondwana can betracked through time, whilst in other cases thesame genus appears at widely separated areas,or latitudes, simultaneously.
Figure 4a shows abundance of bivalve speciesbelonging to each major clade for the faunas ofSouth Wales and the Montagne Noire; compari-son of Figure 3a with Figure 4a shows somesurprising differences. Most notable for Wales isthe fact that the diverse pteriomorphians makeup only 9% of individuals, with the fauna totallydominated by heteroconchs (76.6%). For theMontagne Noire the fauna is again dominatedby heteroconchs (56.6%) whilst the pterio-morphians make up only 0.4% of the bivalvepopulation there.
Early Ordovician bivalves seem particularlycontrolled by facies, occurring virtually exclus-ively in siliciclastic sediments and being mostabundant in very shallow-water silty muds(Cope & Babin 1999). Even the deeper-waterfacies, such as the muds of the Montagne Noire,were certainly deposited in water less than 50 mdeep (Babin 2000). These Early Ordovicianbivalves were predominantly infaunal, althoughthere may have been some semi-infaunal forms.
In the Mid-Ordovician bivalves managed tomigrate from Gondwana and reached bothBaltica and the Laurentian margin in early Mid-Ordovician times. Once established here theyspread to reach all continental shelves by theearliest part of the Late Ordovician. Mid-Ordovician faunas are thus, again, virtually
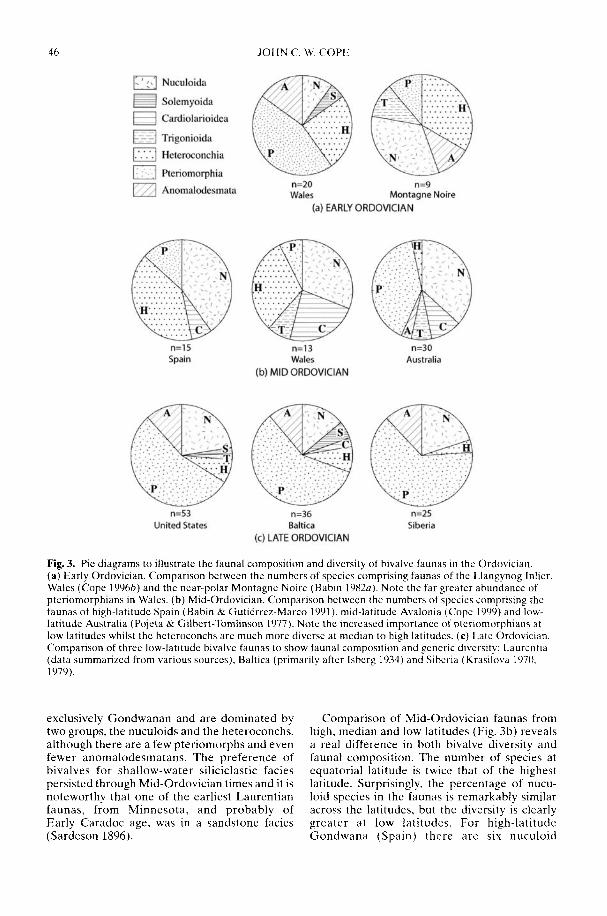
46 JOHN C. W. COPE
Fig. 3. Pie diagrams to illustrate the faunal composition and diversity of bivalve faunas in the Ordovician.(a) Early Ordovician. Comparison between the numbers of species comprising faunas of the Llangynog Inlier.Wales (Cope 1996b) and the near-polar Montagne Noire (Babin 1982a). Note the far greater abundance ofpteriomorphians in Wales, (b) Mid-Ordovician. Comparison between the numbers of species comprising thefaunas of high-latitude Spain (Babin & Gutierrez-Marco 1991), mid-latitude Avalonia (Cope 1999) and low-latitude Australia (Pojeta & Gilbert-Tomlinson 1977). Note the increased importance of pteriomorphians atlow latitudes whilst the heteroconchs are much more diverse at median to high latitudes. (c) Late Ordovician.Comparison of three low-latitude bivalve faunas to show faunal composition and generic diversity: Laurentia(data summarized from various sources), Baltica (primarily after Isberg 1934) and Siberia (Krasilova 1970.1979).
exclusively Gondwanan and are dominated bytwo groups, the nuculoids and the heteroconchs,although there are a few pteriomorphs and evenfewer anomalodesmatans. The preference ofbivalves for shallow-water siliciclastic faciespersisted through Mid-Ordovician times and it isnoteworthy that one of the earliest Laurentianfaunas, from Minnesota, and probably ofEarly Caradoc age, was in a sandstone facies(Sardeson 1896).
Comparison of Mid-Ordovician faunas fromhigh, median and low latitudes (Fig. 3b) revealsa real difference in both bivalve diversity andfaunal composition. The number of species atequatorial latitude is twice that of the highestlatitude. Surprisingly, the percentage of nucu-loid species in the faunas is remarkably similaracross the latitudes, but the diversity is clearlygreater at low latitudes. For high-latitudeGondwana (Spain) there are six nuculoid
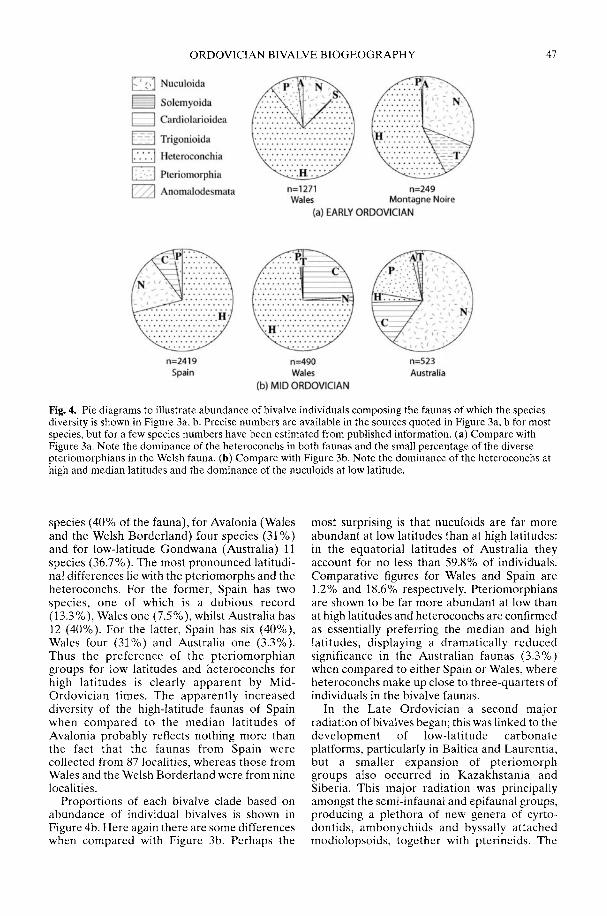
ORDOVICIAN BIVALVE BIOGEOGRAPHY 47
Fig. 4. Pie diagrams to illustrate abundance of bivalve individuals composing the faunas of which the speciesdiversity is shown in Figure 3a, b. Precise numbers are available in the sources quoted in Figure 3a, b for mostspecies, but for a few species numbers have been estimated from published information. (a) Compare withFigure 3a. Note the dominance of the heteroconchs in both faunas and the small percentage of the diversepteriomorphians in the Welsh fauna. (b) Compare with Figure 3b. Note the dominance of the heteroconchs athigh and median latitudes and the dominance of the nuculoids at low latitude.
species (40% of the fauna), for Avalonia (Walesand the Welsh Borderland) four species (31%)and for low-latitude Gondwana (Australia) 11species (36.7%). The most pronounced latitudi-nal differences lie with the pteriomorphs and theheteroconchs. For the former, Spain has twospecies, one of which is a dubious record(13.3%), Wales one (7.5%), whilst Australia has12 (40%). For the latter, Spain has six (40%),Wales four (31%) and Australia one (3.3%).Thus the preference of the pteriomorphiangroups for low latitudes and heteroconchs forhigh latitudes is clearly apparent by Mid-Ordovician times. The apparently increaseddiversity of the high-latitude faunas of Spainwhen compared to the median latitudes ofAvalonia probably reflects nothing more thanthe fact that the faunas from Spain werecollected from 87 localities, whereas those fromWales and the Welsh Borderland were from ninelocalities.
Proportions of each bivalve clade based onabundance of individual bivalves is shown inFigure 4b. Here again there are some differenceswhen compared with Figure 3b. Perhaps the
most surprising is that nuculoids are far moreabundant at low latitudes than at high latitudes:in the equatorial latitudes of Australia theyaccount for no less than 59.8% of individuals.Comparative figures for Wales and Spain are1.2% and 18.6% respectively. Pteriomorphiansare shown to be far more abundant at low thanat high latitudes and heteroconchs are confirmedas essentially preferring the median and highlatitudes, displaying a dramatically reducedsignificance in the Australian faunas (3.3%)when compared to either Spain or Wales, whereheteroconchs make up close to three-quarters ofindividuals in the bivalve faunas.
In the Late Ordovician a second majorradiation of bivalves began; this was linked to thedevelopment of low-latitude carbonateplatforms, particularly in Baltica and Laurentia,but a smaller expansion of pteriomorphgroups also occurred in Kazakhstania andSiberia. This major radiation was principallyamongst the semi-infaunal and epifaunal groups,producing a plethora of new genera of cyrto-dontids, ambonychiids and byssally attachedmodiolopsoids, together with pterineids. The

48 JOHN C. W. COPE
anomalodesmatans, which are primarily (butnot exclusively) infaunal, were also more diverseat low latitudes, as too were the protobranchgroups. The rapid diversification of bivalves inthe Late Ordovician produced a large number ofspecies as well as genera, but there has been littlerevisionary work on the faunas. For this reason,Figure 3c shows the number of genera in theLate Ordovician in three low-latitude areas.Insufficient is known of higher-latitude faunas tomake meaningful comparisons.
The low-latitude shelves contained a highproportion of Late Ordovician endemic genera.Cope & Babin (1999) noted that out of 53genera from the Laurentian Late Ordovician,over 40% were endemic. However, some low-latitude continental shelves were evidently insufficient proximity to each other to allow inter-change of genera. For example the stronglyribbed Paraphtonia Khalfin, 1958 from Kaza-khstan also occurs in Siberia (Krasilova 1979)and genera such as Cyrtodonta are close tocosmopolitan. In stark contrast to these richlow-latitude faunas, the Late Ordovician faunasof high latitudes are quite impoverished andentirely dominated primarily by nuculoids;there are a few heteroconchs, but very fewpteriomorphs or anomalodesmatans. Althoughthere has been a lack of recent work on high-latitude Late Ordovician faunas, it appears thatrates of endemism are lower, with Gondwana atabout 30% and Avalonia at about 14% (Cope &Babin 1999).
These factors strongly suggest that latitudinaltemperature differences became more pro-nounced in the Late Ordovician, heralding theLate Ashgill glaciation. This glaciation pro-duced a major eustatic sea-level fall thatexposed the low-latitude carbonate platforms.The resulting extinction event had a profoundeffect on bivalve stocks. The greatest losses wereamongst the epifaunal and semi-infaunalgroups, with major extinctions amongst thecyrtodontids, ambonychiids and pterineids.Other groups to be affected were the modi-olopsids, amongst which the extinction ofthe coral-boring genera Corallidomus andSemicorallidomus ended a mode of life that wasnot to re-evolve until the Late Jurassic. On theother hand, at higher latitudes, the nuculoid-dominated communities appeared to havesuffered least by the sea level fall, the infaunalforms adapting to the new sea-levels with littleapparent difficulty; several genera survivedapparently little changed into the Silurian.
There has been very little recent work pub-lished on Silurian bivalve faunas; this is a pityas the bivalves have the potential to provide
much information about the recovery from thelatest-Ordovician mass extinction. One faunathat has been studied is that of the Silurian ofWales and the Welsh Borderland (Ratter 1999).which is largely unpublished. Ratter's studiesshow that in the Llandovery Series these mid-latitude Avalonian faunas, predominantly fromsiliciclastic facies, are dominated by nuculoids,which make up half of the total of 18 recordedspecies. The remainder of the bivalve faunaconsists, in order of diminishing importance, ofanomalodesmatans, heteroconchs and trigo-nioids; there are no pteriomorphians. Diversitylevels appear much lower in the Early Silurianand it seems that the Ashgill extinctions essen-tially reduced the bivalves to something liketheir Mid-Ordovician levels of diversity.
Thanks are due to L. C. Norton and L. E. Popov. bothof the Geology Department, National Museum ofWales, for assistance in drafting of figures 1. 3 and 4.and figure 2, respectively.
ReferencesBABIN. C. 1966. Mollusques bivalves et cephalopodes
du Paleozoique armoricain. Imprimerie Commer-ciale et Administrative. Brest.
BABIN. C. 1977. Etude comparee des genres BabinkaBarrande et Coxiconcha Babin (Mollusquesbivalves de l'Ordovicien) Interet phylogenetique.Geobios. 10. 51-79.
BABIN. C. 1982a. Mollusqes bivalves et rostroconches.In: BABIN. C.. COURTESSOLE. R.. MELOU. M..PILLET, J.. VIZCAINO, D. & YOCHELSON. E. L. (eds)Brachiopodes (articules) et mollusques (bivalves,rostroconches, monoplacophores, gastropodes) deI'Ordovicien inferieur (Tremadocien-Arenigien)de la Montagne Noire (France meridionale).Memoire de la Societe des Etudes Scientifiquesde l'Aude. Sival. Carcassonne. 37-49. pls 8-12.
BABIN. C. 1982b. Tromelinodonta nov. gen.. Bivalvelyrodesmatide (Mollusca) de l'Arenigien(Ordovicien inferieur). Geobios. 15. 423—427.
BABIN. C. 1987. Pseudarca. Mollusque Bivalve sin-gulier de l'Ordovicien superieur du Massifarmoricain. Annales de la Societe Geologique duNord. 105, 253-258.
BABIN. C. 1993. Role des plates-formes gondwani-ennes dans la diversification des mollusquesbivalves durant l'Ordovicien. Bulletin de laSociete geologique de France. 164. 141-153.
BABIN. C. 2000. Ordovician to Devonian diversifica-tion of the Bivalvia. American MalacologicalBulletin. 15. 167-178.
BABIN. C. & B R A N I S A . L. 1987. Ribeiria. Peelerophon yotros moluscos del Ordovicico de Bolivia. IVCongreso Latinamerico de Paleontologia, SantaCruz. 1. 119-129.
BABIN. C. & DESTOMBES. J. 1992 (dated 1990).Les Mollusques Bivalves et Rostroconchesordoviciens de l 'Anti-Atlas marocain: Interet

ORDOVICIAN BIVALVE BIOGEOGRAPHY 49
paleogeographique de leur inventaire. GeologieMediterraneenne, 17, 243-261.
BABIN, C. & GUTTIERREZ-MARCO, J. C. 1985. Unnouveau cycloconchide (Mollusca, Bivalvia) duLlanvirn inferieur (Ordovicien) des Monts deTolede (Espagne). Geobios, 18, 609-614.
BABIN, C. & GUTTIERREZ-MARCO, J. C. 1991. MiddleOrdovician bivalves from Spain and their phyleticand palaeogeographic significance. Palaeontol-ogy,34, 109-147
BARRANDE, J. 1881. Systeme silurien du centre de laBoheme. lere partie. Recherches paleon-tologiques. Vol VI: Classe des Mollusques. Ordredes Acephales. Bellman, Prague and Paris.
BARROIS, C. 1891. Memoire sur la faune du gres armor-icain. Annales de la Societe Geologique du Nord,19,134-351, pis 1-5.
BERG-MADSEN, V. 1987. Tuarangia from Bornholm(Denmark) and similarities in Baltoscandian andAustralasian late Middle Cambrian faunas.Alcheringa, 11, 245-259, pl. 7.
BRANSON, E. E. 1909. The fauna of the residuaryAuburn Chert of Lincoln County, Missouri.Transactions of the Academy of Science of StLouis, 18, 39-52.
CARTER, J. G. & ALLER, R. C. 1975. Calcification ofthe bivalve periostracum. Lethaia, 8, 315-320.
CARTER, J. G. & SEED, R. 1998. Thermal potentiationand mineralogical evolution in Mytilus (Mollusca;Bivalvia). In: JOHNSTON, P. A. & HAGGART, J. W.(eds) Bivalves: An Eon of Evolution — Paleobio-logical Studies Honoring Norman D. Newell. Uni-versity of Calgary Press, Calgary, 87-117.
CARTER, J. G., CAMPBELL, D. C. & CAMPBELL, M. R.2000. Cladistic perspectives on early bivalve evol-ution. In: HARPER, E. M., TAYLOR, J. D. &CRAME, J. A. (eds) The Evolutionary Biology ofthe Bivalvia. Geological Society, London, SpecialPublications, 177, 47-79.
CARTER, R. M. 1971. Revision of Arenig Bivalvia fromRamsey Island, Pembrokeshire. Palaeontology,14,250-261.
COPE, J. C. W. 1995 (Dated 1996). The early evolutionof the Bivalvia. In: TAYLOR, J. D. (ed.) Origin andEvolutionary Radiation of the Mollusca. OxfordUniversity Press, Oxford, 361-370.
COPE, J. C. W. 1996a. 8. Bivalves. In: HARPER, D. A. T.& OWEN, A. W. (eds) Fossils of the UpperOrdovician. Palaeontological Association, FieldGuides to Fossils, 7, 95-115.
COPE, J. C. W. 1996b. Early Ordovician (Arenig)bivalves from the Llangynog Inlier, South Wales.Palaeontology, 39, 979-1025.
COPE, J. C. W. 1997a. Affinities of the early Ordo-vician bivalve Catamarcaia Sanchez andBabin,1993 and its role in bivalve evolution.Geobios Memoire Special, 20,126-131.
COPE, J. C. W. 1997b. The early phylogeny of the classBivalvia. Palaeontology, 40, 713-746.
COPE, J. C. W. 1999. Middle Ordovician bivalves frommid-Wales and the Welsh Borderland. Palaeon-tology, 42, 467-499 .
COPE, J. C. W. 2000. A new look at early bivalve phy-logeny. In: HARPER, E. M, TAYLOR, J. D. &CRAME, J. A. (eds) The Evolutionary Biology of
the Bivalvia. Geological Society, London, SpecialPublications, 177, 81-95.
COPE, J. C. W. & BABIN, C. 1999. Diversification ofbivalves in the Ordovician. Geobios, 32,175-185.
Cox, L. R., NEWELL, N. D., BOYD, D. W., et al,1969-1971. In: MOORE, R. C. (ed.) Treatise onInvertebrate Paleontology. Part N. Mollusca 6,Bivalvia, vols 1-3. Geological Society of Americaand University of Kansas Press, Boulder, Col-orado and Lawrence, Kansas.
DEAN, W. T. & MONOD, O. 1970. The Lower Palaeo-zoic stratigraphy and faunas of the Taurus Moun-tains near Beysehir, Turkey. I. Stratigraphy.Bulletin of the British Museum (Natural History),Geology Series, 19,413-426.
DESPARMET;R.,TERMIER,G. &TERMIER, H. 1971. Surun bivalve protobranche ante-arenigien trouve aunord de Wardak (Afghanistan). Geobios, 4,143-150.
DZIK, J. 1994. Evolution of 'small shelly fossils' assem-blages. Ada Palaeontologica Polonica, 39,247-313.
EICHWALD, E. 1856. Beitrag zur geographischen Ver-breitung der fossilen Thiere Russlands AltePeriod. Bulletin de la Societe des Naturalistes deMoscou, 17, 247-339.
FANG, ZONG-JIE & MORRIS, N. J. 1997. The genusPseudosanguinolites and some rnodioliformbivalves (mainly Palaeozoic). Palaeoworld, 7,50-74.
FISCHER, P. H. 1880-1887. Manual de conchyliologie etde paleontologie conchy liologique. F. Savy, Paris.
FOERSTE, A. F. 1910. Preliminary notes on Cincinnat-ian and Lexington fossils of Ohio, Indiana, Ken-tucky and Tennessee. Bulletin of the ScienceLaboratories of Denison University, 16,17—87.
FORTEY, R. A. & MORRIS, S. F. 1982. The Ordoviciantrilobite Neseuretus from Saudia Arabia and thepalaeogeography of the Neseuretus fauna relatedto Gondwanaland in the earlier Ordovician.Bulletin of the British Museum (Natural History),Geology Series, 36, 63-75.
GEYER, G. & STRENG, M. 1998. Middle Cambrian pele-cypods from the Anti-Atlas, Morocco. RevistaEspanola di Paleontologia, no. extra Homenajaeal Prof. Gonzalo Vidal, 83-96.
GROBBEN, K. 1894. Zur Kenntniss der Morphologie,Der Verwandschaftsverhaltnisse und des Systemsder Mollusken. Sitzungsberichte der KaiserlichenAkademie der Wissenschaften, Mathematisch-Naturwissenschaftlichen Classe, 103, 61-86.
Guo, FUXIANG. 1988. New genera of fossil bivalvesfrom Yunnan. Yunnan Geology, 1, 112-144 (inChinese with English abstract).
GUTTIERREZ-MARCO, J. C. & BABIN, C. 1999.Lyrodesma et autres mollusques bivalves desQuartzites Botella (Ordovicien Moyen) de laZone Centre-iberique (Espagne). RevistaEspanola di Paleontologia, no. extra Homenajaeal Prof. J. Truyols, 229-238.
HALL, J. 1847. Palaeontology. Geological Survey ofNew York, 1, 1-338.
HARRINGTON, H. J. 1938. Sobre las faunas del Ordo-viciano inferior del Norte Argentine. Revistadel Museo de La Plata, Nueva Serie, 1, 109-289,14 pls.

50 JOHN C. W. COPE
HARRISON, W. B. & HARRISON, L. K. 1975. AMaquoketa-like molluscan community in theBrassfield Formation (Early Silurian) of AdamsCounty, Ohio. Bulletins of American Paleontol-ogy, 67, 197-231.
HAVLICEK, V. & KRIZ, J. 1978. Middle CambrianLamellodonta simplex Vogel: 'bivalve' turnedbrachiopod Trematobolus simplex (Vogel).Journal of Paleontology, 52, 972-975.
HERTWIG, C. W. T. R. 1895. Lehrbuch der Zoologie.Gustav Fischer, Jena.
HICKS, H. 1873. On the Tremadoc rocks in the neigh-bourhood of St. David's, South Wales and theirfossil contents. Quarterly Journal of the Geo-logical Society of London, 29, 39-52, pls 3-5.
HINZ-SCHALLREUTER, I.1995. Muscheln (Pelecypoda)aus dem Mittelkambrium von Bornholm.Geschiebekunde aktuell, 11, 71-84.
HINZ-SCHALLREUTER, I. 2000. Middle CambrianBivalvia from Bornholm and a review of Cam-brian bivalved Mollusca. Revista Espanola deMicropaleontologia, 32, 225-242.
ISBERG, O. 1934. Studien uber Lamellibranchiaten desLeptaenakalkes in Dalarna. Hakan OhlssonsBuchdruckerei, Lund.
JELL, P. A. 1980. Earliest known pelecypod on Earth.A new Early Cambrian genus from South Aus-tralia. Alcheringa, 4, 233-239.
JOHNSTON, P. A. 1996. Middle Ordovician (Caradoc)pelecypods from the Advance Formation, BritishColumbia. Bulletin of the Geological Survey ofCanada, 491, 113-129.
JOHNSTON, P.A. & ZHANG RENJIE. 1998. Re-interpre-tation of Sinodora Pojeta and Zhang, 1984, anunusual bivalve (Trigonioida) from the Devonianof China. In: JOHNSTON, P. A. & HAGGART,J. W. (eds) Bivalves: An Eon of Evolution—Paleo-biological Studies Honoring Norman D. NewellUniversity of Calgary Press, Calgary. 361-375.
KHALFIN, L. L. 1958. Plastinchatozhabernye mollyuskiordovika Chu-Iliyskikh gor. Trudy Geologich-eskogo Instituta Akademii Nauk SSSR, 9,130-136(in Russian).
KOROBKOV, I. A. 1954. Sprvochnik i metodicheskoerukovodstro po tretichnum Mollyuskam Plastin-chatozhabernye. Nauchno-tecnoi" Isledov,Leningradskoi Otdelenie (in Russian).
KRASILOVA, I. N. 1970. Droustvortchatie MolliouskiSrednego i Verkhnego Ordovika Khr. Sette-Daban. In: ROSMAN, K. S., IVANOVA, V. A.,KRASILOVA, I. N. & MODZALEVSKAIA, E. A. (eds)Biostratigrafiya verkhnego ordovika Severo-Vostoka SSSR. Trudy Geologicheskogo InstitutaAkademii Nauk SSSR, 205.168-187, pls 26-27 (inRussian).
KRASILOVA, I. N. 1976. Noyi rod Siberoctenia(Bivalvia) iz ordovika Sibisrskoi Plarformy.Paleontologicheskii Zhurnal, 10, 144-147 (inRussian).
KRASILOVA, I. N. 1979. In: MENNER, V. V. (ed.) Faunaordovika srednei" Sibiry. Trudy GeologicheskogoInstituta Akademii Nauk SSSR, 330 (in Russian).
KRIZ, J. 1995. Coxiconcha Babin, 1966 from theLlanvirn of the Prague Basin (Bivalvia,
Ordovician, Bohemia) and the function of some"accessoric"' muscles in Recent and fossilBivalvia. Vestnik Ceskeho geologickeho ustavu,70, 45-50.
LAMONT, A. 1946. Lamellibranchs from the LowerDrummuck Group, (Ashgillian), Girvan. Scot-land. Cement, Lime and Gravel, 20, 364-366.
LILJEDAHL, L. 1984. Silurian silicified bivalves fromGotland. Sveriges Geologiska Undersokning,Series C 804. 1-82.
McALESTER, A. L. 1965. Systematics, affinities andlife habits of Babinka, a transitional Ordovicianlucinoid bivalve. Palaeontology, 8, 231-246.
MCALESTER, A. L. 1966. Evolutionary and system-atic implications of a transitional Ordovician luci-noid bivalve. Malacologia, 3, 433-439.
McKINNON, D. L 1982. Tuarangia paparua n. gen. andn. sp. A late Middle Cambrian pelecypod fromNew Zealand. Journal of Paleontology, 56,589-598.
MILLER, S. A. 1874. Monograph of the Lamelli-branchiata of the Cincinnati group. CincinnatiQuarterly Journal of Science, 1,211-231.
MILLER, S. A. 1877. The American Palaeozoic Fossils,a Catalogue of the Genera and Species. Publishedby the author, Cincinnati.
MORRIS, N. J. 1978. The infaunal descendants of theCycloconchidae: an outline of the evolutionaryhistory of the Heteroconchia. SuperfamiliesCycloconchacea to Chamacea. PhilosophicalTransactions of the Royal Society of London,Series B, 284, 259-275.
MORRIS, N. J. & FORTEY, R. A. 1976. The signifi-cance of Tironucula gen. nov. to the study ofbivalve evolution. Journal of Paleontology, 50,701-709.
NEUMAN, R. B., BRUTON, D. L. & POJETA. J. 1997.Fossils from the Ordovician 'Upper HovinGroup' (Caradoc-Ashgill), Trondheim Region,Norway. Norges geologiske undersokelse Bulletin,432, 25-58.
NEUMAYR, M. 1884. Zur Morphologie der Bivalven-schlosses. Kaiserlich-Koniglichen Akademie derWissenschaften, Wien. Mathematische-Naturwis-senschaftliche Klasse, Sitzungberichte, Abteilung1,88,385-418.
NEWELL, N. D. 1965. Classification of the Bivalvia.American Museum Novitates, 2206.
NEWELL, N. D. & BOYD. D. W. 1975. Parallel evo-lution in early trigoniacean bivalves. Bulletin ofthe American Museum of Natural History, 154.53-162.
PELSENEER, P. 1889. Sur la classification phylogene-tique des pelecypodes. Bulletin Scientifique de laFrance et de la Belgique, 20, 27-52.
PFAB, L. 1934. Revision der Taxodonta des bohmis-chen Silurs. Palaeontographica, Abteilung A, 80.195-253, pls 2-4.
PHILLIPS. J. 1848. In: PHILLIPS, J. & SALTER. J. W.Palaeontological appendix to Professor JohnPhillips' memoir on the Malvern Hills comparedwith the Palaeozoic districts of Abberley etc.Memoir of the Geological Survey of GreatBritain, 2, 331-386. pls 4-30.

ORDOVICIAN BIVALVE BIOGEOGRAPHY 51
POJETA, J, 1962. The pelecypod genus Byssonychiaas it occurs in the Cincinnatian at Cincinnati,Ohio. Palaeontographica Americana, 4(30),170-216.
POJETA, J. 1966. North American Ambonychiidae.Palaeontographica Americana, 5(36), 131-238.
POJETA, J. 1971. Review of Ordovician Pelecypods.United States Geological Survey, ProfessionalPapers, 695.
POJETA,J. 1973. Fordilla troyensis Barrande: the oldestknown pelecypod. Science, 180, 866-868.
POJETA, J. 1975. Fordilla troyensis Barrande and earlypelecypod phylogeny. Bulletins of American Pale-ontology, 67(287), 363-384.
POJETA, J. 1978. The origin and early taxonomic diver-sification of pelecypods. Philosophical Trans-actions of the Royal Society of London, Series B,284, 225-246.
POJETA, J. 1980. Molluscan phylogeny. Tulane Studiesin Geology and Paleontology, 16, 55-80.
POJETA, J. 1988. The origin and Paleozoic diversifica-tion of solemyoid bivalves. Memoir of the NewMexico Bureau of Mines and Mineral Resources,44,201-271.
POJETA,J. 1997. Claudeonychia babini nov. gen. et nov.sp. of Ordovician ambonychiid pelecypods fromCincinnati, Ohio, USA. Geobios MemoireSpecial, 20, 433-36.
POJETA, J. 2000. Cambrian Pelecypoda (Mollusca).American Malacological Bulletin, 15,157-166.
POJETA, J. & GILBERT-TOMLINSON, !. 1977. AustralianOrdovician pelecypod molluscs. Bulletin of theBureau of Mineral Resources, Geology and Geo-physics, 174,1-64, pls 1-29.
POJETA, J. & RUNNEGAR, B. 1985. The early evolutionof Diasome Molluscs. In: TRUEMAN, E. R. &CLARKE, M. R. (eds) The Mollusca, Volume10, Evolution. Academic Press, New York,295-336.
POJETA,J. & ZHANG, RENJIE. 1986. Systematic paleon-tology of the Devonian pelecypods of Guangziand Michigan. In: POJETA, J. (ed.) DevonianRocks and Lower and Middle Devonian Pelecy-pods of Guangzi, China and the Traverse Group,Michigan. United States Geological Survey, Pro-fessional Papers, 1394.
RATTER, V. A. 1999. Silurian Bivalvia from Walesand the Welsh Borderland. PhD thesis, Universityof Wales.
RATTER, V. A. & COPE, !. C. W. 1998. New neo-taxodont bivalves from the Silurian of SouthWales and their phylogenetic significance.Palaeontology, 41, 975-991.
REED, F. R. C. 1946. Notes on some lamellibranchsfrom Quarrel Hill, Girvan. Geological Magazine,83, 201-205, pl. 14.
SALTER, J. W. 1859. Fossils from the base of theTrenton Group. In: Figures and Descriptions ofCanadian Organic Remains. Geological Survey ofCanada, Decade 1,1-47.
SANCHEZ, T. M. 1986. La fauna de bivalves en laFormacion Santa Gertrudis (Ordovicico) de laProvincia de Salta (Argentina). Ameghiniana, 23,131-139.
SANCHEZ, T. M. 1990. Bivalvos del Ordovicico medio-tardio de la Precordillera de San Juan(Argentina). Ameghiniana, 27, 251-261.
SANCHEZ, T. M. 1996. Un nuevo genero de Tironuculi-dae (Bivalvia, Palaeotaxodonta) en el Arenigianodel Noroeste argentino. Geobios, 29, 683-690.
SANCHEZ, T. M. 1997a. Natasiinae, a new subfamily ofthe Tironuculidae (Bivalvia, Palaeotaxodonta)from the Acoite Formation (Early-MiddleArenig), Northwestern Argentine. GeobiosMemoire Special, 20, 471-475.
SANCHEZ, T. M. 19976. Additional Mollusca (Bivalviaand Rostroconchia) from the Suri Formation,Early Ordovician (Arenig), western Argentina.Journal of Paleontology, 71,1046-1054.
SANCHEZ, T. M. 1999a. Late Ordovician (EarlyCaradoc) bivalves from the Sierra de Villicum(Argentine Precordillera). Journal of Paleontol-ogy, 73, 66-76.
SANCHEZ, T. M. 1999b. Caradoc bivalves from theArgentine Precordillera: a local radiation-extinc-tion event. Geobios, 32, 333-340.
SANCHEZ, T. M. & BABIN, C. 1993. Un insolite bivalve,Catamarcaia n.g., de 1'Arenig (Ordovicieninferieur) d'Argentine. Comptes rendus de l'A-cademie des Sciences, Paris, Serie II, 316,265-271.
SANCHEZ, T. M. & BABIN, C. 1994. Los generosRedonia y Catamarcaia (Mollusca, Bivalvia) de laFormacion Suri (Ordovicico Temprano, Oeste deArgentina) y su interes Paleobiogeografico.Revista Espanola de Paleontologia, 9, 81-90.
SANCHEZ, T. M. & SUAREZ-SORUCO, R. 1996. Primerregistro de lyrodesmatidos (Bivalvia, Hetero-conchia) en America del sur (Formacion SanBenito) Ordovicico superior de Bolivia. Memo-rias del XII Congreso Geologico de Bolivia,Tarija, Bolivia, 217-219.
SARDESON, F. W. 1896. The Saint Peter Sandstone.Bulletin of the Minnesota Academy of NaturalSciences, 4, 64-88.
SARDESON, F. W. 1939a. Early pelecypod Vanuxemia inMinnesota. Pan-American Geologist, 71,283-293.
SARDESON, F. W. 1939b. Early pelecypod Cyrtodontain Minnesota. Pan-American Geologist, 71,337-346.
SHARPE, D. 1853. Description of the new species ofZoophyta and Mollusca. Appendix B. In:RIBEIRO, C, SHARPE, D. & !ONES, T. R. On theCarboniferous and Silurian formations in theneighbourhood of Bussaco in Portugal. QuarterlyJournal of the Geological Society of London, 9,146-158, pls 7-9.
SHERGOLD, J. H., GORTER, j. D, NICOLL, R. S. &HAINES, P. W. 1991. Stratigraphy of the PacootaSandstone (Cambrian-Ordovician), AmadeusBasin, N. T. Bulletin of the Bureau of MineralResources, Geology and Geophysics, 237,1-14.
SOOT-RYEN, H. 1964. Nuculoid pelecypods from theSilurian of Gotland. Arkiv for Mineralogioch Geologi, Kungliga Svenska Vetenskap-sakademien, Series 3, 28, 489-519.
SOOT-RYEN, H. 1969. A new species of Babinka(Bivalvia) from the Lower Ordovician of Oland,Sweden. Palaeontology, 12,173-177, pls 1-5.

52 JOHN C. W. COPE
SOOT-RYEN, H. & SOOT-RYEN, T. 1960. The MiddleOrdovician of the Oslo Region, Norway. 11, Pele-cypoda. Norsk Geologisk Tidsskrift, 40, 81-122,pls I-IV.
STANLEY, S. M. 1977. Co-adaptation in the Trigoniidae.a remarkable family of burrowing Bivalves.Palaeontology, 20, 869-899.
STAROBOGATOV, YA. L. 1971. Systematic position ofthe Ordovician bivalve Babinka. PaleontologicalJournal, 5, 384-387 (English translation byAmerican Geological Institute published 1972).
STEINMANN, G. & HOEK, H. 1912. Das Silur und Cam-brium des Hochlandes von Bolivia und ihreFauna. Neues Jahrbuch fur Mineralogie, Geologieund Palaontologie,Beilige-Band, 34, 176-252.
STUBBLEFIELD, C. J. 1938. The types and figuredspecimens in Phillips and Salter's palaeonto-logical appendix to John Phillips' memoir on'The Malvern Hills compared with the Palaeo-zoic districts of Abberley. etc.'. Summary ofProgress of the Geological Survey of GreatBritain and the Museum of Practical Geology.1936,27-51.
TAYLOR, J. D. & GLOVER, E. A. 2000. Functionalanatomy, chemosymbiosis and evolution of theLucinidae. In: HARPER, E. M, TAYLOR, J. D. &CRAME, J. A. (eds) The Evolutionary Biology ofthe Bivalvia. Geological Society. London, SpecialPublications, 177, 207-225.
THORAL, M. 1935. Contribution a l'etude paleon-tologique de I'Ordovicien inferieur de la MontagneNoire et revision sommaire de lafaune cambriennede la Montagne Noire. Imprimerie Charite,Montpellier.
TONI. R. T. 1975. Upper Ordovician bivalves from theOslo Region. Norway. Norsk GeologiskTidsskrift, 55, 135-156.
TORSVIK, T. H. 1998. Palaeozoic palaeogeography: ANorth Atlantic viewpoint. GFF, 120, 109-118.
TROMELIN, G. LE G. DE & LEBESCONTE P. 1875. Notesur quelques fossiles des gres Siluriens de Saint-Germain-sur-Ile, la Bouxiere, Champeau. Edi-tions Cottonec, Quimper.
TROMELIN. G. LE G. DE & LEBESCONTE P. 1876. Essaid'un catalogue raisonne des fossiles siluriensdes departements de Maine-et-Loire. de la
Loire-Inferieure et du Morbihan. avec des obser-vations sur les terrains Paleozoiques de 1'Ouest dela France. Comptes rendus du Congres de l'Asso-ciation Francaise pour l'Avancernent des Sciences,Nantes, 1875,601-661.
TROMELIN. G. LE. G. DE & LEBESCONTE P. 1877. Etudesur les terrains Paleozoiques de la Basse-Normandie. Comptes rendus du Congres del'Association Francaise pour l'Avancement desSciences, Le Havre, 1876. 493-501.
TUNNic LIFF. S. P. 1982. A revision of late Ordovicianbivalves from Pomeroy. Co. Tyrone. Ireland.Palaeontology. 25. 43-88.
T U N N i c L I F f . S. P. 1987. Caradocian bivalve molluscsfrom Wales. Palaeontology, 30. 677-690.
U L R i c H , E. O. 1879. New genera and species of fossilsfrom the Lower Silurian about Cincinnati. Journalof the Cincinnati Society of Natural History, 2,1-22.
ULRICH. E. O. 1892. New Lower Silurian Lamelli-branchiata chiefly from Minnesota rocks. Geo-logical and Natural Historv Survey of Minnesota.19.211-248.
U L R i c H . E. O. 1894. The Lower Silurian Lamelli-branchiata of Minnesota. In: Final Report of theGeological and Natural History Survey of Min-nesota, Volume 3. 475-628 (published and distrib-uted under separate cover prior to publication ofthe entire Volume 3 in 1897).
VAN DE POeL. L. 1959. Faune malacologique duHervien. Institut Royal des Sciences Naturelles deBelgique, Bulletin. 35(15) 1-26.35(16) 1-28.
WALLER. T. R. 1990. The evolution of ligamentsystems in the Bivalvia. In: MORTON. B. (ed.) TheBivalvia — Proceedings of a Symposium inHonour of Sir Charles Maurice Yonge. HongKong University Press. Hong Kong. 49-71.
WALLER. T. R. 1998. Origin of the molluscan classBivalvia and a phylogeny of major groups. In:JOHNSTION. P. A. & HAGGART. J. W. (eds)Bivalves: An Eon of Evolution — PaleobiologicalStudies Honoring Norman D. Newell. Universityof Calgary Press. Calgary. 1-45.
W H I I F I E L D . R. P. 1893. Contributions to the paleon-tology of Ohio. Ohio Division of the GeologicalSurvey. 7(2). 407-494. pl. 13.

Phytogeny of the Reedocalymeninae (Trilobita): implications forEarly Ordovician biogeography of Gondwana
SAMUEL T. TURVEYDepartment of Earth Sciences, Oxford University, Parks Road, Oxford OX1 3PR,
United Kingdom (e-mail: [email protected])
Abstract: The shallow-water Neseuretus Association has been recognized by many authorsas an important indicator of the former extent of Gondwana during the Early Ordovician.Phylogenetic relationships of the Reedocalymeninae (Arenig-?Early Silurian) were inves-tigated using cladistic analysis, incorporating 22 species of Neseuretus as well as represen-tatives of all other reedocalymenine genera. The area cladogram derived from this analysiscontains four subclades each containing areas that span much of the palaeogeographicextent of Gondwana, which are interpreted as representing separate biogeographic eventsduring the evolution of the subfamily. As the majority of taxa included in the analysis areassociated with shallow shelf facies, this suggests that no significant environmental barriersexisted across the continent during the Early Ordovician. Consideration of area relation-ships both for different subclades within the area cladogram, and within a consensus areacladogram, also supports the idea of a faunal cline between eastern and western Gondwanaduring this time interval. Further cladistic analysis of different trilobite taxa can be used totest these ideas.
The palaeogeography of the Early Palaeozoic ispoorly understood relative to that of more recentgeological history, and faunal data have beenwidely used to determine continental configur-ations during this time period. Biogeographicsensitivity varies between different taxa: whereassome groups (e.g. graptolites) commonly cannotbe used to detect palaeogeographic boundaries,trilobites have been recognized as being ableto indicate biogeographic patterns which arecongruent with abiotic geological informationsuch as palaeomagnetism, geodynamic featuresand climatically sensitive sediments (Fortey &Mellish 1992).
Faunal provinciality was very high during theEarly Ordovician (Arenig-Llanvirn), as a resultof both continental and climatic differentiation(Cocks & Fortey 1988, 1990), and geographicseparation is regarded as a more importantcontrol on faunal composition than biofaciesduring this interval (Fortey & Mellish 1992). Thedistributions of different trilobite taxa can beused to differentiate Laurentia, Baltica andGondwana, the three main continental regionswhich existed during the Ordovician (Zhou &Dean 1989), although some faunal admixturebetween regions at similar latitudes on thedifferent continents can also be detected, such asthe presence of Baltican genera in carbonates atSobova, south central Turkey (Dean 1973,1975).
Whereas Laurentian and Baltican trilobitefaunas show little intracontinental provinciality
during the Early Ordovician, considerablefaunal differences have long been detectedbetween different geographically separatedGondwanan cratonic faunas during this interval(e.g. Stubblefield 1939). For example, westernand eastern regions of the palaeocontinent arecharacterized by different trilobite groups, andhave been recognized respectively by severalauthors as the Selenopeltis Province' or 'caly-menacean-dalmanitacean Province' and the'Asaphopsis Province' (Whittington & Hughes1972; Cocks & Fortey 1990). Gondwana, thelargest of the Early Ordovician palaeoconti-nents, stretched from polar to equatorial lati-tudes, extending from the South Pole to morethan 30°N (Cocks 2001), so unlike Laurentia orBaltica it represented a multiclimate region(Spjeldnaes 1961, 1981; Cocks & Fortey 1988,1990). As temperature is regarded as a first-order control on faunal distribution (Fortey& Mellish 1992), the corresponding well-developed climatic gradient across the palaeo-continent can account for much of this faunalvariation, with Palmer (1972) and Whittington(1973) being among the first to consider that thedistribution of Early Ordovician trilobitesaround Gondwana may be related to latitudinalbelts around a North African South Pole. Geo-graphic separation must also have determinedpatterns of trilobite provinciality and endemicityto some extent, both across the continentsforming the core of Gondwana, and betweenthese continents and the numerous small
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations, Geological Society, London, Special Publications, 194, 53-68.0305-8719/02/S15.00 © The Geological Society of London 2002.

54 SAMUEL T. TURVEY
tectonic plates and terranes distributed alongits borders or adjacent to it (termed 'peri-Gondwana'). However, the characterization andgeographic location of these smaller tectonicunits during the Early Ordovician is still stronglydebated, even for geographic regions such aswestern Europe which have been exposed toconsiderable study, and few authors agree on thenumber of oceans, tectonic plates and terranesinvolved (Scotese & McKerrow 1990; Paris 1998;Cocks 2001),
Different authors have used trilobite datato propose differing biogeographic patternsfor Early Ordovician Gondwana and peri-Gondwana. Area relationships across thepalaeocontinent have largely been investigatedon the basis of overall faunal similarity, usingboth quantitative (e.g. Whittington & Hughes1972; Fortey & Mellish 1992) and qualitativeapproaches. Cladistic biogeographic analysis ofEarly Ordovician taxa has tended instead toconsider the relationships between Laurentia,Baltica and Gondwana (e.g. Peers 1997). SouthChina and Australia have been regarded asrepresenting a distinct biogeographic regionseparate from central Europe during this timeperiod by some authors (Palmer 1972; Cocks &Fortey 1988, 1990; Peers 1997). An alternativedivision of Gondwana into a Sino-EuropeanRegion and a South American-AustralianRegion has also been advocated (Li 1994).Other authors have emphasized the distinctionbetween SE Asian and European regions.Zhou & Dean (1989) suggested that trilobitefaunas from all parts of Gondwanan and peri-Gondwanan east Asia may belong to a singlefaunal province, in turn divisible into sub-provinces, with further research into SE AsianEarly Ordovician trilobites highlighting faunalsimilarities between South China, Tarim andIndo-China (Zhou et al 1998a, b). A distinctNorthern Gondwanan Province, consisting ofNorth Africa, the Middle East, and western andcentral Europe exclusive of Avalonia, has alsobeen recognized and subdivided into differentdomains partly on the basis of trilobite data(Paris 1998). There seems to be no faunalevidence for the presence of oceanographic orother environmental barriers separatingdifferent areas of Ordovician Gondwana (Zhou& Dean 1989). Spjeldnaes (1961, 1981) andCocks & Fortey (1988,1990) have suggested thatthe relationships between different Gondwanancratonic trilobite faunas are best understood interms of a geographic and climatic clineacross the entire palaeocontinent. Gondwananfaunas can thus be interpreted as being grada-tional across the large palaeocontinent, with
intermediate faunal regions variously allied bydifferent authors to both eastern and westernbiogeographic provinces representing 'mixed'faunas.
The Neseuretus AssociationBiofacies has been recognized as an importantdeterminant of the quality of the biogeographicsignal provided by different trilobite faunas. Thegeneralized palaeogeographic model proposedby Fortey & Owens (1978) suggests that, fora series of different trilobite associations occur-ring in different water depths along a continent-edge profile, the associations occurring nearestinshore should display the highest degree ofendemicity; the environmental barrier separ-ating them from neighbouring geographicregions is higher than for deeper-water associ-ations and so is more likely to act as a barrier togene flow and induce speciation. According tothis model, shallow-water trilobite associationscan thus potentially provide the highest resolu-tion biogeographic information. However,shallow-water trilobite associations can only beof use in biogeographic analysis if the genera orsubfamilies they contain are geographicallywidespread rather than localized.
The trilobite association expected to providethe best biogeographic information according tothese criteria is the Neseuretus Association, firstidentified by the term 'Calymene Tristani-Stufe'(Born 1918; see also Hammann 1983) and alsoreferred to variously as 'Neseuretus shales', the'Neseuretus Community' or the 'NeseuretusFauna'. This association, described by Fortey &Morris (1982), is generally found in coarse- tofine-grained elastics such as decalcified iron-stones, mudrocks and the Gres Armoricainor Armorican Quartzite, which extends fromBrittany to Iberia and also probably occurs ineastern Newfoundland (the Armoricain Grit' ofVan Ingen 1914). The reedocalymenine genusNeseuretus is the dominant faunal component. Itoccurs either on its own, or associated with asparse, low-diversity trilobite fauna, whichincludes genera such as Kerfornella, Plaesia-comia, Eohomalonotus, Iberocoryphe, Cro-zonaspis, Taihungshania, Ogyginus, Merliniaand Liomegalaspides in different parts of theassociation's geographic range (Fortey & Owens1978; Fortey & Morris 1982; El-Khayal &Romano 1985; Rabano 1990; Zhou et al. 1998a).
The Neseuretus Association has the widestgeographic distribution of any Early OrdovicianGondwanan trilobite association, occurring inAvalonia, southern and western Europe, NorthAfrica, the Middle East, Indo-China, South

TRILOBITE BIOGEOGRAPHY OF GONDWANA 55
Fig. 1. Early Ordovician (Arenig) palaeogeography, showing the Arenig-basal Caradoc geographicdistribution of the 22 Neseuretus species included in the cladistic analysis. Key: 1, South China (YangtzePlatform): N. concavus (Arenig), N. elegans (Arenig-Llanvirn), N. intermedius (Arenig), N.planus (Arenig),N. shensiensis (Arenig); 2, Indo-China: N. elegans (Arenig-Llanvirn), N. turveyi (Llanvirn); 3, Arabia: N.tristani (Llanvirn); 4, South Turkey: N. sexangulus (Arenig); 5, Libya: N. tristani (Llanvirn); 6, France(Armorica, Montagne Noire); N. arenosus (Arenig), N. tristani (Llanvirn); 7, Spain (Iberia): N. avus(Llanvirn), N. henkei (Llanvirn), N. leonensis (?Arenig/Llanvirn), N. tristani (Llanvirn-basal Caradoc); 8, Anti-Atlas, Morocco: N. attenuatus (Llanvirn), N. tristani (Llanvirn); 9, Eastern Avalonia: N. caerhunensis (Arenig),N. monensis (Arenig), N. murchisoni (Arenig), N. parvifrons (Arenig), N. ramseyensis (Arenig); 10, WesternAvalonia: N. vaningeni (Arenig); 11: South America: N. chaschuilensis (Arenig, Argentina), N. lipanensis(Arenig-Llanvirn, Argentina), N. sanlucasensis (Llanvirn, Bolivia). Map generated by David Lees (NaturalHistory Museum, London).
China and the Central Andean basin of SouthAmerica, and is regarded as one of the morereliable indicators of the former extent ofGondwana (although species assigned toNeseuretus or its junior synonym Synhomalono-tus from various Central Asian regions, some ofwhich were listed by Fortey & Morris (1982) andRabano (1990) as valid representatives of thegenus, represent different reedocalymeninegenera) (Fig. 1). As early as 1937, Kobayashinoted that the genus (as Synhomalonotus] wascharacteristic of the 'Euro-Meridional' Provincein the Early Ordovician, and later establishedthe term 'Asaphopsis-Taihungshania-Neseure-tus Fauna' to describe the Early and MiddleOrdovician southern faunal province(Kobayashi 1976, 1987). The association isinterpreted as typically occurring in shallow
inner-shelf environments (Fortey & Morris1982). However, various species of Neseuretushave also been recorded in shallow outer-shelfdeposits across much of the geographic range ofthe genus, in faunas composed of slightlydeeper-water trilobite taxa such as Colpo-coryphe, Salterocoryphe, Placoparia, Zelis-zkella, Ectillaenus and Hanchungolithus,indicating that the genus had a wider palaeo-bathymetric tolerance than has previously beensuggested (Dean 1966; Yin & Lee 1978;Courtessole et al. 1981; Beckly 1989; Henry1989; Rabano 1990; Zhou et al. 1998a). TheNeseuretus Association was originally regardedas an indicator of cold water, circumpolar shelfseas (Fortey & Morris 1982). This interpretationhas been revised due to the occurrence of theassociation in Early Ordovician tropical regions

56 SAMUEL T.TURVEY
such as the Yangtze Platform of the South ChinaPlate; Neseuretus is now regarded as a eurytopicgenus with exceptional latitudinal tolerance,adapted to general inshore clastic environments(Cocks & Fortey 1988).
Neseuretus has traditionally been regarded asbasal within the Reedocalymeninae, represent-ing the ancestral stock from which other generaevolved (e.g. Whittington 1966b; Dean 1975).Dean (1967a, b) and Zhou & Dean (1989) inter-preted Neseuretus as having originated insouthern and western Europe, continuing toevolve in its region of origin but also dispersingeastwards around the Gondwanan margins toarrive in South China during the Llanvirn.However, these ideas on faunal migration havebeen biased by differing local preservation indifferent geographic regions of the appropriateshallow-water clastic sediments to whichNeseuretus was restricted; for example, appro-priate transgressive facies are only known fromthe lower Arenig in south Wales (Fortey &Owens 1987). They have also been based oninaccurate stratigraphic correlation betweendifferent Gondwanan regions; for example,although the South Chinese Neseuretus specieswere originally interpreted as occurring inLlanvirn deposits (Lu 1975), these are nowconsidered to have been deposited during theArenig (Mu 1974; Chen et al. 1995; Zhou et al.1998a). Further problems for biogeographicinterpretation have arisen as a result of someauthors regarding Synhomalonotus as a separ-ate genus from Neseuretus (e.g. Moore 1959;Ross 1975). Neseuretus stocks in differentGondwanan regions have alternatively beeninterpreted as representing separate lineages,which may have been environmentally sepa-rated because of the transience of the Neseure-tus Association in a marginal setting (Beckly1989). Fortey & Owens (1987) considered thatindividual Neseuretus species probably rangedwidely over Gondwana, as well as observing,following Hammann (1983), that differentspecies were associated with particular facies,suggesting ecological differentiation; however,so far only N. tristani (Brongniart in Desmarest,1817), the geologically youngest species ofNeseuretus, has been recorded from severalOrdovician palaeoplates.
Phylogenetic analysis
Although Neseuretus is regarded as an import-ant genus both biogeographically and for under-standing the evolution of the Calymenidae, noquantitative phylogenetic analysis has previ-ously been conducted to determine the pattern
of relationships between different Neseuretusspecies. This is in part because of the largenumber of species currently recognized withinthe genus. Whereas the seven other generaassigned to the Reedocalymeninae (Caly-menella, Calymenesun, Neseuretinus, Pradoella.Reedocalymene, Sarrabesia and Vietnamia)contain relatively few species, over 60 differentspecies or subspecies have at various times beenestablished for either Neseuretus or Synhoma-lonotus, many of which are poorly known andmay represent invalid taxa. Representatives ofthe Reedocalymeninae are shown in Figure 2.
Cladistic analysis was conducted on theReedocalymeninae, incorporating 22 of thebetter-known species of Neseuretus and arepresentative species from each of the otherreedocalymenine genera. Type material wasstudied where possible, although additionalmaterial for many taxa was also considered, andtype species were included for most reedocaly-menine genera. Reedocalymene expansa Yi.1957 was used instead of R. unicornis (Reed.1917), as the type species is poorly known andthe genus was revised comprehensively on thebasis of extensive new material of R. expansa byPeng et al. (2000). Calymenella preboisseliBeckly, 1989 was included on stratigraphicgrounds instead of C. boisseli Bergeron. 1890.as this species represents the only Arenigrepresentative of an otherwise Late Ordoviciangenus. Calymenesun was coded using C. granu-losa Lu, 1975 instead of C. tingi (Sun, 1931), aswell-preserved material representing this specieswas available for analysis. Bavarilla hofensis(Barrande, 1868), Pharostomina oepiki Sdzuy.1955 and Protocalymene mcallisteri Ross, 1967were selected as outgroup taxa to determinecharacter polarity within the Reedocalymeni-nae, as they have been interpreted by previousauthors as either closely related to the Caly-menidae or representing basal calymenids. or inthe case of Protocalymene ( = "aff. Calymenidiussp. indet.' of Whittington 1965) as a possiblecalymenid (Whittington 1966b; Fortey 1983;Hammann 1983; D.J.Siveter pers. comni. 2001).
Twenty-five exoskeletal characters werecoded for the analysis. As most Neseuretusspecies are only known from holaspid cranidiaand pygidia, all of the characters used weretaken from these two exoskeletal regions. Thisprevented the cladistic data matrix from con-taining large amounts of missing values, whichcould lower the resolution of the analysis byproducing large numbers of equally parsi-monious trees, or produce spurious theories ofcharacter evolution (Kitching et al. 1998).Certain characters which have been used by
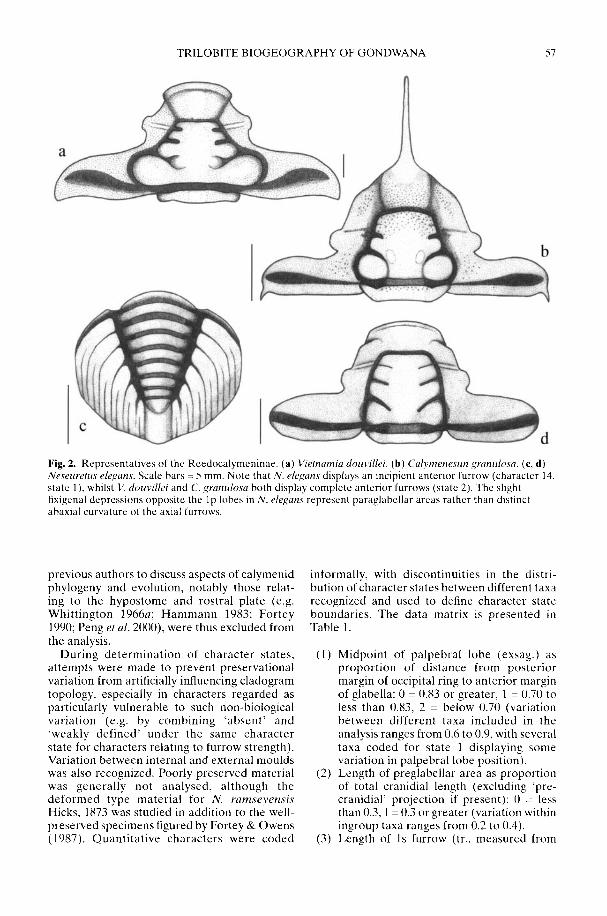
TRILOBITE BIOGEOGRAPHY OF GONDWANA 57
Fig. 2. Representatives of the Reedocalymeninae. (a) Vietnamia douvillei. (b) Calymenesun granulosa. (c, d)Neseuretus elegans. Scale bars = 5 mm. Note that N. elegans displays an incipient anterior furrow (character 14,state 1), whilst V. douvillei and C. granulosa both display complete anterior furrows (state 2). The slightfixigenal depressions opposite the lp lobes in N. elegans represent paraglabellar areas rather than distinctabaxial curvature of the axial furrows.
previous authors to discuss aspects of calymenidphylogeny and evolution, notably those relat-ing to the hypostome and rostral plate (e.g.Whittington 1966a; Hammann 1983; Fortey1990; Peng et al 2000), were thus excluded fromthe analysis.
During determination of character states,attempts were made to prevent preservationalvariation from artificially influencing cladogramtopology, especially in characters regarded asparticularly vulnerable to such non-biologicalvariation (e.g. by combining 'absent' and'weakly defined' under the same characterstate for characters relating to furrow strength).Variation between internal and external mouldswas also recognized. Poorly preserved materialwas generally not analysed, although thedeformed type material for N. ramseyensisHicks, 1873 was studied in addition to the well-preserved specimens figured by Fortey & Owens(1987). Quantitative characters were coded
informally, with discontinuities in the distri-bution of character states between different taxarecognized and used to define character stateboundaries. The data matrix is presented inTable 1.
(1) Midpoint of palpebral lobe (exsag.) asproportion of distance from posteriormargin of occipital ring to anterior marginof glabella: 0 = 0.83 or greater, 1 = 0.70 toless than 0.83, 2 = below 0.70 (variationbetween different taxa included in theanalysis ranges from 0.6 to 0.9, with severaltaxa coded for state 1 displaying somevariation in palpebral lobe position).
(2) Length of preglabellar area as proportionof total cranidial length (excluding 'pre-cranidial' projection if present): 0 = lessthan 0.3,1 = 0.3 or greater (variation withiningroup taxa ranges from 0.2 to 0.4).
(3) Length of 1s furrow (tr., measured from

58 SAMUEL T. TURVEY
Table 1. Data matrix for character state distributions used in phylogenetic analysis
Species
Bavarilla hofensis (Barrande, 1868)Calymenella preboisseli Beckly, 1989Calymenesun granulosa Lu, 1975Neseuretinus turcicus (Dean, 1967a)Neseuretus arenosus Dean, 1966N. attenuatus (Gigout, 1951)N. avus Hammann, 1977N. chaschuilensis Vaccari & Waisfeld, 1994N. caerhunensis Beckly, 1989N. concavus Lu, 1975N. elegans Lee in Yin & Lee, 1978N. henkei Hammann, 1977N. intermedius Lu, 1975N. leonensis Hammann, 1983N. lipanensis Waisfeld, 1997N. monensis (Shirley, 1936)N. murchisoni (Salter, 1865a)N. parvifrons (M'Coy in Sedgwick & M'Coy, 1851)N. planus Lu, 1975N. ramseyensis Hicks, 1873N. sanlucasensis Pribyl & Vanek, 1980N. sexangulus Dean, 1971N. shensiensis (Lu, 1957)N. tristani (Brongniart in Desmarest, 1817)N. turvevi Zhou, Dean & Luo in Zhou et al., 2001N. vaningeni Dean in Dean & Martin, 1978Pharostomina oepiki Sdzuy, 1955Pradoella pradoi Hammann, 1977Protocalymene mcallisteri Ross, 1967Reedocalymene expansa Yi, 1957Sarrabesia teichmuelleri Hammann & Leone, 1997Vietnamia douvillei (Mansuy, 1908)
1 21 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5
1 0 1 0 2 1 0 0 0 0 0 1 0 0 1 1 0 0 0 0 1 0 0 0 02 1 1 0 0 0 0 0 0 1 0 0 0 1 0 0 1 B 0 0 1 0 0 1 02 1 0 0 1 1 1 1 0 0 1 1 0 2 0 1 1 1 0 0 2 2 1 0 01 1 1 0 1 1 1 1 1 0 1 0 0 2 1 1 2 2 0 0 0 1 0 1 01 0 1 0 ? ? 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 1 01 0 1 0 0 0 0 0 0 0 0 0 1 1 0 0 ? ? ? ? ? ? ? ? ?A O 1 0 0 0 1 0 1 0 0 1 1 2 0 0 1 B 0 0 0 0 0 1 01 0 1 0 ??? 0 0 0 0 1? ? 00 1 0 0 1 0 01 ?2 1 1 0 0 0 0 0 0 0 0 1 0 1 0 0 1 2 ? 1 ? 1 0 1 00 1 1 0 0 0 0 0 0 0 0 0 1 2 1 0 1 B 0 0 0 0 1 1 01 0 1 0 0 0 1 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 1 00 1 1 0 0 0 1 0 0 0 0 1 0 2 0 0 1 1 0 0 0 0 0 1 00 A 11 ? ? 1 1 1 0 1 0 0 2 0 0 2 1 0 0 0 0 1 1 01 0 1 1 0 ? 0 0 0 0 0 1 0 2 0 0 ? 0 ? ? 0 0 ? ? 00 1 1 1 0 0 1 0 0 0 0 1 0 2 0 0 1 1 0 0 0 0 1 1 02 1 1 0 0 0 0 0 0 0 0 1 0 1 0 0 1 1 0 7 0 0 0 0 01 A 1 0 0 0 0 0 0 0 0 1 0 B 0 0 1 0 0 ? 0 0 0 1 01 1 1 0 0 0 A 0 0 0 0 1 0 2 0 0 1 A 1 1 0 0 0 1 00 0 1 1 ? ? 1 1 1 0 1 0 0 2 0 0 2 0 0 0 0 0 0 1 01 0 1 0 0 0 0 0 0 A 0 1 0 1 0 0 1 2 1 0 0 1 0 0 10 0 1 ? ? ? 1 0 1 0 0 1 0 2 0 0 2 1 0 ? 0 0 0 1 01 0 1 1 ? 0 0 0 0 0 0 0 0 2 0 0 1 ? ? ? ? ? ? ? ?0 0 1 0 0 0 0 0 0 0 0 0 0 2 0 0 1 1 0 0 1 0 1 0 01 0 1 0 0 0 1 0 0 0 0 1 1 2 0 0 1 1 0 0 0 0 0 1 01 0 1 0 0 0 1 0 0 0 0 0 0 2 0 0 1 1 0 0 0 0 0 1 01 0 1 1 0 0 0 0 0 0 0 1 0 2 0 0 1 0 1 0 0 0 1 1 01 0 1 0 2 1 0 0 1 0 0 0 0 0 1 0 2 0 0 ? 1 0 0 0 01 0 1 1 0 0 0 0 0 1 0 1 0 2 0 0 1 B O O 1 0 0 1 02 0 0 0 1 1 0 1 0 0 0 0 0 0 1 0 2 0 0 0 0 0 0 1 02 0 1 0 1 1 1 0 0 0 0 0 0 2 0 1 1 1 0 0 0 0 1 1 00 0 1 0 1 1 2 1 1 0 0 0 0 2 0 0 1 2 0 0 2 1 0 0 10 1 1 0 ? ? 2 1 1 0 0 1 0 2 0 0 1 2 0 0 2 1 0 0 1
Characters and states are as described in the text. Character numbers are listed at the top of the table. Missingdata are indicated with a question mark. Character states listed as 'A' and 'B' indicate multistate coding,where 'A' = (O&l) and 'B' = (1&2).
base of 2p lobe) as proportion of width ofunfurrowed median lobe of glabella (tr.,along same transverse line as that offurrow width): 0 = 0,40 or less width ofmedian lobe, 1 = above 0.40 width ofmedian lobe (such that combined width ofboth 1s furrows is approximately equal toor greater than width of median lobe).
(4) Position of anterior margin of postocularcheek at fixigenal midlength (tr.): 0 =approximately opposite anterior margin oflp lobe, 1 = opposite or anterior to 2p lobe.
(5) Facial suture: 0 = gonatoparian, 1 = propar-ian, 2 = opisthoparian.
(6) Genal spine: 0 = absent, 1 = present.(7) Shape of posterior border furrow: 0 =
equal width along fixigenal margin ('pty-choparioid condition'), 1 = expanding
abaxially, 2 = sigmoidal.(8) Axial furrow adjacent to 1p lobe: 0 =
straight, 1 = curved outwards (abaxiallyconvex).
(9) Glabellar outline: 0 = subrectangular, 1 =tapering strongly forwards ('truncatedcone'/subtrapezoidal).
(10) Strength of glabellar furrows: 0 = stronglydefined, 1 = absent or weakly defined,
(11) 1s furrow (tr.): 0 = does not reach occipitalfurrow, 1 = reaches occipital furrow.
(12) Strength of intermediate lobe (swelling atadaxial base of 2p lobe): 0 = absent orweakly defined, 1 = strongly defined.
(13) Shape of posterior margin of 4p lobe: 0 =lateral margin at similar abaxial position toanterior margin of 3p lobe, 1 = indentedadaxially relative to 3p lobe.

TRILOBITE BIOGEOGRAPHY OF GONDWANA 59
(14) Anterior furrow (furrow running fromanterior margin to distal portion ofpreglabellar furrow, forming lateralboundary to preglabellar area, sensuWhittard 1960; see Fig. 2): 0 = absent, 1 =incipient, only expressed anterolaterally('fixigenal inflation' or 'fixigenal overhang'condition), 2 = extends posteriorly topreglabellar furrow (may superficiallyappear to be a posteriorly positionedanterior border furrow if true anteriorborder furrow is absent, e.g. Dean &Martin 1978, pl. 4, figs 1, 4,7, pl. 5, fig. 1).
(15) Anterior border furrow: 0 = absent orweakly defined, 1 = strongly defined,
(16) Anterior border (dorsal view): 0 = trans-versely straight to gently anteriorlyconvex, 1 = strongly convex and pointed,possibly forming distinct anterior 'pre-cranidiaP projection.
(17) Surface tubercles (size and density): 0 = notuberculation, 1 = light tuberculation, 2 =heavy tuberculation.
(18) Number of pygidial axial rings: 0 = up toand including five, 1 = six or seven, 2 =eight or more (the majority of Neseuretusspecies have five to seven axial rings, andseveral are described as having 'six orseven', although the number of axial ringsvaries from two to ten between differenttaxa included in the analysis).
(19) Dorsal outline of terminal piece: 0 = con-tinuous with anterior part of pygidial axis,1 = anterior region laterally expanded('swollen').
(20) Lateral outline of terminal piece: 0 = con-tinuous with anterior part of pygidial axis,1 = inflated.
(21) Postaxial pygidial morphology: 0 = weaklydefined terminal piece, with narrowerpostaxial ridge extending to margin('Neseuretus condition'), 1 = posteriormargin of terminal piece defined by dis-tinct posterior border furrow, with nopostaxial ridge present, 2 = posteriorborder furrow absent, narrow terminalpiece fused with prominent postaxial ridgeto form a continuous elongate structurewhich extends posteriorly to the pygidialmargin ('SarrabesialVietnamia condition').
(22) Pygidial axis anterior to terminal piece: 0 =evenly tapered, 1 = funnel-shaped (taper-ing to a point slightly anterior to theterminal piece, posterior to which the axialfurrows become parallel), 2 = swollen(axial furrows abaxially convex anterior toa parallel-sided posterior section).
(23) Pygidial lateral pleural furrow/vincular
furrow ( = 'cincture' of Dean & Zhou1988): 0 = absent, 1 = present.
(24) Pygidial interpleural furrows: 0 = absent oronly present very close to the pygidialmargin, 1 = clearly defined across much ofpleural fields.
(25) Pygidial rib morphology: 0 = flattened, 1 =ridge-like.
Cladistic analysis was conducted onPAUP*4.0b4a (Swofford 2001), using a heuristicsearch with 50 random addition sequence repli-cates. Of the eight multistate characters, four (5,7,21 and 22) were treated as unordered and four(1, 14, 17 and 18) as ordered. Multistate taxawere coded as polymorphisms. All characterswere unweighted. Searching was performedtwice, once using accelerated transformationoptimization (ACCTRAN) and a second timeusing delayed transformation optimization(DELTRAN).
Within these parameters, four minimal lengthcladograms of 106 steps, occupying a singleisland, were recovered (Consistency Index =0.4151, Retention Index = 0.5867). ACCTRANand DELTRAN optimization resulted in identi-cal sets of cladograms, as did subsequentreweighting of the dataset. The strict consensusof these cladograms is shown in Figure 3a (asemistrict consensus has the same topology).Support for individual nodes within the con-sensus cladogram was assessed with bootstrapanalysis (Felsenstein 1985) using PAUP*4.0b4a;bootstrap values are displayed on the cladogramin Figure 3a. Removing the three taxa with fiveor more missing values (Neseuretus attenuatus,N. chaschuilensis and N. sexangulus} from thecharacter matrix and rerunning the analysis doesnot improve resolution in the Neseuretus sub-clade, recovering five different cladograms of103 steps (Consistency Index = 0.4272, Reten-tion Index = 0.5845) (Fig. 3b), and so theconsensus cladogram in Figure 3a is here used asthe basis for further analysis.
The consensus cladogram will provide thebasis for a systematic revision of the Reedocaly-meninae in a separate study, and it is used hereas the basis for biogeographic analysis. Thereliability of the consensus cladogram obviouslyunderpins both any taxonomic or biogeographicassumptions based on it. As several of the nodesare only weakly supported by bootstrap analysis,some caution should be employed whenevaluating biogeographic ideas based on thetopology of the resultant area cladogram. Aqualitative but straightforward method ofinvestigating the validity of the cladogram is aconsideration of the degree of congruence
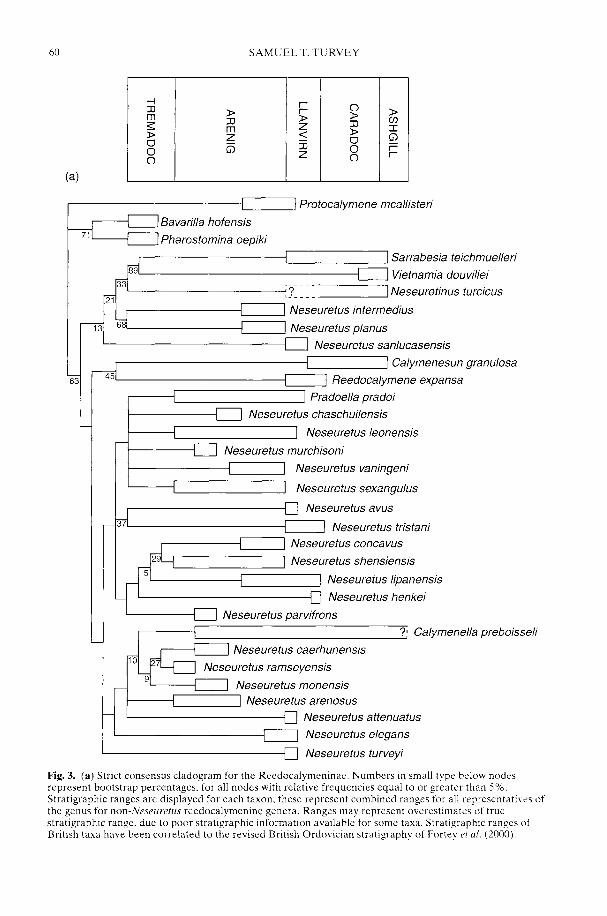
60 SAMUEL T.TURVEY
Fig. 3. (a) Strict consensus cladogram for the Reedocalymeninae. Numbers in small type below nodesrepresent bootstrap percentages, for all nodes with relative frequencies equal to or greater than 5%.Stratigraphic ranges are displayed for each taxon; these represent combined ranges for all representatives ofthe genus for non-Neseuretus reedocalymenine genera. Ranges may represent overestimates of trueStratigraphic range, due to poor Stratigraphic information available for some taxa. Stratigraphic ranges ofBritish taxa have been correlated to the revised British Ordovician stratigraphy of Fortey et al. (2000).

TRILOBITE BIOGEOGRAPHY OF GONDWANA 61
Fig. 3. (b) Neseuretus subclade from second strict consensus cladogram generated by removing N. attenuatus,N. chaschuilensis and N. sexangulus from the dataset.
between the consensus tree and the knownstratigraphic ranges of the ingroup taxa. AsNeseuretus represents a derived member of theReedocalymeninae in the consensus tree, all ofthe major subclades are constrained to haveevolved by the Arenig. This necessitates ghostranges during the Arenig for N. sanlucasensisand the Reedocalymene-Calymenesun subcladeonly, providing fairly good correlation at serieslevel, although the stratigraphic occurrence ofNeseuretinus, Sarrabesia and Vietnamia remainspoorly understood for many Asian localitiesand little confidence can be placed on strati-graphic ranges within this subclade (e.g. Dean1967b; Pillet & de Lapparent 1969; Hammann& Leone 1997; Zhou et al. 1998a).
Cladistic biogeographyThe consensus cladogram in Figure 3a was con-verted into an area cladogram for biogeographicanalysis by substituting the names of thedifferent ingroup taxa with the Early Ordoviciangeographic area(s) from which they have beenrecorded (Fig. 4). The area cladogram is simpli-fied, as area redundancy has been corrected. Forthe non-Neseuretus reedocalymenine taxa, thecombined biogeographic area occupied by allrepresentatives of the genus recognized hereinwas recorded on the area cladogram.
Area delimitation in some cases requires
explanation. France+Spain are regarded as rep-resenting a single biogeographic area, followingthe strong faunal similarities between theseregions noted by previous authors (e.g. Paris1998). Sengor (1984, 1987) proposed thatTurkey, the Sibumasu and Indo-China terranesand areas of Central Asia represented aperi-Gondwanan continent called Cimmeria.However, Turkey represents a complex region,with northern and southern Turkey now inter-preted as having occupied separate palaeogeo-graphic positions during the Ordovician.Southern Turkey, source of the Turkish materialof Neseuretus and Neseuretinus, was tentativelyassigned to a high-latitude position on themargin of Gondwana at the eastern end of thepresent-day Mediterranean (Dean et al, 2000;Cocks 2001), the eastern region of the NorthGondwanan Province of Paris (1998). Followingthese authors, southern Turkey is here pro-visionally interpreted as having been separatefrom Cimmeria during the Ordovician, and isgrouped with Sardinia, another eastern Mediter-ranean component of Paris' (1998) NorthGondwanan Province. As noted above. EarlyOrdovician faunas from South China, Tarim andIndo-China also share strong similarities (e.g.Zhou & Dean 1989), with the two South ChineseNeseuretus species N. elegans Yin in Yin & Lee,1978 and N. tungtzuensis Sheng, 1958 alsorecorded from Indo-China by Zhou et al. (1998a,

62 SAMUEL T. TURVEY
Fig. 4. Area cladogram derived from the strict consensus cladogram in Figure 3a. The nodes numbered 1 to 4are discussed in the text.
2001). These three regions are here interpretedas representing a single Early Ordovician SEAsian palaeogeographic area. A restrictedinterpretation of Cimmeria is thus employed inthis analysis.
Area assignment was generally straight-forward, with the exception of some Asian andAustralian taxa. The Himalayan species Caly-mene nivalis Salter, 1865b was interpreted asrepresenting Neseuretus by Dean (1967b, 1975),Fortey & Morris (1982), Morris & Fortey (1985)and Rabano (1990); examination of the typematerial in the Natural History Museum,London, suggests that this species should insteadbe referred to Sarrabesia. 'Synchomalonotus'sulcatus Kolobova in Sokolov & Yolkin, 1978,from the Ashgill of Uzbekistan, previouslyreassigned to Sarrabesia by Hammann & Leone(1997), is here interpreted as a species ofVietnamia. Calvmenesun longinasuta Dean &Zhou, 1988, from the early Ashgill of the ZapValley, SE Turkey, is extremely similar toNeseuretinus turcicus, and is certainly con-generic. Several different authors (Banks 1988;Corbett & Banks 1974; Legg 1976) haverecorded the presence of reedocalymenine taxa
in Australia, but the taxonomic position of thesespecimens has remained unclear. Edgecombe etal. (1999) tentatively assigned some of thismaterial to Sarrabesia; although other Aus-tralian material may well represent differentreedocalymenine taxa, only Sarrabesia isconsidered to occur in Australia during theOrdovician in this analysis.
The consensus cladogram indicates thatNeseuretus as currently understood representsboth a paraphyletic and a polyphyletic genus.Whereas Neseuretus sensu stricto (i.e. Neseuretusexclusive of N. sanlucasensis, N. intermedius andN. planus) is known to have occurred inshallow inner-shelf conditions amenable to high-resolution biogeographic analysis, and Caly-menella and Pradoella occupied similarshallow-water conditions during the EarlyOrdovician (e.g. Beckly 1989; Rabano 1990), theecological requirements of Middle OrdovicianAsian reedocalymenine taxa, and the speciespreviously assigned to Neseuretus which occur atthe base of the Neseuretinus-Sarrabesia-Vietnamia subclade, are less well understoodor are known to have differed. Neseuretinusturcicus is recorded from offshore shallow shelf

TRILOBITE BIOGEOGRAPHY OF GONDWANA 63
conditions in the Portixeddu and Tuviois for-mations, Sardinia (Leone et al. 1991, as ICaly-menesun sp.; Hammann & Leone 1997), andSarrabesia teichmuelleri occurs in coarse-grained sandstones of the Punta SerpeddiFormation, Sardinia, which are also indicative ofshelf facies (Hammann & Leone 1997). N. long-inasuta occurs alongside a varied trilobiteassemblage in shales and mudstones of the SortTepe Formation in Turkey, suggestive of similarenvironmental conditions (Dean & Zhou 1988).Both Neseuretinus birmanicus and Vietnamiadouvillei are known from siltstones from theupper Na Mo Formation in Vietnam; the for-mation's depositional environment is poorlyunderstood, but both lithofacies and biofaciescomparisons suggest that it represents shelffacies (Tong-Dzuy Thanh, pers. comm. 2001)."Neseuretus" intermedius occurs in outer shelffacies on the Yangtze Platform, in associationwith characteristically shelf-slope taxa such asraphiophorids and Nileus (Wang et al. 1987, as N.xiadongensis). Reedocalymene expansa is abun-dant in shales from the Miaopo Formation of theYangtze Platform, which has been interpreted asrepresenting a deep-water aulacogenic environ-ment by Chen & Qiu (1986). Calymenesun alsooccurs in deep-water facies (e.g. Tripp et al.1989).
However, all of these other reedocalymeninetaxa still show relatively restricted geographicranges, which suggests that their incorporationinto cladistic biogeographic analysis will notlower the resolution of the pattern of arearelationships (although see the discussion onthe possible geographic range of Neseuretinusin Hammann & Leone 1997, p. 114). Similarly,although most of these taxa occurred during theCaradoc or Ashgill, a time period with differentcontinental configurations to the Arenig-Llanvirn interval (Scotese & McKerrow 1990),the consensus cladogram indicates that most ofthe divergences involving these lineages arestratigraphically constrained to have occurredduring the Early Ordovician. All of the reedoca-lymenine taxa included in cladistic analysis aretherefore also included in cladistic bio-geographic analysis. However, the occurrence ofCalymenella in Australia during the EarlySilurian (A. Sandford, pers. comm. 2001) isexcluded from this analysis, as this would appearto represent a range expansion which occurredafter the time period under consideration here.
The area cladogram in Figure 4 contains foursubclades containing areas that span much ofthe palaeogeographic extent of Gondwana(nodes 1-4 in Fig. 4). This is interpreted asindicating that several separate, independent
biogeographic events occurred during theevolution of the Reedocalymeninae, which canthus be compared with one another to reach anunderstanding of Gondwanan area relation-ships. Node 2 consists of an unresolved poly-tomy in the area cladogram, and so can provideno information on the relationships betweenthe four different areas (France/Spain, SouthAmerica, Avalonia and Turkey) included withinthis subclade. The other three nodes are eitherwell-resolved (nodes 1, 4) or adequatelyresolved (node 3). Nodes 1 and 4 display theeasternmost Gondwanan areas at the base ofthe subclade, with progressively more westernGondwanan areas tending to be nested progres-sively further within the subclade. At node 1,the far eastern Gondwanan region of SouthAmerica represents the sister group to CentralAsian and eastern Mediterranean regionsand South China; at node 4, the westernGondwanan regions of Avalonia, France, Spainand North Africa are more closely related toone another than any is to the more easterlysituated SE Asian regions of Indo-China andSouth China. Node 3 shows the opposite patternof area relationships, with South America andSouth China as sister areas, together sister toSpain, with the westernmost region of Avaloniaas the sister area to the rest of the subclade.
The ordering of successively nested areaswithin each of the resolved nodes matchesprevious ideas on the relative geographicpositions of Gondwanan and peri-Gondwanantectonic units in the Early Ordovician. Thesethree sets of area relationships can therefore beinterpreted as supporting the idea of a faunalcline mirroring the climatic cline acrossGondwana during this time interval. However,the three resolved subclades appear to representbiogeographic shifts in opposite directionsacross Gondwana. The opposing nested patternsof area relationships thus represent non-congruent biogeographic events, and containparalogous areas which would conflict withduplications of themselves to generate ambigu-ous data if combined as a consensus cladogram,reducing the information available from the areacladogram in Figure 3a. As it is imperative tolimit paralogy in area cladistics in order to detectbiogeographic congruence (Ebach 1999), com-bining all three into a single consensus areacladogram in an attempt to provide furtherinformation on Early Ordovician Gondwananarea relationships is thus to be avoided.
The subclades contained within nodes 1 and 4appear to represent faunal shifts in the samedirection across Gondwana (Fig. 5a, b). As thesetwo subclades share the area of SE Asia in

64 SAMUEL T.TURVEY
Fig. 5. (a, b) Area cladograms for nodes 1 and 4 of original area cladogram in Figure 4. with revised areaboundaries, (c) Consensus area cladogram of nodes 1 and 4, showing complete pattern of area relationships ofGondwanan and peri-Gondwanan regions.
common, they can be combined to produce aconsensus cladogram, which represents therelationships between different Gondwananareas indicated by the congruent westwardfaunal shifts within nodes 1 and 4 (Fig. 5c). Thisconsensus cladogram shows that the faunal clineextended across the entire region of Gondwanafor which Early Ordovician deposits are avail-able. South America and Avalonia+westernEurope+North Africa respectively represent theeastern and western limits of the cline, with noevidence for a faunal connection linking thesetwo areas directly across the Early Ordoviciansouthern polar region.
Conclusions and future research
The area cladograms generated from the cladis-tic analysis of the Reedocalymeninae supportcertain previously held ideas on Gondwananbiogeography. The presence of multiple sub-clades within the Reedocalymeninae, eachcontaining a wide range of both eastern andwestern Gondwanan areas, suggests that nosignificant environmental barriers existed acrossthe palaeocontinent even for shallow inner-shelfmarine taxa. The pattern of area relationshipsboth within these subclades and as evidenced bythe consensus cladogram in Figure 5c arealso consistent with the idea of a faunal clineoccurring across the palaeocontinent during the
Early Ordovician. No reedocalymenine taxahave been recorded from the Tremadoc, butsome Neseuretus species are known from thebase of the Arenig, suggesting that this patternof area relationships refers approximately to theinterval between these two Ordovician series.
This cladistic biogeographic analysis offers apreliminary hypothesis on the relationshipsbetween different Gondwanan and peri-Gond-wanan geographic areas for the Early Ordo-vician. However, the inter-relationships betweenthe various Asian areas included in the analysisare unresolved and require further study, andsome of the nodes used to generate the areacladograms in Figures 4 and 5 are only weaklysupported by bootstrap analysis. The hypothesisthus requires testing against area cladogramsgenerated with other Early Ordovician taxa.
Cladistic analysis of several other EarlyOrdovician trilobite groups could be used toassess support for this pattern of area relation-ships. Of the shallow-water taxa which some-times co-occur with Neseuretus in the NeseuretusAssociation, several genera (e.g. Kerfornella,Plaesiacomia, Eohomalonotus, Iberocoryphe,Crozonaspis and Taihungshania) are onlyknown from restricted regions of Gondwana,and so their phylogenies cannot be used togenerate comparable area cladograms. Othershallow-water trilobite groups do occur overmuch of the same geographic region as

TRILOBITE BIOGEOGRAPHY OF GONDWANA 65
Neseuretus during the Early Ordovician.Asaphids are a common component of theNeseuretus Association across the entire Gond-wanan region; however, the relationships withinthe family are currently poorly understood, andthorough taxonomic revision is required beforethey can be used to test the hypothesis of arearelationships generated with the Reedocaly-meninae.
Early Ordovician trilobite taxa potentiallyable to provide a test for this hypothesis includethe dikelokephalinid Hungioides, which rangesfrom western Europe to South America (Fortey& Peel 1983), and two different groups oftrinucleids which occur in slightly deeper shelfconditions. The closely related hanchun-golithine trinucleid genera Hanchungolithus andNingkianolithus, regarded by Li (1994) as occur-ring only in South China, have in fact also beenrecorded in Avalonia, France, the Middle East,Indo-China and possibly Spain (Dean 1966; El-Khayal & Romano 1985; Beckly 1989; Rabano1990), although South American materialoriginally assigned to Hanchungolithus (Hugheset al. 1975, as Ichangolithus} has recently beentransferred to the Trinucleinae (Baldis & Pothede Baldis 1995). A second trinucleid groupcomprises the genera Lordshillia, Anebolithus,Famatinolithus and Incaia, which appear to beclosely related to each other within the Trinu-cleinae, and occur in Avalonia, South China,New Zealand and South America (Hughes et al.1975). Future cladistic research on some of thesegroups can hopefully provide further infor-mation on Early Ordovician area relationshipsacross Gondwana and peri-Gondwana.
This project was greatly helped by the encouragementand assistance of D.J. Siveter, who engaged the authorwith considerable discussion on the evolution of theCalymenidae, and provided access to his substantialreference collection. Discussion with T.G. Watersgenerated further ideas on appropriate methods ofbiogeographic analysis. P. Janvier and Tong-DzuyThanh provided useful information on Vietnamia dou-villei, and casts of most of Mansuy's type material ofthis species were provided by P. Racheboeuf. TheEarly Ordovician map used in Figure 1 was generatedby D. Lees, and M.D. Sutton provided welcome tech-nical assistance. W.T. Dean, A.W. Owen and M.Romano reviewed the manuscript and made severaluseful suggestions for its improvement. Funding forthe project was provided by a Natural EnvironmentResearch Council D.Phil studentship.
ReferencesBALDIS, B. & POTHE DE BALDIS, E. D. 1995. Trilobites
Ordovicicos de la Formacion Las Aguaditas (SanJuan, Argentina) y consideraciones estratigrafi-
cas. Boletin de la Academia National de Ciencas,Corboda, 60,409-435.
BANKS, M. R. 1988. The base of the Silurian System inTasmania. In: COCKS, L. R. M. & RICKARDS, R. B.(eds) A global analysis of the Ordovician-Silurianboundary. Bulletin of the British Museum(Natural History), Geology, 43, 191-194.
BARRANDE, J. 1868. Silurische Fauna aus der Umge-bung von Hof in Bayern. Neues Jahrbuchfur Min-eralogie, 6/7, 641-696.
BECKLY, A. J. 1989. A new Arenig trilobite fauna fromthe Bangor area. North Wales. Bulletin of theBritish Museum (Natural History), Geology, 45,1-20.
BERGERON, J. 1890. Sur une forme nouvelle de trilo-bite de la famille des Calymenidae [genre Caly-menella]. Bulletin de la Societe Geologique deFrance, 18,365-371.
BORN, A. 1918. Die Calymene Tristani-Stufe (mittleresUnter-silur) bei Almaden, ihre Fauna,Gliederung und Verbreitung. Abhandlungen dersenckenbergischen naturforschenden Gesellschafi,36, 309-358.
CHEN Xu & QIU JUNYU. 1986. Ordovician palaeo-environmental reconstruction of Yichang area,W. Hubei. Journal of Stratigraphy, 10, 1-15.
CHEN XU, RONG JIAYU, WANG XIAOFENG, WANGZHIHAO, ZHANG YUANDONG & ZHANG RENBIN.1995. Correlation of the Ordovician rocks ofChina. International Union of GeologicalSciences, Publication 31.
COCKS, L. R. M. 2001. Ordovician and Silurian globalgeography. Journal of the Geological Society,London, 158,197-210.
COCKS, L. R. M. & FORTEY, R. A. 1988. Lower Palaeo-zoic facies and faunas around Gondwana. In:AlJDLEY-CHA]'RLES, M. G. & HALLAM, A. (eds)
Gondwana and Tethys. Geological Society,London, Special Publications, 37,183-200.
COCKS, L. R. M. & FORTEY, R. A. 1990. Biogeographyof Ordovician and Silurian faunas. In: Mc-KERROW, W. S. & SCOTESE, C. R. (eds) PalaeozoicPalaeogeography and Biogeography. GeologicalSociety, London, Memoirs, 12, 97-104.
CORBETT, K. D. & BANKS, M. R. 1974. Ordovicianstratigraphy of the Florentine Synelinorium,south-west Tasmania. Papers and Proceedings ofthe Royal Society of Tasmania, 55, 1-55.
COURTESSOLE, R., PlLLET, J. & VIZCAINO, D. 1981.Nouvelles donnees sur la biostratigraphie del'Ordovicien inferieur de la Montagne Noire.Revision des Taihungshaniidae, de Megistaspis(Ekeraspis) et d'Asaphopsoides (Trilobites).Memoire de la Societe des Etudes Scientifiques del'Aude. Carcassonne, France.
DEAN, W. T. 1966. The Lower Ordovician stratigra-phy and trilobites of the Landeyran valley andthe neighbouring district of the Montagne Noire,south-western France. Bulletin of the BritishMuseum (Natural History), Geology, 12,247-353.
DEAN, W. T. 1967a. The correlation and trilobite faunaof the Bedinan Formation (Ordovician) in south-eastern Turkey. Bulletin of the British Museum(Natural History), Geology, 15, 83-123.

66 SAMUEL T.TURVEY
DEAN, W. T. 1967b. The distribution of Ordovicianshelly faunas in the Tethyan region. In: ADAMS, C.G. & AGER, D. V. (eds) Aspects of Tethyan Bio-geography. Systematics Association, Publication7.11-44.
DEAN, W. T. 1971. The Lower Palaeozoic stratigraphyand faunas of the Taurus Mountains nearBeysehir, Turkey. II. The trilobites of theSeydisehir Formation (Ordovician). Bulletin ofthe British Museum (Natural History), Geology.20. 3-24.
DEAN, W. T. 1973. The Lower Palaeozoic stratigraphyand faunas of the Taurus Mountains nearBeysehir, Turkey. III. The trilobites of the SobovaFormation (Lower Ordovician). Bulletin of theBritish Museum (Natural History), Geology. 24,279-348.
DEAN, W. T. 1975. Cambrian and Ordovician corre-lation and trilobite distribution in Turkey. Fossilsand Strata. 4, 353-373.
DEAN. W. T. & MARTIN, F. 1978. Lower Ordovicianacritarchs and trilobites from Bell Island, easternNewfoundland. Geological Survey of CanadaBulletin. 284,1-35.
DEAN, W. T. & ZHOU ZHIYI. 1988. Upper Ordoviciantrilobites from the Zap Valley, south-east Turkey.Palaeontology. 31, 621-649.
DEAN. W. T, MONOD, O., RICKARDS, R. B., OSMANDEMIR & BULTYNCK, P. 2000, Lower Palaeozoicstratigraphy and palaeontology, Karadere-Zirzearea, Pontus Mountains, northern Turkey. Geo-logical Magazine. 137, 555-582.
DESMAREST, A. G. 1817. Crustaces Fossiles. In: BIOT,J. B. et al. (eds) Nouveau Dictionnaire d'HistoireNaturelle. 8 [Cor-Cun]. Deterville, Paris,495-519.
EBACH, M. C. 1999. Paralogy and the centre of originconcept. Cladistics. 15, 387-391.
EDGECOMBE, G. D., BANKS, M. R. & BANKS, D. M. 1999.Upper Ordovician Phacopida (Trilobita) fromTasmania. Alcheringa, 23, 235-257.
EL-KHAYAL, A. A. & ROMANO, M. 1985. LowerOrdovician trilobites from the Hanadir Shale ofSaudi Arabia. Palaeontology. 28, 401-412.
FELSENSTEIN, J. 1985. Confidence limits on phyloge-nies: an approach using the bootstrap. Evolution.39.783-791.
FORTEY, R. A. 1983. Cambrian-Ordovician trilobitesfrom the boundary beds in western Newfound-land and their phylogenetic significance. SpecialPapers in Palaeontology. 30, 179-211.
FORTEY. R. A. 1990. Ontogeny, hypostome attach-ment and trilobite classification. Palaeontologv,33, 529-576.
FORTEY, R. A. & MELLISH, C. J. T. 1992. Aresome fossils better than others for inferringpalaeogeography? The early Ordovician of theNorth Atlantic as an example. Terra Nova. 4.210-216.
FORTEY; R. A. & MORRIS. S. F. 1982. The Ordoviciantrilobite Neseuretus from Saudi Arabia, and thepalaeogeography of the Neseuretus fauna relatedto Gondwanaland in the earlier Ordovician.Bulletin of the British Museum (Natural History),Geology. 36, 63-75.
FORTEY, R. A. & OWENS. R. M. 1978. Early Ordo-vician Arenig) stratigraphy and faunas of theCarmarthen district, south-west Wales. Bulletin ofthe British Museum (Natural History), Geology.30. 225-294.
FORTEY, R. A. & OWENS. R. M. 1987. The ArenigSeries in South Wales. Bulletin of the BritishMuseum (Natural History), Geology. 41. 69-307.
FORTEY. R. A. & PEEL. J. S. 1983. The anomalousbathyurid trilobite Ceratopeltis and its homeo-morphs. Special Papers in Palaeontology. 30.51-57.
FORTEY. R. A.. H A R P E R , D. A. T, I N G H A M . J. K..OWEN. A. W. PARKES. M. A., RUSHTON. A. W. A.& WOODCOCK, N. H. 2000. A revised correlationof Ordovician rocks in the British Isles. Geo-logical Society Special Report. 24. 1-83.
GIGOUT. M. 1951. Etudes geologiques sur la MesetaMarocaine Occidentale (arriere Pays deCasablanca. Mazagan et Safi). Travaux de I'Insti-tut scientifique cherifien. 3. 1-507.
H A M M A N N . W. 1977. Neue Calymenacea (Trilobita)aus dem Ordovizium von Spanien. Senckenber-giana Lethaea. 58. 91-97.
HAMMANN. W. 1983. Calymenacea (Trilobita) aus demOrdovizium von Spanien: ihre Biostratigraphie.Okologie und Systematik. Abhandlungen dersenckenbergischen naturforschenden Gesellschaft.542.1-177.
HAMMANN. W. & LEONE, F. 1997. Trilobites of thepost-Sardic (Upper Ordovician) sequence ofsouthern Sardinia. Part I. Beringeria. 20. 1-217.
HENRY. J-L. 1989. Paleoenvironnements etdynamique de faunes de trilobites dansl'Ordovicien (Llanvirn superieur - Caradocbasal) du massif Armoricain (France). Palaeo-geographv, Palaeoclimatology, Palaeoecologv.73.139-153.
HICKS. H. 1873. On the Tremadoc rocks in the neigh-bourhood of St. David's. Pembrokeshire, withspecial reference to those of the Arenig and Llan-deilo groups and their fossil contents. QuarterlyJournal of the Geological Society of London. 29.39-52.
HUGHES. C. P., I N G H A M , J. K. & ADDISON. R. 1975. Themorphology, classification and evolution of theTrinucleidae (Trilobita). Philosophical Trans-actions of the Royal Society of London. B. Bio-logical Sciences. 272(920), 537-607.
KITCHEN. I. J., FOREY, P. L., HUMPHRIES. C. J. &WILLIAMS. D. M. 1998. Cladistics. The Theory andPractice of Parsimony Analysis (second edition).Oxford University Press. Oxford.
KOBAYASHI. T. 1937. The Cambro-Ordovician shellyfaunas of South America. Journal of the Faculty ofScience, Imperial University of Tokyo, Section II(Geology, Mineralogy, Geography, Seismology).4,369-522.
KOBAYASHI. T. 1976. On the Ordovician TrilobiteProvinces. Proceedings of the Japanese Academy.52, 232-235.
KOBAYASHI. T. 1987. The Cambrian and Ordoviciansystems in the south side of Eurasia and the originof the Tethys Sea. Journal of Geography. 96.39-368.

TRILOBITE BIOGEOGRAPHY OF GONDWANA 67
LEGG, D. P. 1976. Ordovician trilobites and graptolitesfrom the Canning Basin, western Australia. Geo-logica et Palaeontologica, 10, 1-58.
LEONE, F., HAMMANN, W., LASKE, R., SERPAGLI, E.& VILLAS, E. 1991. Lithostratigraphic unitsand biostratigraphy of the post-sardic Ordovi-cian sequence in south-west Sardinia. Bollet-tino della Societa Paleontologica Italiana, 30,201-235.
Li ZHIMING. 1994. Ordovician. In: YIN HONGFU (ed.)The Palaeobiogeography of China. ClarendonPress, Oxford, 64-87.
Lu YENHAO. 1957. Trilobita. In: Index fossils of China.Invertebrates (3). Science Press, Beijing (inChinese).
Lu YENHAO. 1975. Ordovician trilobite faunas ofcentral and southwestern China. PalaeontologicaSinica, New Series B, 11,1-463.
MANSUY, H. 1908. Contribution a la Carte Geologiquede 1'Indo-Chine. Paleontologie. GouvernementGeneral de ITndo-Chine, Direction Generale desTravaux Publics, Service des Mines. Imprimeried'Extreme-Orient, Hanoi-Haiphong.
MOORE, R.C. (ed.) 1959. Treatise on InvertebratePaleontology. Part O. Arthropoda 1. University ofKansas Press and Geological Society of America,Lawrence, Kansas.
MORRIS, S. F. & FORTEY, R. A. 1985. Catalogue ofthe Type and Figured Specimens of Trilobita in theBritish Museum (Natural History). BritishMuseum (Natural History), London.
Mu ENZHI. 1974. Evolution, classification and distri-bution of Graptoloidea and graptodendroids.Scientia Sinica, 17, 227-238.
PALMER, A. R. 1972. Problems of Cambrian biogeog-raphy. 24th International Geological Congress, 7,310-315.
PARIS, F. 1998. Early Palaeozoic palaeobiogeographyof northern Gondwana regions. Acta UniversitatisCarolinae - Geologica, 42, 473-483.
PEERS, S. 1997. The taxonomy, phylogeny and palaeo-biogeography of the trilobite families Pliomeridaeand Encrinuridae - reconstructing the Ordovicianworld using evolving lineages. PhD thesis,Glasgow University.
PENG SHANCHI ,LINTIANRUI & LI YUE. 2000. Notes onthe genus Reedocalymene Kobayashi, 1951 (Trilo-bita, Ordovician). Acta Palaeontologica Sinica,39, 63-75.
PILLET, J. & DE LAPPARENT, F. 1969. Description destrilobites ordoviciens, siluriens et devoniensd'Afghanistan. Annales de la Societe geologiquedu Nord, 84, 323-333.
PRIBYL, A. & VANEK, J. 1980. Ordovician trilobites ofBolivia. Rozpravy Ceskoslovenske AkademieVed, 90,1-90.
RABANO, I. 1990. Trilobites del Ordovicico Medio delsector meridional de la zona Centroibericaespanola. Publicaciones especiales del boletingeologico y minero. Instituto TecnologicoGeominero de Espana, Madrid.
REED, F. R. C. 1917. Ordovician and Silurian fossilsfrom Yun-Nan. Memoirs of the Geological Surveyof India, Palaeontologica Indica, New Series, 6,1-69.
Ross, R. J. Jr. 1967. Some Middle Ordovician brachio-pods and trilobites from the Basin Ranges,western United States. Geological Survey Pro-fessional Paper, 523-D, 1-43.
Ross, R. J. Jr. 1975. Early Paleozoic trilobites, sedi-mentary facies, lithospheric plates, and ocean cur-rents. Fossils and Strata, 4, 307-329.
S ALTER, J. W. 1865a. A Monograph of the BritishTrilobites from the Cambrian, Silurian andDevonian Formations. PalaeontographicalSociety Monograph, 2, 81-128.
SALTER, J. W. 18656. In: SALTER, J. W. &BLANDFORD, H. F. Palaeontology of Niti in theNorthern Himalaya: being descriptions and figuresof the Palaeozoic and Secondary fossils collectedby Colonel Richard Strachey, R. E. Calcutta.
SCOTESE, C. R. & McKERROW, W. S. 1990. Revisedworld maps and introduction. In: McKERROW, W,S. & SCOTESE, C. R. (eds) Palaeozoic Palaeo-geography and Biogeography. Geological SocietyLondon, Memoirs 12,1-21.
SDZUY, K. 1955. Die Fauna der Leimitz-Schiefer(Tremadoc). Abhandlungen der senckenbergis-chen naturforschenden Gesellschaft, 492, 1-74.
SEDGWICK, A. & M'CoY, F. 1851. A synopsis of theclassification of the British Palaeozoic rocks, witha systematic description of the British Palaeozoicfossils in the geological museum of the Universityof Cambridge. Pt. 1. Cambridge.
SENGOR, A. M. C. 1984. The Cimmeride orogenicsystem and the tectonics of Eurasia. GeologicalSociety of America, Special Paper, 195.
SENGOR, A. M. C. 1987. Tectonics of the Tethysides:orogenic collage development in a collisionalsetting. Annual Reviews of Earth and PlanetaryScience, 15,213-244.
SHENG, S. F. 1958. The Ordovician trilobites of south-western China. Acta Palaeontologica Sinica, 6,169-204.
SHIRLEY, J. 1936. Some British trilobites of the familyCalymenidae. Quarterly Journal of the GeologicalSociety of London, 92, 384-422.
SOKOLOV, B. S. & YOLKIN, E. A. (eds). 1978. Pogranich-nye sloi ordovika i silura Altye-Sayanskoy oblastii Tyen-Shanya. Transactions of the Academy ofSciences of the USSR, Siberian Branch, Institute ofGeology and Geophysics, 397, 1-221.
SPJEDNAES, N. 1961. Ordovician climatic zones. NorskGeologisk Tidsskrift, 41, 45-77.
SPJEDNAES, N. 1981. Lower Palaeozoic palaeoclima-tology. In: HOLLAND, C. H. (ed.) Lower PalaeozoicRocks of the World. Volume 3. Lower Palaeozoic ofthe Middle East, Eastern and Southern Africa, andAntarctica. John Wiley and Sons, Chichester,199-256.
STUBBLEFIELD, C. J. 1939. Some aspects of the distri-bution and migration of trilobites in the BritishLower Palaeozoic faunas. Geological Magazine,76, 49-72.
SUN, Y. C. 1931. Ordovician trilobites of central andsouthern China. Palaeontologica Sinica, Series B,7, 1-47.
SWOFFORD, D. L. 2001. PAUP*: Phylogenetic AnalysisUsing Parsimony (*and other methods). Version4.0b4a. Sinauer Associates, Sunderland, MA.

68 SAMUEL T.TURVEY
TRIPP, R. P., ZHOU ZHIYI & PAN ZHENQIN. 1989. Trilo-bites from the Upper Ordovician Tangtou For-mation, Jiangsu Province. China. Transactions ofthe Royal Society of Edinburgh: Earth Sciences.80. 25-68.
VACCARI. N. E. & WAISFELD. B. G. 1994. Nuevos trilo-bites de la Formacion Suri (Ordovicico temprano)en la region de Chaschuil, Provincia de Cata-marca. implicancias bioestratigraficas. Ameghini-ana. 31, 73-86.
VAN INGEN. G. 1914. Cambrian and Ordovician faunasof south-eastern Newfoundland (Abstract).Bulletin of the Geological Society of America. 25.138.
WAISFELD. B. G. 1997. Trilobites calymenaceos de laFormacion Acoite (Arenigiano) en el contra-fuerte occidental de la Cordillera OrientalArgentina. Ameghiniana. 34. 333-343.
WANG XIAOFENG, XIANG LIWEN. NI SHIZHAO, ET AL.1987. Biostratigraphy of the Yangtze Gorges Area(2). Early Palaeozoic Era. Geological PublishingHouse. Beijing.
WHITTARD. W. F. 1960. The Ordovician trilo-bites of the Shelve Inlier. west Shropshire.Palaeontographical Society Monograph. 4,117-162.
WHITTINGTON, H. B. 1965. Trilobites of the Ordo-vician Table Head Formation, western New-foundland. Bulletin of the Museum ofComparative Zoology, Harvard University. 132.275-442.
WHITTINGTON. H. B. 1966a. Trilobites of the HenllanAsh. Arenig Series. Merioneth. Bulletin of theBritish Museum (Natural History), Geology. 11.489-505.
WHITTINGTON. H. B. 1966b Phylogeny and distri-bution of Ordovician trilobites. Journal of Pale-ontology. 40.696-737.
WHITTINGTON. H. B. 1973. Ordovician trilobites. In:H A L I . A M , A. (ed.) Atlas of Palaeobiogeography.Elsevier Science Publications. Oxford. 13-18.
WHITTINGTON. H. B. & HUGHES. C. P. 1972. Ordo-vician geography and faunal provinces deducedfrom trilobite distribution. Philosophical Trans-actions of the Royal Society of London. B. Bio-logical Sciences. 263(850). 235-278.
YI YONGEN. 1957. The Caradocian trilobite fauna fromthe Yangtze-Gorges. Acta Palaeontologica Sinica.5. 527-559.
YIN GONGZHENG & LEE SHANJI . 1978. Trilobita. In:Atlas of Palaeontology of Southwest China,Guizhou (1). Geological Publishing House.Beijing (in Chinese).
ZHOU Z H I Y I & DEAN. W. T. 1989. Trilobite evidencefor Gondwanaland in east Asia during theOrdovician. Journal of Southeast Asian EarthSciences. 3. 131-140.
ZHOU Z H I Y I . DEAN. W.T.&LUO HUILN. 1998a. EarlyOrdovician trilobites from Dali. west Yunnan.China, and their palaeogeographical significance.Palaeontology. 41. 429-460.
ZHOU Z H I Y I . DEAN. W. T. YUAN WENWEI & ZHOUTIANRONG. 1998b. Ordovician trilobites from theDawangou Formation. Kalpin. Xinjiang, north-west China. Palaeontology. 41. 693-735.
ZHOU Z H I Y I , LUO HUILIN. ZHOU Z H I O I A N G & YUANW E N W E I . 2001. Palaeontological constrains on theextent of the Ordovician Indo-China Terrane inwestern Yunnan. Acta Palaeontologica Sinica. 40.310-317.

The spatial and temporal diversification of Early Palaeozoicvertebrates
M. PAUL SMITH, PHILIP C. J. DONOGHUE & IVAN J. SANSOMSchool of Earth Sciences, University of Birmingham, Edgbaston, BirminghamB15 2TT, UK (e-mail: [email protected]; [email protected];
Abstract: Recent discoveries have dramatically altered traditional views of the stratigraphicdistribution and phylogeny of Early Palaeozoic vertebrates and permit a reappraisal ofbiogeographic patterns and processes over the first 120 million years of vertebrateevolution. Stratigraphic calibration of the phylogenetic trees indicates that most of thepre-Silurian record can be inferred only through ghost ranges. Assessment of the availabledata suggests that this is due to a shift in ecological niches after the latest Ordovicianextinction event and a broadening of geographical range following the amalgamation ofEuramerica during the early Silurian, Two major patterns are apparent in the bio-geographic data. Firstly, the majority of jawless fishes with dermoskeletal, plated 'armour'were highly endemic during Cambrian-Ordovician time, with arandaspids restricted toGondwana, galeaspids to China, and anatolepids, astraspids and, possibly, heterostracansconfined to Laurentia. These Laurentian groups began to disperse to other continentalblocks as the 'Old Red Sandstone continent' amalgamated through a series of tectoniccollisions. The second major pattern, in contrast, encompasses a number of microsquamousand naked, jawed and jawless primitive vertebrates such as conodonts, thelodonts, placo-derms, chondrichthyans and acanthodians, which dispersed rapidly and crossed oceanicbarriers to attain cosmopolitan distributions, although many have Laurentian origins. Aclear difference in dispersal potential exists between these two types of fishes. Overall, thedevelopment of biogeographic patterns in Early Palaeozoic vertebrates involved a complexinteraction of dispersal, vicariance and tectonic convergence.
Elliott et al (1991) reviewed pre-Silurian verte- Arandaspis and Sacabambaspis were reliablybrates, concluding that only six Ordovician understood in terms of their anatomy and, ingenera could be recognized unequivocally, and phylogenetic terms, Elliott et al. (1991) con-that none of the reports of Cambrian taxa were sidered Astraspis to be the most derived, withsustainable on available evidence. The oldest Arandaspis + Sacabambaspis as a sister taxon.vertebrates were considered to be Arandaspis The biogeography of Astraspis, Arandaspisand the poorly known taxon Porophoraspis from and Sacabambaspis was examined by Elliott etthe early Llanvirn Stairway Sandstone of central al (1991) in the light of both dispersal andAustralia (Ritchie & Gilbert-Tomlinson 1977). vicariance models. Their dispersal model inter-In addition, Sacabambaspis had been described preted the spatial and temporal data ratherfrom the Caradoc of Bolivia by Gagnier et al. literally in biogeographic terms, invoking a(1986) and was considered to be closely related dispersal route from the older, Australian,to Arandaspis (Gagnier et al. 1986; Elliott et al. locality across Gondwana to Bolivia and thence1991). The most diverse vertebrate fauna was across an oceanic barrier to Laurentia, amount-thought to be that from the Harding Sandstone ing to what might be termed an 'out of(Caradoc) of Colorado, USA, which had been Gondwana' model. It was recognized that thethe first locality to yield pre-Silurian vertebrates, final trans-oceanic migration was an obstacle toWalcott (1892) described three species of fishes a dispersal model given the apparent shallow-from the Harding Sandstone: Astraspis desider- water ecological specialism of these fishes, but itata, Eriptychius americanus and Dictyorhabdus was also noted that a vicariant model was notpriscus. The last is not now considered to be a supported, and that both were in conflictvertebrate, although its affinities remain obscure with the palaeocontinental reconstructions of(Sansom et al. 2001). A third vertebrate taxon Scotese (1986) and Scotese&McKerrow (1990).was, however, known from the unit (Denison Since the review of Elliott et al. (1991), there1967; Smith 1991) and was subsequently has been a dramatic increase in the quality of thedescribed as Skiichthys halsteadi by Smith & fossil record of Early Palaeozoic vertebratesSansom (1997). Of these taxa, only Astraspis, which has arisen both through the discovery of
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic—Cenozoic Radiations. Geological Society, London, Special Publications, 194, 69-83.0305-8719/02/$15.00 © The Geological Society of London 2002.

70 M. P. SMITH, P. C. J. DONOGHUE & I. J. SANSOM
new faunas and from reappraisals of affinity inwell-known faunas. These advances haveincluded the discovery of probable EarlyCambrian vertebrates in the Chengjiang Lager-statte (Shu et al 1999a), the recognition ofpossible vertebrates in the Middle CambrianBurgess Shale Lagerstatte of British Columbia(Simonetta & Insom 1993; Smith et al. 2001). theconfirmation of the Late Cambrian-EarlyOrdovician sclerite Anatolepis as a vertebrate(Smith et al. 1996, 2001) and the discovery ofunanticipated diversity in the Harding Sand-stone of Colorado (Smith et al. 1995; Sansom etal. 1995, 1996, 2001). Perhaps the most signifi-cant discovery in terms of the biodiversity andgeographical range of early vertebrates has beenthe addition of conodonts to the clade. Thepresence of preserved soft tissues had alreadysuggested to Aldridge et al. (1986) that theaffinities of this highly diverse group lay with thevertebrates, and a wealth of new data has nowbeen advanced in support of the hypothesis.These include additional soft tissue finds (Smithet al. 1987; Aldridge et al. 1993; Gabbott et al1995), the recognition of vertebrate hard tissuesynapomorphies in conodont elements (Sansometal 1992, 1994; Sansom 1996; Donoghue 1998;Donoghue & Aldridge 2001) and detailedcladistic analysis (Donoghue et al. 2000). Theinclusion of conodonts as the earliest vertebratesto possess a mineralized skeleton, more derivedthan either hagfishes or lampreys, increases thegeneric and specific biodiversity of Cambro-Ordovician vertebrates by two orders ofmagnitude.
Does the substantial amount of new dataacquired subsequent to the review of Elliottet al. (1991) permit more detailed andbetter supported models of Early Palaeozoicvertebrate biogeography?
Early Palaeozoic palaeogeographyThe determination of biogeographic trends inEarly Palaeozoic vertebrates is highly depen-dent on the accuracy, and choice, of palaeo-continental reconstructions. For Cambrian-Ordovician vertebrates, this is particularly trueof the relative positions of Laurentia, Australiaand South America, whereas constraints on thetiming of collisions that assembled the 'OldRed Sandstone (ORS) continent' are criticalfor interpretations of Silurian-Devonian bio-geography. To avoid circularity, we have usedpalaeocontinental reconstructions constructedfrom palaeomagnetic and tectonic data, andhave eschewed those that already incorporatefaunal data (e.g. Scotese & McKerrow 1990).
Fig. 1. Palaeogeographic reconstructions, (a) Base ofCambrian: (b) mid-Ordovician; (c) Wenlock. BasedonDalziel(1997).

EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 71
Traditionally, the Iapetan margin of southernLaurentia has been reconstructed as the conju-gate margin of NW Africa prior to the openingof Iapetus, following the early reconstructions ofWilson (1966) and an implicit acceptance of an'accordian style' opening and closure model(Dalziel 1997). However, the evidence for pre-lapetan juxtaposition of the Laurentian andWest African cratons is not strong and evidencehas accumulated in favour of a radical alterna-tive which arose out of the 'SWEAT' hypothesisof Moores (1991). Independently, Dalziel (1991)and Hoffman (1991) suggested that, prior to theopening of Iapetus and the final break-up ofthe Neoproterozoic supercontinent Rodinia,Laurentia was located between East Antarctica-Australia (East Gondwana) and South America(West Gondwana) (Fig. la). Following rifting,Laurentia was considered to have followed aclockwise rotating path around the conjoinedSouth American and West African cratons(Dalziel 1991,1997). Strong supporting evidencecomes from the similarities between the fauna,stratigraphy and geological structure of SWUSA and the Argentinean Precordillera (DallaSalda et al. 1992; Astini et al 1995; Thomas &Astini 1996) and there is now general agreementthat the latter terrane originated as part of theLaurentian craton, located within the Ouachitaembayment. Opinion differs, however, on theway in which the terrane was decoupled fromLaurentia and attached to South America.Dalziel (1997) suggested that it originated as apromontory of Falkland-Malvinas Plateau typewhereas Astini et al. (1995) favoured a riftedmicrocontinent that detached from Laurentia,drifted across Iapetus and collided with theSouth American margin. This has somerelevance to early vertebrate biogeography, asdiscussed below, but in both cases the conju-gate nature of the SE Laurentian and SouthAmerican margins is implicit. The reconstruc-tions of Dalziel (1997) are used as the basis forour assessments of Early Palaeozoic palaeo-biogeography (Fig. la-c), although we recognizethat not all details of these are universallyaccepted.
With regard to the assembly of the ORScontinent, evidence has mounted to the pointwhere a reasonably well-constrained andconsistent model is available. The earliestcollisions are of island arcs, with successiveterrane accretion events (see van Staal et al.(1998) and Armstrong & Owen (2001) forreviews) serving as a prelude to the assembly ofAvalonia, Baltica and Laurentia in mid-Siluriantime. On the NE margin of Laurentia, earlyarc collisions are absent, and the first major
assembly event was the collision with Baltica(Elevold et al. 2000). On the basis of palaeo-magnetic data and kinematic indicators, it haslong been recognized that this collision wasoblique to the margins, with a sinistral com-ponent. Despite this obliquity, the collision isremarkably synchronous along the entire 2000km length of this sector of the Caledonides. InNW Scotland, mylonites in the Moine thrustzone were dated by Freeman et al. (1998) as437-430 Ma (Llandovery) and brittle defor-mation extends from 430 to 408 Ma (latest Llan-dovery-earliest Devonian). In comparison,prograde metamorphism in East Greenland hasbeen dated at 435-423 Ma with anatexis at430-422 Ma (Elvevold et al. 2000; Hartz et al.2001) and in eastern North Greenland, latethrusts truncate earliest Wenlock (425 Ma) fore-land basin turbidites (Higgins et al. 2001).
The closure of the Tornquist Sea, betweenAvalonia and Baltica, probably began in thelatest Ordovician by dextral 'soft' amalgamationof the two plates (McKerrow et al. 1991, 2000:Torsvik et al. 1996). Palaeomagnetic dataconfirm that the sector of Iapetus betweenAvalonia and Laurentia was also closed, withinthe limits of palaeomagnetic resolution, by theWenlock (Mac Niocaill 2000). The location ofSiberia during this interval is more open toquestion. There is good evidence that Siberiawas geographically inverted with respect tomodern coordinates during the Early Palaeozoic(Torsvik et al. 1995, 1996; Smethurst et al. 1998)and drifted northwards, apparently in tandemwith Euramerica ( = Laurentia + Baltica -Avalonia) (Smethurst et al. 1998). Rotation ofSiberia and collision with Baltica occurredbetween 360 Ma (earliest Carboniferous) and250 Ma (end-Permian) (Smethurst et al. 1998),and McKerrow et al (1991) interpreted Siberiaas having an almost juxtaposed relationship withEuramerica during Silurian-Early Devoniantime. There is no evidence for collision but theremay not have been a significant oceanic barrierseparating Novaya Zemlya and the northernSiberian margin. Cocks (2001, p. 199) noted thatshelly taxa were mainly endemic at species level,with some endemic genera, indicating that thepalaeocontinent was 'not particularly isolatedduring much of the period'. An additional, andkey, component of Siberia from the standpointof vertebrate biogeography is the terrane ofTuva (Young 1991, 1993), which had amalga-mated with Siberia by the early Silurian(Bachtadse et al. 2000) but does contain lateSilurian and Devonian endemic vertebrates(Afanassieva & Janvier 1985; Young 1993;Janvier 1996a).

72 M. P. SMITH. P. C. J. DONOGHUE & I. J. SANSOM
The stratigraphic and biogeographicrecord
Cambrian vertebrates
Although molecular clock estimates imply anextensive pre-Cambrian history to the vertebrateclade (Kumar & Hedges 1998), at present thereare no palaeontological data to support thisconclusion. The Cambrian record begins with theChengjiang Lagerstatte (basal Atdabanian, 530Ma) which has yielded a number of taxa thathave been interpreted as vertebrates. Shu et al(1999a) described two unmineralized vertebratetaxa, Haikouichthys and Myllokunmingia, fromthe deposit, which they considered to be closelyrelated to lampreys. Other Chengjiang taxahave also been interpreted as vertebrates.Yunnanozoon and Haikouella were consideredto be stem group vertebrates by Chen et al.(1999) and Holland & Chen (2001), but this is notwell-supported by the preserved anatomicalevidence. Xidazoon was likened to Pipiscius, ajawless vertebrate from the CarboniferousMazon Creek Lagerstatte of Illinois, USA, byShu et al. (1999b), but newly discovered materialof the latter indicates that it is not a vertebrate(Conway Morris, pers. comm. 2000).
A more equivocal vertebrate record is presentin the Burgess Shale Lagerstatte of BritishColumbia, Canada. Simonetta & Insom (1993)described an unnamed taxon (Smithsonian Insti-tution specimen USNM 198612) and likened thepreserved structures in the head region to thecranial cartilages and sensory organ supports ofextant lampreys. Testing of this intriguingsuggestion probably awaits the discovery ofadditional material.
The earliest evidence of biomineralizingvertebrates is present in the mid-Late Cambrianwhere the phosphatic sclerite Anatolepis (Bock-elie & Fortey 1976; Repetski 1978; Smith et al.1996) and euconodonts have an almost simul-taneous first occurrence (Smith et al. 2001). Thevertebrate character of earliest euconodont hardtissues has been confirmed in a number ofstudies (e.g. Sansom et al. 1992; Donoghue1998). If a phylogenetic link with paracon-odonts, initially proposed by Bengtson (1976), isdemonstrated, then the origin ofbiomineralizing vertebrates would be extendedback to the Middle Cambrian. Euconodonts arecosmopolitan and their first appearance isglobally synchronous, at least within the limitsof biostratigraphic resolution. In contrast,Anatolepis is entirely confined to the margins ofLaurentia. The affinities of Anatolepis havebeen the subject of some debate, but the
presence of dentine indicates that it is avertebrate (Smith et al. 1996) whilst the presenceof dermoskeletal 'armour' (rather thanvisceroskeletal 'teeth') suggests that it is morederived than conodonts. For ease of reference,groups of jawless fish more derived thanconodonts are referred to in this paper as'ostracoderms', a paraphyletic but nonethelessuseful term in this context.
Ordovician vertebrates
In contrast to the six genera, and seven species,recognized by Elliott et al. (1991). the total bio-diversity of Ordovician vertebrates is nowknown to number several hundred species, ofwhich the vast majority are conodonts. However,the recognized diversity of non-conodont verte-brates has also increased significantly. Analysisof the microvertebrate fauna of the HardingSandstone of Colorado and a number of roughlycoeval clastic units in North America hasrevealed that, in contrast to the three taxarecognized in earlier studies, at least 20 speciesare present. More remarkably, this fauna con-tains a large number of first appearances ofmajor clades, including astraspids. thelodontsand chondrichthyans (Sansom et al. 2001). Theage of these units is thus critical to accurateconstraint of the ghost ranges of vertebrate taxa.Conodonts are present in all units examined andindicate that the Harding Sandstone is ofundatus Chronozone age in the graphicallycorrelated stratigraphic framework of Sweet(1984), and that related units with similarvertebrate faunas span the compressa-undatusChronozone interval (mid-Mohawkian, mid-Caradoc). The chondrichthyan fauna includesmongolepids, which are here considered to bestem-chondrichthyans based on the sharedpresence of neck canals in the scales, but theabsence of other chondrichthyan synapomor-phies. In addition to Laurentia, thelodonts arealso found in Upper Ordovician localities inBaltica (Timan-Pechora; Karatajute-Talimaa1997) and peri-Gondwanan terranes (Austria:Cocks 2000; Bogolepova pers. comm.).
The application of micropalaeontologicalmethodologies in the Amadeus Basin of centralAustralia has also revealed more diverse faunasthan previously suspected. Young (1997)described microvertebrate faunas ranging in agefrom early Arenig to early Caradoc. Some of thesclerites are of doubtful affinity, but Apedolepis(Llanvirn) is a probable vertebrate (Young 1997;Sansom et al. 2001) and the range-base ofPorophoraspis has been extended from theLlanvirn to the lower Arenig. Pirchanchaspis
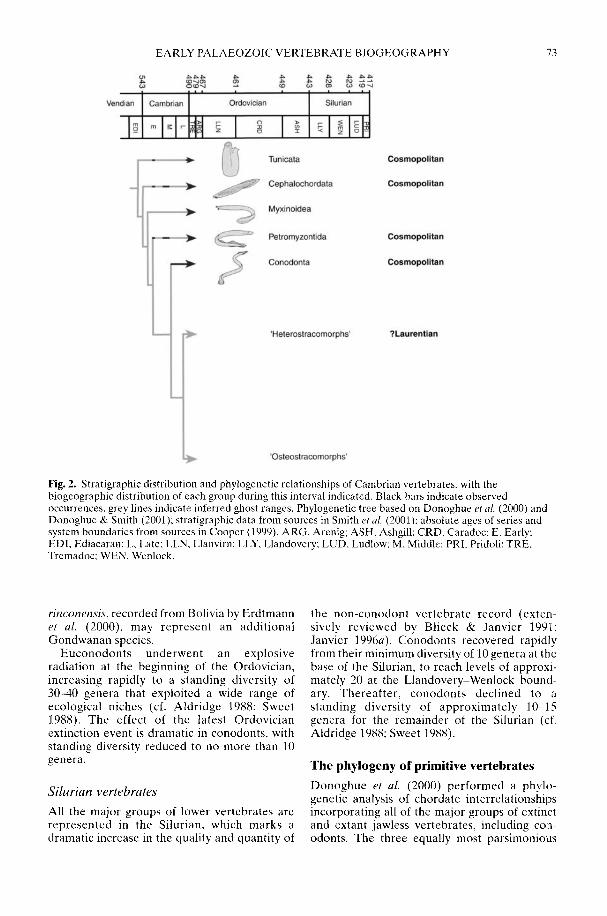
EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 73
Fig. 2. Stratigraphic distribution and phylogenetic relationships of Cambrian vertebrates, with thebiogeographic distribution of each group during this interval indicated. Black bars indicate observedoccurrences, grey lines indicate inferred ghost ranges. Phylogenetic tree based on Donoghue et al. (2000) andDonoghue & Smith (2001); Stratigraphic data from sources in Smith et al. (2001); absolute ages of series andsystem boundaries from sources in Cooper (1999). ARG, Arenig; ASH, Ashgill; CRD, Caradoc; E, Early;EDI, Ediacaran; L, Late; LLN, Llanvirn; LLY, Llandovery; LUD, Ludlow; M, Middle; PRI. Pridoli; TRE,Tremadoc; WEN, Wenlock.
rinconensis, recorded from Bolivia by Erdtmannet al (2000), may represent an additionalGondwanan species.
Euconodonts underwent an explosiveradiation at the beginning of the Ordovician,increasing rapidly to a standing diversity of30-40 genera that exploited a wide range ofecological niches (cf. Aldridge 1988; Sweet1988). The effect of the latest Ordovicianextinction event is dramatic in conodonts, withstanding diversity reduced to no more than 10genera.
Silurian vertebrates
All the major groups of lower vertebrates arerepresented in the Silurian, which marks adramatic increase in the quality and quantity of
the non-conodont vertebrate record (exten-sively reviewed by Blieck & Janvier 1991;Janvier I996a). Conodonts recovered rapidlyfrom their minimum diversity of 10 genera at thebase of the Silurian, to reach levels of approxi-mately 20 at the Llandovery-Wenlock bound-ary. Thereafter, conodonts declined to astanding diversity of approximately 10-15genera for the remainder of the Silurian (cf.Aldridge 1988; Sweet 1988).
The phylogeny of primitive vertebrates
Donoghue et al. (2000) performed a phylo-genetic analysis of chordate interrelationshipsincorporating all of the major groups of extinctand extant jawless vertebrates, including con-odonts. The three equally most parsimonious
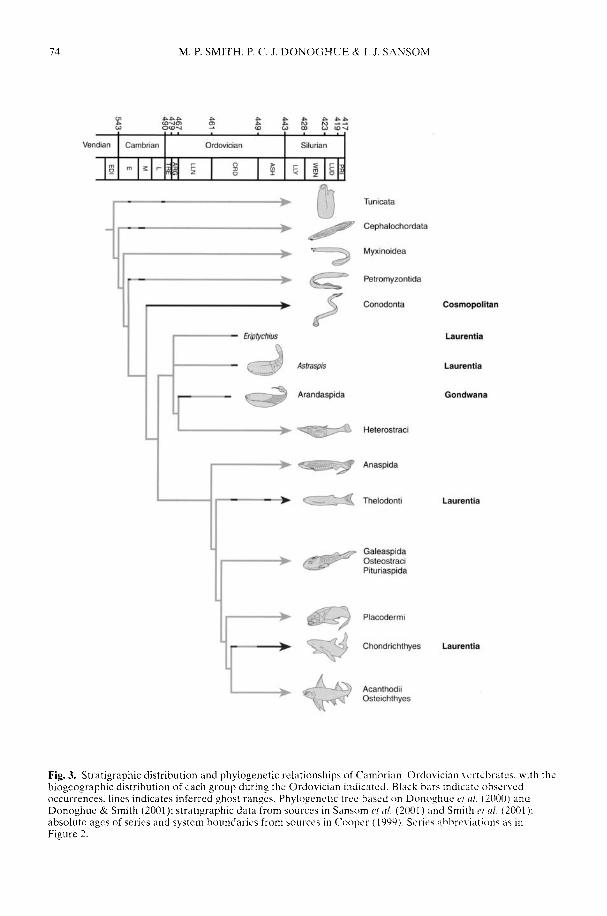
74 M. P. SMITH. P. C. J. DONOGHUE & I. J. SANSOM
Fig. 3. Stratigraphic distribution and phylogenetic relationships of Cambrian-Ordovician vertebrates, with thebiogeographic distribution of each group during the Ordovician indicated. Black bars indicate observedoccurrences, lines indicates inferred ghost ranges. Phylogenetic tree based on Donoghue et al. (2000) andDonoghue & Smith (2001): Stratigraphic data from sources in Sansom et al. (2001) and Smith et al. (2001):absolute ages of series and system boundaries from sources in Cooper (1999). Series abbreviations as inFigure 2.
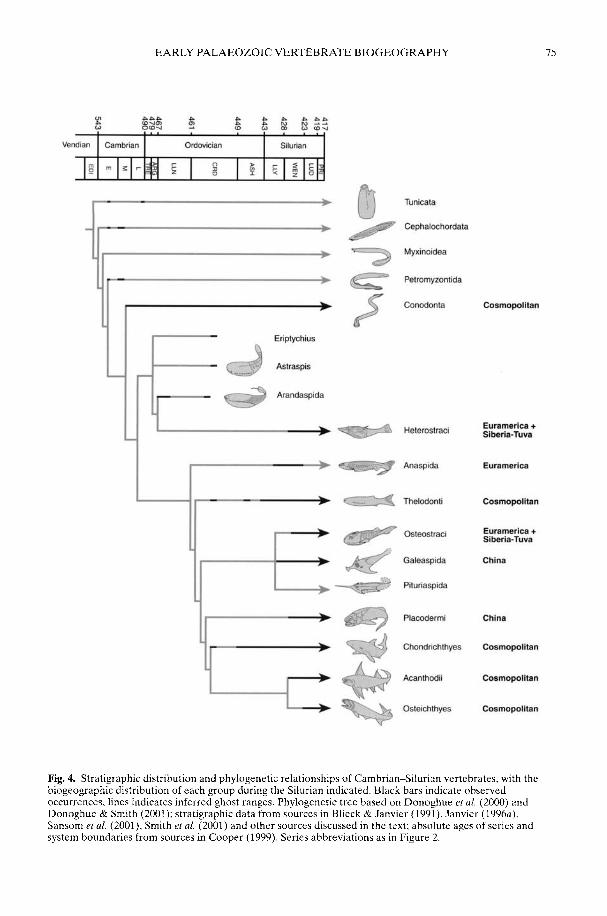
EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 75
Fig. 4. Stratigraphic distribution and phylogenetic relationships of Cambrian-Silurian vertebrates, with thebiogeographic distribution of each group during the Silurian indicated. Black bars indicate observedoccurrences, lines indicates inferred ghost ranges. Phylogenetic tree based on Donoghue et al. (2000) andDonoghue & Smith (2001); Stratigraphic data from sources in Blieck & Janvier (1991), Janvier (1996a),Sansom et al (2001), Smith et al. (2001) and other sources discussed in the text; absolute ages of series andsystem boundaries from sources in Cooper (1999). Series abbreviations as in Figure 2.

76 M. P. SMITH, P. C. J. DONOGHUE & I. J. SANSOM
trees differed only in the relationship ofosteostracans, galeaspids and pituriaspids,which together constitute the sister group tojawed vertebrates. Myxinoids and petromyzon-tids lie at the base of the craniate/vertebrateclade, with conodonts resolved as the sistergroup of all other jawless and jawed vertebrates.The relationships of derived jawless vertebrateswere further resolved by Donoghue & Smith(2001) who argued that the majority ofthelodonts comprise a monophyletic group andconstitute the sister taxon to osteostracans,pituriaspids, galeaspids and jawed vertebrates(Figs 2-4).
Ghost ranges and their implicationsCalibrating cladograms against stratigraphicrange data enables the completeness of the fossilrecord to be assessed as it provides a means ofinferring the existence of unsampled taxa. Suchan approach was first adopted by Gauthier et al(1988), and the concept of 'ghost lineages' or'ghost ranges' was subsequently developed byNorell (1992), amongst others. As useful asghost ranges are, they are no panacea andrequire a number of assumptions upon whichto base the inference of an unsampled (orunsampleable) fossil record. The most basicassumption requires that the cladogram is atleast a reasonable reflection of the true tree ofrelationships. The second requires that the taxarepresented in the cladogram must be mono-phyletic, since the inclusion of paraphyletic taxaleads to spurious inferences of ghost lineages(see e.g. Wagner 1998).
On the first count, we note that although thetree used is only relatively weakly supported atvarious nodes (Donoghue et al. 2000; Donoghue& Smith 2001), a degree of confidence isprovided by the consistency of the signal arrivedat following experimentation with the dataset(Donoghue et al. 2000) and its congruence withanalyses of independent datasets (e.g. Forey &Janvier 1993, 1994; Forey 1995; Janvier 1996b).There is also good evidence to support theassumption that the operational taxa are mono-phyletic given that synapomorphies for each ofthese groups have been identified throughcharacter analysis (see e.g. Janvier 1996a) andnumerical cladistic analysis of lower rank taxacorroborates this assumption (Donoghue &Smith 2001).
On the basis of these assumptions, it ispossible to interpret the stratigraphically cali-brated trees presented in Figures 2-4. The treesindicate that although the fossil record of mostmajor groups of lower vertebrates does not
begin until the Silurian, all have ghost lineagesthat extend into the Ordovician, suggesting thatthe early evolutionary history of these groups isat best unsampled, or at worst unrepresented. Inconsequence, assessments of diversity changecarried out at family level and above (e.g.Benton 1999) cannot be applied with anyconfidence to the analysis of early vertebrateevolution, since entire orders are missing fromthe Ordovician record.
Biogeographic trends - patterns andprocesses
Cam b rian-Ordovician
Evidence for the biogeographic distribution ofEarly and Mid-Cambrian vertebrates is scantand insufficient to derive secure models, otherthan to conclude that, by the Mid-Cambrian,vertebrates were probably present in both Chinaand Laurentia. It is not until the first appearanceof biomineralized vertebrates, Anatolepis andeuconodonts, in the Late Cambrian, that thereare sufficient data to draw firm conclusionsregarding biogeographic patterns. As a clade,euconodonts are cosmopolitan and have asynchronous first appearance within the limits ofbiostratigraphic resolution. At lower taxonomiclevels, many euconodont taxa are also cosmo-politan (Miller 1984) although some endemicgenera and species are present in the Cambrian,and provincialism develops in the Ordovicianbetween high- and low-latitude faunas(Rasmussen 1998; Armstrong & Owen 2002).
In distinct contrast, the early 'ostracoderm'Anatolepis is exclusively circum-Laurentianin its distribution. Indeed, there are noknown cosmopolitan 'ostracoderms' during theCambrian-Ordovician interval, leading tohighly regionalized distribution patterns. Manymajor clades, including astraspids, hetero-stracans, thelodonts and chondrichthyans havetheir earliest occurrences in Laurentia, togetherwith a number of plesiomorphic taxa of un-certain affinity, some as yet unnamed (Sansom etal. 2001), that are known exclusively frommicrovertebrate assemblages. That such taxaare exclusively Laurentian in their distributionduring the Ordovician is supported by theirrepeated occurrence in Upper Ordoviciandeposits throughout Laurentia (Sansom et al.2001, unpublished data) and their absence fromcoeval microvertebrate assemblages fromGondwana and elsewhere (e.g. Young 1997),which does not seem to be an artefact ofsampling (see below). The available dataset thusimplies that the latest common ancestor of all

EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 77
'ostracoderms' and jawed vertebrates wasLaurentian, rather than Gondwanan (contraElliott et al 1991).
In this context, the phylogenetic and spatialrelationship between the arandaspids and other'ostracoderms' is puzzling, since the group isentirely restricted to Gondwana (Fig. 3).Astraspis, the sister taxon of the arandaspids +heterostracans, is present throughout Laurentia,but endemic to it. In turn, heterostracans have afirst record in the Wenlock of Laurentia (Soehn& Wilson 1990) and throughout their strati-graphic range are restricted to Euramerica +Siberia-Tuva. Therefore, despite the strictendemicity exhibited by 'ostracoderm' groups,the presence of arandaspids in Gondwanarequires an absence of barriers betweenLaurentia and Gondwana for at least some ofthe time interval between the Late Cambrianand earliest Ordovician (Fig. la, b), and thisbiogeographic pattern lends some support toDalziel's (1997) model for tectonic interactionbetween Laurentia and South America duringthe Ordovician.
Most groups exhibit endemicity through tothe Wenlock, although significant exceptionsamongst the 'ostracoderms' and jawedvertebrates are thelodonts and mongolepidchondrichthyans. The oldest thelodonts areexclusively Laurentian (Sansom et al 2001), butby the end of the Ordovician they had dis-persed to Baltica (Timan-Pechora) and peri-Gondwana, and were present in Siberia andTuva by the end of the Llandovery. A similarpattern is evident amongst the mongolepidchondrichthyans that have a first occurrence inLaurentia (Sansom et al. 2001) but which, by theend of the Llandovery, were present in NorthChina, South China and Mongolia (Karatajute-Talimaa 1996; Sansom et al. 2000).
Silurian
The ghost lineages suggest that the origin ofmost of the major groups of jawed and jawlesslower vertebrates lies within the Ordovicianand, thus, that the dramatic change in vertebratedistribution during the Silurian is deceptive.There is, nevertheless, an increase in the bio-geographic range, abundance and diversity of allgroups during the Silurian.
Blieck & Janvier (1991) recognized fourSilurian vertebrate provinces: Euramerica(Laurentia, Baltica, Avalonia plus Kara-Taimyr), Siberia, Tuva and China. Within theEuramerican Province, tectonic convergencein the form of successive terrane and conti-nent-continent collisions led to the assembly of
Euramerica. This tectonic setting, with anabsence of deep oceanic barriers, enabled 'ostra-coderms' to disperse across the range of terranesand continental blocks that made up the ORScontinent. The assembly of Euramerica alsocoincides with the increased abundance of arange of vertebrate clades in shallow marineenvironments. The timing of dispersal correlateswell with dates for the collision events derivedfrom other sources (see above), since none ofthe elements of the dispersal predate theWenlock. The assembly of the ORS continentand the dispersal of vertebrates are also coevalwith the breakdown of endemism in inverte-brate faunas (Hallam 1994).
As noted above, Tuva had docked withSiberia by the early Silurian (Bachtadse et al2000), but the exact position of Siberia-Tuvaand its relationship to Euramerica during thelater Silurian and Devonian is the subject ofsome equivocation (cf. Torsvik et al. 1995; Cocks2001). However, stratigraphic data indicate thatosteostracans and heterostracans spread fromEuramerica to Siberia-Tuva, suggesting theabsence of deep water barriers at that time.Once this initial dispersal had taken place,relatively high levels of endemicity were thenmaintained between the two blocks during thelate Silurian and Devonian, with amphiaspidheterostracans and tannuaspid osteostracansbeing unique to Siberia and Tuva respectively(Blieck & Janvier 1991; Young 1991,1993). Thepresence of the distinctive endemic tannuaspidfauna within the Tuva part of the Siberia-Tuvablock (Blieck & Janvier 1991; Janvier 1996a)correlates with the development of the endemicTuvaella brachiopod fauna, and may be relatedto the high-latitude position of Tuva (Cocks2001).The Silurian biogeography of vertebrates inChina and Vietnam contrasts markedly with thatof Euramerica. Osteostracans and heterostra-cans are absent and faunas are dominated byendemic galeaspids from the Llandoveryonwards, together with thelodonts and, from theWenlock, acanthodians and placoderms (Wang1995; P'an et al 1996; Thanh et al 1997). Thehigh degree of endemicity is consistent withsubstantial oceanic separation between Chinaand Euramerica/Siberia, but the palaeogeo-graphic origin of galeaspids is puzzling becausetheir closest known relatives are exclusivelyLaurentian/Euramerican (Donoghue & Smith2001). This posits the conclusion that the latestcommon ancestor of galeaspids and their nearestrelatives was, again, Laurentian. The onlyderived 'ostracoderms' to have dispersed fromLaurentia to China were the thelodonts, and

78 M. P. SMITH, P. C. J. DONOGHUE & I. J. SANSOM
this group may offer a clue to the origin ofendemic galeaspids in China.
In Gondwana, 'ostracoderms' disappear fromthe record after the Caradoc, presumably as aresult of glaciation (Elliott et al. 1991) and theonly reliably documented Silurian vertebratesfrom Gondwana are conodonts, thelodonts andjawed vertebrates (Blieck & Janvier 1991).Indeed, some parts of Gondwana apparentlyremained unpopulated until the dispersal ofjawed vertebrates during the Devonian. This isdemonstrably not a sampling artefact, sinceconodont faunas from Australia and Laurentiaare comparable, but 'ostracoderm' biodiversityis not. By the Early Devonian, an endemicplacoderm province (the 'wuttagoonaspid-phyllolepid' province) is recognizable in EastGondwana which began to break down in theLate Devonian (Young 1991, 1993). WestGondwana, in contrast, is relatively depauperatein placoderms and has chondrichthyan-acanthodian-dominated faunas at that time(Young 1993).
The Silurian thus records a complex history ofdispersal, vicariance and tectonic convergence.Acanthodians, thelodonts and chondrichthyanscontinue to be widely dispersed and almostcertainly had genuine trans-oceanic dispersalcapability (contra Blieck & Janvier 1991, p. 377).Heterostracans, anaspids and osteostracans alsobegan to disperse, but only after the tectonicassembly of the ORS continent had removedoceanic barriers. In Siberia, a degree of vicariantendemism is observed after the original input ofheterostracans and osteostracans fromEuramerica, and in China a significant degree ofendemism is also established following initialdispersal.
Where are all the Cambro-Ordovicianvertebrates?One of the most intriguing aspects of this newperspective on Cambro-Ordovician vertebratebiodiversity does not in itself stem from thediscovery of vertebrate remains in rocks of thisage but, rather, the discovery of significant gapsin the record in the form of ghost ranges. Theimproved stratigraphic constraints and betterunderstanding of phylogenetic relationshipsprovided by Cambro-Ordovician vertebratesreveal long ghost ranges which imply that mostof the major groups of 'ostracoderms' and primi-tive jawed vertebrates have an evolutionaryhistory that extends into the Ordovician. It islikely that many of the newly discoveredCambro-Ordovician microvertebrate remains of
currently uncertain affinity will fulfil theprediction of these ghost lineages, but there aremany more ghost lineages than there aretangible candidates. There are at least threepossible, and non-mutually exclusive, expla-nations for the dramatic improvement in thequality of the vertebrate fossil record during theearly-middle Silurian: (a) the increase in thetaxonomic diversity and disparity of vertebratefossil record accurately reflects an early Siluriancladogenic event (cf. Blieck & Janvier 1991) andthe inferences of ghost lineages are entirelyspurious; (b) Cambrian and Ordovician verte-brates are rare because there is a systematic biasagainst the preservation of the environments inwhich they lived; and/or (c) Cambro-Ordovicianvertebrates were ecologically distinct from theirmiddle Palaeozoic relatives and their fossilrecord is either undersampled or the preser-vation of fossil remains unlikely because ofsystematic bias.
The fossil record of Cambrian and Ordovicianvertebrates (except for conodonts) is so poorthat it is not possible to test comprehensivelywhether or not vertebrates were affected bythe latest-Ordovician extinction event. Never-theless, there is evidence that at least somevertebrate groups were affected. The fossilrecord of conodonts exhibits a dramatic drop intaxonomic diversity during this event such thatthe entire clade almost became extinct(Aldridge 1988; Armstrong 1995). However, thismodel has yet to be tested against phylogenetictrees and it is notable that not one of the majorconodont groups became extinct. Furthermore,all of the Ordovician vertebrate remains discov-ered thus far can be identified either as membersof clades surviving into the middle Palaeozoic,or as possible sister taxa to one or more of theseclades. Thus, although it is likely that thevertebrate clade as a whole was affected by theextinction event locally, especially at lowtaxonomic level and in terms of numericaldiversity, there is no evidence that any majorvertebrate groups arose or met their demise atthis time, a pattern mirrored in invertebratefaunas (Droser et al 2000). The absence ofextinction in major vertebrate groups indicatesthat the increase in the quality of the vertebraterecord during the Silurian is not the result ofmajor evolutionary turnover at high taxonomiclevel.
The second possibility, that the fossil record ofearly vertebrates is poor because of a systematicbias against the preservation of shallow-waternearshore lithofacies, is also doubtful. Suchenvironments are both extensively preservedand crop out extensively throughout the

EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 79
Ordovician of Laurentia in North America. Thisinterval has been intensively sampled forconodonts, which are abundantly preservedthroughout, but the 'ostracoderm' record isconfined to specific intervals that seem tocoincide with highstand events (Sansom et al.2001). It is clear, therefore, that althoughshallow-water, nearshore environments arewidely preserved, they contain no record ofvertebrates, other than conodonts. Thus, itwould appear that the third of the alternatives isworthy of further consideration and investi-gation, namely that Ordovician 'ostracoderms'were ecologically distinct from their middlePalaeozoic relatives and are preserved in verydifferent facies which are either unsampledand/or systematically underrepresented in therock record. If this is the case, the two endmembers that could serve as refugia for Ordovi-cian vertebrates are freshwater and deep marineenvironments. The former may be excluded.Despite continuing appeals for the freshwaterorigin of vertebrates (Graffin 1992; Griffith1994), no Cambro-Ordovician vertebrates arefound in freshwater environments and Blieck &Janvier (1991) argued that most or all Silurianvertebrates were marine. To the contrary,Anatolepis occurs predominantly in outer-shelf,circum-Laurentian settings and the HardingSandstone and Stairway Sandstone faunas maycoincide with sea-level highstands (Sansom et al2001). This suggests that the missing Ordovicianrecord may be found in deeper-water, outboardsettings which, due to their position, tend to bemore deformed and metamorphosed thancratonic interiors. Prospecting these areas forvertebrate faunas may provide additionalinsights into early vertebrate evolution.
What underlies endemism andcosmopolitanism during the Silurian?
There is a notable correlation between thebiogeographic distribution of vertebrates duringthe Silurian and their anatomical design. Forinstance, thelodonts are the only monophyleticgroup of 'ostracoderms' to exhibit cosmo-politanism during the Silurian, the other groupsbeing restricted to the margins of Laurentia and,later, Euramerica plus Siberia-Tuva (with theexception of galeaspids, which are restricted toChina). In common with the thelodonts andconodonts, jawed vertebrates also exhibitcosmopolitanism during this interval. It ispossible that the heavily armoured groups of'ostracoderms' remained endemic to the ORScontinent throughout the middle Palaeozoicbecause they were constrained against active
swimming in deeper water and/or open oceanenvironments. Indeed, it has been argued byGutmann (1967) and Gutmann & Bonik (1983)that, in the absence of a mineralized axialskeleton, the heavily armoured ' ostracoderms'(the Panzerfische of Gutmann) used theirextensive dermal skeleton to brace musculature.The mineralized axial skeleton of jawed verte-brates (a synapomorphy of the clade) provides abiomechanically superior structure againstwhich to brace musculature. As part of their'new head' hypothesis, Gans & Northcutt (1983)noted that the origin of vertebral elementsimmediately precedes the advent of paired fins,as the final stage of their scenario in whichvertebrate evolution was driven by a change infeeding strategies towards active predation. Wesuggest, therefore, that the heavily armoureddermal skeleton of most pre-jawed vertebrateswas a limiting factor in their dispersal. Far fromthere being a direct competitive link betweenthe 'ostracoderms' and their jawed relatives (seee.g. Purnell 2001), it is possible that the greaterdispersal potential of the jawed vertebrates wasa significant factor in the longevity of the group,while the 'ostracoderms' underwent a pro-gressive decline to their demise in the LateDevonian. This greater dispersal potentialwould, in turn, enable the subsequent develop-ment of areas of endemism and concomitantlyhigher levels of global biodiversity which areencountered in Devonian jawed vertebrates.
ConclusionsWe began with the 'out of Gondwana' vicariancemodel of Ordovician vertebrate spatial andtemporal diversification (Elliott et al. 1991) and,with the incorporation of new and more com-plete datasets, conclude with a new model thatincorporates a complex interaction of dispersal,vicariance, and tectonic convergence during theEarly Palaeozoic. New datasets and the elucida-tion of existing datasets lead to the suggestionthat much of the early evolutionary history ofthe major early vertebrate clades lies within theOrdovician, or even the Cambrian. Further-more, it is likely that the record of these eventsis poor because the organisms were ecologicallydistinct from their middle Palaeozoic relatives,rather than because of a systematic bias in thepreservation of environments typical of thesecounterparts.
We provide this assessment of the evol-utionary palaeobiogeography of Early Palaeo-zoic vertebrates based on the data that iscurrently to hand. However, biogeographicanalyses based upon historical evidence are, by

80 M. P. SMITH, P. C. J. DONOGHUE & I. J. SANSOM
their very nature, reliant upon incompletedatasets and it is likely that unexpected discov-eries will lead to radical reinterpretation of thedata presented herein. Nevertheless, we feel thatthe database has grown to an extent that we canbegin to build testable models with the aim ofstimulating the search for new data in both timeintervals and palaeoenvironments in which theremains of the earliest vertebrates might other-wise be unexpected and therefore remainunsampled.
We are grateful to A. Owen and A. Crame for invitingus to contribute to the Lyell Meeting. The paper hasbeen significantly improved by comments from H.Armstrong, T. Hallam and P. Janvier, and the work hasbeen funded by NERC grants GR3/10272 andNER/B/S/2000/OQ284 and by NERC Post-DoctoralResearch Fellowship GT5/99/ES/2.
ReferencesAFANASSIEVA, O. & JANVIER, P. 1985. Tannuaspis,
Tuvaspis and Ilemoraspis, endemic osteostracangenera from the Silurian and Devonian of Tuvaand Khakassia (USSR). Geobios, 18, 493-506.
ALDRIDGE, R. J. 1988. Extinction and survival in theConodonta. In: LARWOOD, G.P. (ed.) Extinctionand Survival in the Fossil Record. SystematicsAssociation Special Volume, 34, 231-256.
ALDRIDGE, R. J., BRIGGS, D. E. G., CLARKSON,E. N. K. & SMITH, M. P. 1986. The affinities ofconodonts - new evidence from the Carbonifer-ous of Edinburgh, Scotland. Lethaia, 19, 279-291.
ALDRIDGE, R. I, BRIGGS, D. E. G., SMITH, M. P..CLARKSON, E. N. K. & CLARK, N. D. L. 1993. Theanatomy of conodonts. Philosophical Transac-tions of the Royal Society of London, Series B.340,405-421.
ARMSTRONG, H. A. 1995. High-resolution biostrat-igraphy (conodonts and graptolites) of the UpperOrdovician and Lower Silurian - evaluation ofthe Late Ordovician mass extinction. ModernGeology, 20, 41-68.
ARMSTRONG, H. A. & OWEN, A. W. 2001. Terraneevolution of the paratectonic Caledonides ofnorthern Britain. Journal of the GeologicalSociety, London, 158, 475-486.
ARMSTRONG, H. A. & OWEN, A. W. 2002.Euconodont diversity changes in a cooling andclosing lapetus Ocean. In: CRAME, J. A. & OWEN,A. W. (eds) Palaeobiogeography and BiodiversityChange: the Ordovician and Mesozoic-CenozoieRadiations. Geological Society, London, SpecialPublications, 194, 85-98.
ASTINI, R. A., BENEDETTO, J. L., & VACCARI, N. E.1995. The early Paleozoic evolution of theArgentine Precordillera as a rifted, drifted andcollided terrane - A geodynamic model. Geo-logical Society of America Bulletin, 107, 253-273.
BACHTADSE, V., PAVLOV, V. E., KAZANSKY, A. Y. &TAIT, J. 2000. Paleomagnetism of the Tuva Terrane(Russia) - evidence for low latitudes of the
Siberian craton in Siluro-Devonian times. Journalof Geophysical Research. 105(B6), 13,509-13,518.
BENGTSON, S. 1976. The structure of some MiddleCambrian conodonts, and the early evolution ofconodont structure and function. Lethaia. 9.185-206.
BENTON, M. J. 1999. The history of life: databases inpalaeontology. In: HARPER. D. A.T. (ed.) Numeri-cal Palaeobiology. John Wiley & Sons.Chichester, 249-283.
BLIECK, A. & JANVIER, P. 1991. Silurian vertebrates.Special Papers in Palaeontology, 44. 345-389.
BOCKELIE, T. & FORTEY, R. A. 1976. An early Ordo-vician vertebrate. Nature, 260. 36-38.
CHEN, JUN-YUAN, HUANG, Di-YiNG & LI, CHIA-WEl.1999. An early Cambrian craniate-like chordate.Nature, 402, 518-522.
COCKS, L. R. M. 2000. The Early Palaeozoic geographyof Europe. Journal of the Geological Society,London. 157, 1-10.
COCKS, L. R. M. 2001. Ordovician and Silurian globalgeography. Journal of the Geological Society.London. 158, 197-210.
COOPER, R. A. 1999. The Ordovician time scale -calibration of graptolite and conodont zones. AdaUniversitas Carolinae - Geologica. 43(1/2), 1-4.
DALLA SALDA, L. H., DALZIEL, I. W. D.. CINGOLANL.C. A. & VARELA, R. 1992. Did the TaconicAppalachians continue into southern SouthAmerica? Geology, 20. 1059-1062.
DALZIEL, I. W. D. 199i. Pacific margins of Laurentiaand East Antarctica-Australia as a conjugate riftpair: Evidence and implications for an Eocam-brian supercontinent. Geology. 19, 598-601.
DALZIEL. I. W. D. 1997. Neoproterozoic-Paleozoicgeography and tectonics: Review, hypothesis,environmental speculation. Geological Society ofAmerica Bulletin. 109. 16-42.
DENISON, R. H. 1967. Ordovician vertebrates fromWestern United States. Fieldiana Geology. 16.131-192.
DONOGHUE, P. C. J. 1998. Growth and patterning inthe conodont skeleton. Philosophical Trans-actions of the Royal Society of London, Series B.353,633-666.
DONOGHUE, P. C. J. & ALDRIDGE, R. J. 2001. Originof a mineralised skeleton. In: AHLBERG. P. E. (ed.)Major Events in Early Vertebrate Evolution:Palaeontology, Phylogeny, Genetics and Develop-ment. Taylor & Francis. London. 85-105.
DONOGHUE, P. C. J. & SMITH, M. P. 2001. The anatomyof Turinia pagei (Powrie) and the phylogeneticstatus of the Thelodonti. Transactions of theRoyal Society of Edinburgh: Earth Sciences. 92.15-37.
DONOGHUE, P. C. J., FOREY, P. L. & ALDRIDGE. R. J.2000. Conodont affinity and chordate phylogeny.Biological Reviews. 75, 191-251.
DROSER, M. L., BOTTJER. D. J., SHEEHAN. P. M. &McGHEE, G. R. 2000. Decoupling of taxonomicand ecologic severity of Phanerozoic marine massextinctions. Geology, 28, 675-678.
ELLIOTT. D. K.. BLIECK. A. R. M. & GAGNIER. P.-Y.1991. Ordovician vertebrates. Geological Surveyof Canada, Paper. 90-9. 93-106.

EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 81
ELVEVOLD, S., ESCHER, J. C, FREDERILSEN, K. S., ETAL, 2000. Tectonic architecture of the East Green-land Caledonides 72°-74°30' N. Danmark ogGronlands Geologiske Undersogelse Rapport,2000/88.
ERDTMANN, B.-D., WEBER, B., SCHULTZE, H.-P. &EGENHOFF, S. 2000. A possible agnathan platefrom the lower Arenig (Lower Ordovician) ofsouth Bolivia. Journal of Vertebrate Paleontology,20, 394-399.
FOREY, P. L. 1995. Agnathans recent and fossil, andthe origin of jawed vertebrates. Reviews in FishBiology and Fisheries, 5, 267-303.
FOREY, P. L. & JANVIER, P. 1993. Agnathans and theorigin of jawed vertebrates. Nature, 361,129-134.
FOREY, P. L. & JANVIER, P. 1994. Evolution of theearly vertebrates. American Scientist, 82,554-565.
FREEMAN, S. R., BUTLER, R. W. H., CLIFF, R. A. &REX, D. C. 1998. Direct dating of mylonite evol-ution: a multi-disciplinary geochronological studyfrom the Moine Thrust Zone, NW Scotland.Journal of the Geological Society, London, 155,745-758.
GABBOTT, S. E., ALDRIDGE, R. J. & THERON, J. N.1995. A giant conodont with preserved muscletissue from the Upper Ordovician of SouthAfrica. Nature, 374, 800-803.
GAGNIER, P. Y., BLIECK, A. & RODRIGO, G. 1986,First Ordovician vertebrate from South America.Geobios, 19, 629-634.
GANS, C. & NORTHCUTT, R. G. 1983. Neural crest andthe origin of vertebrates: a new head. Science, 220,268-274.
GAUTHIER, J, KLUGE, A. G. & ROWE, T. 1988.Amniote phylogeny and the importance of fossils.Cladistics, 4,105-209.
GRAFFIN, G. 1992. A new locality of fossiliferousHarding Sandstone: evidence for freshwaterOrdovician vertebrates. Journal of VertebratePaleontology, 12,1-10.
GRIFFITH, R. W. 1994. The life of the first vertebrates.BioScience, 44, 408-417.
GUTMANN, W. F. 1967. Das Dermalskelett der -fossilen 'Panzerfische' funktionell und phylo-genetisch interpretiert. Senckenbergiana Lethaea,48, 277-283.
GUTMANN, W. F. & BONIK, K. 1983. Die Entstehungdes Dermalskelettes: Biomechanische Grundbe-dingungen des fossilen Schuppenpanzers. Senck-enbergiana Lethaea, 64,179-197.
HALLAM, A. 1994. An Outline of Phanerozoic Bio-geography. Oxford University Press, Oxford.
HARTZ, E. B., ANDRESEN, A., HODGES, K. V. &MARTIN, M. W. 2001. Syncontractional extensionand exhumation of deep crustal rocks in the eastGreenland Caledonides. Tectonics, 20, 58-77.
HIGGINS, A. K., LESLIE, A. G. & SMITH, M. P. 2001.Neoproterozoic-Lower Palaeozoic strati-graphical relationships in the marginal thin-skinned thrust belt of the East GreenlandCaledonides: comparisons with the foreland inScotland. Geological Magazine, 138,143-160.
HOFFMAN, P. F. 1991. Did the breakout of Laurentiaturn Gondwana inside out? Science, 252,1409-1412.
HOLLAND, N. D. & CHEN, JUN-YUAN. 2001. Origin andearly evolution of the vertebrates: new insightsfrom advances in molecular biology, anatomy,and palaeontology. Bioessays, 23,142-151.
JANVIER, P. 1996a. Early Vertebrates. Oxford Uni-versity Press, Oxford.
JANVIER, P. 1996b. The dawn of the vertebrates: char-acters versus common ascent in the rise of currentvertebrate phylogenies. Palaeontology, 39.259-287.
KARATAJUTE-TALIMAA, V. 1996. The Mongolepida:scale structure and systematic position. GeobiosMemoire Special, 19, 35-37.
KARATAJUTE-TALIMAA, V. 1997. Lugalepis ~ a newgenus of elasmobranchs from Devonian of thewestern part of the main Devonian field.Geologija, 21, 24-29.
KUMAR, S. & HEDGES, S. B. 1998. A moleculartimescale for vertebrate evolution. Nature, 392,917-920.
MCKERROW, W. S., DEWEY, J. F. & SCOTESE, C. R.1991. The Ordovician and Silurian developmentof the Iapetus Ocean. In: BASSETT, M.G. et al(eds) The Murchison Symposium. Proceedings ofan International Conference on the SilurianSystem. Special Papers in Palaeontology, 44,165-178.
MCKERROW, W. S., MAC NIOCAILL, C. & DEWEY, J. F,2000. The Caledonian Orogeny redefined.Journal of the Geological Society, London, 157,1149-1154.
MAC NIOCAILL, C. 2000. A new Silurian palaeolatitudefor eastern Avalonia and evidence for crustalrotations in the Avalonian margin of southwest-ern Ireland. Geophysical Journal International,141,661-671.
MILLER, J. F. 1984. Cambrian and earliest Ordovi-cian conodont evolution, biofacies, and provin-cialism. In: CLARK, D.L. (ed.) Conodont Biofaciesand Provincialism. Geological Society ofAmerica, Special Papers, 196, 43-68.
MOORES, E. M. 1991. Southwest U.S.-East Antarctica(SWEAT) connection: A hypothesis. Geology,24, 917-920.
NORELL, M. A. 1992. Taxic origin and temporaldiversity: the effect of Phylogeny. In: NOVACEK,M. J. & WHEELER, Q. D. (eds) Extinction andPhylogeny. Columbia University Press, NewYork, 89-118.
P'AN, JIANG, Lu, LIWU. & JI, SHU'AN 1996. A briefreview of studies on palaeogeography of MiddlePaleozoic vertebrates in China. Journal of South-east Asian Earth Sciences, 13,185-190.
PURNELL, M. A. 2001. Scenarios, selection, and theecology of early vertebrates. In: AHLBERG, P. E.(ed.) Major Events in Early Vertebrate Evolution:Palaeontology, Phylogeny, Genetics and Develop-ment. Taylor & Francis, London, 187-208.
RASMUSSEN, J. A. 1998. A reinterpretation of theconodont Atlantic Realm in the late EarlyOrdovician (Early Llanvirn). Proceedings ofthe Sixth European Conodont Symposium(ECOS VI). Palaeontologica Polonica, 58, 67-77.
REPETSKI, J. E. 1978. A fish from the Upper Cam-brian of North America. Science, 200, 529-531.

82 M. P. SMITH, P. C. J. DONOGHUE & I. J. SANSOM
RITCHIE, A. & GILBERT-TOMLINSON, J. 1977. FirstOrdovician vertebrates from the Southern Hemi-sphere. Alcheringa, 1, 351-368.
SANSOM, I. J. 1996. Pseudooneotodus: a histologicalstudy of an Ordovician to Devonian vertebratelineage. Zoological Journal of the LinneanSociety, 118, 47-57.
SANSOM, I. I, SMITH, M. P., ARMSTRONG, H. A. &SMITH, M. M. 1992. Presence of the earliest verte-brate hard tissues in conodonts. Science, 256,1308-1311.
SANSOM, I. J, SMITH, M. P. & SMITH, M. M. 1994,Dentine in conodonts. Nature, 368, 591.
SANSOM, I.J, SMITH, M. P., SMITH, M. M. & TURNER,P. 1995. The Harding Sandstone revisited - a newlook at some old bones. Geobios, 19, 57-59.
SANSOM, I. J, SMITH, M. P. & SMITH, M. M. 1996.Scales of thelodont and shark-like fishes from theOrdovician. Nature, 379, 628-630.
SANSOM, I. J., ALDRIDGE, R. J. & SMITH, M. M. 2000. Amicrovertebrate fauna from the Llandovery ofSouth China. Transactions of the Royal Society ofEdinburgh: Earth Sciences, 90, 255-272.
SANSOM, I. J., SMITH, M. M. & SMITH, M. P. 2001.The Ordovician radiation of vertebrates. In:AHLBERG, P. E. (ed.) Major Events in EarlyVertebrate Evolution: Palaeontology, Phylogeny,Genetics and Development. Taylor & Francis,London, 156-171.
SCOTESE, C. R. 1986. Phanerozoic reconstructions: anew look at the assembly of Asia. University ofTexas Institute for Geophysics Technical Report,66,1-54.
SCOTESE, C. R. & MCKERROW, W. S. 1990. RevisedWorld maps and introduction. In: MCKERROW, W.S. & SCOTESE, C. R. (eds) Palaeozoic Palaeo-geographyandBiogeography. Geological Society,London, Memoirs, 12,1-21.
SHU, D.-G, Luo, H.-L., CONWAY MORRIS, S., ET AL.1999a. Lower Cambrian vertebrates from southChina. Nature, 402,42-46.
SHU, D.-G, CONWAY MORRIS, S., ZHANG, X.-L., CHEN,L., LI, Y. & HAN, J. 1999b. A pipiscid-like fossilfrom the Lower Cambrian of south China. Nature,400, 746-749.
SIMONETTA, A. M. & INSOM, E. 1993. New animalsfrom the Burgess Shale (Middle Cambrian) andtheir possible significance in understanding theBilateria. Bollettino Zoologica, 60, 97-107.
SMETHURST, M. A., KHRAMOV, A. N. & TORSVIK, T. H,1998. The Neoproterozoic and Palaeozoic palaeo-magnetic data for the Siberian Platform: FromRodinia to Pangea. Earth Science Reviews, 43,1-24.
SMITH, M. M. 1991. Putative skeletal neural crest cellsin early Late Ordovician vertebrates from Col-orado. Science, 251, 301-303.
SMITH, M. M. & SANSOM, I. J. 1997. Exoskeletalmicroremains of an Ordovician fish from theHarding Sandstone of Colorado. Palaeontology,40, 645-658.
SMITH, M. M., SANSOM, I. J. & SMITH, M. P. 1995.Diversity of the dermal skeleton in Ordovician toSilurian vertebrate taxa from North America: his-tology, skeletogenesis and relationships. Geobios,19, 65-70.
SMITH, M. P., BRIGGS, D. E. G. & ALDRIDGE. R. J.1987. A conodont animal from the lower Silurianof Wisconsin, USA, and the apparatus architec-ture of panderodontid conodonts. In: ALDRIDGE,R. J. (ed.) Palaeobiology of Conodonts, EllisHorwood, Chichester, 91-104.
SMITH, M. P., SANSOM, I. J. & REPETSKI, J. E. 1996.Histology of the first fish. Nature, 380, 702-704.
SMITH, M. P., SANSOM, I. J. & COCHRANE, K. D. 2001.The Cambrian origin of vertebrates. In:AHLBERG, P.E. (ed.) Major Events in Early Verte-brate Evolution: Palaeontology, Phylogeny,Genetics and Development. Taylor & Francis.London, 67-84.
SOEHN, K. L. & WILSON, M, V. H. 1990. A complete,articulated heterostracan from Wenlockian(Silurian) beds of the Delorme Group, Macken-zie Mountains, Northwest Territories, Canada.Journal of Vertebrate Paleontology. 10, 405-419.
SWEET, W. C. 1984. Graphic correlation of upperMiddle and Upper Ordovician rocks of theUnited States Midcontinent. In: Bruton, D.L.(ed.) Aspects of the Ordovician System. Palaeon-tological Contributions of the University of Oslo,295, 23-35.
SWEET, W. C. 1988. The Conodonta: Morphology,Taxonomy, Paleoecology, and EvolutionaryHistory of a Long-extinct Animal Phylum. Claren-don Press, Oxford.
THANH, TONG-DZUY, PHUONG,TA HOA, BOUCOT, A. J..GOUJET,D. & JANVIER,P. 1997. Vertebres siluriensdu Viet-nam central. Comptes rendus de l'A-cademie des sciences, Serie II a, 324, 1023-1030.
THOMAS, W. A. & ASTINI, R. A. 1996. The Argen-tine Precordillera: a traveler from the OuachitaEmbayment of North American Laurentia.Science, 283, 752-757.
TORSVIK, T H., TAIT, J., MORALEV. V. M., MCKERROW,W. S., STURT, B. A. & ROBERTS, D. 1995. Ordo-vician palaeogeography of Siberia and adjacentcontinents. Journal of the Geological Society,London, 152,279-287.
TORSVIK, T. H., SMETHURST, M. A., MEERT, J. G. ET AL.1996. Continental break-up and collision in theNeoproterozoic and Palaeozoic - A tale of Balticaand Laurentia. Earth Science Reviews, 40,229-258.
VAN STAAL, C. R., DEWEY, J. F, MACNIOCAILL. C. &MCKERROW, W. S. 1998. The Cambrian-Siluriantectonic evolution of the northern Appalachiansand British Caledonides: history of complex westand southwest Pacific-type segment of Iapetus. In:BLUNDELL, D. J. & SCOTT. A. C. (eds) Lyell: thePast is the Key to the Present, Geological Society,London, Special Publications, 143.199-242.
WAGNER, P. J. 1998. Phylogenetic analysis and thequality of the fossil record. In: DONOVAN, S. K. andPAUL, C. R. C. (eds) The Adequacy of the FossilRecord. John Wiley & Sons. Chichester, 165-187.
WALCOTT, C. D. 1892. Preliminary notes on the dis-covery of a vertebrate fauna in Silurian (Ordo-vician) strata. Bulletin of the Geological Society ofAmerica, 3,153-171.
WANG, N. Z. 1995. Silurian and Devonian jawlesscraniates (Galeaspida, Thelodonti) and theirhabitats in China. Bulletin du Museum nationald'Histoire naturelle. Paris. 17. 57-84.

EARLY PALAEOZOIC VERTEBRATE BIOGEOGRAPHY 83
WILSON, J. T. 1966. Did the Atlantic close and then YOUNG, G. C. 1993. Vertebrate faunal provinces inre-open? Nature, 211, 676-681. the Middle Palaeozoic. In: LONG, J. A. (ed.)
YOUNG, G. C. 1991. Devonian vertebrate distribution Palaeozoic Vertebrate Biostratigraphy and Bio-patterns and cladistic analysis of palaeogeo- geography. Bellhaven Press, London, 293-323.graphic hypotheses. In: McKERROW, W. S. & YOUNG, G. C. 1997. Ordovician microvertebrateSCOTESE, C R. (eds) Palaeozoic Palaeogeography remains from the Amadeus Basin, central Aus-and Biogeography, Geological Society, London, tralia. Journal of Vertebrate Paleontology, 17,Memoirs, 12, 243-255. 1-25.

This page intentionally left blank

Euconodont diversity changes in a cooling and closing IapetusOcean
H. A. ARMSTRONG1 & A. W. OWEN2
1 Department of Geological Sciences, University of Durham, Durham DH1 3LE, UK(e-mail: [email protected])
2Division of Earth Sciences, University of Glasgow, Gregory Building,Glasgow G12 8QQ, UK
Abstract: Constrained seriation of euconodont generic presence-absence matrices for fourtime slices between the late Llanvirn and late Llandovery provides a qualitative methodfor defining shelf and oceanic biofacies, reconstructing biofacies architectures andanalysing biodiversity within a regional context.
We propose many North Atlantic Province taxa had a pelagic mode of life and rangedwidely across the Iapetus Ocean. Oceanic biofacies are considered to reflect water massstructure. Changes in vertical distribution of one such biofacies (including Amorphog-nathus and Spinodus) suggest adaptation to cold, nutrient-rich, oxygen-poor upwellingwater. Biofacies distributions suggest that upwelling occurred along the Avalonian marginthroughout the Ashgill, but was only initiated along the Laurentian margin immediatelyprior to the Hirnantian glacial maximum.
Clade diversities and trajectories differ between biofacies and latitudes, reflectingdifferent causal mechanisms. In Laurentia, diversity fell in the early Ashgill, coincident withthe onset of ocean cooling. Diversity declined in Avalonia when the microcontinent driftedinto tropical latitudes. The stability of euconodont biofacies architecture during the LateOrdovician indicates that global cooling and plate reorganization had a low palaeoecolog-ical impact despite decreases in alpha and beta diversity.
Over the past 20 years the differential success ofclades has been attributed to either intrinsicproperties of the clades themselves (e.g. com-petitive ability or origination rates) or tochanges in their biogeographical, environmentaland palaeoecological context (Erwin 1998;Jablonski 1998). If intrinsic properties of theclade are the primary control on diversity thenclade diversity is considered independent fromgeological setting. Attempting to correlatechanges in clade diversity with independentlyestablished changes in environmental con-ditions would provide a test of these competinghypotheses.
Two recent studies on Palaeozoic cladessupport the hypothesis that diversity reflectschanging palaeoenvironmental conditions at aregional scale. Miller (1997) compared the earlyOrdovician radiation in six palaeocontinentalregions and found regional differences in theevolutionary history of trilobites, brachiopodsand molluscs. Miller & Mao (1995) showed thatin a generic dataset, corrected for includedspecies number, there was a correlationbetween Ordovician diversity trends and theextent of siliciclastic sedimentation, consideredby them to be a proxy for mountain-buildingactivity. However, aggregate map areas of
marine tectonic provinces through the Ordovi-cian indicate a slight increase in the area!extent of the rock record during the period ofmaximum diversity decline (Miller & Mao1995). There is therefore no correlationbetween decrease in diversity and totalpreserved rock record during the LateOrdovician.
The changing patterns of Phanerozoic bioticdiversity have been recognized almost entirelyfrom synoptic global datasets that, by theirnature, average very different palaeoenviron-mental signals (see Miller 2000 and referencestherein). Detailed studies of local successionscan be ecologically well constrained but raisequestions about the quality of the record,particularly with respect to preservation,restricted sampling and the response of the biotato rapidly changing substrates. Regional com-parative studies provide the best compromise oftaxonomic scope and palaeoenvironmentalacuity (Sepkoski 1993; Miller & Mao 1995;Miller 2000).
The late Ordovician was a time of majorglobal environmental change with the lateOrdovician glaciation punctuating a periodof prolonged global greenhouse climate(Hambrey 1985; Berner 1990, 1992; Crowley &
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194,85-98.0305-8719/02/$15.00 © The Geological Society of London 2002.

86 H. A. ARMSTRONG & A. W. OWEN
Baum 1991). Sedimentological evidence fromDob's Linn, located on the Laurentian slope,indicates that at least in low latitudes oceancooling persisted from the early Ashgill, withthe glacial maximum ranging from the lateRawtheyan to mid-Hirnantian (Armstrong &Coe 1997). This glaciation has been considereda major causal factor in the late Ordovicianmass extinction (see reviews in Brenchley 1988;Brenchley et al. 1995a,b; Armstrong 1995;Owen & Robertson 1995). At the same timeplate configurations were changing dramaticallyassociated with the closure of the Iapetus Ocean(Scotese & McKerrow 1990; van Staal et al.1998)
We have conducted an analysis of LateOrdovician euconodont generic diversity in thecircum-Iapetus region. Different diversity trajec-tories are found in different biofacies in eachof low-latitude Laurentia and mid-temperateAvalonia and Baltica, and can be attributedto different causal mechanisms. Laurentiandiversity appears to have been moderated bychanging climate, associated with the onset of thelatest-Ordovician glaciation, whereas Avaloniandiversity reflects the change from temperate totropical conditions as the plate drifted north-wards. This analysis thus supports the hypothesisthat changing palaeoenvironmental setting is theprimary influence on clade diversity within theeuconodonts.
The method we have applied allows regional-scale diversity patterns to be elucidated fromexisting presence-absence data and is thuswidely applicable. The generic associationsestablished by the method also provide anopportunity to analyse biofacies architectureand stability in a number of different regionalsettings and hence examine the palaeoecologicalimpact of global-scale events (see also Droser etal. 1997, 2000).
MethodPresence-absence matrices for genera havebeen compiled from sections in Laurentia andAvalonia for the upper serra and anserinusbiozones (Llandeilian-Aurelucian), velicuspisChron (equivalent to the upper superbusBiozone in the British biostratigraphicalscheme), upper ordovicicus Biozone and thecelloni Biozone. Genera are considered taxo-nomically stable and are more likely to beidentified correctly. Each genus comprises lessthan five species; several are monospecific. Theresults of a cladistic analysis of euconodontgenera are not currently available but many arelikely to be paraphyletic (P. C. J. Donoghue,
pers. comm., 2001). This does not detract fromour analysis as paraphyletic taxa are more likelyto have had narrower palaeoecological rangesthan monophyletic taxa, in the same way thatmany paraphyletic fish genera have highlyrestricted ecological distributions (Sepkoski &Kendrick 1993; Kemp 1999; Miller 2000).
Two major Ordovician conodont faunalprovinces have previously been identified in theIapetus region: the Midcontinent Province,including Laurentia, Siberia and parts of China;and the North Atlantic Province, includingBaltoscandia and eastern Laurentia (Barnes etal. 1973; Sweet & Bergstrom 1974, 1984).Cluster analysis of Jaccard and Dice coefficientsfor euconodont faunas of the lower Llanvirnindicate that the faunal similarities betweenBaltoscandia and eastern Laurentia are dueentirely to the presence of a cosmopolitan deep-water fauna, the Protopanderodus-PeriodonBiofacies (comparable to OB2 herein) (Ras-mussen 1998). Once this biofacies is removedfrom the analysis then the North AtlanticProvince is restricted to Baltoscandia andneighbouring areas. Genera consideredrepresentative of the North Atlantic Province(including coniform taxa) and the MidcontinentProvince have therefore been separated in thepresence - absence matrix for each region.
The original matrices were compiled with thelocalities arranged with an onshore-offshoredistribution (Figs 1-5). The matrices were sub-jected to manual constrained sedation with theonshore to offshore distribution of localitiesmaintained (Brower & Kile 1988). The resulttherefore reflects an offshore deepeningpalaeoecological gradient. Generic biofacies,identified as blocks of taxa within the matrix,were named, and the included genera werecoded for biofacies for each time slice. Theassumption was made that a genus was morelikely to stay within the same biofacies in thesubsequent time slice. Anomalous occurrencesin the initial coding were corrected by a posteri-ori revision of the seriated matrix. The genericcoding is illustrated in Figure 6; biofacies diver-sity counts were made from this matrix for eachtime slice and region. The juxtaposition of bio-facies at each locality enables the constructionof a biofacies architecture diagram (Figs 4b, 5b,7) for each time slice and region. Biofaciesarchitecture can then be compared throughtime. There is no correlation of the shelf bio-facies with the described shelly invertebratebiofacies such as the substrate-controlledpalaeocommunities of the late Llanvirn - earlyCaradoc of the Welsh basin summarized byLockley(1983).

EUCONODONT DIVERSITY CHANGES 87
Fig. 1. Constrained seriation matrix for the Laurentian shelf and marginal terranes for each of the Ordoviciantime slices described in the text. Source of locality data: Oklahoma from Bauer (1994); Alabama fromBergstrom (1990); Girvan from Bergstrom (1990); Culzean from Armstrong (2000); Tweed from Armstrong(1997); velicuspis Chron data from Sweet & Bergstrom (1984); ordovidcus Biozone data from Nowlan &Barnes (1981) and Radcliffe (1998). Localities have been located onshore (to the left) to offshore withreference to palaeogeographical maps in Scotese & McKerrow (1990) and Bevins et al. (1992). Shaded boxesindicate a presence; biofacies for generic association in Llanvin-Caradoc and velicuspis Chron have beendefined by blocks of genera in the matrix. Biofacies were allocated for ordovidcus Biozone genera based ongeneric occurrences in previous time slices (see Fig. 6 and text for explanation).

88 H. A. ARMSTRONG & A. W. OWEN
Fig. 2. Constrained seriation matrix for the Avalonian shelf (Wales) for each of the time slices described in thetext. Source of locality data: (1) Savage & Bassett (1985): (2-7) Bergstrom & Orchard (1985), Bergstrom et al.(1987): (8) Savage & Bassett (1985): (9) Savage & Bassett (1985); (10) Orchard (1980). Bergstrom & Orchard(1985), Savage & Bassett (1985): (11) Bergstrom & Orchard (1985). Savage & Bassett (1985): (12-14) Orchard(1980), Bergstrom & Orchard (1985). Localities have been located onshore (to the left) to offshore withreference to Bevins et al. (1992). Shaded boxes indicate a presence: biofacies for generic associations havebeen defined by blocks of generic associations and occurrences in previous time slices (see Fig. 6 and text forexplanation).
Results taxa, which range onshore to offshore or in thedata matrix have ranges that terminate onshore.
Biofacies architecture These biofacies include taxa considered to rep-resent the North Atlantic Province (NAP) plus
Six biofacies have been identified. Oceanic Bio- many coniform genera. NAP taxa are known infacies (OBI, OB2 and OB3) contain pelagic both low and mid-temperate latitude sections
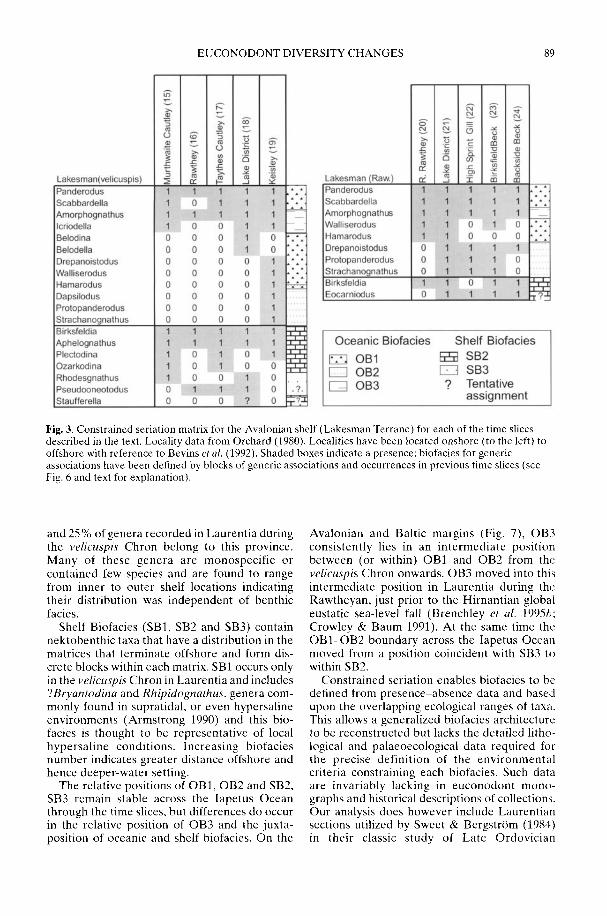
EUCONODONT DIVERSITY CHANGES 89
Fig. 3. Constrained seriation matrix for the Avalonian shelf (Lakesman Terrane) for each of the time slicesdescribed in the text. Locality data from Orchard (1980). Localities have been located onshore (to the left) tooffshore with reference to Bevins et al. (1992). Shaded boxes indicate a presence; biofacies for genericassociations have been defined by blocks of generic associations and occurrences in previous time slices (seeFig. 6 and text for explanation).
and 25% of genera recorded in Laurentia during Avalonian and Baltic margins (Fig. 7), OB3the velicuspis Chron belong to this province, consistently lies in an intermediate positionMany of these genera are monospecific or between (or within) OB1 and OB2 from thecontained few species and are found to range velicuspis Chron onwards. OB3 moved into thisfrom inner to outer shelf locations indicating intermediate position in Laurentia during thetheir distribution was independent of benthic Rawtheyan, just prior to the Hirnantian globalfacies. eustatic sea-level fall (Brenchley et al. 1995b;
Shelf Biofacies (SB1, SB2 and SB3) contain Crowley & Baum 1991). At the same time thenektobenthic taxa that have a distribution in the OB1-OB2 boundary across the Iapetus Oceanmatrices that terminate offshore and form dis- moved from a position coincident with SB3 tocrete blocks within each matrix. SB1 occurs only within SB2.in the velicuspis Chron in Laurentia and includes Constrained seriation enables biofacies to be?Bryantodina and Rhipidognathus, genera com- defined from presence-absence data and basedmonly found in supratidal, or even hypersaline upon the overlapping ecological ranges of taxa.environments (Armstrong 1990) and this bio- This allows a generalized biofacies architecturefacies is thought to be representative of local to be reconstructed but lacks the detailed litho-hypersaline conditions. Increasing biofacies logical and palaeoecological data required fornumber indicates greater distance offshore and the precise definition of the environmentalhence deeper-water setting. criteria constraining each biofacies. Such data
The relative positions of OB1, OB2 and SB2, are invariably lacking in euconodont mono-SB3 remain stable across the Iapetus Ocean graphs and historical descriptions of collections,through the time slices, but differences do occur Our analysis does however include Laurentianin the relative position of OB3 and the juxta- sections utilized by Sweet & Bergstrom (1984)position of oceanic and shelf biofacies. On the in their classic study of Late Ordovician

90 H. A. ARMSTRONG & A. W. OWEN
Fig. 4. (a)Constrained sedation matrices for the Baltica shelf for each of the time slices described in the text.Source of locality data for Estonia from Sweet & Bergstrom (1984); Sweden from Bergstrom (1990): Polandfrom Dzik (1994); Oslo from Smith (1999). Balto-Scandian locations are constrained onshore to offshore fromthe confacies belts proposed by Jaanusson (1976). (b) Biofacies architecture diagrams; note the intermediateposition of OB3 in the velicuspis Chron data.
euconodont biofacies. They defined biofacies three confacies belts in the Middle Ordovician ofbased upon cluster analysis assuming a priori the Appalachians, based upon shelly faunas andthat all euconodonts were pelagic. Their euconodonts. Their inshore Lee and Tazewellapproach clusters localities based on included belts contain almost identical euconodont faunaseuconodonts, grouping both pelagic and nekto- including Belodella, Belodina, Appalachig-benthic taxa. Constrained seriation enables the nathus, Curtognathus, Erismodus, Phragmodusseparation of pelagic and nektobenthic taxa and and Plectodina. This fauna contains a mixture ofour reinterpretation of only North Atlantic OB1 (coniform) and SB2 (non-coniform) taxa.Province and coniform taxa as pelagic makes The offshore Blount Belt contains the eucono-direct comparison with this work difficult. For donts of OB2, plus Protopanderodus (previouslyexample SB2 includes genera common with the Cordylodus) and 'Polyplacognathiis'. They alsoAphelognathus-Oulodus and Plectodina Bio- noted the similarity at species level of theirfacies of Sweet & Bergstrom (1984), SB3 has Blount Belt fauna with Baltoscandian faunas ofelements in common with the Phragmodus- the same age and the horizontal uniformity ofIcriodella-Plectodina Biofacies though is not a this fauna across various confacies belts. Both ofdirect correlative. OB2 has genera in common these observations support our contention thatwith the Hamarodus-Dapsilodus-Scabbardella these taxa were pelagic.(HDS) Biofacies and OB3 has taxa in common Stouge & Rasmussen (1996) identified thewith the Amorphognathus Biofacies, but neither HDS Biofacies in the lower Ashgill of Bornholmis directly comparable. and concluded that widespread geographical
Jaanusson & Bergstrom (1980) recognized distribution indicated that this faunal assemblage
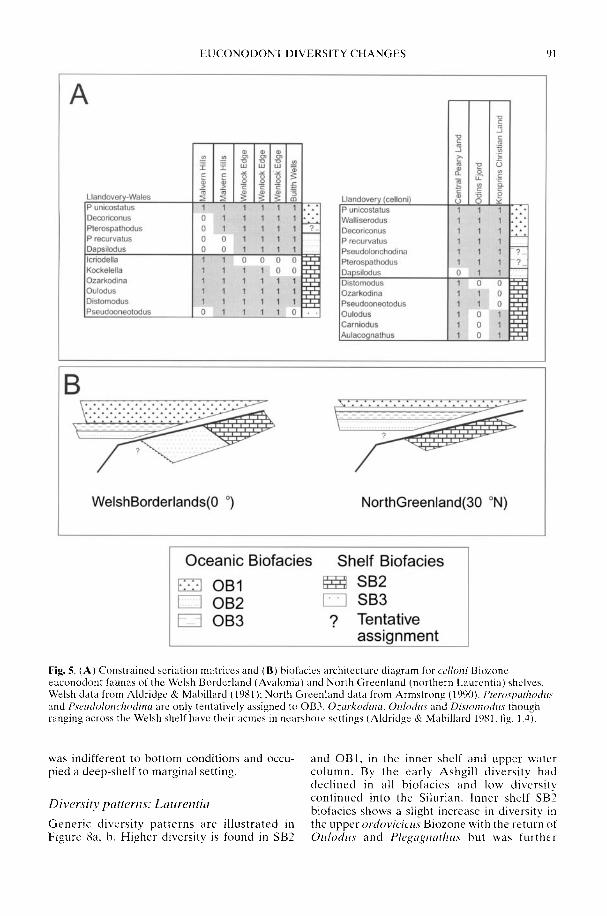
EUCONODONT DIVERSITY CHANGES 91
Fig. 5. (A) Constrained sedation matrices and (B) biofacies architecture diagram for celloni Biozoneeuconodont faunas of the Welsh Borderland (Avalonia) and North Greenland (northern Laurentia) shelves.Welsh data from Aldridge & Mabillard (1981); North Greenland data from Armstrong (1990). Pterospathodwand Pseudolonchodina are only tentatively assigned to OB3. Ozarkodina, Oulodus and Distomodus thoughranging across the Welsh shelf have their acmes in nearshore settings (Aldridge & Mabillard 1981. fig. 1.4).
was indifferent to bottom conditions and occu-pied a deep-shelf to marginal setting.
Diversity patterns: Laurentia
Generic diversity patterns are illustrated inFigure 8a, b. Higher diversity is found in SB2
and OB1, in the inner shelf and upper watercolumn. By the early Ashgill diversity haddeclined in all biofacies and low diversitycontinued into the Silurian. Inner shelf SB2biofacies shows a slight increase in diversity inthe upper ordovicicus Biozone with the return ofOulodus and Plegagnathus but was further

92 H. A. ARMSTRONG & A. W. OWEN
Fig. 6. Generic biofacies for each Ordovician timeslice. The assumption was made that a genus wasmore likely stay within the same biofacies in thesubsequent time slice. Anomalous occurrences in theinitial coding were corrected by a posteriori revisionof the seriated matrix, ? is retained for occurrenceswhere this could not be achieved.
reduced into the Early Silurian. This increasemay be a sampling artefact. The Ordoviciandecline in diversity continued for longer in thedeeper-water SB3 and OB2 biofacies.
Diversity patterns: Avalonia
Generic diversity patterns are illustrated inFigure 8c, d. Higher diversity is found in SB2 andOB1. in the inner shelf and upper water column,as found in the Laurentia data. Diversitydeclined in all biofacies during the mid-Ashgill.coincident with the northward drift of Avaloniain tropical latitudes (Fig. 9). Low diversitycontinued into the early Silurian in oceanicbiofacies whilst diversity increased in shelffaunas. Shelf faunas in the Early Siluriancontained a mixture of incumbents (Icriodellaand Ozarkodina), recruits (including Oulodus}.which had an evolutionary origin in theLaurentian inner shelf, plus Kockelella andDistomodus, new genera with cryptic ancestry.Diversity trajectories in shelf and oceanic bio-facies are parallel and the decline in diversitycontinued for longer in the oceanic biofacies.implying the causal mechanism persisted forlonger in deeper water and affected taxa in allhabitats.
Diversity patterns: Baltica
Generic diversity (Fig. 8e, f) is low in the Balticsuccession, comparable to that in Avalonia, andthe paucity of data makes interpretation difficult.SB2 and SB3 biofacies show slight rises in diver-sity into the early Ashgill, OB1 remains staticand OB2 shows a slight decline. Llandoverydiversity is higher than in the early Ashgill inshelf biofacies and is lower in oceanic biofacies.Diversity patterns in Baltica most closelycompare with those from Avalonia.
Diversity in the early Silurian
Our analysis is restricted to the celloni Biozoneof Avalonia (Welsh Borderland data fromAldridge & Mabillard 1981) and a comparativesection from northern Laurentia (NorthGreenland data from Armstrong 1990; thoughnot part of the lapetus margin at this time thesedata are taken as typical for the Laurentianshelf). A full review of Silurian successions isbeyond the scope of this paper and the IapetusOcean was virtually closed by the Early Silurian(Armstrong & Owen 2001). Oceanic and shelfbiofacies identical to those in the Ordoviciancan be identified in this limited dataset (Fig. 5).Oceanic biofacies comprise predominantlyconiform taxa. Pterospathodus and Pseudolon-chodina are tentatively assigned to OB3 as bothrange across the Greenland shelf. Pseudolon-chodina is not present in the Welsh dataset.Ozarkodina, Oulodus and Distomodus also

EUCONODONT DIVERSITY CHANGES 93
Fig. 7. Biofacies architecture diagrams for Laurentia and Avalonia at each of the Ordovician time slices. SeeFigure 1 for key.
range across the Welsh shelf but Aldridge &Mabillard (1981) recognized these taxa hadtheir acmes in nearshore settings and declinedrapidly in abundance offshore. This patternsuggests that these genera included wide-ranging, nektobenthic, ecological generaliststhat preferred nearshore habitats. Icriodella isrestricted to SB2 in the Silurian and OB3 in theOrdovician. This suggests either a change froma pelagic mode of life, or that Silurian andOrdovician Icriodella are not congeneric.Silurian nektobenthic genera appear to havemuch broader ecological ranges than theirOrdovician counterparts.
Interpretation
Biofacies architecture
In the modern ocean planktonic and pelagicspecies inhabit depth-related water masses
defined by oxygen content, salinity, temperatureand nutrient availability, comparable to bio-facies in continental shelf settings, though oftenof much larger geographical extent. Water masscharacter is partly set by the shape of the basin,the direction of surface winds and patterns ofevaporation and precipitation over the oceans(Emery & Meincke 1986). In general termstemperature and salinity fall with depth whilstnutrients increase (see review in Norris 2000).Specific water masses and their included faunacan move both vertically and horizontallydepending upon oceanography and climaticfactors, and are divided by physiochemicalsurfaces along which species or prey congregatefor growth or reproduction (Norris 2000).Vertical movements of water masses occur as aresult of changes in temperature and density andare most marked in thermally stratified oceansin continental margin upwelling zones. Thestructure of the upwelling zone off the SW
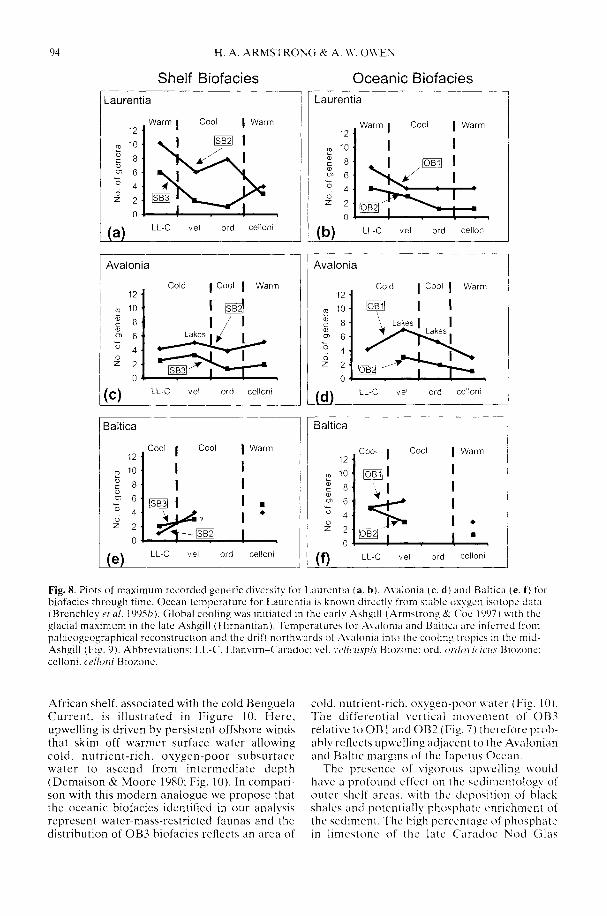
94 H. A. ARMSTRONG & A. W. OWEN
Fig. 8. Plots of maximum recorded generic diversity for Laurentia (a. b). Avalonia (c, d) and Baltica (e, f) forbiofacies through time. Ocean temperature for Laurentia is known directly from stable oxygen isotope data(Brenchley et al. 1995b), Global cooling was initiated in the early Ashgill (Armstrong & Coe 1997) with theglacial maximum in the late Ashgill (Hirnantian). Temperatures for Avalonia and Baltica are inferred frompalaeogeographical reconstruction and the drift northwards of Avalonia into the cooling tropics in the mid-Ashgill (Fig. 9). Abbreviations: LL-C, Llanvirn-Caradoc: vel. vclicuspis Biozone: ord. ordovicicus Biozone:celloni, celloni Biozone.
African shelf, associated with the cold BenguelaCurrent, is illustrated in Figure 10. Here,upwelling is driven by persistent offshore windsthat skim off warmer surface water allowingcold, nutrient-rich, oxygen-poor subsurfacewater to ascend from intermediate depth(Demaison & Moore 1980; Fig. 10). In compari-son with this modern analogue we propose thatthe oceanic biofacies identified in our analysisrepresent water-mass-restricted faunas and thedistribution of OB3 biofacies reflects an area of
cold, nutrient-rich, oxygen-poor water (Fig. 10).The dif ferent ia l vertical movement of OB3relative to OB1 and OB2 (Fig. 7) therefore prob-ably reflects upwelling adjacent to the Avalonianand Baltic margins of the Iapetus Ocean.
The presence of vigorous upwelling wouldhave a profound effect on the sedimentology ofouter shelf areas, with the deposition of blackshales and potentially phosphate enrichment ofthe sediment. The high percentage of phosphatein limestone of the late Caradoc Nod Glas

EUCONODONT DIVERSITY CHANGES 95
Fig. 9. Palaeolatitudes for East Avalonia (based onMacNiocail 2000). These data indicate a closure rateof c.3 cm a-1. Timescale from Tucker & McKerrow(1995). Abbreviations: BR, Browgill Redbeds; BV,Builth Volcanics; MC, Mill Cove Redbeds; MV,Mendips Volcanics; NB, North Builth; Sv, StapleyVolcanics; TL, Tortworth Lavas; Trv, TreffgarneVolcanics; TV, Tramore Volcanics.
Fig. 10. Water mass structure off the SW Africanshelf based upon percentage oxygen content of thewater (based on Demaison & Moore 1980).Upwelling water is generated by onshore to offshorewinds skimming off warm surface water and thusallowing cold, nutrient-rich water to well up fromintermediate depths. A similar structure can be foundoff the Peruvian margin associated with theHumboldt Current (Demaison & Moore 1980). Wehypothesize that oceanic euconodont oceanicbiofacies reflect water masses and have superimposedthese biofacies onto the water mass structure of theSW African shelf. The pattern generated is similar tothat found in the biofacies architecture diagram forAvalonia (Llanvirn-Ashgill) and Laurentia(ordovicicus Biozone) in Figure 7.
Formation of mid-Wales (Cave 1965; Smith1999) and thin phosphorite conglomerate atthe base of the Venst0p Formation in theOslo-Asker district (Williams & Bruton 1983;Owen et al 1990) indicates that the water alongthe southern margin of the lapetus Ocean wasrich in nutrients and supports the upwellinghypothesis.
Upwelling persisted along the southernmargin of the lapetus Ocean at least from thelate Caradoc-early Ashgill and perhaps as earlyas the uppermost Tremadoc or early Arenig(Lindstrom & Vortisch 1983) Upwelling appearsnot to have been initiated at the low-latitudeLaurentian margin until the Rawtheyan, justpredating the glacial maximum and coincidentwith the general upward movement of oceanicbiofacies across the lapetus Ocean.
Regional diversity trajectories
A major reduction in diversity in Laurentianbiofacies correlates with the early Ashgill onsetof ocean cooling along the Laurentian margin(Armstrong & Coe 1997) and represents theextinction of taxa adapted to warm, tropicalconditions. A second decline in diversity in SB2correlates with a return to greenhouse con-
ditions, and extinctions in taxa interpreted asbeing adapted to cooler 'glacial' conditions (seealso Brenchley 1988; Brenchley et al 1995a).
The major decline in diversity in Avalonianbiofacies was coincident with the drift of Ava-lonia into warmer (but cooling) tropical water(Figs 8, 9). This suggests extinction of taxaadapted to cold water conditions, introducedinto cooling but none-the-less warmer water ofthe tropics.
Baltica moved from intermediate latitudes tothe tropics from the Arenig to the Ashgill andsutured to Avalonia during the late Ordovican(see reviews in Cocks & Fortey 1998; Cocks2001). The Late Ordovician euconodont diver-sity trajectories (Figs 8e-f) are more closelysimilar to those of Avalonia, suggesting similarunderlying controls. The onset of upwelling andclose proximity of Avalonia and Baltica in thelate Caradoc-early Ashgill corresponds with arise in diversity largely the consequence ofmigrant shelf taxa from Avalonia.
Conclusions
Constrained seriation of presence-absencematrices provides a method of qualitativelydefining generic associations or biofacies and

96 H. A. ARMSTRONG & A. W. OWEN
hence reconstructing biofacies architectureswithout recourse to extensive recollecting.Upper Ordovician euconodonts can be dividedinto shelf and oceanic biofacies by this method.Euconodonts have traditionally been con-sidered to be markedly provincial during theLate Ordovician with warm, tropical-water taxarestricted to the American MidcontinentProvince and cooler-water taxa representativeof the North Atlantic Province. Our analysisindicates that many genera previously con-sidered to be representative of the NorthAtlantic Province had a pelagic mode of life andranged widely across the Iapetus Ocean. Thisinterpretation challenges the accepted viewof euconodont provinciality and supportsthe province biofacies model introduced byRasmussen (1998) for early Llanvirn eucon-odonts of the lapetus Ocean.
Oceanic biofacies are considered to havereflected water mass structure. The changingvertical distribution of OB3 biofacies taxasuggests that they were adapted to cold, nutrient-rich, oxygen-poor upwelling water. If this isconfirmed then the southern margin of thelapetus Ocean was characterized by upwellingcurrents during the Ashgill. Upwelling wasinitiated along the Laurentian margin during thelate Ashgill, coincided with an upward move-ment of oceanic biofacies and just predates theglacial maximum. Biofacies architecture patternsestablished in the Late Ordovician appear tohave remained into the Early Silurian.
Clade diversities and trajectories in separatebiofacies and latitudes can be attributed todifferent causal mechanisms. In Laurentia, themaximum diversity decline in all biofaciesoccurred in the early Ashgill and was coincidentwith the onset of ocean cooling. Maximumdiversity decline in Avalonia coincided with itsnorthward drift into tropical latitudes. Thestability of euconodont biofacies architectureduring Late Ordovician global cooling and platereorganization indicates that these events hada low palaeoecological impact, causing littledisruption of marine habitats despite a substan-tial decrease in both alpha and beta diversity(see also Droser et al. 1997, 2000).
Late Ordovician euconodont diversityreflects the unique environmental conditionsspecific to a region and not intrinsic propertiesof the clade. The post-extinction recovery ofeuconodonts was largely restricted to nekto-benthic shelf taxa. The failure of many long-ranging pelagic clades to recover from thelatest-Ordovician mass extinction suggestsintrinsic properties of these clades may havesuppressed their rediversification.
Our observations have implications for thealpha taxonomy of euconodonts. Genetic evi-dence for numerous cryptic species in morpho-logically identical pelagic organisms is consistentwith the idea that they are much more specializedthan would be inferred from their geographicaldistributions (Norris 2000). We may now have tofundamentally revise our morphological speciesconcepts for pelagic euconodont clades that arelikely to have been much more diverse thanhas been generally accepted in tradit ionaltaxonomies.
The authors acknowledge funding from the NERC(GR3/11834 to A.W.O.rGR9/02834 to H.A.A. andA.W.O.), The comments provided by the refereesM. P. Smith and J. A. Rasmussen have improved thispaper which is a contribution to IGCP 410. "The GreatOrdovician Biodiversification E v e n t .
ReferencesA I . D R I D G F . R. J. & M A B I L L A R D , J. E. 1981. Local vari-
ations in the distribution of Silurian conodonts: anexample from the amorphognathoides interval ofthe Welsh Basin. In: NFALE, J. W. & BRASIeR. M.D. (eds) Microfossils from Recent and Fossil ShelfSeas Ellis Horwood, Chichester. 10-18.
ARMSTRONG. H. A. 1990. Conodonts from the LowerSilurian of the North Greenland carbonate plat-form. Bulletin Gronlands Geologiske Under-s gelse. 159
ARMSTRONG. H. A. 1995. High-resolution bio-stratigraphy (conodonts and graptolites) of theUpper Ordovician and Lower Silurian - achronometric framework in which to evaluate thecauses of the mass extinction. Modern Geologv,20. 1-28.
ARMSTRONG. H. A. 1997. Conodonts from the ShinnelFormation. Tweeddale Member (middle Ordo-vician). Southern Uplands. Scotland. Palaeontol-ogy 40.763-799.
ARMSTRONG. H. A. 2000. Conodont micropalaeon-tology of mid-Ordovician aged limestone clastsfrom LORS conglomerates. Lanark and Strath-more basins. Midland Valley. Scotland. Journal ofMicropalaeontologv. 19. 45-59.
ARMSTRONG. H. A. & COF. A. L. 1997. Deep sea sedi-ments record the geophysiology of the endOrdovician glaciation. Journal of the GeologicalSociety London, 154. 929-934.
ARMSTRONG. H. A. & OWEN. A. W. 2001. Terraneevolution of the paratectonic Caledonides ofnorthern Britain. Journal of the GeologicalSociety, London. 158.475-486.
BARNES. C. R.. REXROAD. C. B. & MILLER. J. F. 1973.Lower Paleozoic provincialism. In: RHODES. F. H.T. (ed.) Conodont Zoology. Geological Society ofAmerica, Special Papers. 141. 156-190.
BAUER. J. A. 1994. Conodonts from the BromideFormation (Middle Ordovician. south-centralOklahoma. Journal of Paleontology, 68. 358-376.

EUCONODONT DIVERSITY CHANGES 97
BERGSTROM, S. M. 1990. Biostratigraphie and bio-geographic significance of Middle and UpperOrdovician conodonts in the Girvan succession,South-West Scotland. Courier Forschungs Insti-tute Senckenberg, 118,1-43.
BERGSTROM, S. M. & ORCHARD, M. 1985. Conodontsof the Cambrian and Ordovician Systems fromthe British Isles. In: HIGGINS, A. C. & AUSTIN, R.L. (eds) A Stratigraphical Index of Conodonts.Ellis Horwood, Chichester, 32-68.
BERGSTROM, S. M., RHODES, F. H. T. & LINDSTROM,M. 1987. Conodont biostratigraphy of theLlanvirn-Llandeilo and Llandeilo-CaradocSeries boundaries in the Ordovician System ofWales and the Welsh Borderland. In: AUSTIN, R.L. (ed.) Conodonts: Investigative Techniques andApplications. Ellis Horwood, Chichester,294-316.
BERNER, R. A. 1990. Atmospheric carbon dioxidelevels over Phanerozoic time. Science, 249,1382-1386.
BERNER, R. A. 1992. Palaeo-CO2 and climate. Nature,358,114.
BEVINS, R. E., BLUCK, B. J., BRENCHLEY, P. J., FORTEY,R. A., HUGHES, C. P., INGHAM, J. K. & RUSHTON,A. W. A. 1992. Ordovician. In: COPE, J. W. C.,INGHAM, A. J. K. & RAWSON, P. F (eds) Atlas ofPalaeogeography and Lithofacies. GeologicalSociety, London, Memoirs, 13,19-36.
BRENCHLEY, P. J. 1988. Environmental changes closeto the Ordovician-Silurian boundary. Bulletin ofthe British Museum of Natural History (Geology),43, 377-385.
BRENCHLEY, P. J., GARDEN, G. A. F. & MARSHALL, J.D. 1995a. Environmental changes associated withthe "first strike' of the late Ordovician massextinction. Modern Geology, 20, 69-82.
BRENCHLEY, P. J., MARSHALL, J. D., CARDEN, G. A. F.,ET AL. 1995b. Bathymetric and isotopic evidencefor a short-lived late Ordovician glaciation in agreenhouse period. Geologv, 22, 295-298.
BROWER, J. C. & KILE, K. M. 1988. Seriation of anoriginal data matrix as applied to paleoecology.Lethaia, 21, 79-92.
CAVE, R. 1965. The Nod Glas sediments of Caradocage in North Wales. Geological Journal, 4,278-298.
COCKS, L. R. M. 2001. Ordovician and Silurian globalgeology. Journal of the Geological Society,London, 158,197-210.
COCKS, L. R. M. & FORTEY, R. A. 1998. The LowerPalaeozoic margins of Baltica. GeologiskaForeningens i Stockholm Forhandlingar, 120,173-179.
CROWLEY, T. J. & BAUM, S. K. 1991. Towards reconcil-iation of Late Ordovician (~440Ma) glaciationwith very high CO2 levels. Journal of GeophysicalResearch, 96, 22597-22610.
DEMAISON, G. J. & MOORE, G. T. 1980. Anoxicenvironments and oil source bed genesis. AAPGBulletin, 64, 1179-1209.
DROSER, M. L., BOTTJER, D. J. & SHEEHAN, P. M. 1997.Evaluating the ecological architecture of majorevents in the Phanerozoic history of marine inver-tebrate life. Geology, 25, 167-170.
DROSER, M. L., BOTTJER, D. J., SHEEHAN, P. M. &McGEE, G. R. JR. 2000. Decoupling of taxonomicand ecological severity of Phanerozoic marinemass extinctions. Geology, 28, 675-678.
DZIK, J. 1994. Conodonts of the Mojcza Limestone.Palaeontologica Polonica, 53, 43-128.
EMERY, W. J. & MEINCKE, J. 1986. Global watermasses: summary and review. Oceanologica Acta,9,383-391.
ERWIN, D. H. 1998. After the end: Recovery fromextinction. Science, 279, 1324-1325.
HAMBREY, M. J. 1985. The Late Ordovician-Silurianglacial period. Palaeogeography, Palaeoclima-tology, Palaeoecology, 51, 273-289.
JAANUSSON, V. 1976. Faunal dynamics in the MiddleOrdovician (Viruan) of Balto-Scandia. InBASSETT, M. G. (ed.) The Ordovician System,Proceedings of a Palaeontological AssociationSymposium. Birmingham, September 1974. University of Wales and National Museum Wales.Cardiff, 301-326.
JAANUSSON, V. & BERGSTROM, S.M. 1980. MiddleOrdovician faunal spatial differentiation inBaltoscandia and the Appalachians. Alcheringa,4,89-110.
JABLONSKI, D. 1998. Geographic variation in themolluscan recovery from the end-CretaceousExtinction. Science, 279, 1327-1329.
KEMP, T. S. 1999. Fossils and Evolution. Oxford Uni-versity Press, Oxford.
LINDSTROM, M. & VORTISCH W. 1983. Indicationsof upwelling in the Lower Ordovician ofScandinavia. In: THIEDE, J. & SUESS, E. (eds)Coastal Upwelling. Plenum, New York, 535-551.
LOCKLEY, M. G. 1983. A review of brachiopod domi-nated palaeocommunities from the type Ordo-vician. Palaeontology, 26, 111-145.
MACNIOCAILL, C. 2000. A new Silurian palaeolatitudefor eastern Avalonia and evidence for crustalrotations in the Avalonian margin of southwest-ern Ireland. Geophysical Journal International,141,661-671.
MILLER, A. I. 1997. Comparative diversificationdynamics among palaeocontinents during theOrdovician Radiation. Geobios Memoire Special,20, 397-406.
MILLER, A. I. 2000. Conversations about Phanerozoicglobal diversity. In: ERWIN, D. H. & WING, S. L.(eds) Deep Time: Paleobiology's Perspective.Paleobiology (Supplement), 26, 53-74.
MILLER, A. I. & MAO, SHUGUANG. 1995. Associationof erogenic activity with the Ordovician radiationof marine life. Geology, 23, 305-308.
NORRIS, R. D. 2000. Pelagic species diversity, biogeog-raphy and evolution. In: ERWIN, D. H. & WING, S.L. (eds) Deep Time: Paleobiology's Perspective.Paleobiology (Supplement), 26, 236-259.
NOWLAN, G. S. & BARNES, C. R. 1981. Late Ordovicianconodonts from the Vaureal Formation, AnticostiIsland, Quebec. Geological Survey of CanadaBulletin, 329, 1-49.
ORCHARD, M. 1980. Upper Ordovician conodontsfrom England and Wales. Geologica et Palaeonto-logica, 14, 9-44.

98 H. A. ARMSTRONG & A. W. OWEN
OWEN. A. W., BRUTON. D. L.. BOCKELIE, J. F. & BOCK-ELIE. T. G. 1990. The Ordovician successions ofthe Oslo Region, Norway. Norges GeologiskeUnders0kelese, Special Publications, 4.
OWEN. A. W. & ROBERTSON. D. B. R. 1995. Ecologicalchanges during the end-Ordovician extinction.Modern Geology. 20, 21-39.
RADCLIFFE, G. 1998. Biotic Recovery of ConodonisFollowing the end-Ordovician Mass Extinction.PhD thesis. University of Durham.
RASMUSSEN. J. A. 1998. A reinterpretation of theconodont Atlantic Realm in the late EarlyOrdovician (early Llanvirn). In: SZANIAWSKI. H.(ed.) Proceedings of the Sixth European Con-odont Symposium (ECOS VI). PalaeontologicaPolonica. 58, 67-77.
SAVAGE. N. M. & BASSETT, M. G. 1985.Caradoc-Ashgill conodont faunas from Walesand the Welsh Borderland. Palaeontology. 28,679-715.
SCOTESE. C. R. & MCKERROW, W. S. 1990. Revisedworld maps and introduction. In: MCKERROW,W. S. S. & SCOTESE. C. R. (eds) PalaeozoicPalaeogeography and Biogeography. GeologicalSociety, London, Memoirs, No. 12, 1-25.
SEPKOSKI J. J. JR 1993. Ten years in the library: Newdata confirm paleontological patterns. Paleobiol-ogy, 19,43-51.
SEPKOSKI J. J. JR & KENDRICK, D. C. 1993. Numericalexperiments with model monophyletic and para-phyletic taxa. Paleobiology, 19,168-184.
SMITH, C. J. 1999. Evolutionary Palaeobiology of
Deep-water Conodonts. PhD thesis. University ofDurham.
STOUGE. S. & RASMUSSEN. J. A. 1996. Upper Ordo-vician conodonts from Bornholm and possiblemigration routes into the Palaeotethys Ocean.Bulletin of the Geological Society of Denmark. 43.54-67.
SWEET, W. C. & BFRGSTROM, S. M. 1974. Provincialismexhibited by Ordovician conodont faunas. Societyof Economic Paleontologists and Mineralogists.Special Publications. 21. 189-202.
SWEET. W. C. & BFRGSTROM. S. M. 1984. Conodontprovinces and biofacies of the Late Ordovician.Geological Society of America, Special Papers.196. 69-87.
TICKER. R. D. & MCKERROW. W. S. 1995. EarlyPaleozoic chronology: a review in light of new U-Pb zircon ages from Newfoundland and Britain.Canadian Journal of Earth Sciences. 32. 368-379.
VAN STAAL, C. R., DEWEY. J. F., MAC NIOCIALL. C. &MCKERROW, W. S. 1998. The Cambrian-Siluriantectonic evolution of the northern Appalachiansand British Caledonides: history of complex, westand southwest Pacific-type segment of lapetus.In: BLUNDELL. D. J. & Scon. A. C. (eds) Lyell:the Past is the Key to the Present. GeologicalSociety, London. Special Publications, 143.199-242.
WILLIAMS, S. H. & BRITON. D. L. 1983. TheCaradoc-Ashgill boundary in the central OsloRegion and associated graptolite faunas. NorskGeologisk Tidsskrift.23. 147-191.

The role of pyroclastic volcanism in Ordovician diversification
JOSEPH P. BOTTINGDepartment of Earth Sciences, University of Cambridge, Downing Street, Cambridge
CB2 3EQ, UK (e-mail: [email protected])
Abstract: Pyroclastic volcanism has been tentatively correlated with Ordovician faunaldiversification, but it is unclear whether the volcanism itself or associated tectonic effectswere more influential Elevated nutrient flux from uplifted areas has been widely discussed,but the link between enhanced productivity and diversification is unclear. Ecosystem com-partmentalization due to irregular topography was probably significant, but it is debatablewhether this was sufficient to explain the entire diversification. This paper introduces afurther volcanic process, which may have been more significant than these establishedhypotheses, although it is emphasized that all relevant factors acted in combination. Recentstudies of the local ecological effects of ash-fall have revealed dramatic post-depositionalplankton and benthic blooms associated with overturning of a stratified water mass. Thedata are consistent through all studied sections from the Builth Inlier, Central Wales, whiledeeper-water sediments showed plankton blooms only. Repeated bloom events overlocalized dysaerobic shelf areas may have produced temporary benthic 'islands', enhancedgenetic heterogeneity within affected regions, and increased speciation rates. In order tomaximize bloom intensity (and possible effects on evolution), this process requires wide-spread pyroclastic volcanism combined with extensive shelf areas with well-developedstratification. The hypothesis potentially allows explanation of the differential rates ofevolution between different groups, particularly the decoupled planktic-benthic patterns.A preliminary test using Welsh Basin ostracodes is presented, with predictions for theglobal database results.
The Ordovician Period contained the mostdramatic, sustained increases in global diversityduring the Phanerozoic, but its cause (or causes)remains elusive. Unlike most other episodes ofdiversification, it did not follow a major extinc-tion event. There are marked differencesregarding the onset and extent of diversitychanges between palaeocontinents, and a strongecological influence. The majority of the diver-sity increase is accommodated by sessile (andrelatively immobile) benthos, and manifestedpartly as alpha and sometimes gamma diversity(Miller 1997a; Patzkowsky 1995). Beta (inter-community) diversity may even have decreasedduring the Late Ordovician (Miller 1997b).Gamma (inter-regional) diversity increases aregenerally considered to have been limited(Sepkoski 1988), despite the hypothesizedhabitat-fragmentation influence of numerousseparate continents and volcanic islands (but seePatzkowski 1995). Sepkoski (1988) suggestedthat much of the Ordovician diversity increasewas incorporated through the origination ofnovel, biogenic subenvironments such as reefs,bioherms and crinoid thickets. Data presentedby McCormick & Owen (2001), describingOrdovician trilobite diversity in the WelshBasin, show a significant increase in the species:genus ratio during diversity peaks. Genus-level
data are more usually employed in diversityanalyses, but the species approach should beencouraged where possible, since the diversifi-cation operated primarily at low taxonomiclevels. However, the known fossil record ofseveral classes (particularly sessile epifauna)begins in the Ordovician, suggesting some high-level taxonomic origination (Conway Morris1999). This is unlikely to be due entirely to theorigination of preservable skeletons in previ-ously soft-bodied lineages. The relationshipsbetween the different diversity measures arepoorly understood (e.g. Adrain et al. 2000), andthe type of diversification observed (alpha, betaor gamma) may also depend on the taxonomiclevel examined. A similar pattern is apparent inthe post-Permian recovery, in which high-leveltaxonomy showed few changes, with diversifica-tion concentrated at family level and below(Erwin et al 1987).
The primary difficulty in interpreting theOrdovician Radiation (i.e. extreme complexity)may also allow its ultimate solution. Differentialdiversity patterns occur between differentregions and taxonomic groups, and there areputative correlations with changes in climate,sea level, palaeogeography and volcanism. Allthese factors probably affected diversity, inaddition to internal, ecological interactions: to
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations, Geological Society, London, Special Publications, 194, 99-113.0305-8719/02/$15.00 © The Geological Society of London 2002.

100 JOSEPH P. BOTTING
some extent, origination of novel life-habitsprovides additional subsidiary niches, mostobviously in the development of epifaunaltiering. Only through understanding detaileddiversity patterns can these influences possiblybe separated, a procedure that has been advo-cated strongly by Miller (1997a, 1998, 2000).
The approach of Sepkoski (e.g. 1993) andmost others to global diversity changes has beenthat of large-scale datasets, at intermediatetaxonomic levels (usually family or genus), butalso primarily in undivided continental orglobal format. Unless specifically utilized insubsets, global databases will generally provideonly broad patterns, where the influence of eachfactor is disguised by its combination withothers. Complex patterns of local and regionalmigration also confuse data on precise geo-graphic origins; it has not yet been shownwhether high-diversity, stable late Ordoviciancommunities occupied the same environmentsas the centres of diversification, or whether thedevelopment of long-term global diversity isindependent of areas where rapid speciationand overturn take place. Swain (1996), forexample, described exceptionally rapid ostra-code turnover in a bentonite-rich succession,where diversity at the top of the section did notgreatly exceed the initial, but standing diversityat intermediate levels was much higher. Thus, inorder to recognize meaningful diversificationpatterns reliably, overall diversity changes mustbe examined on regional or even intra-basinalscales (Miller 2000). Then, to identify thesignificance of any given process, a preciseecological-evolutionary signature must beestablished, where possible, and then tested.This is best examined by testing hypotheses inlimited regions, under the specific conditionsrequired for the testing. If a meaningful effect isobserved, locally or regionally, then large-scaledatabases (e.g. Owen & McCormick 1999) canallow assessment of the wider significance ofestablished local effects. Although many factorsmay have acted simultaneously in promotingthe Ordovician Radiation, specific predictionsof major regional processes should be recogniz-able on larger scales. Indeed, the ultimatechallenge in understanding the OrdovicianRadiation will be the extrapolation of knownsmall-scale patterns into a global situation,where interactions between regions and pro-cesses will require complex palaeogeography-based analyses.
Among the many possible influences onOrdovician diversity, the apparent correlationwith global pyroclastic volcanism has retained aperennial but uncertain significance. Miller
(1997a) provided a correlation with globaltectonic/volcanic intensity, in which Palaeozoicstanding diversity appeared approximately tomatch levels of orogenic activity. However,correlation of this nature may represent arelationship between standing diversity andinstantaneous tectonism, or between rapidspeciation and tectonism plus another factor,which was restricted to the Ordovician.Extreme fluctuations in tectonism during thePermian, in a relatively stable part of the diver-sity curve, and the lack of correlation in theMesozoic and Tertiary, lend some support to thelatter; the correlation is much more precise forthe Ordovician than for subsequent periods.Similarly, Miller & Mao (1995) presented a geo-graphical correlation of diversification centreswith forearc basins. Assuming some causalrelationship, it is unclear whether the volcanismitself or associated tectonic activity was themore significant influence on diversity. Miller(1997a) suggested habitat partitioning as aprimary cause of diversification, in addition toconsidering substrate transitions and nutrients,while Swain (1996) preferred an increase innutrient supply to explain ostracode diversifica-tion following major ash-falls. Increased nutri-ent supply from run-off in uplifted areas isperhaps inevitable, but the possible link todiversification is unclear; in modern oceans,abundant nutrients generally encourage lowdiversity and very high dominance (e.g. Boyd etal. 2000). In particular, a regional increase innutrient availability would tend to homogenizesubfacies variations for which scarcity of foodsupply was critical for their definition. Para-patric or sympatric speciation based largely onbehavioural food-acquisition procedures is.intuitively, more likely to arise in situationswhere food sources are relatively rare andcompetition most intense.
This chapter introduces a mechanism bywhich pyroclastic volcanism could have directlyinfluenced the speciation rate on regional scales,with predictions for the evolutionary patterns tobe expected. The available global data sub-divided by taxonomy (e.g. Sepkoski 1995)appear to be consistent with these predictionsregarding the differences in diversity changesbetween ecological groups. The hypothesispotentially allows explanation of the differentialevolutionary rates between ecological/taxo-nomic groups, particularly the decoupledplanktic-mobile benthic-sessile benthic diver-sity curves. A preliminary test of the hypothesisis included, using Welsh Basin ostracodes, andsimilar methods are encouraged for thoseinvestigating other hypotheses.
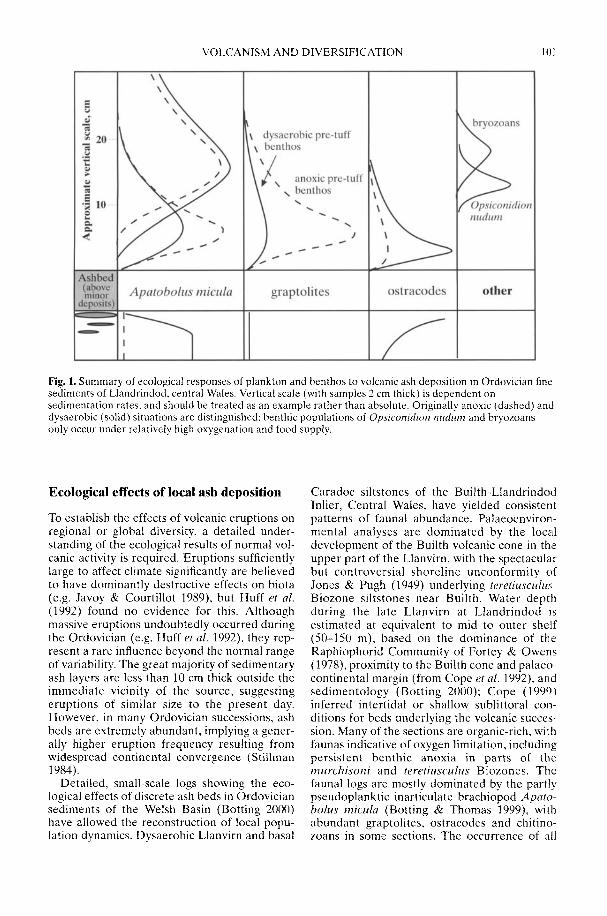
VOLCANISM AND DIVERSIFICATION 101
Fig. 1. Summary of ecological responses of plankton and benthos to volcanic ash deposition in Ordovician finesediments of Llandrindod, central Wales. Vertical scale (with samples 2 cm thick) is dependent onsedimentation rates, and should be treated as an example rather than absolute. Originally anoxic (dashed) anddysaerobic (solid) situations are distinguished; benthic populations of Opsiconidion nudum and bryozoansonly occur under relatively high oxygenation and food supply.
Ecological effects of local ash deposition
To establish the effects of volcanic eruptions onregional or global diversity, a detailed under-standing of the ecological results of normal vol-canic activity is required. Eruptions sufficientlylarge to affect climate significantly are believedto have dominantly destructive effects on biota(e.g. Javoy & Courtillot 1989), but Huff et al.(1992) found no evidence for this. Althoughmassive eruptions undoubtedly occurred duringthe Ordovician (e.g. Huff et al 1992), they rep-resent a rare influence beyond the normal rangeof variability. The great majority of sedimentaryash layers are less than 10 cm thick outside theimmediate vicinity of the source, suggestingeruptions of similar size to the present day.However, in many Ordovician successions, ashbeds are extremely abundant, implying a gener-ally higher eruption frequency resulting fromwidespread continental convergence (Stillman1984).
Detailed, small-scale logs showing the eco-logical effects of discrete ash beds in Ordoviciansediments of the Welsh Basin (Botting 2000)have allowed the reconstruction of local popu-lation dynamics. Dysaerobic Llanvirn and basal
Caradoc siltstones of the Builth-LlandrindodInlier, Central Wales, have yielded consistentpatterns of faunal abundance. Palaeoenviron-mental analyses are dominated by the localdevelopment of the Builth volcanic cone in theupper part of the Llanvirn, with the spectacularbut controversial shoreline unconformity ofJones & Pugh (1949) underlying teretiusculusBiozone siltstones near Builth. Water depthduring the late Llanvirn at Llandrindod isestimated at equivalent to mid to outer shelf(50-150 m), based on the dominance of theRaphiophorid Community of Fortey & Owens(1978), proximity to the Builth cone and palaeo-continental margin (from Cope et al. 1992), andsedimentology (Botting 2000); Cope (1999)inferred intertidal or shallow sublittoral con-ditions for beds underlying the volcanic succes-sion. Many of the sections are organic-rich, withfaunas indicative of oxygen limitation, includingpersistent benthic anoxia in parts of themurchisoni and teretiusculus Biozones. Thefaunal logs are mostly dominated by the partlypseudoplanktic inarticulate brachiopod Apato-bolus micula (Botting & Thomas 1999), withabundant graptolites, ostracodes and chitino-zoans in some sections. The occurrence of all

102 JOSEPH P. BOTTING
Fig. 2. Idealized distribution of oxygenation effects around ash-fall deposits in a shelf region with immobilewater mass: central area of benthic oxygenation by strong overturning, and deoxygenation in lateral regionsdue to partial overturning and plankton bloom. Anoxic regions begin to disperse following decline of planktonbloom (one year), allowing immigration of benthos from exterior.
other faunal elements was noted, although datawere usually insufficient for meaningful abun-dance comparisons. The complex overallpatterns are consistent throughout all studiedsections, minimizing the possibility of an arti-ficial distribution. A generalization of the resultsis shown in Figure 1 and explained below. Fulldetails of these data, including deeper-waterfacies from which benthic blooms were absent,will be published elsewhere, with full interpre-tation, and are also available in Botting (2000).
The ash bed is usually barren, with only oc-casional chitinozoans recorded. A brief, usuallyminor bloom of small mobile benthos (ostra-codes and simple burrows) immediately fol-lowed ash deposition, overlapping with asubsequent dramatic plankton bloom of Apato-bolus micula and graptolites. The planktonbloom showed a pseudo-logarithmic decline,while A. micula also formed a partly overlappingsecond bloom, of longer duration. Thereafter,abundance gradually declined to, or below,initial levels, unless benthic oxygenation becameestablished. The ratio of ostracode to graptoliteabundance varied according to initial con-ditions; in originally anoxic facies, the plankton(Apatobolus micula plus graptolites) bloom wasby far the greater, and vice versa for originallydysaerobic substrates (Fig. 1). A horizontalsequence, sampling the 2 cm immediately abovethe ash upper surface, revealed steep faunal
abundance gradients (>300% over 10m), corre-sponding with up to a 20% change in ash thick-ness. A strong maximum in the immediatelypost-ash plankton abundance overlay the thick-est part of the bentonite.
Multiple lines of evidence were used to con-strain the processes operating, and a coherentinterpretation developed, based on verticalcirculation of a stratified water column. Fine ashdeposited onto a standing body of waterdescended initially as turbid flows rather thandiscrete particles (Carey 1997). Provided acritical water depth (strongly dependent onseveral parameters) was not exceeded, theseflows replaced deep, dysaerobic water withaerobic surface water, enabling a bloom of smallmobile benthos, restricted by low food supplies.Corresponding upwelling induced a large-scaleplankton bloom, continuing until inertial circu-lation ceased. This circulation could have beenmaintained directly by ash deposition for severalweeks, through continuous fragmentation offloating pumice and slow sinking of fineparticles. Following ash deposition, circulationwould be prolonged by the temperature inver-sion of ash-free fluid, and perhaps enhanced bylarge-scale particulate phytoplankton produc-tion at the surface. Upwelling is expected tohave continued weakly for a few months, suf-ficiently long to establish a substantial planktonbloom involving several trophic levels. The

VOLCANISM AND DIVERSIFICATION 103
organic rain allowed a benthic bloom of sessilesuspension-feeders to develop until oxygenationwas depleted below tolerable limits.
The total duration of these bloom events isreconstructed, based on a comparison withmodern bloom events (e.g. Gallardo et al. 1977)and post-seismic sedimentation rates (Goff1997), as five to ten years, with up to one year forthe plankton bloom (Botting 2000). The areaaffected depends on the extent of ash dispersal.Downwelling flows would decay rapidly throughlateral entrainment of fluid, unless concentratedby continued ash input from above. Distaldeposits in deep shelf settings would thus havebeen unable to host benthic blooms, althoughthe plankton bloom would be unaffected. Thiswould have provided elevated organic rain andencouraged benthic anoxia at the ash dispersalmargins, where the high organic input was notoffset by oxygenated downwelling (Fig. 2).While large eruptions affect wide areas, a typicalpyroclastic event may induce benthic blooms inareas of a few hundred to a few thousand squarekilometres. Coarse ash and crystal-tuff beds areless likely to initiate vertical circulation, sincethe critical particle density for mass flow is lesslikely to be exceeded, and individual particlesettling is much more rapid and more varied.
Population geneticsThe presence of repeated regional bloomevents, following the removal of small or sessilebenthos, potentially influenced populationgenetics in several ways. For general discussionof relevant concepts, refer to Harwood & Amos(1999), Amos & Harwood (1998), and refer-ences therein. Primarily, bloom events encour-age the retention of novel characters, via anincreased surviving proportion within eachgeneration. Although the survival probabilitiesof such mutations are not elevated after thebloom maximum, the reduced juvenile mortalityduring the bloom growth phase increases thechance of persistent variations. For example,beneficial genotypic changes that require twomutations, one neutral or detrimental, have verylow probability of arising under normal popu-lation conditions. During a bloom increasephase, carriers of each mutation are much moreabundant, allowing carriers of both to arise, andthereafter become preferentially selected. Ford& Ford (1930) described abundance and vari-ation fluctuations in an isolated butterfly popu-lation (of duration 10-20 years); variationwas maximized during a population increase,with the subsequent maximum and decliningpopulations being more homogenous. The final
population was morphologically distinct fromthe initial. While a single bloom event is prob-ably insufficient for speciation to occur from ahomogeneous population, the result is toproduce genetic heterogeneity on a subregionalscale. This may allow distinct species to appearrapidly under subsequent blooms, as disparateparts of an already heterogeneous populationare brought into immediate contact duringrecolonization (discussed below). Althoughsome intraspecific homogenization would beexpected to occur during intervening quiescentperiods, migration of genetic characteristicsthrough a stable population is much slower thanphysical migration of a population into a barrenregion.
Following the eradication of small mobile andsessile benthos by ash deposition, recolonizationoccurred by some combination of exponentialpopulation increase of rare survivors, and immi-gration from the surrounding area. The delayedonset of the sessile benthic bloom relative to themobile bloom, interpreted above as resultingfrom low food supplies inhibiting suspension-feeders, may also suggest a gradual immigrationIn reality, the bloom population was almost cer-tainly derived from geographically separatedlateral immigrants, combined with a remnantendemic population; lateral separation wasprobably tens to hundreds of kilometres,depending on the eruption size. The geneticdiversity of the benthic bloom biota should thusbe much greater than where the population isderived from a single endemic community. Thisdisparity is further exaggerated by the processdiscussed above, whereby previous bloomevents induced strong regional genetic variabil-ity. The populations at opposite edges of theaffected area would be expected to differ morethan in a stable environment, with these popu-lations becoming immediately adjacent duringrecolonization. Within the resultant population,hybridization and segregation between end-members would result in a higher rate of specia-tion than in more homogeneous communities.Hybridization would further promote theappearance of individuals with distinctivecharacteristics by providing novel gene combi-nations, while the initiation of non-interbreed-ing subpopulations is a necessary first stage insympatric speciation. In a gradually varyingpopulation, there is little incentive for segrega-tion, but behavioural divisions can exist immedi-ately when disparate parts of that population arebrought into contact.
The general situation of colonization of anecologically depauperate region promotesgenetic variation under many circumstances

104 JOSEPH P. BOTTING
(Whitlock & MeCauley 1990; McCauley 1991).Among the most critical parameters controllinggenetic variability is whether recolonizationoccurs through population fusion (differentia-tion decreased) or permanent fission (differenti-ation increased). In many cases, however, itappears that subspecific diversification resultsfrom fission of a population, followed byrefusion, and further factors are presumed to beoperating in order to account for observeddiversification. One possibly is unidirectional'meiotic drive," discussed by Amos & Harwood(1998), but present understanding is verylimited. The critical point when considering theincreases in genetic diversity and potentialspeciation rate is that the bloom events wererepeated. Several thousand events, each sepa-rated by the order of decades to centuries, havethe potential to exaggerate slight probabilisticchanges, perhaps producing noticeable effectson global diversity curves.
A further element that strongly encouragesspeciation in modern faunas (e.g. cichlids:Barlow et al 1984; Russock 1986; Knight et al,1998; Seehausen & van Alphen 1999) is sexualselection based on parental imprinting, whichcan potentially lead to sympatric speciation inheterogeneous populations. However, inter-preting the reproductive habits of extinct organ-isms is notoriously difficult, despite ambiguousevidence that trilobites, for example, providedsome degree of guardianship over their offspring(Davis et al. 2001). Uniformitarian assumptionssuggest that many arthropods provided thepotential for imprinting (e.g. Barnes 1987), butdirect evidence is lacking. Overall, the potentialfor parental imprinting and pure sympatricspeciation by this mechanism in invertebrateswas probably limited.
The effects of subsequent mixing under stableenvironmental conditions must also be con-sidered. Taxa showing highly specific behav-ioural traits may not fall into direct competition,and the increased diversity may be conserved.This is also true of inhabitants of facies with adiverse range of ecological niches, a featureenhanced by the development of tiering duringthe Ordovician (e.g. Droser et al. 1995). Soft-substrate shelf facies with low faunal abundanceand diversity may provide the focus for diversityincreases, but some diversity is likely to havebeen lost by merging or competition duringlong-term quiescence, in the absence of complexecosystems that could maintain niche separ-ation. Although the centres of diversificationwould have been in these environments, sus-tained long-term diversity increases may haveinitially occurred by range expansion into
shallower-water facies, where diverse physicalconditions and complex ecosystems providedpermanent habitats. (Trends of this natureshould be recognizable through comparison oftemporal ranges and morphological variabilitybetween closely related species occurring indifferent environments.) Abundant specificniches could then have allowed preservation oflineages that would rapidly have become extinctin their original environment. The observedonshore-offshore trend during the Palaeozoic(e.g. Bottjer et al. 1988) would reflect the gradualexpansion of this complex, stable ecosystem intodeeper water, while most of the explosive diver-sity increase at speciation centres was subse-quently lost from the original environment.
Since the geographic mixing of distinctivepopulations is critical to this process, plankticorganisms should not be significantly affected.The point relating to preservation of novel char-acters within the blooms would still stand, butconstant mixing of populations would reduce thesignificance of these effects; diversity increasesshould be expected, but to a much lesser extentthan in the benthos.
Implications for Ordovician diversityThe Ordovician Period was characterized bywidespread explosive volcanism in many parts ofthe world, as discussed above. The results of ashdeposition in shelf regions represent a concep-tual argument whereby an increased speciationrate could have operated in volcanically activeareas, and which provides objective predictionsfor several aspects of the diversity patternsexpected. The temporal, geographical andenvironmental distributions should show threebasic correlations; maximum rates of diversifica-tion should correspond to:
(a) areas and times of extensive pyroclasticvolcanism, emphasizing numerous smalleruptions rather than few large ones;
(b) benthic oxygen limitation and well-developed stratification, leading to high-amplitude bloom events;
(c) high eustatic sea level, increasing the shelfarea over which small eruptions couldproduce significant effects.
Volcanism and sea level are usually related, ona global scale, through increased tectonicactivity and ocean ridge volume, while theirrelationship to benthic anoxia is presently un-certain. However, all three factors particularlycharacterize the Ordovician in general, and thecoincidence of all three is predicted to correlate

VOLCANISM AND DIVERSIFICATION 105
with diversification centres on a regional tobasin scale. If the proposed process was signifi-cant, this should be supported by the globaldatabase results; for example, areas of diversifi-cation should be dominated by volcanicallyactive regions showing stratification. The diver-sity increases in other areas should then beshown to occur largely through immigration ofpreviously existing taxa.
A further set of predictions relates to thedifferential rates of evolution between differentecological and taxonomic groups. In these cases,the evolutionary response of each group can bepredicted from the established ecologicalresponse, in that those groups showing thehighest abundance fluctuations and most limitedindividual geographic ranges should respondmost strongly to the variations in populationgenetics outlined above. Temporary local eradi-cation followed by high-amplitude blooms fromrapid recolonization should have maximizedthe evolutionary response. These ecologicalresponses were presumably dependent onseveral factors, including larval and adult lifehabits, trophic group, size, reproductive strategyand developmental time-scales, most of whichcan only be estimated. Observations of palaeo-ecological response remove the complications ofreasoning about factors that may be difficult toassess in extinct taxa. The predicted evol-utionary responses are described according tobroad ecological divisions.
(1) Small sessile benthos and essentially immo-bile epibenthos: extensive responses, appar-ently completely eradicated by substantialash deposition, but with rapid recoloniza-tion in a high-amplitude bloom. Includesbryozoans, small brachiopods ('articulate*and 'inarticulate'), ostracodes, sphenothal-lids. (Ostracodes are included here becausetheir small size allows very restricted move-ment. However, they were apparently ableto escape minor ash deposition, while smallsessile taxa were not, and recolonized aregion more rapidly.)
(2) Intermediate to large sessile benthos: signifi-cant responses, with local eradication andhigh-magnitude blooms. May escape slightash deposition. Includes crinoids, sponges,bryozoans, larger brachiopods, conulariids.Although generally rare, some examples ofthis group (sponges, brachiopods, bryo-zoans) gave rise to brief, intense blooms(Botting 2000).
(3) Weakly mobile epi- and endobenthos(excluding ostracodes): often able to escapedeposition of small ash volumes, but bloom
Fig. 3. Summary of differing diversification patternsby phylum based on Sepkoski (1995), reproduced bykind permission of the Pacific Section - SEPM. (a)Taxa in groups 1, 2 and 3 (part) (see text), exhibitingstrong ecological response to ash-fall; (b) taxa ingroups 3 (part), 4 and 5 (see text), exhibiting lesserecological response to ash-fall, or existing asplankton only.
events not obvious due to small populationsize. Includes bivalves, gastropods, palaeo-scolecids, larger lingulids. Their responsemay be underestimated here because oflower preservation potential of aragoniticand multi-element skeletons.
(4) Mobile epibenthos/nekton: limitedresponse; little noticeable effect on abun-dance across ash beds. Includes trilobites,nautiloids, although aragonitic nautiloidsmay be underestimated because of reducedpreservation.
(5) Plankton and pelagic nekton: affected byextensive blooms, but little or no priordestruction; rapid migration prevents seg-regation into genetically distinct communi-ties, and isolated populations implausible.Includes graptolites, pelagic trilobites,acritarchs (and some chitinozoans?).
Further complications include the toleranceof some taxa to low oxygenation, and ambiguousinterpretations of life habits; 'inarticulate'brachiopods often appear in dysaerobic settings,and some were almost certainly pseudoplanktic
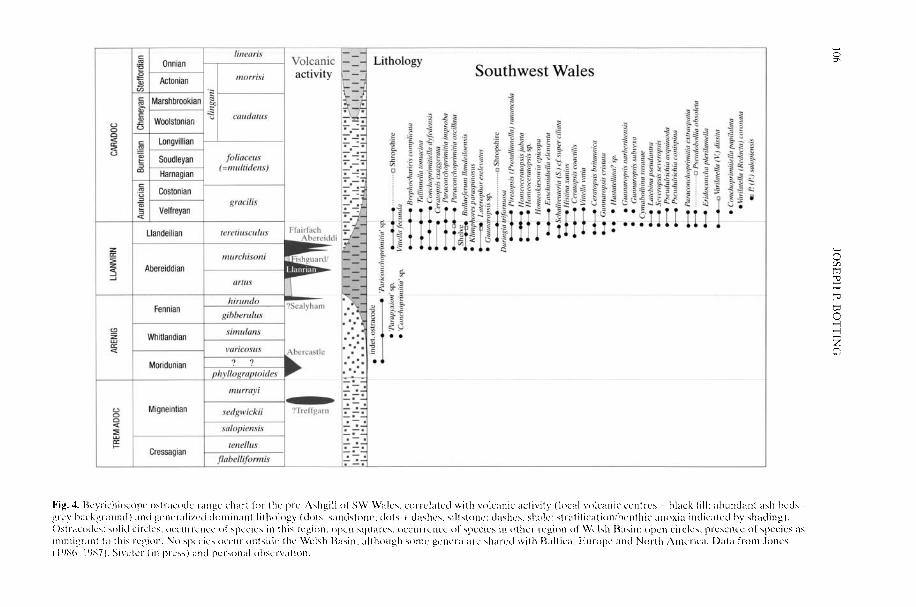
Fig. 4. Beyrichioeope ostracode range chart lor the pre-Ashgill of SW Wales, correlated with volcanic activity (local volcanic centres = black f i l l ; abundant ash beds =grey background) and generalized dominant lithology (dots, sandstone; dots + dashes, siltstone; dashes, shale; s trat i l icat ion/benthic anoxia indicated by shading).Ostracodes: solid circles, occurrence of species in this region; open squares, occurrence of species in other region of Welsh Basin; open circles, presence of species asimmigrant to this region. No species occur outside the Welsh Basin, al though some genera are shared with Ballica, Europe and North America. Data from Jones(1986-1987). Siveter (in press) and personal observation.
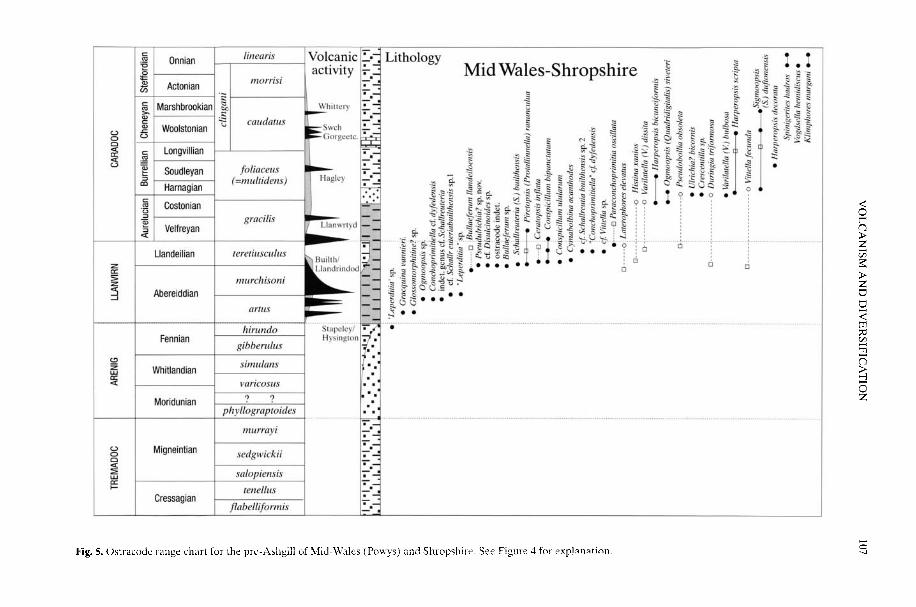
Fig. 5. (Ostracode range chart for the prc-Ashgill of Mid-Wales (Powys) and Shropshire. See Figure 4 for explanation

108 JOSEPH P. BOTTING
(Lockley & Antia 1980; Gabbott 1999; Botting& Thomas 1999). For example, A. miculabenthie populations would have been lessstrongly affected than predicted from thebenthie bloom, because of population mixingunder pseudoplanktic conditions. In contrast,'articulate' brachiopods are generally facies-restricted, and appear among predictable faunalassemblages. It is significant that Sepkoski(1995) found much greater net diversificationamong 'articulates' than 'inarticulates,' sincemany linguloids were capable of burrowing,while 'articulates' were almost exclusivelysessile epibenthos.
Published global diversity curves differenti-ated into taxonomic groups (Sepkoski 1995)appear to record patterns that are consistentwith these predictions (Fig. 3). In general,sessile taxa (brachiopods, bryozoans, corals)show the greatest diversification, with a highlyconsistent pattern between phyla. Bivalves andgastropods first achieved significant diversityduring the Ordovician, but then show veryrapid diversification, while trilobites fluctuatedirregularly (confirmed by Adrain & Westrop2000). Graptolites show little overall diversityincrease, although with significant fluctuations(Cooper 1999); a similar pattern is observedamong acritarchs and chitinozoans (e.g. Kaljo etal 1995). Among the groups predicted to showgreatest diversification are the ostracodes,based on their ecological response. Swain(1996) reported extremely elevated ostracodeturnover, with substantial overall diversifica-tion, in a bentonite-rich Ordovician sequence ofNorth America. He tentatively attributed this tonutrient input but, as discussed above, aplausible precise mechanism is not obvious.
Welsh Basin beyrichiocope ostracodediversityA summary of the Ordovician diversity of pre-Ashgill beyrichiocope ostracodes in the WelshBasin follows; other 'ostracode' groups are notincluded, because of uncertainty over mutualrelationships. The area is subdivided into threesubregions (SW Wales, Mid-Wales and Shrop-shire, North Wales and northern England) andcorrelated with major volcanic episodes withineach area. An estimate of bentonite frequency isincluded, based on limited descriptions instratigraphic literature and personal obser-vation, although in many cases thin beds are notobvious in outcrop, and have not been men-tioned in publications. Where bentonites appearfrequently within an interval lacking a local
volcanic source, they can usually be correlatedwith episodes of major volcanism in one of theother subregions. In addition, a schematic log ofprimary lithology is included for each region,although extreme local variation may occur;apparent stratification (benthie anoxia) isdenoted by shading (Figs 4, 5, 6).
Ostracode data were obtained primarily fromJones (1986-1987), with additional Arenig in-formation from Siveter (in press), and personalobservations. The rarity of pre-Llanvirn taxa isapparently real, although the simple morphol-ogy of described examples may disguise greatertaxonomic variation than is practically identifi-able. However, the same underestimate shouldoperate in later unornamented taxa, suggestingthat the observed morphological diversificationaccurately reflects a real pattern. Data regardingthe stratigraphic position of volcanic episodes,and dominant lithologies were obtained fromSmith & George (1961), George (1970), Forteyet al (2000) and Rushton et al. (2000) as primarysources, supplemented by additional local refer-ences (e.g. Stamp & Wooldridge 1923; Davies1933; Jones & Pugh 1941, 1949; Diggens &Romano 1968; Earp & Haines 1971; Lynas 1983:Cave & Rushton 1996; Davies et al. 1997).
In SW Wales (Fig. 4), massive diversificationover a brief interval (Llanvirn) is correlated withblack shales and extensive volcanism of theFishguard, St David's and Strumble Headregions. Arenig volcanism in aerobic sand faciesproduced little response, although a fewmorphologically conservative taxa appeared atthis time. The low diversity in the upper Caradocmay be partly an artefact; diverse Ashgill faunasare known to exist, but are yet to be studiedintensively (Jones 1987). However, althoughCaradoc ostracodes above the Aurelucian arealmost certainly present (some taxa, such asDuringia triformosa, reappear in Shropshireduring the Burrellian), they are apparently veryrare; no identifiable specimens from the areahave been described.
In Mid-Wales and Shropshire (Fig. 5), specia-tion occurred over the Llanvirn and Caradoc,corresponding to penecontemporaneous vol-canic centres and restricted oxygenation. A fewspecies were immigrants from SW Wales, butmost were endemic.
There are limited data for North Wales andnorthern England (Fig. 6) but the majority ofspecies correspond to the Borrowdale volcanicepisode (Caradoc); a high proportion wasimmigrant. Additional faunas are predicted tobe discovered from the Llanvirn and lowerCaradoc.
Overall, the patterns appear to show

VOLCANISM AND DIVERSIFICATION 109
Fig. 6. Ostracode range chart for North Wales and northern England. See Figure 4 for explanation.
enhanced speciation over the intervals contain-ing frequent ash beds in each area, and lowdiversity outside these episodes. Although nosimilar comparison has been made with areaslacking ash beds, the correlation of diversitywith volcanism appears to be precise in thecurrent data; this view is reinforced by themarked differences between adjacent regions.
The question of sampling bias must beaddressed in this situation, since volcanic areastend to preserve more resistant lithologies,
which may aid the preservation and exposure offossils. However, if this is true, identical patternsshould emerge among taxonomic groups thatshared similar environments and are extractedby similar processes. The ostracode data derivelargely from silicified specimens extracted byacid digestion of limestones, and from mouldicmaterial in siliciclastic sedimentary rocks.Sutton et al. (1999-2000) monographed WelshBasin lingulid brachiopods, using very similarmaterial from a similarly comprehensive
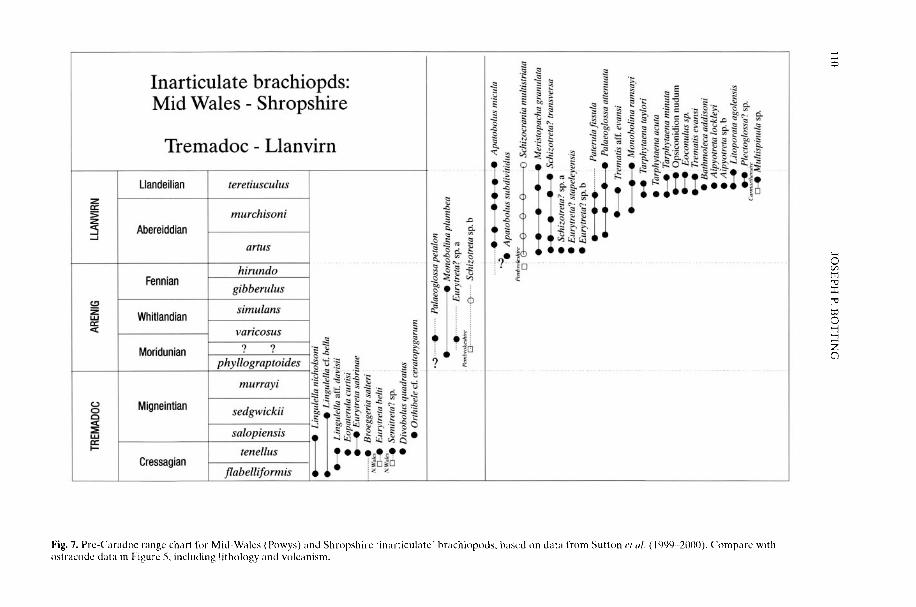
Fig. 7. Pre-Caradoc range chart for Mid-Wales (Powys) and Shropshire Inarticulate' hrachiopods, based on data from Sutton et al. (1999-2000). Compare withostracode data in Figure 5, including lithology and volcanism.

VOLCANISM AND DIVERSIFICATION 111
number of sites. If the patterns observed forostracodes reflect sampling bias resulting fromareal exposure and preservation, the samepatterns should be seen among 'inarticulate'brachiopods. Figure 7 shows the pre-Caradoc'inarticulate' data for Mid-Wales and Shrop-shire. This reveals a more gradual increase indiversity, with a significant number of Tremadocand Arenig species, although the rate of speciesappearance also increased at the time ofLlanvirn volcanism, including similar absoluteorigination. However, it is likely that many moreostracode species remain undiscovered thanbrachiopods, due to their inconspicuousness. Asignificant proportion of the ostracodes encoun-tered during fieldwork were undescribed,whereas brachiopods were almost alwaysidentifiable. Although at an early stage, thisinitial analysis suggests a local influence ofvolcanism on diversification, which affectedecological or taxonomic groups in differentways, these differences being consistent withecological studies of the local effects of volcanicash deposition.
ConclusionsThe local effects of volcanic ash deposition onplanktic and benthic communities in dysaerobicshelf/basin settings have been summarized. Theresultant elimination of benthos, recolonizationfrom lateral areas, and bloom events in bothplankton and benthos, have been analysed interms of population genetics, and shown tocomprise a conceptual method for encouragingspeciation. Physical conditions necessary for thisprocess were maximized during the Ordovician,leading to comparisons with the OrdovicianRadiation. If the process were a significantfactor in this diversification event, predictabledifferences would be expected between taxo-nomic and ecological groups, which may beassessed by comparison with observed palaeo-ecological responses, and reasoning frommodern ecological models. A series of such pre-dictions is provided, with caveats in the case ofambiguous groups. This is illustrated by a sub-basin-scale analysis of ostracode diversity, cor-related with volcanism, and compared with theless dramatic diversification of 'inarticulate'brachiopods to reduce concerns of samplingbias. A possible distinction between those faciesshowing very high speciation and extinctionrates, and others able to adopt more modestlong-term diversity increases is also suggested.Speciation is viewed as occurring largely in themid to outer shelf regions, but within an eco-system insufficiently complex to support this
diversity following environmental stabilization.However, onshore expansion of new speciesmay have contributed to the increasing diversityof more complex shallow-water communitiesthat typify the Palaeozoic Evolutionary Fauna,resulting in a gradual offshore expansion ofthese ecosystems.
This chapter presents a mechanism thatpotentially explains large parts of the Ordo-vician diversification, with predictions. Readersare encouraged to present additional data as atest of the hypothesis, in order to establish itsvalidity. Even if the hypothesis is disproved, theuse of fine-scale diversity and palaeobiogeogra-phy databases is suggested as an essential pro-cedure for understanding processes involved inthe diversity changes. Extrapolation and averag-ing over larger scales will hopefully then allowelucidation of global database results.
Development of the population genetics section wasgreatly aided by discussion with W. Amos (Zoology,University of Cambridge), while D. J. Siveter (Uni-versity of Leicester) and A. W. Owen generouslyprovided 'in press' data regarding ostracodes andtrilobites respectively. The palaeoecological study wasperformed in partial fulfilment of a PhD at theUniversity of Birmingham, during which discussionswith A. T. Thomas and M. P. Smith are particularlyacknowledged. The manuscript was improved byconstructive reviews from A. I. Miller and D. J.Siveter. Research is currently funded by Christ'sCollege, Cambridge. This paper is a contribution toIGCP 410 The Great Ordovician BiodiversificationEvent'.
ReferencesADRAIN, J. M. & WESTROP, S. R. 2000. An empirical
assessment of taxic paleobiology. Science, 289,110-112.
ADRAIN, J. M., WESTROP, S. R., CHATTERTON, B. D. E.& RAMSKOLD, L. 2000. Silurian trilobite alphadiversity and the end-Ordovician mass extinction.Paleobiology, 26, 625-646.
AMOS, W. & HARWOOD, J. 1998. Factors affectinglevels of genetic diversity in natural populations.Philosophical Transactions of the Royal Society ofLondon, B 353, 177-186.
BARLOW, G. W., VALERIO, M & HOLDER, J. 1984.Does the polychromatic midas cichlid showsexual imprinting? American Zoologist, 24, A51(abstract).
BARNES, R. D. 1987. Invertebrate Zoology. SaundersCollege Publishing, USA.
BOTTING, J. P. 2000. Palaeoecology and Systematics ofOrdovician Biotas from Welsh VolcaniclasticDeposits, PhD Thesis, University of Birmingham.
BOTTING, J. P. & THOMAS, A. T. 1999. A pseudoplank-tonic inarticulate brachiopod attached to grapto-lites and algae. A cta Universitatis Carolinae:Geologica, 43, 333-335.

112 JOSEPH P. BOTTING
BOTTJER, D. I, DROSER, M. L. & JABLONSKI, D. 1988.Palaeonenvironmental trends in the history oftrace fossils. Nature, 33, 252-255.
BOYD, P. W., WATSON, A. J.. LAW, C. S., ET AL. 2000.A mesoscale phytoplankton bloom in the polarSouthern Ocean stimulated by iron fertilization.Nature. 407, 695-702.
CAREY, S. N. 1997. Influence of convective sedimen-tation on the formation of widespread tephra falllayers in the deep sea. Geology, 25, 839-842.
CAVE, R. & RUSHTON., A. W. A. 1996. The Llandeilo(Ordovician ) Series in the core of the Tywi Anti-cline, Llanwrtyd, Powys, UK. Geological Journal,31. 47-60.
CONWAY MORRIS, S. 1999. The evolution of diversity inancient ecosystems: a review. In: MAGURRAN, A.E. & MAY, R. M. (eds.) Evolution of BiologicalDiversity, Oxford University Press, New York,282-322.
COOPER, R. A. 1999. Graptolites and the great Ordo-vician biodiversification event. Acta UniversitatisCarolinae: Geologica. 43, 441-442.
COPE, J. C. W. 1999. Middle Ordovician bivalves fromMid-Wales and the Welsh borderland. Palaeontol-ogy. 42, 467-499.
COPE, J. C. W. INGHAM, J. K. & RAWSON, P. F. 1992.Atlas of Palaeogeography and Lithofades. Geo-logical Society, London. Memoirs, 13.
DAVIES, J. R.. FLETCHER, C. J. N., WATERS, R. N,WILSON, D., WOODHALL, D. G. & ZALASIEWICZ.J. A. 1997. Geology of the Country around Llani-lar and Rhayader. British Geological Survey, TheStationery Office, London.
DAVIES, K. A. 1933. The geology of the countrybetween Abergwesyn (Breconshire) and Pump-saint (Carmarthenshire). Quarterly Journal of theGeological Society, London. 89. 172-201.
DAVIS, R. A., FRAAYE, R. H. B. & HOLLAND, C. H.2001. Trilobites within nautiloid cephalopods.Lethaia, 34, 37-45.
DIGGENS, J. N. & ROMANO, M. 1968. The Caradocrocks around Llyn Cowlyd, North Wales. Geo-logical Journal, 6, 31-48.
DROSER, M. L., SHEEHAN. P. M., FORTEY, R. A. &XING Li, 1995. The nature of diversification andpalaeoecology of the Ordovician Radiation withevidence from the Great Basin. In: COOPER, J.D..DROSER, M. L. & FINNEY, S. C. (eds) OrdovicianOdyssey: Short Papers for the Seventh Inter-national Symposium on the Ordovician System.Pacific Section SEPM, Fullerton, California,415-418.
EARP. J. R. & HAINES, B. A. 1971. British RegionalGeology: the Welsh Borderland (third edition).Institute of Geological Sciences, London.
ERWIN, D. H.. VALENTINE, J. W. & SEPKOSKI, J. J. JR.1987. A comparative study of diversificationevents: the Early Palaeozoic versus the Mesozoic.Evolution, 41, 1177-1186.
FORD, H. D. & FORD, E. B. 1930. Fluctuation innumbers, and its influence on variation, in Meli-taea aurinia. Rott. (Lepidoptera). Transactions ofthe Entomological Society of London, 78.345-351.
FORTEY, R. A. & OWENS, R. M. 1978. Early Ordo-vician (Arenig) stratigraphy and faunas of theCarmarthen district, south-west Wales. Bulletin ofthe British Museum Natural History (Geology).30,225-294.
FORTEY, R. A., HARPER, D. A. T. INGHAM. J. K..OWEN, A. W. & RUSHTON, A. W. A. 2000. Arevised correlation of Ordovician rocks in theBritish Isles. Geological Society. London. SpecialReports. 5.
GABBOTT, S. E. 1999. Orthoconic cephalopods andassociated fauna from the late Ordovician SoomShale lagerstatte. South Africa. Palaeontology.42,123-148.
GALLARDO, V. A., CASTILLO. J. G., RETAMAL. M. A..YANEZ, A., MOYANO. H. I., & HERMOSILLA, J. G.1977. Quantitative studies on the soft-bottommacrobenthic animal communities of shallowAntarctic bays. In: LLANO. G. A. (ed.) Adapta-tions within Antarctic Ecosystems. Proceedings ofthe 3rd SCAR Symposium on Antarctic Biology.Smithsonian Institution. Washington D.C..361-387.
GEORGE, T. N. 1970. British Regional Geology: SouthWales. Institute of Geological Sciences, London.
GOFF, J. R. 1997. A chronology of natural and anthro-pogenic influences on coastal sedimentation. NewZealand. Marine Geology. 138. 105-117.
HARWOOD. J. & AMOS. W. 1999. Genetic diversity innatural populations. In: MAGURRAN. A. E. &MAY, R. M. (eds) Evolution of Biological Diver-sity. Oxford University Press. New York. 56-69.
HUFF. W. D.. BERGSTROM, S. M. & KOLATA, D. R.1992. Gigantic Ordovician volcanic ash fall inNorth America and Europe: biological, tectono-magmatic and event-stratigraphic significance.Geology, 20, 875-878.
JAVOY. M. & COURTILLOT. V. 1989. Intense acidic vol-canism at the Cretaceous-Tertiary boundary.Earth and Planetary Science Letters. 94, 409-416.
JONES. C. R. 1986-1987. Ordovician (Llandeilo andCaradoc) beyrichiocope Ostracoda from Englandand Wales. Monograph of the PalaeontographicalSociety, London. 1. 1986: 138. 1-76: 2. 1987: 139.77-114.
JONES. O. T & PUGH, W. J. 1941. The Ordovicianrocks of the Builth district. Geological Magazine.78. 185-191.
JONES. O. T. & PUGH. W. J. 1949. An Early Ordo-vician shoreline in Radnorshire, near BuilthWells. Quarterly Journal of the Geological Societyof London. 104.43-70.
KALJO. D., NOLVAK. J. & UUTELA. A. 1995. Ordo-vician microfossil diversity in the Rapla section,northern Estonia. In: COOPER. J. D.. DROSER. M.L. & F I N N E Y , S. C. (eds) Ordovician Odyssey:Short Papers for the Seventh International Sym-posium on me Ordovician System. Pacific SectionSEPM. Fullerton, California, 415-418.
KNIGHT, M. E., T I R N E R , G. F. Rico. C. VAN OPPEN.M. J. H. & HEWITT, G. M. 1998. Microsatellitepaternity analysis on captive Lake Malawi cich-lids supports reproductive isolation by directmate choice. Molecular Ecology. 7. 1605-1610.

VOLCANISM AND DIVERSIFICATION 113
LOCKLEY, M. G. & ANTIA, D. D. J. 1980. Anomalousoccurrence of the Lower Palaeozoic brachiopodSchizocrania. Palaeontology, 23, 707-713.
LYNAS, B. D. T. 1983. Two new Ordovician volcaniccentres in the Shelve Inlier, Powys, Wales. Geo-logical Magazine, 120, 535-542.
McCAULEY, D. E. 1991. Genetic consequences of localpopulation extinction and recolonization. Trendsin Ecology and Evolution, 6, 5-8.
MCCORMICK, T. & OWEN, A. W. 2001. Assessing trilo-bite biodiversity in the Ordovician of the BritishIsles. GeologicalJournal 36, 279-290.
MILLER, A. I. \997a. Dissecting global diversitypatterns: examples from the OrdovicianRadiation. Annual Review of Ecology and Sys-tematics, 28, 85-104.
MILLER, A. I. 1997b. A new look at age and area: thegeographic and environmental expansion ofgenera during the Ordovician Radiation. Paleo-biology, 23,410-419.
MILLER, A, 1.1998. Biotic transitions in global marinediversity. Science, 281,1157-1160.
MILLER, A. I. 2000. Conversations about Phanerozoicglobal diversity. In: ERWIN, D. G. & WING, S. L.(eds) Deep Time, Paleobiology (supplement), 26,53-73.
MILLER, A. I. & MAO, SHUGUANG. 1995. Association oforogenic activity with the Ordovician radiation ofmarine life. Geology, 23,305-308.
OWEN, A. W. & McCORMICK, T. 1999. Ordovician bio-diversity change in the British Isles: a databaseapproach. I. Rationale and database structure.Acta Universitatis Carolinae: Geologica, 43,523-526.
PATZKOWSKY, M. E. 1995. Ecologic aspects of theOrdovician Radiation of articulate brachiopods.In: COOPER, J. D., DROSER, M. L. & FINNEY, S. C.(eds) Ordovician Odyssey: Short Papers for theSeventh International Symposium on the Ordo-vician System, Pacific Section SEPM, Fullerton,California, 413-414.
RUSHTON, A. W. A., OWEN, A. W, OWENS, R. M. &PRIGMORE, J. K. 2000. British Cambrian to Ordo-vician Stratigraphy, Joint Nature ConservationCommittee, Peterborough.
RUSSOCK, H. I. 1986. Preferential behavior ofSarotherodon (Oreochromis) mossambicus(Pisces, Cichlidae) fry to maternal models and itsrelevance to the concept of imprinting. Behav-iour, 96, 304-321.
SEEHAUSEN, O. & VAN ALPHEN, J. M. 1999. Can sym-patric speciation by disruptive sexual selectionexplain rapid evolution of cichlid diversity inLake Victoria? Ecology Letters, 2, 262-271.
SEPKOSKI, J. J. JR. 1988. Alpha, beta or gamma: wheredoes all the diversity go? Paleobiologv, 14,221-234.
SEPKOSKI, J. J. JR. 1993. Ten years in the library - newdata confirm paleontological patterns. Paleobiol-ogy, 19, 43-51.
SEPKOSKI, J. J. JR. 1995. The Ordovician radiations:diversification and extinction shown by globalgenus-level data. In: COOPER,J. D., DROSER,M. L.& FINNEY, S. C. (eds) Ordovician Odyssey: ShortPapers for the Seventh International Symposiumon the Ordovician System. Pacific Section SEPM,Fullerton, California, 393-396.
SIVETER, D. J. (in press). The Ordovician. In: WILKIN-SON, I. (ed.) A Stratigraphical Index of BritishOstracoda (second edition). British Micro-palaeontological Society.
SMITH, B., & GEORGE, T. N. 1961. British RegionalGeology: North Wales. Her Majesty's StationeryOffice, London.
STAMP, L. D. & WOOLDRIDGE, S. W. 1923. Theigneous and associated rocks of Llanwrtyd(Brecon). Quarterly Journal of the GeologicalSociety, London, 74,16-46.
STILLMAN, C. J. 1984. Ordovician volcanicity. In:BRUTON, D. L. (ed.) Aspects of the OrdovicianSystem. Palaeontological Contributions from theUniversity of Oslo, 295, Universitetsforlaget,183-194.
SUTTON, M. D., BASSETT, M. G. & CHERNS, L.1999-2000. Lingulate brachiopods from theLower Ordovician of the Anglo-Welsh Basin.Monograph of the Palaeontographical Society,London, 1, 1999: 153, 1-60; 2, 2000: 154,61-114.
SWAIN, F. M. 1996. Ostracode speciation followingMiddle Ordovician extinction events, northcentral United States. In: HART, M. B. (ed.) BioticRecovery from Mass Extinction Events. Geo-logical Society, London, Special Publications, 102,97-104.
WHITLOCK, M. C. & MCCAULEY, D. E. 1990. Somepopulation genetic consequences of colonyformation and extinction: genetic correlationswithin founding groups. Evolution, 44,1717-1724.

This page intentionally left blank

The early evolution and palaeobiogeography of Mesozoicplanktonic foraminifera
MALCOLM B. HART, MELISSA J. OXFORD & WENDY HUDSONDepartment of Geological Sciences & Plymouth Environmental Research Centre,
University of Plymouth, Drake Circus, Plymouth PL4 8A A, UK(e-mail: [email protected])
Abstract: The planktonic foraminifera almost certainly evolved from benthonic ancestorsin the early Jurassic. The meroplanktonic genus Conoglobigerina, known from south-central and eastern Europe, appears in the Bajocian and is probably derived from the evenmore geographically restricted Praegubkinella. This genus was represented by a singletaxon in the earliest Toarcian but diversified after the Toarcian anoxic event. At the samelevel Oberhauserella quadrilobata Fuchs, 1967 became more inflated and there is some evi-dence to suggest that the 'anoxic event' was the environmental perturbation that began thetransition to a planktonic mode of life. In the Callovian-Oxfordian interval, the planktonicforaminifera are still restricted to a relatively limited area bounded by the North AtlanticOcean, NW Europe and Eastern Europe and this remained the case even in the earliestCretaceous. It was only in the Aptian-Albian that the palaeogeographical distributionchanged dramatically, probably as a response to the elevated sea levels caused by theincreased rate of ocean crust production which began in the Early Aptian. The principaldiversification events in the Jurassic (Toarcian, Bajocian, Callovian-Oxfordian) also appearto be related to sea level highstands.
The planktonic foraminifera are a distinctiveand abundant part of the modern oceanic fauna.Together with the other planktonic groups(coccolithophorids, diatoms, radiolarians, dino-flagellates, etc), they assist in the chemical/nutrient cycling of the ocean system. While theoceanic plankton, in general, impact on thelevels of atmospheric CO2 in the Mesozoic andCenozoic, the contribution of the planktonicforaminifera is probably difficult to quantifyalthough, as we will demonstrate, the develop-ment of the group was rather limited until themid-Cretaceous. During the Mesozoic it isknown that the planktonic foraminifera:
• evolved from benthonic ancestors;• diversified;• expanded to attain an almost global distri-
bution;• suffered during several major 'events'
(including possible bolide impacts andmethane escapes from gas hydrates); and
• were reduced to two or three species by theterminal Cretaceous 'events' or 'event'(bolide impact(s), sea-level change, tem-perature rise/fall, volcanic fall-out, etc.).
Most micropalaeontologists (e.g. Caron 1983;
Hart 1999; Premoli Silva & Sliter 1999) havetreated the planktonic foraminifera as thoughthey were a single evolutionary plexus, althoughthose working on DNA sequencing of themodern fauna have questioned this basicassumption (e.g. Darling et al. 1996, I999a,b)and suggested that the evolution of the plank-tonic foraminifera may be polyphyletic. It iscertainly true that one cannot claim that theJurassic record of the group (Fig.l) providesanything other than a disjointed set of relation-ships. While preservation and/or collectionfailure is probably a contributing factor, thereare several problems:
• the Praegubkinella - Conoglobigerina link:• the disjointed distribution of Conoglobige-
rina; and• the origins of Globuligerina and other taxa
in the mid- to late Jurassic.
Higher, in the Cretaceous succession, we havelittle evidence for the origin of the Praehed-bergella lineage or the heterohelicids (Fig. 2)and it is clear that the new evidence from thework on DNA must be taken into account and apolyphyletic history considered alongside othersuggestions.
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesoioic-Cenoioic Radiations. Geological Society, London, Special Publications, 194,115-125.0305-8719/02/$15.00 © The Geological Society of London 2002.
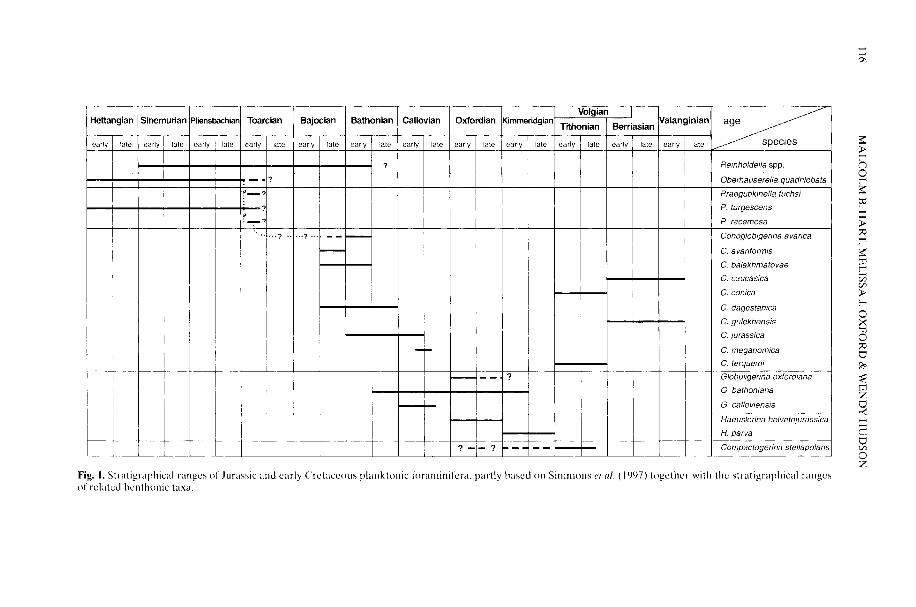
Fig. 1. Stratigraphical ranges of Jurassic and early Cretaceous planktonic foraminifera, partly based on Simmons et al (1997) together with the stratigraphical rangesof related henthonic taxa.
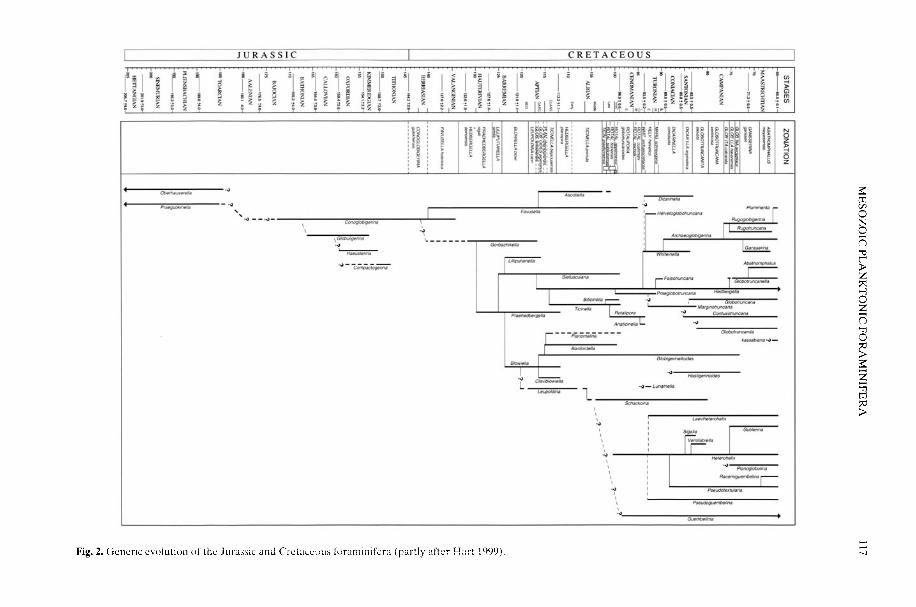
Fig. 2. Generic evolution of the Jurassic and Cretaccous foraminifcra (partly after Hart 1999).

118 MALCOLM B. HART, MELISSA J. OXFORD & WENDY HUDSON
Fig. 3. Distribution of Conoglobigerina spp. in the Middle Jurassic.
Origin of the planktonic for ami niter a
In their review of this subject, Simmons et al.(1997) dismiss the report of Pliensbachianplanktonic specimens (Gorog 1994) as probablecontamination but, more significantly, reject allthe taxa described as planktonic foraminifera byFuchs (1967, 1971, 1973, 1975, 1977). Thesepoorly preserved specimens have been exam-ined by F. Rogl (Natural History Museum,Vienna) and A. Gorog (Budapest, Hungary),both of whom indicate that they are not plank-tonic taxa. Some of this material is currently onloan to The Natural History Museum (London)and it is clear to the senior author that they arebenthonic taxa with flattened umbilical sidesand a complete lack of the inflated chambersnormally associated with a planktonic mode oflife.
If the Triassic and early Jurassic records ofplanktonic foraminifera are rejected then theorigins of the group must be pre-Bajocian (Fig. 1)as the species of Conoglobigerina recorded fromthe Bajocian are clearly meroplanktonic (Wernli& Gorog 1999, 2000). In the Toarcian sedimentsof the Creux de Tours section (Teysachaux,Fribourg, Switzerland) Wernli (1995) hasdescribed a fauna of Praegubkinella spp. that isassociated with Oberhauserella quadrilobata,one of the original 'planktonic' taxa of Fuchs(1967). Wernli (1995) demonstrates the possibleevolution from Praegubkinella racemosa Wernliinto Conoglobigerina (Fig.l), thereby extending
the range of the ancestral forms into theToarcian. The section of the 'Creux de l'ours' hasbeen described by several authors (e.g. Mettrauxet al. 1986, 1989) and the recent commentary(Weidmann 1993) on the Swiss Geological MapNo.1244, Chatel-St-Denis (Weidmann et al.1993) provides much new information. Thefamous section (coordinates 565.07/154.30 onSwiss Geological Map No.1244) was improved in1970 when a new forestry road was constructed.Unfortunately many of the new sections are nowdegraded but one face still exists on a bend wherethe road crosses the stream. Previous workershave identified the elegantulum and exaratunSubzones of the falciferum Zone, placing thesection within the Toarcian 'anoxic event"(Mettraux et al. 1986, 1989, and referencestherein). The senior author has found specimensof Harpoceras and Dactylioceras in this part ofthe section, together with examples of Posidoniaassociated with hard, organic-rich mudstones. M.Mettraux provided the samples for the investi-gation by Wernli (1995), the foraminifera comingfrom a sample of the dark mudstones fromstratigraphically above the 'anoxic event'. ThePraegubkinella/Oberhauserella assemblage ofWernli (1995) is, therefore, of early Toarcian age.
Work on the Toarcian foraminifera of the UKand NW Europe (Hylton 2000; Hylton & Hart2000) has shown that, associated with theToarcian anoxic event, there is a 'bloom' of smallforms of O. quadrilobata which possess slightlymore inflated chambers than the typical forms

MESOZOIC PLANKTONIC FORAMINIFERA 119
Fig. 4. Generic and specific diversity of planktonic foraminifera throughout the Jurassic and Cretaceous (partlyafter Hart 1999).

120 MALCOLM B. HART, MELISSA J. OXFORD & WENDY HUDSON
Fig. 5. Distribution of planktonic foraminifera in the Oxfordian.
described by Fuchs (1967) and which have beenseen by the senior author in The Natural HistoryMuseum. This species appears to be a disaster oropportunist taxon, according to the definition ofHarries et al. (1996). These more inflated formsof O. quadrilobata are found in the UK,Germany and France (Hylton 2000), while theassociated fauna of Praegubkinella spp. is only
known from Switzerland (Wernli 1995) at thepresent time.
Our more inflated specimens of O. quadrilo-bata are probably still benthonic forms as theapertural side remains relatively flat. It is clear,however, that chamber inflation has occurredand it is impossible to deny the possibility that,in mature specimens, some degree of 'floating'
Fig. 6. Distribution of planktonic foraminifera in the Valanginian.

MESOZOIC PLANKTONIC FORAMINIFERA 121
Fig. 7. Distribution of planktonic foraminifera in the mid- to late Albian.
could have taken place. In Wernli's (1995)assemblage of Praegubkinella there are someforms with relatively flat apertural faces while P.racemosa appears much more inflated. Againthere is the possibility that mature individualsmay have been 'planktonic'. The time gapbetween the ranges of P. racemosa and theearliest true Conoglobigerina is significant andthis is one stratigraphic interval that requiresurgent investigation.
The Conoglobigerina fauna is best describedfrom Hungary (Wernli & Gorog 1999, 2000),Morocco (Wernli 1987), the former SovietUnion (Morozova & Moskalenko 1961;Kuznetsova & Gorbatchik 1980; Kasimova &Aliyeva 1984) and Greece (Baumgartner 1985).In the company of A. Gorog and J. Svabo, thesenior author has visited the Somhegy (SomHill) section west of Zirc (Hungary). Here'ammonitico rosso' carbonates are found infissures within the local succession. Samplescollected from the Bajocian/Bathonian sedi-ments have confirmed the presence of abundantConoglobigerina, comprising both thick-walledand thin-walled forms. Whether this is just aspecific difference or whether this variation inwall thickness indicates genuine dimorphismrequires further investigation. The externalmorphology of the thick-walled forms (Wernli &Gorog 2000) is such that they appear almostidentical to the thin-walled forms and it seemsunlikely, therefore, that they are simply ben-thonic/planktonic dimorphic pairs. Species of
Conoglobigerina are known from many loca-tions in central-southern and eastern Europeand this certainly marks a major expansion ofthe planktonic fauna within the Middle Jurassic(Fig. 3). If Wernli (1995) was correct in hisderivation of Conoglobigerina from Praegub-kinella racemosa, then an origin from thePraegubkinella assemblages of the Alpine areawould appear to be quite plausible in palaeo-geographical terms.
Evolution of the Mesozoic planktonicforaminiferaFigure 1 shows the stratigraphical distribution ofthe early planktonic foraminifera and while, inmany cases, the evolutionary lineages/linkagesare unknown it is clear that both specific andgeneric diversity remained low throughout theJurassic and early Cretaceous. Some of thelineages are quite disjointed and it is apparentthat a considerable amount of research remainsto be done on these faunas. One of the principalproblems, however, is the question of preser-vation. The early planktonic foraminifera are allthought to have been aragonitic, very much likethe Reinholdella, Oberhauserella and Epistom-ina faunas with which Jurassic taxa are oftenassociated in faunal assemblages. Aragoniticforaminifera are best preserved in dense claylithologies and are rarely well preserved inporous sandstones and siltstones or some

122 MALCOLM B. HART, MELISSA J. OXFORD & WENDY HUDSON
carbonate-rich environments. The earliestGlobigerinina have been placed in the Super-family Favusellacea (Simmons et al. 1997), agroup which continues through until the mid-Cretaceous (Fig. 2). In the Albian and Ceno-manian the Favusella spp. recorded in the UK(Carter & Hart 1977) do not appear to bearagonitic, being found well-preserved in thechalk facies (in which few other aragonitic fossilsare recorded in such a good state of preser-vation). At the present time there is littleevidence as to when the transition from arago-nitic to calcitic tests is to be found. The lack ofinformation on the faunas of the Tithonian toValanginian interval may be critical in thisrespect.
The major evolutionary changes in theToarcian and Bajocian are potentially related tosea-level highstands (Haq et al. 1988) and appearcoincident with intervals of marked ammonitediversification (O'Dogherty et al. 2000). If oneplots the distribution of the fauna through theJurassic and Cretaceous (Fig. 2) it is clear thatthe major change in evolutionary rate occurs inthe Aptian (Hart 1999; Premoli Silva & Sliter1999). Both specific and generic diversity (Fig. 4)also increase at this level and this changeappears to be coincident with a major increase inocean crust production (Larson 1991a,b; Larsonet al. 1993)
In the earliest Oxfordian the planktonicforaminifera (mainly Globuligerina oxfordiana(Grigelis 1958) and G. bathoniana (Pazdrowa1969)) expanded their distribution (Fig. 5) andare reported from much of Europe, includingthe UK (Oxford et al. in press) and the GrandBanks (Ascoli 1976). The work of Oxford et al(in press) in Dorset has demonstrated thepresence of G. oxfordiana, Haeuslerina helveto-jurassica (Haeusler, 1881) and Compactogerinastellapolaris (Grigelis in Grigelis et al. 1977) inthe Furzedown Clays (Q. mariae Zone) of theOxfordian. This is a comparable level to theoccurrence of G. oxfordiana in the Marnes deVillers of the Vaches Noires cliffs of Normandyand subsurface material from near Le Havre(Bignot & Guyader 1966, 1971; Samson et al.1992). All these occurrences are coincident withthe Q. mariae Zone highstand of the earlyOxfordian (Haq et al. 1988; Jacquin et al. 1998)and appear to represent a distinctive pulse ofmigration into the area of NW Europe. Infor-mation from the latest Jurassic and earliestCretaceous is rather limited and the distributionand evolution of the planktonic foraminiferaare poorly known. Sea levels during this timeinterval are, generally, low and many of theshelf areas covered during the Oxfordian and
Kimmeridgian are either non-marine or exposedland. Even in the Valanginian the distribution ofthe planktonic foraminifera (Fig. 6) is essentiallythat seen in the earliest Oxfordian. In theBarremian stage, however, diversificationbegins (Figs 2 and 4) and continues through theAptian and Albian. By the latest Albian (Fig. 7)the near-global distribution of the planktonicforaminifera is established and continuesthrough until the end-Maastrichtian extinctionevent.
It is during the latest Albian that the distinc-tive 'keeled1 morphotypes appear as, prior tothat time, only 'hedbergellid' taxa are known.Within the early Albian a number of lineagesappear (Ticinella, Globigerinelloides, etc.) thatindicate a diversification of the hedbergellidstock, but it is difficult to assess if these taxa weredepth-stratified in the same way as the youngerfaunas (Hart 1999). The single-keeled morpho-types appear in the latest Albian, possiblyassociated with the sea-level changes in the 5.dispar Zone. The twin-keeled morphotypesappear in the latest Cenomanian which, again,marks a time of significant sea-level rise.Throughout the mid- to late Cretaceous thesuccession is punctuated by a number of'events' beginning with the Faroni Event in theBarremian. All of these events (Selli, Paquier,Amadeus, Bonnarelli, etc.) are associated withblack shales and/or mudstones and have oftenbeen described as 'anoxic events'. Their effectson the planktonic foraminifera have beendocumented by a wide range of authors (e.g.Premoli Silva & Sliter 1999) and while thechanges at some of these levels are significant,they cannot be described as controlling theevolution of the planktonic foraminifera (Hart1999; Premoli Silva & Sliter 1999). Hart (1999)showed that there were significant changes tothe fauna at the level of the Bonnarelli event(latest Cenomanian) with a number of extinc-tions and first appearances, but other events(e.g. the Selli event in the earliest Aptian) recordfew, if any, changes.
The palaeolatitudinal distribution of theCretaceous planktonic foraminifera has recentlybeen documented by Hart (2000) and Hudson(2000). This work continues and a series of mapsfor the Jurassic and Cretaceous is currently inproduction. The changes recorded at any of theevents during the Cretaceous cannot, however,be compared to that at the end of the Cretaceous(Hart 1999, fig. 5). At the end of the Maastricht-ian the fauna was decimated and only two orthree survivors provided the foundation for theCenozoic fauna. Only the tiny, hedgergellid taxasurvived and the evolutionary clock is returned

MESOZOIC PLANKTONIC FORAMINIFERA 123
almost to Oxfordian levels. The diversificationof the Cenozoic fauna following the extinctionevent is, however, much more rapid than thatseen in the Mesozoic.
Summary
The origins of the planktonic foraminifera canbe traced back to the Toarcian, although it isonly in the Bathonian that meroplanktonic taxaappear. The inflated O. quadrilobata andPraegubkinella spp. present in Toarcian strataare clearly not fully planktonic and may havebeen benthonic throughout much of their lifecycle. The environmental disruption caused bythe early Toarcian sea-level rise and associated'anoxic event' are potential triggers for thischange in lifestyle as may have been the massiverelease of methane-derived carbon from sub-seagas hydrates in the falciferum Zone of the earlyToarcian (Hesselbo et al 2000).
These early faunas, including the Conoglo-bigerina spp. assemblage in the Bathonian, areall limited to the northern side of Tethys (Figs 3,5 and 6). With such a limited fauna in therestricted areas shown in these maps it isunlikely that many diversification opportunitieswere available, especially in the latest Jurassicand earliest Cretaceous interval which wascharacterized by reduced sea levels. Once oceanfloor production accelerated during the latestBarremian and earliest Aptian and, as a result,sea levels rose globally, the planktonicforaminifera were able to use the newly formedocean basins (and expanded lengths of conti-nental margin) to attain a near-global distri-bution by the mid-Cretaceous (Fig. 7). Thecontinental fragmentation during the Aptianand Albian allowed the hedbergellid faunas toexpand rapidly and as the various ocean basinsbecame connected, the stratification of the watercolumn generated the opportunity for theevolution of a depth-stratified planktonic faunaby the mid- to late Albian (see Hart 2000).
The authors acknowledge the helpful discussions ofthis topic with M. Simmons (Cambridge), M. Caron(Fribourg), A. Gorog (Budapest), R. Wernli(Geneva), J. Gregory (St Albans), A. Henderson(London) and J. Whittaker (London). A. Gorog and J.Svabo are thanked for their help in the field inHungary and J.-P. Berger is thanked for help in thefield in Switzerland. M.B.H. acknowledges a ResearchGrant from the Royal Society while M.J.O. acknow-ledges receipt of a Research Studentship funded by theUniversity of Plymouth and Amerada Hess (London).J. Abraham is thanked for his patient work in prepar-ing the figures.
References
ASCOLI, P. 1976. Foraminiferal and ostracod bio-stratigraphy of the Mesozoic-Cenozoic ScotianShelf, Atlantic Canada. Maritime Sediments,Special Publication, 1,653-771.
BAUMGARTNER, P.O. 1985. Jurassic sedimentary evol-ution and nappe emplacement in the Argon'sPeninsula (Peloponnesus, Greece). Denkschriftender Schweizerischen NaturforschendenGesellschaft, 69.
BIGNOT, G. & GUYADER, J. 1966. Decouverte deForaminiferes planctoniques dans I'Oxfordien duHavre (Seine Maritime). Revue de Micropaleon-tologie, 9,104-110.
BIGNOT, G. & GUYADER, J. 1971. Observations nou-velles sur Globigerina oxfordiana Grigelis. In:FARINACCI , A. (ed.) Proceedings of the SecondPlanktonic Conference, Roma 1970, Volume 1.Edizioni Tecnoscienza, Rome, 79-81.
CARON, M. 1983. Taxonomie et phylogenie de lafamille des Globotruncanidae. Zitteliana, 10.677-681.
CARTER, D. J. & HART, M. B. 1977. Aspects of mid-Cretaceous stratigraphical micropalaeontology.Bulletin of the British Museum, Natural History(Geology), 29,1-135.
DARLING, K. F., KROON, D., WADE, C. M. & LEIGHBROWN, A. J. 1996. Molecular phylogeny of theplanktic Foraminifera. Journal of ForaminiferalResearch, 26, 324-330.
DARLING, K. F., WADE, C. M., KROON, D. & LEIGHBROWN, A. J. 1999a. Planktic foraminiferalmolecular evolution and their polyphyleticorigins from benthic taxa. Marine Micropaleon-tology, 30, 251-266.
DARLING, K. F., WADE, C. M., KROON, D., LEIGHBROWN, A. J. & BIJMA, J. 1999b. The diversity anddistribution of modern planktic foraminiferalsmall subunit ribosomal RNA genotypes andtheir potential as tracers of present and past oceancirculation. Paleoceanography, 14, 3-12.
FUCHS, W. 1967. Uber Ursprung und Phylogenie deTrias- 'Globigerinen' und die Bedeutung DiesesFormenkreises fur das echte Plankton. Verhand-lungen der Geologischen Bundesanstalt, Wien,1967, 135-176.
Funs, W. 1971. Eine alpine Foraminiferenfauna destieferen Mittel-Barreme aus den Drus-bergschichten von Ranzenberg bei Hohenems inVorarlberg. Abhandlungen der GeologischenBundesanstalt, Wien, 27, 1-49.
FUCHS, W. 1973. Ein Beitrag zur Kenntnis der Jura -"Globigerinen und verwandter Formen an Handpolnischen Materials des Callovien und Oxfor-dien. Verhandlungen der Geologischen Bunde-sanstalt, Wien, 1973, 445-487.
FUCHS, W. 1975. Zur Stammesgeschichte der Plank-ton-foraminiferen und verwandter Formen imMesozoikum. Jahrbuch der Geologischen Bunde-sanstalt, Wien, 118, 193-246.
FUCHS, W. 1977. A contribution to the phylogeny ofthe Mesozoic planktonic foraminifera. In: Actedu Vle Colloque Africain de Micropaleontologie.

124 MALCOLM B. HART, MELISSA J. OXFORD & WENDY HUDSON
Annales des Mines et de la Geologie, Tunis, 28.71-74.
GRIGELIS, A. A. 1958. Globigerina oxfordiana sp. n. -an occurrence of Globigerina in the Upper Juras-sic strata of Lithuania. Nauchnye DokladyVysshei Shkoly, Geologo-Geograficheskie Nauki,1958(3), 109-111 (in Russian).
GRIGELIS, A. A., Mesezhinikov. M. S., Yakovleva, S. P.& Kozlova, G. E. 1977. The first finds of plank-tonic foraminifera in the Upper Jurassic of thePechora River Basin. Doklady Akademii NaukSSSR, 233, 926-927 (in Russian).
GOROG. A. 1994. Early Jurassic planktonicforaminifera from Hungary. Micropaleontology,40, 225-260.
HAEUSLER R. 1881. Note sur une zone a globigerinesdans le terrain jurassique de la Suisse. Annales dela Societe Royale Malacologique de Belgique(Serie 3.1). 16,' 188-190.
HAO, B. U, HARDENBOL, J. & VAIL, P. R. 1988.Mesozoic and Cenozoic chronostratigraphy andcycles of sea-level change. In: WILGUS, C. K..HASTINGS, B. S.. KENDALL, C. G. ST., POSAMEN-TIER. H. W.. Ross, C. A. & VAN WAGONER, J. C.(eds) Sea-level Changes: An Integrated Approach,Society of Economic Paleontologists and Miner-alologists. Special Publications, 42, 71-108.
HARRIES. P. J., KAUFFMAN. E. G. & HANSEN, T. A.1996. Models for biotic survival following massextinction. In: HART, M. B. (ed.) Biotic Recoveryfrom Mass Extinction Events, Geological Society,London, Special Publications, 102, 41-60.
HART, M. B. 1999. The evolution and biodiversity ofCretaceous planktonic Foraminiferida. Geobios,32, 247-255.
HART, M. B. 2000. Climatic modelling in theCretaceous using the distribution of planktonicForaminiferida. In: HART, M. B. (Ed.) Climates:Past and Present. Geological Society, London,Special Publications, 181, 33-41.
HESSELBO, S. P., GROCKE, D. R., JENKYNS, H. C.BJERRUM, C. J.. FARRIMOND, P. MORGANS BELL,H. S. & GREEN, O. R. 2000. Massive dissociationof gas hydrate during a Jurassic anoxic event.Nature, 406, 392-395.
HUDSON, W. 2000. A comparison of the Cretaceous andRecent faunas together with their spatial and tem-poral distributions. MSc Thesis, University ofPlymouth.
HYLTON, M. D. 2000. Microfaunal investigation of theEarly Toarcian (Lower Jurassic) extinction eventin N.W.Europe. PhD Thesis, University ofPlymouth.
HYLTON. M. D. & HART, M. B. 2000. Benthicforaminiferal response to Pliensbachian-Toar-cian (Lower Jurassic) sea-level change andoceanic anoxia in NW Europe. GeoResearchForum, 6, 455-462.
JACQUIN, T, DARDEAU, G., DURLET, C., DE GRACIAN-SKY, P.-C. & HANTZPERGUE, P. 1998. The NorthSea Cycle: an overview of 2nd-Order transgres-sive/regressive facies cycles in Western Europe.In: De GRACIANSKY, P.-C., HARDENBOL, J.,JACOUIN, T. & VAIL, P.R. (eds) Mesozoic andCenozoic Sequence Stratigraphy of European
Basins. Society of Economic Paleontologists andMineralogists. Special Publications. 60. 445—466.
KASIMOVA. G. K. & ALIYEVA. D. G. 1984. Planktonicforaminifera of the Middle Jurassic beds of Azer-baijan. Voprosy Paleontologii i Stratigrafi Azer-baijana, 479. 8-19 (in Russian).
KUZNFTSOVA.K. I. & GoRBATCHiK.T. N. 1980. Novien-akhodki planktonnikh foraminifer. vverkhyurskikh otlzhenii Krime. DokladY Acade-mii Nauk, SSSR, 254. 748-751 (in Russian).
LARSON. R. L. 1991a/. Latest pulse of the Earth: evi-dence for a mid-Cretaceous superplume.Geology, 19. 547-550.
LARSON, R. L. 19915. Geological consequences ofsuperplumes. Geology, 19. 963-966.
LARSON. R. L.. FISCHER. A. G.. ERBA. E. & PREMOLISILVA, 1. 1993. Summary of workshop results. In:LARSON. R. L. et al. (eds) Apticore-Albicore: Aworkshop on global events and rhythms of themid-Cretaceous, Perugia. October 1992.
METTRAUX, M.. DUPASOUIER. C. & HOMEWOOD. P.1986. Conditions de depot et diagenese precocedu Toarcien inferieur des Prealpes medianesromandes (Suisse). Documents BRGM, 110.231-237.
METTRAUX. M.. WEISSERT. H. & HOMEWOOD. P. 1989.An oxygen-minimum palaeoceanographic Signalfrom the Early Toarcian cavity fills. Journal of theGeological Society, London, 146. 333-344.
MOROZOVA. V. G. & MOSKALENKO.T. A. 1961. Plank-tonnie foraminiferi progranichnikh otlozheniiBayosskogo i Batskogo yarusov tsentralnogoDagestana (severo-vostochnii Kavkaz) VoprosyMikropaleontologii, 5. 3-30 (in Russian).
O'DoGHERTY. L.. SANDOYAL. J. & VERA. J. A. 2000.Ammonite faunal turnover tracing sea-levelchanges During the Jurassic (Betic Cordillera,southern Spain). Journal of the GeologicalSociety, London, 157. 723-736.
OXFORD. M. J.. GREGORY. F. J.. HART. M. B..HENDERSON. A. S., SIMMONS. M. D. & WATKINSON.M. P. (in press). Jurassic planktonic foraminiferafrom the United Kingdom. Terra Nova.
PAZDROWA, O.1969. Bathonian Globigerina of Poland.Rocznik Polskiego Towarzystwa Geologicznego,39. 41-56.
PREMOLI SILVA, I. & SLITER. W. V. 1999, Cretaceouspaleoceanography: evidence from planktonicforaminiferal evolution. Geological Societv ofAmerica, Special Paper, 332. 301-328.
SAMSON, Y., JANIN. M.-C. & BIGNOT. G. 1992. Lesglobuligerines (Foraminiferes Planctoniques) deI'Oxfordien Inferieur de Villers-sur-Mer (Calva-dos, France) dans leur gisement. Revue de Paleo-biologie, 11,409-431.
SIMMONS, M. D., BOUDAGHER-FADFL. M. K.. BANNER.F. T. & WHITTAKER, J. E. 1997. The JurassicFavusellacea, the earliest Globigerinina, In:BOUDAGHER-FADEL, M. K.. BANNER, F. T. &WHITTAKER. J. E. (eds) The Early EvolutionaryHistory of Planktonic Foraminifera. Chapman &Hall. London. 17-51.
WEIDMANN, M. 1993. Atlas geologique de la Suisse, 1:25000. Notice explicative, Feuille 1244. Chatel-St-Denis.

MESOZOIC PLANKTONIC FORAMINIFERA 125
WEIDMANN, M., HOMEWOOD, P., MOREL, R. ET AL.1993. Atlas Geologique de la Suisse, Feuille1244, Chatel-St-Denis (Feuille 92 de L'Atlas).Publie par le Service hydrologique et geologiquenational.
WERNLI, R. 1987. Les protoglobigerines(Foraminiferes) du Bajocien inferieur des Sofs(Rif, Maroc). Eclogae Geologicae Helvetiae, 80,817-829
WERNLI, R. 1995. Les Foraminiferes globigeriniformes
(Oberhauserellidae) du Toarcien Inferieur deTeysachaux (Prealpes medianes, Fribourg,Suisse). Revue de Paleobiologie, 14, 257-269.
WERNLI, R. & GOROG, A. 1999. Protoglobigerinids(Foraminifera) acid extracted from Bajocianlimestones (Hungary). Revista Espanola deMicropaleontologia, 31, 419-426.
WERNLI, R. & GOROG, A. 2000. Determination ofBajocian protoglobigerinids (Foraminifera) inthin sections. Revue de Paleobiologie, 19,399-407.

This page intentionally left blank

Opening of the Hispanic Corridor and Early Jurassic bivalvebiodiversity
MARTIN ABERHANMuseum fur Naturkunde, Institut fur Palaontologie, Invalidenstr. 43, D-10115
Berlin, Germany, (e-mail: [email protected]
Abstract: The Hispanic Corridor is a postulated marine seaway linking the eastern Pacificand western Tethyan oceans as early as Early Jurassic times. Two existing hypothesesrelate the Pliensbachian-Toarcian bivalve extinction and recovery to immigration ofbivalve species through the Hispanic Corridor. The extinction hypothesis implies that, inSouth America, the Pliensbachian-Toarcian extinction can be partly explained by theimmigration of bivalves through the Hispanic Corridor and subsequent competitivereplacement. The recovery hypothesis states that, in NW Europe, the renewed rise in diver-sity in the late Toarcian/Aalenian was largely a consequence of immigration of taxa fromAndean South America via the Hispanic Corridor.
To test these hypotheses, I calculated immigration and origination rates of bivalves permillion years. In both regions, early Pliensbachian to Aalenian immigration rates remainedat low levels, thus disproving both hypotheses. By comparison, the origination of newspecies generally played a much more important role than immigration in controllingoverall diversity of both regions. Future research should investigate if this is a more generalpattern in the recovery of post-extinction biotas.
The apparently global Pliensbachian-Toarcian diversity crisis may be best explained bya combination of physicochemical factors, invoking intense volcanism, sea-level highstandand widespread anoxia, as well as biological factors. Recovery from this mass extinctioncommenced when origination rates increased again, which, in the Andean basins, was inthe Aalenian and in NW Europe, the late Toarcian.
Investigations of the history of diversity and ofevolutionary processes that generate biodiver-sity not only improve our knowledge of the fossilpast, but may contribute significantly to ourunderstanding of the course and consequencesof the modern biodiversity crisis. A promisingpalaeontological approach is the analysis of thegeographic variation of patterns observedduring extinction episodes and their subsequentrecoveries. In the present study, I analyse thebiogeography of Early Jurassic bivalves with theaim of testing two hypotheses existing in theliterature. These hypotheses relate the Pliens-bachian-Toarcian mass extinction to theopening of a marine connection (the so-calledHispanic Corridor) between the eastern Pacificand western Tethyan oceans. The first hypothe-sis states that, in South America, the preferentialdisappearance of endemic bivalves across thePliensbachian-Toarcian boundary can be partlyexplained by immigration of cosmopolitanspecies via the Hispanic Corridor and subse-quent competitive replacement (Aberhan &Fursich 1997). The second hypothesis states that,in NW Europe, the renewed rise in bivalvediversity in the late Toarcian/Aalenian waslargely a consequence of immigration of taxafrom Andean South America through the
Hispanic Corridor, presumably filling the eco-space vacated during the extinction event(Hallam 1983, 1996; Hallam & Wignall 1997). Idemonstrate that neither of the two hypothesescan be sustained. Whilst the extinction may bebest explained by a combination of globalpalaeoenvironmental changes and biologicalprocesses, the recovery appears to be largelycontrolled by increasing within-region origina-tion rates rather than immigration of alreadyexisting species.
Early Jurassic bivalve biodiversity and theHispanic CorridorTheir high abundance and taxonomic diversitysince early Mesozoic times, along with anexcellent fossil record, make bivalves an idealgroup for palaeodiversity analysis in the marinerealm. At the species level, Early Jurassicbivalve diversity is known in detail from NWEuropean epicontinental seas (Hallam 1986,1987) and from western South America(Damborenea 1996; Aberhan & Fiirsich 1997,2000), where bivalves occur very abundantly in aseries of backarc basins, the so-called Andeanbasins. In both regions, bivalves experienced a
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194,127-139.0305-8719/02/$15.00 © The Geological Society of London 2002.
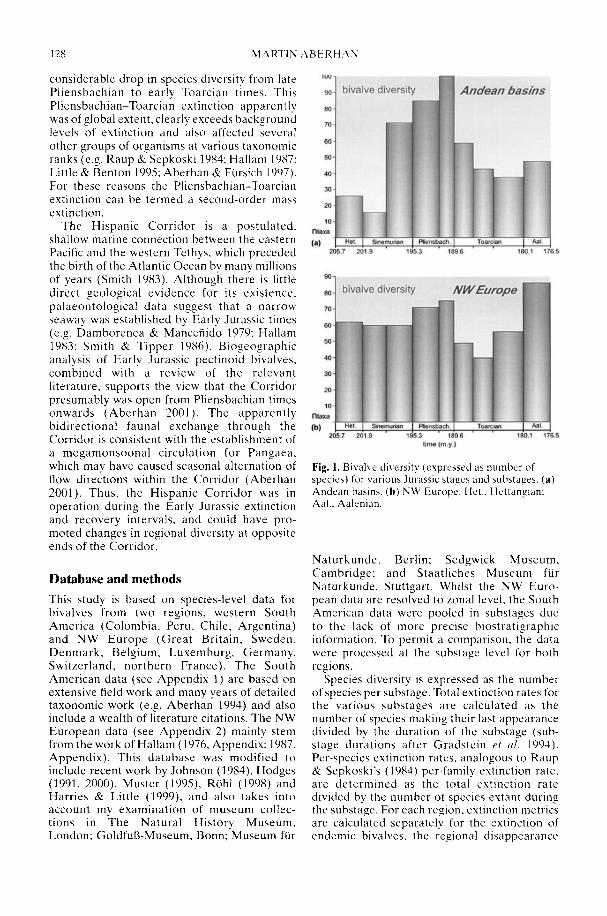
128 MARTIN ABERHAN
considerable drop in species diversity from latePliensbachian to early Toarcian times. ThisPliensbachian-Toarcian extinction apparentlywas of global extent, clearly exceeds backgroundlevels of extinction and also affected severalother groups of organisms at various taxonomicranks (e.g. Raup & Sepkoski 1984; Hallam 1987;Little & Benton 1995; Aberhan & Fursich 1997).For these reasons the Pliensbachian-Toarcianextinction can be termed a second-order massextinction.
The Hispanic Corridor is a postulated,shallow marine connection between the easternPacific and the western Tethys, which precededthe birth of the Atlantic Ocean by many millionsof years (Smith 1983). Although there is littledirect geological evidence for its existence,palaeontological data suggest that a narrowseaway was established by Early Jurassic times(e.g. Damborenea & Mancenido 1979; Hallam1983; Smith & Tipper 1986). Biogeographicanalysis of Early Jurassic pectinoid bivalves,combined with a review of the relevantliterature, supports the view that the Corridorpresumably was open from Pliensbachian timesonwards (Aberhan 2001). The apparentlybidirectional faunal exchange through theCorridor is consistent with the establishment ofa megamonsoonal circulation for Pangaea,which may have caused seasonal alternation offlow directions within the Corridor (Aberhan2001). Thus, the Hispanic Corridor was inoperation during the Early Jurassic extinctionand recovery intervals, and could have pro-moted changes in regional diversity at oppositeends of the Corridor.
Database and methodsThis study is based on species-level data forbivalves from two regions, western SouthAmerica (Colombia, Peru, Chile, Argentina)and NW Europe (Great Britain, Sweden,Denmark, Belgium, Luxemburg, Germany,Switzerland, northern France). The SouthAmerican data (see Appendix 1) are based onextensive field work and many years of detailedtaxonomic work (e.g. Aberhan 1994) and alsoinclude a wealth of literature citations. The NWEuropean data (see Appendix 2) mainly stemfrom the work of Hallam (1976, Appendix; 1987,Appendix). This database was modified toinclude recent work by Johnson (1984), Hodges(1991, 2000), Muster (1995), Rohl (1998) andHarries & Little (1999), and also takes intoaccount my examination of museum collec-tions in The Natural History Museum,London; GoldfuB-Museum, Bonn; Museum fur
Fig. 1. Bivalve diversity (expressed as number ofspecies) for various Jurassic stages and substages. (a)Andean basins, (b) NW Europe. Het.. Hettangian;Aal.. Aalenian.
Naturkunde, Berlin; Sedgwick Museum,Cambridge; and Staatliches Museum furNaturkunde, Stuttgart. Whilst the NW Euro-pean data are resolved to zonal level, the SouthAmerican data were pooled in substages dueto the lack of more precise biostratigraphicinformation. To permit a comparison, the datawere processed at the substage level for bothregions.
Species diversity is expressed as the numberof species per substage. Total extinction rates forthe various substages are calculated as thenumber of species making their last appearancedivided by the duration of the substage (sub-stage durations after Gradstein et al. 1994).Per-species extinction rates, analogous to Raup& Sepkoskrs (1984) per-family extinction rate,are determined as the total extinction ratedivided by the number of species extant duringthe substage. For each region, extinction metricsare calculated separately for the extinction ofendemic bivalves, the regional disappearance
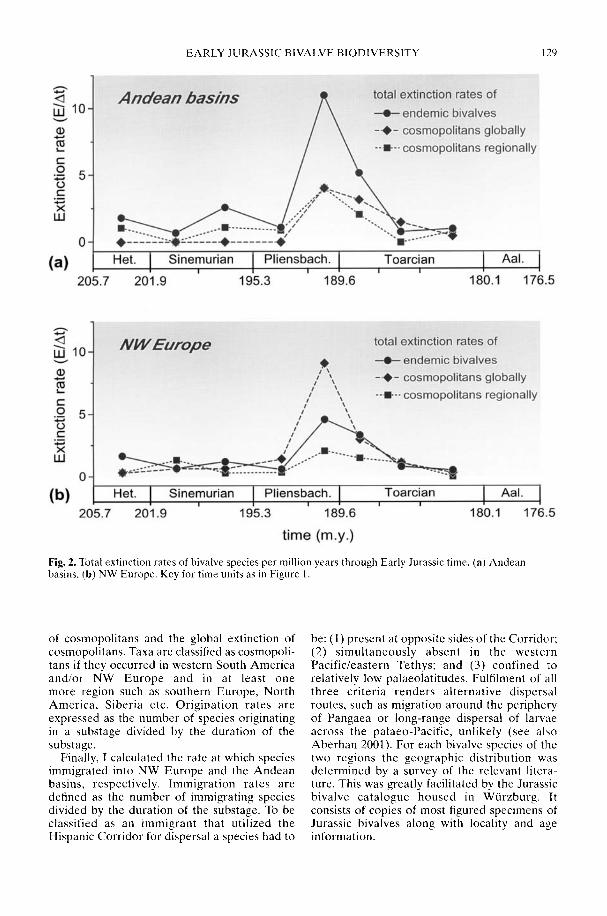
EARLY JURASSIC BIVALVE BIODIVERSITY 129
Fig. 2. Total extinction rates of bivalve species per million years through Early Jurassic time, (a) Andeanbasins, (b) NW Europe. Key for time units as in Figure 1.
of cosmopolitans and the global extinction ofcosmopolitans. Taxa are classified as cosmopoli-tans if they occurred in western South Americaand/or NW Europe and in at least onemore region such as southern Europe, NorthAmerica, Siberia etc. Origination rates areexpressed as the number of species originatingin a substage divided by the duration of thesubstage.
Finally, I calculated the rate at which speciesimmigrated into NW Europe and the Andeanbasins, respectively. Immigration rates aredefined as the number of immigrating speciesdivided by the duration of the substage. To beclassified as an immigrant that utilized theHispanic Corridor for dispersal a species had to
be: (1) present at opposite sides of the Corridor;(2) simultaneously absent in the westernPacific/eastern Tethys; and (3) confined torelatively low palaeolatitudes. Fulfilment of allthree criteria renders alternative dispersalroutes, such as migration around the peripheryof Pangaea or long-range dispersal of larvaeacross the palaeo-Pacific, unlikely (see alsoAberhan 2001). For each bivalve species of thetwo regions the geographic distribution wasdetermined by a survey of the relevant litera-ture. This was greatly facilitated by the Jurassicbivalve catalogue housed in Wurzburg. Itconsists of copies of most figured specimens ofJurassic bivalves along with locality and ageinformation.
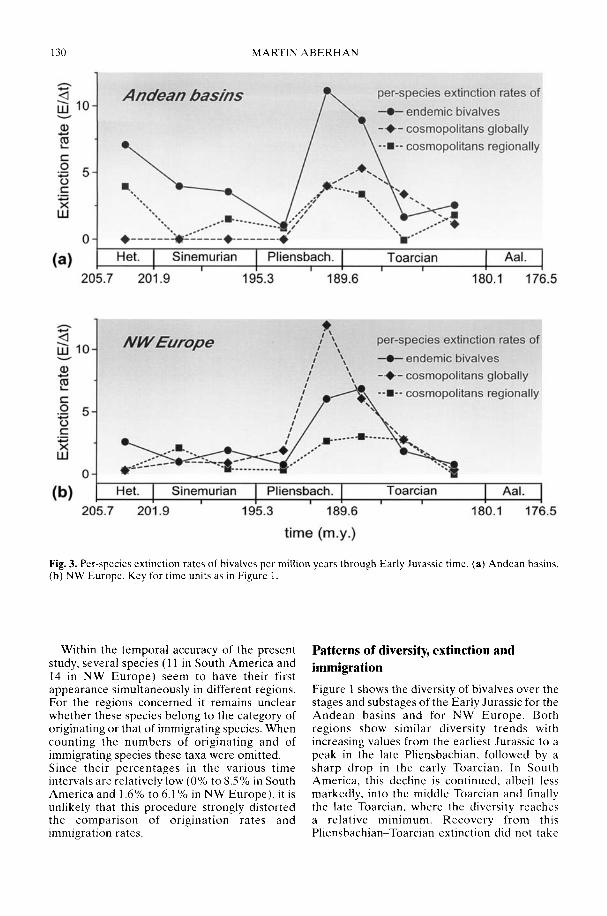
130 MARTIN ABERHAN
Fig. 3. Per-species extinction rates of bivalves per million years through Early Jurassic time, (a) Andean basins,(b) NW Europe. Key for time units as in Figure 1.
Within the temporal accuracy of the presentstudy, several species (11 in South America and14 in NW Europe) seem to have their firstappearance simultaneously in different regions.For the regions concerned it remains unclearwhether these species belong to the category oforiginating or that of immigrating species. Whencounting the numbers of originating and ofimmigrating species these taxa were omitted.Since their percentages in the various timeintervals are relatively low (0% to 8.5% in SouthAmerica and 1.6% to 6.1% in NW Europe), it isunlikely that this procedure strongly distortedthe comparison of origination rates andimmigration rates.
Patterns of diversity, extinction andimmigration
Figure 1 shows the diversity of bivalves over thestages and substages of the Early Jurassic for theAndean basins and for NW Europe. Bothregions show similar diversity trends withincreasing values from the earliest Jurassic to apeak in the late Pliensbachian, followed by asharp drop in the early Toarcian. In SouthAmerica, this decline is continued, albeit lessmarkedly, into the middle Toarcian and finallythe late Toarcian, where the diversity reachesa relative minimum. Recovery from thisPliensbachian-Toarcian extinction did not take
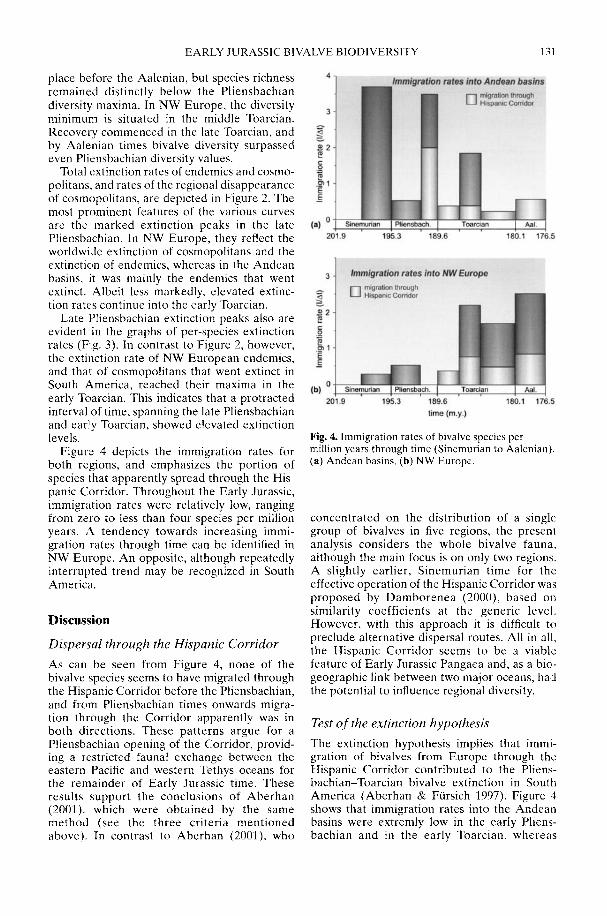
EARLY JURASSIC BIVALVE BIODIVERSITY 131
place before the Aalenian, but species richnessremained distinctly below the Pliensbachiandiversity maxima. In NW Europe, the diversityminimum is situated in the middle Toarcian.Recovery commenced in the late Toarcian, andby Aalenian times bivalve diversity surpassedeven Pliensbachian diversity values.
Total extinction rates of endemics and cosmo-politans, and rates of the regional disappearanceof cosmopolitans, are depicted in Figure 2. Themost prominent features of the various curvesare the marked extinction peaks in the latePliensbachian. In NW Europe, they reflect theworldwide extinction of cosmopolitans and theextinction of endemics, whereas in the Andeanbasins, it was mainly the endemics that wentextinct. Albeit less markedly, elevated extinc-tion rates continue into the early Toarcian.
Late Pliensbachian extinction peaks also areevident in the graphs of per-species extinctionrates (Fig. 3). In contrast to Figure 2, however,the extinction rate of NW European endemics,and that of cosmopolitans that went extinct inSouth America, reached their maxima in theearly Toarcian. This indicates that a protractedinterval of time, spanning the late Pliensbachianand early Toarcian, showed elevated extinctionlevels.
Figure 4 depicts the immigration rates forboth regions, and emphasizes the portion ofspecies that apparently spread through the His-panic Corridor. Throughout the Early Jurassic,immigration rates were relatively low, rangingfrom zero to less than four species per millionyears. A tendency towards increasing immi-gration rates through time can be identified inNW Europe. An opposite, although repeatedlyinterrupted trend may be recognized in SouthAmerica.
Discussion
Dispersal through the Hispanic Corridor
As can be seen from Figure 4, none of thebivalve species seems to have migrated throughthe Hispanic Corridor before the Pliensbachian,and from Pliensbachian times onwards migra-tion through the Corridor apparently was inboth directions. These patterns argue for aPliensbachian opening of the Corridor, provid-ing a restricted faunal exchange between theeastern Pacific and western Tethys oceans forthe remainder of Early Jurassic time. Theseresults support the conclusions of Aberhan(2001), which were obtained by the samemethod (see the three criteria mentionedabove). In contrast to Aberhan (2001), who
Fig. 4. Immigration rates of bivalve species permillion years through time (Sinemurian to Aalenian).(a) Andean basins, (b) NW Europe.
concentrated on the distribution of a singlegroup of bivalves in five regions, the presentanalysis considers the whole bivalve fauna,although the main focus is on only two regions.A slightly earlier, Sinemurian time for theeffective operation of the Hispanic Corridor wasproposed by Damborenea (2000), based onsimilarity coefficients at the generic level.However, with this approach it is difficult topreclude alternative dispersal routes. All in all,the Hispanic Corridor seems to be a viablefeature of Early Jurassic Pangaea and, as a bio-geographic link between two major oceans, hadthe potential to influence regional diversity.
Test of the extinction hypothesis
The extinction hypothesis implies that immi-gration of bivalves from Europe through theHispanic Corridor contributed to the Pliens-bachian-Toarcian bivalve extinction in SouthAmerica (Aberhan & Fursich 1997). Figure 4shows that immigration rates into the Andeanbasins were extremly low in the early Pliens-bachian and in the early Toarcian, whereas

132 MARTIN ABERHAN
Table 1. Relative abundance of selected Early Jurassic bivalve species in northern Chile.
Species
Camptonectes auritus (Schlotheim)Psendopecten equivalvis (J. Sowerby)Modiolus ventricosns (Roemer)Plagiostoma giganteum J. SowerbyPropeamusium pumilum (Lamarck)Wevla alala (von Buch)
Nsanplcs
744_
(5)158
% samples
3.52.02.0-
(2.4)78.2
Relativeabundance ( % )
0.5-3.70.5-3.50.8-3.0-
(0.7-28.6)0.5-69.4
Rel. abundance(mean)
2.01.41.8-
(15.0)10.0
Samples • number of samples in which species occurs (total number of samples = 202); %samples. percentage ofEarly Jurassic samples in which species occurs; relative abundance (%). range of relative abundances ofspecies in samples; rel. abundance (mean), mean of relative abundances of species. Numbers inparentheses indicate data that include information from samples with less than 70 individuals. Forcomparison, the abundant bivalve Weyla alaia has been included. Data based on Aberhan (1992) andunpublished information. For discussion see text.
moderately high values characterize the latePliensbachian. However, during the criticaltime interval, from the early Pliensbachian tothe early Toarcian, only five species possiblyimmigrated through the Corridor (Table 1).Compared to 57 species that went extinct duringthe same time interval, this group of five speciesseems to be too small to contribute significantlyto the extinction.
It could be argued that a single, eurytopic andabundant species might have the potential tocause extinction of many other forms. Table 1summarizes relative abundance data of species,including the five species in question. Thesedata were obtained through detailed palaeo-ecological analysis of benthic associations fromnorthern Chile (Aberhan 1992,1993). It becomesevident that the first three species of Table 1occurred in only a few samples (less than 4% ofall quantitative samples) and in low abundances(on average 2% or less of the total fauna of asample). Another species, Plagiostoma gigan-teum, occurred in very low numbers in a fewsamples, which were too small to be included inthe statistical analysis of Aberhan (1992, 1993).For comparison, the ubiquitous bivalve speciesWeyla alata has been included in Table 1. Itoccurred in nearly 80% of the samples and is oneorder of magnitude more common than the otherfaunal elements (Table 1). In the Andean basins,the four species mentioned above can thereforebe classified as rare and geographically restrictedelements of Early Jurassic benthic communities,and it is indeed very unlikely that they causedcompetitive replacement.
An exception is the pectinoid bivalvePropeamitssium pumilum, which becomeslocally abundant (up to nearly 30%) in
low-diversity, oxygen-controlled environments(Table 1; Aberhan 1993). Its occurrence in theearly Toarcian coincided with the disappearanceof the morphologically related species Posi-donotis semiplicata (Hyatt) , another low-oxygen-tolerant flat clam. P. semiplicata firstappeared in the upper Sinemurian of severalallochthonous terranes of the North AmericanCordillera and by late Pliensbachian times hadspread to the Andean basins, where it persistedas a dominant benthic element of the dysaerobicbiofacies into the early Toarcian (Aberhan &Palfy 1996). The disappearance of P. semiplicatain the Andean basins, followed by the appear-ance of P. pumilum possibly represents a caseof competitive replacement, but the SouthAmerican age ranges of both species suggestthat competition was pre-emptive rather thandisplacive in the sense of Hallam (1990). Also,competition with P. pumilum cannot explain theearly Toarcian extinction of P. semiplicata inNorth America, since P. pumilum seems to beabsent from that region.
It is beyond the scope of the present study toassess the immigration of predators through anopening Hispanic Corridor and their potentialimpact on the regional diversity of bivalves.Nevertheless it is worth mentioning that preda-tion pressure apparently increased strikingly inthe Late Cretaceous and Cenozoic followingadaptive radiation of neogastropods, crabs andteleost fishes (Vermeij 1977), and in this respectpost-dates the Early Jurassic diversity crisis by100 million years and more. In summary, immi-gration into the Andean basins of bivalvespecies through the Hispanic Corridor, followedby competitive replacement, obviously did notcontribute to the observed drop in diversity.
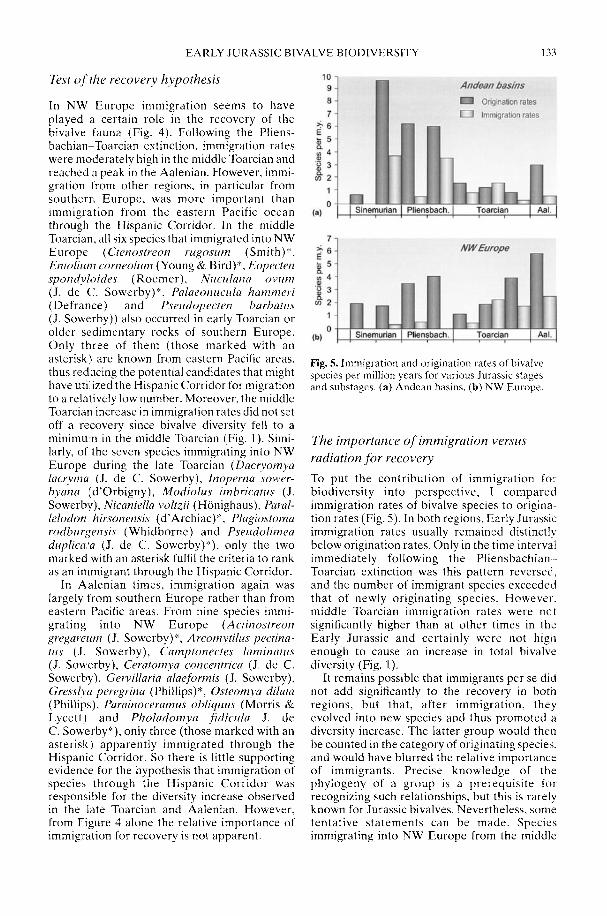
EARLY JURASSIC BIVALVE BIODIVERSITY 133
Test of the recovery hypothesis
In NW Europe immigration seems to haveplayed a certain role in the recovery of thebivalve fauna (Fig. 4). Following the Pliens-bachian-Toarcian extinction, immigration rateswere moderately high in the middle Toarcian andreached a peak in the Aalenian. However, immi-gration from other regions, in particular fromsouthern Europe, was more important thanimmigration from the eastern Pacific oceanthrough the Hispanic Corridor. In the middleToarcian, all six species that immigrated into NWEurope (Ctenostreon rugosum (Smith)*,Entolium corneolum (Young & Bird)*, Eopectenspondyloides (Roemer), Nuculana ovum(J. de C. Sowerby)*, Palaeonucula hammeri(Defranee) and Pseudopecten barbatus(J. Sowerby)) also occurred in early Toarcian orolder sedimentary rocks of southern Europe.Only three of them (those marked with anasterisk) are known from eastern Pacific areas,thus reducing the potential candidates that mighthave utilized the Hispanic Corridor for migrationto a relatively low number. Moreover, the middleToarcian increase in immigration rates did not setoff a recovery since bivalve diversity fell to aminimum in the middle Toarcian (Fig. 1). Simi-larly, of the seven species immigrating into NWEurope during the late Toarcian (Dacryomyalacryma (J. de C. Sowerby), Inoperna sower-byana (d'Orbigny), Modiolus imbricatus (J.Sowerby), Nicaniella voltzii (Honighaus), Paral-lelodon hirsonensis (d'Archiac)*, Plagiostomarodburgensis (Whidborne) and Pseudolimeaduplicata (J. de C. Sowerby)*), only the twomarked with an asterisk fulfil the criteria to rankas an immigrant through the Hispanic Corridor.
In Aalenian times, immigration again waslargely from southern Europe rather than fromeastern Pacific areas. From nine species immi-grating into NW Europe (Actinostreongregareum (J. Sowerby)*, Arcomytilus pectina-tus (J. Sowerby), Camptonectes laminatus(J. Sowerby), Ceratomya concentrica (J. de C.Sowerby), Gervillaria alaeformis (J. Sowerby),Gresslya peregrina (Phillips)*, Osteomya dilata(Phillips), Parainoceramus obliquus (Morris &Lycett) and Pholadomya fidicula J. deC. Sowerby*), only three (those marked with anasterisk) apparently immigrated through theHispanic Corridor. So there is little supportingevidence for the hypothesis that immigration ofspecies through the Hispanic Corridor wasresponsible for the diversity increase observedin the late Toarcian and Aalenian. However,from Figure 4 alone the relative importance ofimmigration for recovery is not apparent.
Fig. 5. Immigration and origination rates of bivalvespecies per million years for various Jurassic stagesand substages. (a) Andean basins, (b) NW Europe.
The importance of immigration versusradiation for recovery
To put the contribution of immigration forbiodiversity into perspective, I comparedimmigration rates of bivalve species to origina-tion rates (Fig. 5). In both regions, Early Jurassicimmigration rates usually remained distinctlybelow origination rates. Only in the time intervalimmediately following the Pliensbachian-Toarcian extinction was this pattern reversed,and the number of immigrant species exceededthat of newly originating species. However,middle Toarcian immigration rates were notsignificantly higher than at other times in theEarly Jurassic and certainly were not highenough to cause an increase in total bivalvediversity (Fig. 1).
It remains possible that immigrants per se didnot add significantly to the recovery in bothregions, but that, after immigration, theyevolved into new species and thus promoted adiversity increase. The latter group would thenbe counted in the category of originating species,and would have blurred the relative importanceof immigrants. Precise knowledge of thephylogeny of a group is a prerequisite forrecognizing such relationships, but this is rarelyknown for Jurassic bivalves. Nevertheless, sometentative statements can be made. Speciesimmigrating into NW Europe from the middle

134 MARTIN ABERHAN
Toarcian to Aalenian belong to 22 genera, andspecies originating in the same time interval aredistributed over 35 genera. Since both groupshave only four genera in common (Actinostreon,Gervillaria, Pholadomya and Plagiostoma) itseems unlikely that immigrants gave rise tomany new species. Moreover, these four generaare not unusually rich in species and compriseonly between one and two species each. In theAndean basins, the corresponding numbers areseven genera for immigrating species, 15 generafor originating species, and the two groups donot share a single genus.
These results demonstrate that in both regionsthe origination of new species generally played amuch more important role than immigration incontrolling overall diversity. Recovery from thePliensbachian-Toarcian mass extinction com-menced when origination rates increasedagain, which, in the Andean basins, was in theAalenian and in NW Europe started in the lateToarcian. Immigration in general, and immi-gration through the Hispanic Corridor inparticular, cannot explain the observed recoveryin diversity.
ConclusionsThe aim of the study was to test two hypotheses,which relate extinction and recovery of EarlyJurassic bivalves to immigration of speciesthrough the Hispanic Corridor. Both hypothesesfailed the test: the Pliensbachian-Toarcianbivalve extinction in South America was notcaused by the immigration of species throughthe Hispanic Corridor and subsequent com-petitive replacement; and recovery from thePliensbachian-Toarcian extinction in NWEurope was not a consequence of the immi-gration of species from the eastern Pacificthrough the Hispanic Corridor.
Concerning the globally increased extinctionrates of bivalves across the Pliensbachian-Toarcian boundary, the synchrony of the bioticcrisis and voluminous continental flood basalteruptions, sea-level highstand, widespreadanoxia and possibly also massive release ofmethane from gas hydrates suggests causal linksof these events (Palfy & Smith 2000; Hesselbo etal. 2000). In addition to physicochemicalchanges, the drop in species diversity may havebeen amplified by biological factors. Thus, in theAndean basins, the extinction peak of endemicbivalves in the late Pliensbachian is preceded byan origination peak in the early Pliensbachian.The time span between these peaks (5,7 Ma)closely corresponds to the longevity peak ofJurassic endemic bivalves (4-6 Ma), and permits
a causal relationship between these two features(Aberhan & Fursich 2000).
Both analysed regions differ in the dynamicsof originating and immigrating clades, andprovide evidence for the biogeographic com-plexity of biotic recoveries. However, a commonfeature of both regions is the observationthat recovery patterns of bivalves from thePliensbachian-Toarcian mass extinction werelargely controlled by increasing origination ratesrather than immigration. Other studies showedthat immigration intensity during the recoveryphase may vary from basin to basin. Forexample, when analysing geographic variationsin the molluscan recovery from the latest-Cretaceous extinction, Jablonski (1998)detected a significantly larger proportion ofimmigrants in the North American Gulf Coastbiota as compared to three other biogeographicprovinces. Future research should furtherinvestigate the relative importance of immi-gration versus radiation for the recovery ofpost-extinction biotas, and whether or notpatterns differ between first-order and second-order mass extinctions.
I thank J. Palfy, Berlin, for critically reading themanuscript and M. Gahr. Wurzburg. for providingunpublished information on the stratigraphic distri-bution of Early Jurassic bivalves from Spain andPortugal. The comments by the journal refereesC. Little and S. Damborenea are greatly appreciated.This study was financially supported by grants fromthe Deutsche Forschungsgemeinschaft (Ab 109/1-1.Ab 109/2-1), which are acknowledged with gratitude.
Appendix 1Stratigraphic distribution of Hettangian to Aalenianbivalves from the Andean basins.
Taxon Range
Palaeonucula cuevitana Aberhan HetPalaeoneilo elliptica (Goldfuss) E.Plb - L.PlbNuculana ovum (J. de C. Sowerby) L.Plb - L.ToaParallelodon groeberi Damborenea E.Plb - M.ToaParallelodon aff, groeberi Het
DamboreneaParallelodon hirsonensis (d'Archiac) L.Sin - L.PlbParalleodon riccardii Damborenea E.Plb - L.PlbParallelodon sp. L.Plb - E.ToaGrammatodon concinnus (Phillips) E.Toa - L.ToaGrammatodon costulatus (Leanza) E.Plb - L.PlbGrammatodon sulcatus Aberhan L.Sin - L.SinGrammatodon sparsicosta Aal
(Gottsche)Grammatodon cf. toyorensis L.Plb
HayamiCucullaea jaworskii Leanza E.Plb - L.PlbCucullaea rothi Leanza E.Plb - E.ToaCucullaeal sp. L.Plb - L.ToaParainoceramus apollo (Leanza) L.Sin - L.Plb

EARLY JURASSIC BIVALVE BIODIVERSITY 135
Parainoceramus? sp. - HetPteria? sp. HetPteroperna sp. L.Sin - E.PlbAguilerella kobyi (de Loriol) L.Sin - L.PlbBakevellia waltoni (Lycett) L.Sin - L.PlbGervillaria alaeformis (J. Sowerby) M.Toa - AalGervillaria ashcroftensis (Crickmay) L.Plb - E.ToaGervillaria hartmanni (Munster) L.Sin - L.ToaGervillaria pallas (Leanza) L.Sin - E.ToaGervillella araucana Damborenea L.Sin - L.PlbGervillia cf. dundriensis Cox L.Sin - E.ToaLithiotis cf. gregarius M.Toa - M.Toa
(Lupher & Packard)Isognomon Jupiter (Leanza) L.Plb - AalGervilleioperna turgida (Leanza) L.Sin - E.ToaGervilleioperna aurita Aal
Aberhan & HillebrandtPulvinites liasicus Damborenea L.Plb - L.PlbBositra buchi (Roemer) M.Toa - AalPosidonotis semiplicata (Hyatt) L.Plb - E.ToaPinna cf. folium Young & Bird L.Sin - E.ToaPinna cf. radiata Miinster L.Plb - L.PlbAntiquilima cf. nagatoensis Hayami L.Sin - E.ToaAntiquilima sp. HetCtenostreon paucicostatum Leanza E.Plb - E.ToaCtenostreon cf. rugosum (Smith) L.Sin - AalPlagiostoma giganteum J. Sowerby L.Plb - L.ToaPlagiostoma punctatum J. Sowerby L.Plb - L.PlbPlagiostoma sp. A Het - L.ToaPlagiostoma sp. B L.Sin - L.SinPlagiostoma sp. C AalPseudolirnea duplicata L.Sin - Aal
(J. de C. Sowerby)Pseiidolirnea hettangiensis Het
(Terquem)Plicatula spinosa (J. Sowerby) L.Sin - L.SinPlicatula rapa Bayle & Coquand L.Sin - E.ToaActinostreon costatum L.Sin - E.Plb
(J. de C. Sowerby)Actinostreon longistriatum L.Sin - E.Toa
(Jaworski)Actinostreon solitarium E.Plb - Aal
(J. de C. Sowerby)Gryphaea darwini Forbes Het - L.SinGryphaea cf. dumortieri Joly L.Sin - L.PlbGryphaea sp. E.Toa - L.ToaGryphaea cf. dilobotes Duff AalGryphaea latior Steinmann L.Sin - L.PlbGryphaea tricarinata Philippi E.Sin - L.SinNanogyra? cf. auricularis (Munster) L.Sin - L.PlbOtapiria neuquensis Damborenea L.Sin - E.PlbOtapiria pacifica Het - E.Sin
Covacevich & EscobarOxytoma inequivalvis (J. Sowerby) Het - AalOxytoma cf. cygnipes Young & Bird HetOxytoma (Palmoxytoma) sp. HetArctotis? frenguellii (Damborenea) L.Toa - AalMeleagrinella cf. echinata (Smith) AalPlacunopsis radiata (Phillips) L.Sin - L.PlbPropeamussium pumilum E.Toa - Aal
(Lamarck)Kolymonectes weaveri Damborenea E.Plb - L.PlbKolymonectes sp. L.Sin - L.SinEntolium corneolum Het - Aal
(Young & Bird)
Agerchlamys wunschae (Marwick) Het - L.PlbCamptonectes auritus (Schlotheim) L.Plb - AalChlamys textoria (Schlotheim) Het - Aal'Chlamys' cf. tingensis (Tilmann) Het - L.PlbChlamys valoniensis (Defrance) HetEopecten abjectus (Phillips) L.Toa - AalEopecten hartzi (Rosenkrantz) E.Plb - E.ToaEopecten velatus (Goldfuss) Het - AalEopecten? sp. E.Plb - E.PlbPseudopecten equivalvis L.Plb - E.Toa
(J. Sowerby)Radulonectites sosneadoensis E.Plb - L.Plb
(Weaver)Weyla alata (Buch) L.Sin - E.ToaWeyla bodenbenderi (Behrendsen) L.Sin - E.ToaWeyla (Weyla?) sp. HetWeyla unca (Philippi) L.Sin - E.ToaWeyla (Lywea?) sp. HetLycettia hypertrigona Damborenea L.Sin - E.ToaModiolus baylei (Philippi) L.Sin - E.ToaModiolus chilensis Aberhan E.Plb - L.PlbModiolus gerthi Damborenea L.Plb - M.ToaModiolus giganteus Quenstedt L.Sin - L.PlbModiolus imbricatus J. Sowerby M.Toa - AalModiolus lonsdalei L.Plb - Aal
(Morris & Lycett)Modiolus ventricosus Roemer L.Plb - L.Plb"Modiolus^ sp. AalPalaeopharus? sp. L.Plb - L.PlbGroeberella neuquensis (Groeber) L.Sin - AalTrigonia sp. A M.Toa - M.ToaTrigonia sp. B M.Toa - M.ToaTrigonia sp. C AalNeuquenitrigonia huenickeni Aal
(Leanza & Garate)Neiiquenitrigonia sp. nov. M.Toa - M.ToaProsogyrotrigonia sp. nov. Het - E.SinProsogyrotrigonia sp. A E.Sin - E.SinProsogyrotrigonia? sp. B L.Sin - L.SinProsogyrotrigonia? sp. C Het - E.SinFrenguelliella chubutensis (Feruglio) E.Plb - L.PlbFrenguelliella inexspectata Jaworski E.Plb - L.PlbFrenguelliella poultoni Leanza L.Sin - L.PlbFrenguelliella tapiai Lambert L.Plb - E.ToaJaworskiella burckhardti (Jaworski) E.Plb - L.PlbJaworskiella gryphitica (Moricke) L.Sin - L.SinJaworskiella sp. L.Sin - L.SinQuadratojaworskiella pustulata E.Plb - L.Plb
Reyes & PerezQuadratojaworskiella sp. nov. Het - L.SinLaevitrigoniinae gen. et sp. nov. L.Sin - L.SinPsilotrigonia sp. nov. L.Sin - E.PlbMyophorella araucana (Leanza) E.Plb - E.ToaMyophorella catenifera (Hupe) E.Plb - L.PlbMyophorella cf. tuberculata (Agassiz) E.Plb - L.PlbMyophorella sp. nov. A AalMyophorella sp. nov. B M.Toa - L.ToaMyophorella sp. A AalMyophorella sp. B L.Plb - L.PlbScaphorella sp. nov. AalVaugonia cf. substriata M.Toa - M.Toa
(Burmeister & Giebel)Vaugonia sp. nov. L.Sin - E.ToaVaugonia sp. A M.Toa - M.ToaVaugonia sp. B L.Plb - L.Plb

136 MARTIN ABERHAN
Vaitgonia sp. C L.Toa - L.Toa"Lucina chubutensis Wahnish L.Sin - L.Plb'Lucina feruglioli Wahnish E.Plb - L.PlbMesomiltha cf. bellona (D'Orbigny) L.Sin - AalMesomiltha huayquimili (Leanza) E.Plb - L.ToaMesornilthal sp. AalMactromya cardioideum (Phillips) L.Sin - E.PlbKalentera sp. L.Sin - E.PlbMyoconcha neuquena Leanza E.Plb - E.ToaMyoconcha steinmanni (Jaworski) Aal"Astarte" chubutensis (Wahnish) E.Plb - E.Toa"Astarte" cf, gregaria Philippi Aal"Astarte" keideli Wahnish E.Plb - L.Plb"Asiarle" cf. subtetragona Munster AalNeocrassina andium (Gottsche) AalNeocrassina aureliae (Feruglio) E.Plb - AalCoelastarte excavata (J. Sowerby) AaiOpisoma excavatum Boehm M.Toa - M.ToaTrigonastarte? steinmanni (Morike) M.Toa - AalAstartidae gen. et sp. nov. L.Plb - L.PlbCardinia andium (Giebel) E.Sin - E.ToaCardinia cf. listen (J. Sowerby) L.Sin - L.SinProtocardia striatula L.Plb - Aal
(J. de C. Sowerby)Protocardia sp. L.Sin - L.SinJurassicardium? asaphum (Leanza) L.Sin - E.PlbQuenstedtial sp. L.Sin - L.PlbCorbicellopsis? sp. L.Sin - L.SinTancredia sp. AalAnisocardia sp. L.Sin - L.PlbIsocyprina ancatruzi (Leanza) L.Sin - L.PlbRolliereHa sp. L.Toa - L.ToaArcticidae gen. et sp. nov. L.Plb - L.PlbPseudisocardia cf. cordata Aal
(Buckmann)Pseudisocardial liasina Wahnish E.Plb - E.Toa" Venus" peregrina Gottsche AalPholadomya corrugata L.Sin - M.Toa
Koch&DunkerPholadomya aff. corrugata Het
Koch & DunkerPholadomya cf. decorata Hartmann L.Sin - E.PlbPholadomya fidicula (J. Sowerby) L.Sin - AalPholadomya multilineata Gabb E.Plb - E.ToaPholadomya sp. nov. L.Sin - L.SinGoniomya cf. proboscidea Agassiz E.Toa - AalGoniomya sp. nov. A E.Plb - L.PlbGoniomva sp. nov. B HetHomomya neuquena Leanza L.Sin - E.ToaOsteomya dilata (Phillips) L.Sin - L.SinPachymya rotundocaudata (Leanza) E.Plb - L.PlbPachymyal sp. L.Sin - L.SinPachymva senckenbergi Moricke AalCeratomya sp. E.Plb - L.PlbGresslya peregrina (Phillips) L.Plb - AalGresslya sp. E.Sin - E.PlbPleuromya galathea Agassiz HetPleuromva uniformis (J. Sowerby) L.Sin - AalCercomya undulata E.Toa - Aal
(J. de C. Sowerby)Cercomya peruviana Cox L.Toa - AalPlatvmvoideal cf. broliensis L.Plb - L.Plb
(Buvignier)
Data based on evaluation of information inHillebrandt (1980). Hillebrandt & Schmidt-Effing (1981). Perez (1982). Hillebrandt &Westermann (1985). Damborenea ( I 9 8 7 a . b .1990a,b 1992. 1993. 1996): Aberhan (1992.1994). Damborenea & Mancenido (1992).Leanza (1993). Perez & Reyes (1994) andunpublished information. Range: Het.Hettangian: Sin, Sinemurian: Plb, Pliensbachian:Toa, Toarcian: Aal. Aalenian: E. Early: M.Middle: L, Late. Note that "Het" does notnecessarily imply first appearance in theHettangian. and 'Aal' does not necessarily implylast appearance in the Aalenian.
Appendix 2Stratigraphic distribution of Hettangian to Aalenianbivalves from NW Europe,
Taxon Range
Palaeonucula hammeri (Defrance) M.Toa - AalPalaeonucula navis (Piette) Het - E.ToaPalaeoneilo elliptica (Goldfuss) Het - L.PlbPalaeoneilo galatea (d'Orbigny) E.Plb - AalNuculana ovum (J. de C. Sowerby) M.Toa - M.ToaDacryomva gaveyi Cox E.Plb - L.PlbDacryomya heberti (Martin) Het - E.PlbDacrvomva lacryma L.Toa - Aai
(J. de C. Sowerby)Rollieria bronni (Andler) Het - L.PlbRyderia doris (d'Orbigny) Het - AalRyderia texturata (Terquem Het - L.Plb
"& Piette)Isoarca subspirata (Minister) AalArea minuta J. de C. Sowerby AalParallelodon buckmani (Richardson) Het - E.ToaParallelodon hirsonensis (d'Archiac) L.Toa - AalParallelodon trapezium Cox E.Plb - E.ToaGrammatodon chapuisi Arkell M.Toa - M.ToaGrammatodon inaequivalvis L.Toa - Aal
(Munster)Grammatodon muensteri (Zieten) E.Sin - AalCucullaea oblonga J. Sowerby L.Toa - AalParainoceramus obliquus Aal
(Morris & Lycett)Parainoceramus ventricosus E.Plb - E.Plb
(J. de C. Sowerby)Pseudomytiloides dubius E.Toa - M.Toa
(J. de C. Sowerby)Mytiloceramus polyplocus (Roemer) M.Toa - AalSteinmannia bronni (Voltz) E.Toa - M.ToaSemuridia dorsetensis (Cox) E.Sin - L.SinPteria carixiensis Cox E.Plb - E.PlbPteroperna deshayesi (Terquem) Het - E.PlbPteropernaplana (Morris & Lycett) AalBakevellia binneyi (Brown) HetBakevellia waltoni (Lycett) Het - AalGervillaria alaeformis (J. Sowerby) AalGervillaria hartmanni (Munster) Het - AalGervillaria tortuosa L.Toa - Aal
(J. de C. Sowerby)Gervillella aviculoides (J. Sowerby) Het - AalGervillella siliqua Aal

EARLY JURASSIC BIVALVE BIODIVERSITY 137
(Eudes-Deslongchamps)Gervillia lanceolata Het - E.Toa
(J. de C. Sowerby)Gervillia metiertensis (Joly 1907) HetGervillia northamptonensis Cox AalIsognomon lugdunensis (Dumortier) L.Sin - L.PlbIsognomon mytiloides (Lamarck) L.Toa - AalLinsdallia quadrata Aal
(J. de C. Sowerby)Bositra buchi (Roemer) E.Toa - AalPinna cuneata Phillips AalPinna folium Young & Bird Het - E.ToaAntiquilima cubifera (Whidborne) E.Toa - AalAntiquilima succincta (Schlotheim) Het - L.PlbCtenostreon rugosum (Smith) M.Toa - AalCtenostreon tuberculatitm Het
(Terquem)Plagiostoma alticosta L.Toa - Aal
(Chapuis & Dewalque)Plagiostoma cardiiformis J. Sowerby AalPlagiostoma giganteum J. Sowerby Het - L.PlbPlagiostoma hersilia (d'Orbigny) AalPlagiostoma punctatum J. Sowerby L.Sin - E.ToaPlagiostoma rodburgensis L.Toa - Aal
(Whidborne)Plagiostoma toarcensis M.Toa - Aal
(Eudes-Deslongchamps)Pseudolimea acuticostata (Minister) E.Plb - E.ToaPseudolimea cristata (Dumortier) E.Plb - L.PlbPseudolimea duplicata L.Toa - Aal
(J. de C. Sowerby)Pseudolimea grettonensis Cox E.Toa - E.ToaPseudolimea harburyiensis Cox Het - E.SinPseudolimea hettangiensis Het - L.Plb
(Terquem)Pseudolimea koninckana Het - E.Sin
(Chapuis & Dewalque)Pseudolimea pectinoides Het - E.Toa
(J. Sowerby)Regalilima oolitica (Lycett) AalPlicatula laevigata (d'Orbigny) L.Plb - L.PlbPlicatula spinosa (J. Sowerby) E.Plb - E.ToaAtreta intusstriata (Emmerich) Het - E.SinActinostreon marshii (J. Sowerby) AalActinostreon solitarium Aal
(J. de C. Sowerby)Gryphaea arcuata Lamarck Het - E.SinGryphaea gigantea J. Sowerby E.Plb - L.PlbGryphaea mccullochi E.Sin - L.Sin
J. de C. SowerbyGryphaea pictaviensis (Hebert) L.Toa - L.ToaCatinula crickleyensis Cox AalLiostrea hisingeri (Nilsson) Het - M.ToaNanogyra monoptera E.Toa - E.Toa
(Eudes-Deslongchamps)Anningella faberi (Oppel) E.Sin - L.SinOxytoma calva (Schlonbach) E.Sin - L.PlbOxytoma inequivalvis (J. Sowerby) Het - AalOxytoma cygnipes (Young & Bird) E.Plb - L.PlbMeleagrinella fallax (Pflucker) HetMeleagrinella papyria (Quenstedt) L.Sin - L.SinMeleagrinella substriata (Miinster) E.Toa - M.ToaTerquemia arietis (Quenstedt) Het - L.PlbPlacunopsis radiata (Phillips) Het - Aal
Propeamussium laeviradiatum L.Plb - Aal(Waagen)
Propeamussium pumilum L.Plb - Aal(Lamarck)
Entolium corneolum (Young & Bird) M.Toa - AalEntolium lunare (Roemer) Het - M.ToaCamptonectes auritus (Schlotheim) Het - AalCamptonectes laminatus (J. Sowerby) AalCamptonectes subulatus (Miinster) Het - L.ToaChlamys pollux (d'Orbigny) Het - E.SinChlamys textoria (Schlotheim) Het - AalChlamys valoniensis (Defrance) Het - E.SinEopecten abjectus (Phillips) L.Plb - AalEopecten spondyloides (Roemer) M.Toa - AalEopecten velatus (Goldfuss) Het - AalPseudopecten barbatus M.Toa - Aal
(J. Sowerby)Pseudopecten dentatus Het - Aal
(J. de C. Sowerby)Pseudopecten equivalvis Het - E.Toa
(J. Sowerby)Pseudopecten veyrasensis Het - L.Plb
(Dumortier)Spondylopecten subspinosus Het - Aal
(Schlotheim)Arcomytilus pectinatus (J. Sowerby) AalMusculus subcancellata (Buvignier) L.Sin - L.PlbInoperna lilliputensis Hodges HetInoperna sowerbyana (d'Orbigny) L.Toa - AalModiolus hillanus (J. Sowerby) Het - L.SinModiolus imbricatus (J. Sowerby) L.Toa - AalModiolus minimus (J. Sowerby) HetModiolus scalprum (J. Sowerby) L.Sin - E.ToaModiolus ventricosus (Roemer) Het - L.PlbLiotrigonia lingonensis (Dumortier) L.Plb - L.PlbTrigonia costata J. Sowerby L.Toa - AalMyophorella striata (J. Sowerby) L.Toa - AalScaphotrigonia navis (Lamarck) M.Toa - AalVaugonia flecta (Morris & Lycett) AalVaugonia literata (Young & Bird) M.Toa - M.Toa'Lucina' plana Zieten L.Plb - L.PlbLuciniola limbata Het - L.Sin
(Terquem & Piette)Mesomiltha lirata (Phillips) L.Toa - AalMactromya cardioideum (Phillips) Het - E.ToaSphaeriola subglobosa (Tate) E.Plb - L.ToaMyoconcha decorata (Miinster) Het - L.PlbMyoconcha psilonoti Quenstedt Het - E.PlbTutcheria cingulata (Goldfuss) Het - E.PlbTutcheria submulticostata E.Plb - L.Plb
(d'Orbigny)Pseudopis deslonchampsis (Tate) E.Plb - L.PlbAstarte guexi d'Orbigny Het - L.PlbAstarte obsoleta Dunker Het - L.PlbNeocrassina elegans (J. Sowerby) M.Toa - AalCoelastarte excavata (J. Sowerby) L.Toa - AalNicaniella voltzii (Honighaus) L.Toa - AalPraeconia rhomboidalis (Phillips) AalCoelopis lunulata (J. Sowerby) L.Toa - AalCardinia attenuata (Stutchbury) L.Sin - E.ToaCardinia concinna (J. Sowerby) Het - E.ToaCardinia listen (J. Sowerby) Het - L.PlbCardinia ovalis (Stutchbury) Het - E.SinNidarica slatted (Wilson & Crick) L.Plb - E.Toa

138 MARTIN ABERHAN
Hippopodium ponderosum E.Sin - L.PlbJ. Sowerby
Protocardia buckmani L.Toa - AalMorris & Lycett
Protocardia phillipiana (Dunker) Het - L.SinProtocardia truncata E.Plb - E.Toa
(J. de C. Sowerby)Quenstedtia laevigata (Phillips) L.Toa - AalSowerbya triangularis (Phillips) L.Toa - AalTancredia donaciformis Lycett L.Toa - AalTancredia securiformis Dunker Het - E.PlbAnisocardia minima (J. Sowerby) AalAnisocardia truncata (Morris) AalIsocyprina menkei (Dunker) HetEotrapezium cucullatum (Munster) E.Plb - E.ToaPronoella beneckei (Rollier) AalPronoella trigonellaris (Schlotheim) L.Toa - AalPseudotrapezinm cordiforme L.Toa - Aal
(Deshayes)Pseudisocardia cordata (Buckman) AalPholadomya ambigua (J. Sowerby) Het - L.PlbPholadomya corrugata Het - E.Plb
Koch & DunkerPholadomya fidicula Aal
J. de C. SowerbyPholadomya lirata (J. Sowerby) AalPholadomya ovalis (J. Sowerby) AalGoniomya hybrida (Miinster) E.Plb - E.ToaGoniomya literata (J. Sowerby) AalHomomya gibbosa (J. Sowerby) L.Plb - AalOsteomya dilata (Phillips) AalPachymya vetusa (Phillips) L.Sin - L.PlbCeratomya concentrica Aal
(J. de C. Sowerby)Ceratomya petricosa (Simpson) L.Sin - L.PlbCeratomya sublaevis (Tate) L.Plb - L.PlbGresslya intermedia (Simpson) L.Plb - AalGresslya peregrina (Phillips) AalPteromya tatei Het
(Richardson & Tutcher)Pleuromya alduini (Brongniart) AalPleuromya calceiformis (Phillips) AalPleuromya costata (Young & Bird) E.Plb - E.ToaPleuromya galathea Agassiz Het - E.SinPleuromya uniformis (J. Sowerby) Het - AalPlatymyoidea broliensis (Buvignier) L.Plb - L.PlbThracia depressa J. de C. Sowerby Aal
Data based on evaluation of information in Hallam(1976, 1987), Johnson (1984), Hodges (1991,2000). Muster (1995), Rohl (1998). Harries &Little (1999), and unpublished information. Forabbreviations see Appendix 1. Note that 'Het'does not necessarily imply first appearance inthe Hettangian. and 'Aal' does not necessarilyimply last appearance in the Aalenian.
ReferencesABERHAN, M. 1992. Palokologie und zeitliche Verbrei-
tung benthischer Faunengemeinschaften imUnterjura von Chile. Beringeria. 5, 1-174.
ABERHAN, M. 1993. Benthic macroinvertebrate associ-ations on a carbonate-clastic ramp in segments ofthe Early Jurassic back-arc basin of northern
Chile (26-29°S). Revista Geologica de Chile. 20,105-136.
ABFRHAN. M. 1994. Early Jurassic Bivalvia of north-ern Chile. Part I. Subclasses Palaeotaxodonta.Pteriomorphia, and Isofilibranchia. Beringeria.13,3-115.
A B E R H A N . M. 2001. Bivalve palaeobiogeography andthe Hispanic Corridor: time of opening and effec-tiveness of a proto-Atlantic seaway. Palaeo-geography, Palaeoclimatologv, Palaeoecology,165. 375-394.
ABERHAN, M. & FURSICH. F. T. 1997. Diversity analy-sis of Lower Jurassic bivalves of the AndeanBasin and the Pliensbachian-Toarcian massextinction. Lethaia, 29, 181-195.
A B E R H A N . M. & FURSICH. F. T. 2000. Mass originationversus mass extinction: the biological contri-bution to the Pliensbachian-Toarcian extinctionevent. Journal of the Geological Society London.157, 55-60.
ABERHAN. M. & PALFY. J. 1996. A low oxygen tolerantEast Pacific flat clam (Posidonotis semiplicata]from the Lower Jurassic of the CanadianCordillera. Canadian Journal of Earth Sciences.33.993-1006.
DAMBORENEA, S. E. 1987 a. Early Jurassic Bivalvia ofArgentina. Part 1: Stratigraphical introductionand superfamilies Nuculanacea, Arcacea. Myti-lacea and Pinnacea. Palaeontographica (A). 199.23-111.
DAMBORENEA. S. E. 1987b. Early Jurassic Bivalvia ofArgentina. Part 2: Superfamilies Pteriacea.Buchiacea and part of Pectinacea. Palaeonto-graphica (A). 199. 113-216.
DAMBORENEA. S. E. 1990a. Bivalves. In: RICCARDI. A.C.. DAMBORENEA, S. E. & MANCENIDO. M. O.(eds) Lower Jurassic of South America andAntarctic Peninsula. Newsletters on Stratigraphy.21. 85-90.
DAMBORENEA, S. E. 1990ft. Bivalves. In: RICCARDI. A.C. WESTERMANN. G. E. G. & DAMBORENEA. S. E.(eds) Middle Jurassic of South America andAntarctic Peninsula. Newsletters on Stratigraphv.21,114,116-119.
D A M B O R E N E A . S. E. 1992. South American Jurassicbivalves. In: WESTERMANN. G. E. G. (ed.) TheJurassic of the Circum-Pacific, Atlas. CambridgeUniversity Press. Cambridge, pls. 115-119.
DAMBORENEA. S. E. 1993. Early Jurassic South Ameri-can pectinaceans and circum-Pacific palaeobio-geography. Palaeogeography, Palaeoclimatologv,Palaeoecology. 100, 109-123.
DAMBORENEA. S. E. 1996. Palaeobiogeography ofEarly Jurassic bivalves along the southeasternPacific margin. XIII Congreso GeologicoArgentino y /// Congreso de Exploracion deHidrocarburos, Actas. 5, 151-167.
DAMBORENEA, S. E. 2000. Hispanic Corridor: Its evol-ution and the biogeography of bivalve molluscs.In: HALL. R. L. & SMITH. P. L. (eds) Advances inJurassic Research 2000. GeoResearch Forum. 6.369-380.
DAMBORENEA. S. E. & M A N C E N I D O , M. O. 1979. Onthe palaeogeographical distribution of the pec-tinid genus Weyla (Bivalvia. Lower Jurassic).

EARLY JURASSIC BIVALVE BIODIVERSITY 139
Palaeogeography, Palaeoclimatology, Palaeo-ecology, 27, 85-i02.
DAMBORENEA, S. E. & MANCENIDO, M. O. 1992. Acomparison of Jurassic marine benthonic faunasfrom South America and New Zealand. Journalof the Royal Society of New Zealand, 22,131-152.
GRADSTEIN, F. M., AGTERBERG, F. P., OGG, J. G., HARD-ENBOL, X, VAN VEEN, P., THIERRY, J. & HlJANG Z.1994. A Mesozoic time scale. Journal of Geophys-ical Research, 99, 24051-24074.
HALLAM, A. 1976. Stratigraphic distribution andecology of European Jurassic bivalves. Lethaia, 9,245-259.
HALLAM, A. 1983. Early and mid-Jurassic molluscanbiogeography and the establishment of thecentral Atlantic seaway. Palaeogeography,Palaeoclimatology, Palaeoecology, 43,181-193.
HALLAM, A. 1986. The Pliensbachian and Tithonianextinction events. Nature, 319, 765-768.
HALLAM, A. 1987. Radiations and extinctions inrelation to environmental change in the marineLower Jurassic of northwest Europe. Paleobiol-ogy. 13,152-168.
HALLAM, A. 1990. Biotic and abiotic factors in theevolution of Early Mesozoic marine molluscs. In:Ross, R. M. & ALLMON, W. D. (eds) Causes ofEvolution. A Paleontological Perspective. TheUniversity of Chicago Press, Chicago, 249-260.
HALLAM, A. 1996. Recovery of the marine fauna inEurope after the end-Triassic and early Toarcianmass extinctions. In: HART, M. B. (ed.) BioticRecovery from Mass Extinction Events. Geo-logical Society, London, Special Publications, 102,231-236.
HALLAM, A. & WIGNALL, P. B. 1997. Mass Extinctionsand their Aftermath. Oxford University Press,Oxford.
HARRIES, P. J. & LITTLE, C. T. S. 1999. The earlyToarcian (Early Jurassic) and the Cenomanian-Turonian (Late Cretaceous) mass extinctions:similarities and contrasts. Palaeogeography,Palaeoclimatology, Palaeoecology, 154, 39-66.
HESSELBO, S. P., GROCKE, D. R., JENKYNS, H. C,BJERRUM, C. J., FARRIMOND, P., MORGANS BELL,H. S. & GREEN, O. R. 2000. Massive dissociationof gas hydrate during Jurassic oceanic anoxicevent. Nature, 406, 392-395.
HILLEBRANDT, A. VON. 1980. Paleozoografia deJurasico marino (Lias hasta Oxfordiana) enSuramerica. In: ZEIL, W. (ed.) Nuevos Resultadosde la Investigacion Geodentifica Alemana enLationoamerica. Deutsche Forschungsgemein-schaft, Bonn, and Instituto de ColaboracionCientifica, Tubingen, 132-134.
HILLEBRANDT, A. VON & SCHMIDT-EFFING, R. 1981.Ammoniten aus dem Toarcium (Jura) von Chile(Stidamerika). Zitteliana, 6,1-74.
HILLEBRANDT, A. VON. & WESTERMANN, G. E. G. 1985.Aalenian (Jurassic) ammonite faunas and zonesof the Southern Andes. Zitteliana, 12, 3-55.
HODGES, P. 1991. The relationship of the Mesozoicbivalve Atreta to the Dimyidae. Palaeontology, 34.963-970.
HODGES, P. 2000. The Early Jurassic Bivalvia from theHettangian and Lower Sinemurian of south-westBritain. Monograph of the PalaeontographicalSociety, London, 154,1-64.
JABLONSKI, D. 1998. Geographic variation in the mol-luscan recovery from the end-Cretaceous extinc-tion. Science, 279,1327-1330.
JOHNSON, A. L. A. 1984. The palaeobiology of thebivalve families Pectinidae and Propeamussiidaein the Jurassic of Europe. Zitteliana, 11,1-235.
LEANZA, H. A. 1993. Jurassic and Cretaceous trigoniidbivalves from west-central Argentina. Bulletins ofAmerican Paleontology, 105, 5-95.
LITTLE, C. T S. & BENTON, M. J. 1995. Early Jurassicmass extinction: a global long-term event.Geology, 23, 495-498.
MUSTER, H. 1995. Taxonomie und Palaobiogeogra-phie der Bakevelliidae (Bivalvia). Beringeria, 14,3-161.
PALFY, J. & SMITH, P. L. 2000. Synchrony betweenEarly Jurassic extinction, oceanic anoxic event,and the Karoo-Ferrar flood basalt volcanism.Geology, 28, 747-750.
PEREZ, E. 1982. Bioestratigrafia del Jurasico deQuebrada Asientos, norte de Potrerillos, Regionde Atacama. Servicio Nacional de Geologia vMineria - Chile, Boletin, 37,1-149.
PEREZ, E. & REYES, R. B. 1994. Catalogo de ejem-plares tipo, conservados en la coleccion paleon-tologica del Sevicio Nacional de Geologia yMineria, Chile, desde 1953 a 1993. ServicioNacional de Geologia y Mineria - Chile, Boletin,46,1-99.
RAUP, D. M. & SEPKOSKI, J. J., Jr. 1984. Periodicity ofextinctions in the geologic past. Proceedings of theNational Academy of Sciences USA, 81, 801-805.
ROHL, H.-J. 1998. Hochauflosende palokologische undsedimentologische Untersuchungen im Posi-donienschiefer (Lias e) von SW-Deutschland.Tiibinger Geowissenschaftliche Arbeiten, Reihe A,47,1-170.
SMITH, P. L. 1983. The Pliensbachian ammonite Day-iceras dayiceroides and Early Jurassic paleogeog-raphy. Canadian Journal of Earth Sciences, 20,86-91.
SMITH, P. L. & TIPPER, H. W. 1986. Plate tectonicsand paleobiogeography: Early Jurassic (Pliens-bachian) endemism and diversity. Palaios, 1,399-412.
VERMEIJ, G. J. 1977. The Mesozoic marine revolution;Evidence from snails, predators and grazers. Pale-obiology, 3, 245-258.

This page intentionally left blank

Cretaceous patterns of floristic change in the Antarctic Peninsula
DAVID J. CANTRILL1 & IMOGEN POOLE2
lBritish Antarctic Survey, Natural Environment Research Council, High Cross, MadingleyRoad, Cambridge CB3 OET, UK (e-mail: [email protected])
2Wood Anatomy Section, National Herbarium of the Netherlands, University of Utrechtbranch, PO Box 80102, 3585 CS Utrecht, The Netherlands
Abstract: Cretaceous radiation of angiosperms from low to high palaeolatitudes, coupledwith the break-up of Gondwana, played a major role in establishing and maintaining bio-geographic patterns across the southern hemisphere. Uncertainties in details of platereconstructions provide conflicting hypotheses about area relationships of Gondwana frag-ments. This has led to a number of competing proposals concerning angiosperm migrationacross Gondwana. Central to this debate is the role of the Antarctic Peninsula, a regionthat is often envisaged as providing the main connection between east and west Gondwana.The initial radiation of angiosperms into the Antarctic Peninsula region, however, post-dates appearances elsewhere in east Gondwana (e.g. Australia), strongly suggesting thatthe Antarctic Peninsula was not the main gateway, at least in the early stages of Gondwanaradiation. A steep climatic gradient in this part of the world probably acted as an effectivebarrier to angiosperm radiation. The peak of floristic replacement coincides with the peakof Cretaceous warmth (Turonian) which in turn suggests that climatic warming acted as aforcing mechanism by pushing latitudinal belts of vegetation southwards. Once into thesouthern high latitudes angiosperms diversified, and as climates cooled during the LateCretaceous a number of important groups seem to have their origins here. Recent investi-gations of Antarctic macro- and microfloras indicate progressive floristic replacementthrough the Cretaceous. Bryophytes, hepatophytes, bennettites and other seed plants allshow a rapid decline in diversity. In contrast, ferns initially decline then recover, whileconifers remain relatively stable. The ecological preferences of the replaced groups implythat angiosperms initially occupied areas of disturbance and were understorey colonizers,only later replacing fern thickets and becoming important in the overstorey. This patternis consistent with those observed elsewhere through the Cretaceous.
Two major events influenced Cretaceous to Antarctica) (Fig. 1). Consequently, the break-uppresent-day biotic history of the southern hemi- of Gondwana underpins many biogeographicsphere: the radiation of angiosperms and the and diversity patterns seen in the southernconcomitant break-up of Gondwana. During hemisphere today. Antarctica is unique amongstmid-Cretaceous times (130-90 Ma BP) terrestrial fragments of Gondwana in that it was attachedvegetation underwent rapid modernization, at some time to all other major components ofwith an explosive radiation of crown group the supercontinent (Fig. 1). For this reasonangiosperms (Lidgard & Crane 1990). In the understanding the geological evolution ofnorthern hemisphere it is well established that Antarctica is essential for evaluating terrestrialangiosperms arose at low palaeolatitudes and connections across Gondwana through time,spread polewards (Crane & Lidgard 1989; Crane Although the timing and pattern of break-up iset al 1995). A paucity of knowledge about relatively well understood for the large conti-southern hemisphere floras, and in particular nental fragments, this is less the case for smallerthose of Antarctica, means that the pattern in microplates (Storey et al 1988). The present-daythe southern hemisphere is less well constrained Antarctic region can be divided into two areas;(Drinnan & Crane 1990). However, a general East Antarctica, a craton; and West Antarctica,pattern of earlier inception in low latitudes is composed of a number of microcontinentalevident from the limited data available (Drinnan fragments (e.g. Antarctic Peninsula, Thurston& Crane 1990). Island-Eights Land, Marie Byrd Land, Haag
During Cretaceous times disintegration of Nunalaks, Ellsworth-Whitmore Mountains)Gondwana was important in maintaining, or (Fig. 2). These microcontinents have a long andcreating, biotic gateways between what are now complicated history (Storey et al 1988) andwidely separated landmasses (Australia, Africa, represent one of the major uncertaintiesIndia, New Zealand, South America and in Gondwana reconstructions. Nevertheless,
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, Londor, Special Publications, 194,141-152.0305-8719/02/$15.00 © The Geological Society of London 2002.
142 DAVID J. CANTRILL & IMOGEN POOLE
Fig, 1. View of Gondwana break-up for selected times in the Cretaceous, (A) Valanginian (135 Ma BP),illustrating the close proximity of Africa and India to the rest of Gondwana, (B) Albian (105 Ma BP). Notethat India and South Africa have separated from the rest of Gondwana. (C) Santonian (84 Ma BP) showingextensive separation between India, Africa and the rest of Gondwana. Australia is beginning to rift fromAntarctica along the southern margin. (D) Maastrichtian (68 Ma BP). New Zealand and Lord Howe rise arerifting from the eastern margin of Gondwana. Note the continued maintenance of connections betweenSouth America and east Gondwana (Australia, Antarctica, New Zealand), Latitude intervals are 30": dashedblack line represents palaeopolar circle. Reconstructions provided by R. A. Livermore. British AntarcticSurvey.
developing a framework that reconstructs theposition of these fragments through time iscritical for understanding terrestrial connectionsbetween various landmasses and subsequentvicariant events.
The same is also true for other regions ofGondwana; for example, large uncertaintiesexist in our knowledge of the separation of India
from Antarctica, due to a lack of seafloor dataand age constraints on magnetic anomaliesalong the Antarctic margin. The development ofmarine conditions in present-day eastern Indiansedimentary basins suggests that the separationof India was diachronous from west to east(Truswell et ai 1999). However, reconstructionsbased on seafloor magnetic anomalies indicate

ANTARCTIC CRETACEOUS BIODIVERSITY CHANGE 143
Fig. 2. AIbian reconstruction of Gondwana courtesyof R. A. Livermore, British Antarctic Survey. Polarview illustrating the importance of the West Antarcticmicrocontinents (A, Antarctic Peninsula; B, ThurstonIsland; C, Marie Byrd Land; D, Ellsworth-WhitmoreMountains; E, Haag Nunataks) in maintainingconnections between east and west Gondwana. Notealso the potential importance of the KergeulenPlateau (F) and the Mozambique Rise (G) in theearly stages of rifting.
Fig. 3. Alternative scenarios for the expansion ofangiosperms across Gondwana plotted on an Aptianreconstruction provided by R. A. Livermore, BritishAntarctic Survey.
opening from east to west (Veevers et al 1991;Royer & Coffin 1992). Although seafloorspreading was well established by Aptian times(c. 120 Ma BP), microcontinental fragments suchas the Kerguelen Plateau (continental crust)(Fig. 2) probably maintained terrestrial connec-tions for a considerable period of time. Recentwork has identified terrestrial deposits ofTuronian age (c. 90 Ma) on the KerguelenPlateau (Shipboard Scientific Party 2000),implying that the microcontinent was above sealevel at this time.
The reconstruction of Africa/Antarctica andthe subsequent early stages of break-up are alsoproblematic due to a paucity of magneticanomaly data (Marks & Tikku 2001). Recon-structions such as those of Lawver et al (1997),Muller et al (1998) and Roeser et al (1996)differ fundamentally from those of Segoufin &Patriat (1980), Livermore & Hunter (1996) andMarks & Tikku (2001), and consequently havemarkedly different implications for the timing ofterrestrial separation. Sea floor spreading hadstarted by 155 Ma BP in, for example, theMozambique Basin (Segoufin & Patriat 1980),and off the Dronning Maud Land Coast (Roeser
et al 1996). However, much of the initialAfrican/Antarctic motion had a large transformcomponent that probably allowed landmasses tobe juxtaposed (e.g. Mozambique Ridge), andrecent reconstructions suggest terrestrial con-nections were present at least until 120 Ma BPand perhaps later (Reeves & de Wit 2000). Therecent discovery of Late Cretaceous dinosaurs inMadagascar (Sampson et al 1998), and theiroccurrence in South America and India, alsosuggests that these landmasses were still con-nected (via Antarctica) in the Late Cretaceous(Sampson et al 1998; Krause et al 1999; Hay etal 1999). The uncertainties in the timing ofterrestrial separation between the componentcontinents of Gondwana remain a majorconstraint in explaining present-day disjunctdistribution patterns.
Despite the present uncertainty, conventionalreconstructions of Gondwana break-up implyseparation of Africa/South America from eastGondwana by at least 155 Ma BP (early LateJurassic times), and therefore substantiallybefore angiosperm radiation and diversification.The only connection between east (Africa,South America) and west (India, Australia,

144 DAVID J. CANTRILL & IMOGEN POOLE
Antarctica, New Zealand) Gondwana is via theAntarctic Peninsula, a region that has beenpostulated as providing the main connecting linkduring the radiation of angiosperms (Fig. 3).Alternative scenarios have also been postulated;palaeogeographic reconstructions of SE Asia,Australia and east Antarctica during theCretaceous indicate land connections betweenthese areas (Metcalfe 1990), and point to apossible invasion from Asia to the north (Burger1981; Truswell et al. 1987; Hill & Scriven 1995,fig. 5) (Fig. 3). Other workers have postulatedthat angiosperms arrived in Gondwana via India(Burger 1990), while more recently Dettmann(1989, 1994) suggested that angiospermsmigrated along rift valleys, as these providedhighly disturbed environments that favouredearly angiosperms (Fig. 3).
To date, relatively little is known ofCretaceous angiosperm floras from Antarctica(Truswell 1991), yet they are critical for testingthe validity of hypothesized radiation routes. Ifthe Antarctic Peninsula acted as the maingateway, then Cretaceous floras from this regionshould have earlier records of angiosperms thanelsewhere in eastern Gondwana (i.e. Australia,New Zealand, India). Alternatively, if angio-sperms radiated into Gondwana via anotherroute then they should appear later in Antarcticfloras than elsewhere in Gondwana.
Floristic change in the Antarctic PeninsulaOver the last ten years our knowledge of thefloristic composition of the Antarctic Peninsulamacro- and microfloras has increased dramati-cally. Well documented macrofloras are nowknown from Aptian (Cantrill 1997, 2000; Cesariet al. 1998, 1999; Torres et al. 1997), Albian(Cantrill 1995, 1996; Cantrill & Nichols 1996;Falcon-Lang & Cantrill 2000; Cantrill & Falcon-Lang 2001), Coniacian (Hayes 1999), Santonian(Hayes 1999), Campanian and Maastrichtian(Poole & Francis 1999, 2000; Poole et al.2000a,b,c; Poole & Gottwald 2001) strata.However, the Maastrichtian record of leaf florasis sparse and could not be used in this study. Incontrast, microfloras have been recorded fromall of the major stages of the Cretaceous (exclud-ing the Hauterivian and Barremian) (Askin1990, 1994; Duane 1996; Riding et al. 1998;Cantrill 2000), and so provide a record for thelatest Cretaceous (Maastrichtian).
Patterns of floristic replacement through theCretaceous were examined using within-floradiversity plots (Lidgard & Crane 1990). Thesewere calculated as a percentage of the totalspecies diversity for each macroflora and
microflora. Within-flora diversity moderatessample size effects (Spicer 1988) but the diver-sity measure for groups is not independent, asthe values of a single flora must sum 100%.Comparisons of absolute within-flora speciesrichness with within-flora diversity measuresdemonstrate that within-flora diversity capturespatterns seen in absolute diversity and does notinduce spurious trends (Lidgard & Crane 1990).Despite taxonomic biases in both the macro-floral and microfloral record that make itdifficult to discriminate certain groups (e.g. ben-nettites in the microflora, lycopods in themacroflora), the datasets show similar patternsthrough time. This gives us some confidence thatthe observed patterns are a real reflection ofchanges in vegetation composition.
Bennettitales and other non-coniferousseed plants
Bennettites were initially reasonably diverse(Aptian, 14%), but then rapidly disappear fromthe floras becoming extinct by the Campanian(Fig. 4A). Unfortunately bennettites do nothave a good pollen record; their monosulcategrains lack distinguishing features at the light-microscope level, which makes discrimination ofspecies difficult. Thus the pollen record appearsto be less diverse than the macrofloral record(see Dettmann 1986b). Like the bennettites.other non-coniferous seed plants (e.g.Pachypteris, Ginkgoales) all show a progressivedecline from the Aptian to Albian, then recoverslightly in Cenomanian times before declining inimportance through the later part of theCretaceous (Fig. 4B).
The decline in bennettites is also reflected intheir abundance through the Cretaceous. Theyare one of the most frequently encounteredfoliage taxa in the Aptian floras, but by Albiantimes, although still accounting for 3.5% of thefloristic diversity, they are very rare. Only ahandful of specimens are known from theAlbian and one from the Coniacian. Thissuggests that not only did they become lessdiverse but that they also became less importantecologically within the vegetation. A similarpattern is seen elsewhere globally. For example,macrofloras in Australia have abundant Bennet-titales in early Aptian floras (e.g. Douglas 1969)but they disappear by Aptian-Albian times(Drinnan & Chambers 1986). Recent work ondispersed cuticles indicates that bennettitesremained a minor constituent of the Australianvegetation at least until the Cenomanian (Pole& Douglas 1999).
ANTARCTIC CRETACEOUS BIODIVERSITY CHANGE 145
Fig. 4. Diversity trends through the Cretaceous for macrofloral (solid line) and mierofloral (dashed line) data.Note the close correspondence between angiosperm diversity and palaeotemperature estimates (F) for theSouthern Ocean. Temperature curve modified and smoothed from Huber (1998). Macrofloral taxic diversitycompiled from Orlando (1968), Hernandez & Azcarate (1971), Lacey & Lucas (1981), Banerji & Lemoigne(1987), Rees & Smellie (1989), Barale et al (1995), Cesari et al. (1999), Hayes (1999), Cantrill (2000) andFalcon-Lang et al. (2001). Mierofloral taxic diversity compiled from Dettmann & Thomson (1987), Askin(1990,1994), Dolding (1992), Keating (1992), Keating et al. (1992), Riding et al (1998), Barreda et al. (1999).Hathway et al. (1999) and Cantrill (2000).

146 DAVID J. CANTRILL & IMOGEN POOLE
Fig. 5. Floristic provincialism across Gondwana derived from Herngreen el al. (1996). Note the regionaldifferences in abundance of key trisaccate species such as Microcachyridites antarcticus within the TrisaccateProvince. Histograms show, percentage abundance of M. antarcticus in Lower Cretaceous strata: (A) fromDettmann & Thomson (1987); (B) southeastern Australia from Dettmann (1986b), central Australia fromBurger (1980).
Bryophytes/Hepatophytes
In the earliest Cretaceous, bryophytes andhepatophytes form an important and character-istic component of the floristic diversity. Not onlyare they diverse, accounting for up to 20% of thespecies (Fig. 4C), but they are also ecologicallyabundant ranging from colonizers of freshsediment to components of established fernthickets and forests (Cantrill 1997). Indeed,many localities are characterized by assemblagescomprising just hepatophytes, where they cancover individual bedding surfaces for hundredsof metres. This is also true for other southernhigh-latitude sites (e.g. southeastern Australia;Douglas 1973) and appears to be a phenomenonof high latitude floras in the early part of theMesozoic (Cantrill 1997). The highest hepato-phyte diversity occurs in the Early Cretaceousbut they rapidly become a minor constituent ofthe vegetation by the Late Cretaceous. Thispattern is also reflected in the microfloraldiversity (Fig. 4C).
Conifer ales
Conifers generally maintain between 10 and20% of the within-flora diversity through theCretaceous interval (Fig. 4D). This is apparentfrom the microfloral record but is less clear inthe macroflora. The macrofloral record shows adistinct drop in within-flora diversity in the
mid-Cretaceous. The differences may be due towind-dispersed conifer pollen representing awider range of plant communities than thoseseen in the macrofloras.
Pteridophytes and lycophytes
Macrofloral remains are most diverse in theEarly Cretaceous but decline towards mid-Cretaceous times (Fig. 4E). Following thisdecline ferns recover through the later partof the Cretaceous (Fig. 4E). This pattern issupported by the microfloral record, whichsuggests that the recovery continues through theLate Cretaceous (Fig. 4E). Although a numberof groups show a marked decline in diversitythrough the Cretaceous (e.g. Dipteridaceae),other families remain relatively stable (e.g.Gleicheniaceae). In terms of abundance, fernsshow a similar pattern to the bennettites: theyare extremely common in Aptian and Albianfloras but are less frequently encountered inLate Cretaceous floras. So, although diverse inthe later part of the Cretaceous, they areprobably less important ecologically.
Angiosperms
Macrofloral remains of angiosperms are notrecorded until the Late Albian (Cantrill &Nichols 1996) where they make up to 12 % of theflora. This includes a diversity of habit ranging

ANTARCTIC CRETACEOUS BIODIVERSITY CHANGE 147
Fig. 6. Palaeolatitudinal gradients through South America and the Antarctic Peninsula highlighting theincrease in diversity of mesic elements (hepatics) and the decrease in abundance of thermophilic species (e.g.Classopollis) from low to high latitudes. Diversity data for hepatics derived from Pons (1988), Riccardi (1988)and Falcon-Lang el al. (2001). Abundance data for Classopollis derived from Herngreen (1975), Dettmann &Thomson (1987), Herngreen et al. (1996) and Pramparo & Volkheimer (1999).
from herbaceous plants to probable trees(Cantrill & Nichols 1996). In the macroflorasangiosperms rapidly increase in importancerising to a peak in the Coniacian (72%); this isfollowed by a decline into the Santonian. In themicrofloras angiosperms appear in the earlyAlbian (Dettmann & Thomson 1987) and showa rapid diversification, reaching up to 75%within-flora diversity by Turonian times (Fig.4F). From this peak of diversity they initiallydecline, before rapidly recovering in theCampanian and Maastrichtian.
Summary
Angiosperm within-flora diversity increases atthe expense of bryophytes/hepatophytes, ben-nettites and other gymnosperms and, initially,ferns and lycophytes. This suggests that theangiosperm invasion took place in the nichesoccupied by these groups. Within the AntarcticCretaceous ecosystem liverworts were colonizersof bare sediment and an understorey componentof the vegetation (Cantrill 1997), whilst ferns andlycophytes were either understorey or thicket-forming plants (Cantrill 1996). So it seems mostlikely that the early angiosperms occupiedcolonization and understorey niches. Furthersupport for this idea can be seen in the over-storey elements: conifers, the main tree species,
maintained a relatively constant diversitythroughout the Cretaceous. Indeed, angiospermwood does not become common until Coniaciantimes, and the early angiosperm floras of theAntarctic Peninsula (Albian) are dominated byherbaceous leafy forms, although some leafmorphologies are suggestive of trees (Cantrill& Nichols 1996). The lack of wood alsosuggests that these early angiosperms occupiedunder/middle-storey niches. We conclude thenthat the angiosperms were invading the under-storey and colonization niches within the eco-system as those are the groups that were beingreplaced. Angiosperms later expanded into theoverstorey, perhaps by filling gaps in the canopyleft by conifers. The recovery of fern diversity(Fig. 4E) suggests that pteridophytes were ableto colonize new niches created by theangiosperms.
Antarctic Peninsula: biotic gateway orbiological backwater?In order to evaluate whether the AntarcticPeninsula acted as a biotic gateway during themid-Cretaceous radiation of angiosperms, thefloral succession needs to be densely sampledand well age-constrained. A pilot study(Dettmann & Thomson 1987) identifiedangiosperm pollen in early Albian samples from

148 DAVID J. CANTRILL & IMOGEN POOLE
the James Ross Basin. Unfortunately these werethe oldest samples examined, but ?Barremian toAptian strata in the South Shetlands lackedangiospermous grains (Askin 1983). Since thesestudies over 250 pre-Albian (e.g. Keating et al.1992; Duane 1996; Riding et al. 1998; Hathway etal. 1999), as well as numerous Albian andyounger samples (e.g. Barreda et al. 1999) havebeen examined from various parts of theAntarctic Peninsula. Despite the intensivesampling of Aptian and older strata, angio-sperms do not appear in the Antarctic Peninsulauntil early Albian times. This event clearly post-dates their occurrence elsewhere in easternGondwana (Burger 1990, 1993). For example,angiospermous pollen grains (Clavatipolleniteshughesii) are known from the basal Cyclo-sporites hughesi Palynozone (early Aptian) ofAustralia (Burger 1988), and macrofossils withattached flowers of Aptian age occur in south-eastern Australia (Taylor & Hickey 1990). Sincethese unequivocal records predate the appear-ance of angiosperms in Antarctica it seemsunlikely that the Antarctic Peninsula could haveacted as the main biotic gateway for dispersalacross Gondwana, at least in the initial stages ofthe radiation. Although it seems certain thatterrestrial land connections occurred betweensouthern South America and the AntarcticPeninsula during the Cretaceous, other factorsmust have been acting as a barrier to radiation.
The Early to mid-Cretaceous in the southernhemisphere is marked by strong floral pro-vincialism (Herngreen et al. 1996). A distinctfloral province dominated by trisaccate pollengrains characterized microfloras of Australia,India, New Zealand, southern Africa andsouthern South America (Herngreen et al. 1996,and references therein) (Fig. 5). In particular,there is a steep floristic gradient through SouthAmerica that has been recognized from micro-floras. For example, in Aptian strata Classopolliscan account for 30-60% of certain palynoflorasby abundance in low latitudes, but drops sharplyin high latitudes, a feature also noted byDettmann & Thomson (1987). In the AntarcticPeninsula region, Classopollis rarely accounts formore than 5% of the residue and is often absent(Fig. 6). This type of pattern is repeated for anumber of groups including Elaterate (Gnetales)and angiosperm pollen. Furthermore the reversepattern is true, with some taxa being morediverse in the high latitudes. For example,hepatophytes make up to 18% of the within-floradiversity in high-latitude sites, yet they arealmost absent from mid-latitude floras (Cantrill1997) (Fig. 6). Similar but less extreme examplesinclude the ferns and lycopsids.
Equally, within the large Trisaccate Provincethat covers Australia, India, New Zealand,Antarctica, southern Africa and southern SouthAmerica there is floristic regionalism. Thisphytogeographic floral province can be sub-divided into two regions: the Cyclusphaera-Classopollis subprovince (Africa plus SouthAmerica), and the Murospora subprovince(Australia, India) (Herngreen et al. 1996).Within these two provinces there are importantvariations. Microcachvridites antarcticus, one ofthe elements that defines the province, typicallyconstitutes up to 25% by abundance of palyno-residues at Australian localities (Fig. 5). Incontrast, M. antarcticus is never more than 5-6%by abundance in the Antarctic Peninsulamicrofloras (Fig. 5). These regional differencesalso extend to the macrofloras and provideindirect support for a late arrival of angiospermsin the Antarctic Peninsula. One group that isapparently replaced by the angiosperms is thebennettites. In Australia, bennettites arecommon in Valanginian to early Aptianmacrofloras of southeastern Australia but dis-appear by the Albian (Douglas 1969). In theAntarctic Peninsula macrofloras, bennettites arecommon in the Aptian, and to a lesser extent theAlbian, but they persist until Coniacian times.This time lag in regional extinction supports theidea that angiosperms arrived later in theAntarctic Peninsula region than elsewhere inGondwana.
One explanation for the steep floristic gradi-ents observed through South America and intothe Antarctic Peninsula is a strong climaticgradient (Dettmann 1986a). Recent climaticmodelling suggests that the thermal and effec-tive precipitation gradients through this latitudi-nal transect were extreme (Valdes et al. 1996),and this is supported by the plant fossils. A com-parison of Aptian macrofloras between theAntarctic Peninsula (Cerro Negro Formation)and southern South America (Baquero For-mation) indicates important differences.Antarctic Peninsula floras are high in cool- andmoisture-loving forms (e.g. hepatophytes. ferns,etc.). In contrast, the Baquero Formation isdominated by desiccation-tolerant, warm-lovingplants (e.g. bennettites and other seed-plants).This can be seen in the cuticular adaptations ofthe Baquero Formation plants with stronglysunken stoma, highly papillate leaves and thickcuticles (Archangelsky et al. 1995). Althoughsome of the plant groups are represented in bothfloras, those such as the bennettites from thehigher latitude sites of the Antarctic Peninsulahave thin cuticles, and appear to lack theadaptations to cope with warm, desiccating

ANTARCTIC CRETACEOUS BIODIVERSITY CHANGE 149
conditions. It could well be that this extremethermal and moisture gradient acted as an effec-tive barrier to migration into the AntarcticPeninsula region, and that the initial radiation ofangiosperms across Gondwana was not via theAntarctic Peninsula but by some other route.
Peak of Cretaceous warmth as a forcingmechanismIt is probably no coincidence that the arrival ofangiosperms and their subsequent rise to domi-nance in the high southern latitudes correspondto a global climatic warming trend. The peak ofthe Cretaceous greenhouse was in the Turonian(Huber 1998) and this coincides with the peak ofangiosperm diversity in the Antarctic Peninsula(Fig. 4E). Certainly if the thermal regime thatexisted in the Early Cretaceous was a barrier tomigration, then global warming would haveresulted in latitudinal range expansion as cli-matic zones widened. This would have pushedfloristic boundaries southwards, and ultimatelyhave led to angiosperm arrival in the AntarcticPeninsula region.
Following the peak of Cretaceous warmth inthe Turonian, the high southern latitudesremained warm until late Santonian to earlyCampanian times when cooling started to occur(Huber 1998). This latest Cretaceous period sawthe origin of a number of groups that todaycharacterize temperate southern floras (e.g.Nothofagus, certain groups in the Proteaceaeand Myrtaceae; Dettmann 1989). Furthermore,events such as the uplift of the Andes (c. 80 MaBP) and a general cooling trend through the latestCretaceous provided opportunities for these taxato expand northwards. As the globe cooledthe tropical biomes contracted equatorwardscreating space for temperate biota to expand. Agood example of this is the fern family Lopho-soriaceae, a group that arose in the AntarcticPeninsula regions during the earliest Cretaceous(Berriasian) (Dettmann 1986a; Cantrill 1998),appeared later in southern South America(Aptian-early Albian), and later still in Colombia(Pleistocene). Although today it occupies anextreme latitudinal range along the Andeanchain, it is clear that the closer to the tropics itoccurs, then the higher (cooler) the altitudinalsetting (Cantrill 1998). Clearly for this group,migration is incumbent upon a suitable habitateither provided by cooling climates or thecreation of high-altitude sites. Similar patterns oflater northward inceptions are seen in the earliestappearance of Nothofagus in the Antarctic Penin-sula (early Campanian) and subsequent spread tosouthern South America (Maastrichtian).
SummaryPatterns of floristic replacement through theCretaceous in the high southern latitudes occuramongst the understorey and colonizer elementsof the vegetation, suggesting that this was theniche that the angiosperms initially occupied.This is a pattern similar to that seen elsewhere inthe world (Lidgard & Crane 1988), and is sup-ported by the lack of angiosperm wood in theseearly deposits, despite the presence of pollen.Later stage invasion occurred in the overstoreywith the appearance of tree species as attested toby abundant angiosperm wood in the LateCretaceous sequences (Chapman & Smellie1992; Poole & Francis 1999, 2000; Poole et al2000a,b,c; Poole & Cantrill 2001; Poole &Gottwald 2001).
The timing of the radiation into the Antarc-tica Peninsula implies that this region was notthe initial gateway for invasion of Gondwana byangiosperms. It seems most likely that the initialinvasion occurred through Africa or India butfurther work is needed to refine not only theplate tectonic reconstructions but also the floris-tic history of these areas. The macro- andmicrofloral record is not well known for south-ern Africa, and is poorly age-constrained forIndia, making it difficult to discriminate betweenthese hypotheses. Despite the terrestrial con-nection between South America and theAntarctic Peninsula, it is suggested that a steepclimatic gradient was probably responsible forblocking the early radiation through this region.However, mid-Cretaceous (Turonian) warmingresulted in latitudinal range expansion of cli-matic belts and appears to have been the forcingmechanism for the radiation of angiosperms intothe Peninsula.
ReferencesARCHANGFLSKY, A., ANDREIS, R. R., ARCHANGELSKY,
S. & ARIA BE, A. 1995. Cuticular charactersadapted to volcanic stress in a new Cretaceouscycad leaf from Patagonia, Argentina. Consider-ations on the stratigraphy and depositionalhistory of the Baquero Formation. Review ofPalaeobotany and Palynology, 89, 213-233.
ASKIN, R. A. 1983. Tithonian (uppermost Jurassic) -Barremian (Lower Cretaceous) spore, pollen andmicroplankton from the South Shetland Islands,Antarctica. In: OLIVER, R. L., JAMES, P. R. &JAGO, J. B. (eds) Antarctic Earth Science. Aus-tralian Academy of Sciences, Canberra, 295-297.
ASKIN, R. A. 1990. Cryptogam spores from the upperCampanian and Maastrichtian of Seymour Island,Antarctica. Micropaleontology, 36, 141-156.
ASKIN, R. A. 1994. Monosulcate angiosperm pollenfrom the Lopez de Bertodano Formation (upper

150 DAVID J. CANTRILL & IMOGEN POOLE
Campanian-Maastrichtian-Danian) of SeymourIsland, Antarctica. Review of Palaeobotany andPalynology, 81, 151-164.
BANERJI, J. & LEMOIGNE, Y. 1987. Significant additionsto the Upper Triassic flora of Williams Point,Livingston Island, South Shetlands (Antarctica).Geobios, 20, 469-487.
BARALE, G., PHILIPPE, M, TORRES, T. & THEVENARD.F. 1995. Reappraisal of the Triassic flora fromWilliams Point, Livingston Island (South Shet-land Islands, Antarctica): systematical, biostrati-graphical and paleogeographical implications.Serie Cientifica del Instituto Antarctica Chileno,45, 9-38.
BARREDA, V., PALAMARCZUK, S. & MEDINA, F. 1999.Palinologia de la Formacion Hidden Lake (Coni-anciano-Santoniano), Isla James Ross, Antartida.Revista Espanola de Micropalaeontologia, 31,53-72.
BURGER. D. 1980. Palynology of the lower Cretaceousin the Surat Basin. Bureau of Mineral Resources,Geology and Geophysics Bulletin, 189,1-106.
BURGER, D. 1981. Observations on the earliestangiosperms development with special referenceto Australia. Proceedings of the 4th InternationalPalynological Conference, Lucknow, 3, 418-428.
BURGER. D. 1988. Early Cretaceous environments inthe Eromanga Basin: palynological evidence fromGSQ Wyandra 1 corehole. Australasian Associ-ation of Palaeontologists Memoir, 5, 173-186.
BURGER, D. 1990. Early Cretaceous angiosperms fromQueensland. Australia. Review of Palaeobotanyand Palynology, 65. 153-163.
BURGER. D. 1993. Early and middle Cretaceousangiosperm pollen grains from Australia. Reviewof Palaeobotany and Palynology, 78, 183-234.
CANTRILL. D. J. 1995. The occurrence of the fernHausmannia Dunker (Dipteridaceae) in theCretaceous of Alexander Island, Antarctica.Alchennga, 19. 243-254.
CANTRILL. D. J. 1996. Fern thickets from theCretaceous of Alexander Island, Antarcticacontaining Alamatus bifarius Douglas and Aculeaacicularis sp. nov. Cretaceous Research, 17.169-182.
CANTRILL, D. J. 1997. Hepatophytes from the EarlyCretaceous of Alexander Island, Antarctica: sys-tematics and palaeoecology. International Journalof Plant Sciences, 158, 476-488.
CANTRILL, D. J. 1998. Early Cretaceous fern foliagefrom President Head, Snow Island, Antarctica.Alchennga, 22, 241-258.
CANTRILL, D. J. 2000. A Cretaceous (Aptian) florafrom President Head, Snow Island, Antarctica.Palaeontographica B, 253. 153-191.
CANTRILL, D. J. & FALCON-LANG. H. J. 2001.Cretaceous (Late Albian) Coniferales ofAlexander Island, Antarctica. Part 2: Foliage,reproductive structures and roots. Review ofPalaeobotany and Palynology, 115, 119-145.
CANTRILL, D. J. & NICHOLS, G. J. 1996. Taxonomy andpalaeoecology of Early Cretaceous (Late Albian)angiosperm leaves from Alexander Island,Antarctica. Review of Palaeobotany and Palvnol-ogy, 92. 1-28.
CESARI, S., PARICA, C. REMESAL. M. & SALANL F.1998. First evidence of Pentoxylales in Antarctica.Cretaceous Research, 19, 733-743.
CESARI, S., PARICA, C., REMESAL. M. & SALANL F.1999. Palaeoflora del Cretacico Inferior de penin-sula Byers, islas Shetland del Sur. Antartida,Ameghiniana, 36. 3-22.
CHAPMAN. J. L. & SMELLIE. J. L. 1992. Cretaceousfossil wood and palynomorphs from WilliamsPoint, Livingston Island, Antarctic Peninsula.Review of Palaeobotanv and Palvnology, 74.163-192. '
CRANE, P. R. & LIDGARD, S. 1989. Angiospermdiversification and paleolatitudinal gradients inCretaceous floristic diversity. Science. 246.675-678.
CRANE, P. R.. FRIIS. E. M. & PFDERSEN. K. R. 1995.The origin and early diversification ofangiosperms. Nature, 374, 27-33.
DETTMANN, M. E. 1986a. Significance of theCretaceous-Tertiary spore genus Cyatheaciditesin tracing the origin and migration of Lophosoria(Filicopsida). Special Papers in Paleontology, 35.63-94.
DETTMANN, M. E. 1986b. Early Cretaceous palyno-flora of subsurface strata correlative with theKoonwarra Fossil Bed. Victoria. AustraliasianAssociation of Palaeontologists Memoir. 3.79-110.
DETTMANN, M. E. 1989. Antarctica: Cretaceous cradleof austral temperate rainforests? In: CRAME J. A.(ed.) Origins and Evolution of the Antarctic Biota.Geological Society. London, Special Publications.47.89-105.
DETTMANN. M. E. 1994. Cretaceous vegetation: themicrofloral record. In: HILL. R. S. (ed.) History ofthe Australian Vegetation, Cretaceous to Recent.Cambridge University Press. Cambridge.143-170.
DETTMANN. M. E. & THOMSON. M. R. A. 1987.Cretaceous palynomorphs from the James RossIsland area - a pilot study. British AntarcticSurvey Bulletin. 77. 13-59.
DOLDING. P. J. D. 1992. Palynology of the MarambioGroup (Upper Cretaceous) of northern HumpsIsland. Antarctic Science, 4. 311-326.
DOUGLAS. J. G. 1969. The Mesozoic floras of Victoria.Parts 1 & 2. Geological Survey of VictoriaMemoir, 28. 1-310.
DOUGLAS. J. G. 1973. The Mesozoic floras of Victoria.Part 3. Geological Survey of Victoria Memoir. 29.1-185.
D R I N N A N , A. N. & CHAMBERS. T. C. 1986. Flora of theLower Cretaceous Koonwarra Fossil Bed(Korumburra Group). South Gippsland. Victoria.Association of Australasian PalaeontologistsMemoir. 3, 1-77.
D R I N N A N , A. N. & CRANE. P. R. 1990. Cretaceouspaleobotany and its bearing on the biogeographyof austral angiosperms. In: TAYLOR, T. N. &TAYLOR. E. L. (eds) Antarctic Paleobiology: itsRole in the Reconstruction ofGondwana. SpringerVerlag. New York. 192-219.
DUANE, A. M. 1996. Palynology of the Byers Group(late Jurassic-Earlv Cretaceous) of Livingston

ANTARCTIC CRETACEOUS BIODIVERSITY CHANGE 151
and Snow Island, Antarctic Peninsula: its bio-stratigraphic and palaeoenvironmental signifi-cance: Review of Palaeobotany and Palynology,91, 241-281.
FALCON-LANG, H. J. & CANTRILL, D. J. 2000,Cretaceous (Late Albian) Coniferales ofAlexander Island, Antarctica. Part 1: Wood tax-onomy: a quantitative approach. Review ofPalaeobotany and Palynology, 111, 1-17.
FALCON-LANG, H. J., CANTRILL, D. J. & NICHOLS, G. J.2001. Biodiversity and terrestrial ecology of amid-Cretaceous, high-latitude floodplain,Alexander Island, Antarctica. Journal of theGeological Society, London, 150, 709-724.
HATHWAY, B., DUANE, A. M., CANTRILL, D. J. &KELLEY, S. P. 1999. 40Ar/39Ar geochronology andpalynology of the Cerro Negro Formation, SouthShetland Islands, Antarctica: a new radiometrictie for Cretaceous terrestrial biostratigraphy inthe Southern Hemisphere. Australian Journal ofEarth Sciences, 46, 593-606.
HAY, W. W., DE CONTO, R. M., WOLD, C. N., ET AL.1999. An alternative global Cretaceous palaeo-geography. In: BARRERA, E. & JOHNSON, C. (eds)Evolution of the Cretaceous Ocean-ClimateSystem. Geological Society of America, SpecialPapers, 332,1-48.
HAYES, P. 1999. Cretaceous Angiosperm Floras ofAntarctica, PhD Thesis, University of Leeds.
HERNANDEZ, P. J. & AZCARATE, V. 1971. Estudiopaleobotanico preliminar sobre restos de unatafoflora de la Peninsula Byers (Cerro Negro),Isla Livingston, Islas Shetland del Sur, Antartica.Serie Cientifica del Instituto Antdrctico Chileno, 2,15-50.
HERNGREEN, G. F. W. 1975. Palynology of Middle andUpper Cretaceous strata in Brazil. MededelingenRijks Geologische Dienst Nieuwe Serie, 26, 39-90.
HERNGREEN, G. F. W., KEDVES, M., ROVINA, L. V. &SMIRNOVA, S. B. 1996. Cretaceous palynofloralprovinces: a review. In: JANSONIUS, J. &MCGREGOR, D.C. (eds) Palynology: Principles,and Applications. American Association ofStratigraphic Palynologists Foundation, 3,1157-1188.
HILL, R. S. & SCRIVEN, L. J. 1995. The angiosperm-dominated woody vegetation of Antarctica: areview. Review of Palaeobotany and Palynology,86,175-198.
HUBER, B. T. 1998. Tropical paradise at theCretaceous poles? Science, 282, 2199-2200.
KEATING. J. M. 1992. Palynology of the LachmansCrags Member, Santa Marta Formation (UpperCretaceous) of north-west James Ross Island.Antarctic Science, 4, 293-304
KEATING, J. M., SPENCER-JONES, M. & NEWHAM, S.1992. The stratigraphical palynology of the KotickPoint and Whisky Bay formations, Gustav Group(Cretaceous), James Ross Island. AntarcticScience, 4, 279-292.
KRAUSE, D. W., ROGER, R. R., FORSTER, C. A.,HARTMAN, J. H. & BUCKLEY, G. A. 1999. TheLate Cretaceous vertebrate fauna of Madagascar:implications for gondwanan palaeobiogeography.GSA Today, 9(8), 1-7.
LACEY, W. S. & LUCAS, R. C. 1981. The Triassic floraof Livingston Island, South Shetland Islands.British Antarctic Survey Bulletin, 53,157-173.
LAWVER, L. A., GAHAGAN, L. M. & DALZIEL, I. W. D.1997. A tight fit early Mesozoic Gondwana, aplate reconstruction perspective. Memoirs of theNational Institute of Polar Research, 53, 214-229.
LIDGARD, S. & CRANE, P. R. 1988. Quantitative analy-ses of the early angiosperm radiation. Nature, 331,344_346.
LIDGARD, S. & CRANE, P. R. 1990. Angiosperm diver-sification and Cretaceous floristic trends: a com-parison of palynofloras and leaf macrofloras.Paleobiology, 16, 77-93.
LIVERMORE, R. A. & HUNTER, R. J. 1996. Mesozoicseafloor spreading in the southern Weddell Sea.In: STOREY, B. C., KING, E. C. & LIVERMORE, R.A. (eds) Weddell Sea Tectonics and GondwanaBreak-up. Geological Society, London, SpecialPublications, 108, 227-241.
MARKS, K. M. & TIKKU, A. A. 2001. Cretaceous re-constructions of East Antarctica, Africa andMadagascar. Earth and Planetary Science Letters,186, 479-495.
METCALFE, I. 1990. Allochthonous terrane processesin Southeast Asia. Philosophical Transactions ofthe Royal Society London A, 331, 625-640.
MULLER, R. D., MIHUT, D. & BALDWIN, S. 1998. A newkinematic model for the formation and evolutionof the west and northwest Australian margin. In:PURCELL, P. G. & PURCELL, P. R. (eds) TheSedimentary Basins of Western Australia. Petrol-eum Exploration Society of Australia, Perth,55-71.
ORLANDO, H. A. 1968. A new Triassic flora fromLivingston Island, South Shetland Islands. BritishAntarctic Survey Bulletin, 16,1-13.
POLE, M. S. & DOUGLAS, J. G. 1999. Bennettitales,Cycadales and Ginkgoales from the mid-Cretaceous of the Eromanga Basin, Queensland,Australia. Cretaceous Research, 20, 523-538.
PONS, D. 1988. Le Mesozoique de Colombie macro-flores et microflores. CNRS, Paris.
POOLE, I. & CANTRILL, D. J. 2001. Fossil woods fromWilliams Point Beds, Livingston Island, Antarc-tica: a Late Cretaceous southern high latitudeflora. Palaeontology 44,1081-1112.
POOLE, L & FRANCIS, J. E. 1999. The first record offossil atherospermataceous wood from the UpperCretaceous of Antarctica. Review of Palaeo-botany and Palynology, 107, 97-107.
POOLE, I. & FRANCIS, J. E. 2000. The first record offossil wood of Winteraceae from the UpperCretaceous of Antarctica. Annals of Botany, 85.307-315.
POOLE, I. & GOTTWALD, H. 2001. Monimiaceae sensulato, an element of gondwanan polar forests: evi-dence from the Late Cretaceous-Early Tertiarywood flora of Antarctica. Australian SystematicBotany, 14, 207-230.
POOLE, L, CANTRILL, D. J., HAYES, P. & FRANCIS, J. E.2000a. The fossil record of Cunoniaceae: new evi-dence from Late Cretaceous wood of Antarctica?Review of Palaeobotany and Palynology, 111.127-144.

152 DAVID J. CANTRILL & IMOGEN POOLE
POOLE, I, GOTTWALD, H. & FRANCIS, J. E. 2000b.Illicioxylon, an element of Gondwanan Polarforests? Late Cretaceous and early Tertiarywoods of Antarctica. Annals of Botany, 86,421-432.
POOLE, L, RICHTER, H. G. & FRANCIS, J. E. 2000c.Evidence for Gondwanan origins for Sassafras(Lauraceae)? Late Cretaceous fossil wood ofAntarctica. International Association of WoodAnatomists Journal, 21, 463-475.
PRAMPARO, M. B. & VOLKHEIMER, W. 1999. Pali-nologia del Miembro Avile (Formacion Agrio,Cretacico Inferior) en la cerro de la Parva,Neuquen. Ameghiniana, 36, 217-227.
REES, P. M. & SMELLIE, J. L. 1989. Cretaceousangiosperms from an allegedly Triassic flora atWilliams Point, Livingston Island, South ShetlandIslands. Antarctic Science, 1, 239-248.
REEVES, C. V. & DE WIT, M. J. 2000. Making ends meetin Gondwana: retracing the transforms of theIndian Ocean and reconnecting continental shearzones. Terra Nova, 12, 272-280.
RICCARDI, A. C. 1988. The Cretaceous system ofsouthern South America. Geological Society ofAmerica Memoir, 168,1-161.
RIDING, J. B., CRAME, J. A., DETTMANN, M. E. &CANTRILL, D. J. 1998. The age of the base of theGustav Group in the James Ross Basin, Antarc-tica. Cretaceous Research, 19, 87-105.
ROESER, H. A., FRITSCH, J. & HINZ, K. 1996. Thedevelopment of the crust of Dronning MaudLand, East Antarctica. In: STOREY, B. C., KING, E.C. & LIVERMORE, R. A. (eds) Weddell SeaTectonics and Gondwana Break-up. GeologicalSociety, London, Special Publications, 108,243-264.
ROYER. J-V. & COFFIN, M. F. 1992. Jurassic to Eoceneplate tectonic reconstructions in the KerguelenPlateau region. Proceedings of the Ocean DrillingProgram, Scientific Results, 120, 917-928.
SAMPSON, S. D., WITMER, L. M., FORSTER, C. A.,KRAUSE, D. W., O'CONNOR, P. M., DODSON, P. &RAVOAVY, F. 1998. Predatory dinosaur remainsfrom Madagascar: implications for theCretaceous biogeography of Gondwana. Science,280, 1048-1051.
SEGOUFIN, J. & PATRIAT, P. 1980. Existence d'anom-alies mesozoiques dans le bassin de Somalie.
Implications pour les relations Afrique-Antarc-tique-Madagascar. Compte Rendu AcademieSciences Paris, 291. 85-88.
SHIPBOARD SCIENTIFIC PARTY. 2000. Leg 183 summary:Kerguelen Plateau-Broken Ridge—A largeigneous province. In: COFFIN. M. F., FREY, F. A.,WALLACE. P. J., ET AL. (eds) Proceedings OceanDrilling Program, Initial Reports. 183, 1-101.
SPICER. R. A. 1988. Quantitative sampling of plantmegafossil assemblages. Paleontological SocietySpecial Publication, 3, 29-51.
STOREY, B. C.. DALZIEL. I. W. D.. GARRETT. S. W..GRUNOW, A. M., PANKHURST. R. J. & VENNUNL W.R. 1988. West Antarctic in Gondwanaland: crustalblocks, reconstruction and break-up processes.Tectonophysics, 155. 381-390.
TAYLOR, D. W. & HICKEY. L. J. 1990. An Aptian plantwith attached leaves and flowers: implication forangiosperm origin. Science, 247. 702-704.
TORRES, T., BARALE, G., THEVERNARD, F.. PHILIPPE.M. & GALLEGUILLOS. H. 1997. Morfologia y sis-tematica de la flora del Cretacico Inferior dePresident Head, isla Snow, archipelago de lasShetland del Sur, Antartica. Serie Cientifica delInstituto Antarctico Chileno, 2, 15-50.
TRUSWELL. E. M. 1991. Antarctica: a history of terres-trial vegetation. In: TINGEY. R, J. (ed.) TheGeology of Antarctica. Oxford Monographs onGeology and Geophysics. 17. Clarendon Press,Oxford, 499-537.
TRUSWELL, E. M., KERSHAW, A. P. & SLUITER, L R.1987. The Australian-south-east Asian connec-tion: evidence from the palaeobotanical record.In: WHITEMORE, T. C. (ed.) BiogeographicalEvolution of the Malay Archipelago OxfordUniversity Press, Oxford, 32-49.
TRUSWELL. E. M.. DETTMANN, M. E. & O'BRIEN. P. E.1999. Mesozoic palynofloras from the Mac.Robertson Shelf, East Antarctica: geological andphytogeographic implications. Antarctic Science,11,239-255.
VALDES. P. J.. SELLWOOD, B. W. & PRICE. G. D. 1996.Evaluating concepts of Cretaceous equability.Palaeoclimates, 2, 139-158.
VEEVERS. J. J.. POWELL, C. McA. & ROOTS. S. R. 1991.Review of seafloor spreading around Australia. 1.Synthesis of the patterns of spreading. AustralianJournal of Earth Sciences, 38. 373-389.

Cenozoic palaeogeography and the rise of modern biodiversitypatterns
J.A. CRAME1 & B.R. ROSEN2
1 British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 OET, UK(e-mail: [email protected],)
2 Department of Palaeontology, The Natural History Museum, Cromwell Road,London SW75BD, UK (e-mail: [email protected])
Abstract: The steepest latitudinal and longitudinal gradients in taxonomic diversity at thepresent day are those associated with tropical high diversity foci. Although there has beena tendency in the past to regard these features as either evolutionary 'cradles' or 'museums'of considerable antiquity, this may not be the case. Within the marine realm, a uniform,pan-tropical fauna was progressively disrupted by a series of plate tectonic events, the mostimportant of which were the Early Miocene (c, 20 Ma) collisions of Africa/Arabia withEurope and Australia/New Guinea with Indonesia, and the Middle Miocene-latestPliocene rise of the Central American Isthmus. This had the net effect of establishing twomain tropical high diversity foci: the Indo-West Pacific and the Atlantic-Caribbean-EastPacific. Similar foci were also established at the same time in the terrestrial realm.
Together with the physical isolation of Antarctica, these same tectonic events con-tributed significantly to global cooling throughout the Cenozoic Era. This in turn led to theimposition of a series of thermally defined provinces, and thus a considerable degree ofbiotic differentiation on a regional scale. However, something else seems to have beeninvolved in the creation of very steep tropical diversity peaks. This could in part be acoincidental radiation of a series of unrelated taxa, or some sort of evolutionary feedbackmechanism between interacting clades. Alternatively, Late Cenozoic rates of originationmay have been enhanced by an external forcing mechanism such as changes in OrbitalRange Dynamics.
Despite the enormous increase in biodiversity diversity are equally impressive contemporaryresearch in recent years we are still a long way biogeographical phenomena (Crame 2000a,b,from understanding the true nature of the and references therein). They are of particularlarger-scale patterns of life on Earth. There is a interest as they are far less likely to co-vary withgeneral appreciation that, for many groups of any known environmental parameter than lati-plants and animals, there are simply more of tudinal gradients (but see Bellwood & Hughesthem in the tropical and low-latitude regions 2001).than in the high-latitude and polar ones, and We also lack a convincing integrated theory orlatitudinal gradients in taxonomic diversity model to account for the origins of large-scaleconstitute one of the most striking macroeco- biodiversity patterns in both time and spacelogical patterns at the present day (e.g. Gaston (Gaston 2000). Are they, as some have argued.,& Blackburn 2000, and references therein). In the product of the 'ecological moment', repre-addition we are also beginning to understand senting no more than the adjustment of regionalthat tropical high diversity is distinctly hetero- biotas to climates and habitats that have devel-geneous. Two tropical hotspots or high diversity oped since the Last Glacial Maximum (i.e. somefoci are often distinguished in the marine realm, 18 ka ago)? Or are there longer-term processesone in the Indo-West Pacific (IWP) and the involved too, stretching back over a geologicalother in the Atlantic, Caribbean and East Pacific timescale of millions or even tens of millions ofregion (ACEP) (sensu Ellison et al. 1999). Anal- years? There has recently been a considerableogous patterns have now been established in development in our understanding of the waysthe terrestrial realm too, and in a global study of in which geological processes could influence thethe family richness of seed plants, amphibians, formation of large-scale biodiversity patterns,reptiles and mammals, Williams et al. (1997) and this has led to an intense debate on thefound hotspots of maximum richness in Colom- relative roles of contemporary versus historicalbia, Nicaragua, Oaxaca (Mexico) and Malaysia, factors in the creation of taxonomic diversityTropical longitudinal gradients in taxonomic gradients (e.g. Turner et al 1996; Francis &
From: CRAME, J.A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations, Geological Society, London, Special Publications, 194,153-168.0305-8719/02/$15.00 © The Geological Society of London 2002.

154 J. A. CRAME & B. R. ROSEN
Currie 1998; Huston 1999; Ricklefs 1999; Qian &Ricklefs 2000).
It is the intention of this study to further thisline of enquiry by focusing on a series of majortectonic and climatic events that occurredthroughout the Cenozoic Era (i.e. the last 65 Maof Earth history). It will be suggested that thiswas a time of major expansion in the size andscale of taxonomic diversity gradients and thequestion asked whether this can be attributeddirectly to the relative isolation of the conti-nents, fundamental reorganization of oceaniccirculation systems and progressive impositionof a steep latitudinal temperature gradient. Thepotential role of biotic factors in promoting themassive Cenozoic radiation of selected tropicaltaxa will not be ignored, but the emphasis herewill essentially be on evolving Cenozoic palaeo-geographies and how they affected such keyprocesses as climate change, the establishmentof new colonization and dispersal routes, and,possibly, pronounced changes in origination(speciation) and extinction rates.
Cenozoic global diversificationThere is general agreement that there was a verysubstantial rise in global biodiversity throughthe Cenozoic Era. This was particularly so at thelower taxonomic levels of species and genuswhere the rise was the steepest and most pro-longed of the entire Phanerozoic interval(Signer 1990; Benton 1999; Foote 2000). Wecould perhaps be looking at an order of magni-tude increase in species diversity at this time(Signer 1990; but see also Alroy et al 2001).Equally striking is the fact that this radiation isrecorded across such a broad range of organismsin both the marine and terrestrial realms. Thiswas the time of widespread proliferation ofgroups such as the neogastropods, heteroconchbivalves, cheilostome bryozoans, decapod crus-taceans and teleost fish in shallow seas, andamphibians, reptiles, birds, mammals, floweringplants and insects on land (e.g. Hallam 1994).Parallel temporal trends in so many differentgroups has been taken as a strong indication thatthere may be a single, basic mechanism under-pinning the process of Cenozoic biodiversifica-tion (Signer 1990; Benton 1999).
The presence of very clear latitudinal gradi-ents at the present day strongly suggests that thispulse of Cenozoic diversification was concen-trated in low-latitude and tropical regions.Unfortunately, we cannot yet demonstrate thisby reconstructing diversity gradients directlyfrom the fossil record, but a number of indirectlines of evidence suggest that this was indeed the
case. For example, in a recent study of latestMesozoic-Cenozoic bivalve diversification,Crame (2000a) demonstrated that the steepestlatitudinal gradients occurred in the youngestclades (and especially in the very large hetero-conch clade). He went on to suggest that thesame phenomenon may occur within the marinegastropods, where the youngest major clade, theneogastropods, shows particularly strong latitu-dinal gradients (Crame 2001). In the terrestrialrealm the three largest core eudicot angiospermclades are amongst the youngest, and verysimilar trends are shown by various insects,birds, mammals, fish and reptiles. There isgrowing evidence to suggest that the age ofcertain major clades and the strength of theirlatitudinal gradients are consistently negativelycorrelated (Crame 2001).
Cenozoic global coolingOne of the most striking and puzzling aspects ofthe Cenozoic biodiversification event is that ittook place against a backdrop of global climaticdeterioration. From an Early Eocene climaticoptimum (55 Ma BP), global climates deterio-rated in a series of pronounced steps to theirpresent-day values. Although the Eocene seemsto have been an epoch of relative global warmth,a 5°C drop in oceanic surface water tempera-tures at the Eocene-Oligocene boundary (33Ma BP) is widely interpreted to mark the onsetof permanent Antarctic glaciation. Furtherabrupt cooling phases occurred in the Middle-Late Miocene (approx. 15-13 Ma BP), latestMiocene-earliest Pliocene (approx. 6.2-4.8Ma BP), Middle-Late Pliocene (3.6-2.4 Ma BP),and Late Pliocene-Recent. Even during theNeogene climatic optimum (early MiddleMiocene, 17-15 Ma BP) average marine globaltemperature values only briefly returned tothose of the Late Eocene (Clarke & Crame 1992;Frakes et al 1992; Zachos et al 2001).
The principal effect of these changes withinthe tropical and subtropical regions was not somuch a reduction in temperature as one inarea. It has been estimated that, since theEarly-Middle Eocene (i.e. the last 45 Ma), thetropical biome has contracted some 40-50% insize (Adams et al 1990). In other words, themarked Cenozoic diversification event tookplace as the tropics actually contracted in sizeand the temperate and polar regions expanded.The principal steps in the physical control of thisprocess are outlined below.
One of the crucial early Cenozoic palaeogeo-graphic events to affect global oceanographiccirculation patterns and climates was the
CENOZOIC BIODIVERSITY 155
Fig. 1. The physical isolation of Antarctica. Late Eocene and Middle Miocene south polar projections redrawnand simplified from Lazarus & Caulet (1993, figs 19 and 22), with some additional detail from maps containedin Lawyer el al. (1992). Key: stipple, continents; grey shading, continental shelf; arrows, principal currentdirections.

156 J. A. CRAME & B. R. ROSEN
physical isolation of Antarctica. Althoughlargely separated from the other southern conti-nents by the end of the Cretaceous Period, therewere still intermittent terrestrial and shallow-water connections between both easternAntarctica and southeasternmost Australia, andthe Antarctic Peninsula and southernmostSouth America (Crame 1999, fig, IB) (Fig. 1).Full, deep-water separation of the SouthTasman Rise from Antarctica was probably notachieved until the late Early Eocene (50 Ma BP),by which time the continent must have beenalmost completely isolated by deep water. Thefirst indications of global cooling occured in theinterval 50-40 Ma BP, and there were significant2°C temperature falls in both the late MiddleEocene and Middle-Late Eocene boundary(Stott et al 1990). These were followed by amajor cooling event at approximately theEocene-Oligocene boundary (37 Ma BP) whereboth surface and bottom waters may havedecreased by as much as 5°C in 75-100 ka(Lazarus & Caulet 1993; Crame 1999; andreferences therein).
The precise time of opening of the DrakePassage is still uncertain. Whereas marinegeophysical evidence indicates that seafloorspreading began in the early Late Oligocene(28 Ma BP), it may have been as late as 23.5 ±2.5 Ma (Oligocene-Miocene boundary) beforefull deep-water circulation was established(Barker et al 1991). On the other hand, there issome micropalaeontological evidence to suggestthat there was an opening, at least at shallow andintermediate depths, as early as 36 Ma BP(Diester-Haas & Zahn 1996), and a taxonomicassessment of mid-Eocene mammal faunas fromSeymour Island (northern tip of the AntarcticPeninsula) shows that they had been isolatedfrom a South American ancestral stock since atleast the Early Eocene (54-51 Ma BP) (Wood-burne & Case 1996). In any event it is apparentthat a full deep-water, circum-Antarctic currentsystem and polar frontal zone had been estab-lished by the Middle Miocene (15 Ma BP)(Lazarus & Caulet 1993) (Fig. 1). This was thetime of development of a latitudinal tempera-ture gradient very similar to that seen today(Loutit et al 1983; Kennett et al 1985).
Nevertheless, it should also be emphasizedthat other Cenozoic tectonic events almostcertainly contributed to significant globalcooling. Of particular importance was theprogressive northward movement of theAfrica/Arabia landmass that led to the constric-tion of the Tethyan Ocean and its eventualclosure in the Early Miocene (20 Ma BP),followed by the Early-Middle Miocene
(20-15 Ma BP) collision of the Australian platewith Indonesia (Kennett 1977; Kennett et al1985; see below). The net effect of these changeswas to shift oceanic circulation from predomi-nantly equatorial to strongly meridional (i.e.north-south) or gyral; this was especially so inthe Pacific Ocean (Kennett et al 1985; Grigg1988). The transport of warm waters into high-latitude regions is thought to have led toincreased levels of precipitation there which inturn contributed to the gradual build-up ofglacial ice (see also below).
Even Late Neogene palaeogeographicalchanges are thought to have had profoundclimatic implications. For example, the gradualuplift of the Central American Isthmus (CAI,i.e. the Isthmus of Panama), which occurredover the interval of 13-1.9 Ma BP, began to havea major effect on oceanographic circulationpatterns by about 4-6 Ma BP (Coates et al 1992;Haug & Tiedemann 1998). At this time theCentral American Seaway had shallowed to<100 m and the Gulf Stream was beginning todeflect warm, shallow waters northward alongthe eastern US seaboard (Fig. 2). Although thisundoubtedly led to some warming in northernhemisphere mid- to high latitudes, it is apparentthat, as a result of evaporation of surface watersin the relatively narrow North Atlantic Oceanby trade winds, this water would have beenslightly hypersaline. When it reached the north-ern high latitudes it began to descend in both theNorwegian and Labrador seas to form NorthAtlantic Deep Water (NADW). This thenspread into both the South Atlantic and centralPacific to initiate a major 'conveyor belt' ofdeep-ocean circulation (Haua; & Tiedemann1998) (Fig. 2).
Further closure of the CAI saw the NorthAtlantic thermohaline circulation system inten-sify by 3.6 Ma BP, and the Arctic Ocean effec-tively isolated from the warm Atlantic waters. Itis this isolation from the oceans to the south thatled to the drastic temperature decline in thenorth polar regions and the eventual onset ofglaciation at 2.5 Ma BP (Stanley 1995). Changesin the Earth's obliquity amplitude fluctuationsbetween 3.1 and 2.5 Ma BP may also have beenan important contributory factor (Haug &Tiedemann 1998). A further important effect ofthe closure of the CAI was to reverse the flow ofwater through the Bering Strait, and this in turninfluenced Pliocene-Pleistocene patterns ofNorth Atlantic thermohaline circulation(Shaffer & Bendtsen 1994; Marincovich 2000).
It can be concluded that, throughout theCenozoic Era, a series of tectonic changesoccurred that led directly to global cooling. First
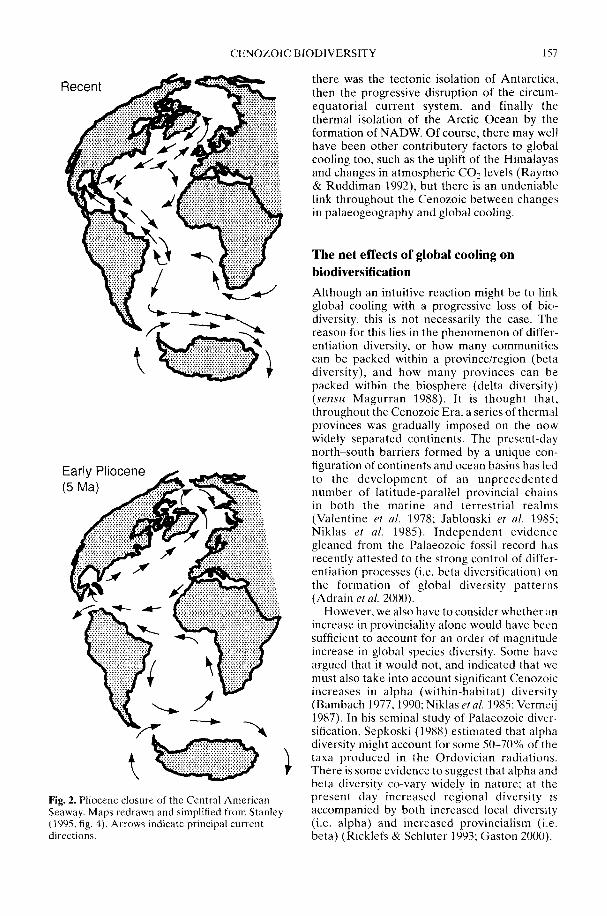
CENOZOIC BIODIVERSITY 157
Fig. 2. Pliocene closure of the Central AmericanSeaway. Maps redrawn and simplified from Stanley(1995, fig. 4). Arrows indicate principal currentdirections.
there was the tectonic isolation of Antarctica,then the progressive disruption of the circum-equatorial current system, and finally thethermal isolation of the Arctic Ocean by theformation of NADW. Of course, there may wellhave been other contributory factors to globalcooling too, such as the uplift of the Himalayasand changes in atmospheric CO2 levels (Raymo& Ruddiman 1992), but there is an undeniablelink throughout the Cenozoic between changesin palaeogeography and global cooling.
The net effects of global cooling onbiodiversification
Although an intuitive reaction might be to linkglobal cooling with a progressive loss of bio-diversity, this is not necessarily the case. Thereason for this lies in the phenomenon of differ-entiation diversity, or how many communitiescan be packed within a province/region (betadiversity), and how many provinces can bepacked within the biosphere (delta diversity)(sensu Magurran 1988). It is thought that,throughout the Cenozoic Era, a series of thermalprovinces was gradually imposed on the nowwidely separated continents. The present-daynorth-south barriers formed by a unique con-figuration of continents and ocean basins has ledto the development of an unprecedentednumber of latitude-parallel provincial chainsin both the marine and terrestrial realms(Valentine et al. 1978; Jablonski et al. 1985;Niklas et al. 1985). Independent evidencegleaned from the Palaeozoic fossil record hasrecently attested to the strong control of differ-entiation processes (i.e. beta diversification) onthe formation of global diversity patterns(Adrain et al. 2000).
However, we also have to consider whether anincrease in provinciality alone would have beensufficient to account for an order of magnitudeincrease in global species diversity. Some haveargued that it would not, and indicated that wemust also take into account significant Cenozoicincreases in alpha (within-habitat) diversity(Bambach 1977,1990; Niklas et al. 1985; Vermeij1987). In his seminal study of Palaeozoic diver-sification, Sepkoski (1988) estimated that alphadiversity might account for some 50-70% of thetaxa produced in the Ordovician radiations.There is some evidence to suggest that alpha andbeta diversity co-vary widely in nature; at thepresent day increased regional diversity isaccompanied by both increased local diversity(i.e. alpha) and increased provincialism (i.e.beta) (Ricklefs & Schluter 1993; Gaston 2000).

158 J. A. CRAME & B. R. ROSEN
Causes of Cenozoic biodiversification
It would appear then that if we are to find a satis-factory explanation for the global Cenozoic bio-diversification event, it must be one that involvesa synchronous increase at all four basic levels(i.e. alpha, beta, gamma and delta diversity). Inother words this must be a process that allowsboth an increase in the numbers of taxa withinindividual habitats as well as increased differen-tiation between these habitats on a variety ofgeographical scales. For example, at one of thevery simplest levels it has been postulated thatthe Cenozoic rise in biodiversity could be linkedto a global rise in trophic resources provided bythe radiation of the angiosperms (Bambach1977). Even in the marine realm the increasedflux of plant detritus through river systems mayhave raised benthic productivity. Similarly, it hasbeen suggested that oceanic nutrient cyclingmay have been influenced by changes in theglobal thermohaline circulation system broughtabout by the formation of Antarctic deepbottom waters at the Eocene-Oligocene bound-ary (Bambach 1977).
Such ideas are indeed appealing, and weshould not lose sight of the fact that there maywell have been times of abrupt increase in bothterrestrial and marine productivity throughEarth history. Nevertheless it is by now wellestablished that there is not necessarily a directlink between increased productivity andincreased biodiversity. An increase in produc-tivity can trigger an increase in biomass, butthere is no reason why this should not, inturn, just be within one or a small number ofspecies. Something else is required to generatea large number of new taxa from a rise inproductivity (Blackburn & Gaston 1996). Wealso have to bear in mind that any potentialmechanism for Cenozoic global biodiversifica-tion must explain the greater relative productionof new taxa in the low- than in the high-latituderegions. In their discussion of the origin anddiversification of major taxonomic groups,Jablonski & Bottjer (1990) identified five basicexplanations for global radiations. We mightrationalize these into two main types: intrinsicand extrinsic.
Intrinsic mechanisms of diversification
One of the simplest explanations for globalCenozoic diversification would be that it rep-resents the coincidental expansion of a series ofunrelated clades. Although key adaptive break-throughs, such as mantle fusion and the develop-ment of posterior siphons in bivalves, or the
development of plants with flowers, undoubtedlyoccurred well before the Cenozoic Era, it mayhave taken periods of tens of millions of years forsuch clades to build up substantial numbers oftaxa (Jablonski & Bottjer 1990). Once certaingroups became established in the latest Meso-zoic/earliest Cenozoic they may in turn havetriggered the automatic radiation of others.Angiosperm assemblages would have provided avariety of novel terrestrial habitats, and the rapiddevelopment of coral reefs, sea-grass beds andmangroves at this time could have promoteddiversification in shallow, tropical seas (Verrneij1977).
It is also possible that intense biological inter-actions between various taxonomic groups werea driving force behind diversification. In particu-lar, if biological hazards due to competitors andpredators have increased through time, then somay the responses of various prey taxa to them.The net result is a sort of 'evolutionary armsrace', with first one group gaining a numericalascendancy, and then the other (Vermeij 1987,1994). This special form of co-evolutionaryrelationship, known as escalation, emphasizesthe importance of enemies as agents of naturalselection: over periods of geological timeenemy-related adaptations bring about long-term evolutionary trends in the morphology,ecology or behaviour of other organisms. It isessentially a tropical phenomenon and isparticularly well represented in species-richgroups, such as angiosperms, arthropods, verte-brates and molluscs, that have highly developedcompetitive and defensive capabilities (Vermeij1987). Striking examples would appear to be theco-radiation of angiosperms and pollinatinginsects in the terrestrial realm, and the linkbetween durophagous predators and bothdeeper-burrowing infauna and more heavilyornamented epifauna in the marine one.
Although escalation seems to be an intuitivelysatisfying explanation for certain tropical radia-tions, some doubts have been raised as to itswidespread efficacy. For example, Vermeij(1987) himself drew attention to the low inci-dence of damage repair within many infaunalbivalve groups, and the high-resolution fossilrecord in general has comparatively fewexamples of gradual morphological changeinduced within a prey lineage (Hansen et al. 1999;Dietl et al 2000; Miller 2000). Many biotic tran-sitions within the fossil record could equallywell be interpreted as the replacement of -incumbent taxa by some form of fundamentalenvironmental change (Jablonski 2000). Non-competitive expansions may be just as commonthrough geological time (Benton 1999).
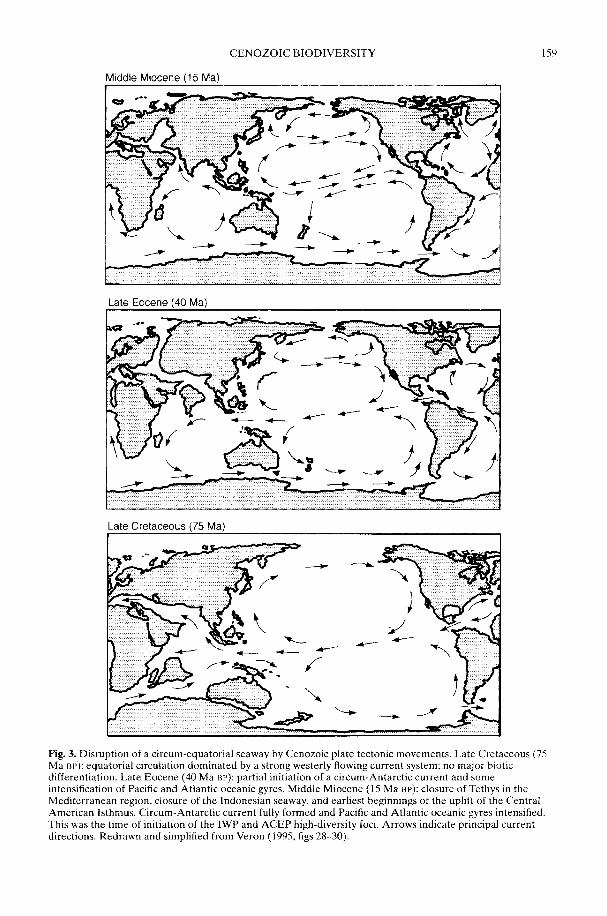
CENOZOIC BIODIVERSITY 159
Fig. 3. Disruption of a circum-equatorial seaway by Cenozoic plate tectonic movements. Late Cretaceous (75Ma BP): equatorial circulation dominated by a strong westerly flowing current system; no major bioticdifferentiation. Late Eocene (40 Ma BP): partial initiation of a circum-Antarctic current and someintensification of Pacific and Atlantic oceanic gyres. Middle Miocene (15 Ma BP): closure of Tethys in theMediterranean region, closure of the Indonesian seaway, and earliest beginnings of the uplift of the CentralAmerican Isthmus. Circum-Antarctic current fully formed and Pacific and Atlantic oceanic gyres intensified.This was the time of initiation of the IWP and ACEP high-diversity foci. Arrows indicate principal currentdirections. Redrawn and simplified from Veron (1995, figs 28-30).

160 J. A. CRAME & B. R. ROSEN
Extrinsic mechanisms of diversification
Closure of the Tethyan Ocean in the Middle East.At the end of the Cretaceous period a vast,circum-equatorial tropical ocean (Tethys) couldbe traced westwards from the Indonesianregion, through the Middle East and southernEurope, to the proto-Atlantic Ocean (Fig. 3).There is thought to have been a predominantlywestward current flow in the ocean and this con-tinued through the Central American Seaway tothe eastern Pacific Ocean (Panthalassa).Although there are some signs of faunal differ-entiation within the Late Cretaceous TethyanOcean, this does not appear to have been such asto create high-diversity foci in any way equival-ent to those seen at the present day (Kauffman1973; Hallam 1994).
The same picture is essentially true for theearly part of the Cenozoic Era (i.e. the Paleo-gene). There was one very large, homogeneoustropical marine realm, with only minor evidenceof physical differentiation in areas such as theCaribbean, Middle East and deep-water easternPacific (Newell 1971; Kay 1984; Rosen 1988)(Fig. 3). A major pulse in coral reef developmentoccurred at the end of the Oligocene epoch, buteven then there was marked similarity, at thegeneric level, between western Tethys (i.e.Mediterranean) and Caribbean/Gulf of Mexicocorals (Frost 1981; Veron 1995). It was not in factuntil the Early Miocene (c. 20 Ma BP) that theTethyan Ocean was closed in its centralregion by the northward movement of theAfrica/Arabia landmass. This dramaticallycurtailed the westward-flowing tropical currentand led, eventually, to the Mediterranean regionbeing excluded from the coral reef belt (Fig. 3).At the same time the eastern Atlantic becamesignificantly cooler and this accentuated thephysical isolation of the Caribbean-easternPacific region (Rosen 1988; Veron 1995). It is tothis time that we can trace the origins of thedistinct IWP and ACEP foci of high tropicalmarine diversity.
The Early Miocene fossil record suggests thatthere was a reduced coral fauna in theCaribbean region that was transitional in com-positional terms between a Late Oligocenecosmopolitan one and a later Miocene one con-taining a number of endemics (Veron 1995). Itwould appear that the overall trend through theNeogene (i.e. the last 23 Ma) was one of relativeimpoverishment of ACEP coral reef faunas andrelative enrichment of IWP ones. For example,whereas there are some 24 coral genera in theACEP region at the present day, there are 87 inthe IWP; a similar comparison at the species
level gives a ratio of approximately 62: 450(Rosen 1988; Veron 1995). Briggs (1995) esti-mated that the vast IWP region contains morethan 6000 species of molluscs, 800 echinoderms,500 hermatypic corals (probably an overesti-mate; Veron 1995) and 4000 species of fishes.This equates to a total species richness that isapproximately 2.5 times that of the westernAtlantic, 3.5 times that of the eastern Pacific, and7.3 times that of the eastern Atlantic (Briggs1996). Ellison et al (1999) estimated that Thespecies richness of trees, shrubs and ferns inmangrove forests is an order of magnitudehigher in the IWP than the ACEP.
Collision of Australia-New Guinea with SEAsia. At the beginning of the Cenozoic Era theAustralia-New Guinea block was separatedfrom mainland SE Asia by a deep-water Indo-Pacific gateway measuring some 3000 km across.This feature was progressively reduced in widththrough the Paleogene (i.e. 65-23 Ma BP),chiefly by the northward subduction of Indian-Australian lithosphere beneath the Sunda-Java-Sulawesi arcs (Hall 1998). By the mid-Oligocene (30 Ma BP) the gap had narrowedsubstantially but there was still a clear deep-water passage floored by oceanic crust of theIndian and Pacific plates (Fig. 4). Major changesin plate boundaries occurred at approximately25 Ma BP when the New Guinea passivemargin collided with the leading edge of theeast Philippines-Halmahera-New Guinea arcsystem and the Australian margin, in the Bird'sHead region, was very close to collision with theEurasian margin in West Sulawesi (Hall 1998).By 20 Ma BP the continent-arc collision hadclosed the deep-water passage between thePacific and Indian oceans, resulting in a majorreorganization of tropical oceanic currentsystems (Kennett et al 1985; Grigg 1988). Con-tinued northward movement of Australia from20 to 10 Ma BP caused the rotation of severalplate boundaries and the formation of tectonicprovinces that are recognizable at the presentday (Fig. 4).
In comparison with both the Mediterraneanand Caribbean regions. Eocene and LowerOligocene hermatypic coral faunas appear tohave been genuinely sparse in the central IWP.However, this situation was dramaticallyreversed in the Early Miocene when there was afour-fold increase in the number of coral generain the IWP focus (Rosen 1988; Wilson & Rosen1998). The Early-Middle Miocene was a time ofwidespread deposition of coral-dominatedcarbonates in northern Australia, New Guineaand throughout SE Asia; reefs proliferated and
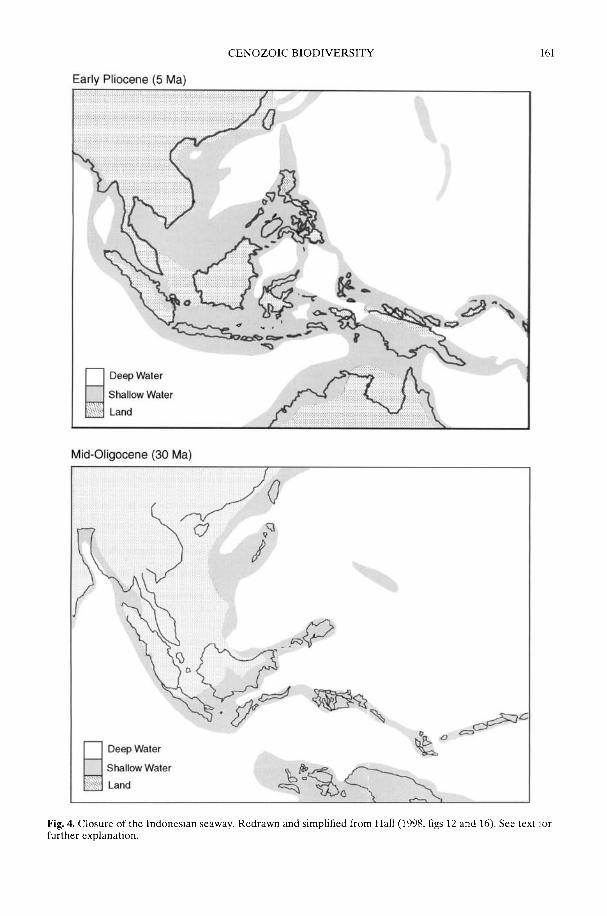
CENOZOIC BIODIVERSITY 161
Fig. 4. Closure of the Indonesian seaway. Redrawn and simplified from Hall (1998, figs 12 and 16). See text forfurther explanation.

162 J. A. CRAME & B. R. ROSEN
many modern genera and species evolved(Veron 1995; Crame 2001, and referencestherein). At the present day some 55% (by area)of the world's coral reefs occur in the SEAsia-New Guinea-Australia region (Wilson &Rosen 1998).
There can be no doubt that this dramatic risein the numbers of both reef and reef-associatedtaxa in the IWP region through the Neogene waslinked to a huge increase in the availability oftropical shallow-water habitats. Besides thenorthward movement of Australia and NewGuinea into the coral reef belt, collision-relateduplift led to the provision of more islands andcarbonate shelves in SE Asia (Fig. 4). This inturn must have greatly increased the amount oflocal habitat heterogeneity and the potential forallopatric speciation between fragmentedshallow-water areas. As a result of the syn-chronous closure of Tethys in the Middle Eastand progressive westward movement of thePacific archipelagos associated with the DarwinRise/Superswell, SE Asia also became a 'cross-roads* for tropical shallow-marine organisms(Rosen 1988; Pandolfi 1992).
Finally, it should be emphasized that theAustralian-New Guinea block continued tomove northwards throughout the Neogene, Ithas been suggested that at approximately 4 MaBP (Early Pliocene) it reached a critical pointwhen it came into close contact with the rapidlygrowing island of Halmahera. This had the effectof deflecting warm south Pacific waters east-wards at the Halmahera eddy to form theNorthern Equatorial Countercurrent (Cane &Molnar 2001). Thus warm waters in the Indo-nesian throughflow were replaced by relativelycold ones from the north Pacific, leading to adrop in sea surface temperatures in the IndianOcean and the aridification of East Africa.These changes were the catalyst for a shift in therelative heat balance between the east and westPacific, which in turn may have helped triggerthe onset of northern hemisphere glaciation(Cane & Molnar 2001).
Neogene climate change andbiodiversification
Following the lines of evidence presented aboveit could be maintained that Cenozoic palaeo-geographic changes were very largely respons-ible for the evolution of some of the majorpatterns of life on Earth. A once-homogeneoustropical biota was disrupted by vicariant eventssuch as the closure of Tethys in the Middle East,the collision of Australia-New Guinea with SE
Asia, and the rise of the CAI, The net effect, inthe marine realm, was to isolate an ACEP centreof high tropical diversity from an IWP one. Evenin the terrestrial realm, which is complicated tosome extent by pockets of high diversity in bothcentral and southern Africa, the effects were toproduce not dissimilar Palaeotropical andNeotropical realms (e.g. Barthlott etal. 1997). Inboth the marine and terrestrial realms thesteepest latitudinal gradients at the present dayare associated with the western margins of theNeotropics/ACEP and eastern margins ofthe Palaeotropics/IWP, respectively (Crame2000a,b).
Perhaps the marked heterogeneity observedin the tropical biota at the present day can beattributed simply to the range in sizes of thevarious subregions imposed by Cenozoictectonics. For example, in the shallow marinerealm the Indo-West Pacific province is approxi-mately four times the area of the WesternAtlantic and Eastern Pacific provinces com-bined (Briggs 1996). If we assume that thegreater species richness of the tropics is a time-invariant feature, caused perhaps by the greatersize of the tropics in comparison with all otherbiomes (e.g. Rosenzweig 1995), or some form ofspecies-energy hypothesis (e.g. Wright et al.1993), then what we see at the present day maybe due as much to tectonic as to biologicalfactors.
Nevertheless, important as these processesundoubtedly are to the generation and mainten-ance of large-scale diversity patterns, there is adistinct impression that something else musthave been involved too. As our knowledge ofthe tropical fossil record slowly improves it isbecoming apparent that much of the very pro-nounced tropical Cenozoic diversification eventactually occurred in the mid- to late Neogene(i.e. last 10-15 Ma) In the marine realm this iscertainly the case for zooxanthellate corals(Veron 1995; Wilson & Rosen 1998), as well ascertain reef-associated molluscan taxa (Crame2001, appendix 2). There is also some palaeon-tological evidence to demonstrate that certainmajor eudicot angiosperm clades are of essen-tially Neogene origin (Magallon et al 1999).
Some of this Neogene rise could well beattributed to differentiation diversity, with com-munities and provinces being distinguished asmuch in a longitudinal sense as a latitudinal one.Nevertheless, there has long been a suspicionthat global climate change was an importantdriver of diversification too, for this was a timeof marked intensification of Milankovitchcyclicity (Bennett 1997). These cycles, which arebased on the obliquity of the Earth's axis and

CENOZOIC BIODIVERSITY 163
eccentricity of orbit, have periods of 41 ka and100 ka, respectively; they are complemented bythe annual timing of the minimum Earth-Sundistance which varies with a 21 ka period. Theseorbital oscillations lead to variations in insola-tion and thus ambient temperatures. Such tem-perature variations would have been greatertowards the poles, but led to eustatic sea-levelchanges that were global in effect (Bennett 1997;Cronin & Raymo 1997).
In a series of previous papers, Rosen (1981,1984,1988) has already suggested how an inten-sification of glacioeustatic cyclicity may havepromoted diversification within the IWP focus.In essence, a type of species diversity pump wasenvisaged whereby taxa created in the outerislands of the western Pacific and eastern Indianoceans during sea level lowstands were sub-sequently 'pumped' into the central Indonesianfocus during highstands. In time, the latterregion became a form of refugium for numeroussympatric species. A similar mechanism ofspecies production by repeated cycles of allo-patry and secondary sympatry over at least a10 Ma period has recently been suggested for thetemperate zone floras of eastern Asia (Qian &Ricklefs 2000). Such floras have twice as manyspecies as their counterparts in eastern NorthAmerica, and it is thought that this is the productof repeated climatic and sea-level changes overa geographically much more heterogeneousterrain (Qian & Ricklefs 2000).
We are also beginning to appreciate morefully the effect of Milankovitch oscillations onspecies ranges; such orbitally forced rangedynamics (ORD) may well be the basis of arange of macroecological phenomena (Gaston& Blackburn 2000). In a wide-ranging review,Dynesius & Jansson (2000) demonstrated thatORD tended to be larger and more pronouncedin high- than low-latitude regions. Although thismight be taken as a strong indication that theformation of ecological isolates, a necessary firststep in the process of allopatric speciation,would be more likely in the high latitudes, theseauthors have argued that this is not in fact thecase. Instead, they contend that the short, stableperiods between high-latitude oscillations werenot long enough for the process of gradualspeciation to be completed; isolates were eitherbrought back together again or they becameextinct. Such a process is essentially reversed inthe tropics, where much smaller ORD promotesthe formation of isolates and drives the processof allopatric speciation (Dynesius & Jansson2000).
In an alternative model, Chown & Gaston(2000) have pointed out that many tropical taxa
are stenotopic and can, in theory, range largedistances within a more or less constant temper-ature belt defined by 25°N and 25°S. High-latitude taxa can achieve large ranges too, butthese tend to be eurytopes buffered to withstanda considerable amount of climate change. Thusit is the tropical taxa that may well be the mostvulnerable to temperature change; repeatedtemperature oscillations through the lateNeogene could have caused far more rangedisjunctions in the low- than high-latituderegions (Chown & Gaston 2000).
Although the precise mechanisms have yet tobe substantiated, it is likely that ORD played akey role in the Neogene diversification event(Bennett 1997). It should also be emphasizedthat their effects were not necessarily confinedto just tropical regions. Some high-latitude andpolar clades obviously radiated through theNeogene too (e.g. Crame & Clarke 1997), whereboth sea-level and temperature oscillationswould have been magnified in certain regions. Itis perhaps the superimposition of ORD on otherfactors, such as greater habitat heterogeneityand productivity, that makes their presence mostkeenly felt in the tropics.
In formulating his diversity pump model,Rosen (1984) supposed that much the same sortof process had operated in the ACEP focus asthe IWP one, though on a somewhat smallerscale. From a comprehensive stratigraphicalstudy of Neogene molluscs in the Caribbeanregion, we know that diversity did indeed risesteadily from approximately 14 to 5 Ma BP, butthen levelled off substantially (Jackson &Johnson 2000). As there is good evidence toshow that over the last 5 Ma extinction rateswere higher in this region than anywhere else inthe tropics, there must also have been anequivalent burst of originations (Allmon et al1996; Jackson et al 1999). For Caribbean corals,we know that diversification increased fromapproximately 16 to 4 Ma BP, but then from 4 to1 Ma BP a peak of originations preceded a peakof extinctions (Budd et al 1996; Allmon 2001).Between 9 and 1 Ma BP there were >120 coralspecies in the Caribbean, as compared with 62 atthe present day. Patterns of faunal change in theWestern Atlantic Neogene are obviouslycomplex and so too must be the environmentalchanges that underpin them. Nevertheless it islikely that origination and extinction patterns inthis region were strongly affected by the pro-gressive closure of the Central AmericanSeaway and rise of the CAI (see above).Although deep-water circulation through theseaway was blocked at 3.6 Ma BP, and shallow-water circulation at 3.0 Ma BP, the slowly rising

164 J. A. CRAME & B. R. ROSEN
CAI may have had an effect since 15-11 Ma BP(Cronin & Dowsett 1996).
Even though temperature changes did occurin the Western Atlantic, they were almost cer-tainly of secondary importance in the formationof diversity patterns. Instead it is likely that themajor reorganization of ocean currents broughtabout by the full emplacement of the CAI led toa reduction in the areas of upwelling and thusprimary productivity (Allmon 2001), At first adecline in productivity may have actually pro-moted the formation of isolates, and thus localspeciation, but in time it would have led to amarked decrease in the rate of isolate persis-tence, decreased origination and increasedextinction (Allmon 2001). The striking turnoverin East Pacific coral faunas since the Plioceneclosure of the CAI can be related to changes intemperature, salinity and nutrient levels (Buddet al 1996; Budd & Johnson 1999). Today thereis only a very depauperate coral fauna in thisregion but a surprisingly rich molluscan one.
DiscussionTaxonomic diversity gradients can be tracedback in the fossil record to the Late PalaeozoicEra, and perhaps even farther than that (Crame2001, and references therein). Despite the factthat it is often difficult to make meaningful com-parisons between these gradients, there is adistinct impression that they were not nearly sosteep as those seen at the present day. Dramaticsteepening appears to have occurred throughthe Cenozoic, and we now suspect that this wasespecially so during the Neogene (i.e. the last23 Ma), Indeed it is unlikely that, in the marinerealm, the present-day IWP and ACEP foci ofhigh taxonomic diversity are any more than20 Ma old. This was the time of crucial platetectonic movements that essentially gave thetropics their modern form.
It is also striking how the imposition of steepdiversity gradients coincided with marked globalclimatic cooling, and tempting to postulate a linkbetween the two. Whilst some form of large-scale differentiation diversity undoubtedlycontributed to the increase in global biodiversityat this time, it is not easy to see how it could havespecifically promoted radiations in two or moretropical foci. There may have been some form oflongitudinal as well as latitudinal biotic differen-tiation, but it is unlikely that this would havebeen on a scale sufficient to generate thefeatures seen at the present day.
Within the tropical marine realm, the richestPaleogene (i.e. 65-23 Ma BP) coral faunasoccurred in Europe and the Caribbean (Rosen
1988, table 2; Wilson & Rosen 1998, fig. 9). In theEarly Miocene, there is evidence to suggest thatat least 12 common IWP genera were formerlymore widespread (Veron 1995). Of particularinterest in this context is the fossil record of thediverse branching genus, Acropora. The oldestoccurrence is in the Paleocene of Somalia, andthere are also numerous European records fromthe Eocene-Miocene. However, Acropora doesnot occur in either the Caribbean or the IWPuntil the Oligocene (Wilson & Rosen 1998, andreferences therein), and a very marked radiationin the IWP only is dated as latest Miocene orPliocene (McCall et al. 1994). Similarly, fungiidcorals, which show their richest diversity at thepresent day in the IWP centre, can be tracedback to records of Cvcloseris in the Paleocene ofSomalia and western Pakistan, as well as otheroccurrences in the Eocene of SE Asia andEurope (Wilson & Rosen 1998).
Similar patterns to those of certain coralgenera are shown by mangroves. The fossilrecord strongly supports a Tethyan origin bothfor mangroves and their associated gastropodfauna in the Maastrichtian/Paleocene (Ellison etal. 1999). A pan-tropical distribution of manykey taxa had been achieved by the MiddleEocene, with, for example, Bruguiera beingrecorded from the London Clay. Range retrac-tions first occurred during a phase of globalcooling at the Eocene-Oligocene boundary anda cosmopolitan biota was then split by a series ofvicariant events. Subsequent in-situ speciationwithin both the IWP and ACEP foci occurredfrom the mid- to late Miocene onwards (Ellisonet al. 1999).
If such trends are repeated in other tropicaltaxa then it will be extremely difficult to classifytropical high-diversity foci as either evol-utionary 'cradles' or 'museums' (Briggs 1992;Jablonski 1993, and references therein); in asense they could be seen as both. A seminalstudy by McCoy & Heck (1976) first emphasizedthe importance of tectonic and climatic events inproducing disjunct tropical distributions in themarine realm and this has since been substanti-ated by a number of other workers (Rosen &Smith 1988; Pandolfi 1992; Veron 1995; Paulay1997). In essence, we would appear to be lookingat a Paleogene phase of range retractions anddisjunctions followed by a Neogene phase ofin-situ radiations from various foci.
It is important to determine whether the midto late Neogene tropical radiations represent anacceleration of evolutionary rates, and, if so,what caused them. Are the various tropicaldiversity patterns that we see at the present daysimply a function of province/region size, or is

CENOZOIC BIODIVERSITY 165
something else involved? It is again tempting tolink a phase of rapid diversification to one ofrapid climate change, and several mechanismshave been proposed whereby enhanced tropicalspeciation could have been caused by changes inORD. These all seem entirely plausible, buthave yet to be subjected to any form of rigorousexamination.
There are also counter-arguments available inthe literature to suggest that at least LatePliocene-Pleistocene climatic cycles weredetrimental to the process of speciation. Forexample, in the marine realm Potts (1984)presented a model whereby Plio-Pleistocenedisturbances (principally sea-level changes)were so frequent in the central IWP region thatevolutionary-scale processes such as speciationwould rarely be seen through to completion.This is, in effect, precisely the opposite of theprediction made by the Dynesius & Jansson(2000) isolate formation model. However, itwould seem likely that most living marine inver-tebrate taxa associated with coral reefs origi-nated well before the Pleistocene and we shouldperhaps be looking at origination events thatoccurred in the mid-Miocene to mid-Pliocenetime frame (i.e. 15-3 Ma BP) (Jackson & Johnson2000).
Evidence for a direct link between enhancedNeogene diversification and climatic changemay be even harder to find in the terrestrialrealm. Intensive stratigraphic study of NorthAmerican Cenozoic mammal faunas has shownvery little obvious correlation with short-termclimate change (e.g. Alroy 1998; Prothero 1999);for many prominent turnover events there is noobvious external trigger. Palynological evidencesuggests that the vast Amazonian lowlandforests were stable throughout much of theCenozoic (Colinvaux & de Oliveira 2001), andmolecular phylogenetic evidence to date doesnot point to any great Pleistocene radiation ofrainforest vertebrate faunas (Moritz et al 2000).Nevertheless, a recent synthesis of phylogeneticand geological data suggests that the species-richCape Flora of South Africa is no more than7-8 Ma old (Richardson et al 2001). In thisparticular instance speciation seems to havebeen driven by a combination of aridification ofa subtropical flora and imposition of regularclimate cycles.
Clearly these are complex issues that will notbe resolved either quickly or easily. However,what is becoming clearer is the major role playedby Cenozoic plate tectonics over the last15-20 Ma. Only when we know much moreabout complex plate interactions in a number ofkey areas, and how these affected parameters
such as oceanic circulation, climate change andorganism dispersal, will we be able to under-stand more fully the nature and origin of ourmajor extant biodiversity patterns.
We are grateful to M. J. Benton, A. Brandt, A. Clarke,A. Hallam and A. W. Owen for reading the manuscriptand suggesting various ways in which it could beimproved. Contribution to British Antarctic SurveyProject, Antarctic marine biodiversity: a historicalperspective'.
ReferencesADAMS, C. G., LEE, D. E. & ROSEN, B. R. 1990. Con-
flicting isotope and biotic evidence for tropicalsea-surface temperatures during the Tertiary.Palaeogeography, Palaeoclimatology, Palaeo-ecology, 77, 289-313.
ADRAIN, J. M,, WESTROP, S. R., CHATTERTON, B. D. E.& RAMSKOLD, L. 2000. Silurian trilobite alphadiversity and the end-Ordovician mass extinction.Paleobiology, 26, 625-646.
ALLMON, W. D. 2001. Nutrients, temperature, disturb-ance, and evolution: a model for the late Cenozoicmarine record of the western Atlantic. Palaeo-geography, Palaeoclimatology, Palaeoecology,166, 7-26.
ALLMON, W. D., ROSENBERG, G., PORTELL, R. W. &SCHINDLER, K. 1996. Diversity of Pliocene-Recent mollusks in the Western Atlantic: extinc-tion, origination, and environmental change. In:JACKSON, J. B. C,, BUDD, A. E & COATES, A. G.(eds) Evolution and Environment in TropicalAmerica. University of Chicago Press, Chicago.271-302.
ALROY, J. 1998. Long-term equilibrium in NorthAmerican mammalian diversity. In: McKINNEY,M. L. (ed.) Biodiversity Dynamics: Turnover ofPopulations, Taxa and Communities. ColumbiaUniversity Press, New York, 272-309.
ALROY, X, MARSHALL, C. R. & BAMBACH, R. K. 2001.Effects of sampling standardization on estimatesof Phanerozoic marine diversification. Proceed-ings of the National Academy of Sciences USA. 98.6261-6266.
BAMBACH, R, K. 1977. Species richness in marinebenthic habitats through the Phanerozoic. Paleo-biology, 3,152-167.
BAMBACH, R. K. 1990, Late Palaeozoic provinciality inthe marine realm. In: McKERROW, W. S. &SCOTESE, C. R. (eds) Palaeozoic Palaeogeographyand Biogeography, Geological Society, London,Memoirs, 12, 307-323.
BARKER, P. E, DALZIEL, I. W. D. & STOREY, B. C.1991. Tectonic development of the Scotia arcregion. In: TINGEY, R. J. (ed) The Geology ofAntarctica, Clarendon Press, Oxford, 215-248.
BARTHLOTT, W., BIEDINGER N., BRAUN, G., FEIG, F.KIER, G, LAUER, W. & MUTKE, J. 1997. Globalbiodiversity: species numbers of vascular plants.Map published on the internet at: http://www.botanik.uni-bonn.de/system/biomaps.htm

166 J. A. CRAME & B. R. ROSEN
BELLWOOD,D. R.& HUGHES, T. P. 2001. Regional-scaleassembly rules and biodiversity of coral reefs.Science, 292,1532-1534.
BENNETT, K. D. 1997. Evolution and Ecology: the Paceof Life. Cambridge University Press, Cambridge.
BENTON, M. J. 1999. The history of life: large data-bases in palaeontology. In: HARPER, D. A. T. (ed.)Numerical Palaeobiology, Wiley, Chichester,249-283.
BLACKBURN, T. M. & GASTON, K. J. 1996. A sidewayslook at patterns in species richness, or why thereare so few species outside the tropics. BiodiversityLetters, 3, 44-53.
BRIGGS. J. C. 1992. The marine East Indies: centreof origin? Global Ecology and BiogeographyLetters, 2,149-156.
BRIGGS, J. C. 1995. Global Biogeographv, Elsevier,Amsterdam.
BRIGGS, J. C. 1996. Tropical diversity and conservation.Conservation Biology, 10, 713-718.
BUDD, A. F. & JOHNSON. K. G. 1999. Originationpreceding extinction during late Cenozoic turnoverof Caribbean reefs. Paleobiology. 25,188-200.
BUDD. A. F., JOHNSON, K. G. & STENMANN, T. A. 1996.Plio-Pleistocene turnover and extinctions in theCaribbean reef-coral fauna. In: JACKSON, J. B. C.,BUDD, A. F. & COATES, A. G. (eds) Evolution andEnvironment in Tropical America. University ofChicago Press, Chicago, 168-204.
CANE, M. A. & MOLNAR, P. 2001. Closing of theIndonesian seaway as a precursor to east Africanaridification around 3-4 million years ago. Nature.411,157-162.
CHOWN, S. L. & GASTON. K. J. 2000. Areas, cradlesand museums: the latitudinal gradient in speciesrichness. Trends in Ecology & Evolution, 15,311-315.
CLARKE, A. & CRAME, J. A. 1992. The SouthernOcean benthic fauna and climate change: ahistorical perspective. Philosophical Transactionsof the Royal Society of London, Series B. 338,299-309. "
COATES, A. G, JACKSON, J. B. C., COLLINS, L. S., ET AL.1992. Closure of the Isthmus of Panama: Thenear-shore marine record of Costa Rica andPanama. Geological Society of America Bulletin,104, 814-828.
COLINVAUX, P. A. & DE OLIVEIRA, P. E. 2001.Amazon plant diversity and climate through theCenozoic, Palaeogeography, Palaeodimatology,Palaeoecology. 166, 51-63.
CRAME. J. A. 1999. An evolutionary perspective onmarine faunal connections between southern-most South America and Antarctica. ScientiaMarina. 63, 1-14.
CRAME, J. A, 2000a. Evolution of taxonomic diversitygradients in the marine realm: evidence from thecomposition of Recent bivalve faunas. Paleobiol-ogy. 26. 188-214.
CRAME, J. A. 2000£>. The nature and origin of taxo-nomic diversity gradients in marine bivalves. In:HARPER, E. M., TAYLOR, J. D. & CRAME, J. A.(eds) The Evolutionary Biology of the Bivalvia.Geological Society, London. Special Publications,177, 347-360.
CRAME, J. A. 2001. Taxonomic diversity gradientsthrough geological time. Diversity and Distri-butions, 7.175-189.
CRAME, J. A. & CLARKE. A. 1997. The historicalcomponent of marine taxonomic diversity gradi-ents. In: ORMOND, R. F. G, GAGE. J. D. & ANGEL.M. V. (eds) Marine Biodiversity: Patterns and Pro-cesses. Cambridge University Press, Cambridge.258-273.
CRONIN. T. M. & DOWSETT, H, J. 1996. Biotic andoceanographic response to Pliocene closing ofthe Central American Isthmus. In: JACKSON.J. B. C. BUDD, A. F. & COATES. A. G. (eds)Evolution and Environment in TropicalAmerica. University of Chicago Press, Chicago.76-104.
CRONIN.T M. & RAYMO. M. E. 1997. Orbital forcing ofdeep-sea benthic species diversity. Nature. 385.624-627.
DIESTER-HAAS. L. & ZAHN. R. 1996. Eocene-Oligocene transition in the Southern Ocean:History of water mass circulation and biologicalproductivity. Geologv, 24. 163-166.
DIETL. G. P., ALEXANDER, R. R. & BIEN, W. F. 2000.Escalation in Late Cretaceous-early Paleoceneoysters (Gryphaeidae) from the Atlantic CoastalPlain. Paleobiology, 26, 215-237.
DYNESIUS. M. & JANSSON. R. 2000. Evolutionary con-sequences of changes in species" geographicaldistributions driven by Milankovitch climateoscillations. Proceedings of the National Academyof Sciences USA. 97. 9115-9120.
ELLISON, A. M.. FARNSWORTH. E. J. & MERKT. R. E.1999. Origins of mangrove ecosystems and themangrove biodiversity anomaly. Global Ecologyand Biogeography, 8, 95-115.
FOOTE, M. 2000. Origination and extinction com-ponents of taxonomic diversity: general prob-lems. In: ERWIN. D. H. & WING.S.L. (eds) DeepTime: Paleobiology's Perspective. Paleobiology.Supplement. 26(4), 74-102.
FRAKES. L. A.. FRANCIS. J. E. & SYKTUS. J. I. 1992.Climate Modes of the Phanerozoic. CambridgeUniversity Press, Cambridge.
FRANCIS, A. P. & CURRIE, D. J. 1998. Global patterns oftree species richness in forests: another look.Oikos. 81, 598-602.
FROST, S. H. 1981. Oligocene reef coral biofacies of theVicentin, northeast Italy. In: TOOMEY. D. H. (ed.)European Fossil Reef Models. Society of Econ-omic Paleontologists and Mineralogists. SpecialPublications. 30. 483-539.
GASTON, K. J. 2000. Global patterns in biodiversity.Nature. 405, 220-227.
GASTON, K. J. & BLACKBURN. T. M. 2000. Pattern andProcess in Macroecology. Blackwell Science.Oxford,
GRIGG. R. W. 1988. Paleoceanography of coral reefs inthe Hawaiian-Emperor Chain. Science. 240.1737-1743.
HALL, R. 1998. The plate tectonics of Cenozoic SEAsia and the distribution of land and sea. In:HALL, R. & HOLLOWAY, J. D. (eds) Biogeographyand Geological Evolution of SE Asia. Backhuys.Leiden. 99-131.

CENOZOIC BIODIVERSITY 167
HALLAM, A. 1994. An Outline of Phanerozoic Bio-geography. Oxford University Press.
HANSEN, T. A., KELLEY, P. H., MELLAND, V. D. &GRAHAM, S. E. 1999. Effect of mass extinctionson escalation in molluscs. Geology, 27,1139-1142.
HAUG, G. H. & TIEDEMANN, R. 1998. Effect of theformation of the Isthmus of Panama on Atlanticthermohaline circulation. Nature, 393, 673-676.
HUSTON, M. A. 1999. Local processes and regionalpatterns: appropriate scales for understandingvariation in the diversity of plants and animals.Oikos, 86, 393-401.
JABLONSKI, D. 1993. The tropics as a source of noveltythrough geological time. Nature, 364, 142-144.
JABLONSKI, D. 2000. Micro- and macroevolution: scaleand hierarchy in evolutionary biology and paleo-biology. In: ERWIN, D. H. & WING, S. L. (eds)Deep Time: Paleobiology's Perspective Paleo-biology, Supplement, 26(4), 15-52.
JABLONSKI, D. & BOTTJER, D. J. 1990. The origin anddiversification of major groups: environmentalpatterns and macroevolutionary lags. In: TAYLOR,P. D. & LARWOOD, G. P. (eds) Major EvolutionaryRadiations. Systematics Association, SpecialVolume 42, Clarendon Press, Oxford, 17-57.
JABLONSKI, D., FLESSA, K. W. & VALENTINE, J. W. 1985.Biogeography and paleobiology. Paleobiology,11, 75-90.
JACKSON, J. B. C. & JOHNSON, K. G. 2000. Life in the lastfew million years. In: ERWIN, D. H. & WING, S. L.(eds) Deep Time: Paleobiology's Perspective.Paleobiology, Supplement, 26(4),221-235.
JACKSON, J. B. C.,TODD, J. A., FORTUNATO, H. & JUNG,P. 1999. Diversity and assemblages of NeogeneCaribbean Mollusca of lower Central America.In: COLLINS, L. S. & COATES, A. G. (eds) TheNeogene of the Isthmus of Panama: a PaleobioticSurvey of the Caribbean Coast. Bulletins ofAmerican Paleontology, 357,193-230.
KAUFFMAN, E. G. 1973. Cretaceous Bivalvia. In:HALLAM, A. (ed.) Atlas of Palaeobiogeography.Elsevier, Amsterdam, 353-383.
KAY, E. A. 1984. Patterns of speciation in the Indo-West Pacific. Special Publications of the BernicePauahi Bishop Museum, 72,15-31.
KENNETT. J. P. 1977. Cenozoic evolution of Antarcticglaciation, the circum-Antarctic ocean, and theirimpact on global paleoceanography. Journal ofGeophysical Research, 82, 3843-3860.
KENNETT, J. P., KELLER, G. & SRINIVASEN, M. S. 1985.Miocene planktonic foraminiferal biogeographyand paleoceanographic development of the Indo-Paciiic region. In: KENNETT, J. P. (ed.) TheMiocene Ocean: Paleoceanography and Biogeog-raphy. Geological Society of America, Memoirs,163,197-236.
LAW VER, L. A., GAHAGAN, L. M. & COFFIN, M. F.1992. The development of paleoseaways aroundAntarctica. In: KENNETT, J. P. & WARNKE, D. A.(eds) The Antarctic Paleoenvironment: A Perspec-tive on Global Change. Antarctic Research Series,56, 7-30.
LAZARUS, D. & CAULET, J. P. 1993. Cenozoic SouthernOcean reconstructions from sedimentologic, radi-olarian and other microfossil data. In: KENNETT, J.
P. & WARNKE, D. A. (eds) The Antarctic Paleoen-vironment: A Perspective on Global Change. PartTwo. Antarctic Research Series, 60,145-174.
LOUTIT, T. S., KENNETT, J. P. & SAVIN, S. M. 1983.Miocene equatorial and Southwest Pacificpaleoceanography from stable isotope evidence.Marine Micropalaeontology, 8, 215-233.
McCALL, J., ROSEN, B. R. & DARRELL, J. G. 1994.Carbonate deposition in accretionary prismsettings: Early Miocene coral limestones andcorals of the Makran mountain range in southernIran. Fades, 31,141-178,
McCoY, E. D. & HECK, K. L. 1976. Biogeography ofcorals, seagrasses, and mangroves: an alternativeto the center of origin concept. SystematicZoology, 25, 201-210.
MAGALLON, S., CRANE, P. R. & HERENDEEN, P. S.1999. Phylogenetic pattern, diversity, and diversi-fication of eudicots. Annals of the MissouriBotanic Garden, 86, 297-372.
MAGURRAN, A. E. 1988. Ecological Diversity and itsMeasurement. Croom Helm, London.
MARINCOVICH, L., JR. 2000. Central American paleo-geography controlled Pliocene Arctic Oceanmolluscan migrations. Geology, 28, 551-554.
MILLER, A. I. 2000. Conversations about Phanero-zoic global diversity. In: ERWIN, D. H. & WING, S.L. (eds) Deep Time: Paleobiology's Perspective.Paleobiology, Supplement, 26(4), 53-73.
MORITZ, C, PATTON, J. L., SCHNEIDER, C. J. &SMITH, T. B. 2000. Diversification of rainforestfaunas: An integrated molecular approach.Annual Reviews of Ecology and Svstematics, 31,533-563.
NEWELL, N. D. 1971. An outline history of tropicalorganic reefs. American Museum Novitates, 2465,1-37.
NlKLAS, K. J., TlFFNEY, B. H. & KNOLL, A. H. 1985.
Patterns in vascular land diversification: Ananalysis at the species level. In: VALENTINE, J. W.(ed.) Phanerozoic Diversity Patterns: Profiles inMacroevolution. Princeton University Press,Princeton, NJ, 97-128.
PANDOLFI, J. M. 1992. Successive isolation rather thanevolutionary centres for the origination of Indo-Pacific reef corals. Journal of Biogeography, 19,593-609.
PAULAY, G. 1997. Diversity and distribution of reeforganisms. In: BIRKLAND, C. E. (ed.) Life andDeath of Coral Reefs. Chapman and Hall,London, 298-352.
POTTS, D. C. 1984. Generation times and the Quater-nary evolution of reef-building corals. Paleobiol-ogy, 10, 48-58.
PROTHERO, D. R. 1999. Does climatic change drivemammalian evolution? GSA Today, 9,1-7.
QIAN, H. & RICKLEFS, R. E. 2000. Large-scale pro-cesses and the Asian bias in species diversity oftemperate plants. Nature, 407,180-182.
RAYMO, R. E. & RUDDIMAN, W. F. 1992. Tectonic-forcing of late Cenozoic climate. Nature, 359.117-122.
RICHARDSON, J. E., WELTZ, F. M., FAY, M. F, CRONK,Q. C .B., LINDER, H. P., REEVES, G. & CHASE,M. W. 2001. Rapid and recent origin of species

168 J. A. CRAME & B. R. ROSEN
richness in the Cape flora of South Africa. Nature,412,181-183.
RICKLEFS, R. E. 1999. Global patterns of tree speciesrichness in moist forests: distinguishing ecologicalinfluences and historical contingency. Oikos. 86.369-373.
RICKLEFS, R. E. & SCHLUTER, D. 1993. Species diver-sity: regional and historical influences. In: RICK-LEFS, R. E. & SCHLUTER, D. (eds) Species Diversityin Ecological Communities: Historical and Geo-graphical Perspectives. University of ChicagoPress, Chicago, 350-363.
ROSEN, B. R. 1981. The tropical high diversity enigma- the corals'-eye view. In: FOREY, P. L. (ed.) TheEvolving Biosphere, British Museum (NaturalHistory), London, and Cambridge UniversityPress, Cambridge, 103-129.
ROSEN, B. R. 1984. Reef coral biogeography andclimate through the late Cainozoic: just islands inthe sun or a critical pattern of islands? In:BRENCHLEY, P. J. (ed.) Fossils and Climate.Wiley, Chichester, 210-262.
ROSEN, B. R. 1988. Progress, problems and patterns inthe biogeography of reef corals and other tropicalmarine organisms. Helgolander Meeresunter-suchungen, 42, 269-301.
ROSEN, B. R. & SMITH, A. B. 1988. Tectonics fromfossils? Analysis of reef-coral and sea-urchindistributions from the late Cretaceous to Recent,using a new method. In: HALLAM, A. & AUDLEY-CHARLES, M. G. (eds) Gondwana and Tethys.Geological Society, London, Special Publications,37, 275-306.
ROSENZWEIG, M. L. 1995. Species Diversity in Spaceand Time. Cambridge University Press, Cam-bridge.
SEPKOSKI, J. J. JR. 1988. Alpha, beta, or gamma:where does all the diversity go? Paleobiology, 14.221-234.
SHAFFER, G. & BENDTSEN, J. 1994. Role of the BeringStrait in controlling North Atlantic ocean circu-lation and climate. Nature, 367, 354-357.
SIGNOR. P. W. 1990. The geologic history of diversity.Annual Reviews of Ecologv and Svstematics, 21,509-539.
STANLEY, S. M. 1995. New horizons for paleontology,with two examples: the rise and fall of theCretaceous Supertethys and the cause of themodern ice age. Journal of Paleontology, 69,999-1007.
STOTT, L. D., KENNETT, J. P., SHACKLETON, N. J. &CORFIELD, R. M. 1990. The evolution of Antarcticsurface waters during the Paleogene: Inferences
from the stable isotopic composition of plank-tonic foraminifers, ODP Leg 113. Proceedings ofthe Ocean Drilling Program Scientific Results. 113.849-863.
TURNER. J. R. G, LENNOX J. J. & GREENWOOD. J. J. D.1996. Does climate cause the global biodiversitygradient? In: HOCHBERG, M. E.. CLOBERT. J. &BARBAULT. B. (eds) Aspects of the Genesis andMaintenance of Biological Diversity. Oxford Uni-versity Press. Oxford. 199-220.
VALENTINE. J. W., FOIN. T. C. & PEART. D. 1978. Aprovincial model of Phanerozoic marine diversity.Paleobiology, 4, 55-66.
VERMEIJ, G. J. 1977. The Mesozoic marine revolution:evidence from snails, predators grazers. Paleobi-ology. 3. 245-258.
VERMEIJ, G. J. 1987. Evolution and Escalation. AnEcological History of Life. Princeton UniversityPress. Princeton. NJ.
VERMEIJ , G. J. 1994. The evolutionary interactionamong species. Selection, escalation, and co-evolution. Annual Reviews of Ecology andSystematics. 25, 219-236.
VERON. J. E. N. 1995. Corals in Space and Time. TheBiogeography and Evolution of the Scleractinia.Comstock/Cornell. London.
WILLIAMS. P. H.. GASTOX K. J. & HUMPHRIES. C. J.1997. Mapping biodiversity values worldwide:combining higher-taxon richness from differentgroups. Proceedings of the Royal Society ofLondon. B264. 141-148!
WILSON, M. E. J. & ROSEN. B. R. 1998. Implications ofpaucity of corals in the Paleogene of SE Asia:plate tectonics or Centre of Origin ? In: HALL. R.& HOLLOWAY. J. D. (eds) Biogeography and Geo-logical Evolution of SE Asia,Backhuys. Leiden.165-195.
WOODBI-RNE. M. O. & CASE. J. A. 1996. Dispersal, vic-ariance, and Late Cretaceous to Early Tertiaryland mammal biogeography from South Americato Australia. Journal of Mammalian Evolution. 3.121-161.
WRIGHT, D. H.. CURRIE. D. J. & MAURER, B. A. 1993.Energy supply and patterns of species richness onlocal and regional scale. In: RICKLEFS. R. E. &SCHLUTER. D. (eds) Species Diversity in Eco-logical Communities: Historical and GeographicalPerspectives. University of Chicago Press.Chicago, 66-74.
ZACHOS, 1. PAGANL M.. SLOAN. L.. THOMAS, E. &BILLUPS. K. 2001. Trends, rhythms, and aberra-tions in global climate 65 Ma to present. Science.292, 686-693.

Palaeontological databases for palaeobiogeography, palaeoecologyand biodiversity: a question of scale
PAUL J. MARKWICK1 & RICHARD LUPIA2
1 Robertson Research International Limited. Llandudno, Conwy, LL30 ISA, UK(e-mail: [email protected]
2 Sam Noble Oklahoma Museum of Natural History and School of Geology andGeophysics, University of Oklahoma, 2401 Chautauqua Avenue, Norman
OK 73072, USA
Abstract: Computerized databases provide an essential tool for investigating large-scalespatial and temporal palaeontological problems. Although advances in both software andhardware have made the logistics of building a database much easier, fundamental prob-lems remain concerning the representation and qualification of the data. Data from thefossil record are highly heterogeneous. Databases must be designed to account for varia-tions in scale (grain, resolution), inconsistency in the data, and potential errors (inaccu-racy). These issues vary with the scope of the study (extent), the biological group, and thenature and scale-dependence of supplementary, non-biological datasets (e.g. climate andocean parameters). With the application of desktop geographic information systems (GIS)to global Earth systems science, and the ability to efficiently integrate and query large,diverse datasets, the need to ensure robust qualification of data, especially scale, hasbecome all the more essential. This chapter examines some of the issues involved, definesterminology and offers pragmatic solutions.
The fossil record is vast, despite its inherentincompleteness, and computerized databasesprovide the only practical means for investi-gating large-scale palaeobiological patterns andthe processes responsible (e.g. Sepkoski 1982;Raup & Sepkoski 1986; Boulter et al. 1991;Benton 1993; Damuth 1993; Labandeira &Sepkoski 1993; Alroy 1995; Krebs et al 1996;Markwick 1996; Lupia 1999; Lupia et al 1999;Alroy et al 2001). But a database is only as goodas the data it contains and the questions asked ofit, and palaeontological data are more complexthan most. Palaeontological databases must bedesigned to take account of heterogeneities inscale (grain, resolution), inconsistencies in thedata, and potential errors (accuracy). As morestudies examine the interplay of diverse datasets(e.g. climate, soil, biodiversity), qualifying theseinherent scaling differences becomes critical, asmixing of incongruent datasets may lead toerroneous results. This has become an importantissue in landscape ecology (see Levin 1992) fromwhich we draw examples, but the problems areexacerbated for palaeontologists by the additionof a temporal dimension in the data (see Kidwell& Behrensmeyer 1993). This paper reviewssome of these issues, examines the potentialconsequences of ignoring scale, and suggestspragmatic solutions that are applicable to thedesign and implementation of palaeontological
databases. For many these concepts will befamiliar, but in the absence of a manual forbuilding palaeontological databases, this isaimed at helping researchers just beginning toconstruct their own databases.
Databases and basic database structure forpalaeontological data
The earliest and most basic form of computerdatabase is the 'flat file database' in which dataare stored as a single set of records of the samekind (Fig.la). Conceptually this is similar to acard index system, and suffers from the sameweaknesses in that data of only one kind can bequeried, and each record must be edited indi-vidually (e.g. Sepkoski 1982). 'Relational data-bases' (Fig.lb) can be viewed as multiple 'flat filedatabases' or tables (relations) linked together(related), such that complex queries can bemade integrating varied and diverse data. Theadvantages of separating data in this way arethat they only need to be entered or updatedonce in one table, but can be utilized by manydifferent records in other tables of the database.
Geographic information systems (GIS)couple the power of relational databases withthe visual efficiency of geographic maps(Fig.lc). In GIS, a record can be represented by
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic—Cenozoic Radiations, Geological Society, London, Special Publications, 194,169-178.0305-8719/02/$l5.00 © The Geological Society of London 2002.
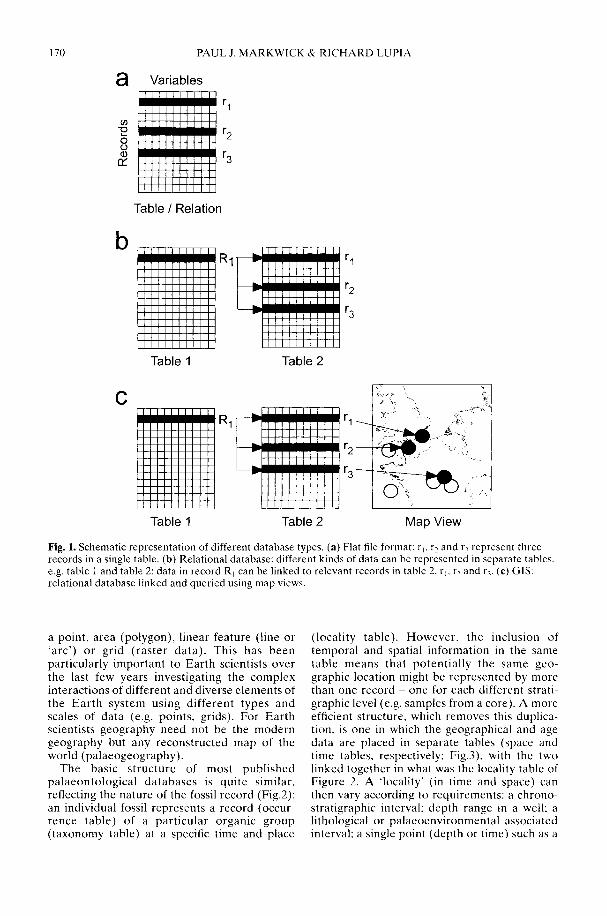
170 PAUL J. MARKWICK & RICHARD LUPIA
Fig. 1. Schematic representation of different database types. (a) Flat file format: r1, r2 and r3 represent threerecords in a single table, (b) Relational database: different kinds of data can be represented in separate tables.e.g. table 1 and table 2; data in record R1 can be linked to relevant records in table 2, r1, r2 and r3. (c) GIS:relational database linked and queried using map views.
a point, area (polygon), linear feature (line or'arc') or grid (raster data). This has beenparticularly important to Earth scientists overthe last few years investigating the complexinteractions of different and diverse elements ofthe Earth system using different types andscales of data (e.g. points, grids). For Earthscientists geography need not be the moderngeography but any reconstructed map of theworld (palaeogeography).
The basic structure of most publishedpalaeontological databases is quite similar,reflecting the nature of the fossil record (Fig.2):an individual fossil represents a record (occur-rence table) of a particular organic group(taxonomy table) at a specific time and place
(locality table). However, the inclusion oftemporal and spatial information in the sametable means that potentially the same geo-graphic location might be represented by morethan one record - one for each different strati-graphic level (e.g. samples from a core). A moreefficient structure, which removes this duplica-tion, is one in which the geographical and agedata are placed in separate tables (space andtime tables, respectively; Fig.3), with the twolinked together in what was the locality table ofFigure 2. A 'locality' (in time and space) canthen vary according to requirements: a chrono-stratigraphic interval; depth range in a well; alithological or palaeoenvironmental associatedinterval; a single point (depth or time) such as a
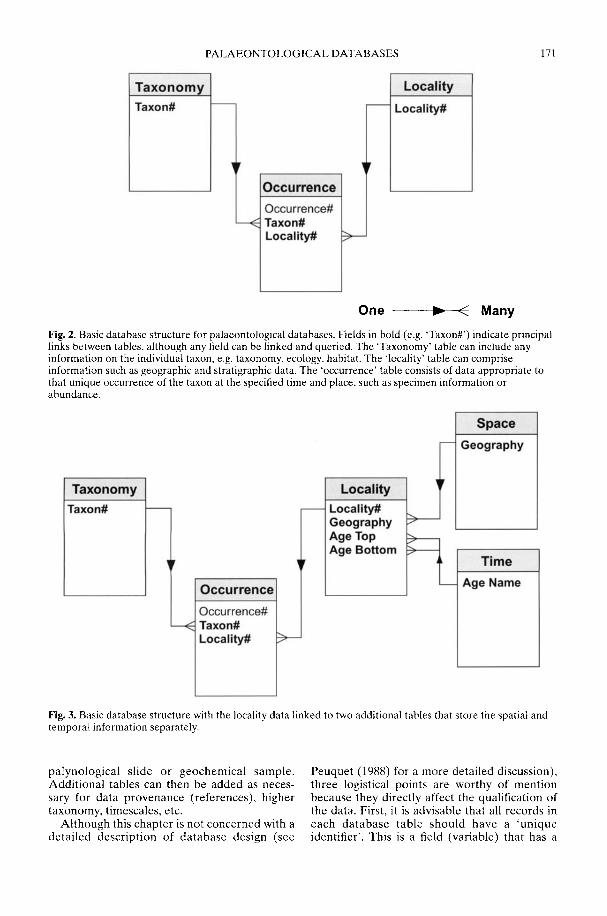
PALAEONTOLOGICAL DATABASES 171
Fig. 2. Basic database structure for palaeontological databases. Fields in bold (e.g. Taxon#') indicate principallinks between tables, although any field can be linked and queried. The 'Taxonomy' table can include anyinformation on the individual taxon, e.g. taxonomy, ecology, habitat. The 'locality' table can compriseinformation such as geographic and stratigraphic data. The 'occurrence' table consists of data appropriate tothat unique occurrence of the taxon at the specified time and place, such as specimen information orabundance.
Fig. 3. Basic database structure with the locality data linked to two additional tables that store the spatial andtemporal information separately.
palynological slide or geochemical sample.Additional tables can then be added as neces-sary for data provenance (references), highertaxonomy, timescales, etc.
Although this chapter is not concerned with adetailed description of database design (see
Peuquet (1988) for a more detailed discussion),three logistical points are worthy of mentionbecause they directly affect the qualification ofthe data. First, it is advisable that all records ineach database table should have a 'uniqueidentifier'. This is a field (variable) that has a

172 PAUL J. MARKWICK & RICHARD LUPIA
unique value for each record and should have noother meaning (i.e. should not include an agecode or taxon name that could potentiallychange in the future). These identifiers canthen be used to link tables (e.g. in Fig. 2 linkinglocality and taxon records to an individualoccurrence). Links can of course be made on anyother fields in a table, but care must be taken inknowing the relationships of the data (one-to-one, many-to-one, one-to-many)
The second logistical consideration is dataprovenance. In order that the data in the data-base can be used with confidence it is essentialto ensure that all data are referenced andaudited. The provenance of information iscritical to ensuring the integrity of the data, suchthat the issues of precision and error can betraced back to source. A distinction also shouldbe drawn between raw data (observations) thatare more or less immutable, and interpretationsbased on those data. If data are to be compiledfrom the published literature, it is also advisableto design the database to record data as it waswritten in the source or to record explicitlychanges made to the data (e.g. correction ofobvious misspellings or selected age assignmentamong disputed alternatives) as it is entered.For example, an author might misspell a taxon'sname and this error may be amended immedi-ately, but because some spelling variants aretruly different taxonomic entities (e.g. Cicatri-cosisporites, a trilete spore, and Cicatricoso-sporites, a monolete spore), any change shouldbe noted in a comment field in case examinationshould verify the 'error'. In the end, originaldata represent facts that can be accepted or dis-puted (and perhaps modified) by different usersof the database according to their scientificopinion. Making corrections or changes at thetime of entry without annotation precludesverification without returning to the originalpublication.
The final logistical point is the treatment oferror (inaccuracy). Errors in a database can beof three types: errors due to mistakes in dataentry; errors due to mistakes in the original data;and errors due to subsequent changes to thatdata (e.g. new phylogenetic hypotheses or agereassignments). In general, the first of these iseasily remedied by systematic checking of thedata. The second and third require that thedatabase be designed to be dynamic and allowupdates as necessary.
ScaleScale is a critical issue in ecology (Levin 1992)and palaeoecology (Kidwell & Behrensmeyer
1993), but frequently obfuscated by ambiguousterminology. In the ecological literature, scalerefers to the spatial and/or temporal dimensionsthat describe an object (e.g. 2 cm tooth or 4 haplot), event (e.g. 4 month rainy season) or obser-vation (e.g. 2 year study of a 4 ha plot) (O'Neill& King 1998). This has the opposite meaning toscale in the cartographic sense, which refers tothe level of detail; thus "large-scale" to an ecolo-gist refers to a large area or duration, but a'large-scale map' is usually of great detail butsmall area. This can lead to confusion whenusing GIS for examining ecology and palaeo-ecology. To combat this we have adopted twoterms from landscape ecology: grain, which isthe minimum resolution/scale of an observation(the smallest spatial or temporal interval ofobservation); and extent, which is the totalamount of space or time observed, usuallydefined as the maximum size of the study area(O'Neill & King 1998). Therefore, a large-scalemap' is fine-grained but of limited extent. Theimportant issue is to specify explicitly what thegrain and extent are for each study.
In studies of the fossil record, scale can betreated in the same manner. The grain of anobservation is equivalent to, for example, a rocksample, or locality, or basin (and the amountof time and space that they represent) and isdetermined by the size - thickness, area orvolume - measured. Which grain is useddepends on the questions asked of the data. Aglobal study (global extent) might only require asummary of the fossil fauna or flora for eachsedimentary basin in the world, and thereforethe grain is defined by the size of each basin.Conversely, a study of a specific basin (basinextent) might require a grain based on localities,or sites, or samples within that basin. The term'resolution' can be taken as a synonym of grain,thus time resolution' refers to the interval ofelapsed time represented by an assemblage (seeKidwell & Behrensmeyer 1993, table 1). "
Precision, strictly defined, is the ability torepeat a result, or the degree of consistencyamong several results, whereas accuracy is theability to achieve the real or true value. Here wemay loosen the definition of precision to refer tohow easily we could return to (literally revisit) asite given the information provided in thedatabase. To record that a site is located in'Yorkshire' may be accurate, that is, true, but itdoes not get us easily to the actual site at the baseof a specific cliff. Likewise, a site might really beof Eocene age, but this would not be helpful totrack down the actual site. Thus precision can beconstrued as uncertainty in the grain or extent ofa sample/analysis of the fossil record.

PALAEONTOLOGICAL DATABASES 173
Table 1. Geographic precision
Code
12345
Explanation
Precise location, within 1 km (equivalent to 'site/locality')Within 10 km (equivalent to 'nearest town')Within 100 km (equivalent to 'US county')Within 500 km (equivalent to 'US state')Very imprecise, not know to within 500 km (equivalent to 'country')
Grain (resolution)
Scaling issues are compounded in palaeoecologyby taphonomic (i.e. preservational) processesthat affect the apparent grain and extent ofanalyses through combining elements of assem-blages that did not co-occur in space (spatialaveraging; e.g. wind-blown pollen from outsidethe depositional area) or in time (time-averaging; e.g. reworking of shells from differentdepositional events), and by inaccuracies in thedata. In terms of a grain represented by'localities' these issues can be summarized intotwo principal questions: Where is the locality?How much 'space' and therefore time, isrepresented by the locality?
Accuracy. A fossil comes from a definitelocation, but it is not always possible to know thelocality with precision, either because the detailsare not reported in the literature, or the locationcould not be known at the time due to poor mapsor difficult terrain. The advent of globalpositioning systems (GPS) has mitigated manyof these problems, but in the older literature,localities were often described with respect toa local geographic feature, e.g. a town, riverconfluence, etc. By using GIS to plot detailedgeographic datasets (topographies, roads,rivers) at various scales, these localities usuallycan be placed in latitude-longitude space.Nonetheless, a simple qualifier can allow forimprecisely known localities to be distinguishedfrom well resolved sites, if that is important inanalyses (geographic precision (Markwick1996); Table 1). It needs to be remembered thatgiven plate motions (and the uncertaintytherein), absolute spatial resolution will deterio-rate the further back in geological time that theinterval under investigation occurred (Fig.4).
Locations can also be misplaced. This can bemitigated by checking locations against thecoordinates given in published gazetteers andatlases, but can be performed most effectivelyusing GIS. Again, detailed map datasets ofrivers, roads, political boundaries, topography,
outcrop geology, etc. can be superimposeddigitally in latitude-longitude (or x-y) spacewith the datapoints to be checked. This providesan immediate visual indication of error. Inten-tionally misplaced localities (for political orsite conservation reasons) can be dealt withsimilarly.
Age assignments can be made incorrectly,based on incorrect radiometric ages or fossilsparcity, or subject to change based on lateranalysis (different timescales). Ziegler et al(1985) tried to qualify confidence in age assign-ments by recording the provenance of the agedate (Table 2). Such a scheme may be refined bydistinguishing between different dating tech-niques within a particular category (e.g. Ar/Aror K/Ar age dating). By keeping the absoluteage data as a separate table, updates, andmultiple timescales can be accommodatedreadily.
Fig. 4. A representation of the uncertainty in spatialand temporal position of a locality at point (x,y) atthe present day (t()), with present uncertainty inspatial location Ax and Ay. The past position at timet1 is more uncertain both spatially ( x1, y1) andtemporally (At) due to uncertainties in the platereconstruction.

174 PAUL J. MARKWICK & RICHARD LUPIA
Table 2. Stratigraphic reliability codes (Paleogeographic Atlas Project, Chicago)
Code
ABCDEFG
Explanation
Complete biostratigraphic controlSome biostratigraphic controlStratigraphic interpolation ( = dating of rocks above or below)Geological inference ( = correlation with other site[s])Radiometric datingSecondary information ( = methods or source unspecified)Guesswork ( = no age provided, or dated to Period only)
Mixing and averaging. Behrensmeyer et al.(2001) provide an up-to-date summary of thefield and implications of taphonomic studiesfor palaeoecological interpretations. In short,taphonomic processes mix assemblages and theamount of space and/or time encompassed by asample is the spatial or temporal resolution ofthat sample. A single locality' may comprisemany taxa and vary spatially from a few centi-metres (such as a palynological preparation) toa few tens of centimetres or metres (e.g. a bed ofrock) to hundreds of kilometres (e.g. a for-mation within a basin). The larger the area orvolume of rock encompassed, the greater theamount of time that might be represented('analytical time averaging'); (Behrensmeyer &Hook 1992). However, biological and tapho-nomic processes specific to a particular group oforganisms reduce generality. A series of palyno-logical samples through a core, each very smalland representing depositional instants, implies atight temporal grain, but mixing and transport ofpollen in wind and water might imply coarsespatial grain for the same samples. Furthermore,the temporal duration of a single palynologicalpreparation from a well core may present adepositional instant if made parallel to bedding,or a few years or tens of years if made perpen-dicular to bedding. This will also be reflected inthe interpretation of the contemporary environ-ment, including climate.
Separate biological and taphonomic pro-cesses produce a distinctly different grainimplied by most vertebrate localities. Because ofthe relative sparsity of specimens in most cases,a vertebrate locality might include an area thatis on the order of kilometres, or even tens ofkilometres, in size, and which may encompass athickness of hundreds of metres of sediment. Assuch, it might represent hundreds (or thou-sands) of years of deposition, depending onthe tectonic setting (Behrensmeyer 1982;Behrensmeyer & Chapman 1993; Rogers 1993),but if the animals are migratory, it would be
necessary to obtain a sample that adequatelyreflects the local fauna.
The physical mixing of earlier faunas withincontemporary faunas ('taphonomic time averag-ing'; Behrensmeyer & Hook 1992; Behrens-meyer & Chapman 1993) further degradesresolution. The consequence of these problemsis that as a palaeontological event (such as anextinction or a response to climate change) orenvironmental interpretation is examined overbroader areas, so the temporal resolution withwhich it can be defined decreases. Conversely,the more finely events are resolved in time, themore difficult it is to know how large a region isaffected. This is referred to as the 'paleonto-logical uncertainty principle', analogous to the'uncertainty principle' in quantum physics(S. Wing, pers. comm. 1991).
One solution is to use only data of a specifiedgrain (resolution), but this can lead to loss ofdata, including information that, althoughpoorly resolved, is nonetheless important. Forexample, if the location of a fossil is given as'India' this may be considered spatially poorlyresolved and therefore ignored, but if it is theonly report of that fossil from India, then it is stilluseful information. However, this requires thatthe precision can be qualified; descriptors suchas 'sample', 'composite locality', 'quarry', 'site'can be used, but each of these terms has numer-ous definitions, and so must be defined for everydatabase. Landscape ecologists, faced with asimilar problem, have derived numerous(mostly hierarchical) classification schemes fordescribing different scales of landscape systembased on the areal extent considered (Huggett1995). A similar approach might be appropriatefor palaeontological databases.
An alternative solution is to include all data atthe resolution at which it is reported (using aqualifier), and then to coarsen all 'localities' tosome standard spatial or temporal scale byconcatenating faunal and floral lists, in order toeliminate local variability. This has been used by

PALAEONTOLOGICAL DATABASES 175
Markwick (2002) for present-day faunas andfloras in order to examine the relationship ofclimate, biogeography and diversity (see alsoMarkwick 1996). The selection of the smallestsampling unit determines the highest resolution(finest grain) possible in analysis based on infor-mation in the database. It is relatively easy tocoarsen the resolution of data at a later date. Itis impossible to refine it.
TaxonomyTaxonomy influences grain, because differentorganisms scale with the environment differ-ently, but this is a matter to consider whenanalysing the data. The major problem to bequalified in database design is taxonomic error(inaccuracy). Errors in taxonomic assignmentscan be due to several causes, among them thefollowing: (i) incomplete preservation (absenceof diagnostic characters); (ii) morphologicaluniformity (e.g. pollen of grasses); (iii) form taxa(e.g. separate genera for leaves, seed, pollen,etc.); and (iv) unreported taxonomy. Classifi-cation schemes for all biological entities aresubject to change and disagreement. This isparticularly true for fossil taxa, which may haveno extant representatives, and which might berepresented by incomplete and/or limitednumbers of specimens. Different workers mayadopt different taxonomic schemes dependingon their own experience and opinions, and therelevant literature may incorporate a longhistory of taxonomic changes. The solution ispartly an issue of accommodating uncertaintybecause assignments at a low taxonomic levelmay be poorly supported and disputed widely,whereas the higher level assignments can bemade with considerable confidence and generalagreement among professionals. Potentialerrors can be minimized by coarsening the datato a more 'confident' taxonomic level, and/or byrecording specimen information as a guide tothe characters used in the taxonomic assign-ment. This will vary according to the groupstudied, such that this method may createproblems when assemblages are compared (thequestion of which taxonomic level to use, andwhether the same level should be applied to allgroups in the analysis). A species assignmentbased on an isolated fossil tooth will probably beof low confidence for a lizard, but significant fora mammal.
Another potential solution is to adopt a 'stan-dard', preferably published, taxonomy and usethis throughout the database. This ensures thatthe higher level taxonomy is at least consistent,although consistency is no guarantee of truth.
Multiple standards can be made available asseparate relations in the database structure.
Synonymy
Synonymization is the method of transferring aspecimen or species to its appropriate taxonomicunit (e.g. species or genus) for any of severalreasons, but usually because it is identical to apreviously designated taxon. This can be dealtwith by adding a 'synonym table' to the databasestructure that is used as a look-up library for alltaxon names entered into the taxonomy table.The links can be structured such that if theentered taxon is found to belong within anotherspecies according to the synonym table, the mostrecent synonymized form replaces it. Again, theissue of data provenance is emphasized asspecies nomenclature is particularly fluid andcontentious.
The rules of biological nomenclature statethat no two animal or plant species may havethe same name, and the rules establish how todesignate and name a new species. Yet differentspecies are often encountered in the literaturethat have the same name given informallyduring a study. This is particularly so in palyn-ology and occurs primarily in the stratigraphicliterature where interest focuses on distinguish-ing rock units from one another by segregationof pollen types. The frequent expression of thisis the designation of many species named bycombining informally a genus name with 'sp. A'or 'sp.l', as in Agasie (1969) and Ravn (1995)who record 'Tricolpites sp. 1' from their sites inArizona and Wyoming. However, sharing thesame name does not imply that these pollentypes represent the same biological entity, whichis implied when formally named species sharethe specific epithet. Indeed, 'Tricolpites sp. 1' inthe paper by Agasie (1969) does not appearsimilar to 'Tricolpites sp. 1' of Ravn (1969). Thesimplest method to overcome this problem is totreat 'sp. 1' etc. of every author as a distinct taxo-nomic unit, distinguished by a unique name, forexample 'Tricolpites sp. 1 of Agasie (1969)'.
DiscussionWith the ready availability of desktop computer-ized relational database and GIS software, thelogistics of building databases to cope with thelarge volumes of palaeontological data is nolonger a major issue. While it is useful to remem-ber certain guidelines as to database structure(Fig. 2) and the physical amount of data to beincluded (a database should be 'simple enoughthat it can be used, but comprehensive enough

176 PAUL J. MARKWICK & RICHARD LUPIA
that it will be useful,'; Markwick 1996, p. 921), theprincipal problem facing designers of palaeonto-logical databases is how to accommodate andqualify heterogeneities within the record,specifically of scale. We have suggested here thatit is always better to collect information at thefinest grain (resolution) possible and to appendthe appropriate confidence estimate (as aqualifier that can be queried on), since higherresolution data can always be degraded to lowerresolution, but the reverse is impossible. Thequestion of how observations made at differentscales can be compared has been discussed bynumerous authors for both modern and fossilsettings (see Signor 1978; Hatfield 1985; Levin1992; Anderson & Marcus 1993; Brown 1995;Rosenzweig 1995). But it is important to under-stand why scale is so important, especially forresearchers integrating datasets from differentfields, which has been made so much easierthrough CIS.
We have already noted how the apparentgrain of a fossil assemblage can be affected byphysical mixing and averaging in time and space,and that this problem worsens as the extent ofthe study increases. Consequently, this problemis greatest for global studies. For example,Markwick (1998), using the global distributionof fossil crocodilians to reconstruct palaeo-climate, calculated that the probability that 100Eocene fossil crocodilian localities representedthe identical 30 year timespan within the Eocene(21 000 000 years) and therefore the same'climate', was 1/700 00099. The problem ofcorrelating age-equivalent samples is furtherexacerbated when multiple lines of evidence areused (e.g. palynology, floras and vertebrates toreconstruct palaeoclimate), each subject todifferent taphonomic processes. Failure torecognize the mixture of biological and environ-mental phenomena operating at different scalescan produce spurious and misleading results.Even within the same biological group, mixingdata of different resolutions can have strongeffects on derived interpretations, especially inquantitative analyses. Lupia et al. (1999)analysed palynological samples from NorthAmerica to investigate the possible replacementof conifers and free-sporing plants byangiosperms. They chose to restrict analyses toindividual palynological samples, from a singlesite and stratigraphic horizon, rather thanincluding samples created by combiningmultiple samples from several sites or strati-graphic horizons. Lupia et aL (1999) foundnearly constant within-flora diversity throughthe Cretaceous compared to previous resultsfrom Lidgard & Crane (1990) that showedincreasing within-flora diversity from Early to
Late Cretaceous. By examining Lidgard andCrane's (1990) dataset, Lupia et al. (1999)concluded that the difference was attributable tothe former's inclusion of combined samples,preferentially of Late Cretaceous age. in theiranalyses.
Likewise, the scale of biotic processesresponding to abiotic conditions combined withresolution may decrease methodological power.For example, published data on using thefoliar physiognomic method for reconstructingpalaeoclimate suggest that the method, whichseems to work well over large geographicgradients (Wolfe 1971,1993), may break down atsmaller scales probably due to the bias of localeffects (Dolph & Dilcher 1979). Such problemsare exacerbated when palaeontological data arecompared with global climate model results,which can be of coarse spatial resolution, on theorder of 4-5° of latitude and longitude(McGuffie & Henderson-Sellers 1997). Suchcoarseness may hide the finer scale variations inthe real contemporary climate system, as experi-enced by the fossil organisms (climate proxies)themselves (Markwick 1998). Precipitation, forexample, is very sensitive to local orography andmoisture sources, and has been found to vary by30% over a matter of a few kilometres (Linacre1992). This may be particularly important inareas of rapid relief changes, such as the Eoceneof the western United States (Sloan 1994).
The effect of error (inaccuracy) in databasesalso depends on the question being addressed.For North American Cambrian trilobites.Westrop & Adrain (2001) found that despite70% of the generic records in the Sepkoskigeneric database being inaccurate (compiledfrom the published literature), when comparedto their own field-based compilation, bothdatasets showed the same large-scale (coarsegrain) patterns in Phanerozoic biodiversity(Adrain & Westrop 2000; Westrop & Adrain2001). With finer grain, such errors becomemore important (Westrop & Adrain 2001).
The consequences of scale (grain) and errordepend on the fossil group or assemblage investi-gated, the extent of the study and the questions tobe asked. Palaeontological databases must there-fore be designed to accommodate these issues.
Conclusions
The fossil record is the only direct evidenceabout the biological evolution of life on Earth.This represents a huge volume of data, andcomputerized databases provide the mostefficient means of storing and examining therecords for large-scale patterns and processes.The quantity and quality of these data are

PALAEONTOLOGICAL DATABASES 177
always less than ideal. But, we suggest that it isthe heterogeneities in scale (grain) in the fossilrecord that are potentially the major cause ofproblems in palaeoecology especially whendifferent and varied datasets are mixed, asincreasingly they are in studies examining theinterplay of biotic and abiotic phenomena usingGIS. Scale can, and should, be qualified in adatabase, and to facilitate this we offer thefollowing suggestions.
(i) Data should be collected at the finest avail-able grain (resolution) since the finest scaleof data stored in the database defines thefinest grain possible in any analysis usingthat data. The grain in each case needs to berecorded (e.g. by a qualifier in a separatedata field). In this way the database can bequeried for data of a specified grain, or dataof mixed resolutions can be coarsened tosome common grain.
(ii) All data in the database should have anaudit trail, recording all sources andchanges made to that data. To this end a dis-tinction should be made between raw data(observations) and interpretation.
(iii) Database structure is dictated by the natureof the fossil record, such that the mostefficient structure is based around separatetables of spatial and temporal location, tax-onomy and occurrence, which can alsoinclude taphonomic information (Fig. 3).The database structure should also facili-tate the inclusion of multiple standards (e.g.alternative timescales and taxonomies).
We would like to thank A. Crame for the opportunityto publish this work, and the reviewers, S. Lidgard andR. Livermore, whose suggestions greatly improvedthis paper. This work draws on our experience ofdesigning and compiling palaeontological databases atThe University of Chicago, where we benefited fromthe advice of P. Crane, S. Lidgard, the late J. Sepkoski,and especially the members of the PalaeogeographicAtlas Project: A. Ziegler, D. Rowley, A. Lottes and M.Hulver. Finally, we wish to thank the libraries of theUniversity of Chicago, especially the staff of the JohnCrerar Science Library - at the heart of all great uni-versities is a great library.
ReferencesADRAIN, J. M. & WESTROP, S. R. 2000. An empirical
assessment of taxic paleobiology. Science, 289,110-112.
AGASIE, J. M. 1969. Late Cretaceous palynomorphsfrom northeastern Arizona. Micropaleontology,15,13-30.
ALROY, J. 1995. Quantitative mammalian biochronol-ogy, biogeography, and diversity history of NorthAmerica, PhD thesis. University of Chicago.
ALROY, J., MARSHALL, C. R., BAMBACH, R. K., ET AL..2001. Effects of sampling standardization on esti-mates of Phanerozoic marine diversification. Pro-ceedings of the National Academy of Sciences,U.S.A., 98/6261-6266.
ANDERSON, S. & MARCUS, L. F. 1993. Effect of quadratsize on measurement of species density. Journal ofBiogeography, 20, 421-428.
BEHRENSMEYER, A. K. 1982. Time resolution in fluvialvertebrate assemblages. Paleobiology, 8,211-227.
BEHRENSMEYER, A. K. & CHAPMAN, R. E. 1993.Models and simulations of time-averaging interrestrial vertebrate accumulations. In:KIDWELL, S. M. & BEHRENSMEYER, A. K. (eds)Taphonomic Approaches to Time Resolution inFossil Assemblages. University of Tennessee,Knoxville, Short Courses in Paleontology, 6,125-149.
BEHRENSMEYER, A. K. & HOOK, R. W. 1992. Paleo-environmental contexts and taphonomic modes.In: BEHRENSMEYER, A. K., DAMUTH, J. D.,DIMICHELE, W. A., POTTS, R., SUES, H.-D. &WING, S. L. (eds) Terrestrial Ecosystems ThroughTime. Evolutionary Paleoecology of TerrestrialPlants and Animals. University of Chicago Press,Chicago, 15-136.
BEHRENSMEYER, A. K., KIDWELL, S. M. & GASTALDO,R. A. 2001. Taphonomy and paleobiology. In:ERWIN, D. H. & WING, S. L. (eds) Deep Time:Paleobiologv's Perspectives. Paleobiology,Supplement, 26 (4), 103-147.
BENTON, M. J. (ed.) 1993. Fossil Record 2. Kluwer Aca-demic Publishers, Dordrecht.
BOULTER, M. C, CHALONER, W G. & HOLMES, P. L.1991. The IOP plant fossil record: are fossil plantsa special case? In: HAWKSWORTH, D. L. (ed.)Improving the Stability of Names: Needs andOptions. Koeltz Scientific Books, Konigstein,Regnum Vegetabile 123, 231-242.
BROWN, J. H. 1995. Macroecology. University ofChicago Press, Chicago.
DAMUTH, J. 1993. ETE Database Manual Evolution ofTerrestrial Ecosystems Consortium, Washington,DC.
DOLPH, G. E. & DILCHER, D. L. 1979. Foliar physion-omy as an aid in determining palaeoclimate.Palaeontographica B, 170,151-172.
HATFIELD, C. B. 1985. Fossil vertebrate taxonomicdiversity correlated with outcrop area. OhioJournal of Science, 85, 64-67.
HUGGETT, R. J. 1995. Geoecology. An EvolutionaryApproach. Routledge, London.
KIDWELL, S. M. & BEHRENSMEYER, A. K. (eds) 1993.Taphonomic Approaches to Time Resolution inFossil Assemblages. Univeristy of Tennessee,Knoxville, Short Courses in Paleontology, 6.
KREBS, J. W, KAESLER, R. L., BROSIUS, E. A., MILLER,D. L. & CHANG, Y.-M. 1996. Paleobank, a rela-tional database for invertebrate paleontology: thedata model. The University of Kansas Paleonto-logical Contributions New Series, 8,1-7.
LABANDEIRA, C. C. & SEPKOSKI, J. J., JR. 1993. Insectdiversity in the fossil record. Science, 261,310-315.
LEVIN, S. A. 1992. The problem of pattern and scalein ecology. Ecology, 73,1943-1967.

178 PAUL J. MARKWICK & RICHARD LUPIA
LIDGARD, S. & CRANE, P. R. 1990. Angiosperm diversi-fication and Cretaceous floristic trends: a com-parison of palynofloras and leaf macrofloras.Paleobiology. 16, 77-93.
LINACRE, E. 1992. Climate Data and Resources. AReference Guide. Routledge, London.
LUPIA, R. 1999. Discordant morphological disparityand taxonomic diversity during the Cretaceousangiosperm radiation: North American pollenrecord. Paleobiology, 25, 1-28.
LUPIA. R.. LIDGARD, S. & CRANE, P. R. 1999. Com-paring palynological abundance and diversity:implications for biotic replacement during theCretaceous angiosperm radiation. Paleobiology.25. 305-340.
McGUFFIE, K. & HENDERSON-SELLERS, A. 1997. AClimate Modelling Primer. Wiley, Chichester.
MARKWICK, P. J. 1996. Late Cretaceous to Pleistoceneclimates: nature of the transition from a 'hot-house' to an 'ice-house' world. PhD Thesis. TheUniversity of Chicago.
MARKWICK, P. J. 1998. Fossil crocodilians asindicators of Late Cretaceous and Cenozoic cli-mates: implications for using palaeontologicaldata for global change. Palaeogeography, Palaeo-climatology, Palaeoecology, 137, 205-271.
MARKWICK, P. J. 2002. Integrating the present andpast records of climate, biodiversity and biogeog-raphy: implications for palaeoecology and palaeo-climatology. In: CRAME, J. A. & OWEN, A. W. (eds)Palaeobiogeography and Biodiversity Change: theOrdovician and Mesozoic-Cenozoic Radiations.Geological society. London, Special Publications,194,179-199.
O'NEILL. R. V. & KING, A. W. 1998. Homage to St.Michael or why are there so many books on scale?In: PETERSON. D. L. & PARKER, V. T. (eds) Eco-logical Scale, Theory and Applications. ColumbiaUniversity Press, New York, 3-15.
PEUQUET, D. J. 1988. Issues involved in selecting appro-priate data models for global databases. In:MOUNSEY, H. (ed) Building Databases for GlobalScience. Taylor and Francis. London, 66-78.
RAUP, D. M. & SEPKOSKI. J. J. 1986. Periodicextinction of families and genera. Science. 231.833-836.
RAVN, R. L. 1995. Miospores from the Muddy Sand-stone (upper Albian). Wind River basin.Wyoming, USA. Palaeontographica, Abteilung B.234.41-91.
ROGERS, R. R. 1993. Systematic patterns of time-averaging in the terrestrial vertebrate record:a Cretaceous case study. In: KIDWELL. S. M.& BEHRF.NSMEYER, A. K. (eds) TaphonomicApproaches to Time Resolution in FossilAssemblages. University of Tennessee, Knoxville.Short Courses in Paleontology. 228-249.
ROSENWEIG, M. L. 1995. Species Diversity in Spaceand Time. Cambridge University Press.Cambridge.
SEPKOSKI, J. J. JR. 1982. A Compendium of FossilMarine Families. Milwaukee. Public Museum,Contributions in Biology and Geology. 51.
SIGNOR. P. W. III 1978. Species richness in thePhanerozoic: an investigation of sampling effects.Paleobiology, 4. 394-406.
SLOAN, L. C. 1994. Equable climates during the earlyEocene: significance of regional paleogeographyfor North American climate. Geology. 22.881-884.
WESTROP. S. R. & ADRAIN. J. M. 2001. Sampling at thespecies level: impact of spatial biases on diversitygradients. Geology. 29. 903-906.
WOLFE. J. A. 1971. Tertiary climatic fluctuations andmethods of analysis of Tertiary floras. Palaeo-geography, Palaeoclimatology, Palaeoecology. 9.27-57.
WOLFE, J. A. 1993. A method of obtaining climatic par-ameters from leaf assemblages. U.S. GeologicalSurvey Bulletin. 2040. 1-71.
ZIEGLER. A. M.. ROWLEY. D. B.. LOTTES, A. L..SAHAGIAN. D. L.. HULVER. M. L. & GIERLOWSKI.T. C. 1985. Paleogeographic interpretation: withan example from the Mid-Cretaceous. AnnualReview of Earth and Planetary Sciences. 13.385-425. '

Integrating the present and past records of climate, biodiversity andbiogeography: implications for palaeoecology and
palaeoclimatology
PAULJ.MARKWICKRobertson Research International Limited, Llandudno, Conwy, LL30 ISA, UK
(e-mail: [email protected])
Abstract: A geographic information system (GIS) based, integrated dataset of RecentNorth American, European, southern African and Australian non-avian tetrapod faunas isused to examine the macroscale relationship between climate, biogeography and terrestrialtaxonomic and functional species diversity (richness). The results support a modified formof the species-energy hypothesis, with the pattern of terrestrial biodiversity reflecting themanner in which species procure energy, rather than only the absolute amount of 'avail-able energy'. Area and history are also found to be important. Ectotherms show thesimplest relationship with environmental variables (and strongest latitudinal diversitygradients), and endotherms the most complex. A strong linear relationship is foundbetween the proportion of each fauna represented by ectotherms and temperature (meanannual temperature and coldest month mean temperature). This relationship is used in anexperiment to retrodict the palaeotemperature for the Middle Eocene lagerstatten faunafrom Messel, Germany. Results compare well with interpretations based on other climateproxies.
Climate interpretations based on fossil datadepend heavily on analogy with recent speciesand to this end an understanding of living groupsand their distribution is crucial. Terrestrialorganisms are neither randomly nor evenly dis-tributed on the globe, and natural historianshave long postulated that this is due, at least inpart, to climate and other environmental factors(Humboldt & Bonpland 1807; Wallace 1876;Matthew 1915; Darlington 1948). Some taxa,such as crocodilians, are demonstrably limitedby temperature and have been used by geolo-gists since the early nineteenth century as toolsfor reconstructing palaeoclimate (Lyell 1830;Colbert et al 1946; Hibbard 1960; Markwick1994, 1998a). A climate origin has also beenpostulated to explain observed 'latitudinal'species diversity (or richness) gradients in manyRecent terrestrial organic groups (Pianka 1966;Stevens 1989; Currie 1991; Rohde 1992; Wrightet al 1993). Ostrom (1970) has suggested thatsuch gradients might provide a better tool forretrodicting palaeoclimate than comparison ofan individual fossil with the climate of its livingrelative (see also Fischer 1961, p. 50).
Many theories have been proposed to accountfor observed latitudinal diversity gradients in theRecent (see Rohde (1992) for review), of whicharea-history and species-energy have perhapsreceived the most attention. Arguments basedon area (Rosenzweig 1995) derive from theobservation that island habitats tend to have
fewer species than non-isolated habitats of thesame area. This is embodied in the island bio-geography theory of MacArthur & Wilson(1967), in which standing diversity reflects thebalance between immigration and extinctionrates, as dictated by island area (smaller areashold smaller populations that are thereforeassumed to be more susceptible to extinction)and the proximity in time (history) and space tothe dispersal source. Historical changes in insu-larity, for example the repeated fragmentationand coalescence of 'islands' during Pleistoceneinterglacial-glacial changes, are envisaged toincrease the opportunity for allopatric specia-tion, and have been used to account for the highspecies diversities in SE Asia (Qian & Ricklefs2000). Similar arguments have been used toexplain patterns in the pre-Pleistocene marineinvertebrate fossil record (Flessa 1975; Sepkoski1976; Flessa & Sepkoski 1978; Crame 2001). Butthe relationship between area and diversity isnot unequivocal, even for well documentedexamples such as SE Asia (Harrison et al. 2001),and the use of area per se must be viewed withcaution. As Rohde (1997) has pointed out inregard to latitudinal diversity gradients, the lowlatitudes today do not necessarily contain thelargest areas, despite being the location of thegreatest species richness. However, islands neednot be geographic entities (ocean islands), butmay include other isolated physiologicalfeatures (mountains, lakes) or distinct habitats
From: CRAME, J. A. & OWEN, A. W. (eds) 2002. Palaeobiogeography and Biodiversity Change: the Ordovicianand Mesozoic-Cenozoic Radiations. Geological Society, London, Special Publications, 194,179-199.0305-8719/02/$15.00 © The Geological Society of London 2002.

180 PAUL J. MARKWICK
(habitat 'islands'), although few habitat 'islands'are as absolute as ocean islands (Rosenzweig1995).
The association of areas of high species diver-sity with low latitudes, high temperatures andhigh incident solar energy fluxes, has given riseto the widely promoted species-energy theory(Pianka 1966; Currie 1991; Rohde 1992; Stevens1992; Currie & Fritz 1993; Latham & Ricklefs1993; Wright et al. 1993; Brown 1995; Rosen-zweig 1995). Currie (1991), comparing NorthAmerican terrestrial species diversity against 21environmental variables, found that speciesdiversity correlated best with potential evapo-transpiration (PET), considered to be an appro-priate measure of ambient energy. However,Kerr & Packer (1997) found that this was onlytrue for mammals in North America where PET< 1000 mm a-1 (Canada and Alaska). Stevens(1989) has suggested that climate (energy) vari-ability is the most important determinant ofspecies numbers, with few taxa being able tosurvive large seasonal variations. Such taxashould therefore have the largest latitudinalranges (Rapoport's rule), but this pattern hasnot been found in all groups (Rohde et al. 1993;Roy et al. 1994). For many this theory remainsequivocal (Pianka 1966; Rohde 1992; Roy et al.1994; Brown 1995; Rosenzweig 1995; Kerr &Packer 1997) because not all groups show cleardiversity gradients and the mechanism by whichenergy can dictate the number of species isuncertain.
The problem is that individual speciesrespond to different environmental factors (andcombinations of factors) and to different extentsdepending on their physiology and ecology(Root & Schneider 1993). Measuring only thenumber of species (taxonomic diversity) ratherthan the distribution of differences betweenorganisms (functional and ecological diversity)may obfuscate the processes dictating diversitypatterns (Gaston 2000). It is therefore essentialto be able to examine the macroscale spatialstructure of diversity in the context of physi-ology and behaviour (what an organism does),as well as taxonomy (what an organism iscalled), although for traditional classificationmethods there is often considerable overlapbetween the two. What is more, species richnessdoes not vary only with latitude (Brown 1995),nor is it independent of history (evolution andpalaeobiogeography). To understand globalpatterns therefore requires large, intercontin-ental datasets. Unfortunately, there have beenfew studies at this scale, and these are restrictedto analyses at relatively coarse taxonomic levelsand resolutions/grain (e.g. Gaston et al. 1995).
This paper presents a new, digital, geographicinformation system-based dataset with whichthe relationships between present-day terres-trial biodiversity, biogeography and climate areexamined. Examples of the observed macro-scale modern patterns are illustrated (both asmaps and bivariate plots) in order to facilitatecomparison with previous studies of NorthAmerica, especially that of Currie (1991). Theconsequences of derived relationships forinterpretations of palaeoclimate and palaeo-ecology are discussed. As a test of Ostrom's(1970) suggestion of using diversity gradientsto retrodict palaeoclimate, an experiment ispresented in which the Middle Eocenepalaeotemperature of Messel, Germany, isreconstructed using modern-day regressionsbetween observed taxonomic diversity andtemperature, and the results compared withvalues obtained from other methods.
MethodsThe dataset used in this study is part of a largecomputer-based ecological database of fossiland modern faunal and floral localities compiledby Markwick (1996). The database is designed tofacilitate analysis at any specified taxonomiclevel, such that differences between theresponse of families, genera and species can besystematically analysed. Data can also bequeried for any combination of parametersincluded in the database. Since this study wasbegun in 1990, the database has been integratedinto a geographic information system (ArcViewGIS and Arclnfo). The fossil data includedetailed specimen, environmental and strati-graphic information on about 6000 Cretaceousand Cenozoic fossil vertebrate localities, withtaxonomic and ecological data for 22 000 extantand fossil vertebrate and floral taxa (includinghabitat, size and diet). The modern dataanalysed here draw on the climate informationfrom 1060 climate stations (Fig. 1) taken fromMuller's (1982) compilation for vegetationstudies. Each station contains monthly data for14 climate variables including mean daily tem-perature, mean precipitation, radiation andpotential evapotranspiration (PET). A largenumber of additional parameters, includingannual metrics and combinations of variables,have been calculated using these data. A list ofvariables mentioned in this paper, their abbrevi-ations and explanations are given in Table 1. Thepropensity for stations to occur in lowland sitesreflects Muller's (1982) original requirements:acceptable stations must contain data for a largearray of climate parameters representing time

BIODIVERSITY AND CLIMATE 181
Fig. 1. The distribution of climate stations from Muller (1982) used in this study. Black circles representstations with complete non-avian tetrapod lists; grey circles represent those stations with only partial data.
series on the order of 30 years (typical of'climate'). Consequently most stations arelocated in large towns or cities that historicallyoccupy lowland sites. This bias closely mimicsthe biases in the geological record that tend toover-represent low-elevation environments.
Complete non-avian tetrapod faunal lists havebeen assigned to 408 of these stations: Europe(204); North America (143); southern Africa(21); and Australia (40). Seventy-two lists fromSouth America and 18 lists from Arabia containmammal data only. Floral data (Little 1977)
have been assigned to the North Americanstations, and the database also includes globalcoverage for all palms (Uhl & Dransfield 1987),turtles (Iverson 1992) and crocodilians (seereferences in Markwick 1998a).
Faunal lists are assigned to each station byoverlaying climate station distributions ontopublished species distribution maps (Hall &Kelson 1959; Van der Brink 1967; Little 1977;Arnold & Burton 1978; Cook 1984; Stebbins1985; Uhl & Dransfield 1987; Branch 1988;Bouchardy & Moutou 1989; Eisenberg 1989;
Table 1. Explanation of environmental variables used in this paper
Abbreviation
MATMARTCMMWMMRadiationCumulative T 0CumulativeT 5Anuual precipitationP rangeMonths T 10 P 40
PETMean annual NDVI
NDVI 1SD
Explanation
Mean annual temperatureMean annual range of temperatureColdest month mean temperatureWarmest month mean temperatureIncident solar radiation measured at surfaceSum of all degrees above 0°C ('degree months')Sum of all degrees above 5°C ('degree months')Mean annual precipitationMean annual range in precipitationNumber of months with mean temperatures greater than 10°C andPrecipitation greater than 40 mmPotential evapotranspirationMean annual negative difference vegetation index: a satellite-based metric usedas a proxy for primary productivityOne standard deviation of mean monthly NDVI
182 PAUL J. MARKWICK
Fig. 2. The relationship between the numbers of species and genera at each site used in the analysis. (a) Non-avian tetrapod species and genera by continent. (b) Species and genera by taxonomic group.
Kingdon 1990; Conant & Collins 1991; Grenard1991; Cogger 1992; Iverson 1992; Redford &Eisenberg 1992; Strahan 1992). A 50 km radiuscircle is drawn around each station, and anoccurrence registered where the taxon's distri-bution intersects this circle. A radial limit of 50km was chosen as it represents a typical decor-relation distance for precipitation, which is themost sensitive climate parameter to spatialheterogeneity. This means that the faunas andfloras can be confidently assumed to haveexperienced the climate assigned to them,except in areas with rapid relief changes (such asthe Alps), where this methodology mixes high-and low-elevation faunas. These points arefound to fall off the derived regressions but donot significantly affect results. Ecologically, thisapproach removes local, small-scale faunal andfloral heterogeneities, and thereby emulates thespatial and temporal time-averaging in the fossilrecord, with which this modern dataset canthereby be directly compared. The 50 km radiusalso approximately equates with the scale ofregional general circulation models (0.5° X 0.5°).
This approach differs from existing diversitygradient studies (e.g. Currie 1991) in two ways.Firstly, the use of point (station) rather thangridded data reduces the area effects implicit inquadrat techniques (Anderson & Marcus 1993),and allows a direct comparison with climate (theuse of latitudinal zones is to be avoided becauseit ignores longitudinal effects such as 'continen-tality'). Secondly, the method integrates datafrom more than one continent, thus reducing thepotential effects of regional biogeographical orhistorical artifacts.
The diversity data have been plotted onpresent-day maps using Arc View GIS for eachtaxonomic group in the database, as well as foreach habitat and diet category. This provides aqualitative indication of spatial similaritiesbetween biogeography, taxonomic diversity andenvironmental factors. The Spearman rank test(using the Statview software; Haycock et al1992-1993) was used to investigate correlationsbetween environmental variables and globaltaxonomic diversity (Table 2), and regionaldiversity (Tables 3-7). The relationship oftaxonomic assemblage composition to environ-mental factors was examined with correspon-dence analysis using the CANOCO software(Ter Braak 1987-1992). The use of genericrather than species-level presence-absenceinformation for this analysis is due to computa-tional limitations. However, analysis of thedataset has shown the close relationshipbetween the numbers of species and genera (Fig.2), such that results derived from either taxo-nomic level are comparable. Generic levelassignments are probably more robust for fossiltaxa.
Results
General patterns
Plots of diversity against absolute latitude areshown in Figure 3. For comparison, a selection ofenvironmental variables is plotted againstabsolute latitude in Figure 4 (see Table 1 for anexplanation of each variable). Total non-aviantetrapod species diversity (amphibians + reptiles
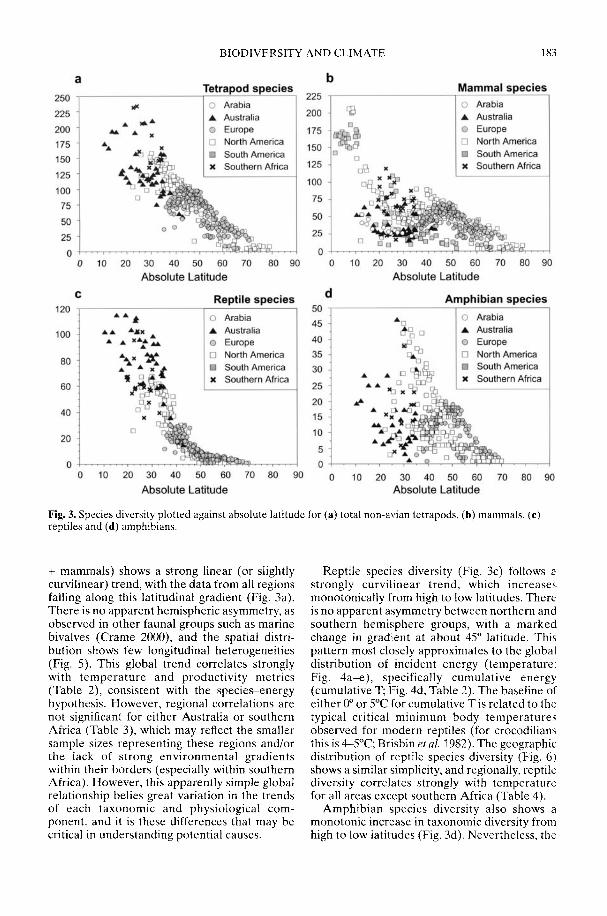
BIODIVERSITY AND CLIMATE 183
Fig. 3. Species diversity plotted against absolute latitude for (a) total non-avian tetrapods, (b) mammals, (c)reptiles and (d) amphibians.
+ mammals) shows a strong linear (or slightlycurvilinear) trend, with the data from all regionsfalling along this latitudinal gradient (Fig. 3a).There is no apparent hemispheric asymmetry, asobserved in other faunal groups such as marinebivalves (Crame 2000), and the spatial distri-bution shows few longitudinal heterogeneities(Fig. 5). This global trend correlates stronglywith temperature and productivity metrics(Table 2), consistent with the species-energyhypothesis. However, regional correlations arenot significant for either Australia or southernAfrica (Table 3), which may reflect the smallersample sizes representing these regions and/orthe lack of strong environmental gradientswithin their borders (especially within southernAfrica), However, this apparently simple globalrelationship belies great variation in the trendsof each taxonomic and physiological com-ponent, and it is these differences that may becritical in understanding potential causes.
Reptile species diversity (Fig. 3c) follows astrongly curvilinear trend, which increasesmonotonically from high to low latitudes. Thereis no apparent asymmetry between northern andsouthern hemisphere groups, with a markedchange in gradient at about 45° latitude. Thispattern most closely approximates to the globaldistribution of incident energy (temperature;Fig. 4a-e), specifically cumulative energy(cumulative T; Fig. 4d, Table 2). The baseline ofeither 0° or 5°C for cumulative T is related to thetypical critical minimum body temperaturesobserved for modern reptiles (for crocodiliansthis is 4-5°C; Brisbin et al 1982). The geographicdistribution of reptile species diversity (Fig. 6)shows a similar simplicity, and regionally, reptilediversity correlates strongly with temperaturefor all areas except southern Africa (Table 4).
Amphibian species diversity also shows amonotonic increase in taxonomic diversity fromhigh to low latitudes (Fig. 3d). Nevertheless, the
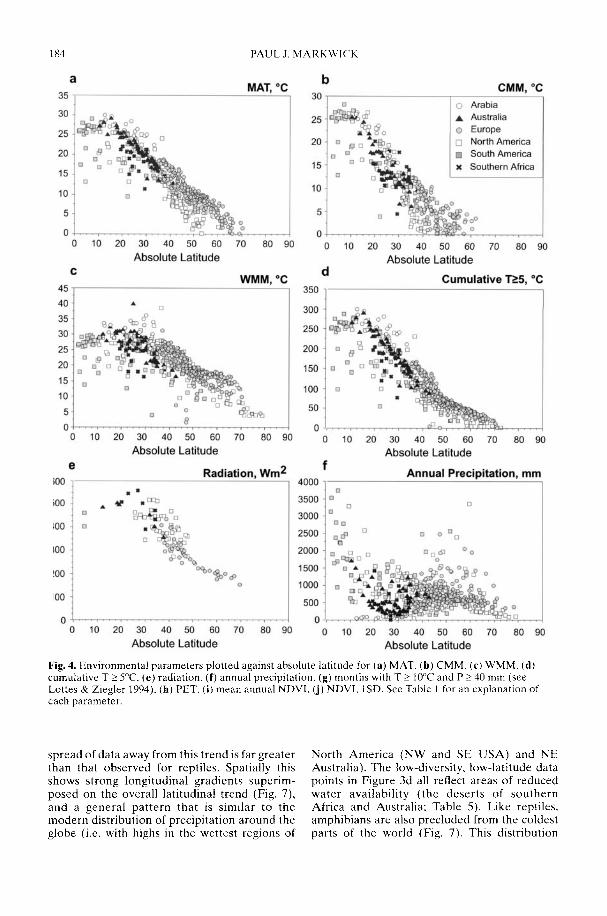
184 PAUL J. MARKWICK
Fig. 4. Environmental parameters plotted against absolute latitude for (a) MAT. (b) CMM, (c) WMM, (d)cumulative T 5°C, (e) radiation, (f) annual precipitation, (g) months with T 10°C and P 40 mm (seeLottes & Ziegler 1994), (h) PET, (i) mean annual NDVI, (j) NDVI, 1SD. See Table 1 for an explanation ofeach parameter.
spread of data away from this trend is far greaterthan that observed for reptiles. Spatially thisshows strong longitudinal gradients superim-posed on the overall latitudinal trend (Fig. 7),and a general pattern that is similar to themodern distribution of precipitation around theglobe (i.e. with highs in the wettest regions of
North America (NW and SE USA) and NEAustralia). The low-diversity, low-latitude datapoints in Figure 3d all reflect areas of reducedwater availability (the deserts of southernAfrica and Australia; Table 5). Like reptiles,amphibians are also precluded from the coldestparts of the world (Fig. 7). This distribution
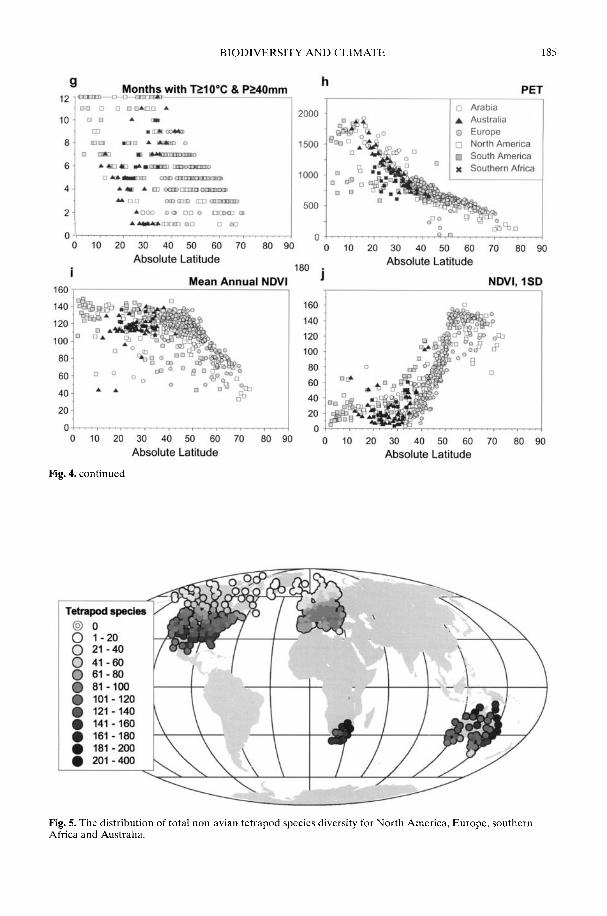
BIODIVERSITY AND CLIMATE 185
Fig. 4. continued
Fig. 5. The distribution of total non-avian tetrapod species diversity for North America, Europe, southernAfrica and Australia.
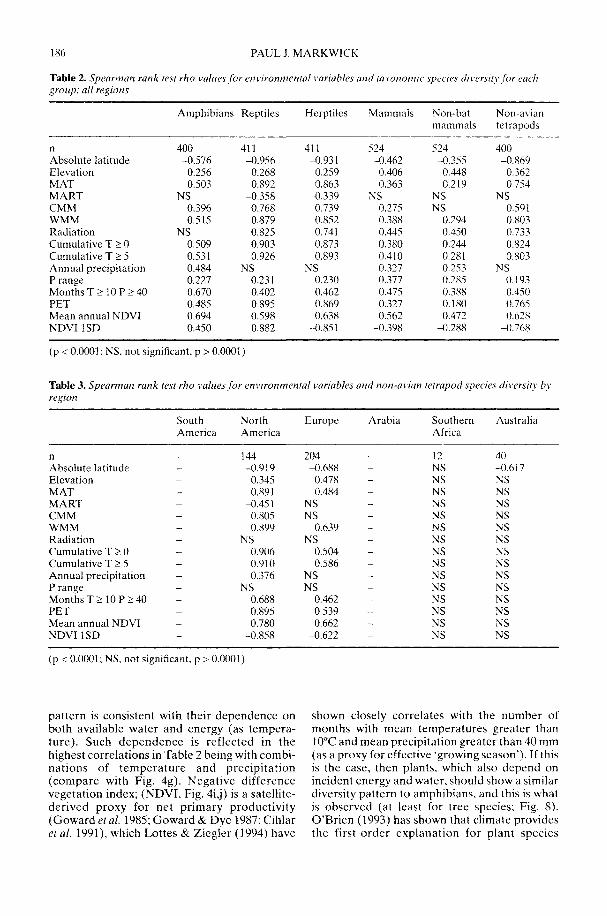
186 PAUL J. MARKWICK
Table 2. Spearman rank test rho values for environmental variables and taxonomic species diversity for eachgroup: all regions
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI 1SD
Amphibians
400-0.576
0,2560.503
NS0,3960.515
NS0.5090.5310.4840.2270.6700.4850.694
-0.450
Reptiles
411-0.9560.2680.892
-0.3580.7680.8790.8250.9030.926
NS0,2310.4020.8950.598
-0.882
Herptiles
411-0.931
0.2590.863
-0.3390.7390.8520.7410.8730.893
NS0.2300.4620.8690.638
-0.851
Mammals
524-0.4620.4060.363
NS0.2750.3880.4450.3800.4100.3270,3770.4750.3270.562
-0.398
Non-batmammals
524-0.3550.4480.219
NSNS
0.2940.4500.2440.2810.2530.2850.3880.1800.472
-0.288
Non-aviantetrapods
400-0.8690.3620.754
NS0.5910.8030.7330.8240.803
NS0.1930.4500.7650.628
-0.768
(p < 0.0001; NS, not significant, p > 0.0001)
Table 3. Spearman rank test rho values for environmental variables and non-avian tetrapod species diversity byregion
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI 1SD
SouthAmerica_
_____
_____
__
___
-
NorthAmerica
144-0.919
0.3450.891
-0.4510.8050.899
NS0.9060.9100.376
NS0.6880.8950.780
-0.858
Europe
204-0.6880.4780,484
NSNS
0.639NS
0.5040.586
NSNS
0.4620.5390.662
-0.622
Arabia
__--____-__
-_____
SouthernAfrica
12NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
Australia
40-0.617NSNSNSNSNSNSNSNSNSNSNSNSNSNS
(p < 0.0001; NS, not significant, p > 0.0001)
pattern is consistent with their dependence onboth available water and energy (as tempera-ture). Such dependence is reflected in thehighest correlations in Table 2 being with combi-nations of temperature and precipitation(compare with Fig. 4g). Negative differencevegetation index; (NDVI, Fig. 4i,j) is a satellite-derived proxy for net primary productivity(Goward et al 1985; Goward & Dye 1987; Cihlaret al 1991), which Lottes & Ziegler (1994) have
shown closely correlates with the number ofmonths with mean temperatures greater than10°C and mean precipitation greater than 40 mm(as a proxy for effective 'growing season'). If thisis the case, then plants, which also depend onincident energy and water, should show a similardiversity pattern to amphibians, and this is whatis observed (at least for tree species; Fig. 8).O'Brien (1993) has shown that climate providesthe first-order explanation for plant species
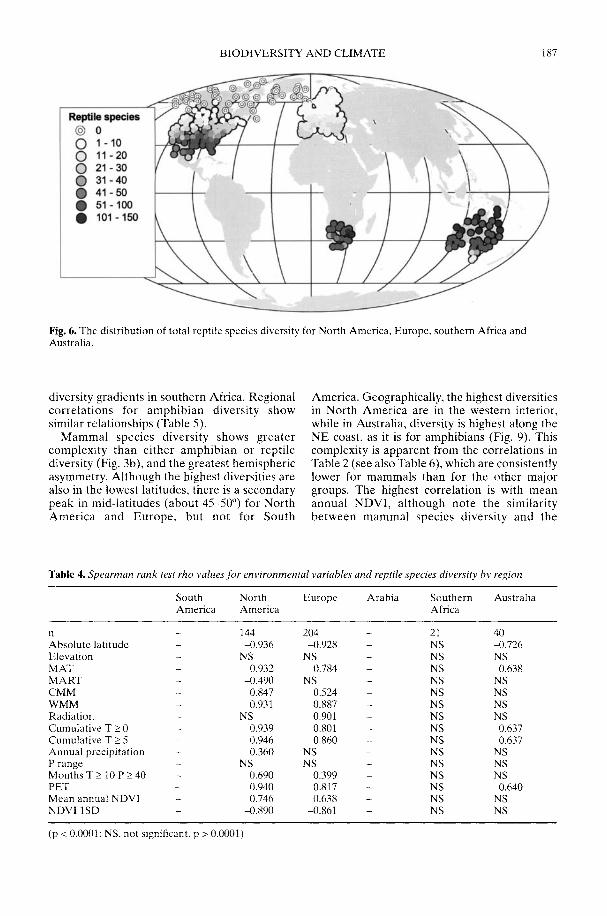
BIODIVERSITY AND CLIMATE 187
Fig. 6. The distribution of total reptile species diversity for North America, Europe, southern Africa andAustralia.
diversity gradients in southern Africa. Regionalcorrelations for amphibian diversity showsimilar relationships (Table 5).
Mammal species diversity shows greatercomplexity than either amphibian or reptilediversity (Fig. 3b), and the greatest hemisphericasymmetry. Although the highest diversities arealso in the lowest latitudes, there is a secondarypeak in mid-latitudes (about 45-50°) for NorthAmerica and Europe, but not for South
America. Geographically, the highest diversitiesin North America are in the western interior,while in Australia, diversity is highest along theNE coast, as it is for amphibians (Fig. 9). Thiscomplexity is apparent from the correlations inTable 2 (see also Table 6), which are consistentlylower for mammals than for the other majorgroups. The highest correlation is with meanannual NDVI, although note the similaritybetween mammal species diversity and the
Table 4. Spearman rank test rho values for environmental variables and reptile species diversity by region
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI 1SD
SouthAmerica
_----_---_----_-
NorthAmerica
144-0.936
NS0.932
-0.4900.8470.931
NS0.9390.9460.360
NS0.6900.9400.746
-0.890
Europe
204-0.928
NS0.784
NS0.5240.8870.9010.8010.860
NSNS
0.3990.8170.638
-0.861
Arabia
____________--___-
SouthernAfrica
21NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
Australia
40-0.726NS0.638
NSNSNSNS0.6370.637
NSNSNS0.640
NSNS
(p < 0.0001; NS, not significant, p > 0.0001)
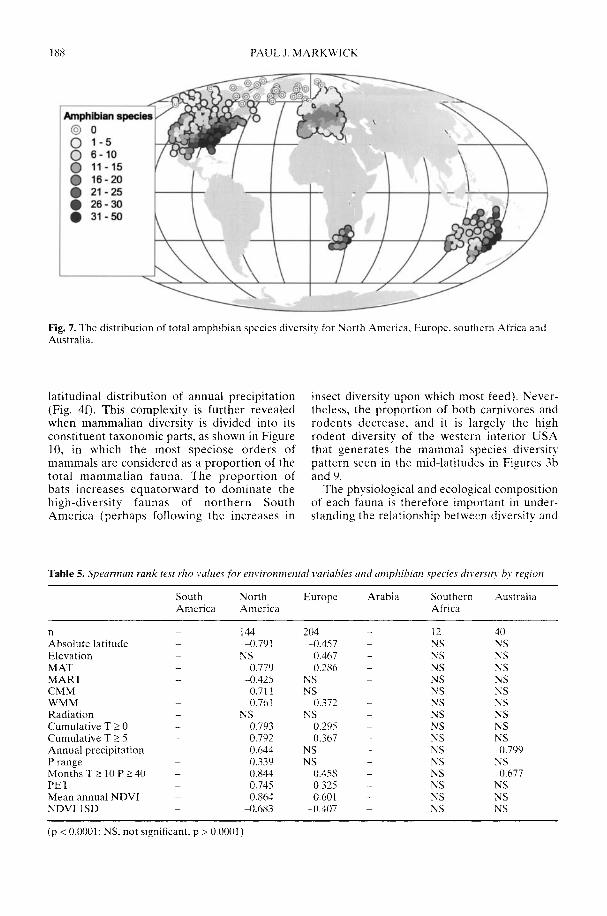
188 PAUL J. MARKWICK
Fig. 7. The distribution of total amphibian species diversity for North America, Europe, southern Africa andAustralia.
latitudinal distribution of annual precipitation(Fig. 4f), This complexity is further revealedwhen mammalian diversity is divided into itsconstituent taxonomic parts, as shown in Figure10, in which the most speciose orders ofmammals are considered as a proportion of thetotal mammalian fauna. The proportion ofbats increases equatorward to dominate thehigh-diversity faunas of northern SouthAmerica (perhaps following the increases in
insect diversity upon which most feed). Never-theless, the proportion of both carnivores androdents decrease, and it is largely the highrodent diversity of the western interior USAthat generates the mammal species diversitypattern seen in the mid-latitudes in Figures 3band 9.
The physiological and ecological compositionof each fauna is therefore important in under-standing the relationship between diversity and
Table 5. Spearman rank test rho values for environmental variables and amphibian species diversity by region
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI ISO
SouthAmerica
_---_----_-_----
NorthAmerica
144-0.791
NS0,779
-0.4250.7110.761
NS0.7930.7920.6440.3390.8440.7450.864
-0.683
Europe
204-0.457
0.4670.286
NSNS
0.372NS
0.2950.367
NSNS
0.4580.3250.601
-0.407
Arabia
__________________
SouthernAfrica
12NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
Australia
40NSNSNSNSNSNSNSNSNS
0.799NS0.677
NSNSNS
(p < 0.0001; NS, not significant, p > 0.0001)
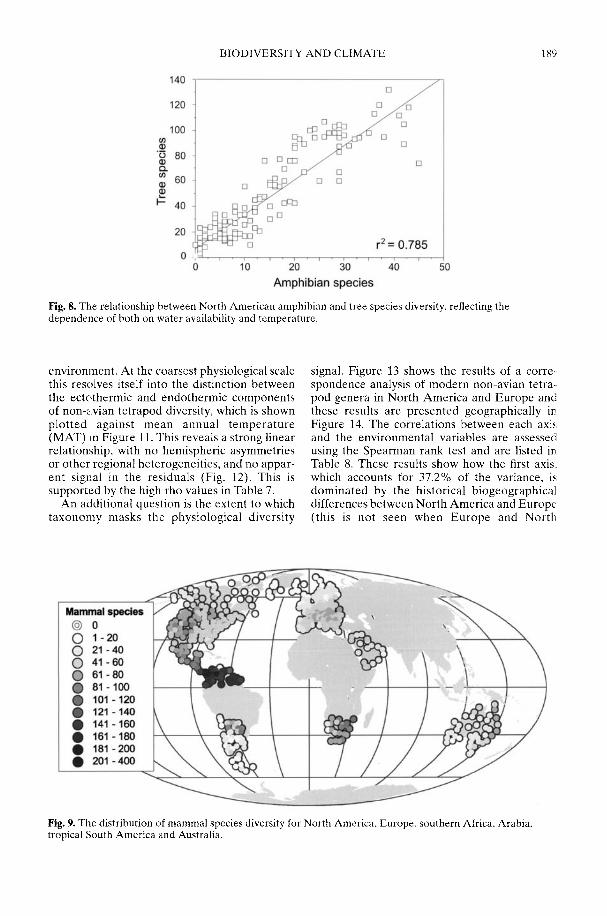
BIODIVERSITY AND CLIMATE 189
Fig. 8. The relationship between North American amphibian and tree species diversity, reflecting thedependence of both on water availability and temperature.
environment. At the coarsest physiological scalethis resolves itself into the distinction betweenthe ectothermic and endothermic componentsof non-avian tetrapod diversity, which is shownplotted against mean annual temperature(MAT) in Figure 11. This reveals a strong linearrelationship, with no hemispheric asymmetriesor other regional heterogeneities, and no appar-ent signal in the residuals (Fig. 12). This issupported by the high rho values in Table 7.
An additional question is the extent to whichtaxonomy masks the physiological diversity
signal. Figure 13 shows the results of a corre-spondence analysis of modern non-avian tetra-pod genera in North America and Europe andthese results are presented geographically inFigure 14. The correlations between each axisand the environmental variables are assessedusing the Spearman rank test and are listed inTable 8. These results show how the first axis,which accounts for 37.2% of the variance, isdominated by the historical biogeographicaldifferences between North America and Europe(this is not seen when Europe and North
Fig. 9. The distribution of mammal species diversity for North America, Europe, southern Africa, Arabia,tropical South America and Australia.
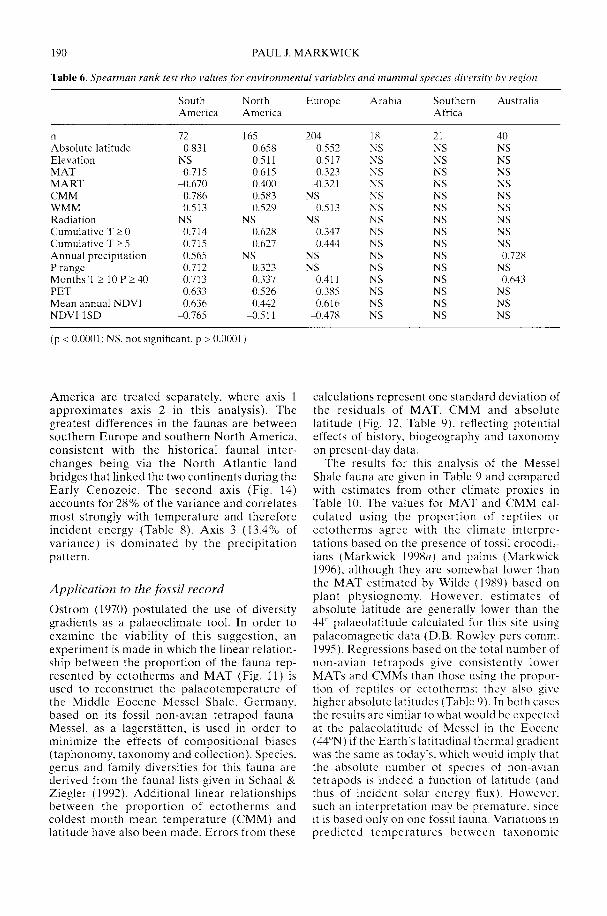
190 PAUL J. MARKWICK
Table 6. Spearman rank test rho values for environmental variables and mammal species diversity by region
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI 1SD
SouthAmerica
72-0.831NS0.715
-0.6700.7860.513
NS0.7140.7150.5650.7120.7130.6330.636
-0.765
NorthAmerica
165-0.6580.5110.615
-0.4000.5830.529
NS0.6280.627
NS0.3230.3370.5260.442
-0.511
Europe
204-0.552
0.5170.323
-0.321NS
0.513NS
0.3470.444
NSNS
0.4110.3850.616
-0.478
Arabia
18NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
SouthernAfrica
21NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
Australia
40NSNSNSNSNSNSNSNSNS0.728
NS0.643
NSNSNS
(p < 0.0001: NS, not significant, p > 0.0001)
America are treated separately, where axis 1approximates axis 2 in this analysis). Thegreatest differences in the faunas are betweensouthern Europe and southern North America,consistent with the historical faunal inter-changes being via the North Atlantic landbridges that linked the two continents during theEarly Cenozoic. The second axis (Fig. 14)accounts for 28% of the variance and correlatesmost strongly with temperature and thereforeincident energy (Table 8). Axis 3 (13.4% ofvariance) is dominated by the precipitationpattern.
Application to the fossil record
Ostrom (1970) postulated the use of diversitygradients as a palaeoclimate tool. In order toexamine the viability of this suggestion, anexperiment is made in which the linear relation-ship between the proportion of the fauna rep-resented by ectotherms and MAT (Fig. 11) isused to reconstruct the palaeotemperature ofthe Middle Eocene Messel Shale, Germany,based on its fossil non-avian tetrapod fauna.Messel, as a lagerstatten, is used in order tominimize the effects of compositional biases(taphonomy, taxonomy and collection). Species,genus and family diversities for this fauna arederived from the faunal lists given in Schaal &Ziegler (1992). Additional linear relationshipsbetween the proportion of ectotherms andcoldest month mean temperature (CMM) andlatitude have also been made. Errors from these
calculations represent one standard deviation ofthe residuals of MAT, CMM and absolutelatitude (Fig. 12, Table 9), reflecting potentialeffects of history, biogeography and taxonomyon present-day data.
The results for this analysis of the MesselShale fauna are given in Table 9 and comparedwith estimates from other climate proxies inTable 10. The values for MAT and CMM cal-culated using the proportion of reptiles orectotherms agree with the climate interpre-tations based on the presence of fossil crocodil-ians (Markwick 1998a) and palms (Markwick1996), although they are somewhat lower thanthe MAT estimated by Wilde (1989) based onplant physiognomy. However, estimates ofabsolute latitude are generally lower than the44° palaeolatitude calculated for this site usingpalaeomagnetic data (D.B. Rowley pers comm.1995). Regressions based on the total number ofnon-avian tetrapods give consistently lowerMATs and CMMs than those using the propor-tion of reptiles or ectotherms; they also givehigher absolute latitudes (Table 9). In both casesthe results are similar to what would be expectedat the palaeolatitude of Messel in the Eocene(44°N) if the Earth's latitudinal thermal gradientwas the same as today's, which would imply thatthe absolute number of species of non-aviantetrapods is indeed a function of latitude (andthus of incident solar energy flux). However,such an interpretation may be premature, sinceit is based only on one fossil fauna. Variations inpredicted temperatures between taxonomic
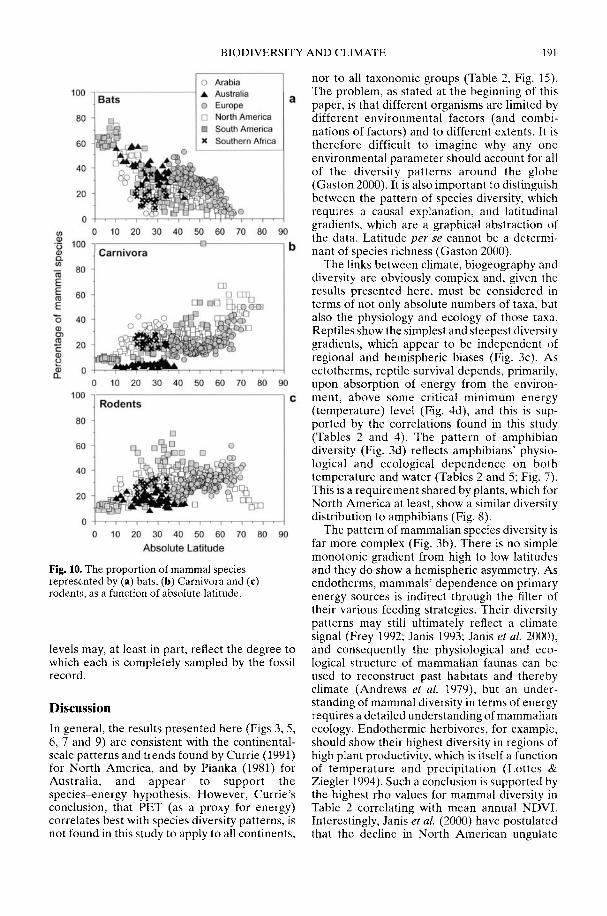
BIODIVERSITY AND CLIMATE 191
Fig. 10. The proportion of mammal speciesrepresented by (a) bats, (b) Carnivora and (c)rodents, as a function of absolute latitude.
levels may, at least in part, reflect the degree towhich each is completely sampled by the fossilrecord.
DiscussionIn general, the results presented here (Figs 3, 5,6, 7 and 9) are consistent with the continental-scale patterns and trends found by Currie (1991)for North America, and by Pianka (1981) forAustralia, and appear to support thespecies-energy hypothesis. However, Currie'sconclusion, that PET (as a proxy for energy)correlates best with species diversity patterns, isnot found in this study to apply to all continents,
nor to all taxonomic groups (Table 2, Fig. 15).The problem, as stated at the beginning of thispaper, is that different organisms are limited bydifferent environmental factors (and combi-nations of factors) and to different extents. It istherefore difficult to imagine why any oneenvironmental parameter should account for allof the diversity patterns around the globe(Gaston 2000). It is also important to distinguishbetween the pattern of species diversity, whichrequires a causal explanation, and latitudinalgradients, which are a graphical abstraction ofthe data. Latitude per se cannot be a determi-nant of species richness (Gaston 2000).
The links between climate, biogeography anddiversity are obviously complex and, given theresults presented here, must be considered interms of not only absolute numbers of taxa, butalso the physiology and ecology of those taxa.Reptiles show the simplest and steepest diversitygradients, which appear to be independent ofregional and hemispheric biases (Fig. 3c). Asectotherms, reptile survival depends, primarily,upon absorption of energy from the environ-ment, above some critical minimum energy(temperature) level (Fig. 4d), and this is sup-ported by the correlations found in this study(Tables 2 and 4). The pattern of amphibiandiversity (Fig. 3d) reflects amphibians' physio-logical and ecological dependence on bothtemperature and water (Tables 2 and 5; Fig. 7).This is a requirement shared by plants, which forNorth America at least, show a similar diversitydistribution to amphibians (Fig. 8).
The pattern of mammalian species diversity isfar more complex (Fig. 3b). There is no simplemonotonic gradient from high to low latitudesand they do show a hemispheric asymmetry. Asendotherms, mammals' dependence on primaryenergy sources is indirect through the filter oftheir various feeding strategies. Their diversitypatterns may still ultimately reflect a climatesignal (Frey 1992; Janis 1993; Janis et al 2000),and consequently the physiological and eco-logical structure of mammalian faunas can beused to reconstruct past habitats and therebyclimate (Andrews et al. 1979), but an under-standing of mammal diversity in terms of energyrequires a detailed understanding of mammalianecology. Endothermic herbivores, for example,should show their highest diversity in regions ofhigh plant productivity, which is itself a functionof temperature and precipitation (Lottes &Ziegler 1994). Such a conclusion is supported bythe highest rho values for mammal diversity inTable 2 correlating with mean annual NDVI.Interestingly, Janis et al. (2000) have postulatedthat the decline in North American ungulate
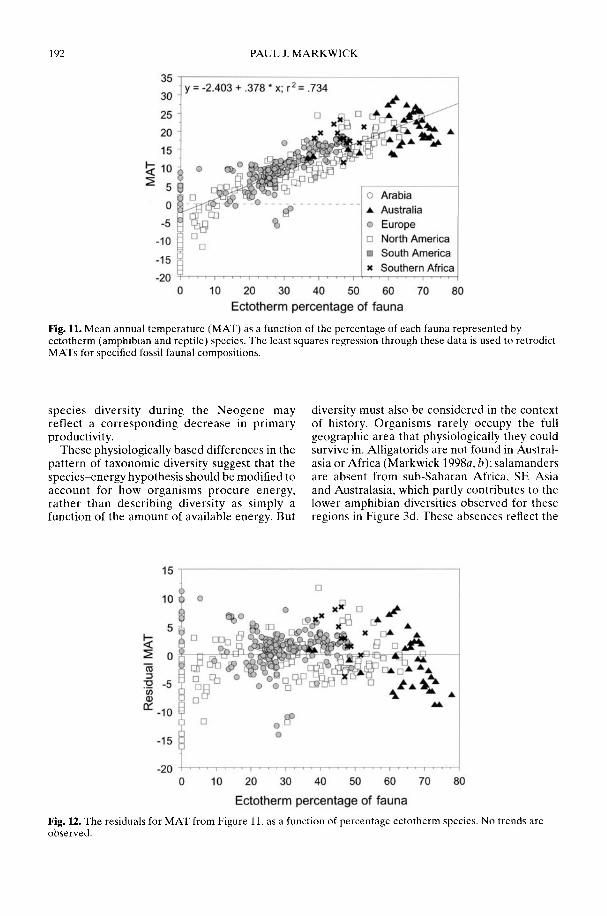
192 PAUL J. MARKWICK
Fig. 11. Mean annual temperature (MAT) as a function of the percentage of each fauna represented byectotherm (amphibian and reptile) species. The least squares regression through these data is used to retrodictMATs for specified fossil faunal compositions.
species diversity during the Neogene mayreflect a corresponding decrease in primaryproductivity.
These physiologically based differences in thepattern of taxonomic diversity suggest that thespecies-energy hypothesis should be modified toaccount for how organisms procure energy,rather than describing diversity as simply afunction of the amount of available energy. But
diversity must also be considered in the contextof history. Organisms rarely occupy the fullgeographic area that physiologically they couldsurvive in. Alligatorids are not found in Austral-asia or Africa (Markwick 1998a,b); salamandersare absent from sub-Saharan Africa, SE Asiaand Australasia, which partly contributes to thelower amphibian diversities observed for theseregions in Figure 3d. These absences reflect the
Fig. 12. The residuals for MAT from Figure 11, as a function of percentage ectotherm species. No trends areobserved.
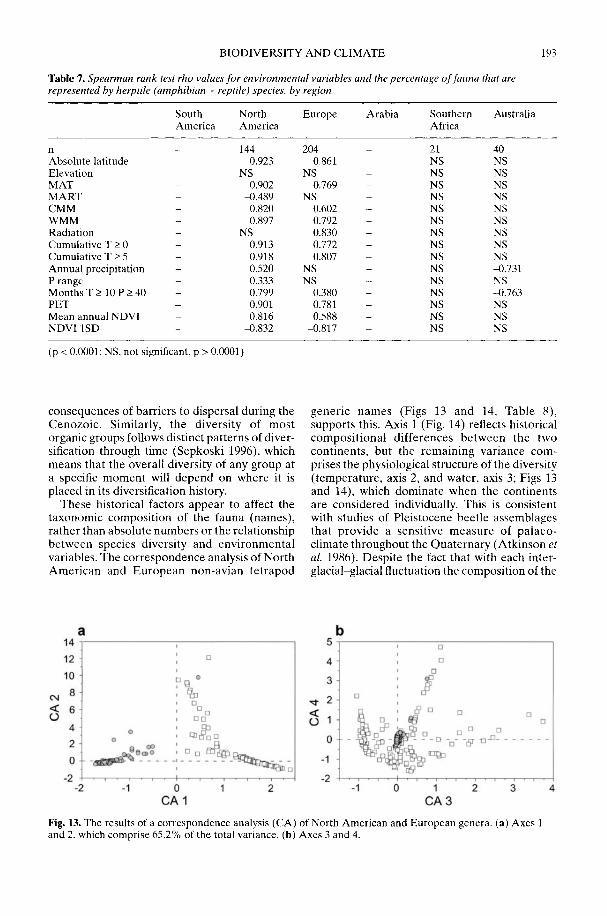
BIODIVERSITY AND CLIMATE 193
Table 7. Spearman rank test rho values for environmental variables and the percentage of fauna that arerepresented by herptile (amphibian + reptile) species, by region
nAbsolute latitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI ISD
SouthAmerica
_---------------
NorthAmerica
144-0.923
NS0.902
-0.4890.8200.897
NS0.9130.9180.5200.3330.7990.9010.816
-0.832
Europe
204-0.861
NS0.769
NS0.6020.7920.8300.7720.807
NSNS
0.3800.7810.588
-0.817
Arabia
_-----------_---
SouthernAfrica
21NSNSNSNSNSNSNSNSNSNSNSNSNSNSNS
Australia
40NSNSNSNSNSNSNSNSNS-0.731NS-0.763NSNSNS
(p < 0.0001; NS, not significant, p > 0.0001)
consequences of barriers to dispersal during theCenozoic. Similarly, the diversity of mostorganic groups follows distinct patterns of diver-sification through time (Sepkoski 1996), whichmeans that the overall diversity of any group ata specific moment will depend on where it isplaced in its diversification history.
These historical factors appear to affect thetaxonomic composition of the fauna (names),rather than absolute numbers or the relationshipbetween species diversity and environmentalvariables. The correspondence analysis of NorthAmerican and European non-avian tetrapod
generic names (Figs 13 and 14, Table 8),supports this. Axis 1 (Fig. 14) reflects historicalcompositional differences between the twocontinents, but the remaining variance com-prises the physiological structure of the diversity(temperature, axis 2, and water, axis 3; Figs 13and 14), which dominate when the continentsare considered individually. This is consistentwith studies of Pleistocene beetle assemblagesthat provide a sensitive measure of palaeo-climate throughout the Quaternary (Atkinson etal 1986). Despite the fact that with each inter-glacial-glacial fluctuation the composition of the
Fig. 13. The results of a correspondence analysis (CA) of North American and European genera, (a) Axes 1and 2, which comprise 65.2% of the total variance, (b) Axes 3 and 4.
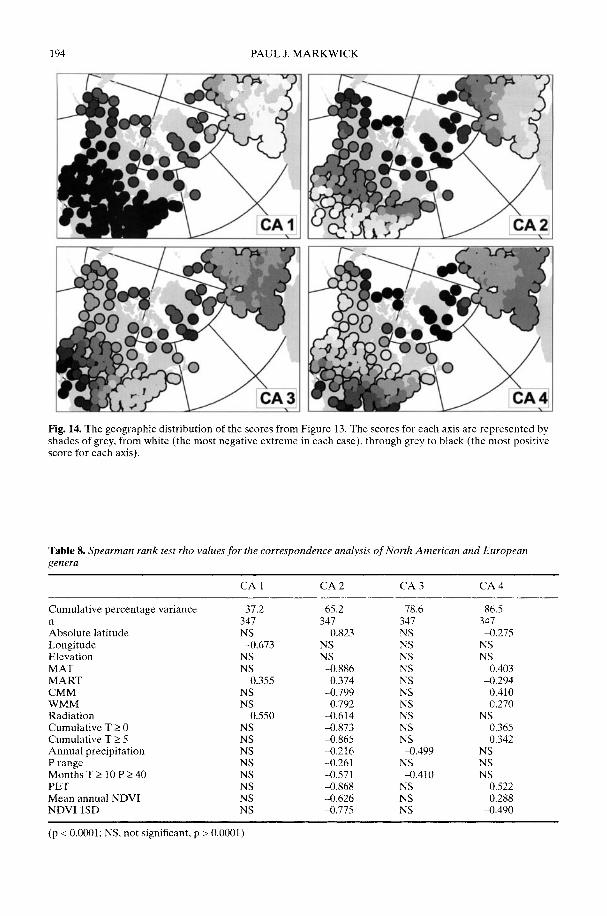
194 PAUL J. MARKWICK
Fig. 14, The geographic distribution of the scores from Figure 13. The scores for each axis are represented byshades of grey, from white (the most negative extreme in each case), through grey to black (the most positivescore for each axis).
Table 8, Spearman rank test rho values for the correspondence analysis of North American and Europeangenera
Cumulative percentage variancenAbsolute latitudeLongitudeElevationMATMARTCMMWMMRadiationCumulative T 0Cumulative T 5Annual precipitationP rangeMonths T 10 P 40PETMean annual NDVINDVI 1SD
CA1
37.2347NS-0.673
NSNS
0355NSNS
0,550NSNSNSNSNSNSNSNS
CA2
65.2347
0.823NSNS-0.8860.374
-0.799-0.792-0.614-0.873-0.865-0.216-0,261-0.571-0.868-0.626-0.775
CA3
78.6347NSNSNSNSNSNSNSNSNSNS-0.499
NS-0.410
NSNSNS
CA4
86.5347-0.275
NSNS
0,403-0.2940,4100.270
NS0.3650.342
NSNSNS
0,5220.288
-0.490
(p < 0.0001; NS, not significant, p > 0.0001)
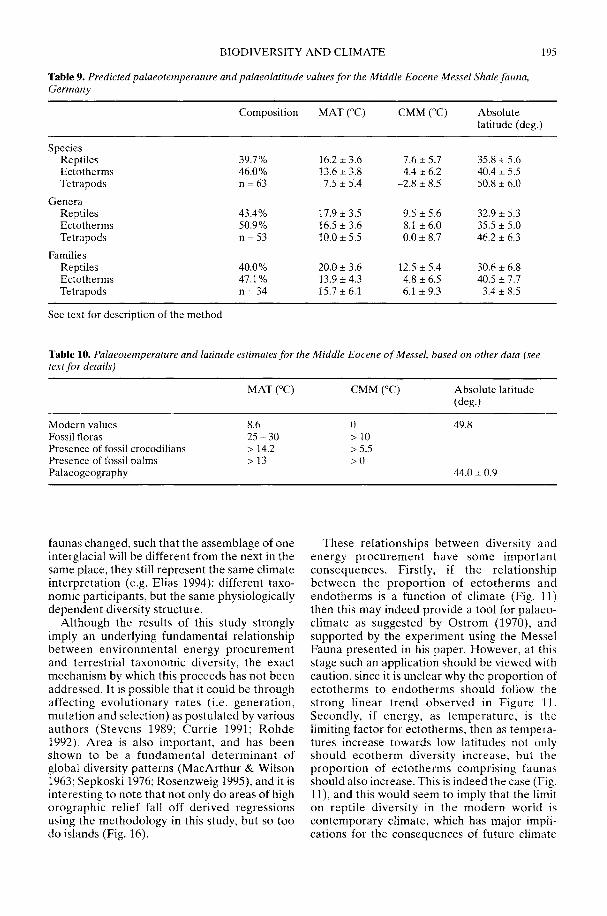
BIODIVERSITY AND CLIMATE 195
Table 9. Predicted palaeotemperature and palaeolatitude values for the Middle Eocene Messel Shale fauna,Germany
SpeciesReptilesEctothermsTetrapods
GeneraReptilesEctothermsTetrapods
FamiliesReptilesEctothermsTetrapods
Composition
39.7%46.0%n = 63
43.4%50.9%n = 53
40.0%47.1%n-34
MAT (°C)
16.2 ±3.613.6 ±3.87.5 ±5.4
17.9 ±3.516.5 ± 3.610.0 ± 5.5
20.0 ± 3.613.9 ±4.315.7 ±6.1
CMM (°C)
7.6 ± 5.74.4 ± 6.2
-2.8 ± 8.5
9.5 ± 5.68.1 ± 6.00.0 ± 8.7
12.5 ± 5.44.8 ± 6.56.1 ± 9.3
Absolutelatitude (deg.)
35.8 ± 5.640.4 ± 5.550.8 ± 6.0
32.9 ± 5.335.5 ± 5.046.2 ± 6.3
30.6 ± 6.840.5 ± 7.73.4 ± 8.5
See text for description of the method
Table 10. Palaeotemperature and latitude estimates for the Middle Eocene of Messel, based on other data (seetext for details)
Modern valuesFossil florasPresence of fossil crocodiliansPresence of fossil palmsPalaeogeography
MAT (°C)
8.625-30>14.2>13
CMM (°C)
0>10>5.5>0
Absolute latitude(deg.)
49.8
44.0 ± 0.9
faunas changed, such that the assemblage of oneinterglacial will be different from the next in thesame place, they still represent the same climateinterpretation (e.g. Elias 1994): different taxo-nomic participants, but the same physiologicallydependent diversity structure.
Although the results of this study stronglyimply an underlying fundamental relationshipbetween environmental energy procurementand terrestrial taxonomic diversity, the exactmechanism by which this proceeds has not beenaddressed. It is possible that it could be throughaffecting evolutionary rates (i.e. generation,mutation and selection) as postulated by variousauthors (Stevens 1989; Currie 1991; Rohde1992). Area is also important, and has beenshown to be a fundamental determinant ofglobal diversity patterns (MacArthur & Wilson1963; Sepkoski 1976; Rosenzweig 1995), and it isinteresting to note that not only do areas of highorographic relief fall off derived regressionsusing the methodology in this study, but so toodo islands (Fig. 16).
These relationships between diversity andenergy procurement have some importantconsequences. Firstly, if the relationshipbetween the proportion of ectotherms andendotherms is a function of climate (Fig. 11)then this may indeed provide a tool for palaeo-climate as suggested by Ostrom (1970), andsupported by the experiment using the MesselFauna presented in his paper. However, at thisstage such an application should be viewed withcaution, since it is unclear why the proportion ofectotherms to endotherms should follow thestrong linear trend observed in Figure 11.Secondly, if energy, as temperature, is thelimiting factor for ectotherms, then as tempera-tures increase towards low latitudes not onlyshould ecotherm diversity increase, but theproportion of ectotherms comprising faunasshould also increase. This is indeed the case (Fig.11), and this would seem to imply that the limiton reptile diversity in the modern world iscontemporary climate, which has major impli-cations for the consequences of future climate
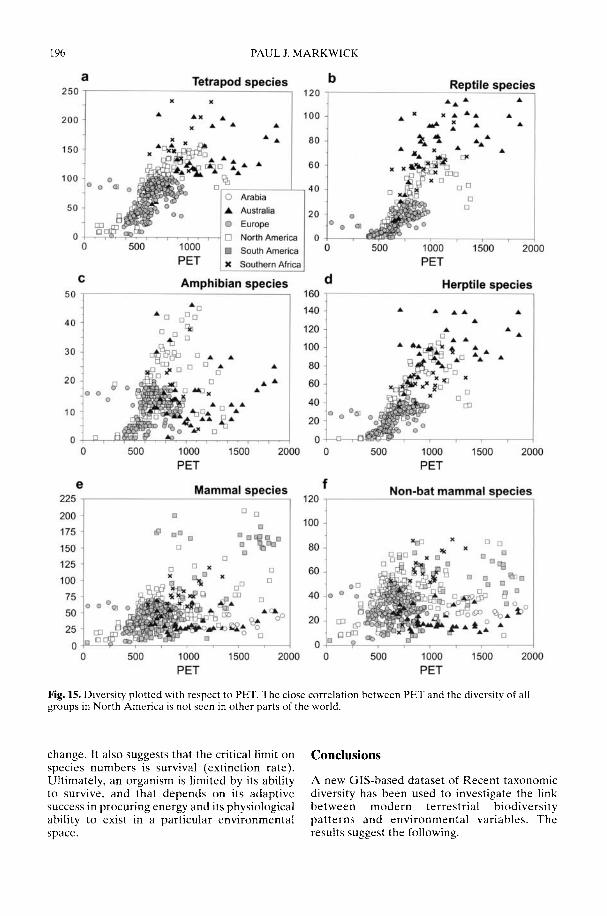
196 PAUL J. MARKWICK
Fig. 15. Diversity plotted with respect to PET. The close correlation between PET and the diversity of allgroups in North America is not seen in other parts of the world.
change. It also suggests that the critical limit onspecies numbers is survival (extinction rate).Ultimately, an organism is limited by its abilityto survive, and that depends on its adaptivesuccess in procuring energy and its physiologicalability to exist in a particular environmentalspace.
Conclusions
A new GIS-based dataset of Recent taxonomicdiversity has been used to investigate the linkbetween modern terrestrial biodiversitypatterns and environmental variables. Theresults suggest the following.
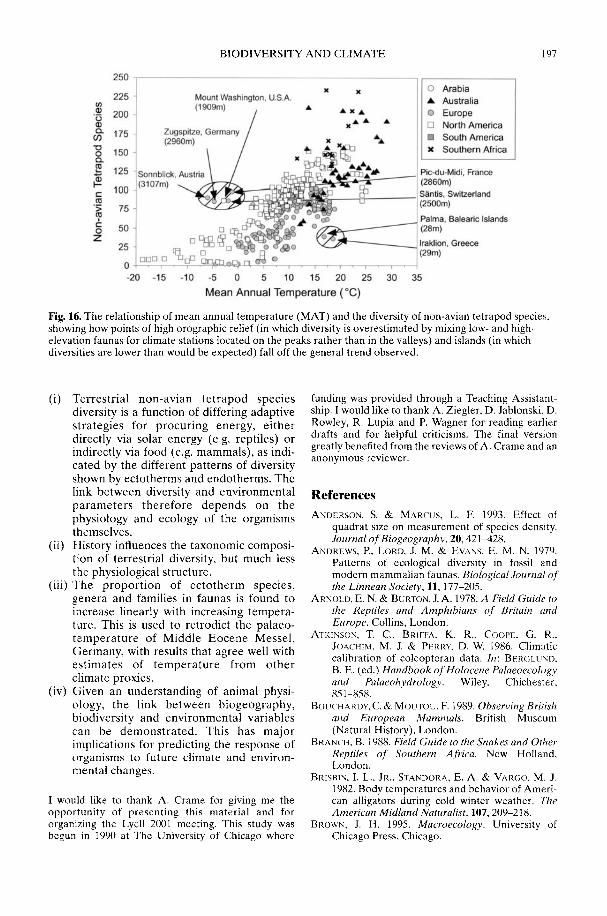
BIODIVERSITY AND CLIMATE 197
Fig. 16. The relationship of mean annual temperature (MAT) and the diversity of non-avian tetrapod species,showing how points of high orographic relief (in which diversity is overestimated by mixing low- and high-elevation faunas for climate stations located on the peaks rather than in the valleys) and islands (in whichdiversities are lower than would be expected) fall off the general trend observed.
(i) Terrestrial non-avian tetrapod speciesdiversity is a function of differing adaptivestrategies for procuring energy, eitherdirectly via solar energy (e.g. reptiles) orindirectly via food (e.g. mammals), as indi-cated by the different patterns of diversityshown by ectotherms and endotherms. Thelink between diversity and environmentalparameters therefore depends on thephysiology and ecology of the organismsthemselves.
(ii) History influences the taxonomic composi-tion of terrestrial diversity, but much lessthe physiological structure.
(iii) The proportion of ectotherm species,genera and families in faunas is found toincrease linearly with increasing tempera-ture. This is used to retrodict the palaeo-temperature of Middle Eocene Messel,Germany, with results that agree well withestimates of temperature from otherclimate proxies.
(iv) Given an understanding of animal physi-ology, the link between biogeography,biodiversity and environmental variablescan be demonstrated. This has majorimplications for predicting the response oforganisms to future climate and environ-mental changes.
I would like to thank A. Crame for giving me theopportunity of presenting this material and fororganizing the Lyell 2001 meeting. This study wasbegun in 1990 at The University of Chicago where
funding was provided through a Teaching Assistant-ship. I would like to thank A. Ziegler, D. Jablonski, D.Rowley, R. Lupia and P. Wagner for reading earlierdrafts and for helpful criticisms. The final versiongreatly benefited from the reviews of A. Crame and ananonymous reviewer.
ReferencesANDERSON, S. & MARCUS, L. F. 1993. Effect of
quadrat size on measurement of species density.Journal of Biogeography, 20,421-28.
ANDREWS, P., LORD, J. M. & EVANS, E. M. N. 1979.Patterns of ecological diversity in fossil andmodern mammalian faunas. Biological Journal ofthe Linnean Society, 11,177-205.
ARNOLD, E. N. & BURTON, J. A. 1978. A Field Guide tothe Reptiles and Amphibians of Britain andEurope. Collins, London.
ATKINSON, T. C, BRIFFA, K. R., COOPE, G. R.,JOACHIM, M. J. & PERRY, D. W. 1986. Climaticcalibration of coleopteran data. In: BERGLUND,B. E. (ed.) Handbook of Holocene Palaeoecologyand Palaeohydrology. Wiley, Chichester,851-858.
BOUCHARDY,C. & MOUTOU, F. 1989. Observing Britishand European Mammals. British Museum(Natural History), London.
BRANCH, B. 1988. Field Guide to the Snakes and OtherReptiles of Southern Africa. New Holland,London.
BRISBIN, I. L., JR., STANDORA, E. A. & VARGO, M. J.1982. Body temperatures and behavior of Ameri-can alligators during cold winter weather. TheAmerican Midland Naturalist, 107, 209-218.
BROWN, J. H. 1995. Macroecology. University ofChicago Press, Chicago.

198 PAUL J. MARKWICK
CIHLAR, J., ST.-LAURENT, L. & DYER, J. A. 1991.Relation between the Normalized DifferenceVegetation Index and ecological variables.Remote Sensing of Environment, 35, 279-298.
COGGER, H. G. 1992. Reptiles and Amphibians ofAustralia. Comstock, Cornell.
COLBERT, E. H., COWLES, R. B. & BOGERT, C. M. 1946.Temperature tolerances in the American alligatorand their bearing on the habits, evolution, andextinction of the Dinosaurs. Bulletin of theAmerican Museum of Natural History, 86,327-374.
CONANT, R. & COLLINS, J. T. 1991. A Field Guide toReptiles and Amphibians. Eastern/Central NorthAmerica. Houghton Mifflin, Boston.
COOK, F. R. 1984. Introduction to Canadian Amphib-ians and Reptiles. National Museums of Canada,Ottawa.
CRAME, J. A. 2000. Evolution of taxonomic diversitygradients in the marine realm: evidence from thecomposition of Recent bivalve faunas. Paleobiol-ogy, 26,188-214.
CRAME, J. A. 2001. Taxonomic diversity gradientsthrough geological time. Diversity & Distri-butions, 7,175-189.
CURRIE, D. J. 1991. Energy and large-scale patterns ofanimal- and plant- species richness. The AmericanNaturalist, 137, 27-49.
CURRIE, D. J. & FRITZ, J. T. 1993. Global patterns ofanimal abundance and species energy use.OIKOS, 67, 56-68.
DARLINGTON, P. J. 1948. The geographical distributionof cold-blooded vertebrates. The QuarterlyReview of Biology, 23,1-26,105-123.
EISENBERG. J. F. 1989. Mammals of the Neotropics. TheNorthern Neotropics. Volume 1. Panama, Colom-bia, Venezuela, Guyana, Suriname, FrenchGuiana. University of Chicago Press, Chicago.
ELI AS, S. A. 1994. Quaternary Insects and theirEnvironments. Smithsonian Institution Press,Washington, DC,
FISCHER, A. G. 1961. Latitudinal variations in organicdiversity. American Scientist, 49, 50-74.
FLESSA, K. W. 1975. Area, continental drift andmammalian diversity. Paleobiology, 1, 189-194.
FLESSA, K. W. & SEPKOSKI, J. J. JR 1978. On the relation-ship between Phanerozoic diversity and changesin habitable area. Paleobiology, 4, 359-366.
FREY, J. K. 1992. Response of a mammalian faunalelement to climatic changes. Journal of Mammal-ogy, 73, 43-50.
GASTON, K. J. 2000. Global patterns in biodiversity.Nature, 405, 220-227.
GASTON, K. J., WILLIAMS, P. H., EGGLETON, P. &HUMPHRIES, C. 1995. Large scale patterns ofbiodiversity: spatial variation in family richness.Proceedings of the Royal Society, London, B, 260,149-154.
GOWARD, S. N. & DYE, D. G. 1987. Evaluating NorthAmerican net primary productivity with satelliteobservations. Advances in Space Research, 7,165-174.
GOWARD, S. N, TUCKER, C. J. & DYE, D. G. 1985.North American vegetation patterns observedwith the NOAA-7 advanced very high resolutionradiometer. Vegetatio. 64, 3-14.
GRENARD, S. 1991. Handbook of Alligators andCrocodiles. Krierger Publishing Company.Malabar, Florida.
HALL, E. R. & KELSON, K. R. 1959. The Mammals ofNorth America. Ronald Press, New York.
HARRISON, S. P., YU, G.,TAKAHARA. H. & PRENTICE, I.C. 2001. Plant diversity and palaeovegetation inEast Asia. Nature, 413, 129-130.
HAYCOCK, K., ROTH, J. & GAGNON, J. 1992-1993.Statview. Abacus Concepts. Inc.. Berkeley.California.
HIBBARD, C. W. 1960. An interpretation of Plioceneand Pleistocene climates in North America.Annual Report of the Michigan Academy ofScience, Arts, and Letters, 62, 5-30.
HUMBOLDT, A. V. & BONPLAND, A. 1807. Essai sur lageographic des plantes; accompagne d'un tableauphysique des regions equinoxiales. Fr. Schoell.Paris.
IVERSON, J. B. 1992. A Revised Checklist with Distri-bution Maps of the Turtles of the World. Privatelypublished, Richmond, Indiana.
JANIS, C. M. 1993. Tertiary mammal evolution in thecontext of changing climates, vegetation, andtectonic events. Annual Review EcologicalSystems, 24, 467-500.
JANIS, C. M., DAMUTH, J. & THEODOR. J. M. 2000.Miocene ungulates and terrestrial primaryproductivity: where have all the browsers gone?Proceedings of the National Academy of ScienceUSA, 97, 7899-7904.
KERR, J.T. & PACKER, L. 1997. Habitat heterogeneityas a determinant of mammal species richness inhigh-energy regions. Nature. 385. 252-254.
KINGDON, J. 1990. Arabian Mammals. A NaturalHistory. Academic Press, London.
LATHAM, R. E. & RICKLEFS. R. E. 1993. Globalpatterns of tree species richness in moist forests:energy-diversity theory does not accountfor variation in species richness. Oikos. 67.325-333.
LITTLE, E. L. 1977. Atlas of United States Trees. UnitedStates Department of Agriculture Forest Service.United States Government Printing Office.Washington. DC.
LOTTES. A. L. & ZIEGLER. A. M. 1994. World peatoccurrence and the seasonality of climate andvegetation. Palaeogeography, Palaeoclimatology,Palaeoecology, 106. 23-37.
LYELL, C. 1830. Principles of Geology, being an attemptto explain the former changes of the Earth'ssurface, by reference to causes now in operation,vol. 1. John Murray. London.
MAC-ARTHUR, R. H. & WILSON. E. O. 1963. An equi-librium theory of insular zoogeography. Evol-ution. 17. 373-387.
MACARTHUR, R. H. 1967. The Theory of Island Bio-geography. Princeton University Press. Princeton.NJ.
MARKWICK. P. J. 1994. 'Equability', continentality andTertiary 'climate': the crocodilian perspective.Geology, 22. 613-616.
MARKWICK, P. J. 1996. Late Cretaceous to Pleistoceneclimates: nature of the transition from a 'hot-house' to an 'ice-house' world. PhD Thesis. Uni-versity of Chicago.

BIODIVERSITY AND CLIMATE 199
MARKWICK, P. J. 19980. Fossil crocodilians as indi-cators of Late Cretaceous and Cenozoic climates:implications for using palaeontological data forglobal change. Palaeogeography, Palaeoclima-tology, Palaeoecology, 137, 205-271.
MARKWICK, P. J. 1998b. Crocodilian diversity in spaceand time: the role of climate in paleoecology andits implication for understanding K/T extinctions.Paleobiology, 24, 470-497.
MATTHEW, W. D. 1915. Climate and evolution. Annalsof the New York Academy of Science, 24,171-318.
MULLER, M. J. 1982. Selected climatic data for a globalset of standard stations for vegetation science.Tasks for vegetation science. Dr W. Junk Publish-ers, The Hague.
O'BRIEN, E. M. 1993. Climatic gradients in woodyplant species richness: towards an explanationbased on an analysis of southerm Africa's woodyflora. Journal of Biogeography, 20,181-198.
OSTROM, J. H. 1970. Terrestrial vertebrates as indi-cators of Mesozoic climates. Proceedings of theNorth American Paleontological Convention,347-376.
PIANKA, E. R. 1966. Latitudinal gradients in speciesdiversity: a review of concepts. The AmericanNaturalist, 100, 33-6.
PIANKA, E. R. 1981. Diversity and adaptive radiationsof Australian desert lizards. In: KEAST, A. (ed.)Ecological Biogeography of Australia. Dr W. JunkPublishers, The Hague, 1377-1392.
QIAN, H. & RICKLEFS, R. E. 2000. Large-scale pro-cesses and the Asian bias in species diversity oftemperate plants. Nature, 407,180.
REDFORD, K. H. & EISENBERG, J. F. 1992. Mammals ofthe Neotropics. The Southern Cone. Volume 2.Chile, Argentina, Uruguay, Paraguay. Universityof Chicago Press, Chicago.
ROHDE, K. 1992. Latitudinal gradients in species diver-sity: the search for the primary cause. Oikos, 65,514-527.
ROHDE, K. 1997. The larger area of the tropics doesnot explain latitudinal gradients in species diver-sity. Oikos, 79,169-172.
ROHDE, K., HEAP, M. & HEAP, D. 1993. Rapoport'sRule does not apply to marine teleosts and cannotexplain latitudinal gradients in species richness.The American Naturalist, 142,1-16.
ROOT,T. L. & SCHNEIDER, S. H. 1993. Can large-scaleclimatic models be linked with multiscale eco-logical studies? Conservation Biology, 7,256-270.
ROSENZWEIG, M. L, 1995. Species Diversity inSpace and Time. Cambridge University Press,Cambridge.
ROY, K., JABLONSKI, D. & VALENTINE, J. W. 1994.
Eastern Pacific molluscan provinces and lati-tudinal diversity gradient: no evidence for'Rapoport's rule'. Proceedings of the NationalAcademy of Science U.S.A., 91, 8871-8874.
SCHAAL, S. & ZIEGLER, W. 1992. Messel. An Insightinto the History of Life and of the Earth. Claren-don Press, Oxford.
SEPKOSKI, J. J. JR. 1976. Species diversity in thePhanerozoic: species-area effects. Paleobiology,2, 298-303.
SEPKOSKI, J. J. JR. 1996. Competition in macro-evolution: the double wedge revisited. In:JABLONSKI, D., ERWIN, D. H. & LIPPS, J. H. (eds)Evolutionary Paleobiology: Essays in Honor ofJames W. Valentine. University of Chicago Press,Chicago, 211-255.
STEBBINS, R. C. 1985. A Field Guide to WesternReptiles and Amphibians. Houghton MifflinCompany, Boston.
STEVENS, G. C. 1989. The latitudinal gradient in geo-graphical range: how so many species coexist inthe tropics. The American Naturalist, 133,240-256.
STEVENS, G. C. 1992. The elevational gradient inaltitudinal range: an extension of Rapoport'slatitudinal rule to altitude. The AmericanNaturalist, 140, 893-911.
STRAHAN, R. 1992. Encylopoedia of AustralianAnimals. Mammals. Angus and Robertson,Pymble, NSW.
TER BRAAK, C. J. F. 1987-1992. CANOCO - aFORTRAN program for Canonical CommunityOrdination. Microcomputer Power, Ithaca, NY.
UHL,N. W. & DRANSFIELD, J. 1987. Genera Palmarum.A Classification of Palms Based on the Work ofHarold E. Moore, Jr. Allen Press, Lawrence.Kansas.
VAN DER BRINK, F. H. 1967. A Field Guide to theMammals of Britain and Europe. Collins.London.
WALLACE, A. R. 1876. The Geographical Distributionof Animals, vol. 1. MacMillan, London.
WILDE, V. 1989. Untersuchungen zur Systematik derBlattreste aus dem Mitteleozan der GrubeMessel bei Darmstadt (Hessen, BundesrepublikDeutschland). Courier Forschungsinstitut Senck-enberg, 115.
WRIGHT, D. H., CURRIE, D. J. & MAURER, B. A. 1993.Energy supply and patterns of species richness onlocal and regional scales. In: RICKLEFS, R. E. &SCHLUTER, D. (eds) Species Diversity in Eco-logical Communities. Historical and GeographicalPerspectives. University of Chicago Press.Chicago, 66-74.

This page intentionally left blank

Index
Page numbers in italics refer to Figures and page numbers in bold refer to Tables.
acanthodians 74, 75, 77, 78accuracy and database management 172,173acrotetides 26Actinodonta 41Actinostereon gregareum 133age of fossils, reliability assessment 173,174Allonychia 43Aloconconcha 38alpha diversity 2, 99Amadeus Basin 72Ambonychia 43Ambonychiopsis 43Amorphognathus Biofacies 90amphibian diversity 183-187,188,188,189,191, 792,
193analytical time averaging 174Ananterodonta 41anaspids 74, 75, 78Anatolepis 70, 72, 76, 79Andean Basin Jurassic bivalve populations
diversity 128,134-136effect of Hispanic Corridor 131-134extinction rates 729,130immigration rates 131
Angarella 19angiosperms 1, 5, 5-6
Cenozoic biodiversity 158Cretaceous radiation 141,143-144
Antarctic Peninsula 145,146-147Anomalocoelia 43Anomalodesmata 44-45Anomalodonta 43anoxic events and planktonic foraminifera 122Antarctic Peninsula
Cretaceous flora 144-147plate setting 141-142role in floristic radiation 147-149
Antarctica, geographic isolation 154-156Anti-Atlas (Morocco) bivalves 35, 36Apatobolus micula 101, 102Apedolepis 72Aphelognathus-Oulodus Biofacies 90Appalachignathus 90Arabia, species diversity 183aragonitic foraminifera 121Arandospis 69Area spp. 37, 40Arcomytilus pectinatus 133Arctic Ocean isolation 156Arctochedra spp. 16,17Arenigomya 44Armorica and rynchonelliformean brachiopods 75Asaphopsis province 53Asaphopsis-Taihungshania-Neseuretus Fauna 55ash deposition effects 4,101-104
ostracode case study 106,107,108-111Astraspis 11athyridides 27Atlantic, Caribbean and East Pacific (ACEP) realm
153,160,163,164atrypides 27Australia 63,183Australia/New Guinea block collision 160-162Autolamellibranchia 39-45Avalonia 19
biodiversity 86, 92, 95brachiopod diversity profiles 31conodonts 88, 89, 91Ordovician bivalves 36Ordovician palaeogeography 28palaeolatitudes 95
Babinka 41-42Baltica
biodiversity 92, 95conodont provinces 90Ordovician 18-19,19-20,25rynchonelliformean brachiopods 75,18
Baltoscandian conodont provinces 86Bambachian megaguilds 25Bavarilla hofensis 56, 58Belodella 90Belodina 90Bennettitales 144, 145beta (between habitat) diversity 2, 99,157Billingsella Association 75,16, 77, 20biodiversity in Cenozoic 154,158-164biofacies
architecture 88-91, 93-95biogeographic cladogram, trilobites 61,62biogeography, Palaeozoic 53-54, 76-78biotic provincialism 1birds radiation 1bivalve molluscs 35-36
Andean Basin - NW Europe studydatabase and analytical methods 128-130results 128,130,131-134
heteroconch evolution 5Mesozoic-Cenozoic diversity 154Ordovician 4,105
Autolamellibranchia 39-45Protobranchia 36-39
Black Mountain eustatic event 20Bohemia, rynchonelliformean brachiopods 75Bornholm 90-91Brachilyrodesma 40brachiopods
Ordoviciandiversity 105

202 INDEX
mid Wales range chart 110radiation 19-20
rhynchonelliformean 4,13,15,16, 26Cambrian 14-16distribution 14,15migration potential 19Ordovician 16-18, 25, 26-27, 29-31radiation and paiaeogeography 19-20
Breviorthodonta 45Bryantodina 89bryophytes 145bryozoans 5,19,26,101,105Burgess Shale Lagerstatte 72Burj Formation 16
Cadomia 39Calymenacean-Dalmanitacean Province 53Calymenella 56, 58, 62, 63Calymenesun spp. 56,57, 58, 63Cambrian
bivalves 35, 45diversity decline 26mid-late period sea level 19paiaeogeography 70rynchonelliformean brachiopods 14-16skeletonized fauna 13vertebrates 69, 72, 74, 76, 78-79
Cambrian Evolutionary Fauna 13,14, 26cameralloideans 18,19Camnantia 42Camptonectes laminatus 133carbonate deposition in Ordovician 20Cardiolaria 39Cardoilarioidea 39 -0Caribbean Cenozoic coral diversity 160,163Carminodonta 41Carotidens 43Catamarcaia 42Celtic faunas 27, 28, 29Celtoconcha 42Cenozoic biodiversity and climate 154-164,160-163Central American Isthmus (CAI) uplift 156,163-164Central Asia, rynchonelliformean brachiopods 15, 18Cephalochordata 74, 75Ceratomya concentrica 133cheilostome bryozoan, evolution 5Chengyiang Lagerstatte 72Chile, Jurassic bivalves 132China fauna 63, 75, 77, 86chondrichthyans 74, 75, 78Cimmeria 61cladograms, Reedocalymeninae 60, 61, 62, 64Clarkella Fauna 15, 17, 18, 20Classopolis 148Claueonychia 43Clavatipollenites hughesii 148Cleionvchia 43climate and biodiversity 4, 154-157, 162-165, 179,184clitambonitidines 18, 19Colpantyx 44Colpocoryphe 55Compactogerina stellapolaris 122Coniferales 145
Coninae 5conodonts 4, 70, 72, 73, 74, 75, 76
see also euconodontsConoglobigerina 118, 121continental breakup and radiation 1, 6Copidens 41Corallidomus 44corals, Cenozoic diversity 160-163Cosmogoniophorina tenuicostata 44Coxiconcha 42craniformeans 26Cretaceous
floristic radiation 143-144,145, 146-149Gondwana breakup 141-143ocean circulation 759planktonic foraminifera 115. 116, 117,120,121,
122-123Cretaceous-Recent biodiversity 1. 5-6. 7Creux de Fours 118crocodilians 176, 179Crozonaspis 54crustacean evolution 5Cryptolichenaria 19Ctenodonta 38Ctenostreon rugosum 133Citneamya 44. 45Curtognathiis 90Cycloconcha 41,42Cymatonota 45Cymbithyris 16Cyrtodonta 43Cvrtodontula 43cyrtomatodonts 26-27
Dacryomya lacrvma 133Dactiloceras 118Dalmanellidina 26databases 6, 180-182
application 175-177use in diversity analysis 182-190use in palaeoclimate analysis 190-197
scale 172-175structure 169-172taxonomy 175
decapod evolution 5Deceptrix 37delta diversity 2deltidoidonts 26-27Denticelox 43Derenjal Formation 16, 77Devonian fauna 2, 25, 78. 79Diaphelasma 18differentiation diversity 2Diraphora Association 75. 16discinoideans 26dispersal potental 19Distomodus 92, 93diversification and Ordovician volcanism 99-100,
103case study 106, 107. 108-111ecology 101-103gene pool 103-104
Dob's Lin 86

INDEX 203
Drake Passage opening 156Dulcineia 41
East Uralian microcontinent 19echinoderms and Ordovician diversity 18, 26,105Ectillaenus 55Ekaterodonta 38energy flux and diversity 180,195Entolium corneolum 133Eocene
Messel Shale 190-191,195ocean circulation 155, 159
Eocene-Oligocene ocean cooling 154,156Eohomalonotus 54Eopecten spondyloides 133epsilon diversity 2Eridodychia 43Erismodus 90Eritropis 39-40escalation, Cenozoic 158euconodonts 72, 73, 76
presence-absence matrices analysis 86-95Euramerican vertebrate phylogeny 75Europe 183
Jurassic bivalve populationdiversity 128,136-138effect of Hispanic Corridor 131-134extinction rates 129,130immigration rates 131
extent defined 172extinction events
late Ordovician 78Pliensbachian-Toarcian 127
methods of analysis 128-130results 130-131testing hypothesis 131-134
extinction rates in bivalves 128-129,130,131-133
Falcatodonta 43Favusella 120,122Fidera 36fish, teleost 5flat file format 169,170floristic development in Cretaceous 1, 5, 5-6,
143-144,145,146-147role of Antarctic Peninsula 147-149role of global warming 149
foraminifera (planktonic) 5evolution 121-123Jurassic-Cretaceous distribution 118,120,121Mesozoic development 115,116,117origins 118-121
Fordilla 35
galeaspids 74, 75,11, 78gamma diversity 2, 99genetic diversity and ash falls 103-104geographical range, role of 3geograpic information systems (GIS) 169-170Gervillaria daeformis 133ghost ranges, vertebrates 76, 78
glaciation 4, 86glacioeustasy and diversity 163global warming and floristic radiation 149Globigerinina 122Globuligerina spp. 122Glyptarca 39, 40, 42Glyptonichia 43Glyptoria spp. 16,17Glyptotrophia 18Gondwana
area cladogram data 61, 63brachiopods 75,16,18, 20,31breakup 141-143Cambro-Ordovician faunal replacement 20floristic provincialism 146,148Ordovician bivalves 36Ordovician palaeogeography 28trilobites 53-54vertebrates 74,77
Goniophorina tenuicosta 44grain 172,173-175graptolite blooms 101,102graptoloids, Ordovician diversity 105Great Basin of Laurentia, brachiopod radiation 25,26Gresslya peregrina 133gymnosperms 145
habitat islands 179-180Haeuslerina helvetojurassica 122Haikouella 72Haikouichthys 72Hamarodus-Dapsilodus-Sabbardella (HDS)
Biofacies 90Hanchungolithus 55Harding Sandstone 69,70, 72Harpoceras 118Hemiprionodonta 40, 42Hepatophytes 145Heteroconchia 40-42Heterodonta 40heterostracans 77, 78Heterostraci 74, 75Hirnantian glaciation 4Hispanic Corridor 5,128
effect on immigration of bivalves 129,131-133,134Homilodonta 37Huenellaspp.17,18Huenella-Palaeostrophia Association 15
lapetus Ocean 27-28, 28-29, 71, 86Iberocoryphe 54Icriodella 92, 93immigration of bivalves 129,131
role of Hispanic Corridor 131-133Inaequidens 40Indo-Pacific gateway 160,161Indo-West Pacific (IWP) realm 153,160,164,165
glacioeustasy and diversity 163Miocene coral diversity 160-162
Inopema sowerbyana 133inventory diversity 2island biogeography theory 179

204 INDEX
Jamesella 16Johnmartinia 39Jurassic
bivalve study in Andean Basin-NW Europedatabase and methods of analysis 128-130results 128,130,131-134
planktonic foraminifera 115,116,117distribution 118,120evolution 121-122origins 118-121
Pliensbachian-Toarcian extinctions 127,128
Kerfornella 54Kerguelen Plateau 143Koagash Formation 17Kockelella 92Kutorginata 14-16Kutorginide Fauna 15,16Kutorina 16Kyjandy Formation 16
Lamellodonta 35land bridges 5latitude and biodiversity 153,154Laurentia
biodiversity 86, 91, 95conodonts 86, 87, 89, 91lapetus margin 71Ordovician
brachiopod radiation 15,18,19-20faunal replacement 18-19palaeogeography 28
vertebrates 74, 76, 77Leconychia 43Lingulide Fauna 15linguliformeans 26linguloideans 26Liomegalaspides 54lithofacies preservation potential 78-79Llangynog (Wales), bivalves 36Lophoconcha 39lycophytes 145Lvrodesma 40
mammals 1, 5,183,187-188,186,189,190,191-192Marnes de Villers 122Maryonychia 43Merlinia 54Mesozoic-Cenozoic boundary and biodiversity 1,
5-6,7Miaopu Formation 63Messel Shale 190-191,195Microcachyridites antacticus 148Midcontinent Province, conodonts 86migration potental 19Milankovitch cyclicity 6,162-163Miocene
coral diversity 160-162ocean circulation 755, 759
Miocene-Pliocene, global cooling 154Modioliodon 44
Modiolopsis 44Modiolus imbricatus 133Modiomorpha 44molluscs 163
see also bivalve molluscsmongolepids 72, 76, 77Montagne Noire bivalves 35. 36Moridunia 41Moyeronia 19Myllokunmingia 72Myodakryotus 43Myoplusia 39Myxinoidea 74, 75, 76
Na Mo Formation 63Natasia 38negative difference vegetation index 185,186.187.
188,190neogastropod evolution 5Neogene climate change 162-165Neseuretinus turcicus 58Neseuretus spp. 54, 56,57, 58, 62. 63Neseuretus Association 54-56Nicaniella voltzii 133Nileus 63Nimra Formation 16Noradonta 40North America species diversity 183North Atlantic Deep Water (NADW) 156North Atlantic Province (NAP) conodont biofacies
88-89Nuculana ovum 133Nuculites 39Nuculoida 36-39
Obelellata 14-16Oberhauserella quadrilobata 118.119Obolid Fauna 75ocean circulation patterns 154-156. 759ocean gateways 5Ogyginus 54Old Red Sandstone continental assembly 70. 71Olenty Formation 18Opisthoptera 43Opsiconidion nudum 101orbital forcing and range dynamics 163Ordovician
biogeography and biodiversity 3-5bivalves 35-45. 46, 47brachiopods 16-18
development in lapetus 29diversification 26-27diversity profiles 30, 31radiation 19-20
carbonate deposition 20diversification 85, 99-100. 105
effect of ash on 101-104. 106, 107,108-111euconodonts presence-absence matrices analysis
86-95faunal replacement 18-19glaciation 86palaeoplate faunal exchange 13

INDEX 205
Ordovician (continued)plate reconstructions 37,55, 70radiation 1,7, 25-26sea-level change 19, 20,30tectonic activity 19trilobites 99, 53-54vertebrates 69-70, 72-73, 74, 76-77, 78-79
organophosphatic forms 26Orthida 26Orthide-Polytoechoidean Association 15Orthide-Syntrophiodean Association 15orthides, endopunctate 18,19Orthidina 26orthotetides 16Ortondla 43Osteichthyes 74, 75osteostracans 74, 75, 77, 78ostracoderm 72, 76-79ostracodes and Ordovician volcanism 106,107,
108-111Oulodus 91, 92oxygenation and ash falls 102,103Ozarkodina 92
palaeoclimate and diversity 190-197Palaeoconcha 36palaeogeography
early Palaeozoic 37, 55, 70-71, 70role in radiation 1, 6
Palaeoheterodonta 40palaeolatitude and planktonic foraminifera 122-123Palaeoneilo 39palaeontological uncertainty principle 174Palaeonucula hammeri 133Palaeopteria 43Palaeostrophia 18Palaeozoic Evolutionary Fauna 13,16,18,19, 26Parallelodon hirsonensis 133Pamllelodus 44Paramytiiarea 43parental imprinting 104Paulinea 36pelromyzontids 76Pensarnia 36Pentamerides 26, 27Perunica. brachiopod diversity profiles 31Petromyxinoidea 74, 75Pharcidoconcha 43Pharostomina oepiki 56, 58Pholadornya fi ducula 133Phragmodus 90Phragmodus-lcriodella-Plectodina Biofacies 90phylogenetic analysis, trilobites 56-61phylogeny, vertebrates 73-76Pipiscius 72Pirchanchaspis rinconensis 72-73Pituriaspida 74, 75placoderms 74, 75, 77, 78Plaesiacomia 54Plagiostoma rodburgensis 133planktonic blooms 102,103planktonic foraminifera see foraminiferaplate reconstructions see palaeogeography
plate tectonics and biodiversity 4plectambonitiodeans 18,26Plectodina 90Plegagnathus 91Plethocardia 43Pliensbachian-Toarcian extinctions 127,128Pliocene ocean circulation 157Pojetaia 35Polyplacognathus 90polytoechiides 18polytoechinoideans 16Porophoraspis 69, 72Posidonia 118Posidonotis semiplicata 132potental evapotranspiration (PET) 180,185,191, 796Pradoella spp. 58, 62Praeanomalodonta 43Praegubkinella spp. 118,119,120Praenucula 36precipitation and diversity 184,186,187,188,190precision defined 172predator pressure 132presence-absence matrices analysis 86-95Prolobella 43Propeamussium pumilum 132Protambonites 17,18Prothyris 45Protobranchia 36-39Protocalymene mcallisteri 56,58Protopanderodus 90Protopanderodus-Periodon Biofacies 86Protorthide Fauna 15,16Protorthis 16provinciality changes 2Pseud arc a 40Pseudolimea duplicata 133Pseudolonchodina 92Pseudopecten barbatus 133Psiloconcha 45Psilonychia 43Psiloria 16Pteridophytes 145Pteriomorphia 42-44Pteronychia 43Pterospathodus 92pyroclastics see ash
radiation and diversity 184,186,187,188,190Ramsey Island (Wales), bivalves 36Rapoport's rule 180Redonia 41Reedocalymeninae 4, 56-61relational databases 169,170reptile diversity 183,186,187,187,191,192,193resolution see grainRhipidognathus 89rhynconelliformean brachiopods see under
brachiopodsRhytimya 44
Sacabambaspis 69Salta Province (Argentina) bivalves 35, 36

206 INDEX
Salterocoryphe 55Sarrabesia spp. 58, 62, 63scale problems in databases 172-175sea level
change and biodiversity 4-5,163,165Palaeozoic 19, 20,30relation to tectonism 104
Selenopeltis Province 53Semicorallidomus 44Shirgesht Formation 17,18Siberia
Cambrian fauna 16conodont provinces 86early Palaeozoic 71, 75, 77Ordovician fauna 18-19,19-20,28
Siberoctenia 38siliciclastic input and diversity 4Silurian
diversity 92-93endemisrn and cosmopolitanism 79palaeogeography 70provincialism 2vertebrates 73, 74, 77-78
Similodonta 38siphonotretides 26skeletonized fauna 13Skiichthys halsteadi 69snakes radiation 5Solemyoida 39Solenomorpha 45Sort Tepe Formation 63South Africa species diversity 183South America 63,148,183spiriferides 27Stairway Sandstone 69Sthenodonta 39stratigraphic reliability codes 174stromatoporoid reefs 25Strophomenata 16Strophomenida 26strophomenides 16,19sympatric speciation 104Synchomalonotus 62Synhomalonotus 55, 56synonymization 175syntrophiidines 18syntrophioideans 18,19
Taconic Orogeny 4Taihungshania 54Tancrediopsis 38taphonomic time averaging 174taxonomy, role in database management 175tectonism, role in radiation 1, 6teleost fish evolution 5temperature and diversity 184,186,187,188,190,192,
197Tethyan Ocean closure 156,160Tetralobula 18tetrapod diversity 182-183,185,186,197thelodonts 72, 74, 75, 76, 77, 78Thorslundia 43Thysanotos-Leptembolon Fauna 15Tironucula 38
Toquima-Table Head faunas 27, 28, 29Tornquist Sea closure 71Trecanolia 42tree diversity 189Tremadoc 19, 20-21Trematobolus 16Trematosia 16Trigonioida 40Trigonoconcha 38trilobites
asaphide 18Cambrian Evolutionary Fauna 26Neseuretus Association 54-56Ordovician 4
diversity 99,105provinciality 53-54
phylogenetic analysis 56-61triplesiides 19Tritoechia 18Tritoechia-Protambonites Association 15Tromelinodonta 40Tuarangia 35Tunicata 74, 75Turkey, trilobites 61, 62, 63Turukhansk-Irkutsk facies 16Tuva, vertebrates 75, 77Tuvaella 11
uncertainty principle 174Uralian margin uplift 19, 20. 21Uskardita 42
Vanuxemia 43Veneridae evolution 5vertebrates
biogeography 76-78early Palaeozoic 69-70, 72-73phylogeny 73-76
Vietnam, trilobites 63Vietnamia douvillei 57, 58, 63Villicumia 38volcanism and diversity 4,19,101-104
ostracode case study 106,107, 108-111
Warburgia 43Welsh Basin 19, 99,101
ostracode development 108-111Whiterock Fauna 4, 26within-habitat diversity see alpha diversity
Xestoconcha 44Xidazoon 72
Yangtze Platform, trilobites 63Yunnanozoon 12
Zadiemrodia fastigata 41Zeehania 39Zeliszkella 55

















