This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
The translational value of rodentgastrointestinal functions:a cautionary taleGareth J. Sanger1, Joanna D. Holbrook2 and Paul L.R. Andrews3
1 Wingate Institute of Neurogastroenterology, Blizard Institute, Barts and The London School of Medicine and Dentistry,
Queen Mary University of London, 26 Ashfield Street, London, E1 2AJ, UK2 Singapore Institute for Clinical Sciences, Agency for Science Technology and Research (A*STAR),
Brenner Centre for Molecular Medicine, 30 Medical Drive, 117609 Singapore3 Division of Biomedical Science, St George’s University of London, Cranmer Terrace, London, SW17 0RE, UK
Understanding relationships between gene comple-ments and physiology is important, especially wheremajor species-dependent differences are apparent. Mo-lecular and functional differences between rodents (rats,mice, guinea pigs) and humans are increasinglyreported. Recently, the motilin gene, which encodes agastrointestinal hormone widely detected in mammals,was found to be absent in rodents where the receptorsare pseudogenes; however, actions of motilin in rodentsare sometimes observed. Although ghrelin shares com-mon ancestry with motilin, major species-dependentabberations are not reported. The apparently specificabsence of functional motilin in rodents is associatedwith specialised digestive physiology, including loss ofability to vomit; motilin is functional in mammals capa-ble of vomiting. The exception is rabbit, the only othermammal unable to vomit, in which motilin might beconserved to regulate caecotrophy, another specialiseddigestive process. Motilin illustrates a need for cautionwhen translating animal functions to humans. Never-theless, motilin receptor agonists are under develop-ment as gastroprokinetic drugs.
Species-dependent variations in gastrointestinalfunctions and molecular pathwaysGastrointestinal functions in different mammalian speciesare broadly similar: we all have a need to eat food, digestand extract nutrients, and excrete the waste. However,marked differences exist in anatomy and controllingmechanisms, perhaps arising during evolution as a resultof different diets and living environments [1]. This appliesparticularly to rodents (exemplified by rats, mice, guineapigs). Compared with humans, differences in functionaland molecular pathways controlling gastrointestinal (GI)functions for rodents are increasingly reported (Table 1);such differences potentially compromise translation ofexperimental data to the clinic unless properly understood.A recent example is provided by reports that cDNAs withintact open reading frames (consistent with functionalstatus) for the GI hormone motilin (and its seven trans-membrane, G protein-coupled receptor, GPCR) are found
in the human, tree shrew, dog, cow, pika and rabbit, but notin rodents, such as the rat, mouse, guinea pig and squirrel[2]. It is now important to understand why this differenceexists, how it impacts the physiological functions of the gutand how it changes the translational value of rodents andother mammals as models of human GI pathophysiology.The importance is exemplified by the increasing interest indeveloping motilin receptor agonists as drugs for GI dis-orders and in understanding the actions of the antibioticdrug erythromycin, also a motilin receptor agonist [3].
Any analysis of the functions of motilin must makecomparisons with ghrelin, because similarities in peptideand precursor structures and similarities in receptor se-quence identity place these two hormones in the samefamily group [4]. Consequently, species-dependent similar-ities and differences between the hormones provide addi-tional information on functions. The five transmembranereceptor splice variants and additional peptides from pro-hormones of ghrelin (des-acyl ghrelin and obestatin) [5] arenot included in this comparison, given the limited infor-mation on their functions.
Motilin and ghrelin receptors: shared functions andcommon ancestry?Further justification for considering motilin and ghrelintogether is provided by several observations which suggestcommon ancestry. The motilin and ghrelin receptor genesare located, respectively, on human chromosomes 13q14.2and 3q26.31. However, unlike most family A GPCRs whichare encoded within one exon [6], the ghrelin and motilinreceptors are encoded within two exons, with no untrans-lated exons. For both, exon 1 is the larger (901 and 796nucleotides, respectively), encoding transmembranedomains 1–5. Exon 2 is smaller (333 and 302 nucleotides,respectively), encoding transmembrane domains 6 and 7.This shared unusual two exon gene structure is consistentwith both receptors deriving from a common ancestor morerecently than other family A GPCRs.
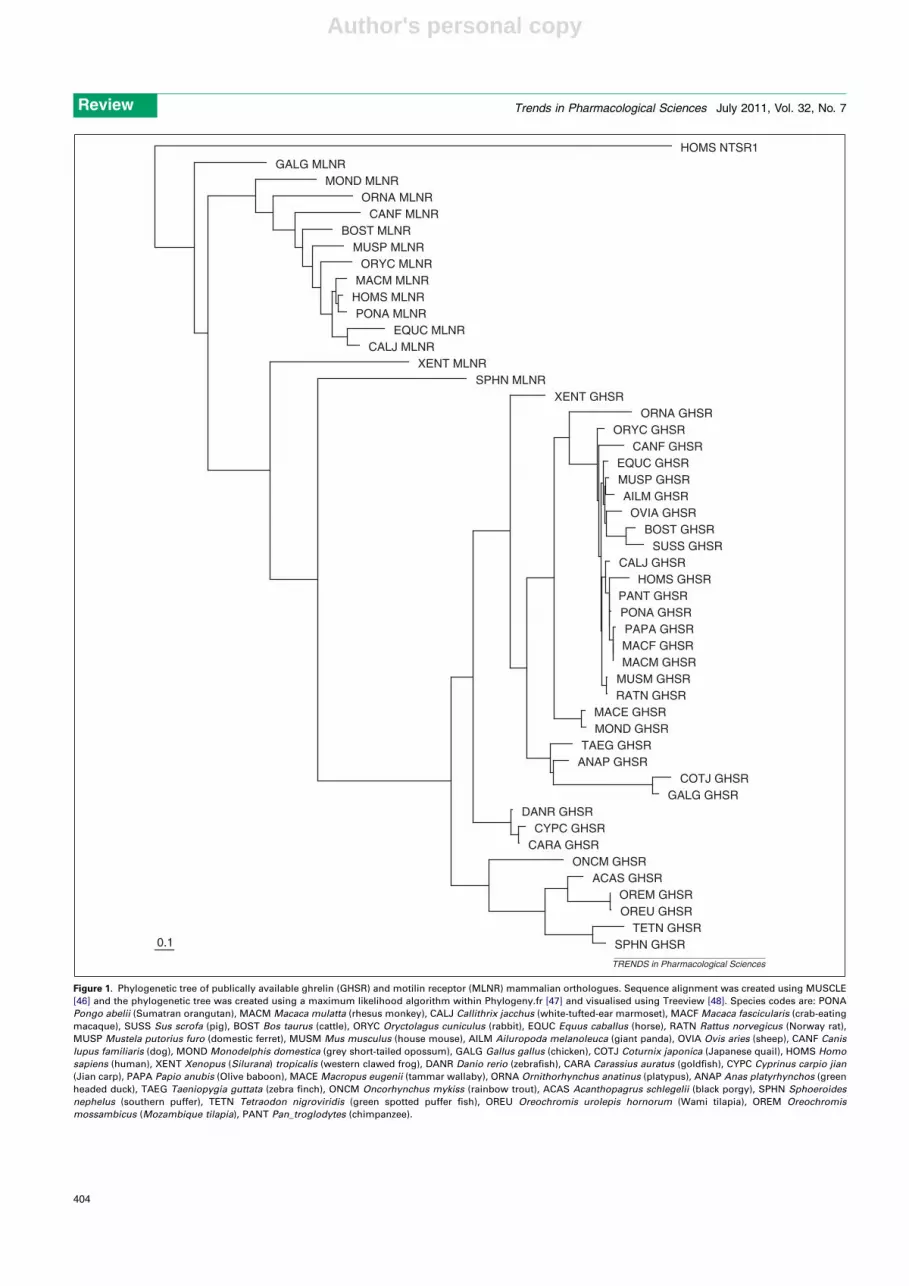
Secondly, all publically available mammalian motilinand ghrelin receptor peptide sequences align easily andcan be represented in a phylogenetic tree (Figure 1). Thisshows sequences for both receptors forming distinct clades,
402 0165-6147/$ – see front matter � 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.tips.2011.03.009 Trends in Pharmacological Sciences, July 2011, Vol. 32, No. 7
Author's personal copy
being more similar to each other than any other GPCR (e.g.the paralogue neurotensin receptor 1). To date, this highhomology has not been linked by shared sensitivity toendogenous ligands, as each receptor is activated by dis-tinct peptides (at least in humans and rabbits [7]). Never-theless, the clear homology between the receptors suggestscommon ancestry and an evolutionary constraint whichhas prevented significant divergence. Such restraintappears to be consistent with sharing of some GI functions.In humans, for example, both hormones are found espe-cially in gastro-duodenal endocrine cells. Furthermore, byacting within the enteric nervous system and/ or on thevagal innervation of the gut, both motilin and ghrelin canincrease gastric emptying of meals and both are able tomimic the burst of propulsive activity which occurs in theupper gut during fasting (known as phase III of the mi-grating motor complex, MMC) [3,4]. A major differencebetween the hormones is that ghrelin is also found in manynon-GI regions, where it exerts additional functions suchas those relating to appetite and metabolism [4,5].
Conservation of ghrelin throughout evolutionTo date, there are no mammalian species in which ghrelinor its receptor has been found to be absent [8] (Figures 1and 2). All ghrelin peptide sequences form a single cladeand cluster in approximate taxonomic order (Figure 2),suggesting functional constraint. Included are rats, mice,dogs and humans, where an ability of ghrelin to stimulategastric emptying of food and/or phase III of the MMC hasbeen demonstrated [3,4]. It seems reasonable to suppose,therefore, that the diverse actions of ghrelin [3–5], affecting
feeding, appetite, metabolism and endocrine control,together with gastric prokinetic activity, provide reasonsfor constraint on its evolution.
Loss of motilin function is unique to rodentsThe common ancestry and the preservation of a functionalghrelin system in all mammalian species make thereported absence of motilin receptors in rodents, such asthe mouse, rat and guinea pig, particularly striking [2]. Todate, it has not been possible to clone the cDNA or detectthe motilin gene in a variety of different rodent species(including mouse, rat, guinea pig and squirrel), althoughan apparently functional motilin gene (defined by an openreading frame) has been detected in one rodent, namely thekangaroo rat. Nevertheless, the motilin receptor gene hasso far only been identified as a pseudogene in genomic DNAsequence in rodent species, including the kangaroo rat,containing frameshifts and stop codons which preventtranslation of the receptor protein. Variations in the mech-anism by which motilin function has been disrupted in thedifferent rodent species might reflect different time pointsat which disruption occurred during the evolution of theseanimals towards a digestive system which does not requirethe functions of motilin [2].
Figure 2 shows alignments of the molecular forms ofmotilin and the phylogenetic tree. It can be seen thatmotilin is generally conserved, with 59–95% sequenceidentity, apart from the absence of sequences for mouseand rat motilin. In these animals, mutation of the motilingene to a pseudogene presumably occurred because ofrelaxation of functional constraint. A similar picture
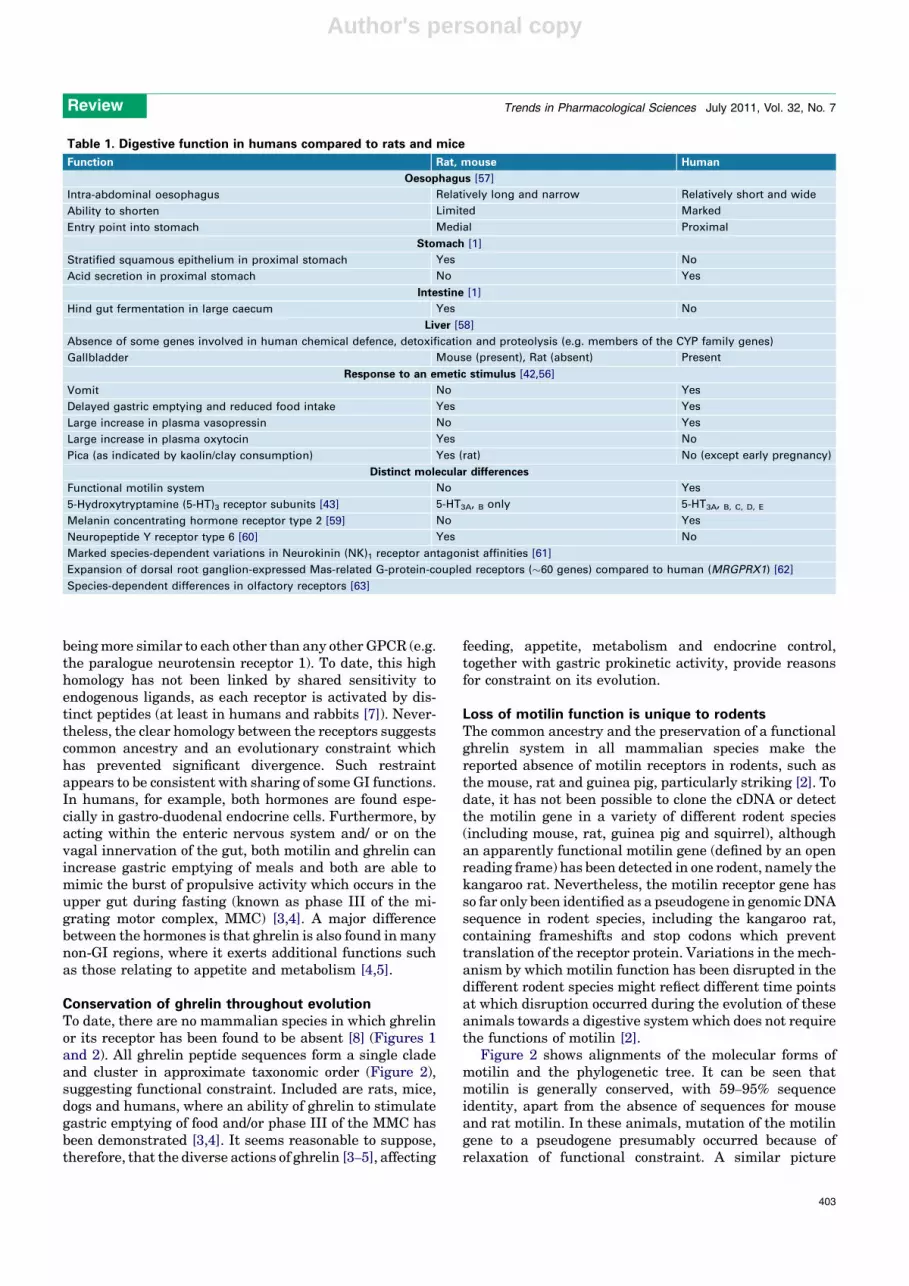
Table 1. Digestive function in humans compared to rats and mice
Function Rat, mouse Human
Oesophagus [57]
Intra-abdominal oesophagus Relatively long and narrow Relatively short and wide
Ability to shorten Limited Marked
Entry point into stomach Medial Proximal
Stomach [1]
Stratified squamous epithelium in proximal stomach Yes No
Acid secretion in proximal stomach No Yes
Intestine [1]
Hind gut fermentation in large caecum Yes No
Liver [58]
Absence of some genes involved in human chemical defence, detoxification and proteolysis (e.g. members of the CYP family genes)
Gallbladder Mouse (present), Rat (absent) Present
Response to an emetic stimulus [42,56]
Vomit No Yes
Delayed gastric emptying and reduced food intake Yes Yes
Large increase in plasma vasopressin No Yes
Large increase in plasma oxytocin Yes No
Pica (as indicated by kaolin/clay consumption) Yes (rat) No (except early pregnancy)
Distinct molecular differences
Functional motilin system No Yes
5-Hydroxytryptamine (5-HT)3 receptor subunits [43] 5-HT3A, B only 5-HT3A, B, C, D, E
Melanin concentrating hormone receptor type 2 [59] No Yes
Neuropeptide Y receptor type 6 [60] Yes No
Marked species-dependent variations in Neurokinin (NK)1 receptor antagonist affinities [61]
Expansion of dorsal root ganglion-expressed Mas-related G-protein-coupled receptors (�60 genes) compared to human (MRGPRX1) [62]
Species-dependent differences in olfactory receptors [63]
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
Figure 1. Phylogenetic tree of publically available ghrelin (GHSR) and motilin receptor (MLNR) mammalian orthologues. Sequence alignment was created using MUSCLE
[46] and the phylogenetic tree was created using a maximum likelihood algorithm within Phylogeny.fr [47] and visualised using Treeview [48]. Species codes are: PONA
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
404
Author's personal copy
emerges for the guinea pig. Thus, a motilin receptor pseu-dogene was found in guinea pig genomic DNA sequence [2],and in silico searches of available guinea pig genomicsequences did not yield a closer match to the humanmotilin receptor sequence. Attempts to repeat the amplifi-cation of the reported guinea pig motilin coding sequence[9] (Figure 2) were also unsuccessful (unpublished obser-vations); the reasons for the different observations areunclear.
Translation from molecular to native functionTo properly understand the molecular data on the evolu-tion of motilin, a first step is to identify supporting, trans-lational evidence in native tissues. In this analysis, it isclear that in all species in which motilin and its receptorhave been identified, a motilin receptor agonist has beenshown to increase gastric motility. These include the hu-man and rabbit, which have been extensively studied [3],and also the dog [10], rhesus monkey (Macaca mulutta)
[11] and several other species not listed here. In somemammals motilin has been identified, and although thematching receptor has not yet been reported, motilin re-ceptor agonists have been shown to stimulate gastric orintestinal motility. Of many examples, three include thesheep [12], pig [13] and house musk shrew [14].
The absence of functional motilin receptors in mice andrats is consistent with several reports demonstrating aninability of motilin receptor agonists to stimulate GI mo-tility in these species (recent examples include [15,16]).However, these reports contrast with several others whichsuggest that, in mice and rats, motilin can stimulategastric motility or increase intracellular calcium in stom-ach muscle (recent examples include [17–19]), excite brainneurons, including the dorsal vagal nucleus, vagal moto-neurons and arcuate nucleus (recent examples include [20–
23]) or provoke feeding [24]. Similarly, the absence of afunctional motilin receptor in guinea pigs is consistentwith several reports which demonstrate an inability of
TRENDS in Pharmacological Sciences
Figure 2. Peptide alignment of publically available ghrelin and motilin mammalian orthologues are aligned using MUSCLE [46]. Phylogenetic analysis was conducted by
maximum likelihood algorithm within phylogeny.fr [47]. Rodent cDNA or EST sequences matching the motilin peptide were not found in GenBank. For both motilin and
ghrelin the Monodelphis domestica (opossum) orthologue sequence has been trimmed at the C terminus. Residues are shaded to denote positional homology, black for
identical residues in all orthologues and grey for identical residue shared by the majority of orthologues. Species codes are as in Figure 1 legend and additional codes are:
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
405
Author's personal copy
motilin receptor agonists to contract guinea pig isolatedstomach (e.g. [25]) but contrasts with other reports ofexcitatory activity in the myenteric plexus of guinea pigileum [26] and inhibition of myenteric ganglionic transmis-sion in the same species, using high nanomolar concentra-tions of motilin [27].
The reasons why some laboratories report activity inrodents with motilin (or with motilin receptor agonists) areunclear. One possibility is that the motilin receptor isabsent in some laboratory rodent strains but present inothers, perhaps acting at a still-functioning, only partlydegenerate receptor. Another possibility is that rodentghrelin receptors are responsive to motilin at high concen-trations. Thus, although rabbit and human ghrelin andmotilin receptors have poor affinity for the opposingligands [7], it is not known if rodent ghrelin receptorscan respond to porcine or human motilin. Finally, it shouldbe noted that certain motilin receptor agonists which havebeen derived from the highly complex macrolide structureof erythromycin (known as motilides) might have at leastsome affinity for the ghrelin receptor. One example is thecompound EM574, where even an IC50 value of 10 mM forthe ghrelin receptor (radioligand binding study) falls with-in the concentration range found after intravenous infu-sion of motilides in humans [28].
It is also unclear why motilin has apparently beendetected in rat tissues (recent examples include [18,29]).Conflicting results have been obtained with radioimmuno-assay or immunohistochemical techniques where the anti-sera were raised against synthetic porcine motilin. Forexample, the use of cloned porcine cDNA encoding pre-promotilin failed to reveal a homologous message in pigbrain regions expressing motilin-like immunoreactivity[30].
Evolution of specialised digestive physiology in rodentsMutation of motilin and its receptor to non-functionalpseudogenes provides a mechanism for loss of motilinfunctions in rodents, but not an explanation. Rodentsare undoubtedly well adapted to diverse environments,with an estimated 2277 living species contributing halfthe diversity of placental mammals [31]. Several reasonscontribute to this success, but we hypothesise that special-isation of rodent digestive physiology, which no longerrequires the functions of motilin, is likely to have madea contribution.
To test the hypothesis, key digestive functions must betracked across a wide range of mammalian orders. Given thewell established relationship between plasma motilin con-centrations and phase III MMC activity [3,4] it seemedreasonable to look for species-dependent variations inMMC activity. However, MMC activity has not been mea-sured in sufficient mammalian species, and where observa-tions exist, a dependence on experimental methodologies[32] make it impossible to draw firm conclusions. A simplerapproach is to look at the ability to vomit, the forceful oralexpulsion of gastric contents. Although the motive force inmammals is provided by the diaphragm and abdominalmuscles (see below) we consider it a gastrointestinalfunction because immediately prior to vomiting the gutundergoes preparatory motility changes (gastric relaxation,
retrograde giant contraction in the small intestine bothunder vagal control) for the purpose of reducing exposureof the gut epithelium to potentially damaging toxins, acci-dentally ingested with the food and then removing thesetoxins prior to absorption (cf. cough reflex). Although vomit-ing can be induced by vestibular, cortical and brainstem(area postrema) inputs it is the abdominal vagal afferentinput from the gut which is considered to be the trigger ofmost relevance to survival [33]. This protective reflex ispresent in humans and reported across a wide range ofother mammalian species, representing nine mammalianorders (Figure 3). Data were not found on representatives ofnine other orders (Montremata, Dermoptera, Macrosceli-dea, Scandentia, Xenarthra, Pholidota, Tubulidentata, Hyr-acoidea and Sirenia), representing only �2% of mammalianspecies. In representatives from the remaining two orders(Rodentia and Lagomorpha) there is no ability to vomit. Ratsare well known to ingest soil in the wild and kaolin in thelaboratory (pica) in response to toxins that would causenausea/vomiting in other species. Pica is argued to be adefensive strategy aimed at ameliorating the effect of toxinswhich in the wild would have been ingested with food and isnot confined to species lacking a vomiting reflex, althoughthe experimental evidence for its existence in mice andrabbits is scant in comparison to rats ([33] for detaileddiscussion]).
The finding that a vomiting capability is widespreadamong mammalian orders suggests that the emetic reflexwas present in mammals ancestral to those which do notpossess the reflex. Thus, the ability to vomit must be lostfrom these lineages, rather than fail to develop. The gas-trointestinal and somatomotor events (see above), togetherwith salivation, increased swallowing, tachycardia andcutaneous vasoconstriction that frequently accompanyemesis, are coordinated in the brainstem by neuronesconceptualised as a ‘vomiting centre/pattern generatorfor emesis’ [34]. The brainstem is a relatively ‘primitive’brain region and as rodents possess the same key nuclei(e.g. nucleus tractus solitarius, dorsal motor vagal nuclei)and motor outputs (e.g. phrenic and vagus nerves) asvomiting species, their inability to vomit could reflectadaptive changes in the brainstem in addition to thedigestive tract itself (Table 1). Together, these argumentsare consistent with observational reports of differences inbrainstem projections to the phrenic nucleus betweenferrets and rats [34], the presence in rats of the gag reflexitself mediated via the brainstem [35] and some ability ofmice to retch but not vomit [36].
Notably, the ghrelin system is conserved irrespective ofwhether vomiting is present or absent (Figure 3). However,it is particularly striking that the capacity to vomit and theexistence of a functional motilin system have both been lostin the Rodentia (rats, mice, guinea pig). By contrast, theLagomorpha (as exemplified by the rabbit) have retainedthe motilin system but lost the ability to vomit [37].Unusually, rabbits are dependent on caecotrophy as partof their normal digestive process [38]. Here, plant materialis partially digested within the caecum and excreted as softfaecal pellets for immediate reingestion, further fermenta-tion and absorption of water and nutrients (providing anessential source of vitamin B [39]), before final excretion as
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
406
Author's personal copy
hard faecal pellets. The process requires sophisticatedcolonic separation mechanisms (e.g. contraction of a spiralmuscle) and coordination between gastric and caecal emp-tying rates, both of which are never fully empty. Notably,motilin receptor agonists facilitate both ends of this diges-tive process – gastric emptying and the defecation of hardpellets [40]. Thus, the essential dependence on caecotrophyin a relatively large animal, utilising a relatively lowenergy density diet with a high fibre residue, could haveprovided a strong selection pressure for the conservation ofan endocrine regulator of gut motility (i.e. motilin).
Thus, by using the emetic reflex as an index and byconsidering the rabbit as a ‘special case’, we can suggestthat the loss of the motilin system in rats, mice and guineapigs occurred as a consequence of the evolutionary pres-sures which shaped their digestive functions (e.g. leadingto the loss of the vomiting reflex and also the unusualexpulsion of faeces as hard, individual pellets), arguably tominimise water loss and thereby promote survival across awide range of environments. In other words, the loss of thevomit reflex removed selective pressure for retention of themotilin system. Alternately, it should be noted that inanimals which vomit, the process of restarting normal,anally directed GI motility following vomiting is an impor-tant requirement, so the metabolic and ionic deficit israpidly restored by feeding [41]. Accordingly, a role forendogenous motilin in this process can be envisaged, be-cause although ghrelin can increase gastric motility, theprokinetic activity of motilin can be especially aggressive
[3]. By contrast, in rodents, toxic or ‘emetic stimuli’ induceprolonged gastric stasis, part of a defensive strategy toprevent further ingestion of toxic material, and delaydelivery of toxins into the intestine [42]. The lack of ahormone which aggressively promotes GI motility is con-sistent with the reduced requirement for such a mecha-nism.
The way forward: implications for use of rodents asmodels in GI researchIt is important to understand the biology of rodents interms of how this might impact on translation to humanphysiology and pharmacology. In the present example, theloss of motilin functions can be related to the loss of theemetic reflex, but the correlation is complex and additionalmarkers of GI functions are needed, in which data isavailable across multiple mammalian species. With regardto motilin, it is noteworthy that motilin receptor agonistsare currently being developed for disorders associated withdelayed gastric emptying (e.g. diabetic and non-diabeticgastroparesis) [3,4], yet these developments stem from theaccidental discovery that an antibiotic drug (erythromycin)was also a gastric prokinetic agent (subsequently identifiedas a motilin receptor agonist) and owe nothing to the studyof rodents, the most common group of laboratory animals.
Given the widespread use of mice, rats and guinea pigsin medical research, it would now be interesting to look forother genes with different patterns of expression acrossthese species, relative to humans, relating the differences
X
X
DogPig
Rat
HorseElephant
Sperm Whale
RabbitGuinea-pig
Eutheria
Primate [50]
Insectivora [51]
Lagomorpha[56 ] Rodentia[51 ,56 ]
Carnivora [50,56]
Gorill avervet monke y
Musk shrew
Camel
Cow
Goa t
Guanac o
Cat
Cheetah
Ferret
Jaguar
Leopard
Lion
Sea lion
Wolf
Zorro
Mouse
hamster
Chiroptera[55]
Vomiting reflex absent
Commonbrushtailpossum
Marsupiala[49]
Vomiting reflex present
Human
Fruit bat
Artiodactyla[50,52,53]
Cetacea[54]
Proboscida[50 ]
Perissodactyla[52]
TRENDS in Pharmacological Sciences
Figure 3. Phylogenetic tree showing the presence or absence of the ability to vomit in members of the 11 major mammalian orders, representing �98% of mammalian
species, based on key reports in the published literature (note that the number of species indicated is limited only by space and not by the number of reports that are
available in the literature) (e.g. Marsupiala [49], Primate [50], Insectivora [51], Perissodactyla [52], Artiodactyla [50,52,53], Cetacea [54], Chiroptera [55], Carnivora [50,56],
Lagomorpha [56], Rodentia [51,56], Note that in a few cases (e.g. elephant) the presence of vomiting is based on a single clinical report). Functional motilin system,
Functional ghrelin system. = point at which it is proposed that the vomiting reflex was lost and = point at which a functional motilin system was lost. Shaded box
indicates orders and species in which the vomiting reflex is absent. The arrow next to the rabbit silhouette indicates that coprophagia is a normal part of digestive behaviour
of this animal.
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
407
Author's personal copy
to species-dependent functions of both the GI tract (Table1) and to other organs. An obvious candidate is the5-hydroxytryptamine3 receptor, which is closely associatedwith emesis and consists of at least five different subunitsin the human and dog genome, compared to only two inrodents [43]. It is essential to highlight these differences,determine if they are widespread or peculiar to rodents anddevelop an understanding of why they occur. This infor-mation informs the level of caution required when trans-lating animal studies on mechanisms (or genes) to humansbut also an opportunity to identify molecular causes ofspecies differences in physiology and disease [44]. Al-though we have focused on the emetic reflex coordinatedin the brainstem, a comparative study of the neurophar-macology of key nuclei in this brain region of emetic andnon-emetic species is also warranted, because of its addi-tional involvement in cough and pain (especially visceral)where rodents are common models. These strands of evi-dence suggest a need for caution in the reliance on the useof laboratory rodent models as representative of humanbiology.
A final note of caution must be directed at the apparentdiscrepancies which continue to be published, showingclear functional activity with motilin receptor agonists inrats, mice and guinea pigs, species in which motilin or itsfunctional receptor has not been identified. It is essential tounderstand these differences and thereby clarify the avail-able literature using several approaches: attempting toclone the ‘motilin receptor’ from those individuals in whicha response to motilin has been demonstrated; carefulexamination of the selectivity of the motilin receptor ago-nist used, against other rodent receptors such as theghrelin receptor (which is known to be present withinthe rat brainstem [45]); and undertaking a comparativegenomic analysis as the majority of the apparently discor-dant studies are performed in rats from different breedingpopulations.
Conflict of interest statementG.J.S. was the recipient of a grant from GlaxoSmithKlineto study the gastrointestinal actions of motilin receptoragonists, including GSK962040. J.D.H. and P.L.R.A. de-clare no conflicts of interest.
AcknowledgementsWe thank Jeffrey Hill, Harjeet van der Keyl, Nabil Elsbourghy and TaniaTesta, who provided early information on the identity of someorthologues, now in public databases. We thank Dr Charles Hoyle forcomments on an early draft of parts of the manuscript.
References1 Stevens, C.E. and Hume, I.D. (1995) Comparative Physiology of the
Vertebrate Digestive System, (2nd edn), Cambridge University Press2 He, J. et al. (2010) Stepwise loss of motilin and its specific receptor
genes in rodents. J. Mol. Endocrinol. 44, 37–443 Sanger, G.J. (2008) Motilin, ghrelin and related neuropeptides as
targets for the treatment of GI diseases. Drug Discov. Today 13,234–239
4 Ohno, T. et al. (2010) The roles of motilin and ghrelin ingastrointestinal motility. Int. J. Pept. 2010, 1–6
5 Kojima, M. and Kangawa, K. (2010) Ghrelin: more than endogenousgrowth hormone secretagogue. Ann. N. Y. Acad. Sci. 1200, 140–148
6 Sakharkar, M.K. et al. (2005) Computational prediction of SEG (singleexon gene) function in humans. Front Biosci. 10, 1382–1395
7 Dass, N.B. et al. (2003) The rabbit motilin receptor: molecularcharacterisation and pharmacology. Br. J. Pharmacol. 140, 948–954
8 Kojima, M. et al. (2008) Structure of mammalian and non-mammalianghrelins. Vitam. Horm. 77, 31–46
9 Xu, L. et al. (2005) Evidence for the presence of motilin, ghrelin, and themotilin and ghrelin receptor in neurons of the myenteric plexus. Reg.Peptides 124, 119–125
10 Ohshiro, H. et al. (2008) Molecular identification and characterizationof the dog motilin receptor. Reg. Peptides 146, 80–87
11 Yogo, K. et al. (2007) Effects of motilin and mitemcinal (GM-611) ongastrointestinal contractile activity in rhesus monkeys in vivo and invitro. Dig. Dis. Sci. 52, 3112–3122
12 De Clercq, P. et al. (1997) Isolation and sequencing of the cDNAencoding the motilin precursor from sheep intestine. Gene 202,187–191
13 Wechsung, E. and Houvenaghel, A. (1993) Influence of motilin ongastrointestinal electrical activity in the conscious piglet. J. Vet.Med. 40, 229–232
14 Tsutsui, C. et al. (2009) House musk shrew (Suncus murinus, order:Insectivora) as a new animal model for motilin study. Peptides 30,318–329
15 Bassil, A. et al. (2005) Prokineticin-2, motilin, ghrelin andmetoclopramide: prokinetic utility in mouse stomach and colon. Eur.J. Pharmacol. 524, 138–144
16 Depoortere, I. et al. (2005) Comparison of the gastroprokinetic effects ofghrelin GHRP-6 and motilin in rats in vivo and in vitro. Eur. J.Pharmacol. 515, 160–168
17 Feng, X. et al. (2007) Motilin activates neurons in the rat amygdala andincreases gastric motility. Peptides 28, 625–631
18 Jia, Y-D. et al. (2007) Expression of motilin in the hypothalamus andthe effect of central erythromycin on gastric motility in diabetic rats.Neurosci. Bull. 23, 75–82
19 Fang, P. et al. (2010) Effects of motilin on intracellular free calcium incultured smooth muscle cells from the antrum of neonatal rats. ActaPhysiol. 199, 53–61
20 Xu, L. et al. (2008) Effect of motilin on the discharge of rat hippocampalneurons responding to gastric distension and its potential mechanism.Peptides 29, 585–592
21 Todak, H. et al. (2009) Motilin acts as a modulator of post-synapticGABA receptor in the medial vestibular nuclear neurons in mice.Neurosci. Res. 65 (Suppl. 1), S1389
22 Liu, H. et al. (2010) Erythromycin inhibited glycinergic inputs togastric vagal motorneurons in brainstem slices of newborn rats.Neurogastroenterol. Motil. 22, 1232–1239
23 Xu, L. et al. (2011) Effect of motilin on gastric distension sensitiveneurons in arcuate nucleus and gastric motility in rat.Neurogastroenterol. Motil. 23, 265, e121
24 Asakawa, A. et al. (1998) Motilin increases food intake in mice. Peptides19, 987–990
25 Furness, J.B. et al. (1999) An action of erythromycin in the intestinethat is not mediated via motilin receptors. Clin. Exp. Pharmacol.Physiol. 26, 100–104
26 Katayama, Y. et al. (2005) Motilin inhibits ganglionic transmission inthe myenteric plexus of the guinea-pig ileum. Neurosci. Res. 53, 156–
16027 Katayama, Y. et al. (2005) Excitatory actions of motilin on myenteric
neurons of the guinea-pig small intestine. Auton. Neurosci. 118, 88–9228 Asakawa, A. et al. (2003) EM574, a motilide, has an orexigenic activity
with affinity for growth-hormone secretagogue receptor. J.Gastroenterol. Hepatol. 18, 881–882
29 Delinsky, D.C. et al. (2004) Quantitative determination of thepolypeptide motilin in rat plasma by externally calibrated liquidchromatography/electrospray ionization mass spectrometry. RapidCommun. Mass Spectrom. 18, 293–298
30 Nilaver, G. et al. (1988) Heterogeneity of motilin immunoreactivity inmammalian tissue. Synapse 2, 266–275
31 Blanga-Kanfi, S. et al. (2009) Rodent phylogeny revised: analysis of sixnuclear genes from all major rodent clades. BMC Evol. Biol. 9, 71
32 Ariga, H. et al. (2008) Fixed feeding potentiates interdigestive gastricmotor activity in rats: importance of eating habits for maintaininginterdigestive MMC. Am. J. Physiol. 294, G655–G659
33 Stern, R.M. et al. (2011) Nausea – Mechanisms and Management,Oxford University Press
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7
408
Author's personal copy
34 Yates, B.J. et al. (1998) Physiological basis and pharmacology of motionsickness. Brain Res. Bull. 47, 395–406
35 Andrew, B.L. (1956) The nervous control of the cervical oesophagus ofthe rat during swallowing. J. Physiol. 134, 729–740
36 Furukawa, T. and Yamada, K. (1980) The a-napthoxyacetic acid-elicited retching involves dopaminergic inhibition in mice.Pharmacol. Biochem. Behav. 12, 735–738
37 Borison, H.L. et al. (1981) Phylogenetic and neurologic aspects of thevomiting process. J. Clin. Pharmacol. 21, 23S–29S
38 Bjornhag, G. (1994) Adaptations in the large intestine allowing smallanimals to eat fibrous foods. In The Digestive System in Mammals(Chivers, D.J. and Langer, P., eds), pp. 287–309, CambridgeUniversity Press
39 Stevens, C.E. and Hume, I.D. (1998) Contribution of microbes invertebrate gastrointestinal tract to production and conservation ofnutrients. Physiol. Rev. 78, 393–427
40 Sanger, G.J. et al. (2009) GSK962040: a small molecule, selectivemotilin receptor agonist, effective as a stimulant of human andrabbit gastrointestinal motility. Neurogastroenterol. Motil. 21, 657–666
41 Andrews, P.L.R. et al. (2005) Potential energetic implications of emesisin the house musk shrew (Suncus murinus). Physiol. Behav. 84,519–524
42 Sanger, G.J. and Andrews, P.L.R. (2006) Treatment of nausea andvomiting: gaps in our knowledge. Auton. Neurosci. Basic Clin. 129, 3–16
43 Holbrook, J.D. et al. (2009) Characterisation of 5-HT3c, 5-HT3d and 5-HT3e receptor subunits: evolution, distribution and function. J.Neurochem. 108, 384–396
44 Vamathevan, J. et al. (2008) The mouse genome as a rodent model inevolutionary studies. In Handbook of Human Molecular Evolution(Vol. 2) Cooper, D.N. and Kehrer-Sawatzki, H.,eds In pp. 1445–1452,John Wiley
45 Faulconbridge, L.F. et al. (2008) Caudal brainstem delivery of ghrelininduces fos expression in the nucleus of the solitary tract, but not in thearcuate or paraventricular nuclei of the hypothalamus. Brain Res.1218, 151–157
46 Edgar, R.C. (2004) MUSCLE: a multuiple sequence alignment methodwith reduced time and space complexity. BMC Bioinform. 5, 113
47 Dereeper, A. et al. (2008) Phylogeny.fr: robust phylogenetic analysis forthe non-specialist. Nucleic Acids Res. 36, W465–W469
48 Page, R.D.M. (1996) TREEVIEW: an application to displayphylogenetic trees on personal computers. Computer Appl. Biosci.12, 357–358
49 Morgan, D.R. (1991) Behavioural responses of brushtail possumsTrichosurus vulpecula, to baits used in pest control. Aust. Wildl.Res. 17, 601–614
50 Dolowy, W.C. et al. (1960) Some observations on vomiting in animals. J.Am. Vet. Met. Assn. 137, 426–427
51 Liu, Y.L. et al. (2005) Pica – a model of nausea? Species differences inresponse to cisplatin. Physiol. Behav. 85, 271–277
52 Smith, B.P. (1990) Large Animal Internal Medicine – Diseases ofHorses, Cattle, Sheep, and Goats, C.V Mosby Co.
53 Milano, S. et al. (1995) The piglet as a suitable animal model forstudying the delayed phase of cisplatin-induced emesis. J.Pharmacol. Exp. Ther. 274, 951–961
54 Clarke, R. et al. (1988) Sperm whales of the southeast pacific. Part IV:fatness food and feeding. In Investigations on Cetacea (Vol. XXI)(Pilleri, G., ed.), In pp. 53–195, Brain Anatomy Institute, Universityof Berne
55 Vogelenest, L. and Woods, R. (2008) Medicine of Australian Mammals,CSIRO Publishing
56 Andrews, P.L.R. and Horn, C.C. (2006) Signals for nausea and emesis:implications for models of upper gastrointestinal diseases. Auton.Neurosci. 125, 100–115
57 Andrews, P.L.R. et al. (2003) Differences in the vagal innervation of theoesophagus may explain the lack of emesis in rodents.Neurogastroenterol. Motil. 15 (Abstr. 142), 28
58 Zawaira, A. et al. (2008) Prediction of sites under adaptive evolution incytochrome P450 sequences and their relationship to substraterecognition sites. Pharmacogenet. Genomics 18, 467–476
59 Antal-Zimanyi, I. and Khawaja, X. (2009) The role of melanin-concentrating hormone in energy homeostasis and mood disorders.J. Mol. Neurosci. 39, 86–98
60 Rose, P.M. et al. (1997) Molecular genetic analysis of a humanneuropeptide Y receptor. J. Biol. Chem. 272, 3622–3627
61 Andrews, P.L.R. and Rudd, J.A. (2004) The role of tachykinins and thetachykinin NK1 receptor in nausea and emesis. In Tachykinins.Handbook of Experimental Pharmacology (Holzer, P., ed.), pp. 359–
440, Springer62 Kinloch, R.A. and Cox, P.J. (2005) New targets for neuropathic pain
therapeutics. Expert Opin. Ther. Targets 9, 685–69863 Gilad, Y. et al. (2003) Human specific loss of olfactory receptor genes.
Proc. Natl. Acad. Sci. U.S.A. 100, 3324–3327
Review Trends in Pharmacological Sciences July 2011, Vol. 32, No. 7