39
SECTION II Understanding Emotional Language Content (Edited by Johanna Kissler)
| Date post: | 14-Nov-2023 |
| Category: |
Documents |
| Upload: | independent |
| View: | 0 times |
| Download: | 0 times |

SECTION II
Understanding Emotional Language Content(Edited by Johanna Kissler)


Anders, Ende, Junghofer, Kissler & Wildgruber (Eds.)
Progress in Brain Research, Vol. 156
ISSN 0079-6123
Copyright r 2006 Elsevier B.V. All rights reserved
CHAPTER 8
Emotional and semantic networks in visual wordprocessing: insights from ERP studies
Johanna Kissler�, Ramin Assadollahi and Cornelia Herbert
Department of Psychology, University of Konstanz, P.O. Box D25, D-78457 Konstanz, Germany
Abstract: The event-related brain potential (ERP) literature concerning the impact of emotional content onvisual word processing is reviewed and related to general knowledge on semantics in word processing:emotional connotation can enhance cortical responses at all stages of visual word processing following theassembly of visual word form (up to 200ms), such as semantic access (around 200ms), allocation ofattentional resources (around 300ms), contextual analysis (around 400ms), and sustained processing andmemory encoding (around 500ms). Even earlier effects have occasionally been reported with subliminal orperceptual threshold presentation, particularly in clinical populations. Here, the underlying mechanisms arelikely to diverge from the ones operational in standard natural reading. The variability in timing of theeffects can be accounted for by dynamically changing lexical representations that can be activated asrequired by the subjects’ motivational state, the task at hand, and additional contextual factors. Through-out, subcortical structures such as the amygdala are likely to contribute these enhancements. Furtherresearch will establish whether or when emotional arousal, valence, or additional emotional propertiesdrive the observed effects and how experimental factors interact with these. Meticulous control of otherword properties known to affect ERPs in visual word processing, such as word class, length, frequency, andconcreteness and the use of more standardized EEG procedures is vital. Mapping the interplay betweencortical and subcortical mechanisms that give rise to amplified cortical responses to emotional words will beof highest priority for future research.
Keywords: emotion; semantics; word processing; event-related potentials; healthy volunteers; clinicalpopulations
Introduction
Influential dimensional approaches to the study ofemotion derive their basic dimensions from analy-ses of written language. Osgood and collaborators,using the ‘semantic differential’ technique, werethe first to empirically demonstrate that affectiveconnotations of words are determined by threeprincipal dimensions, namely evaluation, potency,
and activity (Osgood et al., 1957), the first two ac-counting for the majority of the variance. On thesemantic differential, a word’s evaluative connota-tion is determined by ratings on a multitude ofseven-point scales, spanned by pairs of antonymssuch as hot–cold, soft–hard, happy–sad, etc. Factoranalyses of the judgments of many words on suchscales, given by large subject populations, reveal athree-dimensional evaluative space, whose structurehas been replicated many times and across differentcultures (Osgood et al., 1975). Figure 1 provides anillustration of the evaluative space determined bythe semantic differential.
�Corresponding author. Tel.: +49-7531-884616;
Fax: +49-7531-4601; E-mail: [email protected]
DOI: 10.1016/S0079-6123(06)56008-X 147

Osgood’s principal dimensions are at the core ofother circumplex theories of affect (Lang, 1979;Russel, 1980). For instance, Lang and colleaguesposit that human affective responses are determinedby the dimensions valence, arousal, and dominance;again the first two having the largest impact, leadingthese authors to propose model of affect defined bythe dimensions of arousal and valence. Within such atwo-dimensional affective space, different classes ofstimuli such as pictures, sounds, and words cluster ina u-shaped manner. Highly arousing stimuli usuallyreceive valence ratings as either highly pleasant orhighly unpleasant, low arousing material is generallyregarded as more neutral with regard to valence (see
Fig. 2 for an illustration derived from the ratings ofGerman word stimuli). The impact of valence andarousal on various central nervous and peripheralphysiological indicators of affective processing hasbeen repeatedly validated using picture and soundmedia (Lang et al., 1993; Bradley and Lang, 2000;Junghofer et al., 2001; Keil et al., 2002).
Word stimuli assessed for perceived arousal andvalence such as the ‘Affective Norms for EnglishWords’, ANEW (Bradley and Lang, 1998) havealso been put to use in physiological research (seee.g., Fischler and Bradley, this volume), althoughso far the resulting evidence seems more restrictedthan the one for pictorial material.
Fig. 1. A three-dimensional affective space of connotative meaning as postulated by Osgood (1957, 1975). The figure depicts the three
orthogonal bipolar dimensions, evaluation (E), activity (A) and potency (P), and gives examples of prototypical words for each
dimension and polarity. (Adapted from Chapman, 1979.)
148

Emotions are generally viewed as culturally uni-versal, largely innate evolutionary ‘old’ signalingand activation systems, residing in ‘old’, subcorti-cal parts of the brain. They are designed to pro-mote survival in critical situations, i.e., to signaland activate fight, flight or feeding, attachment,and sexual behavior.
Reading and writing, by contrast, representcomparatively recent developments in the historyof mankind, and in individual development theseare acquired much later than oral language. Con-sequently, these skills are often regarded as
cultural achievements, but during the acquisitionof written language considerable regional special-ization emerges in the human brain (Warringtonand Shallice, 1979, 1980; Dehaene et al., 2005).Reading acts as a secondary process that utilizesthe processing capabilities of the earlier acquiredauditory language system once the analysis of vis-ual word form is completed (Perfetti, 1998; Everattet al., 1999; Perfetti and Sandak, 2000). Languageand emotion share a communicative function butlinguistic communicative functions are obviouslynot restricted to the communication of affect.
Fig. 2. Illustration of a two-dimensional affective space spanned by the dimensions arousal and valence. Examples of the relative
position of some German adjectives and nouns used in our studies are given in English translation. Along the x- and y-axis are depicted
the arousal and valence scales of the Self-Assessment Manikin (SAM, Bradley and Lang, 1994) used to rate individual emotional
responses to the words.
149

How do the ‘emotional brain’ and the ‘linguisticbrain’ interact, when written words with emotionalconnotations are encountered? Emotion theoriesposit that linguistic expressions are stored withinsemantic networks that encompass links to all as-pects of their linguistic and pragmatic usages andemotional connotations (Lang, 1979; Bower, 1981).Thus, the word ‘gun’, for example, not only repre-sents the object itself, but also includes links to itsoperations, use, purposes, and their consequencesas well as their emotional evaluation (Bower, 1981).A converging view is shared in neurolinguistics(Pulvermuller, 1999) and cognitive semantics(Barsalou, 1999): All information related to a wordis stored in a dynamic network. Recent evidencesuggests that subnetworks1 representing differentaspects of a word’s lexical representation can beseparately and dynamically activated. For instance,differential neuromagnetic activations of semanticsubnetworks have recently been shown for sub-classes of plant or animal names (Assadollahi andRockstroh, 2005). Moreover, biasing contextualconstrains can affect the timing of access to thedominant vs. subordinate meaning of homonyms(Sereno et al., 2003), challenging the modular viewthat in word processing initially all lexical entrieshave to be exhaustively accessed.
Functional divisions of the semantic systemmirroring functional divisions in the organizationof the cortex have repeatedly been shown for verbsdenoting different types of actions (Pulvermuller etal., 2001b; Hauk et al., 2004). Investigating verbspertaining to movements carried out with differentparts of the body, these authors demonstrate thatthe meaning of action words is reflected by thecorrelated somatotopic activation of motor andpremotor cortex. These patterns of coactivationspresumably reflect individual learning history,where the so-called referential meaning has beenacquired by repeated coactivation of the body
movement and the descriptive speech pattern, forinstance when a child observes or carries out agesture such as throwing and simultaneously hearsthe caregiver say the respective word. Later, in theacquisition of written language, this phonologicalcode is mapped onto the visual word form (Perfettiand Sandak, 2000).
For emotional concepts, Lang et al. (1993, 1994)assume that not only associated semantic but alsophysiological and motor response information iscoactivated in associative networks. Figure 3 illus-trates such a multilevel network representation ofan emotional scene, encompassing a semantic codeof the given situation as well as associated motorand physiological responses. Thus, the semanticnetwork that codes for ‘emotional semantics’could include the neuronal circuitry processingthe associated emotion (see also Cato and Cross-on, this volume, for a related suggestion).
How does emotional content influence differentstages of visual word processing? Here, the literatureis sparse. Event-related brain potentials (ERPs), thescalp recorded averaged synchronized activity ofseveral thousands cortical pyramidal cells, have suc-cessfully been used to delineate different stages ofvisual word processing (for reviews see e.g., Posneret al., 1999; Tarkiainen et al., 1999). A closer lookalso reveals that a considerable number of electro-physiological studies of emotional processing haveemployed visually presented word-stimuli. Some,particularly early, studies have used the semanticdifferential as their theoretical vantage point. In fact,in the wake of Osgood’s studies, the examination ofERP correlates of emotional semantics generatedsubstantial research interest (Chapman et al., 1978,1980; Chapman, 1979; Skrandies, 1998; Skrandiesand Chiu, 2003). More recently, two-dimensionalvalence � arousal models have been used as aframework for research (see Fischler and Bradley,this volume). However, many studies used wordswith emotional connotations as experimentally con-venient instances of a broader class of emotionalevents (Anderson and Phelps, 2001; Dijksterhuis andAarts, 2003) or conversely, as a semantic class with-out much reference to any particular theory of lan-guage and/or emotion (Begleiter and Platz, 1969). Sofar, little systematic knowledge has been gathered onthe relationship between the emotion and language
1The terms sub-network or sub-representation as used here
are not necessarily intended to imply a fixed hierarchical or-
dering of the neural networks coding for different aspects of
semantics, although a certain degree of hierarchical ordering
may indeed exist. Instead, sub-network or sub-representation
refers to the fact that different neural networks are likely to
code for different aspects of a word’s meaning, such as ani-
macy, emotional connotation, grammatical gender, etc.
150

systems in visual word processing and possible im-plications for the underlying neural implementationof meaning.
ERP recordings have an excellent temporal res-olution, allowing for a fine-grained analysis of thetemporal sequence of different processing stages.Their spatial resolution is more restricted, and in-ferences from the spatial distribution of scalpmeasured ERPs to their neural generators can onlybe made with caution. The number of electrodesand the recording reference used influence theprobability of finding effects, the generalizabilityof these findings, and the accuracy of spatial lo-calization (see also Junghofer et al., this volume).
The present review will summarize and system-atize existing studies on the role of emotion invisual word processing, both in healthy volunteersand in clinical populations and relate this evidence
to the available knowledge on ERP indices ofstages of visual word processing.
First, we will address studies reporting earlyeffects of emotional content on ERP responses, oc-curring within the first 300ms after stimulus onset,separately for healthy volunteers and clinical pop-ulations; second, we will review effects of emotionalcontent on the late ERPs to visual words; and third,discuss how an interplay of subcortical and corticalmechanisms of emotion and word processing maygive rise to the observed effects. To facilitate com-parisons between studies, main methodological pa-rameters and results concerning effects of emotionalcontent, as stated in the reviewed studies, are sum-marized in two tables in the appendix. Table A1describes studies with healthy volunteers; Table A2describes work with clinical populations. There, itbecomes immediately apparent that the studies
Fig. 3. A network representation of a complex emotional scene (exam situation) illustrates how in dynamic emotional processing
perceptual, semantic and response systems are interactively linked. Activation on any level of this system can spread to other
subsystems. (Adapted after Lang, 1994.)
151

described vary considerably in theoretical approachand methodology.
Early effects — occurring within 300ms after
stimulus onset
Healthy volunteers
The extent to which early exogenous components ofthe human event-related potential are subject tomodification by nonphysical stimulus characteristicsis a matter of ongoing controversy. In visual wordprocessing, a traditional view holds that within thefirst 150–200ms after a word has been presented,specific perceptual features of written words but nomeaning-related attributes are extracted (Schendanet al., 1998; Posner et al., 1999) and for many yearsthe N400 potential, a centro-parietal negativity aris-ing around 400ms after stimulus onset has beenviewed as ‘the’ index of semantic processing (Kutasand Federmeier, 2000).
Using other types of visually presented stimuliwith emotional content, such as faces or pictures,remarkable ERP differences between emotionallysignificant and neutral stimuli have been foundwithin the first 300ms after stimulus onset, someeven within the first 100ms.
In his single subject study of ERP responses toemotional words, Lifshitz (1966) failed to find avisually impressive differentiation between emo-tional and neutral words within the first 500msafter word onset, although upon visual inspectionthe difference between the likewise presented eroticand neutral line drawings was sizeable. However, inthe meantime there is a considerable body of evi-dence indicating that even very early ERP responsescan diverge between emotional and neutral words.Thus, the probably first quantitative study on theeffects of emotional content on ERP indices ofword processing found differences between negative‘taboo words’ and neutral words already withinthe first 200ms after word presentation (Begleiterand Platz, 1969). Notably, this study was entitled:‘Cortical evoked potentials to semantic stimuli’,expressing a view of emotion as a vital part ofsemantics. The technological standards at thetime were not very sophisticated and the authors
recorded only from a single right occipital electrode(O2), but ERP differences due to emotional contentappeared in the ERP tracings already in the P1–N1complex: twenty minimally above threshold pre-sented repetitions of each of two clearly negative‘four-letter –words’ led to larger responses thantwenty repetitions of each of the words ‘tile’ and‘page’. This pattern held for both a passive viewingcondition and a naming condition.
Several years later, Kostandov and Azurmanov(1977) contrasted ERP responses to both sublim-inally and supraliminally presented ‘conflict’ andneutral words. In the subliminal condition, theearliest differences between ‘conflict’ and neutralappeared around 200ms after stimulus onset. Sub-liminally presented ‘conflict words’ apparently re-lating to the subjects’ relationship conflicts causedby jealousy led to larger N200 responses than neu-tral words. Supraliminally presented words led tolater differentiations starting around 300ms, in theP3a window. Unfortunately, the description of thestudy is somewhat sparse on details regardingthe materials used.
Chapman et al. (1978) sought to electrophysio-logically validate Osgood’s connotative dimen-sions of meaning (see Fig. 1), namely evaluation(E), potency (P), and activity (A). They recordedfrom one midline electrode (CPz) referenced tolinked mastoids while subjects had to name brieflyflashed (17ms) words. Twenty words each fromthe extreme ends of the three connotative dimen-sions, E, P, and A, yielding six semantic classes(E+/-, P+/-, and A +/-) were used as stimuli.Random sequences of all these words were pre-sented 12–20 times to the subjects (until a set cri-terion of artifact-free trials was entered into theaverage). Although the exact multivariate analysisof the data in the original report is somewhat hardto reconstruct, in essence Chapman et al. (1978)were able to statistically differentiate between allsix connotative dimensions using ERP data from asingle central channel. Whereas the different con-notative dimensions are not mapped onto anyspecific ERP components, in the original grand-averages a second positive peak at around 260msafter word onset is noticeable that clearly differ-entiates between the extreme ends (+ and -) of allthree dimensions but not so much among E, P, and
152

A. In general, the material in this study was wellcontrolled for physical aspects but apparently notfor other linguistic attributes such as word fre-quency, word class, or concreteness. Replicationstudies by the same group investigated the effectsof explicit affective rating tasks on ERPs (Chap-man, 1979) and directly compared the effects ofaffective rating and word naming in one study,yielding similar results (Chapman et al., 1980).Thus, a series of studies demonstrated reliable sta-tistical differentiation between six connotative cat-egories within 500ms after stimulus onset on thebasis of ERP data from a single channel.
Extending their 1969 study across a larger selec-tion of stimuli, Begleiter et al. (1979) recordedERPs elicited by unpleasant, pleasant, and neutralwords, as 10 subjects had to either identify the lastvowel in a presented word or give their personalaffective evaluation of the word. Recordings weremade from three electrodes over each hemisphere,referenced to linked mastoids. However, results arereported only for two of these electrodes, namely P3and P4. Stimuli were the 62 most unpleasant, 62most neutral, and 62 most pleasant five-letter wordsderived from a larger pool of words previously as-sessed with the semantic differential (Osgood et al.,1957). Words were presented very briefly (20ms).The words’ emotional meaning affected N1–P2peak-to-peak amplitude during emotional evalua-tion. ERP responses to words evaluated as pleasant,neutral, or unpleasant could be distinguished sta-tistically at both electrodes. Overall, effects of emo-tional content were somewhat more pronouncedover the left hemisphere and were restricted to theaffective evaluation condition. At the left hemi-spheric electrode, ERPs were also generally largerwhen the words were shown in the emotional eval-uation than in the letter-identification task. Unlikethe previous studies, Begleiter et al. (1979) was thefirst to provide evidence for a major impact of task,showing early ERP effects of emotional contentonly during active evaluation.
Skrandies (1998), studying ‘ERP correlates of se-mantic meaning’, used an approach conceptuallysimilar to Chapman’s (1978, 1979, 1980). A pool of60 nouns representing the bipolar extremes on Os-good’s E, P, and A dimensions were selected andpresented in a rapid serial visual presentation
(RSVP) design, i.e., a continuous stream of alter-nating words without interstimulus interval. Skran-dies (1998) used a comparatively slow RSVP designpresenting each word for 1 s. Ten stimuli per cate-gory and polarity were selected and each stimuluswas repeated 40 times, yielding 400 averages percategory, thus optimizing the signal-to-noise ratio.Subjects were instructed to visualize the words andto remember them for a subsequent memory test inorder to ensure active engagement with the stimuli.EEG was recorded from 30 channels referenced toan average reference. Brain responses between theemotional word categories differed in six distincttime windows, with respect to either their peaklatency, associated global field power or the locationof the centroids of the scalp distribution. Remark-ably, most of the differences occurred within thefirst 300ms after word presentation, starting withP1 at around 100ms. These results were recentlyextended cross-culturally in a virtually identicalstudy of affective meaning in Chinese, yielding asomewhat different but equally complex pattern ofdifferences depending on emotional word content,which were restricted to the first 300ms after wordonset (Skrandies and Chiu, 2003). Together withBegleiter and Platz (1969), Skrandies studies reportprobably the earliest meaning-dependent differenti-ation between words. Consequently, these studiesare frequently cited in the visual word processingliterature, albeit without reference to the particularemotional semantic contents used.
Schapkin et al. (2000) recorded ERPs as subjectsevaluated nouns as emotional or neutral. The stim-uli consisted of a total of 18 words, 6 pleasant, 6unpleasant, and 6 neutral, which were matchedacross emotional categories for word length, fre-quency, concreteness, and initial letters. Wordswere presented peripherally, to the left and rightvisual fields for 150ms, while the EEG was re-corded from 14-linked mastoid referenced elec-trodes, eight of which were consistently analyzed.Stimuli were repeated 32 times, 16 times in eachvisual field. The earliest effect of emotional signifi-cance of the words on ERP responses was observedin the P2 window, peaking at 230ms. At bilateralcentral sites, P2 responses to pleasant words werelarger than responses to unpleasant and neutralones. While the P2 response was generally larger
153

over the left hemisphere, the emotion effect was notlateralized. Similar effects of emotional content onERP responses to words were also observed in latertime windows (see below).
As already suggested by Kostandov and Arzuma-nov (1977), the powerful effects of emotional con-notation on brain responses appear to extend evenbelow the limits of conscious perception. Bernatet al. (2001), recording from six-linked mastoidreferenced scalp electrodes, report a differentiationbetween both subliminally and briefly (40ms) pre-sented unpleasant and pleasant adjectives at left-hemispheric electrode positions already in the P1and N1 time ranges, as participants simply main-tained fixation on a central cross and viewed thecomputer screen without an explicit behavioral re-sponse being required. In the P1 and N1 windows,unpleasant adjectives lead to larger ERP responsesin the left hemisphere. Overall larger responses tounpleasant as compared to pleasant adjectives wereobtained in the subsequent P2, P3a, and LPC timeranges, the main effects of emotional content havingearlier onsets in the subliminal condition. The affec-tive valence of the stimuli had been determined byassessing a larger pool of words on five bipolarscales from the evaluative dimension of the semanticdifferential. Both the subsequent ERP study subjectsand an independent sample had repeatedly rated thestimuli. ERPs to 6 repetitions of the 10 most ex-tremely pleasant and unpleasant as well as to 12neutral words were recorded. Thus, study subjectshad considerable experience with the words. Also, itis unclear, whether the stimuli were assessed onother potentially relevant emotional or linguistic di-mensions such as arousal and dominance or wordlength, frequency, and abstractness. Still, these re-sults as well as data from Kostandov and Azurma-nov (1977) or Silvert et al. (2004) and Naccacheet al. (2005) in principle support the possibility ofmeasurable physiological responses to subliminallypresented emotional words and add to the evidenceof emotional content-dependent P1 differences(Begleiter, 1969; Skrandies, 1998; Skrandies andChiu, 2003).
Recently, Ortigue et al. (2004) also reported avery early effect of word emotionality in a densearray ERP study recording from 123 scalp chan-nels. The task consisted of a lexical decision to
very briefly flashed (13ms) stimuli. Subjects had toindicate which of two simultaneously presentedletter combinations in both visual fields consti-tuted an actual word. Stimuli were half neutral andhalf emotional nouns of both pleasant and un-pleasant valence. They were matched for wordlength and frequency and selected from a largerpool of words pre-rated on a bipolar seven-pointscale spanning the neutral-emotional continuum.Overall, emotional words presented in the rightvisual field were classified most accurately andfastest. However, the relative advantage for emo-tional words was larger for words presented in theleft visual field. Using a source estimation ap-proach (LAURA) the authors identified a stabletopographic pattern from the spatiotemporal dis-tribution of the ERP data that accounted for theprocessing advantage of emotional words in theright visual field. This pattern emerged between100 and 140ms after stimulus onset, i.e., mostly inthe P1/N1 window. Curiously, it was localized toprimarily right-hemispheric extra-striate cortex.Surprisingly, no specific neurophysiological corre-late of the even more pronounced advantage foremotional words in the left visual field was iden-tified within the 250ms after stimulus presentationthat this study restricted its analysis to.
Recent data from our own laboratory also pro-duced evidence for early differences between cor-tical responses to emotionally arousing (bothpleasant and unpleasant) and neutral adjectivesand nouns. The word’s emotional content hadbeen predetermined in a separate experiment, ob-taining valence and arousal ratings on two nine-point rating scales (see Fig. 2) from 45 undergrad-uate students. According to these ratings, highlyarousing pleasant and unpleasant and low arous-ing neutral words were selected. Different subsetsof these words were used in three studies whereERPs from 64 scalp sites were measured. Neutraland highly arousing pleasant and unpleasantwords matched for word length, frequency, andin one experiment also for concreteness were pre-sented in RSVP designs to subjects instructed toread the words. Across three different stimuluspresentation durations (333, 666, 1000ms) and re-gardless of word type (adjectives or nouns), a left-hemispheric dominant occipitotemporal negativity
154

differentiated emotional (both pleasant and un-pleasant) from neutral words. This negativity hadits maximum around 260ms after stimulus onset.The influence of stimulus repetition was assessed,but neither habituation nor sensitization wasfound for the emotional-neutral difference withinthe five repetitions used (Fig. 4 illustrates the effectfor the 666ms presentation rate). In one of thestudies we also manipulated task demands, in-structing subjects to attend to and count one of thetwo word classes (adjective or noun). Interestingly,this manipulation did not affect the enhanced earlynegativity to emotional words but had a significantimpact on the later positivity.
Herbert et al. (2006) also recorded ERPs from64 average-reference linked scalp channels, as 26subjects evaluated the emotional significance of
highly arousing pleasant and unpleasant as well asneutral adjectives. The affective content of thestimuli had been predetermined in a separate pop-ulation using the above-described procedure.Words were presented for a relatively long period,namely 5 s. In this study the P2 component was thefirst index of differential processing of emotionalvs. neutral words. This P2 component primarilyresponded to perceived stimulus intensity/arousaland did not differentiate brain responses to pleas-ant from those to unpleasant words. The same wastrue for the subsequent P3a component, but thepicture changed for a later LPC component andthe simultaneously recorded startle response thatwere more pronounced for pleasant than for un-pleasant and neutral words. This sequence ofeffects is depicted in Fig. 5.
Fig. 4. Early arousal-driven enhancement of cortical responses to emotional words. Uninstructed reading of both pleasant and
unpleasant words in a rapid serial visual stimulation paradigm (RSVP, 666ms stimulus duration) leads to larger occipitotemporal
negativities than reading of neutral words. The effect is illustrated at two occipital sensors (O9, O10) and the scalp topography of the
difference potential emotional–neutral words is depicted. Grand-averages from 16 subjects are shown.
155

Clinical studies
One of the first studies to use emotional words as atool to address processing biases (or a lack thereof)in clinical populations is Williamson et al. (1991)who investigated behavioral and cortical responsesto pleasant, unpleasant, and neutral words in psy-chopathic and nonpsychopathic prisoners. Subjectshad to detect words in a sequence consisting ofwords and nonwords while their EEG was being re-corded from five scalp positions referenced to linkedmastoids. Stimuli were presented vertically for176ms, separately to either visual field, and repeatedthree times. Stimuli had been matched for length,
number of syllables, word frequency, and concrete-ness but differed in emotional connotation. Non-psychopathic subjects had faster reaction times andlarger P2 responses to both pleasant and unpleasantemotional words than to neutral ones. These differ-ences induced by emotional arousal extended intothe late positive component (LPC) time range incontrols but were completely absent in psychopaths.
Weinstein (1995) assessed correlates of enhancedprocessing of threatening and nonthreatening verbalinformation in university students with elevated ornormal trait anxiety levels. Subjects read sentenceswith threatening and pleasant content that served asprimes for subsequently presented threat related,
Fig. 5. Difference maps of cortical activation for emotional minus neutral words in a covert evaluation task. Averaged activity in three
time windows is shown: P2 (180–280ms), P3 (280–400ms), and LPC (550–800ms). For P2 and P3 both pleasant and unpleasant words
are associated with larger positivities than neutral ones. In the LPC window only processing of pleasant words diverges from neutral.
The time course of the activity is shown at electrode Pz. Grand-averages from 26 subjects are shown.
156

neutral, or pleasant words. ERPs in response to thetarget words were assessed at Fz, Cz, and Pz, assubjects had to decide whether the target wordcontextually fit the previously shown sentence.Highly anxious subjects were reported to exhibit alarger frontal N1 in the threat-priming condition andan enhanced P400 (i.e., reduced N400, see below) ina later time window. In retrospect, a number ofmethodological problems seem to exist in this studyor the data presentation may contain errors. Forinstance, a large ERP offset at baseline is shown, thepresented condition means occasionally do notseem to correspond to the ERPs displayed and in-formation on linguistic properties of the stimuliused is missing. However, taken at face value, theresults indicate heightened selective attention to (N1)and facilitated semantic integration of (N400/P400)threat-related information in students with elevatedtrait anxiety levels.
A similar pattern of early ERP differences wasalso found in a study investigating the processingof pain-, body-related, and neutral adjectives inhealthy volunteers and prechronic pain patients.Knost et al. (1997) report enhanced N1 responsesto the pain-related stimuli at a left frontal sensor(F3) in the patient group. ERPs had been recordedfrom 11 mastoid linked scalp positions while sub-jects had to name words that were presented at theindividually determined perceptual threshold. Inboth groups, pain- and body-related words pro-duced larger positivities than neutral ones in alater time window (600–800ms) and were also as-sociated with larger startle eye-blink responses onseparately administered startle probe trials. Theauthors interpret their findings as evidence forpreconsciously heightened attention to unpleasant,pain-related stimuli in the patients.
These results were paralleled in an analogousstudy with chronic pain patients (Flor et al., 1997).Chronic pain patients had a larger frontal N1 re-sponse to pain-related words than comparison sub-jects. Additionally, a general hemispheric asymmetryemerged: N1 responses were larger over the right forpain-related words and over the left side of the headfor neutral words. The enhanced responses for pain-related words in the patient group were also visiblein a centro-parietally maximal N2 component. In theP2 window, right hemispheric responses to pain
words were likewise larger in the patients. In contrastto the first study, no differential effects of emotionalcategory were observed in subsequent time windows(P3 and LPC). The ERP results are taken to reflectheightened preconscious allocation of attention (N1)and stimulus discrimination (N2) to disorder-relatedwords in pain patients, but show no evidence offurther evaluative processing.
In a similar vein, Pauli et al. (2005) studied cog-nitive biases in panic patients and healthy volunteersanalyzing ERP responses from nine scalp channelsto panic-related unpleasant and neutral words thatwere presented, in separate runs, at individually de-termined perceptual thresholds and for 1000ms.Early ERPs differentiated panic patients from com-parison subjects. At threshold presentation, patientsshowed two enhanced early frontal positivities in re-sponse to panic words, one between 100 and 200ms(P2) and the other (P3a) between 200 and 400mspost-stimulus onset. Both early effects were absent atthe longer exposure duration and did not occur at allin the control group. Interestingly, subsequent pos-itivities between 400 and 600ms as well as between600 and 1000ms differentiated between panic wordsfrom neutral words in both groups and for bothpresentation durations. This pattern of data resem-bles the results by Knost et al. (1997).
Kissler and colleagues (in preparation) assessedprocessing biases in depressed patients and com-parison subjects using the above-described RSVPparadigm, recording from 256 scalp electrodes andcomparing the amplitude and scalp distribution ofthe previously described early negativity to pleas-ant, unpleasant, and neutral adjectives matchedfor length and frequency. Around 250ms afterword onset (see Fig. 3), comparison subjects dis-played the above-described left-hemispheric dom-inant enhanced negativity for emotional words,pleasant and unpleasant alike. Depressed patients,by contrast, exhibited this enhanced negativitysolely in response to the unpleasant words andonly in the right hemisphere.
Comparing early emotional and early semantic
processing
In sum, numerous studies have found early(o300ms) amplifications of ERPs in response to
157

words with emotional content compared with neu-tral words. The occurrence of such effects is re-markable since controlled conscious processinghas been suggested to arise only with the P3/N400components (Halgren and Marinkovic, 1995), im-plying that emotion can affect preconscious stagesof word processing.
Such effects appear to be more pronounced invarious clinical populations, reflecting heightenedsensitivity and orienting to unpleasant, disorder-related material (Weinstein, 1995; Flor et al., 1997;Knost et al., 1997; Pauli et al., 2005). Otherpatients groups, by contrast, seem to selectivelylack processing advantages for emotional words,psychopaths showing no cortical differentiationbetween emotional and neutral words (Williamsonet al., 1991) and depressed patients showing pref-erential processing of unpleasant but not pleasantwords (Kissler et al., in preparation). At any rate,processing biases in a number of clinical pop-ulations are reflected in their patterns of earlyresponses to pleasant, unpleasant, and neutralwords.
A debated issue in emotion research pertains towhether, when, and how cortical responses differas a function of arousal, valence, and additionalfactors such as dominance, or complex interac-tions of these. Here, the data must remain some-what inconclusive, as the studies discussed differedvastly on the dimensions included and assessmentmethods used. Studies that assessed their materialswith the semantic differential found that brain re-sponses differentiate between all dimensions andpolarities within the first 300ms after stimulus on-set (Chapman et al., 1978, 1980; Skrandies, 1998;Skrandies and Chiu, 2003). However, the arisingpattern of results is so complex that it is hard togauge the effect of each individual dimension onbrain responses. The vast majority of studies re-port generally larger ERP responses to emotionalthan to neutral words, with some studies reportingthese effects even in the absence of a task thatwould explicitly require processing of emotionalcontent or other types of semantic access (Begleiterand Platz, 1969; Bernat et al., 2001; Kissler et al.,submitted manuscript). However, occasionally,early emotion effects in word processing werefound restricted to situations where explicit
processing of the emotion dimension is requiredby the task (Begleiter et al., 1979).
Directly comparing the impact of pleasant vs.unpleasant word content yields mixed results, withsome studies finding larger early effects for pleas-ant words (Schapkin et al., 2000) and others largereffects of unpleasant ones (Bernat et al., 2001). Asmentioned above, the subjects’ clinical or motiva-tional status may bias their cortical responses ineither direction. Also, task characteristics as wellas the timing of stimulus presentation may have anadditional impact but so far the influence of theseparameters is not well understood.
Of note, some of the described effects occurredeven before 200ms after word onset, in a time rangein which from a traditional theoretical standpointmeaning-related processing differences would notbe expected (Schendan et al., 1998; Cohen et al.,2000).
These very early effects of emotional content onERP indices of visual word processing are ratherheterogeneous with regard to timing, locus, anddirection. Some of the inconsistencies are probablyrelated to differences in instrumentation and re-cording methodology, number of electrodes, andchoice of reference electrode(s) representing butthe most obvious differences.
The described studies also differ vastly in theway emotional content of the stimulus materialwas assessed as well as in the extent to which other,nonemotional, linguistic factors such as wordclass, length, and frequency or concreteness werecontrolled. Nevertheless, the bulk of the evidencesuggests that, indeed, under certain circumstancesthe emotional connotation of words can affecteven the earliest stages of preconscious sensoryprocessing.
Thus, the challenge is to specify under which cir-cumstances such emotional modulation of earliestprocessing may occur and what the underlyingmechanisms are. Two experimental factors arisefrom the reviewed studies that may contribute tothe emergence of very early emotion effects. First,very brief stimulus presentation, near or even belowthe perceptual threshold (Begleiter and Platz, 1969;Kostandov and Arzumanov, 1977; Chapman et al.,1978, 1980; Flor et al., 1997; Knost et al., 1997;Bernat et al., 2001; Ortigue et al., 2004; Pauli et al.,
158

2005) and second, repeated presentation of com-paratively small stimulus sets (Begleiter and Platz,1969; Chapman et al., 1978, 1980; Skrandies, 1998;Skrandies and Chiu, 2003; Ortigue et al., 2004).None of the cited studies have explicitly assessedthe effect of stimulus repetition on the latency ofemotional-neutral ERP differences. Our own stud-ies of repetition effects on negative difference wavesdistinguishing emotional from neutral contentaround 250ms after stimulus onset show no evi-dence of change within five repetitions. However,some of the cited studies used by far more than fivestimulus repetitions and studies of early semanticprocessing indeed suggest an effect of stimulus rep-etition on the timing on meaning-related differencesin cortical activity: Pulvermuller and colleaguesreport neurophysiological evidence of differences insemantic processing from 100ms post word onset(Pulvermuller et al., 2001a). They used a task inwhich a single subject was repeatedly, over severaldays, presented with a set of 16 words that she hadto monitor and hold active memory as responses tooccasionally presented new words were required.Thus, in above threshold presentation, preactiva-tion of the cortical networks coding for meaning byusing tasks that require continuous attention to andworking memory engagement with the stimuli aswell as use of many repetitions may foster earliestsemantic processing differences, nonemotional andemotional alike.
Further, a recent study on repetition effects insymbol processing found an increase in N1 ampli-tude (around 150ms) across three repetitions ofinitially unfamiliar symbol strings (Brem et al.,2005), supporting the view that stimulus repetitioncan amplify early cortical responses to word-likestimuli. Thus, repetition effects affecting emo-tional stimuli more than neutral ones as a conse-quence of differential initial capture of attentionand rapid perceptual learning may account forsome of the very early ERP effects in emotionalword processing.
Early effects of emotional content on brain re-sponses to subliminally or near-subliminally pre-sented stimuli have occasionally been accountedfor by fast, subcortical short-cut routes (see Wiens,this volume, for a discussion of issues of sublim-inal stimulus presentation). Evidence from animal
experiments and functional neuroimaging indeedreveals the existence of such subcortical ‘short-cut’routes in emotional processing, particularly offear-relevant stimuli (Davis, 1992; LeDoux, 1995;Morris et al., 1999). Direct pathways from the su-perior colliculi and the thalamus to the amygdalaand the cortex allow for the automatic processingof relevant stimuli outside of conscious awareness,preparing rapid behavioral responses. In humans,this subcortical pathway has been mapped byfunctional neuroimaging during fear conditioningof subliminally presented faces (Morris et al.,1999) as well as during the subliminal presentationof faces with fearful expressions (Liddell et al.,2005). On a cortical level, its activity may be re-flected in transient early responses. Brief, sublim-inal stimulation with fearful faces has recentlybeen shown to result in a transient enhancement ofthe N2 and early P3a components, which, how-ever, did not continue in later N4/P3b/LPC win-dows. For supraliminal stimulation, conversely,N4/P3/LPC but not N2 components responded toemotional content (Liddell et al., 2004), suggestingthe operation of a slower, conscious processingand evaluation route. Conceivably, subliminalstimuli receive a temporally limited amount ofprocessing that wanes if it is not confirmed byfurther supraliminal input, much like in the case ofsubliminal priming (Greenwald et al., 1996; Kieferand Spitzer, 2000). Recording ERPs during sub-liminal and supraliminal semantic priming, Kieferand Spitzer observe decay of subliminal semanticactivation within 200ms, a delay at which suprali-minal priming effects can still be robustly demon-strated. A plastic, maladaptive downregulation ofsubcortical excitability may account for early re-sponsiveness to unpleasant and disorder-relatedwords in clinical populations (see e.g. Pauli et al.,2005 for supportive evidence).
Clearly, at present the operation of a fast sub-cortical route from the thalamus and the amygdalain emotional word processing that could accountfor near or subthreshold emotion effects in visualword processing remains a speculative conjecture.A most critical point is that such a mechanismwould require at least basic ‘reading abilities’ inthe thalamus. While the case for stimuli such asfaces or threatening scenes that by some are
159

assumed to be part of our ‘evolved fear module’(Ohman and Mineka, 2001) can be made muchmore easily, many would have a hard time believ-ing in rather sophisticated subcortical visual ca-pacities allowing for the discrimination of writtenwords. On the other hand, subcortical structuresare also subject to modifications by learning, andby the time people take part in experiments theywill usually have had about two decades of readingexpertise.
So far, most of the evidence for the subliminalprocessing of emotional stimuli is based on studieswith aversive material. Accordingly, the above-reviewed studies evidence extremely early effectsprimarily for unpleasant words (Flor et al., 1997;Knost et al., 1997; Bernat et al., 2001).
An alternative explanation of some of theseearly effects of enhancement by emotional contentin visual word processing that would not rely onsubcortical by-pass routes and therefore on sub-cortical vision is reentrant connections between theso-called visual word form area (VWFA) and theemotion processing system.
During visual word recognition the earliest ac-tivation of an invariant version of the visual wordform (i.e. the font-, size-, position-invariant repre-sentation of the string of letters) occurs from about100ms after stimulus onset (Sereno et al., 1998;Assadollahi and Pulvermuller, 2003). Form invar-iant, abstract representations of highly over-learned visual objects such as words and faceshave been found to originate in the fusiform gyrus(Haxby et al., 1994; Chao et al., 1999; Cohen et al.,2000; Dehaene et al., 2002). Electrophysiologicalevidence with regard to the onset of word-specificeffects of fusiform activity varies, with some au-thors reporting onsets around 120ms (Tarkiainenet al., 1999; Assadollahi and Pulvermuller, 2001)and others somewhat later around 170ms (Bentinet al., 1999; Cohen et al., 2000). Timing differencesmay be partly attributable to differences in wordfamiliarity across experiments (King and Kutas,1998).
Immediately after access of the visual word form,meaning can be activated: Assadollahi and Rocks-troh (2005) showed that activation differences dueto super-ordinate categorical differences (animalsvs. plants) can be found in left occipitotemporal
areas between 100 and 150ms after word onset,whereas activation differentiating between subordi-nate categories was evident only from 300ms on.Dehaene (1995) observed the earliest ERP differ-ences between words of different categories (verbs,proper names, animals), 250–280ms after word on-set. Semantic category differences were reflected inthe scalp distribution of a left occipitotemporalnegativity. Using RSVP designs a similar occipito-temporal negativity has been identified. This nega-tivity has been termed the ‘recognition potential’(RP). It is sensitive to semantic aspects of visualword processing and has its maximum around250ms after word (Rudell, 1992; Martin-Loeches etal., 2001; Hinojosa et al., 2004). The ‘RP’ respondsto manipulations of depth of semantic analysis, itsamplitude increasing with the meaningfulness andtask-relevance of the presented word. Source anal-ysis has placed the origin of the RP in the fusiformgyrus (Hinojosa et al., 2001). Results from our lab-oratory are consistent with the view that a word’semotional connotation enhances the associated rec-ognition potential (see Fig. 4).
Thus, a word’s emotional connotation could bedirectly connected to the abstract representation ofits visual form. Moreover, the combined evidencesuggests that emotional content amplifies earlystages of semantic analysis in much the same wayan instructed attention enhancing processing taskwould.
If enhanced semantic processing is an importantmechanism by which emotional content affectsvisual word processing, again, the question arisesas to the causative mechanism: back-projectionsfrom the anterior cingulate and the amygdala maygive rise to such processing enhancements. In sup-port, amygdala lesions impair the enhanced detec-tion of unpleasant words in an RSVP attentionalblink paradigm but not of identification enhance-ments caused by manipulation of target color(Anderson and Phelps, 2001). Thus, assuming thatthe amygdala plays a pivotal role in the preferen-tial processing of emotional words as recently sug-gested by several neuroimaging and lesion studies(Isenberg et al., 1999; Anderson and Phelps, 2001;Garavan et al., 2001; Hamann and Mao, 2002;Naccache et al., 2005), an alternative model to theabove described thalamo-amygdalo-cortical route
160

could account for most of the data. Emotionalamplification of semantic processing would occurafter initial stimulus identification, caused by bi-directional reentrant communication between cor-tical regions and the amygdale (Amaral et al.,2003). Crucially, cortical analysis would precedeand spark subcortical amplification of corticalprocessing. Clearly, a theoretically crucial priorityfor future research is to determine the timing ofsubcortical mechanisms in relation to cortical en-hancement of ERP responses to emotional words.
Unlike for other semantic categories such asmovement-related verbs (Pulvermuller et al., 2000,2001b), ERP data for emotional words so far sug-gest little consistent emotion-specific change in to-pography (but see Skrandies, 1998; Skrandies andChui, 2003; Ortigue, 2004). A distinct, emotion-as-sociated topography might point to the existence ofa homogeneous emotion lexicon localizable in dis-tinct neuronal populations of the brain as has beensuggested for other semantic categories (Martin etal., 1996). Rather, emotional content seems to am-plify cortical word processing, much in the sameway as it enhances picture (Junghofer et al., 2001)or face processing (Schupp et al., 2004). However,functional neuroimaging techniques with betterspatial resolution of especially deep cortical andsubcortical structures (see Cato and Crosson, thisvolume) and the more consistent use of dense arrayEEG studies (Ortigue et al., 2004) may provide ad-ditional information.
Late components (after 300ms)
Healthy volunteers
In relation to traditional stages of visual wordprocessing, effects occurring later than 300msafter word onset are less puzzling than the previ-ously discussed early ones. Enhanced late positivi-ties in response to emotional word content havemost often, but not invariably, been found. Inseveral of the already discussed studies reportingearly ERP modulations as a function of emotionalcontent, later enhanced positivities associated withthe emotional content of the word stimuli are alsoapparent. For instance, in the data shown by
Chapman et al. (1978, 1979, 1980; see above) apositivity occurring around 300ms is discernibleand appears to be primarily related to the potencydimension extracted from their data.
Using materials from the Begleiter et al. (1969,1979) and Chapman et al. (1978, 1979) studies,Vanderploeg et al. (1987) assessed ERP responsesto visually presented emotional (20 pleasant, 20unpleasant) and 20 neutral words and face draw-ings (two per emotion category), which were eval-uated during viewing. The EEG was recorded fromsix electrodes referenced to linked ears in 10 malesubjects. During viewing, the visual stimuli werepresented for either 80ms (words) or 100ms (faces).In the conditioning phase, the face drawings wereshown for 1500ms. For both faces and wordsclearly discernible emotion-category dependentdifferences in ERP tracings appear from around300ms after stimulus onset as parietal positivities.A small but significant effect of emotional conno-tation of words but not faces on the spatial distri-bution of the ERP was also evident in the P2window. Interestingly, although sizeable in appear-ance, the P3 effect of emotional connotation did notreach statistical significance for words. For a laterpositivity (positive slow wave/late positive complex)a similar result was obtained; although visible in thepresented grand-averages, the difference in parietalpositivity between emotional, pleasant and unpleas-ant, and neutral words does not reach significancein an analysis of the corresponding PCA factorswhile it does for faces. The authors, in line withLifshitz’ (1966) early finding, suggest that wordsmay be less powerful (or more heterogene-ously evaluated) emotional stimuli than pictures(Vanderploeg et al., 1987). Thus, ERPs from10subjects may not yield enough statistical powerto assess connotation-dependent ERP differences,particularly in studies using comparatively sparseelectrode arrays. Also, the perceptual variance be-tween 20 words of a category may be higher thanamong two faces. Differential effects may, there-fore, also result from the greater consistency orhigher frequency of occurrence of the faces.
Indeed, subsequent studies have found robusteffects of emotional connotation on later ERPcomponents. For instance, Naumann et al. (1992)investigated late positive potentials to adjectives
161

varying in emotional content. Their key idea wasthat using ERPs it should be possible to dissociateemotional and cognitive processing and that, fol-lowing LeDoux (1989), cognitive and emotionalprocessing systems should be functionally andneuronally separable as reflected in distinct ERPscalp topographies. In an initial experiment, 30prerated pleasant, unpleasant, and neutral adjec-tives were presented to 14 subjects who had toeither evaluate the words as pleasant, unpleasant,or neutral (affective task) or determine whether aword’s length was longer, shorter, or equaled sixletters (structural task). The EEG was recordedfrom three midline electrodes (Fz, Cz, and Pz),which were referenced to the left mastoid. ERPswere assessed between 300 and 700ms after wordpresentation for the P3 component and between700 and 1200ms for the later positive slow wave.For both components and all word categories,ERP amplitudes were more positive going for theaffective than for the structural task, particularlyat electrodes Fz and Cz. Moreover, P3 amplitudeswere also generally more positive in response toemotional than neutral adjectives. The spatial dis-tribution of the subsequent slow wave componentvaried with emotional category, displaying largeramplitudes at Pz than at Cz and Fz for pleasantand unpleasant words but having equal amplitudesat all three electrodes for the neutral words. Thispattern was taken as evidence for the hypothesizedseparateness of affective and cognitive processingsystems.
Naumann et al. (1992) replicated this result in asecond experiment having calculated an ideal sam-ple size of 106 subjects, minimizing the likelihoodof false-negative results. In the replication study abetween groups design was used, assigning 53 sub-jects each to either the structural or the affectivetask. Again, for both components and all wordtypes, more positive frontal ERPs were obtainedfor the affective than for the structural task. TheP3 component was larger for both pleasant andunpleasant than for neutral adjectives. Further-more, positivities in response to emotional adjec-tives were particularly pronounced at Cz and Pz,and this gradient was most evident for the pleasantwords. For the slow wave, the scalp distributionlikewise exhibited a parietal maximum and this
was more pronounced for the emotional than forthe neutral adjectives. Thus, overall, an emphasison emotional processing (affective task) caused ananterior shift of the scalp distribution. Further-more, regardless of task, emotional stimuli led tomore pronounced parietal peaks than neutralones. Again, the authors interpreted their resultsas evidence for a functional and structural distinc-tiveness of affective and cognitive functions in thehuman brain as suggested by LeDoux (1989).
However, a third demonstration of dissociableaffective and cognitive processes in visual wordprocessing failed. Naumann et al. (1997) examineda sample of 54 students in three different tasks,namely letter search (structural task), concrete–ab-stract decision (semantic task), and an unpleas-ant–neutral decision (affective task) on a set ofnouns varying in emotional content. Fifty-sixnouns were used that could be divided into sub-sets of seven words unambiguously belonging toone of eight possible combinations of these at-tributes. ERPs were now recorded from nine scalplocations (Fz, Cz, Pz, and adjacent left and rightparallels). There was indeed a considerably largerP3 for unpleasant compared to neutral words, al-beit the effect was not general but restricted to theaffective task. This casts doubt on the assumptionthat cognitive and emotional word processing op-erate along completely separable routes and raisesthe question to what extent larger late positivepotentials to emotional word stimuli occur outsidethe attentional focus. Given the early effects(o300ms) reported above, it would have been in-teresting to analyze the data with a focus on earlyand possibly automatic impacts of emotion onword processing.
Naumann et al. (1997) raise a number of con-ceivable reasons for the reduction of the effect,favoring a familiarity-based explanation. In theexperiments that had yielded ‘uninstructed’ andtopographically distinct late responses, subjectshad been familiar with the stimuli beforehand.Thus, the affective differences between the stimulimay have already attracted the participants’attention. Moreover, the new design reduced theprobability of occurrence for an emotional word,possibly making this dimension less salient;although the converse hypothesis, based on an
162

oddball effect, would be equally plausible. More-over, in their initial studies, Naumann et al. (1992)had used adjectives that may produce somewhatdifferent effects, given that ERP differences be-tween word classes have been reported (e.g. Fed-ermeier et al., 2000; Kellenbach et al., 2002).
Fischler and Bradley (this volume) report on aseries of well-controlled studies where effects ofword emotionality on late positivities are consist-ently found for both pleasantly and unpleasantlyarousing words when the task requires semanticprocessing of the presented words but not otherwise.
Some studies also show larger late positive po-tential effects restricted to pleasant words (Schap-kin et al, 2000; Herbert et al., 2006), which weremissing in Naumann’s 1997 study.
For instance Schapkin et al. (2000, see above) re-port larger P3 and late positive slow wave responsesto pleasant compared to both neutral and unpleas-ant words during evaluative decision. As mentionedabove, Herbert et al. (2006) report a study whereearly ERP responses (P2, P3a) reflected the arousaldimension of the words, differentiating both pleas-ant and unpleasant from neutral words. The laterLPC, however, differentiated pleasant from un-pleasant stimuli and was larger for pleasant words(see Fig. 5). Bernat et al. (2001), on the other hand,report enhanced responses to unpleasant as com-pared to pleasant words across the entire analysiswindow, until 1000ms after word onset encompass-ing P3 and LPC.
Schapkin et al. (2000) additionally assessed latenegativities that were labeled N3 (around 550ms)and N4 (around 750ms), finding no effect of emo-tional content. Data from our own studies doshow a small effect of emotional content on N4amplitudes with larger N4 to neutral than to emo-tional words possibly reflecting a contextual ex-pectancy for an emotional content caused byunequal stimulus probabilities. In both studies(Schapkin et al, 2000; Kissler et al., submitted),two-thirds stimuli had emotional content (pleasantor unpleasant), only one-third was neutral.
Late components — clinical studies
Weinstein (1995) is one of the few reports of amodulation of integration of emotionally charged
words following a sentence context. Students withhigh-trait anxiety levels had a reduced N400 (or, inWeinstein’s terminology enhanced P400) to wordsfollowing a threatening sentence context, indicat-ing facilitated integration of information withinthreatening contexts. An alternative interpretationmight suggest enhanced sustained attention tothreatening information in highly anxious subjects,if the potential described were taken to resemble aP3/LPC component, which is not entirely clear onthe basis of the presented data.
Personality-dependent changes in late corticalresponses to emotional words have been subse-quently replicated: Kiehl et al. (1999) tried to ex-tend Williamson et al.’s (1991, see above) results ofdeficient early (P2) and late (LPC) ERP responsesto emotionally charged words in psychopathicsubjects. They assessed similarities and differencesin the processing of abstract–concrete vs. pleas-ant–unpleasant words in psychopaths and com-parison subjects. To address the processing ofemotional words a pleasant–unpleasant decisiontask was used; although the initial study had notrevealed any valence differences. The altered taskwas apparently motivated by clinical observationssuggesting that psychopaths have difficulty in un-derstanding abstract information and in distin-guishing pleasant from unpleasant valence. Stimuliwere controlled for word length and frequency,syllable number, and concreteness. Word presen-tation was extended to 300ms and words werepresented only once, centrally and in a horizontalformat. EEG was recorded from nine scalp posi-tions, again with a linked mastoids reference.Analyses now focused on a 300–400ms post-stim-ulus window and a LPC window (400–800ms).Behaviorally, in both groups responses to pleasantwords were faster and more accurate than those tounpleasant ones. Cortically, an N350 componentdifferentiated between pleasant and unpleasantwords but not between psychopaths and nonpsy-chopaths, being across groups larger for the pleas-ant words. Moreover, the valence differentiationwas more pronounced over the left hemisphere. Inthe later time window (400–800ms), unpleasantwords elicited more positive going brain wavesthan pleasant ones. This left-hemispheric domi-nant differentiation was absent in psychopaths. In
163

effect, ERPs to unpleasant words were more pos-itive than ERPs to pleasant words across both timewindows, and the differentiation was reduced inpsychopaths. It is unclear how the ERP patternsrelate to the behavioral data (both groups werefaster and more accurate for pleasant). But morepositive-going late potentials for unpleasant stim-uli in a binary pleasant–unpleasant decision are inline with data from Bernat et al. (2001). During alexical decision task, Williamson et al. (1991) re-ported a larger LPC to emotional than to neutralwords in nonpsychopathic subjects and to a lesserdegree in psychopaths, but no differentiation bet-ween the pleasant and unpleasant words. Schapkinet al. (2000) and Herbert et al. (2006), by contrast,report larger late positivities for pleasant in com-parison to both neutral and unpleasant words.Note, that neither the Kiehl et al. (1999) nor theBernat et al. (2001) studies report data on neutralstimuli.
Comparing late emotional and late semantic word
processing
A considerable number of studies have found am-plifying effects of emotional word content onelectrophysiological cortical activity later than300ms after word onset. In contrast to the veryearly effects, they occur in a time range wheremodulation of cortical responses by word meaningis not unusual in itself. By 300–400ms after stim-ulus onset ERP tracings reflect conscious process-ing stages (Halgren et al., 1994a, b) and clearlyvary with semantic expectancy (Kutas and Hill-yard, 1980, 1984), task relevance (Sutton et al.,1967), or depth of mental engagement (Dien et al.,2004). Thus, it is not surprising that ERPs in thistime range can reflect processing differences bet-ween words of different emotional content. Froma semantics perspective, N400 might represent anappropriate ‘classical’ ERP component to assessfor emotion effects. Indeed, some studies havefound modulations of N400- or N400-like ERPresponses to words of emotional categories (Will-iamson et al., 1991; Weinstein, 1995; Kiehl et al.,1999). However, in line with ERP studies of affec-tive processing of faces (Schupp et al., 2004) and
pictures (Keil et al., 2002), most researchers fo-cused on an analysis of late positivities. The com-parative paucity of reports on N400 modulationby emotional word content may partly reflect abias on the part of the investigators and appearsurprising in view of the fact that N400 is oftenregarded as ‘the electrophysiological indicator’ ofsemantic processes in the brain. On the otherhand, it is becoming increasingly clear that theN400 response does not index lexical access or se-mantic processing per se but reflects semantic in-tegration within a larger context, created by eitherexpectations on sentence content or other contex-tual constraints within experiments (Kutas andFedermeier, 2000). Thus, it is reasonable to as-sume that single-word studies will only result inN400 modulations if strong expectations on emo-tional word content are established. Priming stud-ies or experiments establishing an emotionalexpectation within a sentence context may pro-vide a better testing ground for the issue of N400modulations by emotional word content. Indeed,Weinstein (1995) followed this rationale establish-ing emotional expectations on a sentence level.Recent work from our laboratory also shows N400modulation by emotional content in a lexical de-cision task where an emotional expectation (pleas-ant or unpleasant) was established by a precedingemotional picture. Of note, the pictures were ofsimilar emotional connotation as the subsequentadjectives but the words were not descriptive of thepicture content (Kissler and Kossler, in prepara-tion). A transient mood induction may have me-diated the effect; recently, effects of subjects’emotional states on semantic processing have beenreported (Federmeier et al., 2001). When subjectswere in a mildly positive mood, their semanticprocessing was facilitated as reflected by a smallerN400 potential to more distant members of givencategories than when in a neutral mood. Thus, anumber of studies suggest that both a word’semotional content and a subject’s emotionalstate may affect the N400 ERP response (but seeFischler and Bradley, this volume).
Still, so far the most consistently reported latereffects of emotional word categories on the ERPare seen in broadly distributed late positivities witha parietal maximum (see also Fischler and Bradley,
164

this volume). Such late positivities have generallynot been associated with specific aspects of seman-tic processing but rather with task demands suchas attentional capture, evaluation, or memory en-coding. In neurolinguistics, late positivities haverepeatedly been suggested to index syntactic reanal-ysis following morphosyntactic violations (Osterh-out et al., 1994; Friederici et al., 1996; Hagoort andBrown, 2000). Yet, some studies also report mod-ulations of late positivities by semantic attributes oflanguage. For instance, in antonym processingdifferential P3 and LPC responses were found, de-pending on whether a word contained a given at-tribute or lacked it (Molfese, 1985). Contextualsemantic constraints and stimulus abstractness havealso been reported to affect late positivites (Hol-comb et al., 1999). Both contextually expected andunexpected sentence-final words were associatedwith larger positivities than contextually uncon-strained words, the effect being even more pro-nounced when the abstract words were contextuallyunexpected. Munte et al. (1998) also find late pos-itive responses to language semantics, thereby chal-lenging the account of specific morphosyntactic latepositive shifts and corroborating the view that latepositivities reflect mental engagement and effortfulprocessing across a wide range of higher cognitivefunctions (Dien et al., 2004).
Late positivities are likely to share a proportionof neural generators and differ on others, reflectingthe extent to which the tasks that elicit them shareor draw on different neural systems. Thus, topo-graphically distinct late positive shifts may relateto different aspects of cognitive and emotionalfunctioning, as suggested for instance by Nau-mann et al. (1992). However, in order to unam-biguously elucidate topographic changes thatreflect shifts in neural generator structure, simul-taneous recordings from dozens of electrodes andadvanced data analysis techniques are necessary(see also Junghofer and colleagues, this volume).From the extant studies on the emotional modu-lation of late components in word processing, it ishard to gauge the extent to which emotion inducesgenuine topographic changes indicative of the re-cruitment of additional distinct cortical structuresor purely amplifies the activity of a unitaryprocessing system.
In emotion research, larger late positivities haveconsistently been shown during free viewing ofemotional vs. neutral pictures (Keil et al., 2002)and faces (Schupp et al., 2004). If primary tasksdistract participants from the emotional content ofthe visual stimuli, late positivities to emotionalstimuli are often diminished reflecting competitionfor attentional resources. The degree to which andcircumstances under which emotion and attentioncompete for resources, have additive effects, oroperate in parallel is a matter of ongoing debate(see Schupp, et al., this volume, for a discussion).
For visually presented word stimuli the pictureis similar; when the primary task requires an eval-uative emotional decision (pleasant–unpleas-ant–neutral, emotional–neutral) emotional words,like pictures or faces, are consistently associatedwith larger late positivities than neutral ones.When the primary task requires structural stimulusprocessing the evidence is mixed, with some studiesstill finding larger positivities in response to emo-tional stimuli (Naumann et al., 1992) while othersdo not (Naumann, 1997). During free viewing, arecent study (Kissler et al., submitted manuscript)finds a larger LPC to emotional words, suggestingthat when subjects are free to allocate theirprocessing resources as they wish, they processemotional words more deeply than nonemotionalones. Our results also indicate that late responses,around 500ms, may be more affected by explicitattentional tasks than the simultaneously observedearly effects around 250ms. During lexical deci-sion (Wiliamson et al., 1991) and naming tasks(Knost et al., 1997; Pauli et al., 2005), emotionalwords have also been found to be associated withlarger late positivities (but see Fischler and Brad-ley, this volume). Thus, when the task allows for oreven requires semantic processing, emotionalwords are processed more deeply than neutralones. When the task requires structural processingthis processing advantage is considerably dimin-ished (Naumann et al., 1997).
Clearly, the extent to which larger late positivitiesto emotionally relevant words are driven by arousal,valence, or additional subject-, task-, or situation-specific factors is not quite settled. The matter iscomplicated by the fact that studies differed on theinstruments used to assess emotional word content
165

and the extent to which the pleasant and unpleasantdimension were differentiated or collapsed into one‘emotional’ category. A fair number of studies em-ployed the empirically well-founded semantic differ-ential technique to assess emotional content, or thetwo-dimensional arousal � valence space, yet othersdo not even report the criteria by which the emo-tional content of the material has been determined.
Although multidimensional models of affect areempirically well founded and the use of numericalrating scales allows for the rapid assessment oflarge numbers of stimuli on many dimensions, aninherent problem with Likert-type scaling remains.Such scaling techniques assume that subjects willmeaningfully assign numbers to psychologicalstimuli, such that the quantitative relationshipsbetween the numbers will correctly reflect the psy-chologically perceived relationships among thestimuli, including conservation of distance or con-servation of ratio, yielding interval or even ratioscales. But these assumptions do not always hold,such that it is unclear whether the psychologicaldistance between stimuli rated 2 and 4 on a givenscale is really the same as between stimuli rated 6and 8 (Luce and Suppes, 1965; Kissler and Bauml,2000; Wickelmaier and Schmid, 2004). Moreover,the relationship between behavioral ratings andphysiological impact of emotional stimuli is likelyto be nonlinear.
Nevertheless, the bulk of the data corroboratesthe view that during earlier stages of processing,emotion acts as a nonvalence-specific, arousal-driven alerting system (see above). During laterstages of processing (4300ms), the patterns foundare more varied and may reflect flexible adapta-tions to contextual factors. In support, Herbertet al. (2006) recently found arousal-driven ampli-fication of cortical responses to both pleasantand unpleasant words within the first 300ms and adivergent pattern that favors the processingof pleasant material in a later time window (seeFig. 5). Keil (this volume) discusses a number oftask factors that contribute to processing advan-tages for pleasant or unpleasant material in turn.For language material with emotional content, ageneral pleasant–unpleasant asymmetry in emo-tional processing may be important: At low levelsof arousal, a ‘positivity offset’ is often found in
that the approach system responds more stronglyto relatively little input. The withdrawal system inresponse to unpleasant input, in turn, is activatedcomparatively more at high levels of arousal,this latter process being termed ‘negativity bias’(Caccioppo, 2000; Ito and Caccioppo, 2000). Vis-ually presented words are likely to constitute less-arousing stimuli than complex colored pictures,i.e., the word ‘cruel’ will be less arousing than aphotograph of a corresponding scene, even if bothstimuli receive comparable ratings. Therefore, inthe absence of strong unpleasant personalassociations for a given word, which may well bepresent in various clinical populations (see above),a ‘positivity offset’ for written verbal materialmight be expected. Corresponding data are re-ported, for instance, by Schapkin et al. (2000) orHerbert et al. (2006).
Like for the early effects, the question arises howlate effects of emotional word content on ERPscome about; subcortical activity has again been im-plicated. Nacchache et al. (2005) have recently forthe first time recorded directly from the amygdalafield potentials in response to emotional words.Three epilepsy patients with depth electrodes im-planted for presurgical evaluation performed anevaluative decision task (threatening–non-threaten-ing) on a series of threat or nonthreat words pre-sented subliminally or supraliminally. In all threepatients, larger amygdala potentials to threat thanto nonthreat words could be identified around800ms after word presentation in the subliminaland around 500–600ms in the supraliminal condi-tion. The study is pivotal in that it both directlymeasures amygdala activity during emotional wordprocessing and provides clues as to the timing of thisactivity. As detailed before, subcortical, primarilyamygdala activity may be a source of cortical am-plifying mechanisms in response to emotional stim-uli visible in ERPs. Amygdala activity measured bydepth electrodes around 600ms after stimulus onsetmay provide a basis for LPC amplifications evidentin the surface ERP. However, the timing of the re-sponses poses new puzzles. If amygdala activity inemotional word processing onsets around 600ms,how are early effects of emotional word contentgenerated (see discussion above)? Amplified corticalERP responses reflect the activation of larger
166

patches of cortex, indicating spread of activation ina more densely packed neural network. These resultfrom life-long associative learning mechanisms. Theeffects of emotional learning can be seen in ampli-fied cortical ERP tracings whenever the correspond-ing semantic network is accessed. Subcorticalmechanisms might be active primarily in the acqui-sition of emotional semantics, reflecting the role ofthe amygdala in emotional learning even of abstractrepresentations (Phelps et al., 2001). Their impactmay be attenuated once a representation has beenacquired. Clearly, elucidating the mechanisms bywhich amplified responses to emotional words aregenerated is a vital issue for future research.
In sum, a considerable number of studies showenhanced late positive responses when peopleprocess emotionally laden words, pleasant andunpleasant alike. The responses are not as large asfor pictorial or face stimuli, but they have beenreliably demonstrated across numerous studies.Major challenges for future research remain in de-termining the relative role of arousal and valenceand their interactions with task demands. Finally,the question to what extent and at which points intime, processing of emotional words recruits spe-cific cortical and subcortical neural circuitries mer-its further scientific attention.
Processing emotional words — electrophysiological
conjectures
The above review demonstrates that emotionalword content can amplify word processing atall stages from access to word meaning (around200ms), to contextual integration (around400ms), evaluation, and memory encoding(around 600ms). Occasionally, emotionality-de-pendent enhancements have been reported evenbefore 200ms.
In neurolinguistics, the timing of lexical access isheatedly debated. The reports about differentpoints in time where some aspects of the lexicalinformation on a word are accessed vary between100 and 600ms. Importantly, the interpretation ofthe N400 has shifted from an index of semanticaccess to a signature of the interaction betweensingle word semantics and context. Accordingly, a
growing body of evidence demonstrates that someaspects of word meaning must be active before theN400 is elicited.
We propose that there is no fixed point in timewhere all semantic information is equally availa-ble. Instead, subrepresentations can become acti-vated in a dynamic, possibly cascaded manner.Variations in timing of semantic signatures couldbe interpreted in the light of an internal structureof the entry of a mental lexicon. Different aspectsof semantics could be flexibly prioritized, depend-ing on context, task, and motivation of the subject.If the tenet of a monolithic lexical entry is givenup, the internal structure of a word’s lexical rep-resentation can be assessed by investigating thetiming by which each subrepresentation is availa-ble (Assadollahi and Rockstroh, 2005) or the con-textual constraints that lead to the activation of aparticular subrepresentation at a given point intime (Sereno et al., 2003).
Emotional semantics may be special; their con-nection to biologically significant system statesand behavioral output tendencies may ensure mostrapid activation of the underlying neural networkrepresentations. A number of studies endorse a si-multaneous impact of emotional word content onboth cortical and peripheral responses (Flor et al.,1997; Knost et al., 1997; Herbert et al., 2006),corroborating the view that the neural networksrepresenting emotional concepts dynamically linksemantic and response information (Lang et al.,1993, see Fig. 3). Many studies also show surpris-ingly early cortical responses to emotional wordcontent. Thus, the subrepresentation of a word’semotional content may sometimes, though clearlynot always, be activated before other lexical in-formation is available, for example, whether aword denotes an animate or an inanimate entity.Depending on personality, context, and task, emo-tional semantic networks may become activatedrapidly or gradually, operate over several hundredmilliseconds or their activation may decay quicklywhen not currently relevant.
The specification of experimental factors con-tributing to the generation of arousal-, valence-, oreven potency-specific effects or to the temporalgradient of the processing enhancement caused byemotional connotation is lacking detail. As a
167

working hypothesis, emotional content amplifiesword processing at the earliest stage at which thevisual word form is assembled, i.e., no sooner thanabout 100ms after word presentation. Learning,top-down and priming processes may dynamicallymodify the time point at which the activation ofthe representation takes place (see above). Arousalis likely to govern this amplification process withinthe first 300ms, reflecting a general orienting func-tion of emotion. Under specific experimental ma-nipulations, such as very brief stimulus duration,or in populations with pronounced negativeprocessing biases, earliest advantages for unpleas-ant material may be obtained, possibly reflectingmodality-independent operations of a rudimentaryrapid threat detection system (Ohman and Mine-ka, 2001). In general, the neural representations ofabstract linguistic features are less likely to be ac-tivated by such a superfast detection system.
Later than 300ms after stimulus onset the be-havioral relevance of the stimulus in a given
situation is likely to determine its further process-ing, with tasks that require processing of semanticcontent supporting sustained enhancement ofemotional stimuli and contextual effects determin-ing possible processing advantages for either va-lence. Under circumstances where structuraloutweighs semantic processing or where the stim-uli occur very briefly and are not followed by con-firmatory input, only transient enhancement ofearly brain responses and less, if any, amplificationin later time windows will result.
Subcortical structures, most prominently theamygdala, have been implied at all stages of thisemotional content-driven amplification process,but the dynamics of the interplay between subcor-tical and cortical structures in processing emo-tional words await specification. Combiningelectrophysiological, functional magnetic reso-nance, and lesion approaches within a networktheory of emotion and language will be useful toclarify these issues.
168

Appendix
Table
A1.A
summary
ofexperim
entaldesign,recordingparametersandresultsofthereviewed
studiesoneffectsofem
otionalcontentonERP
measuresofword
processingin
healthyvolunteers.
Stu
dy
Subje
cts
N,
sex
Task
(s)
Sti
muli
Presentation
Rec
ord
ing
and
analy
sis
ER
Pef
fect
s
Word
class
Emotionalcontent
Controlofadditional
stim
ulusparameters
Stimulusduration
Positionofpresentation
Stimulusrepetitions
Number
ofelectrodes
Reference
Epoch
duration(post-
stim
ulus)
Statisticalanalysis
Lifshitz(1966)
1,male
Viewing,mem
orizing
Word
class
notindicated
Emotionalcontent
determined
byauthor:40
‘dirty
words’40neutral
words
Controlled
for:word
length
1000–2000ms
Centrally
Number
ofrepetitionsnot
indicated
4 Left-hem
ispheric
leads,
reference
unclear
500ms
Visualinspection
Visualcomparisondirty
wordsversusneutral
words:nomarked
differencesvisible.
Begleiter
andPlatz
(1969)
18,allmales
Viewing,word
naming
Nouns
Emotionalcontent
determined
byauthors:2
‘taboo’words(shit,fuck)2
‘neutral’words(tile,
page)
blankflash
Controlled
for:word
length
10ms
Centrally
20repetitions
1(O
2)
Linked
ears
reference
1024ms
ANOVA
t-test
Amplitude:
‘taboo’4
neutral&
flash
Naming4viewing(100–200msand
200–300ms)
Latency:‘taboo’o
neutral&
flash
(300–400ms)
Kostandovand
Arzumanov(1977)
37,sexnotindicated
(‘allin
state
ofjealousy’)
Experim
ent1supraliminal
23.
Experim
ent2subliminal,
14.
Covertidentificationand
countingofstim
ulusrepetitions
‘Conflict’and‘neutral’
words
Nofurther
detailson
experim
entalstim
uligiven.
Additionalparameter
controlunclear
Experim
ent1:200ms
50repetitions?
Experim
ent2:15ms
50repetitions?
2(C
z,O1)
Leftmastoid
reference
1000ms
t-tests
Experim
ent1:Supraliminal:N2(�
220ms)
Emotionalcontentonpeakamplitude&
latency
n.s.
P3a(270–320ms)
O1:Peakamplitude:
emotional4
neutral
Peaklatency:em
otionalo
neutralCz:
n.s.
Experim
ent2:Subliminal:N2(�
220ms)
Pea
kam
pli
tude
O1
and
Cz:
emoti
onal4
neu
tral
P3a(�
300ms)
O1andCz:
Peakamplitude:
emotional4
neutral
Peaklatency,n.s.
169
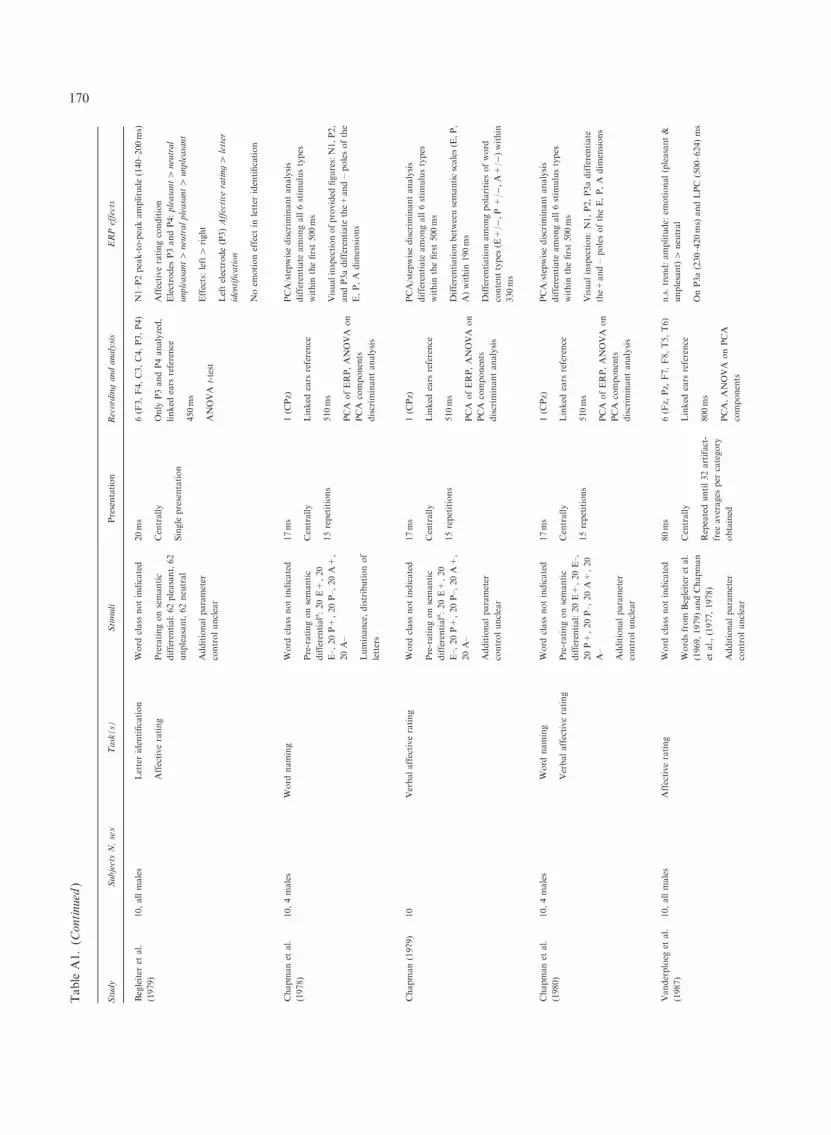
Begleiter
etal.
(1979)
10,allmales
Letteridentification
Affectiverating
Word
class
notindicated
Preratingonsemantic
differential:62pleasant,62
unpleasant,62neutral
Additionalparameter
controlunclear
20ms
Centrally
Single
presentation
6(F3,F4,C3,C4,P3,P4)
Only
P3andP4analyzed,
linked
ears
reference
450ms
ANOVA
t-test
N1–P2peak-to-peakamplitude(140–200ms)
Affectiveratingcondition
Electrodes
P3andP4:
ple
asa
nt4
neu
tral
unple
asa
nt4
neu
tral
ple
asa
nt4
unple
asa
nt
Effects:left4
right
Leftelectrode(P3)
Aff
ecti
vera
ting4
lett
er
iden
tifi
cati
on
Noem
otioneffect
inletter
identification
Chapmanet
al.
(1978)
10,4males
Word
naming
Word
class
notindicated
Pre-ratingonsemantic
differentiala:20E+,20
E–,20P+,20P–,20A+,
20A–
Luminance,distributionof
letters
17ms
Centrally
15repetitions
1(C
Pz)
Linked
ears
reference
510ms
PCA
ofERP,ANOVA
on
PCA
components
discrim
inantanalysis
PCA/stepwisediscrim
inantanalysis
differentiate
amongall6stim
ulustypes
within
thefirst500ms
Visualinspectionofprovided
figures:N1,P2,
andP3adifferentiate
the+
and–polesofthe
E,P,A
dim
ensions
Chapman(1979)
10
Verbalaffectiverating
Word
class
notindicated
Pre-ratingonsemantic
differentiala:20E+,20
E–,20P+,20P–,20A+,
20A–
Additionalparameter
controlunclear
17ms
Centrally
15repetitions
1(C
Pz)
Linked
ears
reference
510ms
PCA
ofERP,ANOVA
on
PCA
components
discrim
inantanalysis
PCA/stepwisediscrim
inantanalysis
differentiate
amongall6stim
ulustypes
within
thefirst500ms
Differentiationbetweensemanticscales(E,P,
A)within
190ms
Differentiationamongpolarities
ofword
contenttypes
(E+/�
,P+/�
,A+/�
)within
330ms
Chapmanet
al.
(1980)
10,4males
Word
naming
Verbalaffectiverating
Word
class
notindicated
Pre-ratingonsemantic
differential:20E+,20E–,
20P+,20P–,20A+,20
A–
Additionalparameter
controlunclear
17ms
Centrally
15repetitions
1(C
Pz)
Linked
ears
reference
510ms
PCA
ofERP,ANOVA
on
PCA
components
discrim
inantanalysis
PCA/stepwisediscrim
inantanalysis
differentiate
amongall6stim
ulustypes
within
thefirst500ms
Visualinspection:N1,P2,P3adifferentiate
the+
and–polesoftheE,P,A
dim
ensions
Vanderploeg
etal.
(1987)
10,allmales
Affectiverating
Word
class
notindicated
Wordsfrom
Begleiter
etal.
(1969,1979)andChapman
etal.,(1977,1978)
Additionalparameter
controlunclear
80ms
Centrally
Repeateduntil32artifact-
free
averages
per
category
obtained
6(Fz,
Pz,
F7,F8,T5,T6)
Linked
ears
reference
800ms
PCA,ANOVA
onPCA
components
n.s.trend:amplitude:
emotional(pleasant&
unplesant)4
neutral
OnP3a(230–420ms)
andLPC
(500–624)ms
Table
A1.(C
on
tin
ued
)
Stu
dy
Subje
cts
N,
sex
Task
(s)
Sti
muli
Presentation
Rec
ord
ing
and
analy
sis
ER
Pef
fect
s
170

Naumannet
al.(1992)
Pilotstudy:17,8males
Replicationstudy:106,50
males
Word
length
decision
Affectiverating
Adjectives
90wordsaffectivenorm
s
(Hager
etal.,1985)
30pleasant,30unpleasant,
30neutral
Controlled
for:length,
concreteness
125ms
Centrally
Single
presentation
3(Fz,
Cz,
Pz)
Leftmastoid
reference
1500mspost
word
ANOVAsonP3and
positiveslow
wave
amplitudeandpeak
latency
Pilotstudy:
P3amplitude(300–700ms)
Aff
ecti
vera
ting4
word
length
dec
isio
n.
Em
oti
onal
conte
nt
(ple
asa
nt
&
unple
asa
nt)
4neu
tral
Positiveslow
waveamplitude(700–1200):
Affectiverating4word
length
decision
Electrode�em
otionalcontent
Larg
erposi
tivi
ties
at
centr
al
and
pari
etal
elec
trodes
for
ple
asa
nt
and
unple
asa
nt
Replicationstudy:
P3amplitude
Emotionalcontent
Em
oti
onal
(ple
asa
nt
&unple
asa
nt)4
neu
tral
Electrode�task:
Fz:
aff
ecti
vera
ting4
word
length
dec
isio
n
Electrode�em
otionalcontent
Larg
eref
fect
for
ple
asa
nt,
part
icula
rly
at
Fz
and
Cz
P3peaklatency:
Neu
tralo
emoti
onal
(ple
asa
nt
&unple
asa
nt)
Positiveslow
waveamplitude
Electrode�em
otionalcontent
No
pari
etal
max
imum
for
neu
tral
Naumannet
al.
(1997)
54(26males,betweendesign,
18per
condition)
Letterdetection
Concretenessdecision
Valence
decision
Nouns
Published
affectivenorm
s
(Schwibbeet
al.,1981)
28unpleasant,28neutral
Controlled
for:length,
concrete-ness
200ms
Centrally
Single
presentation
9(F4,C4,P4,Fz,
Cz,
Pz,
F3,C3,P3)
-Linked
ears
reference
1400mspost
word
ANOVA
andTukey
HSD
test
onP3andpositive
slow
waveamplitudes
and
peaklatencies
Emotionalcontent�task
Unpleasant4
neutral
Effectfrom
valence
decisiontask
only
Valence
decisiontask
P3amplitude:
Unple
asa
nt4
neu
tral
P3peaklatency
Emotionalcontent�task
Letterdetection&
concretenessdecision:
unpleasanto
neutral
Valence
decision:
neutralo
unpleasant
Positiveslow
waveamplitude
Noeffects
171

Skrandies(1998)
22,8males
Visualize
andmem
orize
words
Nouns
Pre-ratingonsemantic
differentiala(different
population):10E+,10
E–,10P+,10P–,10A+,
10A–
Controlled
for:Word
length,word
frequency
1000ms
Centrally
RSVPb
40repetitions
30(10–20system
)
Averagereference
1000ms
ANOVAsoncentroidsof
scalp
distributionsand
peaklatencies
Duncan
post
hoc
Complexsequence
ofeffects,discrim
inating
amongE,P,A
dim
ensionsandtheirpolarity:
P1(80–130ms):
Peaklatency
A4EandP
Rightnegativecentroid
shiftforE-words
Leftpositivecentroid
shiftforE-words
N1(130–195ms):
Increasedpeaklatency
forEandA
words
Reducedglobalfieldpower
(GFP)forA
words
195–265ms:
Peaklatency
PoEandA
Anteriorcentroid
shiftforE-,P-,A-
comparedto
E+,P+,A+
565–635ms:
Peaklatency:
E+,P+,A+
4E-,P-,A-
635–765ms:
Centroids:P+
andP-shiftedanteriorally
andposteriorally
860–975ms:
Peaklatency:
E+
andP+4E-andP-A
+oA-
Skrandiesand
Chiu
(2003)
23,10males
Visualize
andmem
orize
words
Nouns
Pre-ratingonsemantic
differentiala(different
population):10E+,10
E–,10P+,10P–,10A+,
10A–
Controlled
forword
length,word
frequency
1000ms
Centrally
RSVPb
24(?)repetitions
32(10–20system
)
Averagereference
1000mspost
word
ANOVAsonpeaklatency
centroid
positionGlobal
fieldpower
(GFP)
Complexsequence
ofeffects,discrim
inating
betweenE,P,A
dim
ensionsandtheir
polarity:
P1(80–130ms):
Latency
E+,P+,A+
4E-,P-,A-
N1(130–195ms):
GFPP-andA-4
P+andA+
Posterior-anteriorlocationofcentroids:
A+anteriorP+anteriorE+andE-anterior
P-andA-
Left-rightlocationofcentroids:
Right-shiftofA
centroid
Noeffectsafter
300ms
Table
A1.(C
on
tin
ued
)
Stu
dy
Subje
cts
N,
sex
Task
(s)
Sti
muli
Presentation
Rec
ord
ing
and
analy
sis
ER
Pef
fect
s
172

Schapkin
etal.
(2000)
15,7males
Emotionalvs.neutral
classification
Nouns
Pre-ratingonvalence
(differentpopulation)
6pleasant,6unpleasant,6
neutral
Controlled
for:syllables,
word
length,word
frequency,concreteness,
imagery,initialletter
150ms
Leftorrightvisualfield
8repetitions(4
per
visual
field)
14(only
C3,C4,P3,P4,
O1,O2analyzed)
Linked
ears
reference
800mspost
word
ANOVAsonbase-to-peak
measuresofN1,P2,P3,
SPW
post
hoc
t-tests
P2amplitude(200–300ms)
pleasant4
unpleasant
P3amplitude(300–450ms)
pleasant4
unpleasant
Slow
positivewave(1000–1800):
Frontal
pleasant4
unpleasant
Parietal:
pleasant4
neutral
Bernatet
al.
(2001)
17,8males
Focusonfixationpoint
Adjectives
Pre-ratingonsemantic
differentialin
same
population.
10pleasant,10unpleasant,
12neutral
‘word
length,word
frequency,luminance
Subliminal(1
ms)
Supraliminal(40ms)
Centrally
12repetitions(6
subliminal,6supraliminal)
6(F3,F4,P3,P4,CPz,Oz)
Linked
ears
reference
1000mspost
word
ANOVAsonP1,N1,P2,
P3,LPC
meanamplitudes
pleasantvs.unpleasant
(analysisofneutralwords
isnotpresented)
Subliminal:
Amplitudes:
Emotion:
P2(100–210ms),P3(200–500ms),LPC
(500–900ms):
unple
asa
nt4
ple
asa
nt
Emotion�hem
isphere:
P1(40–120ms),N1(80–170ms),P3
(200–500ms):
Lef
t:unple
asa
nt4
ple
asa
nt
Right:unpleasant4
pleasant
Supraliminal:
Amplitudes:
Emotion:
P3(200–500ms)
andLPC
(500–900ms):
unple
asa
nt4
ple
asa
nt
Emotion�hem
isphere,
P1(20–80ms),N1(52–150ms),
Lef
t:unple
asa
nt4
ple
asa
nt
Right:unpleasant4
pleasant
P3(200–500ms)
EmotionalContent�hem
isphere:
unpleasant:left4right
Ortigueet
al.
(2004)
13,allmales
Divided
fieldlexicaldecision
(inwhichvisualfielddid
a
word
appear?)
Nouns
Preratingforem
otionality
onseven-pointscale
(emotional–nonem
otional)
16words,(8
neutral8
emotional)96
pseudowords
Controlled
for:word
length,word
frequency
13ms
Leftandrightvisualfield
simultaneously
30repetitions
123(extended
10–20
system
)
Averagereference
250mspost
word
ANOVAsonspatial
configurationmaps
LAURA
sourceestimation
Amplitude:
100–140ms:
emoti
onal4
neu
tral
Distinct
rightoccipitalspatialmapfor
emotionalwordspresentedin
therightvisual
field
173
Herbertet
al.
(2006)
26,16males
Covertevaluation
Adjectives
Pre-ratedonvalence
and
arousalbydifferent
population
60unpleasant,60pleasant,
60neutral
Controlled
for:word
length,word
frequency
4000ms
Centrally
Single
presentation
64(extended
10–20system
)
Averagereference
1000mspost
word
ANOVAsonchannel
groups
Amplitude:
P2(180–250ms)
andP3a(250–400ms):
emoti
onal
(ple
asa
nt
&unple
asa
nt)4
neu
tral
LPC
(600–750ms):
ple
asa
nt4
neu
tral
Kissler
etal.
(submitted)
16,8males
Silentreading
Nouns
Pre-ratingofvalence
and
arousalbydifferent
population
60unpleasant,60pleasant,
60neutral
Word
length,word
frequency,concreteness
333msand1000ms
RSVPb
10(5
per
presentation
speed)
64(extended
10–20system
)
Averagereference
333ms/1000mspost
word
ANOVA
ongroupsof
electrodes
Amplitude:
200–300msoccipito-tem
poral
electrodes
Em
oti
onal
(ple
asa
nt¼
unple
asa
nt)4
neu
tral
Left:em
otional4
neutral
Right:em
otional4
neutral
aDim
ensionsofthesemanticdifferential:E,evaluation;P,potency;A,activity.+
/-indicate
thepositiveandnegativepolesofthesedim
ensions.
bRSVP,rapid
serialvisualpresentationofstim
uliin
acontinuousconsecutivestream
withoutinterstimulusinterval.
Table
A1.(C
on
tin
ued
)
Stu
dy
Subje
cts
N,
sex
Task
(s)
Sti
muli
Presentation
Rec
ord
ing
and
analy
sis
ER
Pef
fect
s
174

Table
A2.A
summary
ofexperim
entaldesign,recordingparametersandresultsofthereviewed
studiesoneffectsofem
otionalcontentonERP
measuresofword
processingin
clinicalpopulations.
Stu
dy
Su
bje
cts
N,
sex
Ta
sk(
s)S
tim
uli
Pre
sen
tati
on
Rec
ord
ing
an
d
an
aly
sis
ER
Pef
fect
s
clinicalstatus
Electrodes
and
reference
analysis
window
(post-
stim
ulus)
ER
Pef
fect
s
Word
class
Emotional
content
Controlof
additional
stim
ulus
parameters
Stimulus
duration
Positionof
presentation
Stimulus
repetitions
Statistics
Williamsonet
al.
(1991)
16—
8psychopaths,
8non-psychopaths,
allmales
Lexicaldecision
Word
class
not
indicated
Affectivenorm
s
(Togliaand
Battig,1978)
13pleasant,13
unpleasant,13
neutral,39
pseudowords
Word
length,
frequency,
number
of
syllables,
concreteness
176ms
+/-3para-
foveally,
vertical
form
at
6repetitions
5(Fz,
Cz,
Pz,
PT3,
PT4)
Linked
ears
reference
2000ms
ANOVA
t-test
P240:225–300ms
Amplitude:
No
np
sych
op
ath
s:
emo
tio
na
l(
ple
asa
nt
&
un
ple
asa
nt)4
neu
tra
l.
Psy
chop
ath
s:em
oti
on
al
(p
lea
sant
&
un
ple
asa
nt)¼
neu
tra
l
LPC:650–800ms
Amplitude:
Group�em
otional
Content�electrode
site
Mid
lin
e:
No
n-p
sych
op
ath
s:
emo
tio
na
l(
ple
asa
nt
&
un
ple
asa
nt)4
neu
tra
l
Psy
chop
ath
s:em
oti
on
al
(p
lea
sant
&
un
ple
asa
nt)4
neu
tra
l
175

Weinstein(1995)
20students
10
highly
anxious,5
males10low
anxious4males
Decisionon
contextualfit
20threatening,
20pleasant
sentences(5–7
words)
‘Probewords’:20
threat,20neutral
20pleasant
(nouns&
adjectives)2
pairings:fitting-
notfitting.
Threatwords
from
MacL
eod
(1985).
Emotional
contentof
sentencesand
wordsprerated
indifferent
population.
Controlfor
linguistic
parametersnot
indicated
1100msper
word
Centrally
2repetitions
3(Fz,
Cz,
Pz)
Linked
ears
reference
650ms
ANOVA
t-test
N100:90–120ms:Peak
amplitude:
High
anxiety
groupthreat
priming4
low
anxiety
groupthreatpriming.
(Fz,
Cz)
N400/P400:400–500
msPeakamplitude:
Threatcondition:High
anxiety
groupmore
positivethanlow
anxiety
group(C
z,
Pz).Peaklatency:High
anxiety
groupolow
anxiety
group.
(Plots
ofmeansappear
todivergefrom
the
reported
statistics)
Table
A2.(C
on
tin
ued
)
Stu
dy
Sub
ject
sN
,se
xT
ask
(s)
Sti
mu
liPresentation
Rec
ord
ing
an
d
an
aly
sis
ER
Pef
fect
s176

Kiehlet
al.(1999),
Task
3
29prisoners,all
males:8psycho-
paths,9
nonpsychopaths,12
mixed
Pleasant/unpleasant
decision
Word
class
not
indicated
Valence
norm
s
(Togliaand
Battig,1978)
60pleasant,60
unpleasant
Controlled
for:
word
length,
frequency,
number
of
syllabels,
imagery,
concreteness
300ms
Centrally
Single
presentation
5(Fz,
Cz,
Pz,
PT3,
PT4)Linked
ears
reference
1200ms
ANOVA
t-test
on
reactiontimes
and
N350,LPC
amplitudes
N350:300–400ms
Amplitude:
Allgroups:
Emotionalcontent:
ple
asa
nt4
un
ple
asa
nt
Emotional
content�hem
isphere:
Lef
t:
ple
asa
nt4
un
ple
asa
nt
Rig
ht:
ple
asa
nt4
un
ple
asa
nt
LPC:400–800ms
Amplitude
Emotionalcontent:
No
n-p
sych
op
ath
s:
Un
ple
asa
nt4
ple
asa
nt
Psy
chop
ath
s:
Un
ple
asa
nt¼
ple
asa
nt
Emotional
content�hem
isphere:
Lef
t:
un
ple
asa
nt4
ple
asa
nt
Rig
ht:
un
ple
asa
nt4
ple
asa
nt
Reactiontimes:
Pleasanto
unpleasant
Knost
etal.(1997)
38:19
prechronicpain
patients,11
males
19controls,11
males
Detectionand
naming
Adjectives
Pre-ratingon
familiarity,body
andpain
relatedness
40pain-related,
40body-related,
40neutral
Controlled
for:
word
frequency,
word
length
Individual
perceptual
threshold
Centrally
Single
presentation
11(Fz,
F3,F4,Cz,
C3,C4,Pz,
P3,P4,
T3,T4)
Linked
ears
reference
800ms
ANOVA
t-test
N100(80–180ms)
at
electrodeF3
Prechronic
pain
patients:
Am
pli
tude:
pain
wo
rds4
bo
dy
rela
ted
&
neu
tra
l
LPC2:600–800ms
Am
pli
tude
both
gro
ups:
pa
in/b
od
y-
rela
ted4
neu
tra
l
177

Floret
al.(1997)
24:12chronic
pain
patients,5males12
controls,5males
Detectionand
naming
Adjectives
Pre-ratingon
familiarity,body
andpain
relatedness
40pain-related,
40body-related,
40neutral
Controlled
for:
word
frequency,
word
length
Perceptual
threshold
Centrally
Single
presentation
11(Fz,
F3,F4,Cz,
C3,C4,Pz,
P3,P4,
T3,T4)
Linked
ears
reference
800ms
ANOVA
t-test
N100(80–140ms)
at
electrodes
F3,Fz,
C3,
P3,Pz
Chronic
pain
patients
only:
pa
inw
ord
s4b
od
y
rela
ted
&n
eutr
al
Both
groups:
N100(80–140ms)
Pain
words:left4
right
Neutral:right4
left
N200(140–200ms)
at
allelectrodes:
Ch
ronic
pa
in
pa
tien
ts:p
ain
wo
rds4
bo
dy
rela
ted
&
neu
tra
l
P200(180–280ms):
Chronic
pain
patients:
Larger
responsesto
pain
wordsover
right
thanleft
LPC1(400–600ms):
n.s.All:pain
words¼
body
related¼
neutral
LPC2(600–800ms)
n.s.All:pain
words¼
body
related¼
neutral.
Table
A2.(C
on
tin
ued
)
Stu
dy
Sub
ject
sN
,se
xT
ask
(s)
Sti
mu
liPresentation
Rec
ord
ing
an
d
an
aly
sis
ER
Pef
fect
s178

Pauliet
al.(2005)
50:25panic
patients,
9males,25controls,
9males
Detectionand
naming
Adjectives
and
verbs
Pre-ratingfor
descriptiveness
forpanic-
disorder
symptomsby2
psychiatrists
and
2clinical
psychologists
48panic
related,
48neutral
Controlled
for:
word
frequency,
length,syllables,
distributionof
word
classes
Perceptual
threshold
and
1000ms,
2presentations
(oneper
duration)
21(only
9analyzed:
Fz,
Cz,
Pz,
F3,F4,
C3,C4,P3,P4)
Linked
ears
pnce
1000ms
ANOVA
t-test
Threshold
presentation:
Panic
patients:
N200/P200
(100–200ms)
Frontaleffect
pa
nic
wo
rds4
neu
tra
l
P3a:(200–400ms):
pa
nic
word
s4n
eutr
al
LPC1:400–600ms
Amplitudeboth
groups:
pa
nic4
neu
tra
l
1000mspresentation:
LPC1:400–600ms
Amplitudeboth
groups:
pa
nic4
neu
tra
l
LPC2:600–800
Amplitudeboth
groups:
pa
nic4
neu
tra
l
Emotional
content�hem
isphere:
Lef
t:p
an
ic4
neu
tra
l
Rig
ht:
pa
nic4
neu
tra
l
179

Acknowledgments
This work was supported by a grant from theHeidelberg Academy of Sciences (Mind and BrainProgram). We thank Anne Hauswald for help inpreparation of this manuscript and ChristianeBeck, Susanne Kossler, Bistra Ivanona and IreneWinkler for assistance in the experimental workdescribed.
References
Amaral, D.G., Behniea, H. and Kelly, J.L. (2003) Topographic
organization of projections from the amygdala to the visual
cortex in the macaque monkey. Neuroscience, 118:
1099–1120.
Anderson, A.K. and Phelps, E.A. (2001) Lesions of the human
amygdala impair enhanced perception of emotionally salient
events. Nature, 411: 305–309.
Assadollahi, R. and Pulvermuller, F. (2001) Neuromagnetic
evidence for early access to cognitive representations. Ne-
uroreport, 12: 207–213.
Assadollahi, R. and Pulvermuller, F. (2003) Early influences of
word length and frequency: a group study using MEG. Ne-
uroreport, 14: 1183–1187.
Assadollahi, R. and Rockstroh, B. (2005) Neuromagnetic brain
responses to words from semantic sub- and supercategories.
BMC Neurosci., 6: 57.
Barsalou, L.W. (1999) Perceptual symbol systems. Behav. Brain
Sci., 22: 577–609 discussion 610–660.
Begleiter, H. and Platz, A. (1969) Cortical evoked potentials to
semantic stimuli. Psychophysiology, 6: 91–100.
Begleiter, H., Projesz, B. and Garozzo, R. (1979) Visual evoked
potentials and affective ratings of semantic stimuli. In: Beg-
leiter, H. (Ed.), Evoked Brain Potentials and Behavior. Ple-
num Press, New York, pp. 127–143.
Bentin, S., Mouchetant-Rostaing, Y., Giard, M.H., Echallier,
J.F. and Pernier, J. (1999) ERP manifestations of processing
printed words at different psycholinguistic levels: time course
and scalp distribution. J. Cogn. Neurosci., 11: 235–260.
Bernat, E., Bunce, S. and Shevrin, H. (2001) Event-related
brain potentials differentiate positive and negative mood ad-
jectives during both supraliminal and subliminal visual
processing. Int. J. Psychophysiol., 42: 11–34.
Bower, G. (1981) Mood and memory. Am. Psychol., 36:
129–148.
Bradley, M.M. and Lang, P.J. (1994) Measuring emotion: the
Self-Assessment Manikin and the Semantic Differential. J.
Behav. Ther. Exp. Psychiatry, 25(1): 49–59.
Bradley, M., and Lang, P.J., 1998. Affective norms for English
words (ANEW): Instruction manual and affective ratings.
Technical report A-8, The Center for Research in Psycho-
physiology, University of Florida.
Bradley, M.M. and Lang, P.J. (2000) Affective reactions to
acoustic stimuli. Psychophysiology, 37: 204–215.
Brem, S., Lang-Dullenkopf, A., Maurer, U., Halder, P., Buc-
her, K. and Brandeis, D. (2005) Neurophysiological signs of
rapidly emerging visual expertise for symbol strings. Neuro-
report, 16: 45–48.
Caccioppo, J.T. (2000) Asymmetries in affect laden information
processing. In: Banaji, R. and Prentice, D.A. (Eds.), Per-
spectivism in Social Psychology: The Yin and Yang of Sci-
entific Progress. American Psychological Association Press,
Washington, DC, pp. 85–95.
Chao, L.L., Haxby, J.V. and Martin, A. (1999) Attribute-based
neural substrates in temporal cortex for perceiving and
knowing about objects. Nat. Neurosci., 2: 913–919.
Chapman, R.M. (1979) Connotative meaning and averaged
evoked potentials. In: Begleiter, H. (Ed.), Evoked Brain Po-
tentials and Behavior. Plenum Press, New York, pp. 171–197.
Chapman, R.M., McCrary, J.W., Chapman, J.A. and Bragdon,
H.R. (1978) Brain responses related to semantic meaning.
Brain Lang., 5: 195–205.
Chapman, R.M., McCrary, J.W., Chapman, J.A. and Martin,
J.K. (1980) Behavioral and neural analyses of connotative
meaning: word classes and rating scales. Brain Lang., 11:
319–339.
Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-
Lambertz, G., Henaff, M.A. and Michel, F. (2000) The visual
word form area: spatial and temporal characterization of an
initial stage of reading in normal subjects and posterior split-
brain patients. Brain, 123(Pt 2): 291–307.
Davis, M. (1992) The role of the amygdala in fear and anxiety.
Annu. Rev. Neurosci., 15: 353–375.
Dehaene, S. (1995) Electrophysiological evidence for category-
specific word processing in the normal human brain. Neuro-
report, 6: 2153–2157.
Dehaene, S., Cohen, L., Sigman, M. and Vinckier, F. (2005)
The neural code for written words: a proposal. Trends Cogn.
Sci., 9: 335–341.
Dehaene, S., Le Clec, H.G., Poline, J.B., Le Bihan, D. and
Cohen, L. (2002) The visual word form area: a prelexical
representation of visual words in the fusiform gyrus. Neuro-
report, 13: 321–325.
Dien, J., Spencer, K.M. and Donchin, E. (2004) Parsing the late
positive complex: mental chronometry and the ERP compo-
nents that inhabit the neighborhood of the P300. Psycho-
physiology, 41: 665–678.
Dijksterhuis, A. and Aarts, H. (2003) On wildebeests and hu-
mans: the preferential detection of negative stimuli. Psychol.
Sci., 14: 14–18.
Everatt, J., McCorquidale, B., Smith, J., Culverwell, F., Wilks,
A., Evans, D., Kay, M. and Baker, D. (1999) Association
between reading ability and visual processes. In: Everatt, J.
(Ed.), Reading and Dyslexia. Routledge, London, pp. 1–39.
Federmeier, K.D., Kirson, D.A., Moreno, E.M. and Kutas, M.
(2001) Effects of transient, mild mood states on semantic
memory organization and use: an event-related potential in-
vestigation in humans. Neurosci. Lett., 305: 149–152.
180

Federmeier, K.D., Segal, J.B., Lombrozo, T. and Kutas, M.
(2000) Brain responses to nouns, verbs and class-ambiguous
words in context. Brain, 123(Pt 12): 2552–2566.
Flor, H., Knost, B. and Birbaumer, N. (1997) Processing of
pain- and body-related verbal material in chronic pain pa-
tients: central and peripheral correlates. Pain, 73: 413–421.
Friederici, A.D., Hahne, A. and Mecklinger, A. (1996) Tem-
poral structure of syntactic parsing: early and late event-re-
lated brain potential effects. J. Exp. Psychol. Learn. Mem.
Cogn., 22: 1219–1248.
Garavan, H., Pendergrass, J.C., Ross, T.J., Stein, E.A. and
Risinger, R.C. (2001) Amygdala response to both pos-
itively and negatively valenced stimuli. Neuroreport, 12:
2779–2783.
Greenwald, A.G., Draine, S.C. and Abrams, R.L. (1996) Three
cognitive markers of unconscious semantic activation. Sci-
ence, 273: 1699–1702.
Hagoort, P. and Brown, C.M. (2000) ERP effects of listening to
speech compared to reading: the P600/SPS to syntactic vi-
olations in spoken sentences and rapid serial visual presen-
tation. Neuropsychologia, 38: 1531–1549.
Halgren, E., Baudena, P., Heit, G., Clarke, J.M., Marinkovic,
K., Chauvel, P. and Clarke, M. (1994a) Spatio-temporal
stages in face and word processing. 2. Depth-recorded po-
tentials in the human frontal and Rolandic cortices. J.
Physiol. (Paris), 88: 51–80.
Halgren, E., Baudena, P., Heit, G., Clarke, J.M., Marinkovic,
K. and Clarke, M. (1994b) Spatio-temporal stages in face and
word processing. I. Depth-recorded potentials in the human
occipital, temporal and parietal lobes [corrected]. J. Physiol.
(Paris), 88: 1–50.
Hamann, S. and Mao, H. (2002) Positive and negative emo-
tional verbal stimuli elicit activity in the left amygdala. Ne-
uroreport, 13: 15–19.
Hauk, O., Johnsrude, I. and Pulvermuller, F. (2004) Somato-
topic representation of action words in human motor and
premotor cortex. Neuron, 41: 301–307.
Haxby, J.V., Horwitz, B., Ungerleider, L.G., Maisog, J.M.,
Pietrini, P. and Grady, C.L. (1994) The functional organi-
zation of human extrastriate cortex: a PET-rCBF study of
selective attention to faces and locations. J. Neurosci., 14:
6336–6353.
Herbert, C., Kissler, J., Junghofer, M., Peyk, P. and Rockstroh,
B. (2006) Processing emotional adjectives: evidence from
startle EMG and ERPs. Psychophysiology, 43(2): 197–206.
Hinojosa, J.A., Martin-Loeches, M., Munoz, F., Casado, P.
and Pozo, M.A. (2004) Electrophysiological evidence of au-
tomatic early semantic processing. Brain Lang., 88: 39–46.
Hinojosa, J.A., Martin-Loeches, M. and Rubia, F.J. (2001)
Event-related potentials and semantics: an overview and an
integrative proposal. Brain Lang., 78: 128–139.
Holcomb, P.J., Kounios, J., Anderson, J.E. and West, W.C.
(1999) Dual-coding, context-availability, and concreteness
effects in sentence comprehension: an electrophysiological
investigation. J. Exp. Psychol. Learn. Mem. Cogn., 25:
721–742.
Isenberg, N., Silbersweig, D., Engelien, A., Emmerich, S., Ma-
lavade, K., Beattie, B., Leon, A.C. and Stern, E. (1999) Lin-
guistic threat activates the human amygdala. Proc. Natl.
Acad. Sci. USA, 96: 10456–10459.
Ito, T.A. and Caccioppo, J.T. (2000) Electrophysiological ev-
idence of implicit and explicit categorization processes. J.
Exp. Soc. Psychol., 36: 660–676.
Junghofer, M., Bradley, M.M., Elbert, T.R. and Lang, P.J.
(2001) Fleeting images: a new look at early emotion discrim-
ination. Psychophysiology, 38: 175–178.
Keil, A., Bradley, M.M., Hauk, O., Rockstroh, B., Elbert, T.
and Lang, P.J. (2002) Large-scale neural correlates of affec-
tive picture processing. Psychophysiology, 39: 641–649.
Kellenbach, M.L., Wijers, A.A., Hovius, M., Mulder, J. and
Mulder, G. (2002) Neural differentiation of lexico-syntactic
categories or semantic features? Event-related potential ev-
idence for both. J. Cogn. Neurosci., 14: 561–577.
Kiefer, M. and Spitzer, M. (2000) Time course of conscious and
unconscious semantic brain activation. Neuroreport, 11:
2401–2407.
Kiehl, K.A., Hare, R.D., McDonald, J.J. and Brink, J. (1999)
Semantic and affective processing in psychopaths: an event-
related potential (ERP) study. Psychophysiology, 36:
765–774.
King, J.W. and Kutas, M. (1998) Neural plasticity in the dy-
namics of human visual word recognition. Neurosci. Lett.,
244: 61–64.
Kissler, J. and Bauml, K.H. (2000) Effects of the beholder’s age
on the perception of facial attractiveness. Acta Psychol.
(Amst.), 104: 145–166.
Kissler, J., Herbert, C., Peyk, P. and Junghofer, M. (submitted
manuscript) Sex, crime and videotape—enhanced early cor-
tical responses to rapidly presented emotional words.
Kissler, J., and Kossler, S., (in preparation) Pleasant pictures
facilitate lexical decision.
Knost, B., Flor, H., Braun, C. and Birbaumer, N. (1997) Cer-
ebral processing of words and the development of chronic
pain. Psychophysiology, 34: 474–481.
Kostandov, E. and Arzumanov, Y. (1977) Averaged cortical
evoked potentials to recognized and non-recognized verbal
stimuli. Acta Neurobiol. Exp. (Wars), 37: 311–324.
Kutas, M. and Federmeier, K.D. (2000) Electrophysiology re-
veals semantic memory use in language comprehension.
Trends Cogn. Sci., 4: 463–470.
Kutas, M. and Hillyard, S.A. (1980) Reading senseless sen-
tences: brain potentials reflect semantic incongruity. Science,
207: 203–205.
Kutas, M. and Hillyard, S.A. (1984) Brain potentials during
reading reflect word expectancy and semantic association.
Nature, 307: 161–163.
Lang, P.J. (1979) Presidential address, 1978A bio-infor-
mational theory of emotional imagery. Psychophysiology,
16: 495–512.
Lang, P.J. (1994) The motivational organization of emotion:
Affect-reflex connections. In: Van Goozen, S.H.M., Van de
Poll, N.E. and Sergeant, J.E. (Eds.), Emotions: Essays on
181

Emotion Theory. Lawrence Erlbaum Associates, Hillsdale,
NJ, pp. 61–93.
Lang, P.J., Greenwald, M.K., Bradley, M.M. and Hamm, A.O.
(1993) Looking at pictures: affective, facial, visceral, and be-
havioral reactions. Psychophysiology, 30: 261–273.
LeDoux, J.E. (1989) Cognitive-emotional interactions in the
brain. Cogn. Emotion, 3: 267–289.
LeDoux, J.E. (1995) Emotion: clues from the brain. Annu. Rev.
Psychol., 46: 209–235.
Liddell, B.J., Brown, K.J., Kemp, A.H., Barton, M.J., Das, P.,
Peduto, A., Gordon, E. and Williams, L.M. (2005) A direct
brainstem-amygdala-cortical ‘alarm’ system for subliminal
signals of fear. Neuroimage, 24: 235–243.
Liddell, B.J., Williams, L.M., Rathjen, J., Shevrin, H. and
Gordon, E. (2004) A temporal dissociation of subliminal
versus supraliminal fear perception: an event-related poten-
tial study. J. Cogn. Neurosci., 16: 479–486.
Lifshitz, K. (1966) The averaged evoked cortical response to
complex visual stimuli. Psychophysiology, 3: 55–68.
Luce, R.D. and Suppes, P. (1965) Preference, utility and sub-
jective probability. In: Luce, R.D., Bush, R.R. and Galanter,
E.H. (Eds.) Handbook of Mathematical Psychology, Vol. 3.
New York, Wiley, pp. 249–410.
Martin, A., Wiggs, C.L., Ungerleider, L.G. and Haxby, J.V.
(1996) Neural correlates of category-specific knowledge. Na-
ture, 379: 649–652.
Martin-Loeches, M., Hinojosa, J.A., Gomez-Jarabo, G. and
Rubia, F.J. (2001) An early electrophysiological sign of se-
mantic processing in basal extrastriate areas. Psychophysiol-
ogy, 38: 114–124.
Molfese, D. (1985) Electrophysiological correlates of semantic
features. J. Psycholinguistic Res., 14: 289–299.
Morris, J.S., Ohman, A. and Dolan, R.J. (1999) A subcortical
pathway to the right amygdala mediating ‘unseen’ fear. Proc.
Natl. Acad. Sci. USA,, 96: 1680–1685.
Munte, T.F., Heinze, H.J., Matzke, M., Wieringa, B.M. and
Johannes, S. (1998) Brain potentials and syntactic violations
revisited: no evidence for specificity of the syntactic positive
shift. Neuropsychologia, 36: 217–226.
Naccache, L., Gaillard, R., Adam, C., Hasboun, D., Clemenc-
eau, S., Baulac, M., Dehaene, S. and Cohen, L. (2005) A
direct intracranial record of emotions evoked by subliminal
words. Proc. Natl. Acad. Sci. USA, 102: 7713–7717.
Ohman, A. and Mineka, S. (2001) Fears, phobias, and prepar-
edness: toward an evolved module of fear and fear learning.
Psychol. Rev., 108: 483–522.
Ortigue, S., Michel, C.M., Murray, M.M., Mohr, C., Carbon-
nel, S. and Landis, T. (2004) Electrical neuroimaging reveals
early generator modulation to emotional words. Neuroim-
age, 21: 1242–1251.
Osgood, C.E., Miron, M.S. and May, W.H. (1975) Cross-Cul-
tural Universals of Affective Meaning. University of Illinois
Press, Urbana, Chicago, London.
Osgood, C.E., Suci, G.J. and Tannenbaum, P.H. (1957) The
measurement of meaning. University of Illinois Press, Ur-
bana, Chicago, and London.
Osterhout, L., Holcomb, P.J. and Swinney, D.A. (1994) Brain
potentials elicited by garden-path sentences: evidence of the
application of verb information during parsing. J. Exp. Psy-
chol. Learn. Mem. Cogn., 20: 786–803.
Pauli, P., Amrhein, C., Muhlberger, A., Dengler, W. and
Wiedemann, G. (2005) Electrocortical evidence for an early
abnormal processing of panic-related words in panic disorder
patients. Int. J. Psychophysiol., 57: 33–41.
Perfetti, C.A. (1998) Comprehending written language: a blue-
print of the reader. In: Brown, C.M. and Hagoort, P. (Eds.),
The Neurocognition of Language. University Press, Oxford.
Perfetti, C.A. and Sandak, R. (2000) Reading optimally builds
on spoken language: implications for deaf readers. J. Deaf.
Stud. Deaf. Educ., 5: 32–50.
Phelps, E.A., O’Connor, K.J., Gatenby, J.C., Gore, J.C., Grill-
on, C. and Davis, M. (2001) Activation of the left amygdala
to a cognitive representation of fear. Nat. Neurosci., 4:
437–441.
Posner, M.I., Abdullaev, Y.G., McCandliss, B.D. and Sereno,
S.C. (1999) Neuroanatomy, circuitry and plasticity of word
reading. Neuroreport, 10: R12–R23.
Pulvermuller, F. (1999) Words in the brain’s language. Behav.
Brain Sci., 22: 253–279 discussion 280–336.
Pulvermuller, F., Assadollahi, R. and Elbert, T. (2001a) Ne-
uromagnetic evidence for early semantic access in word rec-
ognition. Eur. J. Neurosci., 13: 201–205.
Pulvermuller, F., Harle, M. and Hummel, F. (2000) Neuro-
physiological distinction of verb categories. Neuroreport, 11:
2789–2793.
Pulvermuller, F., Harle, M. and Hummel, F. (2001b) Walking
or talking? Behavioral and neurophysiological correlates of
action verb processing. Brain Lang., 78: 143–168.
Rudell, A.P. (1992) Rapid stream stimulation and the recog-
nition potential. Electroencephalogr. Clin. Neurophysiol., 83:
77–82.
Russel, J. (1980) A circumplex model of affects. J. Pers. Soc.
Psychol., 39: 1161–1178.
Schapkin, S.A., Gusev, A.N. and Kuhl, J. (2000) Categoriza-
tion of unilaterally presented emotional words: an ERP anal-
ysis. Acta. Neurobiol. Exp. (Wars), 60: 17–28.
Schendan, H.E., Ganis, G. and Kutas, M. (1998) Neurophys-
iological evidence for visual perceptual categorization of
words and faces within 150 ms. Psychophysiology, 35:
240–251.
Schupp, H.T., Ohman, A., Junghofer, M., Weike, A.I., Stock-
burger, J. and Hamm, A.O. (2004) The facilitated processing
of threatening faces: an ERP analysis. Emotion, 4: 189–200.
Sereno, S.C., Brewer, C.C. and O’Donnell, P.J. (2003) Context
effects in word recognition: evidence for early interactive
processing. Psychol. Sci., 14: 328–333.
Sereno, S.C., Rayner, K. and Posner, M.I. (1998) Estab-
lishing a time-line of word recognition: evidence from eye
movements and event-related potentials. Neuroreport, 9:
2195–2200.
Silvert, L., Delplanque, S., Bouwalerh, H., Verpoort, C. and
Sequeira, H. (2004) Autonomic responding to aversive words
182

without conscious valence discrimination. Int. J. Psycho-
physiol., 53: 135–145.
Skrandies, W. (1998) Evoked potential correlates of semantic
meaning — a brain mapping study. Brain Res. Cogn. Brain
Res., 6: 173–183.
Skrandies, W. and Chiu, M.J. (2003) Dimensions of affective
semantic meaning — behavioral and evoked potential
correlates in Chinese subjects. Neurosci. Lett., 341:
45–48.
Sutton, S., Tueting, P., Zubin, J. and John, E.R. (1967) Infor-
mation delivery and the sensory evoked potential. Science,
155: 1436–1439.
Tarkiainen, A., Helenius, P., Hansen, P.C., Cornelissen, P.L.
and Salmelin, R. (1999) Dynamics of letter string perception
in the human occipitotemporal cortex. Brain, 122(Pt 11):
2119–2132.
Vanderploeg, R.D., Brown, W.S. and Marsh, J.T. (1987) Judg-
ments of emotion in words and faces: ERP correlates. Int. J.
Psychophysiol., 5: 193–205.
Warrington, E.K. and Shallice, T. (1979) Semantic access dys-
lexia. Brain, 102: 43–63.
Warrington, E.K. and Shallice, T. (1980) Word-form dyslexia.
Brain, 103: 99–112.
Weinstein, A. (1995) Visual ERPs evidence for enhanced
processing of threatening information in anxious university
students. Biol. Psychiatry, 37: 847–858.
Wickelmaier, F. and Schmid, C. (2004) A Matlab function to
estimate choice model parameters from paired-comparison
data. Behav. Res. Methods Instrum. Comput., 36: 29–40.
Williamson, S., Harpur, T.J. and Hare, R.D. (1991) Abnormal
processing of affective words by psychopaths. Psychophys-
iology, 28: 260–273.
183

















