Page 1
What genomics of arbuscular mycorrhizal symbiosis teach us about root development
Formey Damien, Jourda Cyril, Christophe Roux and Pierre-Marc Delaux
Université de Toulouse ; UPS ; UMR 5546, Laboratoire de Recherche en Sciences Végétales;
BP 42617, F-31326, Castanet-Tolosan, France
CNRS ; UMR 5546 ; BP 42617, F-31326, Castanet-Tolosan, France
Soil is a biotic environment where plant roots are faced to myriad of microbes. It has
been estimated that one gram of soil contains up to 1010-1011 bacteria (Horner-Devine et al.,
2003) and up to 200 mg of fungal hyphae (Leake et al., 2004). This biotic environment has a
great incidence on plant diversity and productivity (van der Heijden et al., 2007) as several of
these microbes are pathogenic or symbiotic and directly interact with plants, like arbuscular
mycorhizal (AM) fungi.
AM fungi are a particular monophyletic fungal group called Glomeromycota
(Schüβler et al., 2001, Schwarzott et al., 2001). This group has been described as basal and/or
sister of the Dikarya (Helgason et al., 2003, James et al., 2006, Liu et al., 2009, Liu et al.,
2006, Redecker and Raab 2006). These fungi are ancient and fossil records showed that they
were already in association with the first land plants (Remy et al., 1994, Redecker et al.,
2000). AM fungi are currently present in different ecosystems where they are of great
incidence in the organisation of plant communities (Klironomos et al., 2011).
AM symbiosis is considered as the most widespread interaction on earth between plant
and microbes. This interaction has indeed been described for the great majority of plant
species and ca 80% of land plants are called mycotrophic as they are associated with AM
fungi (Smith & Reads, 1997). The mycorrhizal status of plants is the standard condition in the
phylum of green plants, although some species are not able to interact with AM fungi.
Intriguingly, this inability to form this symbiosis is shared by all the species of a considered
family, like Caryophyllaceae or Brassicaceae. In Brassicaceae for instance, Arabidopsis
thaliana is no host. Although some species have been described as able to be infected by AM
fungi, any symbiotic structures like arbuscules were observed (see DeMars & Boerner, 1995).
Except these non mycotrophic plant families, the ca universal host spectrum and the
Page 2
ancestrally of the AM symbiosis led several authors to hypothesize that AM fungi could have
co-evolved with the green lineage since the first land plants, participating to the apparition of
root tissues (Brundet, 2002).
The establishment of the AM symbiosis initiate from germination of spores in soil or,
more frequently in non-perturbed field, from mycelium issuing from an AM symbiotic root.
At the vicinity of a host root, both partners exchange symbiotic signals. The plant releases a
mix of molecules that stimulate hyphal branching and growth. A class of molecules
participating to this stimulation are strigolactones (Akiyama et al., 2005; Besserer et al.,
2006). On the other side, the fungus produces diffusible compounds that activate the
symbiotic program into the host plant (Kosuta et al., 2003; Olah et al., 2005). A family of
lipo-chito-oligosaccharides, which mimics part of the diffusible compound effects on the host
plant, has been recently identified and called Myc-LCOs (Maillet et al., 2011). Following this
pre-symbiotic phase, contact between both partners occurs: the AM fungus forms a
hyphopodia on the root epidermis. In response, the host cell develops a Pre-Penetration
Apparatus within which AM fungus colonizes the root (Genre et al., 2005). Intraradical
hyphae then grow inter- or intra-cellularly and symbiotic exchanges take place at in cortical
root cells of the host plant by hosting highly branched fungal structures called arbuscules.
Exchanges occurring in arbuscules are bidirectional. As obligate biotroph, the fungus is host-
dependent to obtain its carbon; in return the AM fungus provides nutrients as phosphate and
water to the host plant (Smith & Read, 2009). These exchanges are intense and plant
dependence to AM fungi can be important as it was demonstrated that in some case all the
plant phosphate can be provided by the fungal partner (Pearson & Jacobson, 1993). The
symbiotic phosphate uptakes require specific phosphate transporters localized in the peri-
arbuscular membrane of the host cell (Rausch et al. 2001; Harrison et al. 2002). Highlighting
the significance of these exchanges, it has been proposed that the mutualistic traits of the
symbiosis have been widely retained during the evolution of the green lineage as its
development is tightly tuned by both partners (Kiers et al., 2011). The fact that this trophic
symbiosis promotes plant growth gives to AM fungi a strong agronomic interest in
sustainable agriculture. Different works were developed to enhance infectivity in commercial
mycorrhizal inoculants (Corkidi, 2004). However, the biology of AM fungi is still an enigma.
The difficulty of cultivation of these obligate fungi, the inability to transform these
polykaryotic organisms, the lack of knowledge on their genetics or sexuality makes its study
difficult. Recent works showing transient genetic transformation (Helber and Requena 2008)
Page 3
and fungal host induced RNAi (Helber et al., 2011) are significant technical advances and
allow a glimpse of the possibility for functional analyses on these fungi. Although different
species of Glomeromycetes are used in laboratories (Glomus mosseae, Gigaspora rosea,
Gigaspora gigantea, Gigaspora margarita), the species Glomus irregulare was chosen as a
model due to its aggressiveness and its easy multiplication in root organ cultures (Bécard and
Fortin, 1988). Particularly, an international sequencing program was launched to sequence
genome of the strain DAOM197198 of G. irregulare (Martin et al. 2008).
Beside the trophic incidence of AM symbiosis, or associated to this, it was observed
long ago that the AM fungi modify plant root system (Guether et al., 2009). Recent results on
early AM symbiosis signals strengthen this observation: myc-LCOs induce lateral root
formation (Olah et al., 2005, Maillet et al., 2011); AM fungi perceive molecules,
strigolactones, which were found to have hormonal incidence on plant architecture (Gomez-
Roldan et al., 2008; Umehara et al., 2008). In the same vein, the establishment of mycorrhizal
structures represses the exudation of strigolactones, as high concentration of phosphorus does
(Yoneyama et al., 2007; Lopez-Raez et al. 2010). These data argue for cross-talks between
mechanisms involved in AM symbiosis and root development. We here screen genetics and
genomics data on AM fungus-plant interaction that could provide new insights on these cross
talks.
Forward and reverse genetics for identifying myc mutants
The difficulty to identify the discrete structures of AM symbiosis (intraradical
arbuscules) was a great limitation to develop forward genetic programs. In the last decade, it
has been observed an overlap in Legume plants of symbiotic pathways between AM and
nitrogen-fixing symbioses from the characterization of plant mutants that are unable to
interact both with rhizobia (Nod- phenotype) and AM fungi (Myc- phenotype) (Marsh and
Schultze, 2001). This overlap was of great interest to identify the first Myc- mutants and the
orthologous genes in other plant families. Thereafter, forward and reverse genetics approaches
were developed. Due to the broad host spectrum for AM fungi, these studies were carried out
in a large variety of plant species.
Common Symbiotic Pathways among Legumes
Because of their requirement in both mycorrhizal and nodulating symbioses, the genes
involved in the early signaling pathway during symbiotic interaction was called “Common
Symbiosis Pathway” (CSP, Parniske et al., 2008). In Medicago truncatula, the CSP
Page 4
components are involved in the signal transduction after the perception of Myc-LCOs (Maillet
et al., 2011). To date, nine members of the CSP have been identified, in Medicago truncatula
and/or Lotus japonicas: a LysM receptor-like kinase of the MtNFP clade (Op den Camp et al.,
2010; Maillet et al., 2011), another receptor like kinase (MtDMI2 / LjSymRK, Endre et al.,
2002; Stracke et al., 2002), a cation channel (MtDMI1 / LjCASTOR-LjPOLLUX, Imaizumi-
Anraku et al., 2005) and three nucleoporins (LjNUP85, LjNUP133 and LjNENA, Kanamori
et al., 2006; Saito et al., 2007, Groth et al., 2010) involved in a specific nuclear calcium
spiking (Oldroyd & Downie, 2006). Downstream, a calcium and calmodulin-dependent
protein kinase (CCAMK) has been proposed to decode this specific calcium signature
(MtDMI3 / LjSYM15, Levy et al., 2004, Tirichine et al., 2006). NSP2, a transcription factor
of the GRAS family (Heckman et al., 2006, Maillet et al., 2011), and MtIPD3 / LjCYCLOPS,
which respectively physically interact with MtDMI3 and LjCYCLOPS, are downstream
targets of CCAMK in the signaling pathway (Messinese et al., 2007; Yano et al., 2008,
Parniske et al., 2008, Bonfante & Genre, 2010, Maillet et al., 2011). Mutants of the CSP show
a strong AM symbiosis defective phenotype with alteration in root penetration, intraradical
colonization and arbuscules formation.
Screening of mutant collections to phenotype AM symbiosis defective mutants
Systematic phenotyping of a large collection of mutants to identify AM symbiosis-
defective phenotype (Myc-) is a difficult approach as symbiotic markers are discrete
microscopic intraradical structures. In some species like maize and Brachypodium distachyon
(C. Roux, pers. com.), the accumulation of a yellow pigment during AM symbiosis, identified
as the apocarotenoid-derivate mycorradicin (Walter et al., 2000) can be used as a specific
marker for phenotyping. For these approaches, monocots are of great interest for screening
Myc mutants due to available mutagenesis system like transposon collections. Using these
features, two maize mutants -nop1 and taci1- were identified (Pazskowsky et al., 2006). The
nop1 mutant did not support hyphopodia formation, highlighting the role played by the host
plant to favor root colonization (Pazskowsky et al., 2006). The taci1 mutant was affected in
latter stage: fungal hyphae were septate and intraradical colonization was altered (Pazskowsky
et al., 2006). Mutagenesis strategies were also developed on Dicots. In Medicago truncatula
screening of an ethyl methanesulfonate collection enable the identification of two half-ABC
transporters STR1 and STR2 required for AM symbiosis (Zhang et al., 2010). In tomato two
mutants (pmi and pmi2) were characterized through similar screening (David-Schwartz et al.,
2001, 2003). These mutants showed a defect in pre-symbiotic steps, resulting in lower level of
Page 5
colonization. Finally, transposon mutagenized population of Petunia hybrida was also
screened for Myc phenotype (Reddy et al., 2007). The pam1 mutant issuing from this
screening was strongly affected in root penetration, colonization, in arbuscule formation and
in expression of the specific phosphate transporter (Feddermann et al., 2011). Very
interestingly, the pam1 insertion was map-cloned in a gene encoding an ortholog of the
VAPYRIN protein of Medicago truncatula (see below).
Candidates identified through specific transcriptomic and metabolic analyses
SbtM1 and SbtM3 are two subtilisin-like proteases upregulated in the mycorrhizal
roots of Lotus japonicas and RNAi of these genes reduce the number of arbuscules and
intraradical hyphae (Takeda et al., 2009). The CDPK1, a calcium dependent protein kinase
(Ivashuta et al., 2005), and VAPYRIN, a protein containing two protein-binding domains
(Pumplin et al., 2009), were found highly upregulated in Medicago truncatula during AM
symbiosis (Gomez et al., 2009). Corresponding RNAi lines showed strongly altered
mycorrhizal phenotype (Ivashuta et al., 2005; Pumplin et al., 2010). Components required in
the early steps of the interaction have also been identified by RNAi of genes previously
identified in specific transcriptomic analyses performed on Medicago truncatula. Thus,
MSBP1, which encodes a steroid binding protein, was upregulated at the vicinity of highly
branched fungal hyphae and the corresponding RNAi lines were strongly affected in
arbuscule formation (Kuhn et al., 2009).
As mentioned above, another strategy was phenotyping of mutants already
characterized for a specific metabolic function. The phosphate transport from the fungus to
the host plant is for instance a crucial metabolic trait of the AM symbiosis. Since the first
description of the relevance of a specific plant phosphate transporter in potato, StPT3 (Rausch
et al., 2001), similar transporters have been characterized in other dicots like Populus
trichocarpa (Loth-Pereda et al., 2011) and tomato (Nagy et al., 2005) and also in monocot
like rice (Paszkowsky et al., 2006). In Medicago truncatula the phosphate transporter, MtPT4
is specifically expressed in arbuscules of mycorrhizal roots (Harrison et al., 2002). The
corresponding mutant, Mtpt4 was impaired both in root colonization and arbuscule formation
(Javot et al., 2007). Interestingly, the formed arbuscules are aborted and extraradical fungal
growth is strongly limited. These works got the first evidence of the undeniable requirement
of “symbiotic” phosphate uptake for the maintenance of the AM symbiosis. The study of the
metabolic pathway of apocarotenoid illustrated the overlap of plant development process and
Page 6
AM symbiosis. Carotenoid Cleavage Dioxygenase 7 (CCD7) and 8 (CCD8) have been
previously identified in Pisum sativum, Oryza sativa, Arabidopsis thaliana, and Petunia
hybrida for their highly branched shoot phenotype (Sorefan et al., 2003; Booker et al., 2004,
Morris et al., 2001, Arite et al., 2007, Snowden et al., 2005, Drummond et al., 2009).
Independently, Matusova and co-authors (2005) proposed that the biosynthesis of
strigolactones, a compound released by host plants that induces seed germination of parasitic
weeds like Striga species, involved the cleavage of some carotenoid substrates by CCD
enzymes. Strigolactones were then describe as inducers of fungal growth and branching
(Akiyama et al., 2005, Besserer et al., 2006). Based on these observations, Gomez-Roldan
and co-authors (2008) and Umehara and coauthors (2008) tested respectively the ability of the
CCD7 and CCD8 mutants of pea (Psrms1, Psrms5) and rice (D10, D17) to produce
strigolactones. They demonstrated that strigolactone synthesis is dependent of these two
enzymes. In addition, it was demonstrated on pea that both mutants are unable to be colonized
by AMF species (Gomez-Roldan et al., 2008). These works, validated on two host species
belonging to Legumes and Poaceae, pointed out the role played by strigolactones as a new
plant hormone in the establishment of the AM symbiosis.
Mutant analysis reveals a biological integration between AM symbiosis and root
development.
As previously mentioned, the establishment of AM symbiosis modifies root
architecture. This modification of root architecture is not limited to AM interaction as
ectomycorrhizal fungi are able to induce modification of lateral root (LR) density by
producing plant hormones like auxin and ethylene (Felten et al., 2009; Splivallo et al., 2009).
By contrast, lateral root density modifications induced by AM fungi seem to be on the control
of AM fungal-specific signals. Treatment of A17 WT lines of Medicago truncatula with spore
exudates (Kosuta et al., 2003; Olah et al., 2005, Mukerjhe & Ané 2011) or synthetic Myc-
LCOs (Maillet et al., 2011) stimulated the development of new LR. The same treatments
performed on different mutants of the CSP did not affect the number of LR, arguing that the
LR response is dependent of the perception and transduction pathway of LCOs/COs. These
observations suggest the occurrence of a symbiotic program targeting the mechanisms
directing root development. The biological overlap of AM symbiosis and LR formation is
perfectly illustrated by the maize mutant lrt1 (Pazskowsky et al., 2002). lrt1 displays growth
defect by the lack of LR. Interestingly, this growth defect can be fully reversed after AM
fungal inoculation. Stimulation of LR density was fairly described in several species like rice
Page 7
or cherry plum (Gutjahr et al., 2009, Berta et al., 1995). A complementary illustration of this
overlap is that several of the AM defective mutants showed altered root architecture. For
instance, the root length of the cdpk1 mutant of Medicago truncatula was described as
significantly lower than WT plants and root hairs were also shorter (Ivashuta et al., 2005).
This growth defect is correlated to shorter cortical cells and altered cell wall organization and
composition. As root colonization by AMF requires strong cell wall remodeling (Genre et al.,
2005), altered cell wall could interfere on Myc phenotype.
The strigolactone story is a paradigm of the integration of the AM symbiosis and root
development mechanisms. In angiosperms, strigolactones are involved in the control of shoot
branching (Gomez-Roldan et al., 2008, Umehara et al., 2008). The root development of the
non mycotrophic Arabidopsis thaliana were analysed using the ccd7 and ccd8 mutants,
defective in strigolactone biosynthesis. Both mutants displayed a higher lateral root density
(Kapulnik et al., 2011) and shorter primary root (Ruyter-Spira et al., 2011). In addition,
exogenous treatment with the strigolactone synthetic analog GR24 on the two mutants
restored a WT phenotype. LR phenotype was also observed on corresponding pea mutants
(Delaux & Combier, unpublished data). It must be pointed out that LR are the main entry
point for AM fungal colonization. The increase of the global lateral root length would result
in higher mycorrhizal level by increasing the probability of contact between both partners.
The ccd7 and ccd8 pea mutants displayed lower root colonization, suggesting that the Myc-
phenotype of these mutants was more related to the lack of fungal stimulation by exogenous
stimulation than to plant hormonal function.
Comparative transcriptomics of AM symbiosis: towards identification of genes involved
in root development
First comparative transcriptomes to investigate root symbioses were performed on
Medicago truncatula in nodule and mycorrhizal roots (Journet et al., 2002). Due to overlap of
signaling mechanisms on nodulation and AM symbiosis in Legumes, nodulation as a positive
selection marker could have induced a specific evolution of the ancestral mechanism. For this
reason, comparative transcriptomics on Legumes and non-Legume species would provide new
informations about species-related and general mechanisms involved in AM symbiosis.
Transcriptomic profiles of AM symbiosis were produced in whole root system using different
strategies - EST analysis, suppressive-subtractive cDNA libraries and cDNA array
Page 8
hybridizations - during interaction with different AM fungi such as Glomus intraradices,
Glomus mossae and Gigaspora margarita (Journet et al., 2002; Küster et al., 2004; Siciliano
et al., 2007; López-Ráez et al., 2010 as examples). More recently, specific profiles during
AM symbiosis have been performed in arbuscule-containing cortical cells of mycorrhizal
roots using laser capture microdissection combined with microarray hybridization (Gaude et
al., 2011). Although all these data are informative independently, a comparative analysis of
transcriptomes during AM symbiosis is a complementary approach that could allow
identification of specific and shared mechanisms among plant species. We compared the
transcriptomes of mycorrhizal roots from different plant species by selecting data obtained in
similar experimental settings: transcriptomes in response to Glomus irregulare 197198 from
whole root system of Medicago truncatula (Hohnjec et al., 2005), Lotus japonicus (Guether et
al., 2009), Oryza sativa (Güimil et al., 2005) and Zea mays (Jourda et al., unpublished).
Comparing gene expression of different species needs to identify orthologous genes. The
comparative analysis was performed using OrthoMCL analysis and showed 236 orthologous
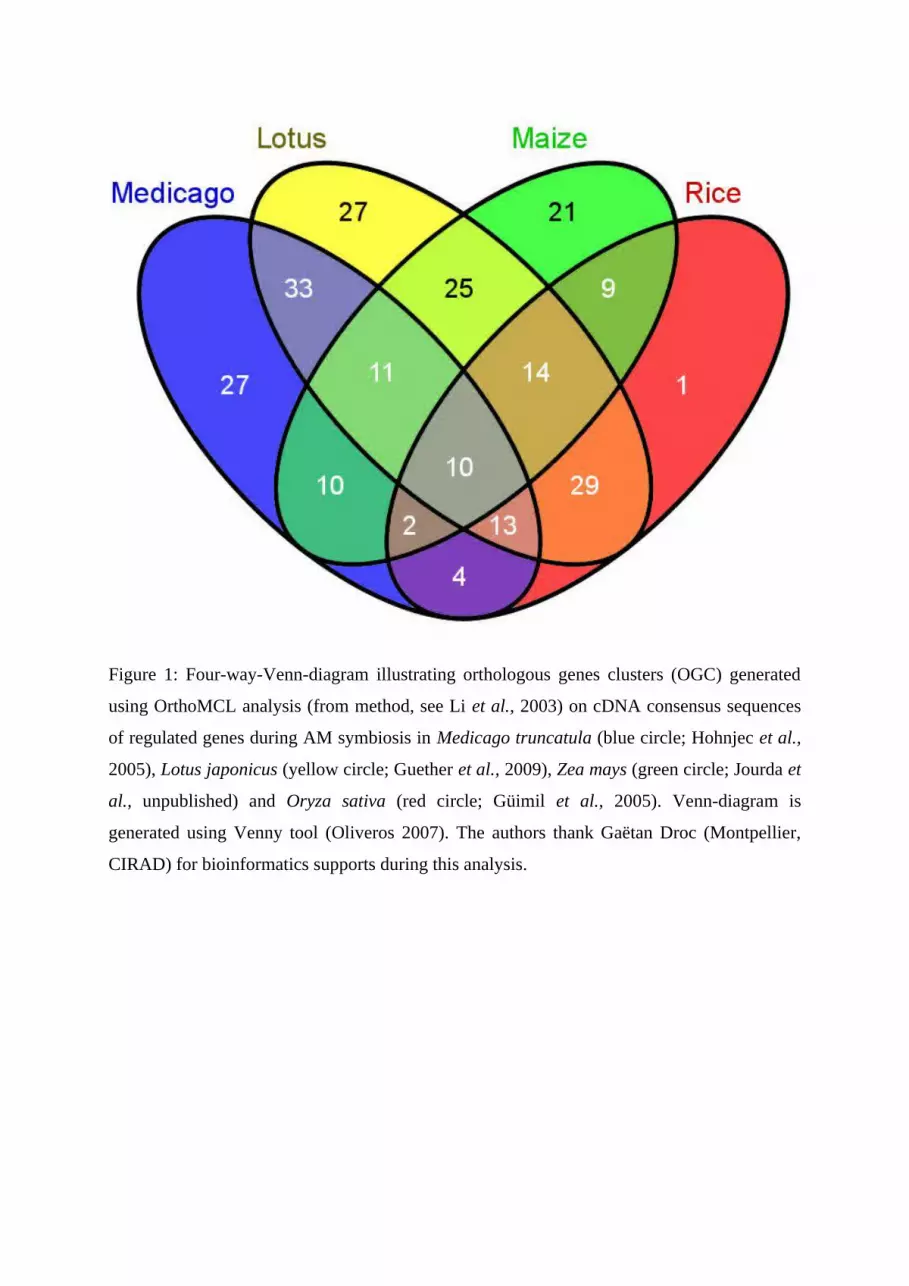
gene clusters (OGC; Fig. 1). It must be underlined that these data are partial as the
microarrays used for these different analyses were not representative of all the genome for
these species. From these data, 10 clusters are common to the 4 species, 54 clusters are
common to 3 species, and 76 were found only in one species. On the two Legume species,
202 clusters were identified, 64 were common to Medicago and Lotus and 87 were Legume-
or Dicot-specific (not found in Poaceae). On maize and rice, 139 clusters were found among
which 35 were common to the two species and 31 were Poaceae specific (not found in
Medicago and Lotus).
If we consider OGC putatively involved in root development that expression is
modified during AM symbiosis, 40 OGC comprising 105 ESTs were identified (Suppl._table
I) among which 10 (55 ESTs) were found in all plant species. Surprisingly none rice-specific
OGC putatively involved in root development has been found. Among the 40 OGC, 5 encode
for transcription factor families (MYB, NAC and GRAS families) potentially involved in the
control of root development, comprising 12 elements at the exception of rice gene. In Lotus
for example, 3 copies of OGC encoding for GRAS members are scarecrow protein coding
genes. Scarecrow members are considered as genetic determinants of root identity (Dolan
2007). All GRAS family members clustered were induced during AM symbiosis and could be
involved in modulation of root development during AM symbiosis. The OGC encoding NAC
transcription factors comprises only two repressed Medicago paralogs. Members of NAC
Page 9
transcription factor family could have conserved functions in secondary cell wall biosynthesis
and in root development (Christiansen et al., 2011) and so could be play a role in root
development by secondary cell wall regulation during AM symbiosis. MYB transcription
factors could be involved in controlling many diverse processes (Allan et al., 2008) such as
root hair formation in Arabidopsis thaliana (Slabaugh et al., 2011). 3 OGC encoding MYB
transcription factors have been detected with different expression profiles during AM
symbiosis. Members of MYB family could be involved in control of root hair formation
during AM symbiosis.
Root development is indeed related to cell elongation and then to cell wall
modifications. A set of 13 OGC that could be involved in degradation, modification,
biosynthesis and structure of cell wall were identified. The OGC related to cell wall
metabolism encode for proline-rich cell wall protein (extensin-like), 4-coumarate:coenzyme A
ligase, beta-glucanase, cinnamoyl-CoA reductase-like protein, expansin-related protein,
thaumatin-like protein, Caffeic acid O-methyltransferase, putative xyloglucan
endotransglycosylase and pectinesterase-like protein with different expression profiles
according to host plant. These differences could arise from diversity of plant cell walls
(Popper et al., 2011), especially between dicotyledonous and monocotyledonous species
(Yokoyama and Nishitani 2004). A good example of these differences is about OCG encoding
for proline-rich cell wall protein (extensin-like) constituted of only two Medicago paralogous
genes induced during AM symbiosis. Yokoyama and Nishitani (2004) have reviewed the
presence of extensin in model of type I walls but not in model of type II walls respectively
represented by Arabidopsis and rice cell walls. Structural cell wall proteins (such as
expansins and extensin-like proteins) are involved in the cleavage and the reassembly of cell
wall polymers necessary for cell elongation (Cosgrove, 2005). Regulation of these potential
agents for cell wall loosening (Cosgrove, 2001) could be a driver of root elongation during
AM symbiosis. An important subset of plant peroxidase coding genes clustered in an OGC
comprising 10 elements, but in absence of orthologous rice gene. However, in rice
mycorrhizal root, the strongest induced gene corresponded to a type III peroxidase which
showed a specific expression pattern in AM condition (Güimil et al., 2005). Peroxidase
enzymes are involved in hydrogen peroxide production and were described for their role in
cell wall organization and root elongation (Liszkay et al., 2004). These proteins could then be
involved in cell wall reorganization and lateral root formation during AM symbiosis. Lastly,
nodulin genes have been identified in these transcriptomic approaches. These genes have been
Page 10
identified as molecular markers of root nodule organogenesis (Crespi and Galvez 2000) and
could be common elements of signaling pathway between mycorrhization and nodulation of
plant roots. However the function of several nodulins are still to be described. Recently,
MtN21-like proteins were described as potentially transporter required for secondary cell wall
formation (Ranocha et al., 2010). Except in rice, at least one MtN21-like protein encoding
gene was found to be regulated during AM symbiosis. Three other OGC putatively encoding
for nodulin proteins were identified. For example, NOD26-like membrane intrinsic protein
(NIP) coding genes have been clustered. NIPs belong to the aquaporin superfamily and are
plant-specific with different functions and expression profiles (Liu et al., 2009).
Root development is obviously under hormone metabolism control (see Osmont et al., 2007).
Four OGC involved in hormone metabolism have been found and encode for Gibberellin-20-
oxidase 2, Zeatin O-xylosyltransferase, cytokinin-O-glucosyltransferase 2 and gibberellins
regulatory protein-like families. In this context, it is interesting to note that no common OGC
encoding auxin specific genes has been found. However, it could be relevant of different
auxinic response patterns for each plant species. The transcript Zm.5919.1.S1_at encodes a
putative auxin response factor and is induced during AM symbiosis in maize. Similar
observation can be noticed for rice and Medicago -induction respectively of OsAM173 and
MT000634 for example- but no putative auxin response factor has been regulated during AM
symbiosis in Lotus.
Nutrient transporter coding genes are an important subset of regulated genes in roots during
AM symbiosis with 12 OGC identified. The OGC encoding for putative phosphate transporter
includes MtPT4 (MT009707), OsPT11, Ljwgs_014433.2_at and Zm.1921.1.S1_at. In
Medicago, MtPT4 is a mycorrhiza-specific gene required for arbuscule development and
function (Javot et al., 2007). In rice, OsPT11 is a high affinity phosphate transporter
specifically induced during AM symbiosis (Paskowski et al., 2002). Moreover, in Lotus,
Ljwgs_014433.2_at is a phosphate transporter required specifically for AM symbiosis (Maeda
et al., 2006). In maize, a mutant lacking lateral root can be complemented by high phosphate
nutrition (Paskowski and Boller, 2002). Improvement of phosphate nutrition by phosphate
transporters specific to AM symbiosis could be involved in lateral root formation. In Lotus, a
putative ammonium transporter-encoding gene (Ljwgs_016680.1_at) is the strongest up-
regulated during AM symbiosis but this observation is not common to the 4 plant species.
Other OGC encoding hexose transporters, oligopeptide transporters, water transporters,
sulfate transporters and nitrate transporters have been found. Over-all, OGC involved in
Page 11
transport show an induction could be involved in an improvement of exchanges during AM
symbiosis in plant roots which could explain a part of branching process observed during AM
symbiosis (Berta et al., 1995).
Despite the low number of available transcriptome and that transcriptomic data are
incompletes, this comparative analysis is a first overview of potential important conserved
mechanisms involved in root development among plant species during AM symbiosis. This
approach can drive the definition of new key components involved in root development. It
must be noticed that transciptomes of AM symbiosis were performed using mycorrhizal root
system where the symbiosis is well established. Although AM symbiosis is a continuous
mechanism where extraradical hyphae can infect new developing roots, the use of well
mycorrhizal root system lead to a dilution of genes involved in early steps in favor to later
ones. The recent and exponential development of next generation sequencing (NGS)
technology gives new perspectives to perform global comparative transcriptome analysis of
plant root system using RNA-seq applications (Ozsolak and Milos, 2011).
Microtranscriptomics of AM symbiosis
Transcriptome analyses illustrated that the establishment of AM symbiosis requires a
massive reprogramming of the host genetic expression program. In the same time, there was
little information on the regulators controlling pathways as crucial as nutrient uptake and root
development during AM symbiosis (Krajinski and Frenzel, 2007). It can be suggested that
master regulator genes but also fine tuning regulators can be involved. The microRNAs are
small (about 21nt) negative regulators of gene expression present in plants. These small
RNAs are involved in developmental processes, hormonal signaling, nutrient balance, abiotic
and biotic stresses (Bartel, 2004; Jones-Rhoades et al., 2006; Mallory and Vaucheret, 2006,
Ruiz-Ferrer and Voinnet, 2009). MicroRNAs also play a role in plant-microorganism
interactions (Navarro et al., 2006; Subramanian et al., 2008). During nodule development in
Medicago truncatula, the miR169 is a regulator acting by targeting the transcriptional factor
MtHAP2-1 (Combier et al., 2006). It is also known that miR166 plays a crucial role in the
root and nodule development and its overexpression in Medicago truncatula leads to a
reduced number of lateral roots and symbiotic nodules (Boualem et al., 2008). As mentioned
above, the nodulation and the mycorrhizal symbioses share a common gene pathway called
CSP. Several analogies are found in the presymbiotic and symbiotic phases of these two
symbioses. Hence, it can be proposed that the regulation pathway could be either shared or
Page 12
discriminant between the symbioses according to the regulators. Recently, Maillet and co-
authors (2011) demonstrated that the miR171h-targeted GRAS transcription factor, MtNsp2,
essential for the signaling during the root nodule development (Oldroyd and Long 2003), is
also involved during AM fungal colonization. These results support the hypothesis that like
the nodulation, AM symbiosis could be finely regulated by microRNAs net. The microRNAs-
mediated phosphate (Pi) regulation can illustrate this hypothesis. Pi regulation has been
widely studied by the AM fungi research community because of the role of this symbiosis in
Pi acquisition. As illustrates in pea or petunia, AM symbiosis is suppressed under high Pi
condition (Balzergue et al., 2010; Breuillin et al., 2010). Some Pi transporters are specifically
expressed in Pi-starvation condition while some are specifically expressed during the AM
symbiosis (Chiou et al., 2001; Rausch et al. 2001; Harrison et al., 2002). The Pi and AM
signaling pathways share upstream regulatory factors and downstream structural genes as
revealed previously by transcriptomic analyses. One of these regulators is the MYB
transcription factor PHR1 that is a key element in signaling of primary Pi responses (Rubio et
al., 2001). PHR1 binds to a cis element and causes the transcription of many Pi starvation-
inducible genes, including members of the miR399 family (Bari et al., 2006). Some of these
microRNAs target PHO2 transcript and negatively regulate its expression (Allen et al. 2009).
PHO2 is responsible for the down-regulation of a subset of genes induces in response to Pi
starvation (Aung et al., 2006, Bari et al., 2006, Chiou et al., 2006). As a consequence,
miR399 can be considered as the Pi starvation signal that de-represses Pi starvation response
and the Pi uptake capacity (Branscheid et al. 2010). These authors showed the correlation
between miR399 expression, Pi homeostasis and AM development in plants. Interestingly,
they found that A. thaliana, a non-mycotrophic Brassicaceae, possesses less than half miR399
members than species capable of enhancing Pi uptake through AMF symbiosis, like M.
truncatula, rice or poplar. In parallel, they observed an accumulation increase of mature
miR399 in mycorrhizal roots comparing with non-mycorrhizal roots in M.truncatula and
tobacco. This suggests that, in mycorrhizal roots, PHO2 is involved in the suppression of AM
symbiosis in Pi-depleted plants because miR399 prevents its accumulation and activity in
response to the fungal Pi uptake. Thus, miR399 could be a regulator in maintaining of the AM
colonization. Gu and co-authors (2009) completed these analyses using microarray-based
approach on tomato. They identified a total of 14 miRNAs differentially expressed in a
mycorrhizal compared to non-mycorrhizal condition. Seven of these microRNAs were up-
regulated in both Pi sufficient level or AM symbiosis (miR158, miR169g*, miR172,
miR172b*, miR319, miR771 and miR775) while two are down-regulated (miR319 and
Page 13
miR394). Four are specifically up-regulated by AM symbiosis and their expression pattern
was similar in Pi sufficient or deficient conditions (miR395, miR779.1, miR840 and miR867).
Some of these miRNAs have been identified as regulators of the sulphate and water transport,
like miR395 (Kawashima et al., 2009) and miR840 (Gu et al., 2009), respectively.
Interestingly, Gu and co-authors (2009) observed that miR837-3p was completely suppressed
by AMF inoculation in leaves. In MTGI10 database, the putative miR837-3p targeted
transcript (TC143374) is similar to a Pathogenesis-related transcriptional factor and ERF1.
These data suggest a role of systemic defense of the miR837-3p that permits the accumulation
of the TC143374 in tomato during AMF colonization.
The availability of powerful genomic approaches in the small-RNAs world (Lu et al.,
2005) allowed a global view of the miRNA expression profile during the establishment of the
symbiosis. An analysis of the microtranscriptome in parallel of the degradome analysis has
been performed in the Krajinski’s lab (Devers et al., 2011). This approach has allowed
comparing the miRNA populations and the degradation of the corresponding potential targets
between AM and mock condition in the Medicago truncatula model. They discovered that a
lot of miRNAs are involved in AM symbiosis, as expected with this interaction targeting
hormonal, architectural, and molecular root processes. These authors propose that the
mycorrhizal symbiosis “leads to a reprogramming of the miRNA target network in roots,
including miRNA strand preference”. In this study, it has been showed that several
microRNAs are regulated during the AM symbiosis but, also, that many miRNA-targeted
genes are relevant to the symbiosis, like proteins implicated in the cellular phosphate
homeostasis. The nutrients uptake network is not the only regulatory system that is
overlapped with the mechanisms implemented by the AM symbiosis. As described before in
this chapter, AM fungi alter the root architecture of the host plant at both pre-symbiotic and
symbiotic stages. The in planta mechanisms required for these alteration remain unclear and
these results lead us to wonder whether some miRNAs implicated in the AM symbiosis can
also regulate the root architecture. The miR171h modulates the Nsp2 expression and then
could indirectly modify the architecture of the root system. The incidence of microRNA-
mediated regulation on symbiosis and root development was already demonstrated. The
microRNA166 for instance has a role on the nodule development and also in lateral root
formation (Boualem et al., 2008). Devers and co-authors (2011) have found that the miR166
is differentially regulated during the AM symbiosis suggesting that mir166 is playing a role in
this symbiosis, probably by regulating the lateral root formation. In previous studies, the
Page 14
microRNAs that regulate root development and architecture has been identified and reviewed
for Arabidopsis (dicotyledonous) and rice (monocotyledonous) (Meng et al., 2010), and more
globally (Kahn et al., 2011). In parallel, Devers and co-authors (2011) also found microRNAs
implicated in root architecture and development that are differentially expressed in the
mycorrhizal condition. A set of 6 microRNAs is common to the two processes (fig. 2). These
microRNAs are targeting transcription factors (HAP2, HD-ZipIII), auxin response factor
(ARF 6, 8, 10, 16, 17) and nutrient metabolism genes (SULTR2; APS1,4; CSD1,2). Some of
these microARNs are also overlapping with the nutrition signaling like miR169 (P), miR167
(N), miR395 (P, S) and miR398 (Zn, P).
As illustrated in Figure 2, the microRNAs expression pattern defined during the AM
symbiosis largely overlapped with specific microtranscriptome of root developmental
processes, as the nutrient homeostasis or root development and architecture. Other metabolic
pathways that are pertubed by the AMF colonization, like strigolactone regulations, remain to
be explored at the microtranscriptomic level. To investigate the relation between
mycorrhization and root development, the fine description of the AM symbiosis pre-symbiotic
steps will bring the opportunity to stress the involvement of microRNA regulation during
these steps. Similarly, specific investigations at the arbuscule-hosting cells and non-colonized
cortical cells, using laser dissection, cell sorting or reporter gene strategy, will allow to define
the identify locally expressed and repressed microRNAs. Finally, as informative are the
global approach of microRNA identification, long road remains to describe the functional
role of microRNAs in the fine tuning of AM symbiosis.
Page 15
Due to the ancestrally of this interaction, it was suggested that root development of early land
plants was accompanied by AM symbiosis. The evolutionary origin of these traits is often
discussed by authors and it will be difficult to resolve whether AM fungi promoted root tissue
differentiation as hypothesized by mycorrhizologists (see Brendett, 2002), or hijacked
previously formed mechanisms for root development. Whatever the evolutionary origin, as
physiopathology teaches us on cell physiology by highlighting perturbed cell mechanisms, the
study of the AM symbiosis can bring us new insights on root development. Genomics of AM
symbiosis are powerful approaches to identify overlaps of symbiotic pathways and root
developmental mechanisms.
References
Akiyama K, Matsuzaki K-ichi, Hayashi H. (2005). Plant sesquiterpenes induce hyphal
branching in arbuscular mycorrhizal fungi. Nature 435: 824-827.
Allan A.C., Hellens R.P. & Laing W.A. (2008). MYB transcription factors that colour our
fruit. Trends Plant Sci., 13, 99–102.
Allen, E., Xie, Z.X., Gustafson, A.M. and Carrington, J.C. (2005) microRNA-directed
phasing during trans-acting siRNA biogenesis in plants. Cell, 121, 207-221.
Ané J-M, Kiss GB, Riely BK, Penmetsa RV, Oldroyd GED, Ayax C, Lévy J, Debellé F, Baek
J-M, Kalo P, et al. (2004). Medicago truncatula DMI1 required for bacterial and fungal
symbioses in legumes. Science 303: 1364-1367.
Arite T, Iwata H, Ohshima K, Maekawa M, Nakajima M, Kojima M, Sakakibara H, Kyozuka
J. (2007). DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth
in rice. Plant Journal 51: 1019-1029
Aung, K., Lin, S.-I., Wu, C.-C., Huang, Y.-T., Su, C.-L. and Chiou, T.-J. (2006) pho2, a
phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 target
gene. Plant Physiology, 141, 1000-1011.
Page 16
Balzergue, C., Puech-Pagès, V., Bécard, G. and Rochange, S.F. (2011) The regulation of
arbuscular mycorrhizal symbiosis by phosphate in pea involves early and systemic
signalling events. J Exp Bot, 62, 1049-1060.
Bari, R., Pant, B.D., Stitt, M. and Scheible, W.-R. (2006) PHO2, microRNA399, and PHR1
define a phosphate-signaling pathway in plants. Plant Physiology, 141, 988-999.
Bartel, D.P. (2004) MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell, 116,
281-297.
Bécard, G. and Fortin, J.A. (1988) Early events of vesicular–arbuscular mycorrhiza formation
on Ri T-DNA transformed roots. New Phytologist, 108, 211-218.
Berta G., Trotta A., Fusconi A., Hooker J.E., Munro M., Atkinson D., Giovannetti M., Morini
S., Fortuna P., Tisserant B., Gianinazzi-Pearson V. & Gianinazzi S. (1995) Arbuscular
mycorrhizal induced changes to plant growth and root system morphology in Prunus
cerasifera. Tree Physiol., 15, 281-293.
Besserer A, Puech-Pagès V, Kiefer P, Gomez-Roldan V, Jauneau Alain, Roy S, Portais J-C,
Roux C, Bécard Guillaume, Séjalon-Delmas N. (2006). Strigolactones stimulate
arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biology 4: e226.
Booker J, Auldridge M, Wills S, McCarty D, Klee H, Leyser O. (2004). MAX3/CCD7 is a
carotenoid cleavage dioxygenase required for the synthesis of a novel plant signaling
molecule. Current Biology 14: 1232-1238.
Boualem, A., Laporte, P., Jovanovic, M., Laffont, C., Plet, J., Combier, J.-P., Niebel, A.,
Crespi, M. and Frugier, F. (2008) MicroRNA166 controls root and nodule development
in Medicago truncatula. Plant Journal, 54, 876-887.
Branscheid, A., Sieh, D., Pant, B.D., May, P., Devers, E.A., Elkrog, A., Schauser, L.,
Scheible, W.-R. and Krajinski, F. (2010) Expression Pattern Suggests a Role of MiR399
in the Regulation of the Cellular Response to Local Pi Increase During Arbuscular
Mycorrhizal Symbiosis. Molecular Plant-Microbe Interactions, 23, 915-926.
Breuillin, F., Schramm, J., Hajirezaei, M., Ahkami, A., Favre, P., Druege, U., Hause, B.,
Bucher, M., Kretzschmar, T., Bossolini, E., Kuhlemeier, C., Martinoia, E., Franken, P.,
Scholz, U. and Reinhardt, D. (2010) Phosphate systemically inhibits development of
Page 17
arbuscular mycorrhiza in Petunia hybrida and represses genes involved in mycorrhizal
functioning. Plant J, 64, 1002-1017.
Brundrett, M. C. (2002). Coevolution of roots and mycorrhizas of land plants. New
Phytologist 154: 275-304.
Chiou, T.J., Aung, K., Lin, S.I., Wu, C.C., Chiang, S.F. and Su, C.L. (2006) Regulation of
phosphate homeostasis by microRNA in Arabidopsis. Plant Cell, 18, 412-421.
Chiou, T.J., Liu, H. and Harrison, M.J. (2001) The spatial expression patterns of a phosphate
transporter (MtPT1) from Medicago truncatula indicate a role in phosphate transport at
the root/soil interface. Plant Journal, 25, 281-293.
Christiansen M.W., Holm P.B. & Gregersen P.L. (2011) Characterization of barley (Hordeum
vulgare L.) NAC transcription factors suggests conserved functions compared to both
monocots and dicots. BMC Res Notes., 4, 302-314.
Combier, J.-P., Frugier, F., de Billy, F., Boualem, A., El-Yahyaoui, F., Moreau, S., Vernie, T.,
Ott, T., Gamas, P., Crespi, M. and Niebel, A. (2006) MtHAP2-1 is a key transcriptional
regulator of symbiotic nodule development regulated by microRNA169 in Medicago
truncatula. Genes & Development, 20, 3084-3088.
Corkidi L, A.E., Merhaut D, Allen MF, Downer J, Bohn J, Evans M. (2004) Assessing the
infectivity of commercial mycorrhizal inoculants in plant nursery conditions. Journal of
Environmental Horticulture, pp. 149-154.
Cosgrove D.J. (2001) Wall structure and wall loosening. A look backwards and forwards.
Plant Physiol., 125, 131-134
Cosgrove D.J. (2005) Growth of the plant cell wall. Nat Rev Mol Cell Biol., 6, 850-861
Crespi M. and Galvez S. (2000) Molecular mechanisms in root nodule development. J Plant
Growth Regul., 19, 155-166.
DeMars B.G. and Boerner R.E.J. (1995) Arbuscular mycorrhizal development in three
crucifers. Mycorrhiza, 5(6): 405-408.
Devers, E.A., Branscheid, A., May, P. and Krajinski, F. (2011) Stars and Symbiosis:
MicroRNA- and MicroRNA*-Mediated Transcript Cleavage Involved in Arbuscular
Mycorrhizal Symbiosis. Plant Physiology, 156, 1990-2010.
Page 18
Dolan L. (2007). Plant science. SCARECROWs at the border. Science, 316, 377–378.
Drummond RSM, Martínez-Sánchez NM, Janssen BJ, Templeton KR, Simons JL, Quinn BD,
Karunairetnam S, Snowden KC. (2009). Petunia hybrida CAROTENOID CLEAVAGE
DIOXYGENASE7 is involved in the production of negative and positive branching
signals in petunia. Plant Physiology 151: 1867-1877.
Endre G, Kereszt A, Kevei Z, Mihacea S, Kaló P, Kiss GB. (2002). A receptor kinase gene
regulating symbiotic nodule development. Nature 417: 962-966.
Feddermann N, Muni RRD, Zeier T, Stuurman J, Ercolin F, Schorderet M, Reinhardt D.
(2010). The PAM1 gene of petunia, required for intracellular accommodation and
morphogenesis of arbuscular mycorrhizal fungi, encodes a homologue of VAPYRIN.
The Plant Journal 64: 470-481.
Felten J, Kohler A, Morin E, Bhalerao RP, Palme K, Martin F, Ditengou FA, Legué V.
(2009). The ectomycorrhizal fungus Laccaria bicolor stimulates lateral root formation in
poplar and Arabidopsis through auxin transport and signaling. Plant Physiol. 151:1991-
2005.
Finlay, R. D. (2008). Ecological aspects of mycorrhizal symbiosis: with special emphasis on
the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot.
59, 1115–1126 .
Foo E, Bullier E, Goussot M, Foucher F, Rameau C, Beveridge Christine Anne. (2005). The
branching gene RAMOSUS1 mediates interactions among two novel signals and auxin
in pea. The Plant Cell 17: 464-474.
Gaude N., Bortfeld S., Duensing N., Lohse M. & Krajinski F. (2011) Arbuscule-containing
and non-colonized cortical cells of mycorrhizal roots undergo a massive and specific
reprogramming during arbuscular mycorrhizal development. Plant J., [Epub ahead of
print]
Genre A, Chabaud M, Timmers T, Bonfante P, Barker DG. (2005). Arbuscular mycorrhizal
fungi elicit a novel intracellular apparatus in Medicago truncatula root epidermal cells
before infection. The Plant Cell 17: 3489-3499.
Page 19
Gomez-Roldan V, Fermas S, Brewer PB, Puech-Pagès V, Dun EA, Pillot J-P, Letisse F,
Matusova R, Danoun S, Portais J-C, et al. (2008). Strigolactone inhibition of shoot
branching. Nature 455: 189-194.
Groth M, Takeda N, Perry J, Uchida H, Dräxl S, Brachmann A, Sato S, Tabata S, Kawaguchi
M, Wang TL, et al. (2010). NENA, a Lotus japonicus homolog of Sec13, is required for
rhizodermal infection by arbuscular mycorrhiza fungi and rhizobia but dispensable for
cortical endosymbiotic development. The Plant Cell 22: 2509-2526.
Gu, M., Xu, K., Chen, A., Zhu, Y., Tang, G. and Xu, G. (2010) Expression analysis suggests
potential roles of microRNAs for phosphate and arbuscular mycorrhizal signaling in
Solanum lycopersicum. Physiologia Plantarum, 138, 226-237.
Guether M., Balestrini R., Hannah M., He J., Udvardi M.K. & Bonfante P. (2009). Genome-
wide reprogramming of regulatory networks, transport, cell wall and membrane
biogenesis during arbuscular mycorrhizal symbiosis in Lotus japonicas. New Phytol.,
182, 200-212.
Güimil S., Chang H.S., Zhu T., Sesma A., Osbourn A., Roux C., Ioannidis V., Oakeley E.J.,
Docquier M., Descombes P., Briggs S.P. & Paszkowski U. (2005) Comparative
transcriptomics of rice reveals an ancient pattern of response to microbial colonization.
Proc Natl Acad Sci U S A., 102, 8066-8070.
Gutjahr C, Casieri L, Paszkowski U. (2009). Glomus intraradices induces changes in root
system architecture of rice independently of common symbiosis signaling. The New
Phytologist 182: 829-837.
Imaizumi-Anraku H., Takeda N., Charpentier M., Perry J,. Miwa H., Umehara Y., Kouchi H.,
Murakami Y., Mulder L., Vickers K., Pike J., Downie J.A., Wang T., Sato S., Asamizu
E., Tabata S., Yoshikawa M., Murooka Y., Wu G.J., Kawaguchi M., Kawasaki S.,
Parniske M., Hayashi M. (2005) Plastid proteins crucial for symbiotic fungal and
bacterial entry into plant roots. Nature 433: 527-531.
Harrison, M.J., Dewbre, G.R. and Liu, J.Y. (2002) A phosphate transporter from Medicago
truncatula involved in the acquisiton of phosphate released by arbuscular mycorrhizal
fungi. Plant Cell, 14, 2413-2429.
Page 20
Helber, N. and Requena, N. (2008) Expression of the fluorescence markers DsRed and GFP
fused to a nuclear localization signal in the arbuscular mycorrhizal fungus Glomus
intraradices. New Phytol, 177, 537-548.
Helber, N., K. Wippel, N. Sauer, S. Schaarschmidt, B. Hause and N. Requena (2011) A
Versatile Monosaccharide Transporter That Operates in the Arbuscular Mycorrhizal
Fungus Glomus sp Is Crucial for the Symbiotic Relationship with Plants." The Plant
Cell Online.
Helgason, T., Watson, I.J. and Young, J.P. (2003) Phylogeny of the Glomerales and
Diversisporales (fungi: Glomeromycota) from actin and elongation factor 1-alpha
sequences. FEMS Microbiol Lett, 229, 127-132.
Hohnjec N., Vieweg M.F., Puhler A., Becker A. & Kuster H. (2005) Overlaps in the
transcriptional profiles of Medicago truncatula roots inoculated with two different
Glomus fungi provide insights into the genetic program activated during arbuscular
mycorrhiza. Plant Physiol., 137, 1283-1301
Horner-Devine, M.C., Leibold, M.A., Smith, V.H. & Bohannan, B.J.M. (2003). Bacterial
diversity patterns along a gradient of primary productivity. Ecol. Lett., 6, 613–622.
James, T.Y., Kauff, F., Schoch, C.L., Matheny, P.B., Hofstetter, V., Cox, C.J., Celio, G.,
Gueidan, C., Fraker, E., Miadlikowska, J., Lumbsch, H.T., Rauhut, A., Reeb, V.,
Arnold, A.E., Amtoft, A., Stajich, J.E., Hosaka, K., Sung, G.H., Johnson, D., O'Rourke,
B., Crockett, M., Binder, M., Curtis, J.M., Slot, J.C., Wang, Z., Wilson, A.W.,
Schüssler, A., Longcore, J.E., O'Donnell, K., Mozley-Standridge, S., Porter, D.,
Letcher, P.M., Powell, M.J., Taylor, J.W., White, M.M., Griffith, G.W., Davies, D.R.,
Humber, R.A., Morton, J.B., Sugiyama, J., Rossman, A.Y., Rogers, J.D., Pfister, D.H.,
Hewitt, D., Hansen, K., Hambleton, S., Shoemaker, R.A., Kohlmeyer, J., Volkmann-
Kohlmeyer, B., Spotts, R.A., Serdani, M., Crous, P.W., Hughes, K.W., Matsuura, K.,
Langer, E., Langer, G., Untereiner, W.A., Lücking, R., Büdel, B., Geiser, D.M.,
Aptroot, A., Diederich, P., Schmitt, I., Schultz, M., Yahr, R., Hibbett, D.S., Lutzoni, F.,
McLaughlin, D.J., Spatafora, J.W. and Vilgalys, R. (2006) Reconstructing the early
evolution of Fungi using a six-gene phylogeny. Nature, 443, 818-822.
Javot H, Penmetsa RV, Terzaghi N, Cook DR, Harrison MJ. (2007). A Medicago truncatula
phosphate transporter indispensable for the arbuscular mycorrhizal symbiosis.
Page 21
Proceedings of the National Academy of Sciences of the United States of America 104:
1720-1725.
Javot H., Penmetsa R.V., Terzaghi N., Cook D.R. & Harrison M.J. (2007) A Medicago
truncatula phosphate transporter indispensable for the arbuscular mycorrhizal
symbiosis. Proc Natl Acad Sci U S A., 104, 1720-1725.
Jones-Rhoades, M.W., Bartel, D.P. and Bartel, B. (2006) MicroRNAs and their regulatory
roles in plants. Annual Review of Plant Biology, 57, 19-53.
Journet E.-P., van Tuinen D., Gouzy J., Crespeau H., Carreau V., Farmer M.-J., Niebel A.,
Schiex T., Jaillon O., Chatagnier O., Godiard L., Micheli F., Kahn D., Gianinazzi-
Pearson V. & Gamas P. (2002) Exploring root symbiotic programs in the model legume
Medicago truncatula using EST analysis. Nucl. Acids Res., 30, 5579-5592.
Kaló P, Gleason C, Edwards A, Marsh J, Mitra RM, Hirsch S, Jakab J, Sims S, Long SR,
Rogers J, et al. (2005). Nodulation signaling in legumes requires NSP2, a member of the
GRAS family of transcriptional regulators. Science. 308: 1786-1789.
Kanamori N, Madsen Lene Heegaard, Radutoiu S, Frantescu M, Quistgaard EMH, Miwa H,
Downie JA, James EK, Felle HH, Haaning LL, et al. (2006). A nucleoporin is required
for induction of Ca2+ spiking in legume nodule development and essential for rhizobial
and fungal symbiosis. Proceedings of the National Academy of Sciences of the United
States of America 103: 359-364.
Kapulnik Y, Delaux P-M, Resnick Natalie, Mayzlish-Gati E, Wininger S, Bhattacharya C,
Séjalon-Delmas N, Combier J-P, Bécard Guillaume, Belausov E, et al. (2011).
Strigolactones affect lateral root formation and root-hair elongation in Arabidopsis.
Planta 233: 209-216.
Kawashima, C.G., Yoshimoto, N., Maruyama-Nakashita, A., Tsuchiya, Y.N., Saito, K.,
Takahashi, H. and Dalmay, T. (2009) Sulphur starvation induces the expression of
microRNA-395 and one of its target genes but in different cell types. Plant Journal, 57,
313-321.
Khan, G.A., Declerck, M., Sorin, C., Hartmann, C., Crespi, M. and Lelandais-Briere, C.
(2011) MicroRNAs as regulators of root development and architecture. Plant Molecular
Biology, 77, 47-58.
Page 22
Kiers E.T., Duhamel M., Beesetty Y., Mensah J.A., Franken O., Verbruggen E., Fellbaum
C.R., Kowalchuk G.A., Hart M.M., Bago A., Palmer T.M., West S.A.,
Vandenkoornhuyse P., Jansa J. and Bücking H. (2011) Reciprocal rewards stabilize
cooperation in the mycorrhizal symbiosis. Science. 12;333(6044):880-2.
Klironomos J, Zobel M, Tibbett M, Stock WD, Rillig MC, Parrent JL, Moora M, Koch AM,
Facelli JM, Facelli E, Dickie IA, Bever JD. (2011) Forces that structure plant
communities: quantifying the importance of the mycorrhizal symbiosis. New Phytol.
189(2):366-70.
Krajinski, F. and Frenzel, A. (2007) Towards the elucidation of AM-specific transcription in
Medicago truncatula. Phytochemistry, 68, 75-81.
Kuhn H, Küster H, Requena N. (2010). Membrane steroid-binding protein 1 induced by a
diffusible fungal signal is critical for mycorrhization in Medicago truncatula. New
Phytologist 185: 716-733.
Küster H., Hohnjec N., Krajinski F., El Yahyaoui F., Manthey K., Gouzy J., Dondrup M.,
Meyer F., Kalinowski J., Brechenmacher L., van Tuinen D., Gianinazzi-Pearson V.,
Pühler A., Gamas P. & Becker A. (2004) Construction and validation of cDNA-based
Mt6k-RIT macro- and microarrays to explore root endosymbioses in the model legume
Medicago truncatula. J Biotechnol., 108, 95-113.
Leake, J.R., Johnson, D., Donnelly, D.P., Muckle, G.E., Boddy, L. & Read, D.J. (2004).
Networks of power and influence: the role of mycorrhizal mycelium in controlling plant
communities and agroecosystem functioning. Can. J. Bot., 82, 1016–1045.
Lévy J, Bres C, Geurts René, Chalhoub B, Kulikova O, Duc G, Journet E-P, Ané J-M, Lauber
E, Bisseling T, et al. (2004). A putative Ca2+ and calmodulin-dependent protein kinase
required for bacterial and fungal symbioses. Science. 303: 1361-1364.
Li L., Stoeckert C.J. Jr. & Roos D.S. (2003). OrthoMCL: identification of ortholog groups for
eukaryotic genomes. Genome Res., 13, 2178-89.
Liszkay A., van der Zalm E. & Schopfer P. (2004) Production of reactive oxygen
intermediates (O(2)(.-), H(2)O(2), and (.)OH) by maize roots and their role in wall
loosening and elongation growth. Plant Physiol., 136, 3114-3123.
Page 23
Liu Q., Wang H., Zhang Z., Wu J., Feng Y. & Zhu Z. (2009) Divergence in function and
expression of the NOD26-like intrinsic proteins in plants. BMC Genomics., 10, 313-
325.
Liu, Y., Leigh, J.W., Brinkmann, H., Cushion, M.T., Rodriguez-Ezpeleta, N., Philippe, H. and
Lang, B.F. (2009) Phylogenomic analyses support the monophyly of Taphrinomycotina,
including Schizosaccharomyces fission yeasts. Mol Biol Evol, 26, 27-34.
Liu, Y.J., Hodson, M.C. and Hall, B.D. (2006) Loss of the flagellum happened only once in
the fungal lineage: phylogenetic structure of kingdom Fungi inferred from RNA
polymerase II subunit genes. BMC Evol Biol, 6, 74.
López-Ráez J.A., Verhage A., Fernández I., García J.M., Azcón-Aguilar C., Flors V. & Pozo
M.J. (2010) Hormonal and transcriptional profiles highlight common and differential
host responses to arbuscular mycorrhizal fungi and the regulation of the oxylipin
pathway. J Exp Bot., 61, 2589-601.
Loth-Pereda V, Orsini E, Courty P-E, Lota F, Kohler A, Diss L, Blaudez D, Chalot M, Nehls
U, Bucher M, et al. (2011). Structure and expression profile of the phosphate Pht1
transporter gene family in mycorrhizal Populus trichocarpa. Plant Physiology.
Lu, C., Kulkarni, K., Souret, F.F., MuthuValliappan, R., Tej, S.S., Poethig, R.S., Henderson,
I.R., Jacobsen, S.E., Wang, W., Green, P.J. and Meyers, B.C. (2006) MicroRNAs and
other small RNAs enriched in the Arabidopsis RNA-dependent RNA polymerase-2
mutant. Genome Research, 16, 1276-1288.
Maeda, D., Ashida, K., Iguchi, K., Chechetka, S.A., Hijikata, A., Okusako, Y., Deguchi, Y.,
Izui, K. and Hata, S. (2006) Knockdown of an arbuscular mycorrhiza-inducible
phosphate transporter gene of Lotus japonicus suppresses mutualistic symbiosis. Plant
Cell Physiol. 47, 807–817.
Maillet, F., Poinsot, V., Andre, O., Puech-Pages, V., Haouy, A., Gueunier, M., Cromer, L.,
Giraudet, D., Formey, D., Niebel, A., Martinez, E.A., Driguez, H., Becard, G. and
Denarie, J. (2011) Fungal lipochitooligosaccharide symbiotic signals in arbuscular
mycorrhiza. Nature, 469, 58-U1501.
Mallory, A.C. and Vaucheret, H. (2006) Functions of microRNAs and related small RNAs in
plants. Nature Genetics, 38, S31-S36.
Page 24
Marsh, J. F., and Schultze, M. 2001. Analysis of arbuscular mycorrhizas using symbiosis-
defective plant mutants. New Phytol. 150:525-532.
Martin, F., Gianinazzi-Pearson, V., Hijri, M., Lammers, P., Requena, N., Sanders, I.R.,
Shachar-Hill, Y., Shapiro, H., Tuskan, G.A. and Young, J.P.W. (2008) The long hard
road to a completed Glomus intraradices genome. New Phytologist: Wiley Online
Library.
Matusova R, Rani K, Verstappen FWA, Franssen MCR, Beale MH, Bouwmeester HJ. (2005).
The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche
spp. are derived from the carotenoid pathway. Plant Physiology 139: 920-934.
Meng, Y., Ma, X., Chen, D., Wu, P. and Chen, M. (2010) MicroRNA-mediated signaling
involved in plant root development. Biochemical and Biophysical Research
Communications, 393, 345-349.
Messinese E, Mun J-H, Yeun LH, Jayaraman D, Rougé P, Barre A, Lougnon G, Schornack S,
Bono J-J, Cook DR, et al. (2007). A novel nuclear protein interacts with the symbiotic
DMI3 calcium- and calmodulin-dependent protein kinase of Medicago truncatula.
Molecular Plant-Microbe Interactions: MPMI 20: 912-921.
Morris SE, Turnbull CG, Murfet IC, Beveridge C A. (2001). Mutational analysis of branching
in pea. Evidence that Rms1 and Rms5 regulate the same novel signal. Plant Physiology
126: 1205-1213.
Mukherjee A, Ané J-M. (2011). Germinating spore exudates from arbuscular mycorrhizal
fungi: molecular and developmental responses in plants and their regulation by
ethylene. Molecular Plant-Microbe Interactions: MPMI 24: 260-270.
Nagy R, Karandashov V, Chague V, Kalinkevich K, Tamasloukht M’barek, Xu G, Jakobsen
I, Levy AA, Amrhein N, Bucher M. (2005). The characterization of novel mycorrhiza-
specific phosphate transporters from Lycopersicon esculentum and Solanum tuberosum
uncovers functional redundancy in symbiotic phosphate transport in solanaceous
species. The Plant Journal. 42: 236-250.
Navarro, L., Dunoyer, P., Jay, F., Arnold, B., Dharmasiri, N., Estelle, M., Voinnet, O. and
Jones, J.D.G. (2006) A plant miRNA contributes to antibacterial resistance by
repressing auxin signaling. Science, 312, 436-439.
Page 25
Olah, B., Briere, C., Becard, G., Denarie, J. and Gough, C. (2005) Nod factors and a diffusible
factor from arbuscular mycorrhizal fungi stimulate lateral root formation in Medicago
truncatula via the DMI1/DMI2 signalling pathway. Plant Journal, 44, 195-207.
Oldroyd, G.E.D. and Long, S.R. (2003) Identification and characterization of nodulation-
signaling pathway 2, a gene of Medicago truncatula involved in Nod factor signaling.
Plant Physiology, 131, 1027-1032.
Oliveros J.C. (2007) VENNY. An interactive tool for comparing lists with Venn Diagrams.
(http:// bioinfogp.cnb.csic.es/tools/venny/index.html).
Osmont K.S, Sibout R. and Hardtke C.S. (2007) Hidden branches: developments in root
system architecture. Annu Rev Plant Biol., 58, 93-113.
Ozsolak F. and Milos P.M. (2011) RNA sequencing: advances, challenges and opportunities.
Nat Rev Genet., 12, 87-98.
Parniske M. (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat Rev
Microbiol. 26:763-775.
Paszkowski U and Boller T. (2002) The growth defect of lrt1, a maize mutant lacking lateral
roots, can be complemented by symbiotic fungi or high phosphate nutrition. Planta, 214,
584-590.
Paszkowski U., Kroken S., Roux C. and Briggs S.P. (2002) Rice phosphate transporters
include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal
symbiosis. Proc Natl Acad Sci U S A., 99, 13324-13329.
Pearson JN & Jakobsen I. (1993). The relative contribution of hyphae and roots to phosphorus
uptake by arbuscular mycorrhizal plants measured by dual labelling with 32P and 33P.
New Phytologist 124: 489–494.
Popper, Z.A., Michel, G., Hervé, C., Domozych, D.S., Willats, W.G., Tuohy, M.G., Kloareg,
B. and Stengel, D.B. (2011) Evolution and diversity of plant cell walls: from algae to
flowering plants. Ann. Rev. Plant Biol., 62, 567-590
Price, N.S., Roncadori, R.W. and Hussey, R.S. (1989) Cotton root-growth as influenced by
phosphorus-nutrition and vesicular arbuscular mycorrhizas. New Phytologist, 111, 61-
66.
Page 26
Pumplin N., Mondo S.J,. Topp S., Starker C.G., Gantt J.S. and Harrison M.J. (2010).
Medicago truncatula Vapyrin is a novel protein required for arbuscular mycorrhizal
symbiosis. The Plant Journal: For Cell and Molecular Biology 61: 482-494.
Ranocha P., Denancé N., Vanholme R., Freydier A., Martinez Y., Hoffmann L., Köhler L.,
Pouzet C., Renou J.P., Sundberg B., Boerjan W. & Goffner D. (2010) Walls are thin 1
(WAT1), an Arabidopsis homolog of Medicago truncatula NODULIN21, is a
tonoplast-localized protein required for secondary wall formation in fibers. Plant J., 63,
469-483.
Rausch, C., Daram, P., Brunner, S., Jansa, J., Laloi, M., Leggewie, G., Amrhein, N. and
Bucher, M. (2001). A phosphate transporter expressed in arbuscule-containing cells in
potato. Nature 414, 462–466.
Redecker, D. and Raab, P. (2006) Phylogeny of the glomeromycota (arbuscular mycorrhizal
fungi): recent developments and new gene markers. Mycologia, 98, 885-895.
Redecker, D., Kodner, R. and Graham, L.E. (2000) Glomalean fungi from the Ordovician.
Science, 289, 1920-1921.
Remy, W., Taylor, T.N., Hass, H. and Kerp, H. (1994) Four hundred-million-year-old
vesicular arbuscular mycorrhizae. Proc Natl Acad Sci U S A, 91, 11841-11843.
Rubio, V., Linhares, F., Solano, R., Martin, A.C., Iglesias, J., Leyva, A. and Paz-Ares, J.
(2001) A conserved MYB transcription factor involved in phosphate starvation
signaling both in vascular plants and in unicellular algae. Genes & Development, 15,
2122-2133.
Ruiz-Ferrer, V. and Voinnet, O. (2009) Roles of Plant Small RNAs in Biotic Stress
Responses. Annual Review of Plant Biology, 60, 485-510.
Ruyter-Spira C, Kohlen W, Charnikhova T, van Zeijl A, van Bezouwen L, de Ruijter N,
Cardoso C, Lopez-Raez JA, Matusova R, Bours R, et al. (2011). Physiological Effects
of the Synthetic Strigolactone Analog GR24 on Root System Architecture in
Arabidopsis: Another Belowground Role for Strigolactones? Plant Physiology 155:
721-734.
Page 27
Saito K, Yoshikawa M, Yano K, Miwa H, Uchida H, Asamizu E, Sato S, Tabata S, Imaizumi-
Anraku H, Umehara Y, et al. (2007). NUCLEOPORIN85 is required for calcium
spiking, fungal and bacterial symbioses, and seed production in Lotus japonicus. The
Plant Cell 19: 610-624.
Schnitzer SA, Klironomos J. (2011) Soil microbes regulate ecosystem productivity and
maintain species diversity. Plant Signal Behav. 1;6(8)
Schüβler, A., Schwarzott, D. and Walker, C. (2001) A new fungal phylum, the
Glomeromycota: phylogeny and evolution. Mycological Research, 105, 1413-1421.
Schwarzott, D., Walker, C. and Schüssler, A. (2001) Glomus, the largest genus of the
arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Mol Phylogenet Evol,
21, 190-197.
Siciliano V., Genre A., Balestrini R., Cappellazzo G., deWit P.J.G.M. & Bonfante P. (2007)
Transcriptome Analysis of Arbuscular Mycorrhizal Roots during Development of the
Prepenetration Apparatus. Plant Physiol., 144, 1455-1466.
Slabaugh E., Held M. & Brandizzi F. (2011) Control of root hair development in Arabidopsis
thaliana by an endoplasmic reticulum anchored member of the R2R3-MYB
transcription factor family. Plant J., 67, 395-405
Smith S.E. and Read D.J. (1997) Mycorrhizal symbiosis. Academic Press, 2: 409-452
Snowden KC, Simkin AJ, Janssen BJ, Templeton KR, Loucas HM, Simons JL,
Karunairetnam S, Gleave AP, Clark DG, Klee HJ. (2005). The Decreased apical
dominance1/Petunia hybrida CAROTENOID CLEAVAGE DIOXYGENASE8 gene
affects branch production and plays a role in leaf senescence, root growth, and flower
development. The Plant Cell 17: 746-759.
Sorefan K, Booker Jon, Haurogné K, Goussot M, Bainbridge K, Foo E, Chatfield S, Ward S,
Beveridge C, Rameau C, Leyser O. (2003). MAX4 and RMS1 are orthologous
dioxygenase-like genes that regulate shoot branching in Arabidopsis and pea. Genes &
Development 17: 1469-1474.
Splivallo R, Fischer U, Göbel C, Feussner I, Karlovsky P. (2009). Truffles regulate plant root
morphogenesis via the production of auxin and ethylene. Plant Physiol. 150:2018-2029
Page 28
Stracke S, Kistner C, Yoshida S, Mulder L, Sato S, Kaneko T, Tabata S, Sandal N, Stougaard
J, Szczyglowski K, et al. (2002). A plant receptor-like kinase required for both bacterial
and fungal symbiosis. Nature 417: 959-962.
Subramanian, S., Fu, Y., Sunkar, R., Barbazuk, W.B., Zhu, J.-K. and Yu, O. (2008) Novel
and nodulation-regulated microRNAs in soybean roots. Bmc Genomics, 9.
Takeda N, Sato S, Asamizu E, Tabata S, Parniske M. (2009). Apoplastic plant subtilases
support arbuscular mycorrhiza development in Lotus japonicus. Plant Journal 58: 766-
777.
Tirichine L, Imaizumi-Anraku H, Yoshida S, Murakami Y, Madsen Lene H, Miwa H,
Nakagawa T, Sandal N, Albrektsen AS, Kawaguchi M, et al. (2006). Deregulation of a
Ca2+/calmodulin-dependent kinase leads to spontaneous nodule development. Nature
441: 1153-1156
Umehara, M., A. Hanada, S. Yoshida, K. Akiyama, T. Arite, N. Takeda-Kamiya, H. Magome,
Y. Kamiya, K. Shirasu, K. Yoneyama, J. Kyozuka and S. Yamaguchi (2008). Inhibition
of shoot branching by new terpenoid plant hormones. Nature 455(7210): 195-200.
van der Heijden MG, Bardgett RD and van Straalen NM. Ecol Lett. 2008 The unseen
majority: soil microbes as drivers of plant diversity and productivity in terrestrial
ecosystems. 11(3):296-310.
van Rhijn P., Fang Y., Galili S., Shaul O., Atzmon N., Wininger S., Eshed Y., Lum M., Li Y.,
To V., Fujishige N., Kapulnik Y. and Hirsch A.M. (1997) Expression of early nodulin
genes in alfalfa mycorrhizae indicates that signal transduction pathways used in forming
arbuscular mycorrhizae and Rhizobium-induced nodules may be conserved. Proc Natl
Acad Sci U S A, 94, 5467-5472.
Walter, M. H., T. Fester and D. Strack (2000). Arbuscular mycorrhizal fungi induce the non-
mevalonate methylerythritol phosphate pathway of isoprenoid biosynthesis correlated
with accumulation of the 'yellow pigment' and other apocarotenoids. The Plant Journal
21(6): 571-578.
Yano, K., Yamauchi, A. and Kono, Y. (1996) Localized alteration in lateral root development
in roots colonized by an arbuscular mycorrhizal fungus. Mycorrhiza, 6, 409-415.
Page 29
Yokoyama R. and Nishitani K. (2004) Genomic basis for cell-wall diversity in plants. A
comparative approach to gene families in rice and Arabidopsis. Plant Cell Physiol., 45,
1111-1121.
Yoneyama, K., K. Yoneyama, Y. Takeuchi and H. Sekimoto (2007). Phosphorus deficiency
in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts
and germination stimulant for root parasites. Planta 225: 1031-1038.
Page 30
Figure 1: Four-way-Venn-diagram illustrating orthologous genes clusters (OGC) generated
using OrthoMCL analysis (from method, see Li et al., 2003) on cDNA consensus sequences
of regulated genes during AM symbiosis in Medicago truncatula (blue circle; Hohnjec et al.,
2005), Lotus japonicus (yellow circle; Guether et al., 2009), Zea mays (green circle; Jourda et
al., unpublished) and Oryza sativa (red circle; Güimil et al., 2005). Venn-diagram is
generated using Venny tool (Oliveros 2007). The authors thank Gaëtan Droc (Montpellier,
CIRAD) for bioinformatics supports during this analysis.
Page 31
Figure 2: Venn diagram of the microRNA families repartition and overlapping. Blue circle
contains the microRNAs involving in nutrition signaling, red circle contains the microRNAs
involved in the root architecture and development and green circle contains the microRNAs
involved in the mycorrhization (see text for references)
Root architecture and development
Nutrition signalingMycorrhization
miR160, miR166
miR390, miR393,
miR164
miR167, miR169, miR395, miR398
miR156, miR778, miR827, miR2111 miR158,
miR172, miR319, miR399, miR771, miR775,miR840
miR162, miR171, miR396, miR837, miR867, miR2086, miR4414, miR5204, miR5206, miR5213, miR5229, miR5232, miR5244, miR5250, miR5281
Page 32
Supp. Data Table I: Table of a subset of 40 orthologous genes clusters (OGC) generated using
OrthoMCL analysis (from method, see Li et al., 2003) on cDNA consensus sequences of
regulated genes putatively involved in root development during AM symbiosis in Medicago
truncatula (Hohnjec et al., 2005), Lotus japonicus (Guether et al., 2009), Oryza sativa
(Güimil et al., 2005) and Zea mays (Jourda et al., unpublished). Cluster annotation
(metabolism and function), plant species with gene identifying, annotation and regulation
during AM symbiosis are indicated. Signs + and - indicate gene is up-regulated or down-
regulated, respectively. Gene ID correspond to Affymetrix Gene Chip identifying available on
http://www.affymetrix.com .
Metabolism Annotation Species Gene ID RegulationCell wall metabolism
4-coumarate--CoA ligase - like protein
Lotus Ljwgs_075474.1_at +Maize Zm.15085.1.A1_at +Medicago MT000912 -
Caffeic acid O-methyltransferase Medicago MT005666 -Medicago MT007074 -
Cinnamoyl CoA reductase - like protein
Lotus Ljwgs_100777.1_s_at +Medicago MT000507 -Rice OsAM163 +Rice OsAM49 +
endo-1,3(4)-beta-glucanase Lotus chr4.CM0119.5_at +Medicago MT003194 +
Expansin protein Lotus Ljwgs_041638.1_s_at +Maize Zm.17372.1.A1_at -
Expansin-related protein Lotus Ljwgs_012501.1_at -Medicago MT000967 -
Extensin like protein Medicago MT007031 +Medicago MT007032 +
Osmotin-like protein/ Putative thaumatin
Medicago MT001796 -Medicago MT007463 +
Pectinesterase like protein Lotus Ljwgs_031539.1_at +Medicago MT001341 +Medicago MT007664 +
Pectinesterase like protein Lotus chr1.TM1573.5_at -Maize Zm.7152.1.A1_at +Medicago MT006917 +
Putative Endo-1,3;1,4-beta-D-glucanase
Medicago MT000950 +Rice OsAM222 -
Putative xyloglucan endotransglycosylase
Lotus Ljwgs_022629.1_at -Maize Zm.704.1.S1_at +Rice OsAM220 -
Zeamatin/ thaumatin-like protein Maize Zm.281.1.S1_s_at +Medicago MT000966 -
Page 33
Hormone metabolism
Cytokinin-O-glucosyltransferase 2
Maize Zm.9570.1.A1_a_at +Maize Zm.9570.2.A1_x_at +
Gibberellin 20 oxidase 2 Lotus chr5.CM0953.24_at +Lotus Ljwgs_018141.1_at +Maize Zm.13480.1.S1_at +Medicago MT003693 +
Gibberellin regulatory protein like
Lotus chr5.CM0239.51_at +Lotus chr6.CM0539.6_at +
ZEATIN O-XYLOSYLTRANSFERASE
Lotus Ljwgs_037161.1_at +Lotus Ljwgs_037161.1_x_at +
MISC MtN19-like protein Lotus chr6.CM0437.7_at +Medicago MT015318 -Medicago MT015656 -Rice OsAM197 +Lotus chr4.CM0337.32.7_at +Medicago MT001044 +Medicago MT001597 +
Multifunctional Nodulin 26-like aquaporin
Lotus chr4.CM0046.51_at +Medicago MT007526 +Rice OsAM143 +
Peroxidase Lotus Ljwgs_018430.1_s_at -Maize Zm.11214.1.S1_at +Maize Zm.10660.1.A1_at +Maize Zm.5170.1.S1_at -Medicago MT001052 -Medicago MT010277 -Medicago MT002659 +Medicago MT007740 +Medicago MT007030 +Medicago MT008600 +
Putative nodulin Lotus Ljwgs_119620.1_at +Medicago MT013567 +Rice OsAM129 +
Transcription factor
GRAS family Lotus Ljwgs_016263.1_at +Lotus Ljwgs_023888.1_at +Lotus Ljwgs_027761.2_at +Maize Zm.6402.2.A1_at +
MYB family Lotus chr1.BM1732.4_at +Medicago MT001930 +
MYB family Lotus Ljwgs_014616.1_at -Medicago MT007392 +
MYB family Maize Zm.13885.1.S1_s_at -Medicago MT002218 -
NAC family Medicago MT000799 -Medicago MT009487 -
Transport Aquaporin Maize Zm.606.1.A1_at -Maize Zm.607.1.A1_at -
Aquaporin Lotus Ljwgs_149324.1_at +Maize Zm.612.1.A1_at -
Page 34
Aquaporin Lotus TM0748.11_at +Maize Zm.602.1.A1_a_at -
High affinity nitrate transporter / Membrane transporter
Medicago MT002501 +Medicago MT009589 +
Oligopeptide transporter-like protein / Putative nitrate transporter
Lotus chr6.CM0118.43_at +Medicago MT006556 +Rice OsAM60 +
Peptide transporter Lotus chr1.CM0295.1_s_at +Lotus chr1.CM0295.2.1_at +
Peptide transporter like Lotus chr2.CM0903.44_at -Maize Zm.17744.1.A1_at +
Phosphate transporter Lotus Ljwgs_014433.2_at +Maize Zm.1921.1.S1_at +Medicago MT009707 +Rice OsPT11 +
Putative ammonium transporter Lotus Ljwgs_016680.1_at +Rice OsAM76 +
Putative sucrose transport protein Lotus chr5.CM0344.52_at -Maize Zm.199.1.S1_s_at +
Sugar transporter like protein Lotus Ljwgs_024490.1_s_at -Medicago MT000349 -
Sulfate tansporter Lotus chr6.CM0314.34_at +Lotus Ljwgs_011755.1_at +Maize Zm.11651.1.A1_at -