

Absorption Spectroscopy/Protein Function
Topic 4 Part 2
Biophysics
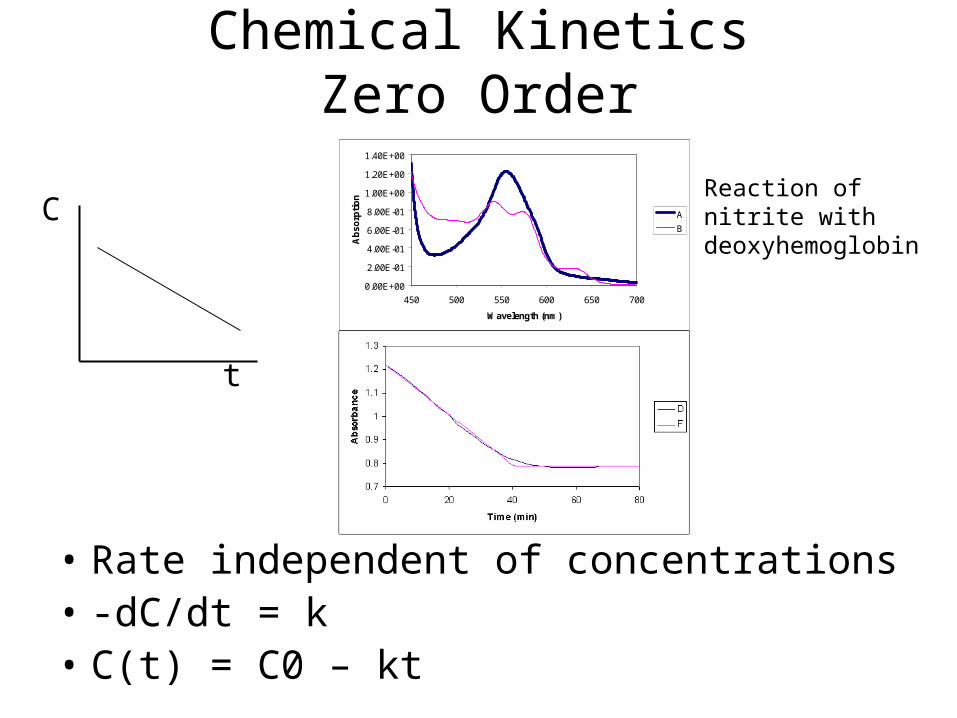
Chemical KineticsZero Order
• Rate independent of concentrations• -dC/dt = k • C(t) = C0 – kt
C
t
0.00E+00
2.00E-01
4.00E-01
6.00E-01
8.00E-01
1.00E+00
1.20E+00
1.40E+00
450 500 550 600 650 700
Wavelength (nm)
Ab
sorp
tio
n
A
B
Reaction of nitrite with deoxyhemoglobin

Chemical KineticsFirst Order
• -dCA/dt = kCA , CA = CA0 e-kt
• t1/2 = ln(2)/k; = 1/k = lifetime
ln(CA)
t
NO binding to Hb
ZA k

Chemical KineticsSecond Order
• -dCA/dt = -dCB/dt = kCACB
• Make one species in excess so get pseudofirst order kinetics, kobs = kCB so
CA = CA0 exp(-kobst)
0B
0A0
B0A
B
A
CClnktCC
C
Cln
oductsPrBA k

Hemoglobin
Cooperative Binding of Oxygen
Linked to quaternary structure
Explained by MWC Model

On the Nature of Allosteric Transitions:
A Plausible ModelJacques Monod, Jeffries Wyman,
Jean-Pierre Changux
J. Mol. Biol. 1965

“Molecular Amplifiers”

ATCase
The goal is control – want a switch.
“Indirect interactions between distinct specific binding sites (allosteric effects)”

Definitions and Generalizations
• Homotrophic effects – identical ligands (eg. for Hb: O2, CO, NO)
• Heterotrophic effects –different ligands (eg. for Hb: DPG, IHP, Cl-, NO as SNO, NEM)
• Most allosteric proteins are oligomers (several subunits or protomers)
• Allosteric changes often involve quaternary stucture
• Heterotrophic - positive or negative, Homotrophic – only positive (exception of Hg reductase?)

Model in English
1. Allosteric proteins are oligomers where the protomers are arranged symmetrically
2. There is one and only one identical ligand-binding site on each protomer
3. Tertiary structure of protomers affected by quaternary structure
4. There are two quaternary states (R and T) which dictate ligand affinities on all protomers
5. Transitions between states preserve symmetry

Model in Math
• T0 = L R0, , L is the allosteric constant, (Big L = Big allostery)
• Only also define
RT
R
K
Fc
K
K
c defines relative affinities of quaternary states and defines absolute affinity of one
When L is smalln
n
F LY
)1(
)1( 1

Compare Hill Equation
n
n
F LY
)1(
)1( 1
n
n
Q
YFvs
Q is a constant, n is the number of ligand sites, n is Hill coefficient

Hb, L, c …

Hb Sigmoidal
See satsimple.mw and sat.mw

Heterotrophic effectorsAffect L
Activators decrease L (push to R) and
Inhibitors increase it
AIn
n
n
n
F
K
A
K
ILL
LY
, ,)1(
)1('
)1('
)1(
1

See sat.mw

Hb – microstate predictions (vs sequential)
Unlike in sequential model, no R2 or T2 – see states.mw

ATCase and inhibitor
• At low concentrations of substrate, low concentrations of analogue activate (by promoting R-state) upper curve
• Desensitized enzyme (quaternary interactions suppressed) shows no increase in activity by analoque (maleate)
• Generally, desensitized enzymes lose cooperativity. Hb dimers are R-state like and like Mb. Homotrophic ligands promote tetramer stabilization (hard to dissociate oxyHb), as predicted

Activators can decrease cooperativity
Fig 6a is theoretical (see yf.mw )
Fig 6b and c show activations in real systems

Confirmations of MWCATCase
• Model predicts fraction in R-state > fraction ligand bound. Schachman lab (1966) shows this using sedimentation to examine quaternary state (size) and spectroscopy for ligation.
• They also showed (like Gerhart lab) low concentration of inhibitor activate ATCase

Confirmations - Hb
• MWC’s prediction of concomitant changes in tertiary structure in protomers with known symmetry of tetramer confirmed by more refined X-ray structures.
• Perutz provides mechanism of allosteric transitions• Szabo and Karplus show quantitative agreement of
MWC/perutz model with equilibrium data (Eg Lc4 constant after all salt bridges broken).
• Equilibrium oxygen binding to Hb trapped in T-state crystal non-cooperative (Eaton lab).
• CO rebinding following photolysis of HbCO (R-state) much faster than CO binding to Hb (T-state) – Gibson.








