

Arbitrary primed PCR ¢ngerprinting and serotyping of clinical
Pseudomonas aeruginosa strains
Javier Hernaèndez
a;
*, Mar|èa A. Ferruès
a, Manuel Hernaèndez
a, Robert J. Owen
b
aDepartamento de Biotecnolog|èa, Universidad Politeècnica, Camino de Vera 14, 46022 Valencia, Spain
bNational Collection of Type Cultures, Central Public Health Laboratory, 61 Colindale Avenue, London NW9 5HT, UK
Received 16 August 1996; revised 30 October 1996; accepted 31 October 1996
Abstract
Arbitrary primed PCR (AP-PCR) analysis was compared with serotyping as a means of high-resolution typing of
Pseudomonas aeruginosa. Seventy-four isolates from 3 different hospitals and 18 reference strains were studied. Serotyping
provided good index of discrimination, although eleven isolates could not be serotyped. Genomic DNA was amplified with a
single 10 nucleotide primer (sequence 5P-AGG GGT CTT G-3P). The strains were genetically diverse and 61 different AP-PCR
profiles of 2^7 bands between 0.3 and 2.4 kb were obtained. AP-PCR profiles were not consistently associated with serotypes,
but they clearly subtyped strains of the same serotype. Numerical analysis of AP-PCR patterns defined 7 groups at the 55%
similarity level, and identified predominant strains in each hospital. The results show that AP-PCR analysis provides a simple
and practical approach to typing P. aeruginosa that is more discriminatory than traditional serotyping scheme. We suggest that
maximum discrimination can be achieved by a combination of both methods.
Keywords: Pseudomonas aeruginosa ; DNA; Polymerase chain reaction; AP-PCR; Serotyping; Fingerprinting
1. Introduction
Pseudomonas aeruginosa is a Gram-negative bac-
terium widely distributed in nature and causing op-
portunistic infections in humans. The organism is an
important nosocomial pathogen of hospitalized pa-
tients, frequently associated with infections, such as
meningitis, septicemia, ocular and burn wound infec-
tions, otitis, urinary tract infections and cystic ¢bro-
sis (CF). Colonized patients can be an important
reservoir, and the organism is maintained in the hos-
pital environment by person-to-person transmission
and by its saprophytic survival and growth in a vari-
ety of hospital environments [1].
P. aeruginosa strains are identi¢ed on the basis of
phenotypic characteristics, but the nosocomial epide-
miology was hampered until recently by the impreci-
sion of most typing methods available. The most
commonly used epidemiological technique is serotyp-
ing, based on an agglutination reaction between bac-
terial lipopolysaccharide (LPS) and type-speci¢c rab-
bit antisera [2]. In 1983, an International Antigenic
Typing System (IATS) was developed [3], establish-
ing the existence of at least 17 major heat-stable so-
matic antigens, and 3 new antigens have been re-
cently added to the existing groups [4]. This
serotyping scheme characterizes most strains of
0928-8244 / 97 / $17.00 Copyright ß 1997 Federation of European Microbiological Societies. Published by Elsevier Science B.V.
PII S 0 9 2 8 - 8 2 4 4 ( 9 6 ) 0 0 1 0 3 - 4
FEMSIM 713 9-4-97
* Corresponding author. Tel. : +34 (6) 387 7423;
fax: +34 (6) 387 7429; e-mail: [email protected]
FEMS Immunology and Medical Microbiology 17 (1997) 37^47
P. aeruginosa possessing smooth LPS, although
shows poor discrimination against rough LPS iso-
lates which are common in patients with cystic ¢bro-
sis [5,6]. Other conventional available systems based
on phenotypic markers, such as biotyping, pyocin-
typing, phage-typing, or antimicrobial susceptibility
may not give satisfactory answers to basic epidemio-
logical questions as they rely on the analysis of char-
acteristics that are not stably expressed [7,8].
In contrast, high-discriminative DNA-based meth-
ods, have been recently used to ¢ngerprint P. aeru-
ginosa isolates, particularly, analysis of DNA restric-
tion fragment length polymorphism of rRNA genes
(ribotyping) [9^14], restriction endonuclease analysis
(REA) [15], and macrorestriction of digested DNA
by pulsed-¢eld gel electrophoresis (PFGE) [16^18].
However, the use of those methods in the clinical
microbiology laboratory is restricted to specialized
centres because of the technical complexity and their
labour-intensive nature [19].
An alternative and simpler typing technique based
on PCR with a single short arbitrary primer (AP-
PCR) has been used successfully for the typing of
various medically important microorganisms [20] in-
cluding P. aeruginosa [17,21,22]. In AP-PCR analy-
sis, the genomic DNA is ampli¢ed using single oli-
gonucleotides of arbitrarily chosen sequences and
non-stringent annealing temperatures, resulting in a
series of PCR ampli¢cation products with di¡erent
sizes that provide a characteristic electrophoretic ¢n-
gerprint. Although some factors need to be standard-
ized to achieve high reproducibility, it is one of the
easiest typing methods, because the template DNA
need not be of high molecular weight, double-
stranded, or highly puri¢ed, and only ng quantities
of DNA are required [23,24].
A comparison of phenotypic and genotypic meth-
ods for typing clinical strains of P. aeruginosa has
been done by The International Pseudomonas aeru-
ginosa Typing Study Group [25], but none PCR-
based typing technique was investigated in that
study.
We report here the use of AP-PCR ¢ngerprinting
and serotyping to study reference strains of the dif-
ferent P. aeruginosa serotypes, and clinical isolates
from hospitalized patients in Spain. The aim was
to determine the genetic diversity among such bac-
teria in order to detect DNA polymorphisms which
can indicate if there were any common strain types
associated with P. aeruginosa infections in 3 hospi-
tals in the Valencia area, and to evaluate the discri-
minatory power of both typing methods for future
epidemiological studies.
2. Materials and methods
2.1. Bacterial strains and culture conditions
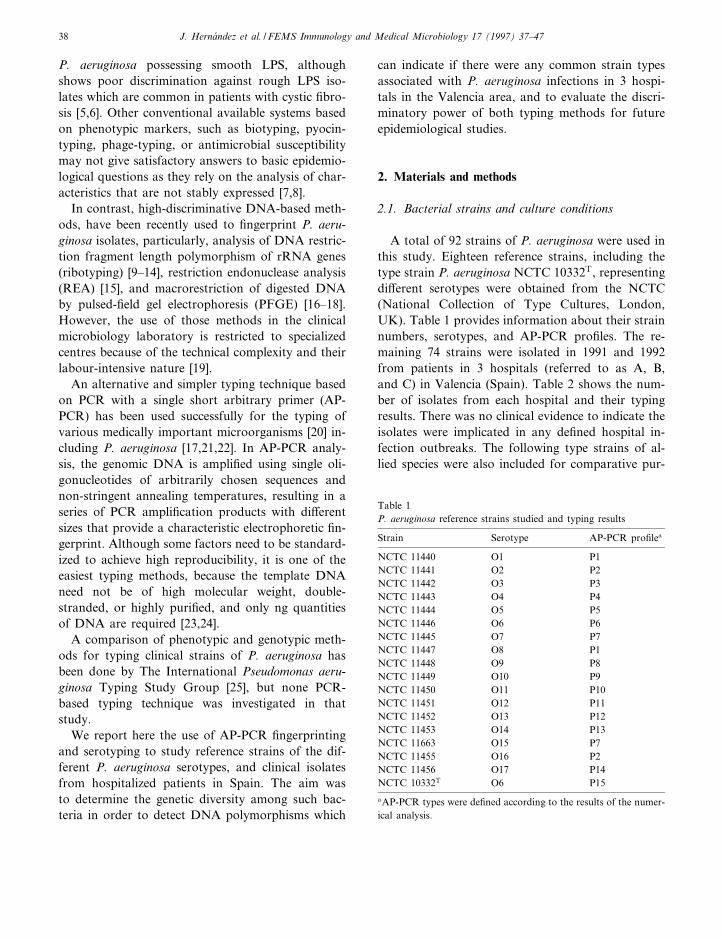
A total of 92 strains of P. aeruginosa were used in
this study. Eighteen reference strains, including the
type strain P. aeruginosa NCTC 10332T, representing
di¡erent serotypes were obtained from the NCTC
(National Collection of Type Cultures, London,
UK). Table 1 provides information about their strain
numbers, serotypes, and AP-PCR pro¢les. The re-
maining 74 strains were isolated in 1991 and 1992
from patients in 3 hospitals (referred to as A, B,
and C) in Valencia (Spain). Table 2 shows the num-
ber of isolates from each hospital and their typing
results. There was no clinical evidence to indicate the
isolates were implicated in any de¢ned hospital in-
fection outbreaks. The following type strains of al-
lied species were also included for comparative pur-
FEMSIM 713 9-4-97
Table 1
P. aeruginosa reference strains studied and typing results
Strain Serotype AP-PCR pro¢lea
NCTC 11440 O1 P1
NCTC 11441 O2 P2
NCTC 11442 O3 P3
NCTC 11443 O4 P4
NCTC 11444 O5 P5
NCTC 11446 O6 P6
NCTC 11445 O7 P7
NCTC 11447 O8 P1
NCTC 11448 O9 P8
NCTC 11449 O10 P9
NCTC 11450 O11 P10
NCTC 11451 O12 P11
NCTC 11452 O13 P12
NCTC 11453 O14 P13
NCTC 11663 O15 P7
NCTC 11455 O16 P2
NCTC 11456 O17 P14
NCTC 10332T
O6 P15
aAP-PCR types were de¢ned according to the results of the numer-
ical analysis.
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^4738

poses: P. £uorescens NCTC 10038
T, P. putida
NCTC 10936
T, and Burkholderia (Pseudomonas) ce-
pacia NCTC 10661
T.
All bacteria were cultivated at 37³C for 24 h on
Pseudomonas agar base (Oxoid CM559), with Pseu-
domonas C-N supplement (Oxoid SR102E) con-
taining cetrimide (200 mg/l) and sodium nalidixate
(15 mg/l). Strains were preserved at 370³C on glass
beads on nutrient broth No. 2 (Oxoid CM67) con-
taining 10% (v/v) glycerol.
2.2. Conventional bacteriological tests
The identity of all clinical isolates were con¢rmed
previously in a range of standard conventional bac-
teriological tests: gram stain, growth at 42³C, oxi-
dase production, DNA hydrolysis, pigment (pyocya-
nin and pyoverdin) production, haemolytic activity,
and the glucose oxidation-fermentation (O-F) test.
The isolates were identi¢ed by the API 20NE system
(bioMeérieux).
2.3. Serotyping
All strains were serotyped according to the scheme
proposed by the International Antigenic Typing Sys-
tem [3]. Qualitative slide agglutination of each strain
was done with autoclaved cells (120³C for 30 min)
and commercial antisera (Sano¢ Diagnostics Pasteur,
Madrid, Spain). All agglutinations were repeated 3
times to assess reproducibility.
Strains showing agglutination with standard saline
were coded as autoagglutinable. Strains reacting with
s 1 antiserum were referred as polyagglutinable.
Non-typeable strains were those which did not react
with any antiserum.
FEMSIM 713 9-4-97
Fig. 1. Arbitrarily primed polymerase chain reaction (AP-PCR) patterns of Pseudomonas aeruginosa strains representing all serotypes of
the scheme proposed by IATS. Genomic DNA was ampli¢ed using a 10-mer primer with sequence 5P-AGGGGTCTTG-3P and electro-
phoresed on a 1.2% agarose gel. Lanes 1^17 contain reference strains of serotypes O1-O17 (see Table 1 for NCTC strain numbers). Sizes
(in kb) of characteristic fragments are indicated on the right.
Table 2
Clinical strains studied and typing results
Source
aStrains (n) O serotypes
bAP-PCR pro¢les
c
Hospital A 13 1, 6^9, 11, PA 16^26
Hospital B 37 1^8, 10^12, NA, PA 1, 7^9, 19, 26^49
Hospital C 24 1, 2, 5^7, 10^13, 16, AA, PA, NA 1, 7, 15, 27, 28, 33, 49^61
aA, Hospital La Fe; B, Hospital General Universitario; C, Hospital Cl|ènico Universitario (Valencia, Spain).
bAA, autoagglutinable; NA, non-agglutinable; PA, polyagglutinable.
cAP-PCR types were de¢ned according to the results of the numerical analysis.
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^47 39

2.4. AP-PCR analysis
Bacterial cells were cultivated at 37³C for 24 h on
plate count agar (Oxoid) and suspended in 200 Wl of
ultrapure water. Each suspension was boiled for 10
min and centrifuged for 5 min. The concentration of
DNA in the supernatant was adjusted to 1.5 Wg/ml
(A260 =0.03). DNA ampli¢cation was performed
using 20 di¡erent primers (OPA 1^20, kit A; Operon
Technologies, Alameda, CA, USA) according to the
FEMSIM 713 9-4-97
Fig. 2. AP-PCR patterns of representative Pseudomonas aeruginosa strains from infected patients. Lanes 1^5 contain strains from hospital
A; lanes 6^11 contain strains from hospital B; lanes 12^17 contain strains from hospital C. Sizes (in kb) of characteristic fragments are
indicated on the right.
Table 3
Relation between serotypes and the origin of strains
Strains (n) from:
Serotypea
NCTC Hospital A Hospital B Hospital C Total
O1 1 3 5 4 13
O2 1 1 2 4
O3 1 3 4
O4 1 1 2
O5 1 1 2 4
O6 2 1 2 3 8
O7 1 1 3 1 6
O8 1 2 1 4
O9 1 1 2
O10 1 3 2 6
O11 1 4 8 3 16
O12 1 2 1 4
O13 1 1 2
O14 1 1
O15 1 1
O16 1 2 3
O17 1 1
AA 1 1
NA 2 1 3
PA 1 5 1 7
aAA, autoagglutinable; NA, non-agglutinable; PA, polyagglutinable.
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^4740

manufacturer's instructions.
A reaction volume of 100 Wl contained 2.5 U of
Taq polymerase (DynaZyme thermostable DNA
polymerase; Finnzymes, Finland), 10 mM Tris-
HCl, pH 8.8, at 25³C, 50 mM KCl, 1.5 mM MgCl2,
0.1% Triton-X100, 200 WM of each deoxynucleotide
triphosphate, 0.8 WM of the primer, 10 Wl of the cell
suspension (15 Wg of DNA) and sterile distilled
water. A negative control in which DNA was re-
placed with sterile distilled water was also included.
The solutions were overlaid with 100 Wl of para¤n
oil and cycled through the following temperature
pro¢le: an initial denaturation step at 95³C for
5 min, 45 cycles of 95³C for 1 min (denaturation),
37³C for 3 min (annealing) and 72³C for 7 min
(DNA chain extension), and a ¢nal elongation step
at 72³C for 10 min. Incubation was performed in a
thermocycler (PHC-3 Thermal Cycler, Techne Cor-
poration, Cambridge, UK). Standard procedures
were adopted to eliminate contamination [26]. Dupli-
cated AP-PCR analysis was performed for each
strain to assess reproducibility. The ampli¢ed DNA
products were electrophoresed on 1.2% (w/v) agarose
gels (ultrapure electrophoresis grade, Gibco-BRL) at
90 V for 3 h and stained in ethidium bromide. A 123
bp DNA ladder (Gibco-BRL) was used as a molec-
ular weight marker. Two lanes (outside left and
right) containing the markers were included on
each gel to correct for within and between gel dis-
tortions.
2.5. Fragment size estimation
Molecular size of fragments was estimated by
comparison with the molecular weight markers using
the computer program HOWBIG [27], based on the
reciprocal correlation of size to migration distance of
DNA. To perform the calculations, the program
chose the 3 marker bands migrating closest to the
test band, so reducing the error to 9 0.5%.
2.6. Computation of strain similarities
To compare AP-PCR patterns from di¡erent gels,
the bands were coded according to size to minimize
errors when determining similarities by computer-
assisted methods of analysis. The ampli¢cation pat-
terns were screened for bands within 16 di¡erent size
ranges from 0.3 to 2.4 kb and positive (presence) or
negative (absence) results were recorded. Computed
similarities among strains were estimated by means
of the Dice coe¤cient (negative matches excluded)
and clustering of strains was based on the un-
weighted average pair group method (UPGMA) to
facilitate the plotting of a dendrogram [28]. All com-
putations were performed using the NTSYS-PC pro-
gram [29].
2.7. Discrimination power
The discriminatory power of AP-PCR analysis
was calculated by application of Simpson numerical
index of diversity [30].
3. Results
3.1. Typeability and reproducibility
To assess primer e¤ciency and discrimination, an
initial screening with the 20 random primers was
FEMSIM 713 9-4-97
Table 4
Numbers of P. aeruginosa strains of di¡erent serotype in each
group based on numerical analysis of AP-PCR pro¢les
AP-PCR group
Serotype I II III IV V VI VII
O1 5 1 2 3 1 1
O2 3 1
O3 4
O4 1 1
O5 2 2
O6 5 3
O7 2 1 1 1 1
O8 2 1 1
O9 1 1
O10 6
O11 9 2 2 1 2
O12 2 1 1
O13 2
O14 1
O15 1
O16 1 2
O17 1
AA 1
NA 1 2
PA 5 2
Total 53 4 19 8 1 4 3
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^47 41

FEMSIM 713 9-4-97
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^4742

performed to determine di¡erences between 3 se-
lected P. aeruginosa strains. As primer OPA-5 (se-
quence 5P-AGG GGT CTT G-3P) gave a clear and
reproducible AP-PCR ¢ngerprint, it was selected for
use in comparative studies. All the 18 reference
strains and the 74 clinical isolates subjected to AP-
PCR analysis yielded ampli¢cation products (type-
ability of 100%). No product was detected in any
negative (distilled water) control. To check the re-
producibility of the method, DNA of each strain
was tested on at least 2 di¡erent subcultures. Iden-
tical AP-PCR banding patterns (relative position and
intensity of bands) were observed. Although similar
results were obtained for most repeated subcultures
of the same strain, some minor di¡erences were ob-
served in the relative amounts of certain fragments,
and variability in the presence of faint bands. There-
fore, only the major ampli¢cation fragments that
appeared in both analysis were used in the numerical
analysis. The reproducibility of the method was also
demonstrated by the fact that identical patterns were
obtained with epidemiologically related strains.
3.2. Distribution of serotypes
Table 3 lists the serotypes of all reference and
clinical strains and the distribution according their
origin. Eleven strains (12%) could not be assigned
to any international serotype and were coded as
autoagglutinable (1 strain), non-agglutinable (3
strains), or polyagglutinable (7 strains). The test re-
sults of the non-serotypeable strains were checked
for growth at 41³C, oxidation of gluconate, and ni-
trate reduction, to con¢rm the identity as typical P.
aeruginosa. The discrimination index of serotyping
analysis referred to clinical isolates was 0.886. Over-
all, the predominant serotypes were O11 and O1,
representing 24 and 19%, respectively, of serotype-
able strains. Three serotypes (O14, O15, and O17)
were not found among the strains isolated from in-
fected patients. The type strain of P. aeruginosa
(NCTC 10332) was serotype O6. No association
was observed between strain serotype and hospital.
3.3. AP-PCR patterns of clinical isolates and
reference strains
The 92 di¡erent strains of P. aeruginosa listed in
Tables 1 and 2 were subjected to AP-PCR analysis
using primer OPA-5. Examples of AP-PCR patterns
of reference strains from all 17 serotypes of the IATS
scheme, and of representative strains from infected
patients are shown diagrammatically in Figs. 1 and
2, respectively. The number of ampli¢ed DNA prod-
ucts detected by electrophoresis varied between 2
and 7, with most having relatively uniform intensi-
ties. The AP-PCR patterns of the reference strains of
3 allied species (P. £uorescens NCTC 10038T, P. pu-
tida NCTC 10936T, and B. cepacia NCTC 10661
T)
were di¡erent from each other and from all the
strains of P. aeruginosa (data not shown).
3.4. Analysis of AP-PCR patterns
Overall 16 di¡erent DNA fragments were detected
among the patterns from P. aeruginosa. The esti-
mated sizes ranged from 0.3 to 2.4 kb. The 1.48 kb
fragment was found in AP-PCR patterns of all iso-
lates. Two additional fragments of 0.96 and 1.71 kb
were found in 89 and 65% of strains, respectively.
The ampli¢cation products of 1.48 and 1.71 kb
were not detected in patterns of the type strains of
3 allied Pseudomonas species, but the 0.96 kb frag-
ment could be observed in the type strain of B. ce-
pacia.
Based on the AP-PCR patterns, a total of 61 dif-
ferent pro¢les were obtained. The pro¢le assigned to
each strain is listed in Tables 1 and 2. Most pro¢les
(45/61) were unique, but 16 pro¢les were common
for 2 or more strains that were not necessarily
from the same hospital. The 61 di¡erent AP-PCR
pro¢les were compared by numerical methods and
the resultant dendrogram (Fig. 3) based on the per-
centage similarity between isolates, shows a high de-
gree of genetic diversity. The numerical index of dis-
crimination was high (D=0.984). Seven major
groups were de¢ned at the 55% similarity level, and
FEMSIM 713 9-4-97
Fig. 3. Dendrogram of the numerical analysis based on the AP-PCR patterns of strains. The numbers on the horizontal axis indicate the
percentage similarities as determined by the Dice coe¤cient. The vertical axis shows the main groups de¢ned at the 55% similarity level.
P, AP-PCR pro¢le.
6
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^47 43

all strains of P. aeruginosa were linked at the 40%
homology level.
Group I, the overall predominant cluster, com-
prised 53 (58%) di¡erent strains representing 34
(56%) diverse AP-PCR pro¢les, linked at the 60%
similarity level. No strain isolated at Valencia hospi-
tal A fell in this group. The more frequent serotypes
represented were O11 (9 strains) and O10 (6 strains).
Only serotype O4 was not included.
Group II, de¢ned at the 70% similarity level, was
heterogeneous and contained only 4 (5%) clinical
strains isolated in the 3 hospitals. Serotypes O1, O7
and O11 were represented among the strains.
Group III was de¢ned at the 55% similarity level
and comprised 19 (21%) strains from all sources (ref-
erence and clinical isolates), with 13 (21%) di¡erent
AP-PCR pro¢les. The type strain of P. aeruginosa
(NCTC 10332) was included in this cluster. No single
serotype predominated among the isolates.
Groups IV (8 strains, 4 AP-PCR pro¢les), V
(1 strain), VI (4 strains, 3 AP-PCR pro¢les), and
VII (3 strains, 2 AP-PCR pro¢les), were relatively
homogeneous, comprising 1 reference strain and 15
di¡erent isolates from patients at hospitals A and B,
but not hospital C.
The inclusion in the study of reference strains re-
presenting all international serotypes revealed no re-
lationship between AP-PCR pro¢les and serotypes.
Some strains assigned to the same AP-PCR pro¢le
had di¡erent serogroups, and conversely, some
strains of the same serogroup had di¡erent AP-
PCR pro¢les.
3.5. Distribution of strain types
The distribution of the di¡erent serotypes and AP-
PCR types within and between each hospital are
shown in Table 3. The number of strains in each
group belonging to di¡erent serogroups is shown in
Table 4. The results show that AP-PCR ¢ngerprint-
ing provides signi¢cantly improved discrimination
within several serotypes; for example, see serotypes
O1, O7 and O11.
4. Discussion
Typing to evaluate the probability of cross-trans-
mission is commonly advised in epidemiological in-
vestigations, and the use of very discriminatory typ-
ing methods is recommended for di¡erentiating
epidemiologically unrelated isolates. It is widely ac-
cepted that traditional phenotypic methods of typing
isolates of P. aeruginosa [5,7,8] do not permit accu-
rate investigation of nosocomial infections because
phenotypic markers are relatively unstable. In con-
trast, molecular subtyping methods o¡er high type-
ability and reproducibility, can de¢ne clonal relation-
ships between isolates, and have contributed
signi¢cantly to a better understanding of the epide-
miology of P. aeruginosa infections [9^18,21,22,25,
31].
In our study, a high degree of technical reprodu-
cibility was achieved by both typing methods, but
AP-PCR ¢ngerprinting provided higher typeability
(100%) and discrimination power (0.984), and conse-
quently, was more useful for epidemiological inves-
tigations. AP-PCR-generated DNA fragments were
relatively easy to interpret because of the low num-
ber of bands. Also, visual comparisons of gels were
facilitated by the presence of common bands that
could be used as internal markers to allow normal-
ization of slight gel-to-gel variations. Our AP-PCR
¢ngerprinting results show considerable genomic di-
versity among the P. aeruginosa isolated from hospi-
tals in Valencia (Spain), and most strains have dis-
tinct genotypes.
The high degree of discrimination associated with
DNA-based typing methods is dependent upon an
evolutionary genetic divergence arising from point
mutations, insertions or deletions of chromosomal
DNA, and if such random genetic events occur fre-
quently in a strain, the applicability of these methods
would be decreased [32,33]. These features are espe-
cially relevant for typing P. aeruginosa isolates as
genetic rearrangements associated with the horizon-
tal transfer of genetic elements from habitats have
been demonstrated [34]. In fact, investigators have
recently proposed several criteria for interpreting
chromosomal DNA restriction patterns generated
by molecular methods [35].
Ribotyping has provided invaluable information
during the investigation of many nosocomial infec-
tions, but its apparently low sensitivity for the study
of P. aeruginosa strains might be related to an insuf-
¢cient search for the most appropriate restriction
FEMSIM 713 9-4-97
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^4744

endonuclease [10]. Generally, the use of single en-
zymes provide less discrimination to that reached
by other genotyping methods [9,10,14,25]. Addition-
ally, when the circumstances and results indicate the
need for further characterization, as in the investiga-
tion of strains from a suspected outbreaks, extra
discrimination must be obtained by combining data
from several endonucleases [10,12]. Thus, Blanc et al.
[11] critically assessed the typeability, reproducibility
and discriminatory power of ribotyping applied to
epidemiologically unrelated P. aeruginosa isolates in
Switzerland and concluded the system compared well
with serotyping. When they used single endonu-
cleases, a discriminatory index of W0.7 was re-
ported, which could be increased to 0.958 with com-
bined data of 4 enzymes.
In a study of 102 P. aeruginosa isolates infecting
23 CF patients in France [12], a comparison was
made of results from esterase electrophoresis and
combined ribopatterns for 3 enzymes. All strains
were typeable with this molecular approach, whereas
only 17% of the isolates could be serotyped. A total
of 16 ribotypes and 25 zymotypes were di¡eren-
tiated, but it was demonstrated that by combining
the 2 molecular methods, precise characterization
of P. aeruginosa could be achieved. An index of dis-
crimination was not calculated.
Macrorestriction analysis of digested DNA by
pulse-¢eld gel electrophoresis, as a molecular ap-
proach to studying P. aeruginosa epidemiology, was
found to be more discriminatory than ribotyping
[16], but it was equally e¡ective when compared to
AP-PCR ¢ngerprinting [17]. In a recent work, Ren-
ders et al. [18], demonstrated the enhanced resolution
of AP-PCR and PFGE when compared to serotyp-
ing and pyocin-typing. Both DNA typing techniques
generated concordant results, although di¡erences in
resolution (15%) are apparent.
Other investigators used AP-PCR analysis to show
genomic diversity among unrelated strains in hospi-
tal environments, and that it could be used to di¡er-
entiate such strains from other isolates in the same
hospital during an outbreak [21,22]. Pitt et al., using
ribotyping for the characterization of multiresistant
O12 P. aeruginosa strains, provided evidence of a
common strain in Europe [31].
In the present work, there were no common strain
AP-PCR types detected in the 3 hospitals in the Va-
lencia area, whereas the majority of types were only
associated with 1 hospital. For instance, just 2 AP-
PCR pro¢les (P19 and P26) were found in hospitals
A and B, whereas 6 di¡erent pro¢les (P1, P7, P27,
P28, P33, and P49) were associated with hospitals B
and C. Because of the relatively small sample size in
each hospital, it is di¤cult to draw any general con-
clusions about the signi¢cance of the strain type dis-
tributions among the hospitals. In the main, they
appear to be associated with sporadic randomly oc-
curring infections. However, it is uncertain from
these data if patient to patient spread of P. aerugi-
nosa with the same type occurred because such strain
types were found independently in 2 hospitals, and
no clinical outbreaks were reported. Nevertheless,
the distinctive feature of the AP-PCR-typing results
shown in Fig. 3 is that no strains from patients at
hospital A were in group I, even though it was the
predominant group with s 50% of strains. In addi-
tion, groups IV^VII were also relatively homoge-
neous, encompassing only strains isolated from pa-
tients at hospitals A and B.
Several parameters can alter AP-PCR pro¢les and
must be previously standardized in order to obtain
reproducible results. Selection of an appropriate
primer, the annealing temperature, the primer-tem-
plate concentration ratio, the magnesium concentra-
tion, and the type of thermocycler have been
reported to produce variations in the pro¢les
[36,37], and are of great importance for maximizing
the discriminatory power of AP-PCR analysis. For
the correct interpretation and classi¢cation of di¡er-
ent but often similar pro¢les, a set of major bands
must be previously de¢ned, and strains with similar
pro¢les have to be analyzed on the same gel. It is
possible to increase the discriminatory power of AP-
PCR analysis by using 2 primers together [38,39]
although we achieved good discrimination with just
1 primer.
This study shows that AP-PCR analysis is a mol-
ecular method that provides a useful and practical
means of typing P. aeruginosa strains. AP-PCR ana-
lysis is clearly more discriminatory and o¡ers higher
typeability than serotyping. It also has the advan-
tages of rapidity, technical simplicity, and relative
ease of ¢ngerprint interpretation compared with
other molecular methods, such as ribotyping and
PFGE. In addition, whole bacterial cells can be
FEMSIM 713 9-4-97
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^47 45

used without the requirement for DNA extraction,
and there is no need for multiple primer pairs, nested
PCR or DNA sequencing to di¡erentiate between P.
aeruginosa strains. Consequently, AP-PCR is an ex-
cellent ¢rst-line typing procedure, especially where
large numbers of strains need molecular typing.
Further investigations on outbreaks are needed to
validate AP-PCR in clinical investigations, even so
we would recommend its use in conjunction with
serotyping for improving discrimination in the epide-
miological surveillance of P. aeruginosa.
Acknowledgments
We are grateful to the following for providing
strains: Dr. M. Gobernado (Ciudad Sanitaria La
Fe); Dr. R. Llucian (Hospital General Universita-
rio); and Dr. J. Garc|èa de Lomas (Hospital Cl|ènico
Universitario, Valencia, Spain).
References
[1] Gilardi, G.L. (1991) Pseudomonas and related genera. In:
Manual of Clinical Microbiology (Balows, A., Hausler,
W.J., Herrmann, K.L., Isenberg, H.D. and Shadomy, H.J.,
Eds.), pp. 429-441. American Society for Microbiology, Wash-
ington, DC, USA.
[2] Fisher, M.W., Devlin, H.B. and Gnabasik, F.J. (1969) New
immunotype schema for Pseudomonas aeruginosa based on
protective antigens. J. Bacteriol. 98, 835^836.
[3] Liu, P.V., Matsumoto, H., Kusama, H. and Bergan, T. (1983)
Survey of heat-stable, major somatic antigens of Pseudomonas
aeruginosa. Int. J. Syst. Bacteriol. 33, 256^264.
[4] Liu, P.V. and Wang, S. (1990) Three new major somatic anti-
gens of Pseudomonas aeruginosa. J. Clin. Microbiol. 28, 922^
925.
[5] Hancock, R.E.W., Mutharia, L.M., Chan, L., Darveau, R.P.,
Speert, D.P. and Pier, G.B. (1983) Pseudomonas aeruginosa
isolates from patients with cystic ¢brosis: a class of serum-
sensitive non-typable strains de¢cient in lipopolysaccharide O
side-chains. Infect. Immun. 42, 170^177.
[6] Speert, D.P. and Campbell, M.E. (1987) Hospital epidemiol-
ogy of Pseudomonas aeruginosa from patients with cystic ¢-
brosis. J. Hosp. Infect. 9, 11^21.
[7] Pitt, T.L. (1988) Epidemiological typing of Pseudomonas
aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 7, 238^247.
[8] Schable, B., Olsen, D.R. and Smith, P.B. (1986) Improved
computer-generated system for pyocin typing of Pseudomonas
aeruginosa. J. Clin. Microbiol. 24, 1017^1022.
[9] Bennekov, T., Colding, H., Ojeniyi, B., Bentzon, M.W. and
Hoiby, N. (1996) Comparison of ribotyping and genome ¢n-
gerprinting of Pseudomonas aeruginosa isolates from cystic
¢brosis patients. J. Clin. Microbiol. 34, 202^204.
[10] Bingen, E.H., Denamur, E. and Elion, J. (1994) Use of ribo-
typing in epidemiological surveillance of nosocomial out-
breaks. Clin. Microbiol. Rev. 7, 311^327.
[11] Blanc, D.S., Siegrist, H.H., Sahli, R. and Francioli, P. (1993)
Ribotyping of Pseudomonas aeruginosa : discriminatory power
and usefulness as a tool for epidemiological studies. J. Clin.
Microbiol. 31, 71^77.
[12] Denamur, E., Picard, B., Goullet, P., Bingen, E., Lambert, N.
and Elion, J. (1991) Complexity of Pseudomonas aeruginosa
infection in cystic ¢brosis: combined results from esterase
electrophoresis and rDNA restriction fragment length poly-
morphism analysis. Epidemiol. Infect. 106, 531^539.
[13] Grattard, F., Pozzetto, B., Ros, A. and Gaudin, O.G. (1994)
Di¡erentiation of Pseudomonas aeruginosa strains by ribotyp-
ing: high discriminatory power by using a single restriction
endonuclease. J. Med. Microbiol. 40, 275^281.
[14] Grundmann, H., Schneider, C., Hartung, D., Daschner, F.D.
and Pitt, T.L. (1995) Discriminatory power of three DNA-
based typing techniques for Pseudomonas aeruginosa. J. Clin.
Microbiol. 33, 528^534.
[15] Maher, W.E., Kobe, M. and Fass, R.J. (1993) Restriction
endonuclease analysis of clinical Pseudomonas aeruginosa
strains: useful epidemiologic data from a simple and rapid
method. J. Clin. Microbiol. 31, 1426^1429.
[16] Hla, S.W., Hui, K.P., Tan, W.C. and Ho, B. (1996) Genome
macrorestriction analysis of sequential Pseudomonas aerugino-
sa isolates from bronchiectasis patients without cystic ¢brosis.
J. Clin. Microbiol. 34, 575^578.
[17] Kersulyte, D., Struelens, M.J., Deplano, A. and Berg, D.E.
(1995) Comparison of arbitrarily primed PCR and macrores-
triction (pulse-¢eld gel electrophoresis) typing of Pseudomonas
aeruginosa strains from cystic ¢brosis patients. J. Clin. Micro-
biol. 33, 2216^2219.
[18] Renders, N., van Belkum, A., Barth, A., Goessens, W., Mou-
ton, J. and Verbrugh, H. (1996) Typing of Pseudomonas
aeruginosa strains from patients with cystic ¢brosis: phenotyp-
ing versus genotyping. Clin. Microbiol. Infect. 1, 261^265.
[19] Owen, R.J. and Hernandez, J. (1993) Ribotyping and arbi-
trary-primer PCR ¢ngerprinting of campylobacters. In: New
Techniques in Food and Beverage Microbiology (Kroll, R.G.,
Gilmour, A. and Sussman, M., Eds.), pp. 265^285. SAB Tech.
Ser. 31. Blackwell, Oxford, UK.
[20] van Belkum, A. (1994) DNA ¢ngerprinting of medically im-
portant microorganisms by use of PCR. Clin. Microbiol. Rev.
7, 174^184.
[21] Elaichouni, A., Verschraegen, G., Claeys, G.,
Devleeschouwer, M., Godard, C. and Vaneechoutte, M.
(1994) Pseudomonas aeruginosa serotype O12 outbreak studied
by arbitrary primer PCR. J. Clin. Microbiol. 32, 666^671.
[22] Mahenthiralingam, E., Campbell, M.E., Foster, J., Lam, J.S.
and Speert, D.P. (1996) Random ampli¢ed polymorphic DNA
typing of Pseudomonas aeruginosa isolates recovered from pa-
tients with cystic ¢brosis. J. Clin. Microbiol. 34, 1129^1135.
[23] Welsh, J. and McClelland, M. (1990) Fingerprinting genomes
FEMSIM 713 9-4-97
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^4746

using PCR with arbitrary primers. Nucleic Acids Res. 18,
7213^7218.
[24] Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A.
and Tingey, S.V. (1990) DNA polymorphisms ampli¢ed by
arbitrary primers are useful as genetic markers. Nucleic Acids
Res. 18, 6531^6535.
[25] International Pseudomonas aeruginosa Typing Study Group
(1994) A multicenter comparison of methods for typing strains
of Pseudomonas aeruginosa predominantly from patients with
cystic ¢brosis. J. Infect. Dis. 169, 134^142.
[26] Kwok, S. and Higuchi, R. (1989) Avoiding false positives with
PCR. Nature (London) 339, 237^238.
[27] Grabner, M. and Hofbauer, R. (1991) A computer program
for molecular weight determination of DNA fragments
(HOWBIG). CABIOS 7, 317^319.
[28] Sneath, P.H.A. and Sokal, R.R. (1973) Numerical Taxonomy.
W.H. Freeman, San Francisco, CA, USA.
[29] Rohlf, J. (1987) Numerical Taxonomy and Multivariate Ana-
lysis System for the IBM PC Microcomputer. pp. 1^36. Ap-
plied Biostatistics, Setauket, NY, USA.
[30] Hunter, P.R. and Gaston, M.A. (1988) Numerical index of the
discrimination ability of typing systems: an application of
Simpson's index of diversity. J. Clin. Microbiol. 26, 2465^
2466.
[31] Pitt, T.L., Livermore, D.M., Pitcher, D., Vatopoulos, A.C.
and Legakis, N.J. (1989) Multiresistant serotype O12 Pseudo-
monas aeruginosa : evidence for a common strain in Europe.
Epidemiol. Infect. 103, 565^576.
[32] Maslow, M.N., Mulligan, M.E. and Arbeit, R.D. (1993) Mo-
lecular epidemiology: application of contemporary techniques
to the typing of microorganisms. Clin. Infect. Dis. 17, 153^
164.
[33] Hall, L.M.C. (1994) Are point mutations or DNA rearrange-
ments responsible for the restriction fragment length poly-
morphisms that are used to type bacteria?. Microbiology
140, 197^204.
[34] Sokol, P.A., Luan, M., Storey, D.G. and Thirukkumaran, P.
(1994) Genetic rearrangement associated with in vivo mucoid
conversion of Pseudomonas aeruginosa PAO is due to inser-
tion elements. J. Bacteriol. 176, 553^562.
[35] Tenover, F.C., Arbeit, R.D., Goering, R.V., Mickelsen, P.A.,
Murray, B.E., Persing, D.H. and Swaminathan, B. (1995) In-
terpreting chromosomal DNA restriction patterns produced
by pulsed-¢eld gel electrophoresis: criteria for bacterial strain
typing. J. Clin. Microbiol. 33, 2233^2239.
[36] Ellsworth, D.L., Rittenhouse, K.D. and Honeycutt, R.L.
(1993) Artifactual variation in randomly ampli¢ed poly-
morphic DNA banding patterns. BioTechniques 14, 214^217.
[37] Penner, G.A., Bush, A., Wise, R., Kim, W., Domier, L.,
Kasha, K., Laroche, A., Scoles, G., Molnar, S.J. and Fedak,
G. (1993) Reproducibility of random ampli¢ed polymorphic
DNA (RAPD) analysis among laboratories. PCR Methods
Applications 2, 341^345.
[38] Micheli, M.R., Bova, R., Calissano, P. and D'Ambrosio, E.
(1993) Randomly ampli¢ed polymorphic DNA ¢ngerprinting
using combinations of oligonucleotide primers. BioTechniques
15, 388^390.
[39] Welsh, J. and McClelland, M. (1991) Genomic ¢ngerprinting
using arbitrarily primed PCR and a matrix of pairwise com-
binations of primers. Nucleic Acids Res. 19, 5275^5279.
FEMSIM 713 9-4-97
J. Hernaèndez et al. / FEMS Immunology and Medical Microbiology 17 (1997) 37^47 47







