

BACTERIAL INJURY AND RECOVERY*
F . F. BUSTA AND L. B. SMITH University of Minnesota
INTRODUCTION
Recognit ion of sublethally-damaged microorganisms is imperative f o r accurate in te rpre ta t ions of microbiological evaluations of meat and meat products. Treatments such as thermal processing, cooking, warming, cooling, ch i l l i ng , freezing-thawing, drying, adding salt or sugar or i r rad ia t ing ; or slmple exposure t o spec i f i c food components, s an i t i z ing *
agents or various preservatives may cause suble tha l damge i n c e l l s o r spores of various microorganisms. Sublethal injury induced by exposure t o processing treatments of ten is observed as a loss of one or w r e of t he a b i l i t i e s of t h e microorganisms t o function normally under conditions t h a t a r e sa t i s f ac to ry f o r c e l l s that have not been exposed t o the processing treatment. suscep t ib i l i t y t o ant imicrobial agents t h a t are used i n se l ec t ive media or i n cont ro l of microorganisms. Injury is i n part characterized and defined by t h e a b i l i t y of t h e microorganisms t o re turn t o i ts normal s t a t e during a resusc i ta t ion period in which t h e damaged components a r e repaired. Understanding suble tha l damage i n microbial c e l l s is absolutely e s s e n t i a l t o in te rpre ta t ion of laboratory data, t o t h e development of various meat processes, and t o the preserving of s t a r t e r cu l tures f o r fermented meats.
C e l l in jury may be observed as a n increased
His tor ica l ly , many invest igators ident i f ied ce r t a in media and c u l t u r a l conditions as superior f o r growth of ce r t a in microorganisms re t r ieved from various environments. An ove ra l l review and compre- hensive c i t a t i o n of s ign i f icant l i t e r a t u r e on the p r a c t i c a l implications of i n ured microorganisms i n food has been published recent ly by Busta (19763
MEAT PROCESSING TREATMENTS THAT INDUCE CELL INJURY
Elevated Temperatures. Processing procedures that involve elevated temperatures nay s t r e s s microbial c e l l s o r spores. temperatures as found In low temperature long time cooking and warming can damage vegetative c e l l s . concentration and dehydration processes also may apply such s t r e s s . Processlng p lan t cleaning procedures have t h e same po ten t i a l . Heat injury of t h e following species has been reported: Staphylococcus aureus, Streptococcus f aeca lis, Salmonella typhirmur ium, Escherichia c 011, Aerobacter aerogenes, Pseudomonas fluorescens, Vibrio marinus, Bacil lus s u b t i l i s , Clostridium botulinum, Candida u t i l i s , and Saccharowces
Relatively low
Similar temperatures associated w i t h
* Presented a t t h e 29th Annual Reciprocal Meat Conference of t he American Meat Science Association, 1976

cerevis iae (Stevenson and Richards, 1976) Thermal processing a t conventional and ult rahigh temperatures damages b a c t e r i a l spores from Bacillus s u b t i l i s , Clostridium perfringens, Clostridium sporogenes, and Clostridium botulinum.
Reduced Temperatures. Reduced temperatures above 0 C can induce injury i n S. aureus, Streptococcus l a c t i s , and E. perfrigens
Processes a t temperatures below 0 C f o r f reezing preservation can induce i n j u r y i n numerous bac ter ia such as Shigel la sonnei, S. faecal is , and Pseudomonas f luorescens . The extensive information on freeze - injury of bac te r ia has been w e l l reviewed by Ray and Speck (1973) and by Fennema, Powrie and f i r t h (1973 ) . *
Removal of moisture from meat i n the frozen s t a t e is a sophis t icated and gent le preservation technique Nevertheless, the many microorganisms that exhib i t damage from the r e su l t an t s t r e s s include S . anatum, E. - -7 c o l i Streptococcus thermophilus, and 2. aureus. prepared by freeze-drying could contain injured microorganisms, and cu l tures f o r food manufacture or f o r laboratory use preserved by t h i s procedure may contain damaged c e l l s . Unfortunately, t o da te the spec i f i c influences of the individual s t r e s ses of thawing, freezing, dehydration, and rehydration have not been wel l d i f f e ren t i a t ed .
Thus food s p e c i a l i t i e s
Moisture Reduction. Removal of part of most of the avai lable water occurs i n many meat processes and m y in jure microorganisms. Incidental dehydration during asp i ra t ion or during chance drying on equipment may a l s o generate a damaging e f f e c t . Reduction of water i n intermediate moisture foods through addi t ion of humectants damages b a c t e r i a l c e l l s (Hunt and Busta, 1974). This damage may be'caused by mechanisms similar t o those observed i n other types of dehydration or f reezing injury.
Nut r i t iona l Environment. Placement of c e l l s i n to a new environment may generate s t r e s ses t h a t r e s u l t i n injury. A growth environment of new or d i f f e ren t subs t ra te concentrations or osmotic s t rengths could produce s t r e s ses that resenible dehydration or rehydration. I n i t i a t i o n of growth i n a f r e sh complex medium m y produce c e l l s that respond t o se l ec t ive or minimal media i n a fashion that resembles injury. Place- ment In a spent medium may expose c e U s t o end products such as acids that promote damage. apparent m t of injury. Ekposure t o d i luents of v a r i m s types and especial ly d i s t i l l e d water may in jure c e l l s . It is reasonable t o suspect that c e l l s exposed t o s ta rva t ion conditions that a r e a p p r e n t l y l e t h a l m i g h t also display suble tha l damage if survivors were determined appropriately .
Incidental exposure t o a i r may influence t h e
I r r a d h t i o n . Use of W i r r ad ia t ion f o r san i t iza t ion , inc identa l i r r ad ia t ion of surfaces i n sunlight, or use of gamma i r r ad ia t ion f o r meat preservation have po ten t i a l f o r generating repairable suble tha l damage i n microorganisms associated w i t h foods. This may be bes t demonstrated in rad ia t ion- res i s tan t strains. Recently Annellis e t a l . (1976) indicated t h e presence of irradiation-danaged C - . botulinum spores i n irradiated.ham.

303
Sani t izers . Chemicals employed t o san i t i ze items in the food industry readi ly induce injury i n bac t e r i a if t he san i t i ze r s a r e used under conditions and a t concentrat ions that promote suble tha l damage and minimize complete inact ivat ion of t h e c e l l s (Scheusner e t al . , 1971)
Preservatives and Acidulants. End products of food fermentations and food addi t ives that a r e preservatives or acidulants frequently serve as inhib i tors of unwanted growth rather than as l e t h a l agents f o r unwanted microorganisms. Ekposure t o acids i n combination with other s t r e s ses damaged 2. aureus, whereas exposure t o alkali a l t e r ed spores of C,. perf r ingens .
Combinations of Treatments and Interact ions of S t resses . Some processing treatments such as freeze-drying are by de f in i t i on canbinations of severa l s t r e s s e s . Unfortunately, invest igat ions on c e l l injury of ten neglect the measuremnt of interact ions and synerg is t ic e f f ec t s among the various individual components of t he treatment. The presence of low levels of N a C l in t h e heating llledium reduces the amount of heat injury of E. c o l i measured as increased suscep t ib i l i t y t o N a C l i n t he p la t ing mezium, whereas sorbic acid enhanced thermal in jury i n C,. u t i l i s . in jury i n S,. aureus. Combinations of heat treatments and severa l humectants i n the heating or growth media resu l ted in various responses that indicated c e l l injury and the responses were dependent on t h e type of humectant and the species of bac te r i a .
Freezing i n combination with law pH enhanced t h e extent of
DEE13NSTRATION OF I X m Y
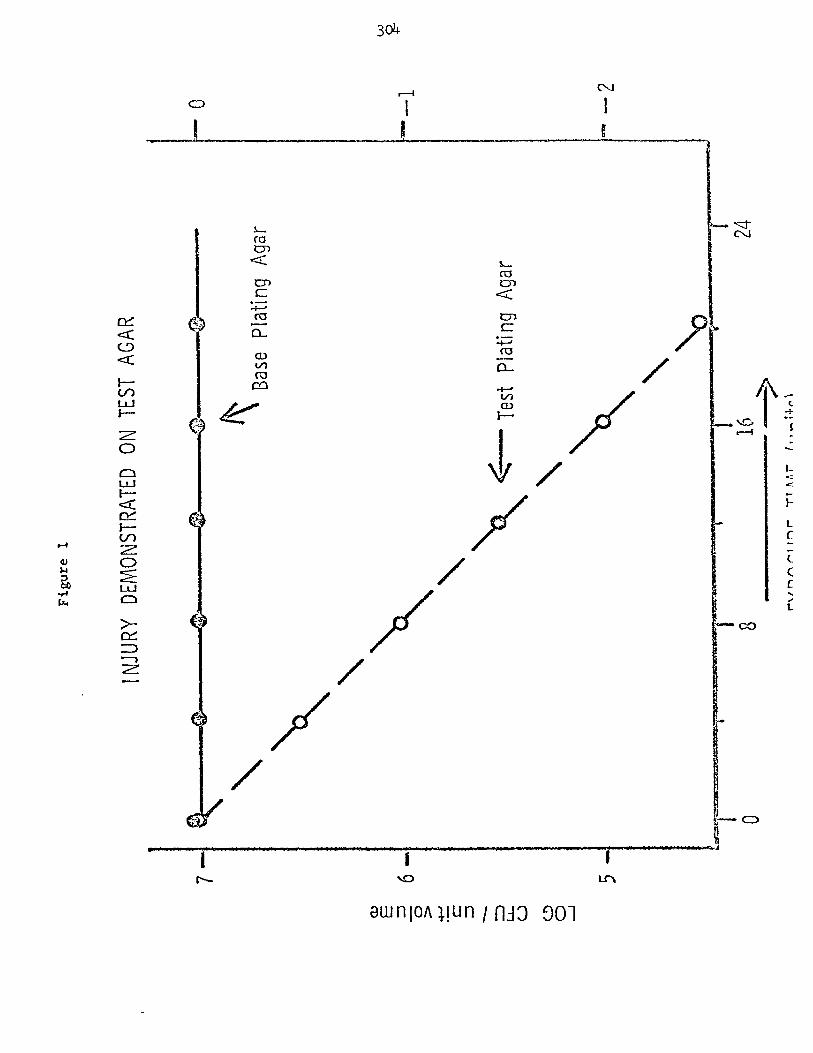
Exposure t o a sub le tha l t r e a t m n t that imposes an environmental s t r e s s w i l l r e s u l t i n the loss of a cha rac t e r i s t i c a b i l i t y t o grow mrnoally under conditions that were sa t i s f ac to ry f o r untreated c e l l s . In Figure 1, a t 0 exposure tlme, equivalent numbers of colony forming u n i t s were observed on a base p la t ing medium and a test plat ing medium. An example of a base p l a t ing medium m i g h t be Trypticase Soy Agar and an example of t he t e s t p la t ing agar m i g h t be Try-pticase Soy Agar w i t h 7.5% N a C 1 , An example of a t e s t microorganism might be 2. aureus exposed t o a heat treatment. Upon exposure t o the environmental s t r e s s , a portion of t he c e l l s lose their cha rac t e r i s t i c tolerance t o a se l ec t ive const i - t uen t (e.g. N a C 1 ) that is present in the t e s t p la t ing agar. r e t a in t h e i r a b i l i t y t o grow in t he absence of t h e se lec t ive agent. The hypothetical data presented in Figure 1 indicate that a f t e r 16 uni t s of exposure time, gg$~ of the init ial population of test microorganisms were unable t o form colonies on the t e s t p la t ing agar . A l l of these organisms were capable of forming colonies on the base p la t ing agar; therefore , the loss in tolerance t o the se l ec t ive agent i n the t e s t p la t ing agar would be interpreted as suble tha l injury in the s t ressed c e l l s . Injury may a l s o be observed as an i n a b i l i t y of an organism t o form colonies i n a defined minimal medium while re ta in ing the a b i l i t y t o form colonies when complex nut r ien ts a r e supplied. The reverse was a l s o observed when t h e c e l l s have been grown i n a minimal medium pr io r t o the s t r e s s . be related t o the loss i n res i s tance t o se lec t ive agents.
The c e l l s
The manifestation of t h i s type of damage may or may not
304
N r-i
1 I 0
II B E
rx 3 7 7
d
P

305
A host of ce l lu l a r changes have been implicated and related t o depressed growth capabi l i t i es of injured c e l l s , but often injury is observed as an increased or new sens i t i v i ty t o se lec t ive agents, a n t i - microbials, or similar substances i n the growth medium of the damaged c e l l s . S a l t tolerance is l o s t by S . aureus, B. s u b t i l i s , E. coli , and - S . ty-phlmuriun. Actinomycin D can penetrate and a c t on damged 5. anatum. Heat-altered spores of C,. perfringens and E. botulinum a re susceptible t o lysozyme-mediated germination.
Deoxycholate inhiFits injured E_. c o l i .
Frozen - S . faeca l i s become sens i t ive t o sodium azide.
Injured c e l l s frequently lose some of t h e i r ce l lu la r material through leakage in to the surrounding medium. release amino acids, small molecular weight ribonucleic acids, and peTtides. Heat-injured S. aureus c e l l s re lease potassium, amino acids, proteins, and 260-IIBI absorbing material . observed i n other damaged bacter ia . Release of lipopolysaccharide indicated damage t o the outer membrane and was correlated w i t h sublethal injury, and preceded death i n heated E. c o l i .
Frozen c e l l s of E_. c o l i
Similar pat terns have been
Mwromlecules within the c e l l a r e modified by s t r e s s . Ribosomal ribonucleic acid was degraded in heated c e l l s of 2. aureus and i n S . typhimurium. induced DNA breaks and DNA repa i r , they have related these changes t o death ra ther than i n j u r y . with heat Injury of salmonellae that responded negatively t o complex nu t r i t i ona l media.
Although several investigators have reported on t h e r k l y
Recently, however, DNA breaks were correlated
Metabolic a c t i v i t i e s associated with the c e l l a r e affected by damaging s t r e s ses . catabol ic capabi l i t i es and a c t i v i t i e s of selected enzymes . Glucose t ransport in heatdamaged S_. t?yphimurium was a l te red . proteinase of - S , Lactis was inactivated by low temperature storage.
Sublethally heat-injured S_. aureus have decreased
A membrane
RESTORATION OF ORIGINAL CAPABILITSES
Damaged c e l l s can undergo subsequent resusci ta t ion where damaged components a r e repaired and the c e l l regains i t s normal capabi l i t i es . By def ini t ion, as described with hypothetical data i n Figure 2, the injured c e l l s have the capabi l i ty t o function in an unres t r ic t ive environment and be restored t o . t he i r normal physiological s t a t e . damage must disappear pr ior t o c e l l divis ion t o indicate injury and ru l e out the poss ib i l i t y of permanent changes i n t h e form of mutation.
The
The hypothetical data in Figure 2 represent a population of damaged c e l l s introduced in to a medium that w i l l support resusci ta t ion or r e p a h of the damage a s w e l l as growth of the microorganism. A t 0 incubation time, the sample w i t h injured c e l l s introduced into the repair medium contained damaged c e l l s (99$) that could not form colonies on the test plat ing agar ( r e s t r i c t i v e or s e l ec t ive ) yet a r e capable of forming colonies on the base plat ing agar (non-selective or non-restr ic t ive) . After 1 hour of incubation in the repa i r medium, about 8@ of the surviving c e l l s have regained t h e i r a b i l i t y t o form colonies i n the

7
a
3 0 >
E - w .- c J
3 0
c.3 0 J
- 6 LL
5
I 1 I I 1
0 1 2 3 4 1 NCUBATI ON TI ME (Hrs)
Figure 2

307
t e s t p la t ing agar. After 2 hours of incubation, essent ia l ly a l l of the damaged c e l l s have regained t h e i r tolerance t o the r e s t r i c t i v e component of the t e s t plat ing agar and can form colonies equally wel l i n the base and t e s t plat ing agar. observed with the base p la t ing agar i n the first 4 hours of incubation indicates t h a t there was no growth during that time; therefore, the increase in colony forming uni t s on the t e s t p la t ing agar must be repa i r of the damaged c e l l s . The untreated c e l l s represented by the upper curve of Figure 2 exhibited a short l ag time but equivalent growth r a t e i n the exponential phase. The extended Lag time is a frequent observation w i t h injured c e l l s .
The absence of increased colony forming un i t s
The need of an appropriate resusci ta t ion medium is demonstrated w i t h the hypothetical data i n Figure 3 . In the absence of appropriate consti tuents which a r e present i n the growth medium but a r e not present i n water, no repa i r and no growth was observed. In the gruwth medium, ant ic ipated repa i r w a s observed during the f i rs t 2 hours of incubation, whereas 2n water, t h e populat ion of damged ce l l s remained a t 9 8 of the survivors during the 6 hours of incubation.
The resusci ta t ion process c lear ly must be related t o some or most of t he ce l lu l a r changes that are present a f t e r a sublethal treatment which impirs the microorganism's a c t i v i t y . Ribosomes that have been degraded during a heat treatment are regenerated. Phospholipids are synthesized during the recovery period. Protein synthesis appears t o be necessary i n repa i r of some frozen, freeze-dried, or heated c e l l s . The repa i r process is dependent upon energy synthesis apparently i n the form of ATP. Heat-induced s ingle strand breaks must be repaired t o prevent death. Synthetic a c t i v i t i e s i n one s t r a i n damaged by a spec i f ic s t r e s s may be unique and not required i n other organisms damaged by d i f f e ren t s t r e s ses .
Recognition of t he presence of po ten t ia l ly damged microorganisms and tkie subsequent value of a resusc i ta t ion period i n the i so la t ion and enumeration procedures is evident in many new methodologies. Most salmonellae i s o h t i o n procedures involve or should use pre- enrichment s teps which allow repa i r of damage. bacteriaceae i n dried foods u t i l i z e s a res tora t ion treatment. A recent ly published plat ing procedure f o r inJured coliforms u t i l i z e s a resusc i ta t ion period before exposure t o the se lec t ive agents. instances, cer ta in se lec t ive media a r e not antagonistic t o damaged c e l l s . Unfortunately, many laboratory procedures continue t o disregard the presence of sublethal ly impaired microorganisms i n ana ly t i ca l samples.
A rapid method t o t e s t f o r Entero-
In some
FACTORS THAT INl?LUEMCE INJURY
Growth conditions that influence the physiological s t a t e and composition of the c e l l or spore may af fec t sucep t ib i l i t y t o damge by subsequent exposure t o one o r more s t r e s ses . Repair of heated S . typhimurium emphasizes that growth media and conditions influence

Base Plating Agar O A 'Test Plating Agar 0 A
I I
I 0 I
I I 1 t I
IN
I
WATER
I i 11 I c I 0 1 2 3 4 5
1 NCUBATION TlME 0 - h )
Figure 3

309
0 cv I
/
d /
p'
p'
F
/

7
E" 3 0 >
LII 3
3 L 0
(3 0
- ct.' .I
- 6
5
3 10
R E P A I R AND GROWTH I N GROWTH MEDIUM
UNTREATED CELLS
Base Plat ing Agar ,A Test P lat ing Agar A
Base P la t ing Agar 0 Test P la t ing Agar
I I
ol I
I I I
I I
0
J I I I 1 0 1 2 3 4
I NC43ATI ON TI ME (Hrs)

NO REPAIR I N WATER
Base Plat ing Agar @A Test Plat ing Agar 6) A
d-dlN GROWTH MEDIUM
I dl I
1 8 IN I
b
WATER
I J 1 v I. I G 1 2 3 4 5
INCUBATION TIME (Hrs.)

suscep t ib i l i t y t o injury, the way injury is expressed, and the mode of r epa i r by the damaged c e l l s . Specific nu t r ien ts , pH, redox poten t ia l , osmolality, water a c t i v i t y , ionic s t rength, surface tension, temperature, ag i ta t ion , gaseous atmosphere, cu l ture age, and any other var iables of a spec i f ic growth system could a c t upon t h e s t ruc ture and functions of the c e l l , and therefore must be considered f o r po ten t i a l influence on t h e express ion of i n jury.
Most f ac to r s that influence c e l l qua l i t y a l s o may have an e f f e c t during appl icat ion of stress. the amount of injury i n freeze-dried 5. typhimurium. Solutes including phosphate buffers, Mg*, and sucrose a l t e r ed the percentages of thermally injured S . ty-phimurium. Lar pH increased damge in freeze-thaw injured S. a u r e G but had l i t t l e e f f e c t on heat injury of t h e same bac ter ia . !&e pH and N a C 1 concentration i n the heating system influenced the amount of heat in jury in 2. aureus. agents a r e directed a t minimizing the extent of freeze-thaw or f reeze- drying damge of c e l l s .
Storage temperature and humidity influenced
Many of t h e frequent ly used cryoprotective
Many of t h e f ac to r s that a f f e c t c e l l qua l i t y and growth m y influence r e susc i t a t ion of injured c e l l s . The r epa i r of heat-injured S. aureus r epa i r of injury in freeze-dried S . ana tm, repa i r of freezerthaw dakged - S . anatum, r ev iva l of heated E. a, r epa i r of dehydration damage in E. -, c o l i minim1 recovery of heated S . ty-phimurium, and in te rac t ion of pH- and N a C 1 on recovery of heat-stressed 2. aureus a r e examples of the extensive in fo rmt ion avai lable on f ac to r s that influence the re turn of injured cells t o normalcy.
BACTERIAL SPORES
Spores a r e d i f f e ren t and have spec ia l charac te r i s t ic components that a r e not present i n vegetative c e l l s . Generally, res i s tance of spores t o s t r e s ses such as heat is considerably greater t h a n that of vegetat ive forms. To demonstrate v i a b i l i t y , the spore must i n i t i a t e germination, germinate, lose its spore-like charac te r i s t ics , grow in to a vegetative c e l l , and multiply i n this form. These many d i f f e ren t b io logica l transformat ions during spore f ormt ion , germinat ion, and outgrowth when coupled w i t h high resis tance t o s t r e s ses make it necessary t o consider spore injury spec ia l ly .
Any measurement of res i s tance depends on a c t u a l or t rue surv iva l and "the a b i l i t y of the surviving spore t o germinate, reproduce, and lead t o a s u f f i c i e n t number of generations t o be recognized as a survivor under the subculture conditions used" (Schmidt, 1955). cul ture media dramatically influenced the apparent surv iva l of damaged spores.
Sub-
Heat-induced suscep t ib i l i t y t o antagonis t ic materials i n t he cu l ture media was evident i n heated spores of c l o s t r i d i a (Olson and Scot t , 1950) and b a c i l l i (Murre11 e t - al. , 1950). (Roberts and Ingram, 196n and of an t ib io t i c s (Flowers and Adam, 1975) on heated spores fu r the r document the occurrence of injury. Chemical treatments a l s o generate apparent damage (Russell and Loosemore, 1964).
The antagonis t ic e f f ec t s of salts

When B. subtilis spores were exposed t o ul t rahigh temperatures fo r short rimes, heat injury was observed by detecting damaged survivors through addition t o the medium of a non-nutritive gecminating agent (Edwards e t a l . , 1965). A spec i f ic germination system was inactivated i n t he spores but the spores remained viable if germinated i n other ways (Adam and Busta, 1972; Adams and Busta, 1972; Busta and Adams, 1972
Apparent loss of v i a b i l i t y a f t e r severe heat treatments of c l o s t r i d i a l spores was reversed with the addition of lysozyme (Cassier and Sebald, 1969; Adams, 1974; Duncan e t a l . , 1972). Lysozyme apparently replaced the normal l y t i c a c t i v i t y of germinating spores. spores repaired during outgrowth (Barach e t a l . , 1975).
These heat-injured The s i te of
heat damage Fn C . spores appears t o be associated with the spore mexribrane(F) Adams, 1976).
Spores of Clostridium botulinum that a r e exposed t o a severe sublethal heat t r e a t m n t are sensi t ized t o NaCl and NaN02 (Jarvis e t e., 1976). than t o unheated n i t r i t e added a f t e r heating.
These spores were m o r e sens i t ive t o n i t r i t e heated i n y e a t
The frequently observed heat-induced s h i r t in optimal temperature f o r maximal enumeration of surviving spores was re la ted t o outgrowth capabi l i t i es ra ther than germination (Prentice and Clegg, 1974). Recently Busta -- e t al. (1976) observed a heat-induced requirement f o r sucrose i n the recovery m e d i u m of forespores of E. cereus. This requirement disappeared after the spores had germinated and reached the f i r s t divis ion.
With data accumulating at an increasing r a t e , heat damage i n b a c t e r i a l spores is being considered in the study of mechanisms of heat res is tance ( G m l d and k i n g , 1974) of spores by any antagonistic agents mst consider po ten t ia l injury when there is an a m r e n t lack of v i a b i l i t y .
Obviously, any research on inact ivatfon
In conclusion, recognition of sublethal injury induced by common meat processing s i tua t ions is imperative. Knowledge on sublethal inJury and resusc i ta t ion lnay be used t o enhance the l e t h a l or control l ing act ion of a processing treatment, m y be used t o minimize damage during preservation of a s t a r t e r cul ture f o r sausage fermentation, may permit a prediction of e f f ec t s of formula modification on subsequent microbial problems, and/or may eliminate or minimize inadequacies within exis t ing or proposed methodology f o r the detection and enumeration of spec i f ic microorganisms. This understanding and knowledge can only increase the competence of decision makers involved in microbiological assessment of meats.
Adams, D . M. 1974. Requirement fo r and sens i t i v i ty t o lysozyme by Clostridium perfringens spores heated a t ultrahigh temperatures-. Appl. Microbiol. 27:797.

3 14
Adams, D. M. and F. F. Busta. 1972. H e a t in jury as the se lec t ive inact ivat ion of a Bacillus s u b t i l i s spore germination system. 377. American SOC. f o r Microb., Washington, D.C.
p. 368- In H. 0. Halvorson, R. Hanson and L. L. Campbell (ed.) Spores V .
Adams, D . M. and F. F. Busta. 1972. Ultrahigh-temperature ac t iva t ion of a low-temperature Bacillus s u b t i l i s spore germination system. Appl. Microbiol. 24:418.
Anellis , A., E . Shattuck, T . La t t , S. Songpasertchai, D . B. Rmley, and E. W, Ross, Jr. 1976. l eve l n i t r i t e / n i t r a t e h a m .
Gamma i r r ad ia t ion a t -30 2 10 C of low Abstr. Ann. Mtg. Amer. SOC. Microbiol.
1976. p. 189.
Barach, J. T. , R . S. Flowers and D . M. Adams. 1975. Repair of heat- injured Clostridium perfringens spores during outgrowth. Microbiol. 30:873.
Appl.
Busta, F. F. 1976. P rac t i ca l implications of injured microorganisms in food. J. Milk Food Technol. 39:138.
Busta, F. F., E. B a i l l i e and W. G . Murrell. lethal damage of Bacillus forespores . Microbiol. 1976. p. 188.
1976. Heat-induced sub- A b s t r . A n n . Mtg. Amer . SOC.
Busta, F. F. and D. M. Adams. 1972. Ident i f ica t ion of a germination system involved i n t he heat injury of Bacillus s u b t i l l i s spores. Appl . Microb io1 . 24 : 412.
Cassier, M. and M. Sebald. 1969. Germination lysozyme-dependente des spores de Clostridium perfringens ATCC 3624 apres traitement thermique . Ann. Inst. Pasteur (Pa r i s ) 117:312.
Duncan, C . L., R . G . Labbe and R . Reich. 1972. Germination of hea t - and al lcs l i -a l tered spores of Clostridium perfringens Type A by lysozyme and an i n i t i a t i o n protein. J. Eiacteriol. 109:550.
Edwards, J. L., Jr., F. F. Busta and M. L. Speck. 1965. Heat injury of Bacillus s u b t i l i s spores a t ul t rahigh temperatures Appl. Microbiol. 13 :858 .
Fennem, 0. R., W . D. Powrie and E. H. Marth. 1973. Lar-temperature preservation of foods and l i v ing mtter. Marcel Dekker, Inc., New York. pp. 386.
Flowers, R . S . and D . M. Adams. 1976. Spore menibrane(s) as t h e s i te of damage wi th in heated Clostridium perfringens spores J. Eiacteriol. 125 :429.
Flowers, R . S. and D. M. Adams. 1975. Inhibi t ion of heat injured Clostridium perfringens spores by polymyxin and neoqc in . Abstr . Ann. Mtg. h e r . Soc . Microbiol, 1975. p. 202.

3 1 5
G a r l d , G. W . and G . J. Dring. 1974. Mechanisms res i s tance . Adv. Microbiol. F’hysiol. 11:137.
Hunt, C . J. and F. F. h s t a . 1974. EScposure t o
of spore heat
N a C l or glycerol as an in jury system coanparable t o f reezing. Soc . Microbiol. 1974. P. 9.
Abstr. Ann. Mtg. Amer.
Ja rv i s , B., A. C . Rhodes, S . E. King and M. Pate l . 1976. Sens i t iza t ion of heat-damged spores of Clostridium botulinum, type B, t o sodium chloride and sodium n i t r i t e . J. Food Technol. 11:41.
Murrell, W G ., A. M. Olsen and W J. Scot t . 1950. The enumeration of heated b a c t e r i a l spores. 11. Experiments with Bacillus species. A u s t r . J. Sc i . Res. B3:234.
Olsen, A. M. and W. J. Scot t . 1950. The enumeration of heated bac te r i a l spores. of Clostridium. Aust. J. Sci . Res. B3:219.
I. Experiments with Clostridium botulinum and other species
Prentice, G. A. and L.F.L. Clegg. 1974. The e f f e c t of incubation temperature on the recovery of spores of Bacillus s u b t i l i s 8057. J. Appl. Bacter iol . 37:501.
Ray, B. and M. L. Speck. 1973. Freeze-injury i n bac te r i a . C.R.C. C r i t i c a l Rev., C l in i ca l Laboratory Sciences. p. 161,
Roberts, T. A . and M. Ingram. 1966. The e f f ec t s of N a C 1 , IUVO3, and NaN02 on recovery of heated b a c t e r i a l spores. 1: 147.
J. Food Technol.
Russell, A . D. and M. Loosemore. s u b t i l i s spores a t elevated temperatures . 1964. Effect of phenol on Bacillus
Appl. M i c r o b i o l m .
Scheusner, D. L., F. F. B u s t a and M. L. Speck. 1971. Injury of bac ter ia by san i t i ze r s . Appl. Microbiol. 21 :k l .
Schmidt, C . F , 1955. The res i s tance of b a c t e r i a l spores with reference Ann. Rev. Microbiol. 9:387. t o spore germination and its inh ib i t ion .
Stevenson, K. E. and L. J. Richards. 1976. Thermal injury ~ n d recovery of Saccharomyces cerevis iae . J. Food Sc i . 41:136.

Bruce Langlois: Thank you, D r . B u s t a . Are there any questions.
F. F . Busta: I 'd l i k e t o point out th ree spec i f ic areas t h a t a r e If you remember, being used now that a r e by-passing suble tha l injury.
Bruce sa id that VFU3 is s t i l l a very popular method f o r the evaluation of c o l i f o r m and we a r e w e l l aware of the damage t o coliforms being observed with VRB medium.
One of t he a l t e rna t ive approaches of p la t ing and detect ing damaged coliforms that has been published recent ly , is the method u t i l i z i n g Try-pticase Soy Agar, where the organism and the 'I4ypticase S o y Agar a r e plated and incubated fo r , I believe, four hours, then they a r e overlaid with an equal amount of Violet Red Bile Agar t o get t he equivalent s e l e c t i v i t y of v i o l e t Red Bile Agar, but t he resusc i ta t ion period i n Try-pticase Soy Agar. So, i n t h i s way we s t i l l have the s e l e c t i v i t y of t he Bile Sa l t s , and y e t permit damaged organisms t o r epa j r . There is a great increase i n the u t i l i z a t i o n of Rose Bengal and a n t i b i o t i c agar r a the r than ac id i f i ed agar f o r the enumeration of the molds. I bel ieve a l o t of t h i s i s based on possible in ju r i e s as wel l as possible detachment f o r the organisms that can' t t o l e r a t e the low PH ac id i f i ed medium. I t h h k the increased use of pre-enrichment f o r the detect ion of Salmonella is an indicat ion of possible suspicion of injury and an attempt t o have a period of resusc i ta t ion before se l ec t ive media a r e used.
Bruce Langlois: Any questions? This brings t o a close the session of Microbiological Techniques. I w i l l t u r n it over t o G a r y Smith f o r t he second up-date.
Gary Smith: Our second up-date speaker i s Mike McClory with Jewel Food Stores . Mike graduated in 1948 from Ohio S ta t e University with a degree of Bacteriology. there f i v e years; moved t o Oscar Mayer and worked there two years . He moved t o Rath Packing Company, worked the re six years. He went t o Marhoefer Packing Company in Muncie, Indiana, and worked the re f o r seven years. and Qua l i ty f o r Jewel Food Stores in Melrose Park, Illinois.
H e s t a r t ed with Armour and Company, worked
Final ly , he wound up as Director of Assurance Sani ta t ion
We're interested i n him, spec i f i ca l ly t h i s morning, because he serves on the two more important committees associated with the areas we ta lked about t h i s morning. One, he is Chairman of the Food, D r u g and Cosmetic Division of t he American Society of Qua l i ty Control and, more importantly, on t h e top ic he is going t o t a l k about t h i s morning, he 's Chairmn of Qua l i ty Assurance and Sani ta t ion fo r t he National Association of Food Chains. t o t r y t o define, as they o r ig ina l ly envisioned, t he problem of Food Qua l i ty Standards. Standards. they ' re down t o j u s t working with ground beef. member of t he American Meat Science Association and the I n s t i t u t e of Food Technologists. We're pleased f o r our second up-date session t o br ing Mike t o the f loo r t h i s morning.
They've been working f o r t he last two years
They narrowed that down t o Meat Microbiological I'm sure, as h e ' l l t a l k t o you about i n a few minutes,
He i s a long time







