

INSTITUTO POLITÉCNICO NACIONAL
ESCUELA NACIONAL DE CIENCIAS BIOLÓGICAS SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN
Estudio de la microbiota asociada a la mariposa monarca Danaus plexippus durante la hibernación en los bosques mexicanos
T E S I S
QUE COMO UNO DE LOS REQUISITOS PARA OPTAR AL GRADO DE
DOCTOR EN CIENCIAS EN BIOMEDICINA Y BIOTECNOLOGÍA MOLECULAR
P R E S E N T A :
J O R G E B R A V O M A D R I G A L
DIRECTORES DE TESIS:
DR. HECTOR M. ZEPEDA LÓPEZ
DR. CARLOS WONG RAMÍREZ
M É X I C O, D. F. ENERO 2 0 0 9


ii
El presente trabajo se realizó en el laboratorio de Medicina de Conservación de la Escuela Superior de Medicina bajo la dirección del Dr. Héctor M Zepeda López y la dirección del Dr Carlos Wong Ramírez. Se llevo a cabo con el apoyo financiero de la dirección de Estudios de Posgrado e Investigación bajo proyecto de investigación registrado SIP 20050576 Estado de Salud de la Mariposa Monarca en México y por el apoyo financiero del Consejo Nacional de Ciencia y Tecnología, otorgado al Becario No. 175431. Se agradece a la Secretaría del Medio Ambiente y Recursos Naturales, a la Procuraduría Federal de Protección al medio Ambiente y a la Comisión Nacional de Áreas Naturales Protegidas, en particular a la Reserva de la Biosfera Mariposa Monarca, por los permisos de colecta, así como el apoyo y asistencia recibidos durante los monitoreos en los santuarios de hibernación. Se agradece al programa de aseguramiento de la calidad PACAL por las cepas por las cepas amablemente donadas para llevar a cabo este trabajo. Un agradecimiento especial a todos los guías y guardabosques, así como a la población en general de los ejidos el Rosario, Sierra Chincua, La mesa y San Antonio Albarranes, por su apoyo en la ubicación de las colonias de hibernación, así como por la información compartida.

iii
ÍNDICE GENERAL
Página
Índice de cuadros vi
Índice de figuras viii
Abreviaturas x
Abstract xi
Resumen xii
1. Introducción 1
1.1 Distribución mundial de la mariposa monarca 1
1.2 Ciclo de vida de la mariposa monarca 1
1.3 Sitios de Hibernación 2
1.4 Amenazas potenciales en los sitios de hibernación de
la mariposa monarca 2
1.5 Enfermedades infecciosas en la mariposa monarca 3
1.6 Efecto de la infección parasitaria sobre la salud de la
mariposa monarca 4
1.7 Relación entre parasitosis y la migración de la mariposa monarca 5
1.8 Ciclo de vida de O. elektroscirrha 5
1.9 Patógenos virales de la mariposa monarca 6
1.10 El papel de las bacterias asociadas a la mariposa y a los insectos 7
1.11 La microbiota bacteriana en los insectos 8
2. Antecedentes 9
3. Justificación 13
4. Objetivo general 14
5. Objetivos particulares 14
6. Materiales y métodos 15
6.1 Muestreos 15
6.2 Colecta de mariposas para estudios parasitológicos. 15
6.3 Colecta de mariposas para estudios bacteriológicos 16
6.4 Obtención de muestras de suelo, corteza, agua y aire 16

iv
6.5 Análisis parasitológico 18
6.5.1 Obtención de improntas de abdomen para análisis
parasitoscópico. 18
6.5.2 Búsqueda de Ophryocystis elektroscirrha en improntas de
abdomen de mariposas 18
6.6 Transporte de muestras 19
6.7 Análisis Bacteriológico y Micológico 19
6.7.1 Procedimiento de elección de medio de cultivo 19
6.7.2 Selección de mariposas para el análisis 20
6.7.3 Cultivo y procesamiento de las muestras 20
6.8 Análisis Bacteriológico de corteza, suelo, agua 21
6.9 Condiciones de incubación 21
6.10 Aislamiento y construcción de cepario 21
6.11 Identificación de los microorganismos aislados 21
6.12 Cepas de referencia 23
6.13 Identificación de géneros y especie 24
6.14 Análisis Estadístico 25
7. Resultados 26
7.1 Análisis Parasitológico 26
7.1.1 Análisis de la distribución de árboles con mariposas infectadas. 26
7.1.2 Variación de la prevalencia de infección con Ophryocystis
elektroscirrha 31
7.1.3 Comparación de la prevalencia entre diferentes sitios
de hibernación. 31
7.1.4 Comparación de la parasitosis entre mariposas muertas y vivas,
así como entre hembras y machos. 33
7.1.5 Influencia de las heladas en la prevalencia de O. elektroscirrha. 36
7.2 Análisis Bacteriológico. 38
7.2.1 Diferencias en la carga microbiana encontrada en diversos sitios
anatómicos. 38

v
7.2.2 Caracterización bioquímica. 39
7.2.3 Carga bacteriana de cada género aislado en la mariposa monarca 43
7.2.4 Descripción de la microbiota y comparación en los diferentes
sitios anatómicos 48
7.2.5 Temporada de hibernación. 49
7.2.6 Santuarios de hibernación. 51
7.2.7 Asociación entre O. elektroscirrha con grupos microbianos. 53
7.2.8 Diferencias en la microbiota presente en mariposas vivas
y mariposas muertas. 55
7.2.9 Análisis microbiológico de muestras ambientales. 56
7.3 Análisis micológico 57
8. Discusión 61
8.1 Análisis parasitológico. 62
8.2 Análisis Bacteriológico. 66
8.2.1 Diversidad microbiana en diferentes sitios y fases de hibernación. 75
8.2.2 Asociación entre O. elektroscirrha con grupos microbianos 77
8.2.3 Diferencias entre la microbiota presente en las mariposas
vivas, mariposas muertas y muestras ambientales. 79
8.3 Análisis micológico. 81
9. Conclusiones. 83
10. Bibliografía. 84
Apéndice. 96

vi
ÍNDICE DE CUADROS
Página
Cuadros
1 Relación de las muestras obtenidas en los sitios de hibernación,
temporada y fase de la hibernación. 17
2 Pruebas preliminares para la identificación presuntiva de
bacterias aisladas 22
3 Modelo de distribución de árboles con mariposas infectadas,
considerando 4 regiones en la temporada de hibernación
2006-2007. 30
4 Modelo de distribución de árboles con mariposas infectadas,
considerando la periferia y el centro de la colonia durante
2007 a 2008 30
5 Descripción general de las bacterias encontradas en este trabajo. 42
6 Comparación entre la microbiota bacteriana gram
negativa, encontrada en abdomen, alas y cefalotórax de
mariposas monarca. 49
7 Comparación entre la microbiota bacteriana gram
positiva, encontrada en abdomen, alas y cefalotórax de
mariposas monarca. 50
8 Variación en la frecuencia de aislamientos a partir de
abdomen de mariposa monarca en función de la temporada
de hibernación 51
9 Géneros bacterianos cuyas frecuencias de aislamiento fueron
similares durante las tres fases de hibernación de la
mariposa monarca 51
10 Distribución de especies y géneros bacterianos gram
negativo en diferentes santuarios de hibernación de la
mariposa monarca 52
11 Distribución de especies y géneros bacterianos gram positivo en
diferentes santuarios de hibernación de la mariposa monarca. 53

vii
12 Resultados de aislamientos obtenidos a partir de muestras
ambientales 56
13 Descripción de las bacterias presentes en fuentes ambientales. 57
14 Tipos de hongos y levaduras aislados en el abdomen de la
Mariposa Monarca. 60
15 Géneros bacterianos propuestos para ser candidatos a la
microbiota de abdomen en la mariposa monarca. 78
A Anexo A. Total de bacterias encontradas en abdomen, alas y
cefalotórax de mariposas monarca 96

viii
ÍNDICE DE FIGURAS
Página
Figuras
1 Mariposa monarca migratoria adulta. 2
2 Superficie de ocupación de la Mariposa Monarca en hectáreas
desde 1993 a 2007 registradas en los santuarios de hibernación
en México. 10
3 Sitios seleccionados para la colecta de agua 17
4 Procedimiento para realizar las improntas de escamas del
abdomen de la mariposa 18
5 Microfotografías de ooquistes en algunas improntas. 27
6 Patrones de adherencia ó asociación de Ophryocystis
elektroscirrha con las escamas de la mariposa monarca.. 28
7. Distribución de la frecuencia de parasitosis de mariposa monarca 29
8 Prevalencia de parasitosis con Ophryocystis elektroscirrhae
desde 2003 a 2008 31
9 Variación en prevalencia de parasitosis de la mariposa
monarca en los sitios de hibernación. 32
10 Variación en prevalencia de parasitosis de la mariposa
monarca por temporada en cada sitio de hibernación 32
11 Parasitosis media desde 2003 a 2008 en mariposas vivas
y mariposas muertas. 34
12 Diferencia en la prevalencia de parasitosis entre machos y
hembras vivas. 35
13 Diferencia en la presencia de parásitos en mariposas muertas 36
14 Efecto de las nevadas en la parasitosis de la mariposa monarca. 37
15 Carga microbiana total en diferentes sitios anatómicos de
la mariposa monarca. 39
16 Dendograma simplificado que muestran el agrupamiento de
los aislados en grupos fenotípicos 40
17 Análisis de la carga microbiana recuperada del abdomen 44

ix
18 Análisis de la carga microbiana en las alas de la
mariposa monarca 45
19 Análisis de la carga microbiana en el cefalotórax de
la mariposa monarca. 46
20 Carga microbiana y géneros frecuentemente aislados, a
partir de diferentes sitios anatómicos de la mariposa monarca. 47
21 Diferencias observadas en la microbiota presente en mariposas
parasitadas y no parasitadas con ooquistes de O. elektroscirrha. 54
22 Comparación entre la frecuencia de aislamiento de
bacterias en mariposas vivas y mariposas muertas. 55
23 Frecuencia relativa de aislamiento de hongos en tres sitios de
hibernación de la mariposa monarca. 58
24 Morfología microscópica de los principales hongos identificados. 59
25 Morfología macroscópica de los principales hongos
encontrados en la mariposa monarca. 59
26 Morfología macroscópica y microscópica de Cladosporium sp. 60

x
ABREVIATURAS
ACPAM Agar caseína peptona almidón monarca
ANOVA Análisis de varianza
ASTLM Agar soya tripticaseína levadura monarca
BGNF Bacilos gram negativo fermetadores
BGNNF Bacilos gram negativo no fermetadores
BGP Bacilos gram postivo
CCE Comisión de cooperación ambiental
CGPCN Cocos gram positivo catalasa negativa
CSS Coeficiente simple de similitud
CTA Agar Cisteína Triptosa
G.L. Grados de libertad
LMC Laboratorio de Medicina de Conservación
msnm metros de altura sobre el nivel del mar
OF Medio oxidación-fermetación de Hugh Leffson
PBS solución reguladora de fosfatos
p/v Peso/volumen
RBMM Reserva de la Biosfera Mariposa Monarca
R.V. Razón de varianzas
SIM Medio Sulfuro Indol Movilidad
UFC Unidades formadoras de colonias
UPGMA Método de comparación de pares y grupos
VP/RM Medio Voges Proskaure-Rojo de Metilo
v/v volumen / volumen

xi
Resumen
La mariposa monarca Danaus plexippus, migra cada año desde Canadá y Estados Unidos a los
bosques de Oyamel en México, donde hibernan, sobreviviendo así las bajas temperaturas del
invierno. A pesar de que esta especie está protegida por los gobiernos de los tres países,
recientemente se han observado grandes mortandades, principalmente atribuibles a tormentas o
nevadas, sin embargo, aún no se ha investigado el papel que juegan otros factores en la salud de
la monarca, como los microorganismos. El objetivo de este trabajo fue analizar la prevalencia de
microorganismos patógenos como el protozoario Ophryocystis elektroscirrha en la población de
mariposa monarca hibernante así como identificar la diversidad microbiana cultivables asociadas
a la mariposa en 4 sitios de hibernación en México. Se realizaron 74 muestreos de diciembre de
2003 a noviembre de 2008, en los cuales se obtuvieron mas de 29500 improntas, así mismo se
colectaron 96 mariposas vivas y 96 muertas que se dividieron en alas, cefalotórax y abdomen
para realizarse cultivos bacteriológicos y micológicos. Se utilizaron procedimientos
convencionales de bacteriología para el cultivo, cuenta y aislamiento de bacterias, así mismo se
utilizó la taxonomía numérica considerando 96 pruebas fenotípicas para la identificación. Los
resultados indican que la prevalencia de parasitosis desde 2005 a 2008 se incrementó de 2.5 a
9.5% en tres años; aparentemente la prevalencia de infección no esta relacionada con el sexo, ni
con el sitio de hibernación de la mariposa monarca; así mismo no hubo diferencia en la
parasitosis entre mariposas vivas y muertas. Los resultados bacteriológicos muestran que el
abdomen es el sitio con mayor cantidad de bacterias, siendo los principales grupos microbianos
encontrados en abdomen Enterobacter spp, Staphylococcus spp y Pasteurella spp; en alas se aisló
frecuentemente Enterobacter spp, y Staphylococcus spp mientras que en cefalotórax se aislaron:
Cromobacterium violaceum, Chryseobacterium meningosépticum, Enterobacter aerogenes,
Micrococcus cristinae y Turicella otitidis. De acuerdo con los estudios ambientales los géneros
Serratia spp, Bacillus spp y Klebsiella spp podrían ser parte de la microbiota transitoria. El
análisis micólogico mostró que posiblemente el hongo Cladosporium sp es una especie
frecuentemente asociada a la mariposa monarca.

xii
Abstract
Monarch butterfly Danaus plexippus migrates every year from Canada and USA to Oyamel fir
trees in the Mexican forest, where it hibernates to avoid low temperatures prevalent in North
America. Despite of this specie is protected by the three countries, sometimes high mortality has
been seen, mainly due to storms or snowstorms; however the roll that other agents, such as
microorganisms, play in the monarch health, has not been investigated yet. The aim of this work
was analyze the prevalence of pathogen microorgnaims such as the protozoo Ophryocystis
elektroscirrha in the overwintering monarch pupulation as well as to identify all cultivable
bacteria belloging to the microbiota associated to monarch butterfly collected in four Mexican
overwintering sites. For this purpose, 74 samplings were taken from December 2003 to March
2008, in where more than 29500 scales slades were taken 96 alive butterflies and 96 dead, that
were dissected in abdomen, wings and cephalothorax to carrie out bacteriological and
micological cultures. Conventional bacteriological techniques for getting cultures, microbial load
and isolated bacteria were applied; moreover numerical taxonomy considering 96 phenotipic test,
was used to identify all strains recovered. The results showed that the prevalence of infection
with Ophryocystis elektroscirrha has increased since 2005 from 2.5 to 9.5%, apparently the
infection prevalence was no related neither the sex nor the overwintering sanctuary; as the same
way there was no difference in parasitosis frequency between alive or dead butterflies.
Bacteriological results showed that abdomen is the part with more bacterial load. The main
bacterial groups found in abdomen were Enterobacter spp, Pasteurella spp, and Staphylococcus
spp. In the wings, Enterobacter spp and Staphylococcus spp were frequently isolated; whereas
Cromobacterium violaceum, Chryseobacterium meningosepticum, Enterobacter aerogenes,
Micrococcus cristinae, and Turicella otitidis, were mainly isolated in cephalothorax. Considering
the environmental results, genus sucha Serratia spp, Bacillus spp y Klebsiella spp could be part
of the transient microbiota. Micological analysis showed that Cladosporium sp fungus was a
specie frequently associated to monarch butterfly.

1
1. Introducción.
1.1 Distribución mundial de la mariposa monarca.
La mariposa monarca, (Danaus plexippus) es un Lepidóptero que pertenece al orden Hexapodos,
se encuentra distribuido en varias partes del mundo; en América se encuentra en Canadá, Estados
Unidos de América (EUA), México, Cuba, Perú y Argentina; también se ha encontrado
continente, en Australia y Hawai. Sin embargo, en Norteamérica se produce un fenómeno que no
ocurre en los demás sitios, la migración de la mariposa monarca a través de más de 3000km
(Malcolm, 1987), a este grupo de mariposas se le llama población de mariposas migrantes del
Este, ya que inician su viaje desde Minnesota y Wisconsin. También hay otra población ubicada
al oeste de EUA en los estados de Nevada, Utah, Oregon, Washington y Colorado que migra una
distancia corta hacia la costa central de California a la que se le denomina población de
mariposas migrantes del oeste (Altizer, 1999).
1.2 Ciclo de vida de la mariposa monarca.
El ciclo de vida la mariposa monarca, inicia cuando la hembra deposita sus huevos sobre la parte
posterior de una hoja de Asclepia syrica conocida comúnmente como algodoncillo o asclépia,
estas plantas, además de brindarle seguridad, también sirven como alimento para las larvas en el
momento de la eclosión del huevo. Cuando las larvas han salido, éstas comienzan una etapa de 4
mudas, en este período la monarca únicamente consume hojas de asclépia, las larvas presentan
bandas de color amarillo, blanco y negro que la hacen muy llamativa, esta característica las
protege de las aves, sus principales depredadores ya que se ha visto que el consumo de estas
hojas confiere a las mariposas de un fuerte alcaloide venenoso para las aves (Erickson, 1973). Por
otro lado, las larvas son el estadio más susceptible tanto a insectos depredadores (Koch y cols.,
2003) como a infecciones por microorganismos, dentro de los cuales encontramos; virus
(Herniou y cols., 2004; Urquhart, 1966), hongos (Hajek y cols., 1999), parásitos (Leong y cols.,
1997) y bacterias (Sikorowski and Lawrence 1998). Al término del cuarto estadio de larva, se
inicia la formación de la crisálida, la cual es un saco color verde que presenta característicamente
una especie de rostelo dorado que asemeja una corona de ahí la denominación de “monarca”.
Conforme pasan los días la cubierta de la crisálida se hace cada vez más delgada hasta que
incluso se pueden notar los colores característicos de las alas, este proceso toma lugar de 8 a 13

2
días. Al finalizar este período, el adulto rasga la crisálida y emerge de ella liberando primero sus
patas y posteriormente sus antenas y alas, las cuales deben primero endurecerse para que éste
pueda volar (Galindo-Leal y cols., 2000).
El adulto de la mariposa monarca mide aproximadamente 10-12 cm con las alas extendidas, su
cuerpo consta de cabeza, tórax y abdomen, las dos primeras partes presentan un color negro con
puntos blancos, las alas son de color anaranjado y presentan una serie de nervaduras negras. El
peso de la mariposa no llega a un gramo, los machos son ligeramente más grandes que las
hembras y presentan dos puntos negros en las vénulas (stigmata), (Figura 1) estas estructuras
producen feromonas que atraen a las hembras. Después de 8 días de haber emergido del capullo o
crisálida, la mariposa está lista para reproducirse, aparearse y depositar los huevos, el tiempo total
de este ciclo es de 30 días, de tal manera que un adulto vive aproximadamente 5 semanas; estos
ciclos cortos se repiten cuatro veces en la primavera, período en el cual la mariposa monarca está
regresando al norte de los EUA. Después de la cuarta generación nace un tipo especial de
mariposas con la capacidad de poder vivir hasta 8 meses, tiempo suficiente para preparase para la
migración de otoño, hibernar, y reemigrar iniciando entonces la reproducción (Galindo-Leal y
cols. 2000).
Figura 1. Mariposa monarca migratoria adulta. El sexo de las mariposas monarca se diferencia fácilmente por la presencia de 2 stigmatas en el macho (flecha). A) macho B) hembra. (fuente LMC, 2004)
1.3 Sitos de hibernación.
Durante la migración que realiza la población del este, se ha estimado que mas de 100 millones
de mariposas arriban a nuestro país (Wassenar,1998), particularmente a los bosques de oyamel
A B

3
ubicados a 3000 msnm en la zona montañosa del Estado de Michoacán y del Estado de México,
en sitios llamados santuarios, donde se llega a estimar la presencia de más de 10 millones de
mariposas por hectárea, Se han documentado consistentemente 12 sitios o santuarios en los
cuales las mariposas forman racimos en las ramas o troncos de oyamel (Abies religiosa),
formando colonias de mariposas en hibernación (Calvert and Brower, 1986).
1.4 Amenazas potenciales en los sitios de hibernación de la mariposa monarca.
La colonia más numerosa se encuentran en la sierra El Campanario en el ejido El Rosario
(Martínez, 2007; Rendón-Salinas y cols., 2006), tanto esta colonia como la ubicada en Sierra
Chincua, son las que tienen mayor impacto de las poblaciones humanas, ya que ambas son
explotadas para actividades ecoturísticas (SEMARNAT, 2001) que han originado el aumento de
polvo en los santuarios, el desvio de riachuelos, la disminución de humedad y el aumento de
temperatura, lo cual ha ocasionado que las mariposas se desplacen a distancias mayores en busca
de agua (CEC, 2008a). Por otra parte, hay otras colonias menos perturbadas como la que se ubica
en la sierra El Campanario, en el ejido de La Mesa, en el borde entre los Estados de México y
Michoacán, y en el cerro de las Palomas, ubicado en el ejido San Antonio Albarrales, a un
costado del volcán Nevado de Toluca, en estos sitios aunque también reciben visitas de turistas,
la cantidad es considerablemente menor.
Otro fuerte impacto causado por las comunidades humanas en el hábitat de este lepidóptero, ha
sido la disminución de sus zonas de hibernación causadas por la tala ilegal. Estudios realizados
desde 1971 hasta 1999 han mostrado la reducción del bosque debida a esta actividad (Brower y
cols., 2002), la cual, aun con los decretos de protección a los hábitat de la mariposa monarca
promulgados desde 1980 a 2000 (SEMARNAT, 2001), no ha logrado suspender esta actividad al
100% y esta sigue afectando la hibernación de la mariposa, aunque en menor escala (WWF,
2004).
1.5 Enfermedades infecciosas en la mariposa monarca.
En cuanto a las amenazas causadas por agentes infecciosos, actualmente se ha documentado
principalmente la susceptibilidad de la larva de la monarca frente a un parásito: Ophryocystis
elektroscirrha, ésto usualmente sucede durante la reemigración, apareamiento y reproducción, ya

4
que, durante toda la ruta migratoria se encuentra una gran variedad de plantas entre ellas las
asclépias que en gran medida afectan el ciclo de la mariposa monarca. Estas plantas
principalmente se les encuentra en grandes extensiones de pastizales, sin embargo debido al gran
poder de adaptación, se ha encontrado incluso en los bosques donde los árboles son muy
abundantes y prácticamente obstruyen el paso de luz, lo anterior ha ocasionado que las asclépias
sean consideradas una plaga, ya que, incluso han invadido los cultivos de maíz. Los granjeros, en
afán de eliminarlas, han utilizado una gran variedad de herbicidas que han tenido un fuerte
impacto en la salud de la mariposa monarca, de igual manera grupos de agricultores con objeto de
eliminar plagas de ciertas polillas, han utilizado métodos de control biológico, empleando al
Bacillus thuringiensis, conocida bacteria productora de toxinas contra lepidópteros (Sears y cols
2001; Sampson y cols., 1998; Tamez-Guerra y cols., 2004).
Esto ha afectado a las poblaciones de la mariposa en su migración, ocasionado que las mariposas
modifiquen sus rutas y entren a otras zonas, muchas de estas zonas están contaminadas con
parásitos, como Ophryocystis elektroscirrha. Este parasito pertenece al orden neogregarinida,
clase Sporozoa Phillum Protozoa, las gregarinas, grupo al cual pertenece este parásito, se
caracterizan por ser parásitos de invertebrados, la fase infectante es el ooquiste (Reyes, 2004) el
cual es de color café oscuro, tiene una forma ovoide y dimensiones de 14 x 9 µm, este ooquiste
comúnmente se encontra en las escamas del abdomen aunque también se distribuye en las alas y
el tórax (Leong y cols., 1992).
1.6 Efecto de la infección parasitaria sobre la salud de la mariposa monarca.
Aunque se tiene bien documentado que la infección de las larvas conduce a serias secuelas que
dependen de la dosis, el efecto en los adultos no está bien claro, algunos experimentos indican
que las mariposas altamente infectadas no migran distancias grandes, sin embargo estas
conclusiones no están completamente probadas ya que se ha visto que, incluso mariposas
altamente infectadas, tienen la capacidad de llegar hasta los sitios de hibernación en nuestro país
(Bradley y cols,. 2005), no obstante hay enormes diferencias en cuanto a la prevalencia de O.
elektroscirrha en las poblaciones migratorias del Oeste y las poblaciones que llegan a nuestros
bosques.

5
1.7 Relación entre parasitosis y la migración de la mariposa monarca.
Se ha encontrado una relación de disminución de la prevalencia conforme aumenta la distancia de
migración, así las poblaciones migratorias del este, migran enormes distancias, y presentan una
prevalencia de parasitosis menores al 8%. Por otra parte, las poblaciones del oeste, aunque
también realizan migración, la distancia es considerablemente menor y mantienen tasas de
parasitosis del 30 al 50%, finalmente, una tercer población en Florida, no realiza migración, es
decir, la población es residente, aquí la prevalencia incluso alcanza el 96% (Altizer, 2001).
Una hipótesis señala que la distancia de la migración ocasiona esta diferencia, producto de la
dispersión del parasito, es decir las mariposas de la población de Florida tienen altas tasas de
infección, dado que es altamente probable encontrar asclépias que previamente estuvieron en
contacto con otra mariposa posiblemente infectada, por otro lado en las poblaciones migratorias
este evento es menos probable, ya que la colonia está en movimiento, por la gran distancia de
migración, lo que origina que las probabilidades de que una mariposa llegue a una asclépia
parasitada sea mínima, en consecuencia las posibilidades de transmisión por esta vía podrían ser
mínimas.
Al considerar lo anterior, resulta claro, que las asclépias juegan un papel muy importante en la
transmisión del parásito. Recientemente se ha constatado la presencia cada vez mayor de
asclépias en los bosques de oyamel donde hiberna la mariposa monarca, esto ha llevado a
sostener la posibilidad de que se forme una colonia residente permanente, según el reporte de
algunos pobladores se han visto colonias residentes cercanas a las faldas del volcán Popocatepetl
esto en vez de ser benéfico, podría tener efectos negativos ya que casi todas las colonias no
migratorias muestran tasas muy altas de infección con O. elektroscirrha.( Altizer 2001; Bradley y
cols., 2005).
1.8 Ciclo de vida de O. elektroscirrha.
El ciclo de vida de este parásito inicia cuando se ingieren las esporas por una larva, esto es
frecuente en zonas de alta prevalencia de O. elektroscirrha, para que una larva puede ingerir las
esporas, éstas deben estar sobre las asclépias, esto es frecuente ya que las hembras infectadas al
momento de la ovoposición contaminan sus huevos, de tal manera que cuando las larvas

6
eclosionan, éstos consumen el resto del huevo o corion, ingiriendo así los ooquistes, a esta vía de
infección se le conoce como vertical. Cuando el parásito entra al intestino de la mariposa, lisa las
células y salen esporozoitos que penetran los tejidos; el parasito lleva a cabo su replicación a
través de una merogonia, en la que se producen de 32 a 64 esporozoitos por cada ooquiste
ingerido. Se ha observado que durante los 5 estadios que presenta la monarca antes de convertirse
en adulto, el parásito lleva a cabo 2 merogonias, de tal manera que se pueden producir mas de
4000 esporozoitos por cada ooquiste. Durante el período de maduración de la mariposa en la
crisálida los parásitos completan una fase sexual y forman alrededor de las escamas las
estructuras latentes llamadas ooquistes, los cuales acompañarán a la mariposa durante toda su
vida pudiendo incluso infectar a otra mariposa en una transmisión de tipo horizontal. (Altizer y
cols 1999: Leong y cols., 1992; Reyes, 2004).
Cuando una larva se infecta con los ooquistes, pueden pasar dos eventos: o muere antes de
emerger de la crisálida o bien emerge como una mariposa con un nivel de infección
sorprendentemente alto (Leong y cols, 1997) sin embargo es común encontrar también mariposas
con niveles de infección menores, ésto sucede durante el apareamiento, en donde los machos
altamente infectados, trasfieren el ooquiste a las hembras en el momento del apareamiento, esto
se ha considerado una transmisión vertical venérea y aunque esta infección no le genera
problemas a la hembra, es suficiente para infectar los huevos y así amplificar la infección (Altizer
y cols., 2000; Reyes, 2004).
1.9 Patógenos virales de la mariposa monarca.
Además del protozoario O. elektroscirrha, hay antecedentes que muestran la susceptibilidad de
las larvas de la mariposa monarca a un virus del grupo de los Baculovirus. Estudios de
microscopía electrónica muestran a las partículas virales dentro de la oruga. Sin embargo, aún no
se conoce sí estos virus están presentes en el adulto o sólo en larvas, tampoco se sabe nada acerca
de la prevalencia (Urquhart y cols., 1966).
Los Baculovirus son virus de DNA, específicos de artrópodos, los cuales se han aplicado en
estrategias de control biológico y como vectores de clonación específicos para eucariontes, Éstos
han sido herramientas biotecnológicas muy útiles en la expresión de genes en células

7
eucarióticas, sin embargo en vida libre pueden ocasionar daño y muerte principalmente a la larvas
de lepidópteros en donde pueden causar lesiones granulocíticas o producir cuerpos de inclusión,
en ambos casos estos efectos conducen a la muerte de la mariposa o polilla (Arnott y cols., 1968;
Herniou y cols., 2004).
1.10 El papel de las bacterias asociadas a la mariposa y a los insectos.
Otro grupo microbiano frecuentemente asociado con artrópodos los constituyen las bacterias.
Dentro de este grupo encontramos reconocidos patógenos de insectos, pero también se encuentra
una gran cantidad de microorganismos no patógenos e incluso benéficos. Los estudios previos en
los que se ha analizado la microbiota de insectos muestran que las especies comúnmente
encontradas son miembros de la familia Enterobacteriaceae tales como Serratia, Enterobacter,
Klebsiella, Xenorhabdus, especies de Bacillus, Staphylococcus, Streptococcus, Acinetobacter, y
Pseudomonas (Greenberg y Klowden, 1972; Demaio y cols., 1996; Thaler y cols., 1998;
McLemore y cols., 2000; González-Ceron y cols., 2003Zayed y Bream, 2004).
En la mariposa monarca se ha determinado algunos grupos de bacterias asociadas a los diferentes
estadios larvales bajo condiciones controladas de laboratorio, y se han encontrado bacterias
pertenecientes a las familias: Corynebacteriaceae, Propionibacteriaceae, Bacillaceae,
Lactobacillaceae Micrococcaceae, Azotobacteriaceae, Acromobacteriaceae y
Enterobacteriaceae (Kingsley, 1972). En dicho estudio a diferencia de los realizados en otros
artrópodos, no se encontró a Serratia spp, enterobacteria que ha demostrado ser entomopatógena
para varios lepidópteros y otros insectos en la fase de larva (Chadwick y cols., 1990; Glare y
cols., 1993; O’Callaghan y cols., 1996; Jackson y cols., 2001; Gatehouse y cols., 2008).
Las propiedades de virulencia de Serratia parecen estar relacionadas con la producción de
enzimas hidrolíticas como quitinasas y lipasas, que no sólo están presentes en estas bacterias,
también se han descrito estas actividades en diversos hongos y otras bacterias(Kirsh, 1934; Fu y
cols., 1997; Akhtar y cols., 1980). Como en todos los artrópodos, la mariposa monarca presenta
un exoesqueleto de quitina que le brinda soporte y protección, sí esta estructura fuese dañada
podría quedar a merced de bacterias u hongos con capacidad lipolítica ocasionándole la pérdida
de sus reservas energéticas, esto toma especial relevancia en los sitios de hibernación de la

8
mariposa monarca, ya que en esos sitios la mariposa no consume alimentos, únicamente
sobrevive con agua y las reservas almacenadas antes de la migración (Cenedella, 1971; Brown,
1974).
1.11 La microbiota bacteriana en los insectos.
El establecer las diferentes relaciones simbióticas que ocurren entre un hospedero y los
microorganismos asociados en un momento dado, permitiría determinar para el hospedero cuales
organismos podrían ser una amenaza a la salud, desgraciadamente sólo para algunos animales y
para el hombre es que estas relaciones se han investigado, al tal punto que existen criterios para
clasificar a los microorganismos como pertenecientes o no a la microbiota (Savage, 1977), por
otra parte en los insectos este tipo de relaciones únicamente se han investigado para algunos
organismos de este grupo (Dillon and Dillon, 2004), de tal manera que resulta complicado
interpretar correctamente lo que significa encontrar diferentes especies de microorganismos
asociadas en un lepidóptero como la mariposa monarca, sobre todo porque no se conocen los
factores involucrados en la permanencia de los microorganismos en las diferentes estructuras que
presenta este hexápodo en sitios como el abdomen, tórax, cabeza, patas o alas. Por otra parte, una
limitante importante en el cultivo de microorganismos de fuentes ambientales, es que sólo una
pequeña proporción se ha podido cultivar en medios sintéticos, con el creciente avance de las
técnicas moleculares se ha podido revelar la presencia de múltiples microorganismos simbióticos
en diversos insectos (Dasch y cols., 1984; Jiggins y cols., 2000; Fukatsu y cols., 2001;
Kittayapong y cols., 2002; Dunn y Stabb, 2005; Lindh y cols., 2005), sin embargo, la desventaja
de las pruebas moleculares es la incapacidad de investigar las interacciones, tanto positivas como
negativas entre el microorganismo y su hospedero. Dado que se desconoce el efecto y tipo de
microorganismos asociados con la mariposa monarca, es indispensable identificarlos y
estudiarlos.

9
2. Antecedentes.
El fenómeno de la migración de las mariposa monarca constituye uno de los eventos más
sorprendentes; actualmente las agencias gubernamentales y de conservación de EUA, Canadá y
México comparten esfuerzos para protegerla. Esto se ha facilitado con el establecimiento en
1996, del Comité Trilateral de Vida Silvestre Conservación y Manejo de Ecosistemas, cuyo
objetivo fue facilitar y fortalecer la cooperación y coordinación entre las agencias de vida
silvestre de los 3 países, así las tres naciones se han enfocado a la conservación del ciclo de la
mariposa monarca y sus diferentes hábitats.
En México, se creó la Reserva de la Biosfera Mariposa Monarca (RBMM) que depende del
Consejo Nacional de Áreas Naturales Protegidas (CONANP), y como una de sus funciones, se
encargó de estimar el número de mariposas que arriban durante la migración a los diferentes
sitios de hibernación, la estimación la realizó de manera indirecta al determinar la superficie de
ocupación total de las colonias de la Mariposa Monarca. Desde 1993 a la fecha se ha observado
una tendencia hacia la baja en el número de hectáreas ocupadas (Figura 2), (Rendón-Salinas y
cols., 2006).
Por otra parte, los factores ambientales, tales como nevadas o tormentas, en ocasiones han
causado mortandades de hasta el 80% de las mariposas, tal como sucedió durante la temporada de
hibernación 2001-2002, en donde dentro y fuera de la reserva, se registraron fenómenos de
mortandad masiva, que generaron una serie de controversias en cifras y llevaron a especular
sobre las causas que originaron este fenómeno. Aunque éstas no fueron investigadas del todo, se
atribuyó la muerte a factores climáticos, particularmente a las tormentas invernales que se
presentaron en el mes de enero (Brower y cols., 2004), ya que las bajas temperaturas registradas
en asociación con alta humedad relativa, puede causar la muerte de numerosos individuos por
congelación (Alonso-Mejía y cols., 1992), sin embargo en esta mortandad no se investigó el
papel de otros factores.
En 2001, la Procuraduría Federal de Protección al Medio Ambiente (PROFEPA) en conjunto con
varios grupos de investigación y de conservación, estableció el Grupo de Trabajo Técnico-

10
Científico Multidisciplinario para la Atención a Contingencias de la Mariposa Monarca. Este
grupo se creó como uno de los compromisos en materia de conservación acordado en las
reuniones del Comité Trilateral de Vida Silvestre Conservación y Manejo de Ecosistemas
México-EUA-Canadá, entre los objetivos del comité está: el aclarar las causas de muerte masiva
de la mariposa monarca, por lo que varios grupos enfocan su investigación hacia las amenazas de
la salud en la mariposa monarca, (CEC. 2008b).
México-EUA-Canadá.
Figura 2. Superficie de ocupación de la Mariposa Monarca en hectáreas desde 1993 a 2007 registradas en los santuarios de hibernación en México. Fuente (Rendón-Salinas, y cols., 2006)
El grupo de trabajo técnico-científico multidisciplinario para la atención a contingencias de la
Mariposa Monarca, se conformó con las siguientes instituciones:
PROFEPA: Órgano oficial coordinador del Grupo de Atención a Contingencias.

11
RBMM: Se encargó de la vigilancia en las áreas protegidas, así como del monitoreo de
mortandad y áreas de ocupación de la Mariposa Monarca, por otra parte participo en el enlace
entre las comunidades que realizan ecoturismo sustentable.
FMV-UNAM A través del Laboratorio de Toxicología, se dedicó a buscar sustancias toxicas en
las mariposas tales como metales, así como análisis de lípidos de las mariposas.
WWF México. World Wildlife Fund-Program. Organización no gubernamental (ONG), se
encargó de estimar áreas de ocupación de la mariposa monarca en colaboración con la RBMM
ESM-IPN: Escuela Superior de Medicina del Instituto Politécnico Nacional a través del
Laboratorio de Medicina de Conservación (LMC) laboratorio de referencia en vida silvestre de la
Procuraduría de Protección al Medio Ambiente, recibió la solicitud de analizar y determinar la
presencia de patógenos que impacten en la salud de la Mariposa Monarca.
Actualmente, sólo se ha reconocido como patógeno de la mariposa monarca un parasito
perteneciente a la familia neogregarinidae llamado Oprhyocystis electroscirrha cuyas formas de
resistencia son llamadas ooquistes que se encuentran en las escamas de la mariposa monarca.
Este parásito fue descrito por primera vez en 1970 en mariposa monarca y mariposa reina en
Florida (McLaughlin, 1970). En 1992, Leong determinó que las larvas de la mariposa monarca
son susceptibles a la infección con el ooquiste, ocasionándoles la muerte en altas dosis del
parásito. En 2000, Altizer reportó la prevalencia de este parasito en tres diferentes zonas de
hibernación y encuentró que en nuestro país sólo se presentó un 8% de parasitosis a diferencia de
California y Florida que la presentan en un 50% y 96% respectivamente. En 2001 Altizer propuso
que la distancia de migración juega un papel importante en la parasitosis.
Como se ha mencionado en trabajos previos la parasitosis en los sitios de hibernación de nuestro
país es baja (Altizer y cols., 2000) por otra parte, aun no hay evidencia contundente que indique
susceptibilidad del adulto a O. elektroscirrha, es decir aunque puede albergar los ooquistes, ésta
aparentemente no es afectada (Leong y cols., 1997; Bradley y cols., 2005).

12
Con estos datos se podría pensar que la mariposa solo es susceptible en los sitios de
reproducción, sin embargo en los sitios de hibernación de nuestro país se siguen reportando
muertes masivas de la mariposa monarca, esto ha llevado a cuestionar sí es causado por otros
patógenos.
Desde 2003, el LMC-IPN se enfocó a la tarea de identificar sí hay patógenos presentes en la
mariposa monarca. El dictaminar qué agentes microbianos pueden ser patógenos para la mariposa
monarca no es sencillo sobre todo porque no se conoce la microbiota comensal asociada al
organismo, de tal manera que no hay valores de referencia que permitan interpretar de manera
atinada los resultados, es importante mencionar que hay un trabajo donde se describen las
bacterias asociadas a la mariposa monarca, sin embargo estos estudios fueron realizados sólo en
larvas cultivadas en laboratorio (Kingsley, 1972), y por tanto no reflejan la microbiota en el
adulto libre.
La investigación de la microbiota asociada y presencia de patógenos en la mariposa monarca es
tópico central de este trabajo.

13
3. Justificación.
La mortandad masiva de una especie protegida, demanda conocer cuáles fueron las causas; esto
permitirá tomar medidas que eviten o disminuyan la frecuencia e intensidad de estos fenómenos.
Por otra parte, es indispensable la participación interdisciplinaria, ya que muchas veces las causas
que desencadenan estos fenómenos son multifactoriales. Uno de estos factores lo constituye los
agentes infecciosos que tienen un importante efecto en la salud de su hospedero pudiendo incluso
causarle la muerte.
Recientemente, se ha mostrado una disminución en la población de la mariposa monarca que
hiberna en nuestro país, este fenómeno se ha visto acompañado con un incremento en la
mortandad lo cual podría amenazar el ciclo biológico de esta mariposa. Diversos grupos de
investigación han enfocado sus objetivos a determinar las causas de este fenómeno, analizando
factores ambientales y humanos tales como el clima y el impacto de la tala en los bosques de
hibernación.
Dado que se desconoce el efecto y tipo de microorganismos asociados con la mariposa monarca,
es indispensable identificarlos y estudiarlos. Esto permitirá, por un lado, establecer los límites de
referencia útiles para realizar el diagnóstico microbiológico y por otra parte, la investigación
posterior de los factores de virulencia en las cepas candidatas a ser patógenas permitirá orientar
mejor el diagnostico microbiológico, enfocado a la búsqueda de patógenos. Finalmente, estos
resultados permitirán evaluar el impacto de los microorganismos en las poblaciones de la
mariposa monarca.

14
4. Objetivo General.
Estudiar la prevalencia de microorganismos patógenos en Danaus plexippus, así como identificar
posibles especies pertenecientes a la microbiota asociada a la mariposa en los santuarios de
hibernación El Rosario, Sierra Chincua, La Mesa y Cerro Palomas.
5. Objetivos Particulares.
Realizar un análisis parasitológico de improntas del abdomen obtenidas de mariposas vivas y
mariposas muertas para determinar la prevalencia de infección por Ophryocystis elektroscirrha
en los santuarios de hibernación El Rosario, Sierra Chincua, La Mesa y Cerro Palomas.
Aislar las especies cultivables de bacterias y hongos asociados a Danaus plexippus, tanto del
interior del abdomen y del cefalotórax como de la superficie de las alas.
Aislar las especies cultivables de bacterias de fuentes ambientales, tales como aire, agua suelo y
corteza de oyamel, en los sitios de hibernación de la mariposa monarca.
Correlacionar los hallazgos y proponer la microbiota asociada a la mariposa monarca, así como
los posible microbiota transitoria.

15
6. Materiales y Métodos
6.1 Muestreos.
Se realizaron 74 muestreos en los sitios de hibernación de la mariposa monarca, tales como Cerro
el Campanario en El Rosario (N19.59589, O100.26363, A3294msnm), y Sierra Chincua
(N19.6724, O100.30278, A3048), ubicados en Michoacán y la Mesa (N19.56580, O100.24131,
A3277) y Cerro las Palomas (N19.10072, O99.87134, A3391) en el Estado de México.
En El Rosario, La Mesa y el Cerro de Las Palomas los muestreos se realizaron de manera
periódica con intervalos de 15 a 30 días, se seleccionaron estos sitios con el objeto de comparar
parámetros en 4 colonias relativamente diferentes de la Mariposa Monarca, la elección se realizó
considerando que, en El Rosario y Sierra Chincua, las mariposas están expuestas en mayor grado
a la población humana, debido a las actividades de ecoturismo que realizan los pobladores de
manera sustentable; por otra parte, se eligió otra colonia cercana a ésta, ubicada en la Mesa, Edo.
de México, la cual comparte muchas de las características climáticas presentes en el Rosario,
excepto que casi no tiene contacto con poblaciones humanas. Finalmente se eligió un tercer sitio
alejado de los dos anteriores, el cual está ubicado a las faldas del volcán nevado de Toluca, en
este sitio las condiciones climáticas son diferentes a las que prevalecen en la Mesa y el Rosario
además de que en este sitio al igual que en la Mesa, la mariposa casi no tiene contacto con la
población humana. Por otra parte, es importante mencionar que todos los sitios muestreados
reciben mariposas de la región Este de los Estados Unidos y Canadá (Calvert y cols., 1986;
Malcolm, 1987).
6.2 Colecta de mariposas para estudios parasitológicos.
Los muestreos se realizaron entre las 8:00 y las 9:30 AM. Una vez que se llegó al borde de la
colonia, se instaló un campamento donde se colocó el material e instrumentos; el equipo técnico
se dividió en dos grupos, uno de ellos se encargó del análisis parasitológico en campo y otro se
encargó de la colecta de las mariposas.
Antes de colectar las mariposas, se realizó un reconocimiento del lugar, se seleccionaron 10
árboles en cada sitio muestreado, esto se realizó considerando los siguientes criterios: Árboles

16
cuyas ramas con mariposas fuesen accesibles; es decir, ramas a una altura máxima de 7 m por
otra parte se consideró que la distancia entre cada árbol muestreado fuera de al menos 15 m De
cada árbol se obtuvo su posición geográfica, y de una las ramas se colectaron las mariposas
presentes en ella, la cuales fueron capturadas con una red entomológica, adicionalmente, se
colectaron 10 mariposas muertas cercanas a cada árbol muestreado, el criterio para colectar estas
mariposas fue el siguiente: mariposas muertas que presenten el abdomen, alas y cefalotórax, sin
lesiones aparentes. Se descartaron las mariposas con el abdomen seco, mojado o con moho; las
mariposas se colectaron con guantes y posteriormente se guardaron en bolsas estériles
individuales. Después de cada colecta, las mariposas se entregaron al equipo encargado de
realizar el análisis parasitológico.
6.3 Colecta de mariposas para estudios bacteriológicos.
Los estudios bacteriológicos se realizaron con el propósito de determinar la diversidad bacteriana
presente en la mariposa monarca bajo alguna condición o variable característica, dichas variables
fueron las siguientes: mariposas vivas aparentemente sanas colectadas en árboles, mariposas
muertas colectadas en el suelo pero dentro de la colonia, mariposas previamente identificadas
como infectadas con O. elektroscirrha; del mismo modo se consideró como variable la fase de
hibernación pudiendo ser ésta: inicial, media o final; así mismo se consideró el sitio de
hibernación donde fue colectada la mariposa cuadro 1, muestra algunas de las características de
las mariposas incluidas en este trabajo.
6.4 Obtención de muestras de suelo, corteza, agua y aire.
Con objeto de relacionar los microorganismos encontrados en la mariposa con los
microorganismos presentes en el ambiente, se colectaron muestras de suelo y corteza de cada
árbol de donde se colectaron mariposas. Para obtener el suelo, se utilizó un pala pequeña y se
obtuvo una muestra de aproximadamente 500 g de rizosfera. Para colectar la corteza, se utilizó
una navaja con la cual se retiró una muestra de cuatro sitios diferentes por árbol, lo anterior, para
tener una muestra de aproximadamente 10 g. Tanto la corteza como el suelo se colocaron en
bolsas con cierre (Ziploc) y se transportaron al laboratorio. Por otra parte también se colectó una
muestra de 100 mL de agua de 3 sitios donde la mariposa se abastece (Figura 3).

17
Cuadro 1. Relación de las muestras obtenidas en los sitios de hibernación, temporada y fase de la hibernación. a Sitios con alta frecuencia de contacto con la población humana b Sitios con baja frecuencia de contacto con la población humana c Cantidad de mariposas vivas aparentemente saludables que se incluyeron en el estudio al cumplir los criterios descritos en materiales y métodos. d Cantidad de mariposas muertas colectadas e incluidas en el estudio e La fase inicial se considera desde 1 de noviembre al 15 de diciembre. f La fase media se consideró desde el 15 de diciembre al 31 de enero. g La fase final se consideró desde el 1 de febrero al 15 de marzo. * Mariposas parasitadas con O. elektroscirrha.
Figura 3. Sitios seleccionados para la colecta de agua. Se monitorearon varios sitios de abastecimiento de agua para la mariposa monarca fueron monitoreados con el fin de relacionar la microbiota en agua con la aislada en el lepidóptero, estos sitios corresponden a pequeños riachuelos cercanos a las colonias de hibernación. (fuente LMC, 2007)
Se realizó un monitoreo microbiológico del aire, empleando el procedimiento de placa expuesta,
que consiste en exponer la superficie de un medio de cultivo al aire durante un tiempo
determinado, en este caso los medios de cultivo se expusieron 5 min a una altura de 1.5 m, los
Temporada 2004-2005Fase inciale 10 al 12 Noviembre 5 / 5 2 / 2
10 al 12 Diciembre 5 / 5 2 / 2Fase mediaf 14 al 15 Enero 10 / 10 4 / 4 2 / 2Fase finalg 11 al 12 Febrero 10 / 10 5 / 5
Temporada 2005-2006Fase inicial 2 al 8 Diciembre 3 / 3 2 / 2 3 / 3Fase media 3 al 10 Enero 2 / 3 3 / 3 3 / 3Fase final 28 febrero al 7 Marzo 2 / 3 2 / 3 1 / 1
Temporada 2006-2007Fase media 9 de Enero 30 / 30
Fase final 9 de Febrero 15* / 15*77 / 77 11 / 11 7 / 7 11 / 11
Etapa de la hibernación
Total
Sitio de hibernación
Mariposas vivasc / mariposas muertasd
Fecha de colecta La mesab Cerro Palomasb Sierra chincuaa El Rosarioa

18
medios que se emplearon fueron los mismos que se utilizaron para el aislamiento de bacterias
asociadas a la mariposa monarca (ver mas adelante).
6.5 Análisis parasitológico.
6.5.1 Obtención de improntas de abdomen para análisis parasitoscópico.
Para el análisis parasitoscópico se siguió el protocolo descrito por Leong en 1992, que consiste en
sujetar con los dedos la mariposa controlando el movimiento de las alas, y presionar el abdomen
de la mariposa con la superficie adhesiva de un trozo de cinta auto adherente “diurex”, este
procedimiento se realizó con cuidado para no lastimar a la mariposa, y obtener una buena
impronta del abdomen, posteriormente la cinta fue adherida a un portaobjeto limpio y
desengrasado, previamente identificado con los datos de la ubicación y localización de la
monarca (Figura 4), después de realizar la impronta, ésta fue colocada en un recipiente, mientras
se determinó la presencia de ooquistes de Ophryocystis elektroscirrha mediante la observación de
la impronta en un microscopio estereoscópico. Las mariposas positivas a este parasito fueron
colectadas y llevadas al laboratorio, en el caso contrario las mariposas fueron liberadas.
Figura 4. Procedimiento para realizar las improntas de escamas del abdomen de la mariposa. Se debe sujetar por las alas a la mariposa, y presionar el abdomen con la parte adhesiva de un trozo de cinta transparente, esta última debe pegarse sobre un portaobjetos perfectamente limpio y desengrasado. El número de improntas por portaobjeto puede ser de 1 a 6. (fuente LMC, 2004)
6.5.2 Búsqueda de Ophryocystis elektroscirrha en improntas de abdomen de mariposas.
Para el análisis parasitoscópico, las improntas se observaron en un microscopio de campo claro
(Nikon) con el objetivo de seco débil (10x), y seco fuerte (40x) a fin de buscar estructuras de
forma ovoide, de color café aproximadamente de 14 µm de largo por 9 µm de diámetro, cada
impronta se revisó por completo cuantificando los ooquistes y se determinó en cada una, el nivel

19
de infección en una escala equivalente a la descrita por Altizer 1999, pero con una modificación,
de acuerdo con lo siguiente:
Negativo cuando no se observan ooquistes en la impronta.
Positivo Nivel bajo hasta 100 ooquistes en toda la impronta
Positivo Nivel alto más de 100 ooquistes en la impronta
Las improntas que resultaron positivas se identificaron y se guardaron en un lugar seco y fresco.
6.6 Transporte de muestras.
Después del análisis parasitoscópico en campo, se seleccionaron 10 mariposas vivas sanas de
cada árbol, las cuales se colocaron en bolsas individuales para ser transportadas al laboratorio,
donde fueron mantenidas en refrigeración hasta su análisis bacteriológico y micológico.
6.7 Análisis bacteriológico y micológico.
6.7.1 Procedimiento de elección de medio de cultivo.
Se siguió el protocolo descrito por Kingsley en 1972, para seleccionar el medio de cultivo, este se
basó en determinar el número de colonias recuperadas en varios medios de cultivo incluyendo el
agar sangre, agar soya tripticaseína, agar nutritivo, y 7 diferentes modificaciones de agar caseína
peptona almidón, en los cuales se adicionó alguno o algunos de los siguientes componentes:
sangre de carnero, extracto de levadura (Difco), triturado de mariposa monarca fresca y estéril;
por otra parte se probaron los siguientes medios para hongos: agar papa dextrosa, agar rosa de
bengala, agar de maltosa sabouraud y agar harina de maíz. Para este fin, una muestra de 10
mariposas sanas vivas se sacrificaron, se retiraron las alas y el cuerpo se sumergió en alcohol al
70% durante 1 min, posteriormente se realizaron 3 lavados de 30 s en agua estéril, finalmente el
abdomen se separó del cefalotórax, y ambas muestras fueron trituradas por separado en un
mortero de porcelana estéril; cada muestra fue homogeneizada con PBS estéril y fue colocada en
un volumen de 10 mL, a partir de esto se realizaron diluciones decimales desde 10-3 a 10-7. Por
otra parte, las alas fueron introducidas en un tubo con 10 mL de PBS, después se
homogeneizaron y se diluyeron igual que las muestras de abdomen y cefalotórax. De cada
dilución se tomó una alícuota de 0.1mL con la cual se inoculó por duplicado cada uno de los 10
medios de cultivo para bacterias y 4 para hongos indicados arriba. El triturado de mariposa se

20
realizó con 10 g de abdómenes frescos de mariposa monarca, que fueron homogeneizados con 90
mL de regulador salino de fosfatos, la mezcla fue diluida a 1000 mL y posteriormente esterilizada
a 121ºC/15 libras por pulgada cuadrada durante 15 min. Se seleccionaron los medios que
mostraron los mejores resultados en cuanto a diversidad recuperada y carga microbiana
encontradas. Los medios seleccionados fueron: Agar Caseína Peptona Almidón Monarca,
ACPAM (1.5% agar Difco, 0.5% caseína Difco, 0.5% almidón Difco, 0.2 g/L de K2HPO4 JT
Baker, 0.05 g/L de MgSO4.7H2O JT Baker, 0.004 g/L de FeCl3 Merck, 0.1% glicerol Merck,
0.1% de extracto de levadura Difco, 0.1% v/v triturado fresco del abdomen de mariposa monarca)
y agar sangre tripticaseína levadura monarca, ASTLM (4% Base agar soya tripticaseína Difco,
0.1% extracto de levadura Difco, 0.1% v/v triturado fresco del abdomen de mariposa monarca).
6.7.2 Selección de mariposas para el análisis.
Para estudios bacteriológicos sólo se seleccionaron las mariposas que presentaron un aspecto
saludable, sin presencia de manchas, decoloración o ruptura de alas, así mismo se incluyeron solo
mariposas cuyo peso se encontró entre 0.5 g ± 0.05 g. Con objeto de evitar la predisposición,
todas las muestras se trabajaron a doble ciego de tal manera que todas las mariposas se trabajaron
de igual manera independientemente de la condición presentada.
6.7.3 Cultivo y procesamiento de las muestras.
Cada mariposa se analizó de manera independiente, primeramente se colocó en refrigeración a
4ºC durante 30 min; posteriormente se obtuvo su peso en una balanza digital de precisión,
aquellas mariposa que cumplieron con los criterios arriba señaladas se incluyeron en el estudio.
Con objeto de evitar al máximo el sufrimiento de las mariposas, éstas se sacrificaron rápidamente
presionando firmemente la cabeza con unas pinzas estériles, las alas fueron separadas, y
homogeneizadas con PBS. El resto de la mariposa se sanitizó en etanol 70% v/v durante 30 s,
posteriormente se realizaron 4 lavados en agua desionizada estéril, el cuerpo se seccionó en tres
partes: abdomen, cefalotórax y un segmento de 2 mm que une ambos sitios anatómicos, este
último se descartó del análisis ya que contiene microorganismos de ambas partes de la mariposa.
Cada corte se homogenizó en un mortero de porcelana, utilizando PBS, a partir de esta
suspensión se realizaron diluciones con las cuales se determinó la carga microbiana de cada tipo

21
de bacteria aislada, los cultivos se realizaron en los medios de cultivo seleccionados de acuerdo
con lo antes mencionado.
6.8 Análisis bacteriológico de aire, corteza, suelo, agua.
Para estos análisis se pesaron 10 g de suelo, corteza, o 10 ml de agua y se colocaron en 90 ml de
PBS estéril, se homogenizó la muestra y se efectuaron diluciones decimales hasta 10-6, a partir de
las últimas 4 diluciones se colocó 0.1 ml sobre cada uno de los medios previamente
seleccionados.
6.9 Condiciones de incubación.
Una de las dos series se incubó en atmósfera anaerobia (sistema generador de anaerobiosis
Gaspack, Becton & Dickinson), a fin de aislar bacterias anaerobias obligadas, la otra se mantuvo
en condiciones aerobias al igual que la serie de medios para el aislamiento de hongos. Todas las
placas se mantuvieron a 20ºC durante 7 días en caso de bacterias y de 7 a 21 días para los hongos.
6.10 Aislamiento y construcción de ceparios.
Cada cepa diferente recuperada en los medios de cultivo se aisló y contabilizó, lo anterior con
objeto de determinar la carga bacteriana cultivable. Las colonias recuperadas se resembraron en
agar sangre y agar caseína peptona almidón suplementado con glicerol, y se mantuvieron en
refrigeración hasta su identificación fenotípica. Alternativamente algunas de las cepas se
conservaron mediante el sistema Microbank® siguiendo los procedimientos y recomendaciones
especificados por el fabricante.
6.11 Identificación de los microorganismos aislados.
La identificación se realizó mediante análisis de taxonomía numérica, empleando como
resultados positivos o negativos, la capacidad o no de utilizar diversos sustratos (Sokal, 1962;
Holder-Franklin y cols., 1992), este procedimiento se llevó a cabo en dos etapas, en la primera de
ellas se ejecutaron una serie de pruebas básicas con las que se ubicó cada aislado en un grupo,
familia ó genero, las pruebas que se emplearon se observan en el cuadro 2.

22
Cuadro 2. Pruebas preliminares para la identificación presuntiva de bacterias aisladas.
Morfología microscópica (tinción) Tinción de Gram
Morfología microscópica (cultivo líquido) Tinción de Ziehl Neelsen
Movilidad en medio semisólido de nitratos Tinción de Shaeffer y Fulton para esporas
Crecimiento anaerobio Glucosa en medio CTA
Crecimiento aerobio Tipo de metabolismo OF
Oxidasa Morfología colonial en agar sangre
Catalasa Crecimiento en medio Mac Conkey
Posteriormente, cada cepa fue sometida a 95 ensayos fenotípicos empleando pruebas
morfológicas, Las pruebas incluyeron: características morfológicas en agar sangre, Müller
Hinton, arreglo microscópico, afinidad tintorial (Gram y Ziehl Neelsen) movilidad (medio SIM
Difco), movilidad por microscopia de contraste de fases, ambas a 37 y a 22ºC, presencia, forma y
ubicación de esporas (tinción de Shaeffer and Fulton); así mismo, en cada cepa se determinó la
actividad hidrolítica sobre almidón, caseína, lecitina, DNA, esculina, gelatina, tirosina, y urea
(Difco); también la actividad catalasa, oxidasa (Difco), tipo de hemólisis en base agar sangre
(Difco) con 5% de sangre de carnero, lisina descarboxilasa, ornitina descaboxilasa y arginina
dihidrolasa en Base Møller descarboxilasa (Difco) con y sin 1% de NaCl, fenilalanina
desaminasa (BBL) reducción de nitratos a nitritos (Medio Nitrato-movilidad, Difco); producción
de ácido a partir de los siguientes carbohidratos: glucosa, lactosa, sacarosa, manitol, xilosa
fructosa, maltosa, sorbitol, manosa, melobiosa, inositol, dulcitol, rafinosa, arabinosa, galactosa,
almidón (medio base rojo fenol y medio CTA (BBL) con y sin 1.0% de NaCl); producción de:
H2S (medios SIM, Kligler (BBL) y tiras de papel con acetato de plomo (Difco), indol (SIM,
caldo tripticasa nitrato BBL); crecimiento en: Mac Conkey (BBL), agar tiosulfato citrato sales
biliares TCBS (BBL), agar sal y manitol SM, caldo malonato (Difco), medio citrato de Simons
(BBL), NaCl al 0, 1, 6, 6.5, 8, 10 y 12% (base caldo nutritivo BBL); También se determinó la
aerotolerancia en medio de sangre en condiciones aerobias y anaerobias (Sistema generador de
anaerobiosis Gaspack Becton and Dickinson); así como el tipo de metabolismo: fermentativo,
oxidativo o inerte (Medio OF BBL), la producción de acetilmetil carbinol y la prueba del rojo de
fenol (medio VP/RM BBL); se realizaron pruebas serológicas incluyendo diversos antisueros

23
(Difco). Para todas las pruebas anteriores se siguieron las recomendaciones y criterios de
interpretación correspondiente (Mac Faddin, 1985).
6.12 Cepas de referencia.
Para validar los resultados de las pruebas anteriores, así como fundamentar el analisis de
taxonomía numerica, se emplearon cepas de referencia, la selección de estas se basó en el tipo de
microorganismos que usualmente se espera recuperar en cultivos de bacterias asociadas a
insectos. Las cepas fueron obtenidas del American Type Culture Collection: Escherichia coli
ATCC 25922, Enterobacter aerogenes ATCC 29008, Klebsiella pneumoniae ATCC 10031,
Salmonella enterica subespecie enterica serovar typhimurium ATCC 13311, Salmonella enterica
subespecie enterica serovar choleraesuis ATCC 10708, Salmonella enterica subespecie enterica
serovar typhi ATCC 7251, Stenotrophomonas maltophila ATCC 13637, Pseudomnas aeroginosa
ATCC 9027, Staphylococcus aureus ATCC 25923. Así mismo se incluyeron cepas control
utilizadas para realizar evaluaciones externas de calidad en bacteriología, dichas cepas
previamente fueron caracterizadas con pruebas fisiológicas y bioquímicas. Las cepas fueron
amablemente donadas por el Programa de Aseguramiento de la Calidad (PACAL):Vibrio
cholerae No O1 PECEL 0110, Vibrio parahaemolyticus PECEL 9912, Aeromonas hydrophila
PECEL 0103, Acinetobacter calcoaceticus PECEL 0105, Moraxella catarrhalis PACAL 0503,
Plesiomonas shigelloides PACAL 0212, Enterobacter cloacae PACAL 0505, Serratia
marcescens PECEL 0005, Citrobacter freundii PECEL 0004 Bacillus Cereus PECEL 9911,
Streptococcus mitis PACAL 0302, Streptococcus agalactiae PECEL 0011, Lactobacillus sp
PACAL 0210 Enterococcus feacalis PACAL 0504 Listeria monocytogenes 0607. Finalmente, se
incluyeron 4 cepas obtenidas de mariposas monarca, (Zepeda y cols., 2005): Pasteurella sp LMC
15, Micrococcus sp LMC23, Corynebacterium sp LMC93, Kurthia sp LMC56; las cepas
anteriores se identificaron utilizando métodos automatizados (BD Phoenix, Becton Dickinson
Diagnostic Systems, Sparks, Md. USA, así como Vitek Systems, bioMérieux Hazelton, Mo,
USA), sólo se consideró la identificación a nivel de género debido a que estos sistemas
fundamentan la identificación en patrones bioquímicos de aislados clínicos, no de aislados de
insectos.

24
6.13 Identificación de géneros y especies.
Con los resultados de las pruebas fenotípicas se determinó la similitud entre los aislados mediante
el coeficiente simple de similitud (CSS). Los resultados positivos y negativos fueron considerados
como 1 y 0, respectivamente. Con los resultados se construyó una base de datos en Microsoft
Excel R y a partir de ella se exportaron datos al programa MEGA 3.0 donde fueron agrupadas las
cepas, en un dendograma basado en el Método UPGMA (unweighted pair-group method of
association), (Rohlf and Sokal, 1981; Sneath, 2005), los patrones obtenidos con las cepas control
y de referencia permitieron agrupar a la cepas problema dentro de alguna rama, dichas ramas se
consideraron como grupos fenotípicos, sí entre ellas mostraron de un 80% a 95% de similitud.
Los patrones bioquímicos de las cepas de cada grupo fenotípico obtenido se compararon con los
patrones de cepas reportados en el Bergey’s Manual of Systematic Bacteriology, utilizando el
CSS, (Krieg y cols 1984; Brenner y cols 2005). Se consideró que una cepa pertenece a una especie
sí: es la que presenta mayor similitud con alguna de las especies reportadas y si el valor de esta
fue mayor de 85%; en aquellos casos en los que la similitud se encontró entre el 80 y 85%, éstas
solo fueron consideradas como especies atípicas, por otra, parte valores entre el 70 y 80% solo se
identificaron como pertenecientes al género.
Los medios de cultivo que se emplearon par las pruebas fenotípicas se prepararon en el
laboratorio, a partir de medios deshidratados (Difco, BBL y Bioxon) siguiendo las
recomendaciones del fabricante. La preparación se realizó en tubos de 10x100 mm. Para asegurar
la calidad en los resultados, los medios se elaboraron 4 días antes de ser inoculados, y
adicionalmente cada medio de cultivo fue probado con las cepas de referencia antes indicadas,
para evaluar las reacciones esperadas, solo se utilizaron los medios de cultivo que mostraron las
reacciones esperadas.
La inoculación se realizó a partir de una colonia procedente de agar sangre o algún medio
enriquecido, ésta fue suspendida en 2 ml de solución salina fisiológica, con dicha suspensión se
inoculó cada uno de los medios de cultivo seleccionados, la incubación se realizó a 35.5ºC ±1°C,
excepto las pruebas de hidrólisis que se realizaron a 20ºC y de movilidad a 22 ºC, los resultados
se registraron después del periodo recomendado para cada prueba, variando desde 18 h para el
TSI a 10 días para las pruebas de hidrólisis de caseína, tirosina y almidón.

25
6.14 Análisis Estadístico.
Para analizar los resultados sobre la prevalencia de parasitosis en hembras, machos, mariposas
vivas o muertas, así como sí hay diferencia en los santuarios o temporada de hibernación se
realizó un análisis de varianza (ANOVA) bilateral por bloques. Este análisis permitió establecer
en que grado afecta la varianza individual de cada variable en la varianza total del parámetro
analizado (Daniel, 2006). Se utilizó la prueba F para evaluar diferencia en la variación.
Se empleó la prueba de Mann Whitney para determinar diferencias entre las medianas de la carga
microbiana presente en cada sitio anatómico. Se utilizó estadística no paramétrica para probar la
hipótesis de dependencia entre la presencia de las especies aisladas con: el sitio anatómico, la
temporada de hibernación y el sitio de hibernación para tal fin se realizó la X2 como prueba
estadística, considerando P<0.05 como significativo. Todos los cálculos estadísticos fueron
realizados utilizando las herramientas estadísticas del paquete SigmaPlot 11.0 (Systat Software,
Inc 2008).

26
7. Resultados.
7.1 Análisis Parasitológico.
El análisis parasitológico se realizó para determinar la prevalencia de Ophryocystis
elektroscirrha, dado que este es un microorganismo patógeno frecuentemente encontrado en la
mariposa monarca, los resultados confirmaron la presencia del ooquiste del parásito en varias
mariposas, dichas estructuras presentaron las descripciones características, tales como: la forma
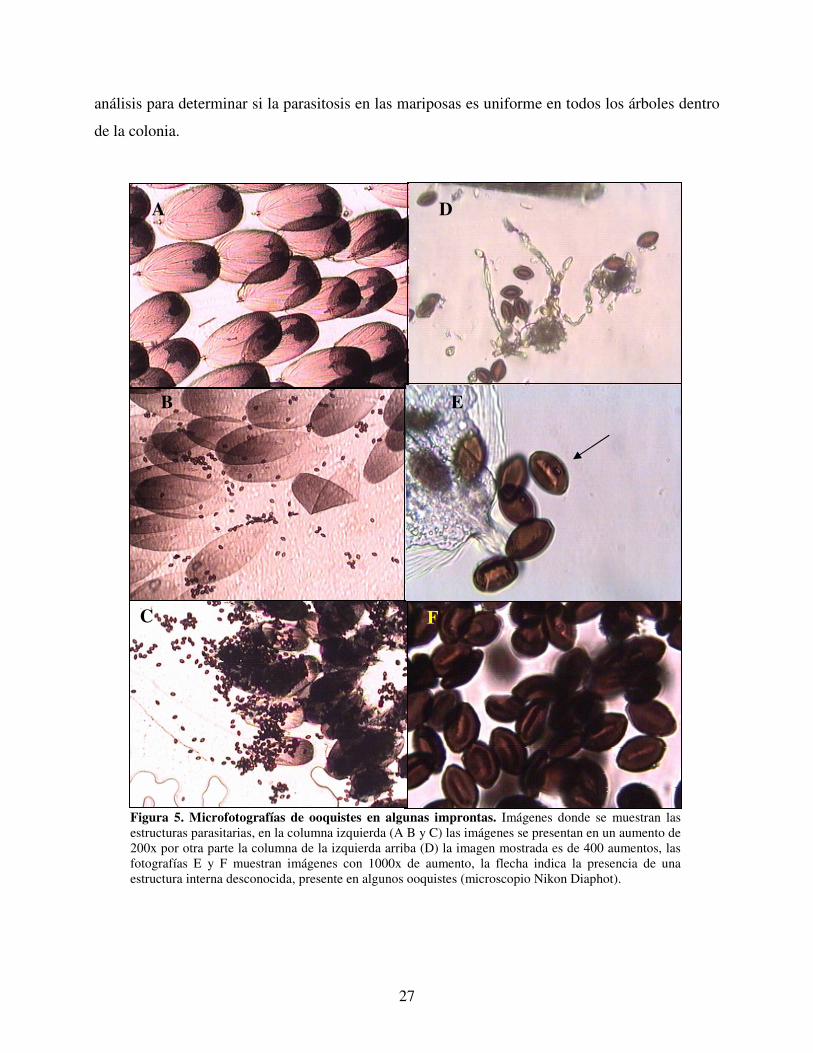
ovoide, el color café y las dimensiones esperadas, 14 µm de largo por 9 µm de diámetro, (Figura
5).
Las mariposas que presentaron infección con Ophryocystis elektroscirrha, no presentaron
diferencias notables en la apariencia física al compararlas con las negativas a este parásito. Las
mariposas no parasitadas presentaron escamas como las que se muestra en la figura 5A; las
mariposas infectadas mostraron diferente nivel de infección, desde aquellas con pocos ooquistes
hasta otros totalmente saturadas con las estructuras parasitarias (Figuras 5B y 5C).
En algunos casos se observaron estructuras aparentemente fúngicas, así como polen, asociado
con los ooquistes, (Figura 5D), también se observaron estructuras desconocidas dentro de algunos
de los ooquistes, (Figura 5E).
La ubicación de la mayoría de estructuras parasitarias fue encontrada sin mostrar algún patrón
característico en las escamas, sin embargo en algunas de las improntas se pudo observar
claramente un patrón de adherencia o asociación sobre la base de la escama, es decir el sitio de
unión de la escama a la mariposa figura 6.
7.1.1 Análisis de la distribución de árboles con mariposas infectadas.
Todas las mariposas que se analizaron en este trabajo se colectaron en ramas de árboles de
oyamel; para este fin se seleccionaron árboles tanto de la periferia de la colonia como dentro de
ella, los resultados mostraron variación en cuanto al porcentaje de mariposas infectadas con
Ophryocystis elektroscirrha, Debido a que se desconoce si la parasitosis de la mariposa monarca
en los sitios de hibernación presenta o no una distribución característica, se procedió a realizar un
27
análisis para determinar si la parasitosis en las mariposas es uniforme en todos los árboles dentro
de la colonia.
Figura 5. Microfotografías de ooquistes en algunas improntas. Imágenes donde se muestran las estructuras parasitarias, en la columna izquierda (A B y C) las imágenes se presentan en un aumento de 200x por otra parte la columna de la izquierda arriba (D) la imagen mostrada es de 400 aumentos, las fotografías E y F muestran imágenes con 1000x de aumento, la flecha indica la presencia de una estructura interna desconocida, presente en algunos ooquistes (microscopio Nikon Diaphot).
A D
B E
C F

28
Figura 6. Patrones de adherencia ó asociación de Ophryocystis elektroscirrha con las escamas de la mariposa monarca. Se puede observar la asociación del ooquiste al sitio de unión de la escama a la mariposa.
Para determinar el tipo de distribución de mariposas infectadas en los árboles dentro de una
colonia de hibernación, se colectaron mariposas a partir de 117 árboles en el periodo 2005-2006,
88 árboles en el ciclo 2006-2007 y 51 en el ciclo 2007-2008; para todos los árboles, se determinó
el porcentaje de mariposas altamente parasitadas, y con estos resultados se construyeron las
gráficas de la figura 7, que muestra la frecuencia de árboles con diferentes intervalos de
mariposas parasitadas.
Debido a que en los resultados mostrados en la figura 7 se observa variación en la parasitosis, se
procedió a determinar si esta variación es homogénea en las diferentes colonias de hibernación o
bien si la parasitosis es mayor en árboles orientados hacia algún punto cardinal o bien dentro o en
la periferia de la colonia.
Para determinar homogeneidad en toda la colonia se colectaron mariposas de árboles distribuidos
en toda la colonia y se formaron cuatro grupos de acuerdo a la ubicación, así se obtuvieron
árboles ubicados en la región noroeste, noreste, sureste y suroeste de la colonia; de cada árbol se
determinó el porcentaje de mariposas infectadas y con estos resultados se construyó el cuadro 3.
Estos datos fueron evaluados con la prueba de X2, para homogeneidad. Se planteó la hipótesis
nula que propone que las cuatro regiones de árboles con mariposas infectadas muestran
homogeneidad en la parasitosis. Los resultados de la evaluación estadística aceptaron la hipótesis
nula, por tanto se acepta homogeneidad para esta variable (α = 0.05).
A B

29
Figura 7. Distribución de la frecuencia de parasitosis de mariposa monarca. De cada árbol se colectó una muestra de entre 100 y 150 mariposas, las cuales fueron analizadas para obtener el porcentaje de parasitosis, la distribución de parasitosis encontrada en los árboles aparentemente corresponde al tipo normal, A) Monitoreo realizado de 2005 a 2006 considerando 117 árboles, B) monitoreo realizado de 2006-2007 donde se incluyen 88 árboles diferente, C) monitoreo realizado en la temporada de hibernación 2007-2008 en donde se incluyeron 51 árboles.
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15Marca de clase ( % parasitosis )
No. de á
rbole
s
0
5
10
15
20
25
30
35
1 3 5 7 9 11 13 15
Marca de clase ( % parasitosis )
No
. d
e á
rbo
les
0
3
6
9
12
15
18
21
1 3 5 7 9 11 13 15
Marca de clase (% de parasitosis)
No
de á
rbo
les
A
B
C

30
Cuadro 3. Modelo de distribución de árboles con mariposas infectadas, considerando 4 regiones en la
temporada de hibernación 2006-2007.
No hay relación entre los porcentajes de parasitosis y la ubicación de las colonias, por lo tanto se acepta que la distribución de mariposas parasitadas en los árboles es homogénea, (α = 0.05).
Sin embargo, debido a que los resultados anteriores dividieron la colonia en cuatro partes, es
posible que se pueda incurrir en un error tipo 2, es decir aceptar una hipótesis nula cuando esta es
falsa, lo anterior debido a que si hubiera mayor o menor parasitosis al centro de la colonia, esta
no sería homogénea, pero al dividir en 4 regiones el sitio de hibernación cada región incluiría una
parte del centro de la colonia por lo tanto al evaluar mediante la prueba de homogeneidad se
aceptaría la hipótesis nula.
Con el fin de corroborar nuestros resultados y evitar incurrir en el error tipo 2, adicionalmente se
evaluó si habría homogeneidad considerando tanto la periferia como el centro, para lo cual se
realizaron monitoreos en árboles ubicados en la periferia de la colonia al norte, sur, este y oeste, y
en el centro de la colonia (cuadro 4).
Cuadro 4. Modelo de distribución de árboles con mariposas infectadas, considerando la periferia y el centro de la colonia durante 2007 a 2008.
hasta 6% mas de 6 hasta 9% mas de 9 hasta 12% mas de 12%Norte 0 1 7 1 1 10Sur 0 2 6 2 1 11Este 0 1 6 3 1 11Oeste 0 1 4 4 1 10Centro 0 1 3 2 3 9Total 0 6 26 12 7 51
Región de la colonia
Número de arboles monitoreados
negativosArboles que donde se encontraron mariposas con O. elektroscirrha
Total
No hay relación entre los porcentajes de parasitosis y la ubicación de las mariposas en los árboles, por lo tanto se acepta que la distribución de mariposas parasitadas en el centro y periferia de la colonia es homogénea, (α = 0.05).
Estos resultados confirman que la distribución de parasitosis es homogénea en toda la colonia,
por otra parte en estos resultados es claro observar tanto en la figura 7 como en los cuadros 3 y 4,
que la parasitosis de la mariposa monarca con Ophryocystis elektroscirrha, aumentó a lo largo de
este estudio.
menos de 4% de 4 a menos de 6% de 6 a menos de 8% mas de 8%Noroeste 13 15 13 3 1 45Suroeste 26 25 11 5 2 69Noreste 6 18 10 4 4 42Sureste 10 7 10 4 0 31Total 55 65 44 16 7 187
Región de la colonia
Número de árboles monitoreados
negativosÁrboles que donde se encontraron mariposas con O. elektroscirrha Total

31
7.1.2 Variación de la prevalencia de infección con Ophryocystis elektroscirrhae
Se analizaron mariposas provenientes de los últimos 5 años durante el periodo de hibernación en
4 santuarios. Claramente se observa un aumento en la prevalencia (Figura 8).
Figura 8. Prevalencia de parasitosis con Ophryocystis elektroscirrhae desde 2003 a 2008. Se observa un aumento sostenido en la parasitosis media entre la población de mariposa monarca en hibernación. El análisis de varianza ANOVA muestra diferencia entre las 5 temporadas de hibernación (R.V=55.27 > 19 α=0.05), las barras muestran el intervalo de confianza del 95%.* Medias cuya diferencia fue tan alta como para ser considerada producto del azar, comparación realizada entre pares utilizando la prueba de Tukey ( α =0.05).
7.1.3 Comparación de la prevalencia entre diferentes sitios de hibernación.
El análisis de la parasitosis en cada sitio de hibernación se observa en las figura 9 y 10; en las
cuales se puede observar que aparentemente la diferencia entre la parasitosis media en cada sitio
de hibernación no fue suficientemente diferente como para ser producido por el azar (R.V.
=0.45<19 . α=0.05) Para realizar esta comparación se consideró el análisis ANOVA por bloques
o bilateral, conformando como bloques cada uno de las temporadas de hibernación.
0%
2%
4%
6%
8%
10%
12%
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
pa
ras
ito
sis
me
dia
(%
)
**
*

32
Figura 9. Variación en prevalencia de parasitosis de la mariposa monarca en los sitios de hibernación. No hay diferencia significativa en la media de parasitosis entre los sitios de hibernación de la mariposa monarca. Se incluyeron todas las temporadas, considerando un análisis ANOVA por bloques considerando la temporada de hibernación, (R.V=0.45 < 19 α=0.05), las barras muestran el intervalo de confianza del 95%. Figura 10. Variación en prevalencia de parasitosis de la mariposa monarca por temporada en cada sitio de hibernación. No hay diferencia significativa en la media de parasitosis entre los sitios de hibernación de la mariposa monarca., las barras muestran la desviación estándar.
0%
2%
4%
6%
8%
10%
El Rosario Sierra chincua Palomas La mesa
Sitio de hibernación
para
sitosis
media
(%
)
0%
2%
4%
6%
8%
10%
12%
14%
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
Para
sitosis
media
(%
)
El Rosario Sierra chincua Palomas La mesa

33
7.1.4 Comparación de la parasitosis entre mariposas muertas y vivas, así como entre
hembras y machos.
Se realizaron muestreos en cuatro sitios con intervalos de un mes esto se realizó durante los dos
primeros años del estudio posteriormente se aumentó la frecuencia a 15 días con el fin de
determinar variaciones en intervalos de tiempo menor.
La figura 11A muestra las diferencias en parasitosis entre mariposas vivas y muertas; cada punto
representa un monitoreo, el cual usualmente se realizó aproximadamente a 500 mariposas
provenientes de 5 a 10 árboles, en esta figura se conglomeraron todos los resultados sin importar
el sexo y/o lugar de hibernación de la mariposa, asimismo fue construida con el análisis
parasitoscópico de 36,300 mariposas provenientes de 74 muestreos realizados desde noviembre
de 2003 a Marzo de 2008 que incluyen 29550 improntas de mariposas vivas y 6750 improntas
realizadas con mariposas muertas. En estos resultados se aprecia también el aumento de la
prevalencia de parasitosis de un 2.3 a 9.3%. Aunque el análisis de varianza no muestra diferencia
en la frecuencia media de parasitosis en mariposas vivas y muertas, se observa una tendencia
hacia el incremento de la frecuencia de parásitos en las mariposas.
Los resultados de la figura 11B muestran los porcentajes de parasitosis obtenidos en cada
temporada tanto en mariposas vivas como muertas. El análisis de varianza muestra que no hay
diferencia entre la parasitosis media encontrada en mariposas vivas colectadas de árboles y las
mariposas muertas colectadas en el suelo. Para este análisis se consideró como bloques a las
temporadas de hibernación (R. V. = 0.43 < 9.29 α=0.05)
El análisis de los resultados anteriores, pero, considerando el sexo de las mariposas, se presenta
en la figura 12, en ella, además de apreciar la tendencia al aumento de la parasitosis, se observa
aparentemente una mayor prevalencia en machos que en hembras. Sin embargo el análisis de
varianza ANOVA muestra que la variación en la parasitosis entre hembras y machos podría ser
causa del azar (R. V. = 0.43 < 9.29 α=0.05). En la figura 13 se incluye el análisis de parasitosis
encontrado en mariposas muertas considerando el sexo de la mariposa, en esta figura también se
confirma que aparentemente no hay diferencia estadística entre la frecuencia de parasitosis en
hembras y machos.

34
A
B
Figura 11. Parasitosis media desde 2003 a 2008 en mariposas vivas y mariposas muertas. A) Los resultados muestran el porcentaje de mariposas con ooquistes, cada punto representa un monitoreo en el que aproximadamente se analizaron 500 improntas provenientes de 5 a 10 árboles. B) No se encontró diferencia estadísticamente significativa, las temporadas fueron incluidas como variable de bloque, (R.V=0.43 < 9.29 α=0.05), las barras muestran el intervalos de confianza del 95%.
0%
5%
10%
15%
20%
25%
28/06/2003 14/01/2004 01/08/2004 17/02/2005 05/09/2005 24/03/2006 10/10/2006 28/04/2007 14/11/2007 01/06/2008
Fecha
Para
sit
osis
med
ia (
%)
vivas muertas
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
0%
2%
4%
6%
8%
10%
12%
14%
16%
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
Para
sit
osis
me
dia
(%
)
vivas muertas

35
A
B Figura 12. Diferencia en la prevalencia de parasitosis entre machos y hembras vivas. A) Los resultados muestran el comportamiento en cada uno de los monitoreos. B) No se encontró diferencia estadísticamente significativa entre la parasitosis en mariposas hembras y machos. Las temporadas fueron incluidas como variable de bloque, (R.V=0.43 < 9.29 α=0.05), las barras muestran el intervalos de confianza del 95%.
0%
5%
10%
15%
20%
25%
28/06/2003 14/01/2004 01/08/2004 17/02/2005 05/09/2005 24/03/2006 10/10/2006 28/04/2007 14/11/2007 01/06/2008
Fecha
Para
sitosis
media
(%
)hembras machos
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
0%
2%
4%
6%
8%
10%
12%
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación
Pa
rasit
os
is m
ed
ia (
%)
Hembras Machos

36
Figura 13. Diferencia en la presencia de parásitos en mariposas muertas. Se observó que la variación en la presencia de parásitos en mariposas muertas hembras y machos no fue estadísticamente significativa como para considerar que no es producto del azar Las temporadas fueron incluidas como variable de bloque, (R.V=0.43 < 9.29 α=0.05).
7.1.5 Influencia de las heladas en la prevalencia de O. elektroscirrha.
Para determinar si los cambios en la temperatura, afectan o incrementan la mortalidad de las
mariposas parasitadas, se realizó un experimento que consistió en un monitoreo de mariposas
tanto antes de que se presentase una helada pronosticada por los sistemas meteorológicos, como
después de la misma. Para este fin se realizaron dos monitoreos en el cerro Palomas, el primer día
el 3 de febrero de 2007 antes de que se presentase una nevada, y el segundo se realizó 5 días
después, justo un día después de un episodio de nevada. En ambos casos se colectaron mariposas
vivas provenientes de ramas de 13 árboles ubicados tanto en la periferia como dentro de la
colonia de los cuales previamente se obtuvo su posición geográfica, así mismo se recolectaron
otras 50 mariposas muertas, con las que se obtuvieron improntas de abdomen. Los resultados de
este experimento se muestran en la figura 14.
0%
5%
10%
15%
20%
25%
28/06/2003 14/01/2004 01/08/2004 17/02/2005 05/09/2005 24/03/2006 10/10/2006 28/04/2007 14/11/2007 01/06/2008
Fecha
Para
sitosis
media
(%
)hembras muertas machos muertos
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008
Temporada de hibernación

37
Figura 14. Efecto de las nevadas en la parasitosis de la mariposa monarca. Se observan diferencias en la frecuencia de parasitosis antes (A) y después de la nevada (B) cada numero representa un árbol monitoreado, así como su posición dentro de la colonia, en azul marino se representa la parasitosis en machos, en rojo es la parasitosis en hembras, V= mariposas vivas M=mariposas muertas. A) los niveles de parasitosis encontrados en el monitoreo realizado antes de las nevadas mostraron una alta tasa de parasitosis, nunca antes registrada. B) Después de la nevada hubo una disminución significativa en la parasitosis (α=0.005).
03-02-20071
20
40
60
80
0
100N
S
EO
Palomas
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
5
2
7
9
4
6
3
8
MV
MV
MV
MV
MV
V
MV
MV
MV21.421.4
4.54.5
21.721.7
36.036.0
30.830.8
7.77.7 8.38.3
88.588.5
20
40
60
80
0
100MV
58.358.3
6.36.320
40
60
80
0
100MV
80.080.0
0.00.0
10 11
20
40
60
80
0
100MV
34.634.6
13
20
40
60
80
0
100MV
80.880.8
12
15.415.4
33.333.3
8.08.0
12.512.5
11.511.5
48.148.1
0.00.0
15.415.4
4.24.2
M
0.00.0
03-02-20071
20
40
60
80
0
100N
S
EO
Palomas
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
5
2
7
9
4
6
3
8
MV
MV
MV
MV
MV
V
MV
MV
MV21.421.4
4.54.5
21.721.7
36.036.0
30.830.8
7.77.7 8.38.3
88.588.5
20
40
60
80
0
100MV
58.358.3
6.36.320
40
60
80
0
100MV
80.080.0
0.00.0
10 11
20
40
60
80
0
100MV
34.634.6
13
20
40
60
80
0
100MV
80.880.8
12
15.415.4
33.333.3
8.08.0
12.512.5
11.511.5
48.148.1
0.00.0
15.415.4
4.24.2
M
0.00.0
20
40
60
80
0
100N
S
EO
08-02-2007
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
1
5
2
7
9
4
6
3
8
MV
MV
MV
MV
MV
V
MV
MV
MV
0.00.0 0.00.09.19.19.59.5
4.24.20.00.0
8.78.7
20
40
60
80
0
100MV
16.716.720
40
60
80
0
100MV
4.34.3
40.040.0
10 11
20
40
60
80
0
100MV
13
20
40
60
80
0
100MV
6.36.3
12
4.84.80.00.0
4.24.28.38.3
5.35.3 0.00.0 0.00.08.38.3
0.00.0
M
8.38.3
4.24.2 8.78.70.00.0
20
40
60
80
0
100N
S
EO
08-02-2007
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
20
40
60
80
0
100
1
5
2
7
9
4
6
3
8
MV
MV
MV
MV
MV
V
MV
MV
MV
0.00.0 0.00.09.19.19.59.5
4.24.20.00.0
8.78.7
20
40
60
80
0
100MV
16.716.720
40
60
80
0
100MV
4.34.3
40.040.0
10 11
20
40
60
80
0
100MV
13
20
40
60
80
0
100MV
6.36.3
12
4.84.80.00.0
4.24.28.38.3
5.35.3 0.00.0 0.00.08.38.3
0.00.0
M
8.38.3
4.24.2 8.78.70.00.0
A
B

38
Para evaluar si hubo asociación entre la nevada con los cambios en el nivel de parasitosis se
ejecutó la prueba estadística Mantel-Haenszel, en la cual se considera en la hipótesis nula, que no
hay asociación entre la ocurrencia de nevadas con la disminución de la parasitosis en mariposas
vivas o muertas. Después de aplicar la pruebas, al comparar los resultados, se descarto la
hipótesis nula, de tal manera que sí hay relación entre estas dos variables ( X2MH= 93.54 > 7.879
α=0.005).
7.2 Análisis Bacteriológico.
El análisis bacteriológico consistió en el aislamiento de todas las especies cultivables en los
medios de cultivo indicados en la sección de materiales y métodos, así mismo se cuantificó la
carga bacteriana de cada cepa diferente, la cual subsecuentemente fue caracterizada
bioquímicamente hasta identificar género y especie.
7.2.1 Diferencias en la carga microbiana encontrada en diversos sitios anatómicos.
Para este ensayo se seleccionaron 30 mariposas vivas, de obtenidas en El Rosario durante la
temporada de hibernación 2006-2007 (cuadro 1), las mariposas se obtuvieron de 5 árboles
diferentes, que se eligieron considerando su posición dentro de la colonia, es decir se tomó un
árbol en el norte de la colonia, uno en el sur, otro en el este y otro más en el oeste, finalmente el
quinto árbol se encontró en el centro de la colonia. Todas las mariposas analizadas se
manipularon con guantes de látex y se mantuviron en bolsas estériles, sólo se aceptaron
mariposas no parasitadas de peso entre 0.50g ± 0.05 g Se siguieron los protocolos antes descritos
para el análisis bacteriológico.
La cantidad de cada una de las bacterias encontradas fue diferente de acuerdo con el sitio
anatómico a partir del cual se realizaron los aislamientos En la figura 15 se muestra la
comparación entre las medianas, en dicha figura se observa que el abdomen presentó mayor
cantidad de bacterias, (mediana 1.11X109 ufc/mariposa), en comparación con los otros dos sitios
(p<0.001), por otro lado, la carga bacteriana encontrada en el cefalotórax, (mediana 4.0X107
ufc/mariposa), y en las alas (mediana 1.49X107 ufc/mariposa) no mostraron diferencia en la carga
microbiana al compararse entre si, (prueba de Mann-Whitney (p=0.565)).

39
Figura 15. Carga microbiana total en diferentes sitios anatómicos de la mariposa monarca. Los resultados mostrados corresponden a la mediana de unidades formadoras de colonias (UFC) recuperadas de cada uno de los sitios anatómicos de la mariposa monarca, *Indica diferencia estadísticamente significativa entre las medianas (p<0.001), prueba de Mann-Whitney.
7.2.2 Caracterización bioquímica.
La cuenta e identificación se realizó a todas las cepas independientemente del sitio donde fueron
aisladas; para este fin se trabajo a doble ciego, en cada cepa se realizaron todas las pruebas
descritas en materiales y métodos. Los resultados mostraron una gran variedad de diferencias
fenotípicas entre las cepas. Considerando que los métodos convencionales de identificación
bacteriana basan sus resultados en datos obtenidos de cepas de origen clínico o veterinario, en
este trabajo, se prefirió utilizar taxonomía numérica (Holder-Franklin y cols., 1992; Prado y cols.,
2001; Sneath, 2005), el análisis de los patrones bioquímicos utilizando el CSS y el método
UPGMA, generó un dendograma que permitió agrupar a los 767 aislados en 24 grupos
fenotípicos considerando un nivel de similitud del 80%; esta comparación se realizó incluyendo
todas las cepas aisladas, (figura 16).
1.00E+05
1.00E+06
1.00E+07
1.00E+08
1.00E+09
1.00E+10
Abdomen Alas Cefalotórax
Sitio anatómico
UF
C/m
ari
po
sa
*
*

40
Figura 16 Dendograma simplificado que muestran el agrupamiento de los aislados en grupos fenotípicos, El dendograma se baso en el coeficiente simple de similitud CSS entre las 767 cepas aisladas y 31 cepas de referencia incluidas en el presente estudio, para el agrupamiento se utilizó el método de la UPGMA, todos los aislamientos quedaron incluidos en 24 grupos fenotípicos a nivel del 80% al 95% de similitud, BGNF=Bacilos gram negativo fermentadores, BGNNF= Bacilos gram negativo no fermentadores, BGP= Bacilos gram positivo, CGPCN=Cocos gram positivo catalasa negativos.
100908070
100908070
Gru
po f
enot
ípic
o
No.
de
cepa
s
Grupo Identificado
A 191 Enterobacteriaceae
B 15 Aeromodaceae
E 4 Neisseriaceaceae
F 35 Pasteurella spp
H 17 BGNNF 1
M 51 Pseudomonadales
G 77 BGNF
L 8 Alcaligenaceae
K 6 Xantomonadaceae
I 8 BGNNF 2
J 9 BGNNF 3
N 64 Corynebacteriaceae
Ñ 26 Micrococcaceae
O 47 Staphylococcaceae
Q 44 Bacillaceae
W 38 CGPCN
V 7 Lactobacillus spp
P 12 BGP Irregulares
S 13 Listeria spp
U 14 Microbacteriacea
C 17 Vibrionaceae
R 18 Planococcaceae
T 5 Nocardiaceae
Cepas de referencia
Vibrio parahaemolyticus PECEL 0110
Vibrio cholerae No O1 PECEL 9912
Aeromonas hydrophila PECEL 0103
Acinetobacter calcoaceticus PECEL 0105
Moraxella catarrhalis PACAL 0502
Pseudomonas aeroginosa ATCC 9027
Plesiomonas shigelloides PACAL 0212
Pasteurella sp LMC 15
Bacillus cereus PECEL 9911
Staphylococcus aureus ATCC 25923
Micrococcus sp LMC 23
Lactobacillus sp PACAL 0210
Streptococcus mitis PACAL 0302
Streptococcus agalactiae PECEL 0011
Enterococcus faecalis PACAL 0504
Listeria monocytogenes PACAL 0607
Yersinia enterocolitica PACAL 0506Escherichia coli ATCC 25922
Proteus mirabilis PACAL 0205Enterobacter aerogenes ATCC 29008Enterobacter cloacae PACAL 0505Serratia marcescens PECEL 0005Klebsiella pneumoniae ATCC 10031
Citrobacter freundii PECEL 0004Salmonella enterica sub typhimurium ATCC 13311Salmonella enterica sub typhi ATCC 7251Salmonella enterica sub choleraesuis ATCC 10708
Corynebacterium sp LMC 93
Kurthia sp LMC 56
D 6 Acinetobacter spp
100908070 100908070
100908070 100908070
Gru
po f
enot
ípic
o
No.
de
cepa
s
Grupo Identificado
A 191 Enterobacteriaceae
B 15 Aeromodaceae
E 4 Neisseriaceaceae
F 35 Pasteurella spp
H 17 BGNNF 1
M 51 Pseudomonadales
G 77 BGNF
L 8 Alcaligenaceae
K 6 Xantomonadaceae
I 8 BGNNF 2
J 9 BGNNF 3
N 64 Corynebacteriaceae
Ñ 26 Micrococcaceae
O 47 Staphylococcaceae
Q 44 Bacillaceae
W 38 CGPCN
V 7 Lactobacillus spp
P 12 BGP Irregulares
S 13 Listeria spp
U 14 Microbacteriacea
C 17 Vibrionaceae
R 18 Planococcaceae
T 5 Nocardiaceae
Cepas de referencia
Vibrio parahaemolyticus PECEL 0110
Vibrio cholerae No O1 PECEL 9912
Aeromonas hydrophila PECEL 0103
Acinetobacter calcoaceticus PECEL 0105
Moraxella catarrhalis PACAL 0502
Pseudomonas aeroginosa ATCC 9027
Plesiomonas shigelloides PACAL 0212
Pasteurella sp LMC 15
Bacillus cereus PECEL 9911
Staphylococcus aureus ATCC 25923
Micrococcus sp LMC 23
Lactobacillus sp PACAL 0210
Streptococcus mitis PACAL 0302
Streptococcus agalactiae PECEL 0011
Enterococcus faecalis PACAL 0504
Listeria monocytogenes PACAL 0607
Yersinia enterocolitica PACAL 0506Escherichia coli ATCC 25922
Proteus mirabilis PACAL 0205Enterobacter aerogenes ATCC 29008Enterobacter cloacae PACAL 0505Serratia marcescens PECEL 0005Klebsiella pneumoniae ATCC 10031
Citrobacter freundii PECEL 0004Salmonella enterica sub typhimurium ATCC 13311Salmonella enterica sub typhi ATCC 7251Salmonella enterica sub choleraesuis ATCC 10708
Corynebacterium sp LMC 93
Kurthia sp LMC 56
D 6 Acinetobacter spp
Css

41
La distribución de los grupos fenotípicos en la figura 16 muestra la separación de bacterias en dos
grandes ramas: gram positivos y gram negativos, así mismo la ubicación de cada grupo mostró
relación con las propiedades metabólicas, así los grupos A, B y C al igual que los grupos F y G
son fermentadores, por otra parte los grupos K y M son oxidativos, mientras que los grupos D, E,
H, I J y L son bacterias no fermentadoras. Las cepas de referencia utilizadas en este estudio
quedaron incluidas dentro de los grupos fenotípicos correspondientes, el grupo fenotípico A fue
el más numeroso, en este se agrupó la familia Enterobacteriaceae, con 191 cepas, incluyendo 11
cepas de referencia, en este grupo se concentraron los siguientes géneros: Enterobacter spp,
Serratia spp, Klebsiella spp, se concentraron también un grupo compuesto por 13 cepas que
aparentemente son enterobacterias, aunque dichas cepas mostraron patrones atípicos; éstas
mostraron similitud con cepas de Klebsiella spp y Serratia spp, sin embargo, será necesario
realizar otro tipo de pruebas para identificar a que especies pertenecen; también se agruparon
cepas de Citrobacter spp y Salmonella spp. En este último grupo se agruparon 4 cepas de
Salmonella spp de las cuales 3 fueron cepas de referencia.
El grupo E se conformó con cepas de cocos gram negativo, incluyendo a los géneros Kingella sp
y Moraxella spp; el grupo G se conformó de bacilos gram negativo fermentadores, dentro de los
cuales se identificó a Plesiomonas shigelloides que recientemente fue incluido en la familia
Enterobacteriaceae (Janda, 2005), junto a este género se encontraron cepas identificadas como
Aeromonas salmonicida atípica y Vibrio spp atípicos. En el grupo H, I y J se encontraron bacilos
gram negativo no fermentadores, dentro de los cuales se identificó a los géneros Eikenella spp y
Chryseobacterium spp; Comomonas spp y Bordetella spp; Shewanella spp, Burkholderia spp y
Pseudomonas spp atípicas.
En el grupo fenotípico P se agruparon bacilos gram positivos irregulares tales como Turicella spp
y algunas especies atípicas de Corynebacterium spp. En el grupo R se agrupó el género Kurthia
spp. El grupo T se conformó de los géneros Rhodococcus spp y Nocardia spp. Por otra parte en el
grupo U se encontraron bacilos gram positivo pertenecientes a la familia Microbacteriaceae,
dentro de la cual se aislaron especies tales como Microbacterium leavaniformans y Agromyces
ramosus así como Curteobacterium spp y algunas especies atípicas de Corynebacterium spp.

42
Finalmente, el grupo fenotípico W se conformó con cepas de cocos gram positivo catalasa
negativa, tales como Streptococcus spp y Enterococcus spp.
Cada uno de los 24 grupos fenotípicos encontrados se analizó individualmente empleando los
esquemas del manual Bergey correspondientes a la familia o género. A partir de los resultados
reportados se obtuvo el Css entre cada aislado y el patrón bioquímico reportado. Se asignaron las
especies siguiendo los criterios indicados en materiales y métodos. A partir de este análisis, se
obtuvieron para cada cepa aislada tanto su identificación bioquímica como su carga microbiana.
El análisis de todas cepas permitió agruparlas en 153 especies de 53 géneros ubicados en 26
familias, (cuadro 5).
Cuadro 5. Descripción general de las bacterias encontradas en este trabajo.
Famila generos Cepas Famila generos CepasAcinetobacter spp 22 Flavobacteriaceae Chryseobacterium spp 10Moraxella spp 4 Comamonadaceae Comamonas spp 1Acromobacter spp 1 Corynebacteriaceae Corynebacterium spp 71Alcaligenes spp 7 Eikenella spp 7Bordetella spp 6 Kingella spp 2
Actinomycetaceae UK 10 Chromobacterium spp 2Aeromonadaceae Aeromonas spp 18 Enterococcaceae Enterococcus spp 17
Bacillus spp 43 Planococcaceae Kurthia spp 20Exiguobacterium spp 1 Lactobacillaceae Lactobacillus spp 7
Burkholderiaceae Burkholderia spp 1 Listeriaceae Listeria spp 13Cedacea spp 11 Agromyces spp 3Citrobacter spp 5 Curtobacterium spp 4Edwarsiella spp 2 Microbacterium spp 2Enteric Group 69 y 68 14 Micrococcaceae Micrococcus spp 25Enterobacter spp 60 Rhodococcus spp 2Escherichia spp 1 Nocardia spp 3Hafnia spp 1 Actinobacillus spp 12Klebsiella spp 24 Pasteurella spp 53Kluyvera spp 4 Pseudomonas spp 36Moellerella spp 1 Chryseomonas spp 1Obesumbacterium spp 1 Alteromonadaceae Shewanella spp 3Plesiomonas spp 22 Staphylococcaceae Staphylococcus spp 47Rahnella spp 3 Xanthomonadeacea Stenotrophomonas spp 4Salmonella spp 1 Fusobacteriaceae Streptobacillus spp 8Serratia spp 73 Streptococcaceae Streptococcus spp 23Yokenella spp 1 Vibrionaceae Vibrio spp 19
Total de Familas = 26Total de Generos = 53
Total de Especies = 153Total de cepas = 767
Pseudomonadaceae
Pasteurellaceae
Nocardiaceae
Microbacteriaceae
Neisserieaceae
Moraxellaceae
Alcaligenaceae
Bacillaceae
Enterobacteriaceae

43
7.2.3 Carga bacteriana de cada género aislado en la mariposa monarca.
Los resultados arriba mostrados toman en cuenta la suma de todas las cargas individuales de cada
cepa recuperada; no todas las cepas se encontraron en igual cantidad, un análisis detallado de la
carga microbiana individual puede observarse en las figuras 17, 18 y 19, donde se muestra la
carga encontrada por cada género bacteriano en cada sitio anatómico, en cada figura se indican
los géneros en los que se encontró mayor carga microbiana, así como aquellos que fueron
recuperados en al menos el 20% de las mariposas analizadas.
Por otra parte en las figuras 17, 18 y 19 también se muestran a la izquierda las concentraciones
relativas de los géneros con respecto del total de bacterias aisladas de una muestra de 30
mariposas. Se conglomeraron todos los resultados, ya que en la mayoría de las mariposas sólo se
obtuvieron en promedio de 4 a 7 aislamientos, de tal manera que el conjuntar lo encontrado en 30
mariposas, permitió observar la concentración relativa de los géneros en la mariposa monarca.

44
Figura 17. Análisis de la carga microbiana recuperada del abdomen. Se observó diferencia en la carga microbiana en algunos de los géneros bacterianos recuperados en abdomen, las barras de desviación indican el intervalo de confianza del 95%.. A) En abdomen de mariposas vivas las especies de Serratia spp,
Enterobacter spp, Plesiomonas sp, Aeromonas spp, Pasteurella spp y Streptococcus spp fueron las mas abundantes en cuanto a UFC/mariposa, este grupo presentó diferencia estadística significativa (* α = 0.05 ) al compararla con la concentración de las especies de Staphylocccus spp, Micrococcus spp,
Corynebacterium spp y Pseudomonas spp. B) representación de la distribución cuantitativa de los géneros encontrados en abdomen de mariposas vivas, para cada uno se incluye la concentración en UFC encontradas en 30 mariposas, la sección en color azul oscuro representa otros géneros, encontrados en concentraciones menores. C) La concentración de bacterias en mariposas muertas no mostró diferencia entre ellas (α=0.05). D) Representación relativa de las concentraciones de cada bacteria encontrada en un conjunto de 30 mariposas, se incluye para cada género la concentración en UFC/30 mariposas.
N = 30 mariposas
1.00E+05
1.00E+06
1.00E+07
1.00E+08
1.00E+09
1.00E+10
Se
rra
tia
sp
p
En
tero
ba
cte
r sp
p
Ple
sio
mo
na
s s
pp
Aero
mo
na
s s
pp
Pas
teu
rella
sp
p
Str
ep
toco
ccu
s
sp
p
Sta
ph
ylo
co
cc
us
sp
p
Mic
roco
cc
us
sp
p
Co
ryn
eb
ac
teri
um
sp
p
Ps
eu
do
mo
na
s
sp
p
UFC/mariposa
**
Plesiomonas spp,
4.04E+09, (13%)
Otros, 6.69E+09,
(20%)
Pseudomonas spp,
1.78E+08, (1%)
Corynebacterium
spp, 2.24E+08, (1%)
Micrococcus spp,
5.04E+08, (2%)
Staphylococcus spp,
6.18E+08, (2%)
Streptococcus spp,
9.16E+08, (3%)
Pasteurella spp,
1.54E+09, (5%)
Aeromonas spp,
1.81E+09, (6%)
Enterobacter spp,
4.92E+09, (15%)
Serratia spp,
1.04E+10, (32%)
B A
1.00E+05
1.00E+06
1.00E+07
1.00E+08
1.00E+09
1.00E+10
Se
rra
tia
sp
p
Ps
eu
do
mo
nas
sp
p
Ple
sio
mo
nas
sp
p
Co
ryn
eb
ac
teri
um
sp
p
Lis
teri
a s
pp
UFC/mariposa
C Enterobacter spp,
2.04E+09, 8%
Stenotrophomonas
spp, 2.20E+09, 8%
Vibrio spp, 2.02E+09,
8%
Otros, 7.44E+09,
29%
Serratia spp,
3.91E+09, 15%
Pseudomonas spp,
2.25E+09, 9%
Plesiomonas spp,
2.14E+09, 8%
Corynebacterium
spp, 2.01E+09, 8%
Listeria spp,
1.92E+09, 7%
D
N = 30 mariposas
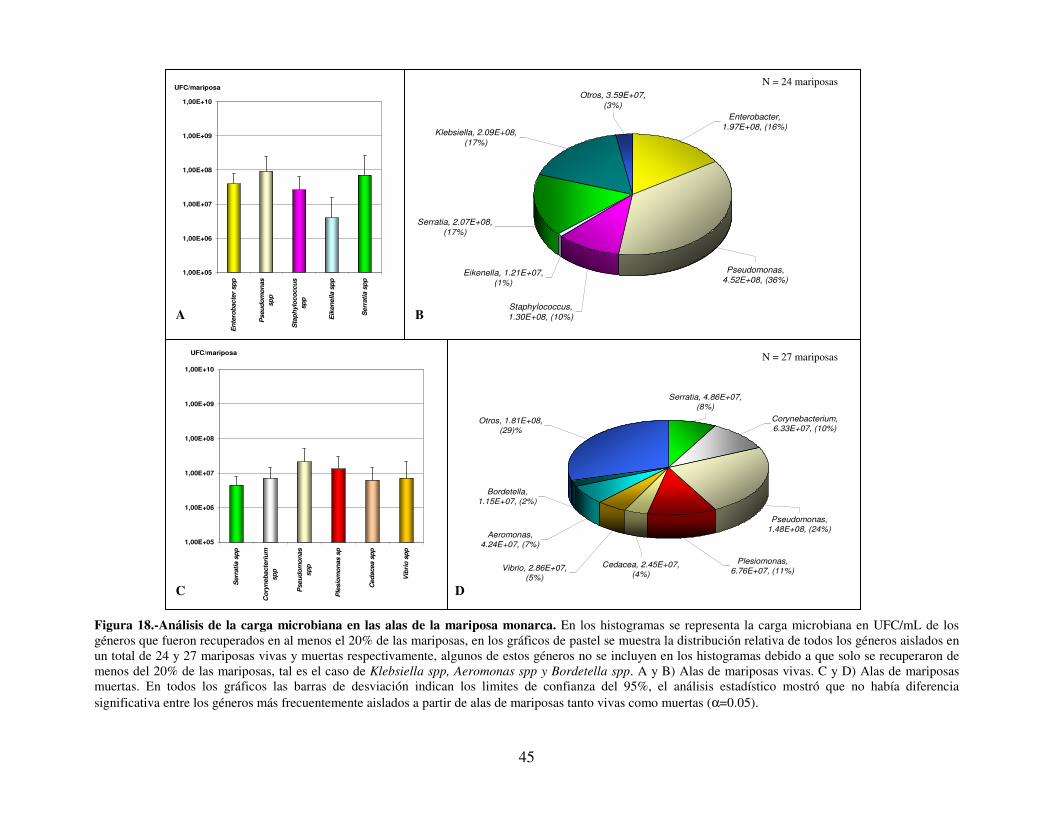
45
Figura 18.-Análisis de la carga microbiana en las alas de la mariposa monarca. En los histogramas se representa la carga microbiana en UFC/mL de los géneros que fueron recuperados en al menos el 20% de las mariposas, en los gráficos de pastel se muestra la distribución relativa de todos los géneros aislados en un total de 24 y 27 mariposas vivas y muertas respectivamente, algunos de estos géneros no se incluyen en los histogramas debido a que solo se recuperaron de menos del 20% de las mariposas, tal es el caso de Klebsiella spp, Aeromonas spp y Bordetella spp. A y B) Alas de mariposas vivas. C y D) Alas de mariposas muertas. En todos los gráficos las barras de desviación indican los limites de confianza del 95%, el análisis estadístico mostró que no había diferencia significativa entre los géneros más frecuentemente aislados a partir de alas de mariposas tanto vivas como muertas (α=0.05).
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
En
tero
bacte
r sp
p
Pseu
do
mo
nas
sp
p
Sta
ph
ylo
co
ccu
s
sp
p
Eik
en
ella s
pp
Serr
ati
a s
pp
UFC/mariposa
Pseudomonas,
4.52E+08, (36%)
Enterobacter,
1.97E+08, (16%)
Otros, 3.59E+07,
(3%)
Klebsiella, 2.09E+08,
(17%)
Serratia, 2.07E+08,
(17%)
Eikenella, 1.21E+07,
(1%)
Staphylococcus,
1.30E+08, (10%)
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
Serr
ati
a s
pp
Co
ryn
eb
acte
riu
m
sp
p
Pseu
do
mo
nas
sp
p
Ple
sio
mo
nas s
p
Ced
acea s
pp
Vib
rio
sp
p
UFC/mariposa
Serratia, 4.86E+07,
(8%)
Corynebacterium,
6.33E+07, (10%)
Pseudomonas,
1.48E+08, (24%)
Plesiomonas,
6.76E+07, (11%)
Bordetella,
1.15E+07, (2%)
Cedacea, 2.45E+07,
(4%)Vibrio, 2.86E+07,
(5%)
Aeromonas,
4.24E+07, (7%)
Otros, 1.81E+08,
(29)%
B A
C D
N = 24 mariposas
N = 27 mariposas

46
Figura 19.- Análisis de la carga microbiana en el cefalotórax de la mariposa monarca. El cefalotórax fue la región anatómica que mostró menor numero de aislamientos. A) Histograma donde se encuentra la carga microbiana de los géneros que al menos fueron recuperados del cefalotórax de un 20% de las mariposas vivas, se encontró que Corynebacterium spp, Chryseobacterium spp y Serratia spp presentaron una mayor concentración por mariposa que la presentada por Micrococcus spp Turicella spp y Bacillus spp esta diferencia fue estadísticamente significativa (α = 0.05) B) gráfico que muestra la proporción de carga bacteriana de cada genero en 23 mariposas. C) En el cefalotórax de las mariposas muertas no se encontró diferencia en cuanto a carga microbiana de los géneros recuperados en al menos el 20% de las mariposas. D) Gráfico que muestra la proporción relativa de los géneros encontrados en cefalotórax de mariposas muertas, la sección azul integra las especies que mostraron menor frecuencia de recuperación.
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
Ente
robacte
r spp
Chry
seobacte
rium
spp
Serr
atia s
pp
Mic
rococcus
Turi
cella
Bacillu
s
UFC/mariposa
*
Serratia spp,
6.60E+08, (29%)
Chryseobacterium
spp, 2.12E+08, (9%)
Enterobacter spp,
4.28E+08, (19%)
Listeria spp,
3.00E+08, (13%)
Klebsiella spp,
2.30E+08, (10%)
Otros, 4.52E+08,
(20%)
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
Serr
atia s
pp
Ple
sio
monas s
p
Paste
ure
lla s
pp
Cory
nebacte
rium
spp
UFC/mariposa
Serratia, 3.00E+09,
(57%)
Plesiomonas,
5.00E+08, (10%)
Pasteurella,
3.40E+08, (7%)
Corynebacterium,
2.20E+08, (4%)
Otros, 1.16E+09,
(22%)
N = 23 mariposas
N = 20 mariposas
A B
C D

47
Los resultados mostrados en las tres figuras anteriores pueden compararse entre sí con mayor
detalle en la figura 20, en la cual se muestran los 16 géneros con mayor carga bacteriana
encontrada en cada sitio anatómico (ufc / sito anatómico), del mismo modo se muestra que la
tendencia a encontrar mayor cantidad de bacterias en el abdomen es similar entre cada género
aislado, también se puede observar que algunos géneros se encontraron en mucho mayor
proporción en el abdomen que en los otros dos sitios anatómicos, tal es el caso de: Aeromonas
spp, Plesiomonas spp, Pasteurella spp, Bacillus spp, Streptococcus spp, Micrococcus spp y
Corynebacterium spp los cuales fueron mayores en al menos un logaritmo.
Figura 20. Carga microbiana y géneros frecuentemente aislados, a partir de diferentes sitios anatómicos de la mariposa monarca. Los resultados mostrados corresponden al promedio de unidades formadoras de colonias (UFC)/mariposa encontradas en el abdomen (barras negras), alas (barras grises) y cefalotórax (barras blancas). Algunos de los géneros mostrados en la figura 20 únicamente se encontraron en dos de los sitios
anatómicos, tal es el caso de Eikenella spp, Aeromonas spp, Plesiomonas sp Pasteurella spp o
Staphylococcus spp, los cuales no se encontraron en el cefalotorax de la mariposa monarca, del
mismo modo, los géneros Micrococcus spp y Turicella spp no se recuperaron de alas.
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
1.00E+09
Eik
enella
spp
Pseudom
onas s
pp
Vib
rio s
pp
Aero
monas s
pp
Ente
robacte
r spp
Ple
sio
monas s
p
Serr
atia s
pp
Paste
ure
lla s
pp
Bacill
us s
pp
Lis
teria s
pp
Sta
phylo
coccus s
pp
Str
epto
coccus s
pp
Mic
rococcus s
pp
Cory
nebacte
rium
spp
Turicella
spp
Chry
seobacte
rium
spp
UF
C/m
ari
po
sa

48
7.2.4 Descripción de la microbiota y comparación en los diferentes sitios anatómicos.
Con objeto de facilitar el análisis de los resultados, en los cuadros del 6 al 11 se pueden observar
las familias, géneros y las especies que al menos mostraron una frecuencia de recuperación
mayor al 5%, el concentrado de todos los resultados puede consultarse en el apéndice A. En los
cuadros 6 al 11, cada uno de los géneros incluye a todas las especies caracterizadas más aquellas
identificadas sólo a nivel de género.
En el grupo de gram negativos, los géneros Enterobacter spp y Serratia spp se encontraron
frecuentemente en los tres sitios anatómicos (cuadro 6), por otra parte hubo géneros recuperados
principalmente en uno o dos sitios anatómicos, así, en el abdomen, Pasteurella spp fue el género
que mostró una frecuencia mayor a la observada en otros sitios anatómicos (P<0.05). Por otro
lado, se observó una alta frecuencia de aislamiento de Eikenella corrodens, y Klebsiella
terrigena, en alas (p<0.05). Asímismo Chromobacterium violaceum, Enterobacter aerogenes y
Chryseobacterium menigosepticum muestran frecuencias de aislamiento mayores en el
cefalotórax que en los otros sitios anatómicos (p<0.05).
Otros géneros gram negativo, que presentaron diferencia menores en la frecuencias de
aislamiento en los distintos sitios anatómicos fueron: Pseudomonas spp, Vibrio spp, Aeromonas
spp, Acinetobacter spp, Actinobacillus spp, Klebsiella spp así como la especie Plesiomonas
shigelloides (p>0.05).
En el grupo de bacterias gram positivo, (cuadro 7) al comparar los aislamientos obtenidos en el
abdomen, las alas y el cefalotórax, se observó que aparentemente en el primero de estos sitios se
aísla con mayor frecuencia a los géneros Corynebacterium spp, Staphylococcus spp y Kurthia spp
(p<0.05). En el cefalotórax hay mayor frecuencia de aislamiento de Turicella otitidis y
Micrococcus kristinae (P<0.05).

49
Cuadro 6. Comparación entre la microbiota bacteriana gram negativa, encontrada en abdomen, alas y cefalotórax de mariposas monarca.
A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje. * Bacterias y grupos cuya frecuencia de aislamiento fue dependiente del sitio anatómico (X2 > 5.991, 2GL, p=0.05)
Bacillus spp Listeria spp y Streptococcus spp, mostraron bajas frecuencias de aislamiento,
menores al 10%, y aunque sus frecuencias de recuperación variaron con el sitio anatómico, dicha
variación no fue estadísticamente significativa (P>0.05).
7.2.5 Temporada de hibernación.
Se determinó si durante la hibernación de la mariposa monarca, había cambios en la microbiota
presente en el abdomen, se agruparon los aislamientos de obtenidos de mariposas colectadas en
alguna de tres diferentes fase de hibernación: la inicial fue considerada desde el 1 de noviembre
al 15 de diciembre, la intermedia desde el 16 de diciembre al 31 de enero, en tanto que la fase
Abdomen Alas Cefalotorax
Neisseriaceae
Eikenella spp * 2B ( 2.1% )C 3 ( 12.5% ) 0 ( 0.0% )Eikenella corrodens * 2 ( 2.1% ) 3 ( 12.5% ) 0 ( 0.0% )
Chromobacterium spp * 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 8.7% )Chromobacterium violaceum * 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 8.7% )
Pseudomonadaceae
Pseudomonas spp 9 ( 9.4% ) 5 ( 20.8% ) 1 ( 4.3% )Pseudomonas aeroginosa 6 ( 6.3% ) 3 ( 12.5% ) 1 ( 4.3% )
Moraxelaceae
Acinetobacter spp 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )Acinetobacter calcoaceticus 5 ( 5.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Vibrionaceae
Vibrio spp 4 ( 4.2% ) 3 ( 12.5% ) 1 ( 4.3% )Aeromonadaceae
Aeromonas spp 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )Aeromonas hydrophila 7 ( 7.3% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterobacteriaceae
Enteric Group 69 5 ( 5.2% ) 1 ( 4.2% ) 0 ( 0.0% )Enterobacter spp 25 ( 26.0% ) 9 ( 37.5% ) 11 ( 47.8% )
Enterobacter aerogenes * 10 ( 10.4% ) 8 ( 33.3% ) 10 ( 43.5% )Klebsiella spp 8 ( 8.3% ) 2 ( 8.3% ) 2 ( 8.7% )
Klebsiella terrigena * 0 ( 0.0% ) 2 ( 8.3% ) 1 ( 4.2% )Plesimonas sp 7 ( 7.3% ) 2 ( 8.3% ) 0 ( 0.0% )
Plesiomonas shigelloides 7 ( 7.3% ) 2 ( 8.3% ) 0 ( 0.0% )Serratia spp 24 ( 25.0% ) 3 ( 12.5% ) 4 ( 17.4% )
Serratia entomophila 4 ( 4.2% ) 1 ( 4.2% ) 2 ( 8.7% )Serratia marcescens 10 ( 10.4% ) 0 ( 0.0% ) 0 ( 0.0% )
Pasteurellaceae
Actinobacillus spp 6 ( 6.3% ) 0 ( 0.0% ) 0 ( 0.0% )Pasteurella spp * 29 ( 30.2% ) 3 ( 12.5% ) 0 ( 0.0% )
Pasteurella multocida 2 1 ( 1.0% ) 2 ( 8.3% ) 0 ( 0.0% )Flavobacteriaceae
Chryseobacterium meningosepticum * 0 ( 0.0% ) 1 ( 4.2% ) 4 ( 17.4% )
Bacterias gram negativo aisladasN=96 A N=24 N=23

50
final se consideró desde el 1 de febrero al 15 de marzo. Para esta comparación se incluyeron
solamente los resultados del análisis bacteriológico realizado en el abdomen de 96 mariposas.
Cuadro 7. Comparación entre la microbiota bacteriana gram positiva, encontrada en abdomen, alas y cefalotórax de mariposas monarca.
A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje. * Bacterias y grupos cuya frecuencia de aislamiento fue dependiente del sitio anatómico, (X2 > 5.991, 2GL, p=0.05).
Los resultados presentados en el cuadro 8 se muestran los géneros y especies bacterianas que
presentaron una frecuencia de aislamiento superior al 5%, además de mostrar variación en la
frecuencia de aislamiento en función a la fase de hibernación. En dicho cuadro se puede observar
que son pocas las bacterias cuya frecuencia de recuperación depende del periodo de hibernación,
del mismo modo, se observa que hay bacterias que muestran disminución significativa en la tasa
de aislamiento (P<0.05), ya sea en el periodo que comprende del inicio de la hibernación a la fase
media (Pasteurella bettyae) o de la fase media a la final (Serratia spp). Por otra lado, también se
observa el comportamiento contrario, es decir un aumento en la frecuencia de aislamiento durante
la fase intermedia y final de la hibernación para la especie Enterobacter taylorae y el género
Enterococcus spp (p<0.05).
Abdomen Alas Cefalotorax
Bacillaceae
Bacillus spp 11A ( 11.5% )C 3 ( 12.5% ) 0 ( 0.0% )Bacillus popilliae 1 ( 1.0% ) 2 ( 8.3% ) 1 ( 4.3% )
Planococcaceae
Kurthia spp * 13 ( 13.5% ) 0 ( 0.0% ) 0 ( 0.0% )Listeriaceae
Listeria spp 1 ( 1.0% ) 2 ( 8.3% ) 2 ( 8.7% )Listeria welshimeri 1 ( 1.0% ) 1 ( 4.2% ) 2 ( 8.7% )
Staphylococcaceae
Staphylococcus spp * 29 ( 30.2% ) 5 ( 20.8% ) 0 ( 0.0% )Staphylococcus lentus 8 ( 8.3% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus warneri 1 ( 1.0% ) 2 ( 8.3% ) 0 ( 0.0% )
Streptococcaceae
Streptococcus spp 9 ( 9.4% ) 0 ( 0.0% ) 2 ( 8.7% )Micrococcaceae
Micrococcus spp * 10 ( 10.4% ) 0 ( 0.0% ) 5 ( 21.7% )Micrococcus kristinae * 1 ( 1.0% ) 0 ( 0.0% ) 3 ( 13.0% )
Corynebacteriaceae
Corynebacterium spp * 34 ( 35.4% ) 1 ( 4.2% ) 2 ( 8.7% )Afiliación desconocida
Turicella spp * 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )Turicella otitidis * 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )
Bacterias gram positivo aisladasN=23N=96 A N=24

51
Cuadro 8. Variación en la frecuencia de aislamientos a partir de abdomen de mariposa monarca en función de la temporada de hibernación. A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje. * Bacterias y grupos cuya frecuencia de aislamiento fue dependiente de la fase de hibernación. (X2 > 5.991, 2GL, p=0.05) + Bacterias que mostraron diferencia en la frecuencia de recuperación durante la etapa inicial y media de la hibernación. (X2 > 3.481, 1GL, p=0.05).ç Bacterias que mostraron diferencia en la frecuencia de recuperación durante la etapa media y final de la hibernación (X2 > 3.481, 1GL, p=0.05).
En el cuadro 9 se presentan los géneros que no mostraron variación conforme avanzó la
temporada de hibernación, en esta se muestran solo aquellos con frecuencia de aislamiento mayor
al 5%.
Cuadro 9. Géneros bacterianos cuyas frecuencias de aislamiento fueron similares durante las tres fases de hibernación de la mariposa monarca. Los géneros incluidos en esta tabla fueron al menos recuperados en frecuencia mayor al 5% del total de mariposas incluidas en cada fase de la hibernación La variación en la frecuencia de recuperación de cada genero durante las tres fases de hibernación no fue estadísticamente significativa (X2 < 3.481, 1GL p=0.05) 7.2.6 Santuarios de hibernación.
El total de muestras analizadas en este trabajo fue obtenido de cuatro sitios de hibernación, El
Rosario, Sierra Chincua, La mesa y Cerro Palomas, estos dos últimos a diferencia de los primeros
son sitios con baja frecuencia de contacto con las comunidades humanas. En los cuadros 10 y 11
se muestra el análisis que se obtuvo al comparar los aislamientos obtenidos de abdomen de
acuerdo con el sitio de hibernación donde fueron colectadas las mariposas. La comparación entre
la diversidad bacteriana presente en el abdomen de mariposa monarca en fase de hibernación en
estos sitios mostró que la mayoría de los bacterias presentan variaciones menores, no
Inicial Media Final
Serratia spp * ç 5B ( 22.7% )C 18 ( 33.3% ) 1 ( 5.0% )Pasteurella spp 9 ( 40.9% ) 13 ( 24.1% ) 7 ( 35.0% )
Pasteurella bettyae * + 2 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterococcus spp * ç 0 ( 0.0% ) 1 ( 1.9% ) 3 ( 15.0% )Enterobacter spp 6 ( 27.3% ) 17 ( 31.5% ) 3 ( 15.0% )
Enterobacter taylorae ç 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 10.0% )
Bacterias que disminuyeron su frecuecia de aislamiento durante la hibernación
Bacterias que aumentaron su frecuecia de aislamiento durante la hibernación
N=20Bacterias
N=22 A N=54
Generos gram negativo Generos gram positivoAcinetobacter spp Bacillus spp
Aeromonas spp Kurthia spp
Pasteurella spp Staphylococcus spp
Enterobacter spp Micrococcus spp
Klebsiella spp

52
dependientes del santuario donde se colectaron las mariposas. Sin embargo, en el grupo de
bacterias gram negativo (cuadro 10), se observó que la especie Serratia odorifera y los géneros
Actinobacillus spp, Acinetobacter spp y Kingella spp se encontraron en mayor frecuencia en la
Mesa y Cerro Palomas, por lo que es posible que esté asociada con estos sitios de hibernación
(p<0.05), por otra lado, también se encontró que aparentemente está relacionada la alta frecuencia
de recuperación de Klebsiella spp, con el sitio de hibernación en Sierra Chincua (p<0.05).
Cuadro 10. Distribución de especies y géneros bacterianos gram negativo en diferentes santuarios de hibernación de la mariposa monarca. A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje. * Especies y/o géneros cuya frecuencia de aislamiento fue dependiente de los sitios de hibernación de la mariposa monarca. (X2 > 7.815, 3GL, p=0.05)
Alcaligenaceae
Alcaligenes spp 0 B( 0.0% )C 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Alcaligenes faecalis 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Neisseriaceae
Kingella spp * 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 14.3% ) 0 ( 0.0% )Pseudomonadaceae
Pseudomonas spp 6 ( 9.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 2 ( 18.2% )Pseudomonas aeroginosa 5 ( 7.5% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Pseudomonas stutzeri 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 9.1% )
Moraxelaceae
Acinetobacter spp * 6 ( 9.0% ) 1 ( 9.1% ) 3 ( 42.9% ) 0 ( 0.0% )Acinetobacter calcoaceticus 5 ( 7.5% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Moraxella spp 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Vibrionaceae
Vibrio spp 4 ( 6.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Aeromonadeaceae
Aeromonas spp 10 ( 14.9% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Aeromonas hydrophila 7 ( 10.4% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterobacteriaceae
Citrobacter spp 2 ( 3.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Enteric Group 69 5 ( 7.5% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterobacter spp 18 ( 26.9% ) 1 ( 9.1% ) 3 ( 42.9% ) 3 ( 27.3% )
Enterobacter aerogenes 10 ( 14.9% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterobacter cancerogenus 2 ( 3.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 9.1% )Enterobacter sakazakii 4 ( 6.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterobacter taylorae 1 ( 1.5% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Klebsiella spp * 3 ( 4.5% ) 4 ( 36.4% ) 0 ( 0.0% ) 1 ( 9.1% )Klebsiella planticola 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Klebsiella pneumoniae 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Plesimonas sp 7 ( 10.4% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Plesiomonas shigelloides 7 ( 10.4% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Serratia spp 20 ( 29.9% ) 1 ( 9.1% ) 1 ( 14.3% ) 2 ( 18.2% )Serratia entomophila 4 ( 6.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia liquefaciens 4 ( 6.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia marcescens 7 ( 10.4% ) 1 ( 9.1% ) 0 ( 0.0% ) 1 ( 9.1% )Serratia odorifera * 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 14.3% ) 0 ( 0.0% )
Pasteurellaceae
Actinobacillus spp * 2 ( 3.0% ) 0 ( 0.0% ) 2 ( 28.6% ) 2 ( 18.2% )Pasteurella spp 18 ( 26.9% ) 5 ( 45.5% ) 3 ( 42.9% ) 3 ( 27.3% )
Sierra ChincuaN=11
Sitio de hibernación Bacterias gram negativo aisladas
N=67 A N=7 N=11El Rosario La Mesa Palomas

53
En los gram positivos, cuadro 11, a pesar de encontrar algunas diferencias en la frecuencia de
recuperación, en los géneros como Bacillus spp, Kurthia spp Enterococcus spp, Streptococcus spp
y Micrococcus spp, éstas no fueron estadísticamente significativas (p<0.05) como para considerar
que estuvieran relacionadas con el sitio de hibernación.
Integrando los resultados anteriores se puede observar que Enterobacter spp, Serratia spp
Pasteurella spp, Staphylococcus spp y Corynebacterium spp están presentes en el abdomen de la
mariposa monarca en todos los sitios de hibernación.
Cuadro 11. Distribución de especies y géneros bacterianos gram positivo en diferentes santuarios de hibernación de la mariposa monarca.
A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje. No se encontraron especies y/o géneros cuya frecuencia de aislamiento fue diferente entre los sitios de hibernación de la mariposa monarca. (X2 > 7.815, 3GL, α=0.05)
7.2.7 Asociación entre O. elektroscirrha con grupos microbianos.
Aunque se sabe que por si solo, el ooquiste de O. elektroscirrha no es capaz de matar o dañar a la
mariposa monarca adulta (Leong y cols., 1997), aun no se ha investigado, si la asociación de este
parásito, pudiese facilitar la infección con algún otro microorganismo, que sí puede causarle
daño. Con objeto de buscar si había asociación entre diversos grupos bacterianos con la presencia
de ooquistes, se decidió comparar la microbiota asociada en mariposas parasitadas y no
parasitadas, adicionalmente, también se realizó la comparación entre mariposas muertas y vivas,
de tal manera que, para este experimento, se formaron 2 grupos de 30 mariposas, uno de ellos
Bacillaceae
Bacillus spp 7 B( 10.4% )C 1 ( 9.1% ) 0 ( 0.0% ) 3 ( 27.3% )Planococcaceae
Kurthia spp 9 ( 13.4% ) 0 ( 0.0% ) 1 ( 14.3% ) 3 ( 27.3% )Staphylococcaceae
Staphylococcus spp 22 ( 32.8% ) 2 ( 18.2% ) 1 ( 14.3% ) 4 ( 36.4% )Staphylococcus auricularis 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus intermedius 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus lentus 8 ( 11.9% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterococcaceae
Enterococcus spp 2 ( 3.0% ) 2 ( 18.2% ) 0 ( 0.0% ) 0 ( 0.0% )Enterococcus faecalis 0 ( 0.0% ) 1 ( 9.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Streptococcaceae
Streptococcus spp 6 ( 9.0% ) 0 ( 0.0% ) 2 ( 28.6% ) 0 ( 0.0% )Micrococcaceae
Micrococcus spp 10 ( 14.9% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% )Corynebacteriaceae
Corynebacterium spp 22 ( 32.8% ) 2 ( 18.2% ) 4 ( 57.1% ) 6 ( 54.5% )
Sierra ChincuaN=11
Bacterias gram postivo aisladas Sitio de hibernación
N=67 A N=7 N=11El Rosario La Mesa Palomas

54
correspondiente a mariposas parasitadas, otro de ellos correspondiente a mariposas no
parasitadas.
Las mariposas se obtuvieron en El Rosario. Se seleccionó para cada grupo, 6 mariposas de cada
uno de 5 árboles ubicados al Norte, Sur, Este, Oeste o Centro de la colonia. Asímismo también se
siguieron los cuidados y criterios antes señalados con el fin de uniformar las condiciones. Debido
a que en este experimento se trabajo con mariposas parasitadas, éstas se seleccionaron después
del análisis parasitoscópico en campo. Los resultados se presentan en la figura 21 que muestra y
compara las diferencias en los aislamientos.
Figura 21. Diferencias observadas en la microbiota presente en mariposas parasitadas y no parasitadas con ooquistes de O. elektroscirrha. Se encontró diferencia significativa en el aislamiento de especies como Listeria spp, Pasteurella spp, y Corynebacterium spp, Corynebacterium spp, Eikenella spp, Enterobacter spp y Turicella spp (α = 0.05) el número total de mariposas analizadas fue de 30 en cada grupo.
Como se observa, hay especies que están presentes en proporción similar tanto en mariposas
parasitadas como no parasitadas, estas corresponden a las especies Plesiomonas sp,
0
5
10
15
20
25
30
35
40
45
Lis
teri
a s
pp
Paste
ure
lla s
pp
Co
ryn
eb
acte
riu
m
sp
p
Serr
ati
a s
pp
Ple
sio
mo
nas s
p
Sta
ph
ylo
co
ccu
s
sp
p
Mic
roco
ccu
s s
pp
Str
ep
toco
ccu
s s
pp
Vib
rio
sp
p
Eik
en
ella s
pp
En
tero
bacte
r sp
p
Tu
ricella s
pp
frecu
en
cia
de a
isla
mie
nto
(%
)
No parasitada Parasitada
*
**
*
*
*
*

55
Staphylococcus spp, Micrococcus spp, Streptococcus spp y Vibrio spp. Por otra parte, se puede
observar que hay especies que principalmente se encuentran en las mariposas parasitadas, éstas
especies pertenecen los géneros Eikenella spp, Enterobacter spp y Turicella spp, la evaluación de
estas diferencias se realizó con la prueba de independencia entre géneros y parasitosis usando X2
(α = 0.05). También se observó menor frecuencia de aislamiento de los géneros Listeria spp,
Pasteurella spp, Corynebacterium spp, y Serratia spp en mariposas infectadas con el ooquiste
(α=0.05).
7.2.8 Diferencias en la microbiota presente en mariposas vivas y mariposa muertas.
La mayoría de los géneros y especies encontrados en mariposas vivas y mariposas muertas, no
mostraron diferencias estadísticamente significativa, sin embargo, hubo tres géneros que se
encontraron en mayor proporción ya sea en mariposas vivas o bien en mariposas muertas, estos
resultados se muestran en la figura 22.
Figura 22. Comparación entre la frecuencia de aislamiento de bacterias en mariposas vivas y mariposas muertas. Se encontró diferencia significativa en el aislamiento de especies como Enterobacter spp, Staphylococcus
spp y Cedacea spp (α = 0.05).
0.0
10.0
20.0
30.0
40.0
50.0
60.0
En
tero
bacte
r sp
p
Bacillu
s s
pp
Ced
acea s
pp
En
tero
co
ccu
s s
pp
Paste
ure
lla s
pp
Ple
sio
mo
nas s
pp
Pseu
do
mo
nas s
pp
Sta
ph
ylo
co
ccu
s
sp
p
Str
ep
toco
ccu
s s
pp
Fre
cu
en
cia
de a
isla
mie
nto
s (
%)
Vivas Muertas
*
*
*

56
Enterobacter spp y Staphylococcus spp fueron encontrados en mucho mayor proporción de
mariposas vivas que muertas (p<0.05), por otra parte Cedacea spp fue encontrado en mayor
frecuencia en mariposas muertas (p<0.05).
7.2.9 Análisis microbiológico de muestras ambientales.
Los resultados microbiológicos ambientales se realizaron con el fin de correlacionar los hallazgos
bacteriológicos presentes en la mariposa monarca en el sitio de hibernación, para este fin se
realizó el estudio bacteriológico en muestras de aire, agua, corteza y suelo. En el Cuadro 12 se
presenta un resumen de las cepas encontradas en las muestras obtenidas.
.
Cuadro 12. Resultados de aislamientos obtenidos a partir de muestras ambientales.
Estos resultados muestran una alta tasa de aislamiento de bacterias en aire y suelo, y menor
cantidad en corteza de oyamel y agua. Todos estos aislamientos corresponden a bacterias
aerobias. En el Cuadro 13 se presenta el análisis detallado de las especies encontradas en fuentes
ambientales.
Los resultados del cuadro 13, muestran el número y porcentaje de muestras ambientales en las
cuales se logró identificar las especies y géneros mostrados en la columna de la izquierda. Varias
cepas únicamente fueron identificadas a nivel de género o familia, ya que los patrones
encontrados fueron atípicos, estas cepas deberán ser sometidas a pruebas adicionales con objeto
de lograr la identificación definitiva.
Se observa también que aproximadamente un 10% de todas las cepas obtenidas de fuentes
ambientales solo pudieron recuperarse a partir del primoaislamiento, no así de las resiembras, aun
empleando el mismo medio de cultivo.
Aire 5 18 17034 UFC / 46 cm2 de medio
expuesto 5 minutosSuelo 5 6 86 1.3 X 106 UFC/ gramoCorteza 5 3 47 4.4 X 104 UFC/gramoAgua 3 6 57 6.0 X 104 UFC/ mL
tipo de muestraNumero de
muestras
Aislamientos
promedio por muestraUFC promedio
No. Total de cepas
aisladas

57
Cuadro 13. Descripción de las bacterias presentes en fuentes ambientales.
A Cantidad total de muestras analizadas B número de muestras con el microorganismo C frecuencia en porcentaje.
7.3 Análisis micológico.
Se realizó un análisis micológico a la mariposa monarca, lo anterior con el fin de identificar las
levaduras y hongos asociados con dicho lepidóptero. Para dicho análisis, se trabajó con el
abdomen de la mariposa monarca y se seleccionaron 10 mariposas vivas y 10 muertas
Alcaligenaceae
Alcaligenes spp 0 B( 0.0% )C 4 ( 80.0% ) 0 ( 0.0% ) 0 ( 0.0% )Alcaligenes faecalis 0 ( 0.0% ) 4 ( 80.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Pseudomonadaceae
Pseudomonas spp 4 ( 80.0% ) 5 ( 100.0% ) 1 ( 20.0% ) 3 ( 60.0% )Pseudomonas aeroginosa 3 ( 60.0% ) 5 ( 100.0% ) 1 ( 20.0% ) 1 ( 20.0% )Pseudomonas stutzeri 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 9.1% )
Bacilos oxidativos atípicos 5 ( 100.0% ) 4 ( 80.0% ) 3 ( 60.0% ) 2 ( 40.0% )Neisseriaceae
Chromobacterium spp 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 3 ( 60.0% )Chromobacterium violaceum 0 ( 0.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 40.0% )
Moraxelaceae
Acinetobacter spp 2 ( 40.0% ) 3 ( 60.0% ) 1 ( 20.0% ) 2 ( 40.0% )Moraxella spp 0 ( 0.0% ) 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Aeromonadeaceae
Aeromonas spp 2 ( 40.0% ) 4 ( 80.0% ) 1 ( 20.0% ) 4 ( 80.0% )Enterobacteriaceae
Enterobacteria atípica 5 ( 100.0% ) 5 ( 100.0% ) 3 ( 60.0% ) 2 ( 40.0% )Citrobacter spp 2 ( 20.0% ) 3 ( 60.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterobacter spp 5 ( 100.0% ) 5 ( 100.0% ) 2 ( 40.0% ) 4 ( 80.0% )
Enterobacter aerogenes 4 ( 80.0% ) 5 ( 100.0% ) 0 ( 0.0% ) 3 ( 60.0% )Enterobacter cancerogenus 1 ( 20.0% ) 0 ( 0.0% ) 2 ( 40.0% ) 1 ( 20.0% )
Klebsiella spp 5 ( 100.0% ) 4 ( 80.0% ) 2 ( 40.0% ) 2 ( 40.0% )Klebsiella planticola 0 ( 0.0% ) 2 ( 40.0% ) 0 ( 0.0% ) 0 ( 0.0% )Klebsiella pneumoniae 0 ( 0.0% ) 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Plesimonas sp 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 40.0% )Plesiomonas shigelloides 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 40.0% )
Serratia spp 3 ( 60.0% ) 5 ( 100.0% ) 2 ( 40.0% ) 3 ( 60.0% )Serratia liquefaciens 1 ( 20.0% ) 3 ( 60.0% ) 0 ( 0.0% ) 1 ( 20.0% )Serratia marcescens 2 ( 40.0% ) 4 ( 80.0% ) 2 ( 40.0% ) 1 ( 20.0% )
Bacillaceae
Bacillus spp 5 ( 100.0% ) 5 ( 100.0% ) 3 ( 60.0% ) 3 ( 60.0% )Bacillus cereus 4 ( 80.0% ) 5 ( 100.0% ) 2 ( 40.0% ) 2 ( 40.0% )Bacillus subtilis 3 ( 60.0% ) 4 ( 80.0% ) 1 ( 20.0% ) 0 ( 0.0% )Bacillus licheniformis 1 ( 20.0% ) 1 ( 20.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Staphylococcaceae
Staphylococcus spp 2 ( 40.0% ) 2 ( 40.0% ) 1 ( 20.0% ) 2 ( 40.0% )Micrococcaceae
Micrococcus spp 3 ( 60.0% ) 2 ( 40.0% ) 1 ( 20.0% ) 2 ( 40.0% )Corynebacteriaceae
Corynebacterium spp 2 ( 40.0% ) 4 ( 80.0% ) 3 ( 60.0% ) 2 ( 40.0% )
Bacteria aislada SueloN=5
Muestra ambiental AguaN=5N=5
CortezaN=5 AAire

58
provenientes del El Rosario, la Mesa y cerro de las Palomas dichas mariposas se colectaron en la
temporada 2005-2006 (tabla 1).
Los resultados muestran que los grupos de hongos y las levaduras son los que se encuentra en
menor proporción asociada a la mariposa monarca (Figura 23). Por otra parte se puede apreciar
que prácticamente no hay diferencia entre la frecuencia de aislamiento de hongos en los
diferentes sitios de hibernación y la frecuencia de recuperación de hongos a partir de mariposas
vivas y mariposas muertas.
Figura 23. Frecuencia relativa de aislamiento de hongos en tres sitios de hibernación de la mariposa monarca. En este estudio se incluyeron 4 mariposas vivas y 4 muertas en el rosario, 3 mariposas vivas y 3 muertas en la mesa y 3 mariposas vivas y 3 muertas en el cerro de las Palomas. Los números indicados sobre las barras indican el número de aislamientos realizados.
Debido a que la identificación de los hongos se fundamente en características morfológicas, se
realizaron microcultivos de los hongos aislados que fueron observados por microscopía, En la
figura 24 se muestran la imágenes obtenidas de diversos tipos de hongos aislados.
En la figura 25 se observan las morfologías macróscopicas de los principales hongos
encontrados. Estos cultivos se realizaros en agar harina de maíz; las fotografías se tomaron 10
días después de la incubación a 28°C.
0
6
0 01
4
30
17
24
20 20
15
1
34
1
3 3
12
32
3
1
0
5
10
15
20
25
30
35
vivas muertas vivas muertas vivas muertas
El Rosario La Mesa Palomas
Fre
cu
en
cia
ais
lam
ien
tos
Actinomicetos Bacterias Hongos Levaduras

59
Figura 24. Morfología microscópica de los principales hongos identificados. Las imágenes fueron obtenidas de microcultivos realizados en agar harina de maíz, las observaciones se realizaron con el objetivo de 40 aumentos. A) Cladosporium spp B) Hongo filamentoso de la familia Mucoraceae. C y D) Hongos septados atípicos. E) Hongo atípicos de crecimiento lento después de 30 días de incubación.
Figura 25. Morfología macroscópica de los principales hongos encontrados en la mariposa monarca. Los cultivos fueron realizados en agar harina de maíz y se incubaron a 28°C. A y B) Se observa el crecimiento y pigmentación característicos del hongo Cladosporium spp C) Hongo filamentoso de la familia Mucoraceae que muestra el clásico crecimiento que invade todo el medio de cultivo en 72 hrs. D, E, F y G) Hongos septados atípicos. H e I) Hongos atípicos de crecimiento lento después de 30 días de incubación.
A B C D E
A D G
B E H
C F I

60
La comparación entre aislamientos obtenidos en mariposas vivas y mariposas muertas puede
observarse en el cuadro 14.
Cuadro 14. Tipos de hongos y levaduras aislados en el abdomen de la Mariposa Monarca.
A Cantidad total de muestras analizadas B número de mariposas en donde se aisló el hongo C frecuencia en porcentaje.
Aunque la cantidad de muestras que se han analizado es muy pequeña para establecer
comparaciones, se puede observar mayor cantidad de hongos aislados en las mariposas muertas;
por otra parte, las levaduras aparentemente se encuentran en proporción similar entre ambos
grupos de mariposas. La identificación definitiva aún no se ha completado los resultados
presentados en el cuadro 14 muestran una alta frecuencia de aislamiento del hongo Cladosporium
sp, el cual fue identificado basándose la morfología microscópica y microscópica (Ainsworth,
1973), cabe señalar que este hongo también se ha observado directamente en las escamas de las
mariposas (Figura 26).
Figura 26. Morfología macroscópica y microscópica de Cladosporium sp. Este hongo encontrado se ha encontrado frecuentemente durante los aislamientos, e incluso se ha observado directamente en improntas de abdomen de la Mariposa Monarca.
Hongos filamentososCladosporium sp 4 B( 40.0% )C 2 ( 20.0% )Mucoraceae 2 ( 20.0% ) 4 ( 40.0% )Hongo septado atípico 1 ( 10.0% ) 3 ( 30.0% )Hongo de crec. lento atípico 1 ( 10.0% ) 5 ( 50.0% )
LevadurasPichia sp 2 ( 20.0% ) 2 ( 20.0% )Levaduras atípicas 4 ( 40.0% ) 3 ( 30.0% )
hongo aislado muertasN=10
Mariposas
N=10 Avivas
A B

61
8. Discusión.
La mariposa monarca es una especie cuyo complejo ciclo de vida, involucra varias etapas
realizadas en Canadá EUA y México; por lo que en los tres países, se han desarrollado estrategias
para asegurar que el ciclo continue, sin embargo, el desarrollo de la actividad humana, ha
impactado de diferente manera a la mariposa monarca, ya sea al disminuir la cobertura de los
bosques, mediante la tala ilegal (Alonso-Mejía y cols, 1997; Brower y cols 2002; WWF 2004), o
al utilizar productos biológicos para el control de plagas en cultivos (Hellmich y cols., 2001;
Sears y cols., 2001; Tames-Guerra y cols., 2004). Por otra parte, las poblaciones de la mariposa
monarca, al igual que todas las especias animales están sujetas al control natural, ya sea a través
de sus depredadores (Calvert y cols., 1976; Glendinning y cols., 1988), a través de cambios
ambientales, como la temperatura y humedad (Alonso-Mejía y Arellano-Guillermo, 1992) o bien
por la presencia de parásitos (Leong y cols., 1997; Altizer., 2001; Bradley y cols., 2005; De
Roode y cols., 2008). Todos estos factores influyen año con año en el nivel de mortandad en la
mariposa monarca. Estimar la mortandad o el impacto de los factores antes mencionados en la
población de la mariposa, ha sido una tarea complicada, una de las maneras indirectas que se ha
realizado, ha sido la medición de áreas de ocupación de la mariposa durante la hibernación en los
bosques mexicanos, los resultados de estos estudios desde la temporada de 1993, muestran que
las poblaciones de la monarca fluctúan considerablemente (Rendón-Salinas y cols., 2006), la
razón de esta variabilidad es desconocida, por otro lado, durante algunos periodos, se ha
documentado la muerte aparentemente inexplicable de grandes cantidades de mariposas. Por tal
motivo es necesario considerar otros factores, tales como la presencia de parásitos o de bacterias
patógenas, sin embargo no se ha investigado qué bacterias asociadas a la mariposa son patógenas
y cuales no los son. Este trabajo se enfocó a determinar tanto la prevalencia de parásitos
patógenos, como las bacterias y hongos asociados a las mariposas monarca aparentemente sanas
y a mariposas muertas; para esto, se cuantificaron y caracterizaron las especies asociadas al
abdomen, alas y cefalotórax en la mariposa. Aunque la mayoría de los trabajos se enfocan al
análisis intestinal, en este trabajo se optó por el estudio de tres áreas de la mariposa, una externa
como son las alas, mismas que reflejan el tipo de microorganismos presentes en el ambiente y
dos internas como el cefalotórax y abdomen, sitios que incluyen los segmentos de tubo digestivo
anterior y medio-posterior respectivamente (Horn, 1978; Richards and Davies, 1977).

62
8.1 Análisis parasitológico.
Ophryocystis. Electroscirrha (Figura 5) es un parásito que usualmente puede ser detectado sobre
la mariposa en hibernación en nuestro país, y aun no se sabe con certeza si este parásito puede
causar daño al adulto, sólo se ha determinado que la fase larvaria de la mariposa monarca es la
susceptible a la infección, ya que en las células intestinales, en esta etapa son invadidas por O.
elektroscirrha, el cual después de que se multiplica en dos fases de merogonia produce sus
estructuras de resistencia, los ooquistes, éstos emergen hacia las células que formarán las escamas
(Reyes, 2004). Las imágenes de la figura 6 aparentemente confirman que los parásitos surgen
asociados a las escamas, ya que se puede observar que hay una distribución particular en el inicio
de la escama, sin embargo, será necesario realizar estudios posteriores enfocados a confirmar
estos hallazgos.
Aunque quedan muchas preguntas por responder acerca de la patogenicidad de O. electroscirrha
sobre la diferentes etapas del ciclo de vida de la mariposa monarca, actualmente se sabe que una
alta frecuencia de infección podría disminuir sustancialmente la población de la mariposa que
arriba a nuestro país, por tal motivo es indispensable realizar una vigilancia epidemiológica
constante acerca de la infección con este patógeno. Hay estudios previos que muestran que en los
sitios de hibernación en nuestro país, la parasitosis no supera el 8%, este bajo nivel contrasta con
la población migratoria de oeste, la cual migra una distancia menor a California y suele presentar
niveles de infección de hasta un 50%, finalmente en la población de mariposas monarca no
migratoria en Florida, la prevalencia de infección es superior al 90% (Altizer y cols., 2000). Estos
datos han conducido a establecer hipótesis acerca del efecto benéfico que tiene la mariposa
monarca al realizar la gran migración ya que aparentemente le ayudaría a mantener baja
prevalencia de parasitosis. Sin embargo, los resultados presentados en este trabajo muestran un
claro aumento de la prevalencia. Esta afirmación es muy seria ya que tiene implicaciones para los
tres países que comparten las etapas del ciclo de vida de la mariposa monarca, de tal manera que
es imperativo demostrar que estas observaciones sean correctas.
En primer lugar debemos acotar los resultados presentados en este trabajo, ya que se ha
determinado que la infección con este parasito puede ser clasificada en dos: nivel de infección
alto y nivel de infección bajo, Esta clasificación está relacionada con la carga de estructuras

63
parasitarias presentes en la mariposa monarca, Los datos de prevalencia mencionados antes
corresponden al nivel alto de infección, este nivel es el que usualmente se emplea para la
vigilancia en los programas de salud en EUA. En este trabajo, se evaluó la prevalencia de
infección en nivel alto, ya que este tipo de mariposas son las causantes de un fuerte impacto
negativo ya que son la principales responsables de transmitir la infección a mariposas sanas. Por
otra parte, nuestros resultados representan los datos del programa de salud en México para la
mariposa monarca, y dichos resultados deben ser comparables en procedimiento y unidades con
los reportados a nivel Trilateral (CEC, 2008).
Nuestros resultados, desde hace tres años, muestran un aumento en el promedio de parasitosis en
la mariposa monarca, (la figura 7 muestra claramente la modificación de la media de parasistosis
desde la temporada de hibernación 2005-2006 a la temporada 2007-2008). Estos resultados
fueron obtenidos tras determinar el porcentaje de parasitosis en la población de mariposas
hacinada por cada arbol, Los resultados fueron consistentes ya que la distribución en campana
prácticamente no se modificó, por otra parte se puede inferir que la distribución de mariposas
parasitadas en los diferentes árboles que conforman la colonia es azarosa, para demostrar esta
hipótesis, se aplicó una prueba estadística basada en la x2 llamada prueba de homogeneidad, La
hipótesis de homogeneidad indicó que no hay predilección de parasitosis hacia alguna región
geográfica. Estos resultados se confirmaron con un estudio similar en la temporada 2007-2008,
pero, en este caso se incluyeron árboles de los bordes de la colonia divididos en los cuatros
puntos cardinales y se compararon con árboles ubicados al centro de la colonia. Los resultados
nuevamente ratificaron que la ubicación de las mariposas infectadas dentro de la colonia es
homogénea en toda la colonia, tanto en el centro como en los bordes de tal manera que la
selección de árboles para los fines de este trabajo no presento ningún sesgo. Por lo anterior es
claro observar un aumento estadísticamente significativo en la parasitosis, las razones de este
aumento son desconocidas, aparentemente no es dependiente del sitio de hibernación, una
posibilidad es el que una mayor proporción de la población migrante, se infectó con O.
elektroscirrha, debido a una reducción en las áreas de alimentación de la mariposa; esto es real ya
que a diferencia de México, en EUA la migración y retorno prácticamente se produce en la mitad
del país, en la región este (Malcolm, 1987) de tal manera que las mariposas para cumplir su ciclo
de vida necesitan de asclépias las cuales son consideradas plaga por los granjeros de EUA (CEC,

64
2008), de tal manera que estos sistemáticamente están eliminando estas plantas, lo cual podría
originar que las mariposas se desvíen hacia zonas en donde la prevalencia de infección por este
parasito es alta, por ejemplo Florida (Altizer y cols. 2000), esta hipótesis tendría que considerar
que las mariposas que arriban a nuestro país llegan ya con alta incidencia de parasitosis, nuestros
resultados permite aseverar que ésta no depende de los sitios de hibernación ya que en todos los
sitios la mariposa esta llegando con el mismo porcentaje de parásitos, lo cual conduce a reforzar
la hipótesis de que el aumento de infección de las mariposas se produjo en Canadá y/o EUA.
Una variable que se decidió considerar fue la proporción de mariposas muertas parasitadas, con el
fin de establecer sí las mariposas parasitadas son mas susceptibles a morir que las no parasitadas.
Nuestros resultados mostraron que aunque en algunas temporadas hay mayor frecuencia de
parasitosis en las mariposas muertas, esta variación no es estadísticamente significativa, lo cual
desecha la hipótesis que propone mayor susceptibilidad de las mariposas parasitadas a morir que
las no parasitadas. Al analizar la distribución de la parasitosis en relación con el sexo, se observa
que en los machos aparentemente hay mayor prevalencia.
Estudios previos indicaron que la frecuencia de mariposas altamente infectadas tiende a
mantenerse sin cambio durante el período de hibernación (Altizer y cols., 2000); sin embargo
nuestros resultados muestran gran variación en la prevalencia de parasitosis de un monitoreo a
otro. Una posibilidad que origine esta gran variabilidad, es que, factores climáticos y/o
microbiológicos originen que las mariposas altamente infectadas sean más susceptibles y éstas
mueran predominantemente. Esta hipótesis se basa en el hallazgo de mayores frecuencias de
parasitosis en mariposas muertas que en vivas.
Una de las ventajas de las poblaciones que realizan migración tal como se ha planteado antes es
que mantiene tasas de parasitosis mínimas en comparación de las otras poblaciones (Altizer y
cols., 2000), Se ha planteado por nuestro grupo que los factores climáticos pudieran participar en
el control de la mariposa infectada, para analizar esta posibilidad se realizaron dos monitoreos en
un santuario donde se había pronosticado una nevada, en el primer monitoreo se observó una alta
tasa de parasitosis, que nunca antes se había reportado, aun no es posible explicar este fenómeno
que se discutirá mas adelante. Por otra parte 5 días después al realizar el segundo monitoreo, se

65
encontró una disminución significativa en la parasitosis tanto de mariposas vivas como muertas
(α=0.005), este segundo monitoreo fue realizado un día después del pronosticado de nevada.
Estos resultados sugieren que, las nevadas, una de las principales amenazas a las que está
expuesta la mariposa monarca (Galindo y cols., 2000) pueden ocasionar mayor muerte a las
mariposas infectadas que a las no infectadas con el parasito. Lo anterior también se correlaciona
por el hecho de encontrar un incremento en la parasitosis en las mariposas muertas tal como se
muestra en el árbol no. 11 de la figura 14B, sin embargo no en todos los árboles se encontró este
aumento, una posible explicación es la presencia de depredadores como roedores, en los cuales se
ha observado, que consumen el abdomen de las mariposas muertas o moribundas (Glendinning y
Brower, 1990), de tal manera que si una nevada ocasionó una mortandad importante en las
mariposas parasitadas, al ser éstas consumidas por los ratones, se impide establecer si
presentaban o no la parasitosis, otra hipótesis que explique estos hallazgos es la posibilidad de
que al moverse la colonia a otra zona en dicha zona se halla producido la muerte de las mariposas
parasitadas, así después de la tormenta, al retornar la colonia a la posición original, las mariposas
muertas presentes en esa zona no serían las parasitadas.
Los resultados de disminución de parasitosis encontrados después de la nevada reforzarían la
hipótesis de que el estrés climático sobre la población de la mariposa hibernante, puede ser una
ventaja para la mariposa migrante, que explique el porque las poblaciones que llegan hasta
México mantienen un tasa de infección menor a las otras dos poblaciones del norte de América.
Por otra parte el hallazgo de alta parasitosis encontrado en el cerro de la palomas días antes de la
nevada, aun no se puede explicar, tal como se muestra en la figura 14A, se encontraron 6 de 13
árboles con un nivel de parasitosis equivalente al que se encontraría en las poblaciones
migratorias del oeste de California (Altizer, 2001). Desde que inició este proyecto en 2003 en
ninguna ocasión se ha detectado un nivel tan alto de parasitosis, y mucho menos en casi el 50%
de los árboles monitoreados, tal como se muestra en los cuadros 3 y 4 la frecuencia de parasitosis
en la mariposa monarca es homogénea en toda la colonia (α=0.05) de tal manera que los
hallazgos encontrados no pueden ser atribuidos a que el monitoreo se realizó en un punto donde
se aglomeren la mariposas infectadas, sin embargo aun no se han evaluado diferencias en la
parasitosis asociadas con la altura en la que se ubican las mariposas, en todos los monitoreos se

66
han obtenido los racimos de mariposas de alturas menores a los 8 metros, aunque éstos pueden
incluso ubicarse en alturas mucho mayores. De acuerdo a lo anterior, si hubiese diferencias de
parasitosis en cada altura de los árboles podría haber la posibilidad de que las mariposas ubicadas
en racimos en alturas mayores estén más parasitadas o menos parasitadas, es decir, que la
proporción de mariposas parasitadas se ubique en forma no homogénea sobre el árbol. Por otra
parte se ha observado que la colonia de mariposas puede desplazarse hacia otras zonas de bosque,
estos movimientos suelen ser lentos, sin embargo en condiciones de amenaza, como las
climáticas, se observan cambios de posición muy rápido. Tomando en cuenta todo lo anterior,
una hipótesis que argumente las razones de encontrar estos niveles tan altos en la parasitosis
puede apoyarse en el hecho de que la colonia se había desplazado al detectar cambios climáticos
próximos a una nevada, lo cual ocasionaría una desorganización en los estratos (racimos de
mariposa a diferente altura) de mariposas, de tal manera que al ser colectadas en esa etapa, se
encontrarían las diferencias observadas. Para probar esta hipótesis sería necesario analizar los
racimos de mariposas a diferentes alturas, así como probar que durante un cambio de posición
influenciado por cambios climáticos, ocurre esta desorganización en los estratos.
8.2 Análisis Bacteriológico.
Los resultados del análisis bacteriológico muestran que el interior del abdomen es el sitio con
mayor cantidad de microorganismos (p<0.001), la carga microbiana mostrada en la figura 15 es
similar a la encontrada en otros artrópodos (Cazemier y cols., 1997). Durante la hibernación
realizada por la mariposa monarca, las fuentes de energía, la constituyen el néctar que obtenido
de flores, sin embargo durante el invierno prácticamente desaparecen las flores, de tal manera que
la mariposa sólo puede disponer de las reservas de lípidos, obtenidas durante la fase larvaria
(Masters y cols., 1988). Estas constituyen hasta un 45% del peso seco del insecto, y se encuentran
en mayor proporción en el abdomen que en cefalotórax o alas (Cenedella, 1971; Brown y
Chippendale 1974), la carga microbiana encontrada en este trabajo muestra que podría haber una
relación con la distribución de lípidos en la mariposa monarca.
El análisis detallado de la diversidad microbiana en los sitios anatómicos muestra que hay 16
géneros bacterianos que fueron recuperados en cuentas mayores a 1×106 ufc/mariposa ya sea en
uno o mas sitios anatómicos, figuras 17,18 y 19. Quince de estos géneros han sido previamente

67
encontrados en estudios bacteriológicos realizados en insectos (Kingsley 1972; Dugas y cols.,
2001; Kuzina y cols., 2001; Zayed y Bream 2004; Dunn and Stabb 2005; Tóth y cols, 2005;
Wertz and Breznak 2007; Indiragandhi y cols., 2008) por otra parte el genero Turicella spp,
únicamente se ha reportados en casos de otitis media en humanos (Funke y cols., 1994; Funke y
cols., 1997). Los géneros gram negativos tales como Serratia spp, Enterobacter spp,
Plesiomonas sp, Aeromonas spp y Pasteurella spp aparentemente se encuentran en mayor
cantidad en el abdomen que los gram positivos tales como, Corynebacterium spp, Micrococcus y
Staphylococcus spp, figura 17, estos resultados, a diferencia de los encontrado en mariposas
monarcas crecidas en laboratorio (Kingsley 1972) muestran que las enterobacterias
aparentemente se encuentran en altas cargas en el abdomen de la mariposa en vida silvestre,
(Figura 20). Aunque en insectos no se ha determinado qué criterios pueden ser usados para
determinar si una especie pertenece o no a la microbiota, se espera que la microbiota incluya
poblaciones microbianas numerosas y estables en un determinado hospedero (Dillon and Dillon
2004). Por otra parte, se espera que las especies pertenecientes a la microbiota se encuentren
asociadas con la mayoría de individuos de una misma especie, los resultados de este trabajo
muestran en alta variabilidad en el aislamiento de cepas sin embargo enterobacterias como
Enterobacter spp o Serratia spp, se encontraron frecuentemente en los tres sitios anatómicos,
figura 20 Enterobacter spp, el género microbiano más frecuentemente aislado en todos los sitios
anatómicos es común en diversos tipos de insectos como: Dípteros (Demaio y cols., 1996;
Kuzina y cols., 2001; González-Cerón y cols., 2003), Lepidópteros (Lighthart, 1988; Broderick y
cols., 2004;), Coleópteros (Yilmaz y cols., 2006) y Neurópteros (Woolfolk and Inglis 2004; Dunn
and Stabb 2005), así mismo se ha visto que a nivel ambiental este género se encuentra
frecuentemente en aire, agua y suelo (Grimont and Grimont, 2005a) lo cual podría explicar la alta
presencia de este microorganismo en alas, cefalotórax y abdomen de la mariposa monarca.
Serratia spp es un reconocido patógeno de varias especies de insectos, propiedad que ha sido
considerada como agente de control biológico (Lecey y cols., 2001), en este estudio las mariposas
analizadas aparentemente no mostraron daño alguno o signo de enfermedad, sin embargo fue
frecuente el aislamiento de este género, particularmente en abdomen se encontró Serratia
entomophila y Serratia marcescens en frecuencias de 4.2 al 10.4% respectivamente, ambas
especies han sido reconocidas como entomopatógenas principalmente en larvas de lepidópteros

68
(Sikorowski and Lawrence 1998), coleópteros (Glare y cols., 1993; Jackson y cols., 2001; Bahar
and Demirbag 2007; Gatehouse y cols., 2008) y en Dípteros adultos (O’Callaghan y cols., 1996)
la virulencia de estas especies está relacionada con la capacidad de adherirse a las células de los
insectos (Chadwick y cols., 1990) y de producir quitinasas (Lysenko 1976) sin embargo también
se han encontrado estas especies en insectos adultos sanos (Moore 1972; Demaio y cols., 1996),
este trabajo reporta el primer aislamiento de S. marcescens y S. entomophila a partir de mariposa
monarca adulta, así como el aislamiento de otras especies de Serratia spp, cuyas frecuencias de
aislamiento fueron inferiores al 5% (14 cepas, pertenecientes a otras especies: S. fonticola, S
liquefaciens, S odorifera, S. plymuthica y S rubidia), estos resultados contrastan con los
obtenidos en adultos de mariposa monarca (Kingsley, 1972) en donde no se encontraron ni
siquiera Enterobacterias, la variación entre estos resultados es probable que se deba al ambiente
en el cual se obtuvieron las muestras, ya que en este trabajo se colectaron mariposas directamente
del sitio de hibernación a diferencia de que las mariposas analizadas por Kingsley fueron
desarrolladas en condiciones controladas de laboratorio, esta hipótesis se apoya en estudios
previos, que han mostrado cambios en la microbiota de larvas de lepidópteros cuando éstas son
mantenidas con alimento controlado en el laboratorio a diferencia de aquellas obtenidas en vida
silvestre (Lighthart 1988).
En este trabajo se encontraron otros géneros que mostraron mayor frecuencia de aislamiento en
algún o algunos de los sitios anatómicos analizados, en el abdomen los géneros mas
frecuentemente aislados (p<0.05) fueron: Pasteurella spp, Corynebacterium spp, Staphylococcus
spp y Kurthia spp. El abdomen de los lepidópteros puede dividirse en dos regiones: el intestino
medio (midgut), dentro del cual se encuentra el estomago y el intestino posterior (hindgut) el cual
incluye al recto y ano (Horn 1978), de tal manera que las cepas reportadas como aisladas en
abdomen pueden estar presentes en una u otra región.
Pasteurella spp, se aisló en mas de un 30% de las mariposas incluidas en este estudio, la
frecuencia al compararla con lo otros dos sitios fue estadísticamente significativa (p<0.05),
dentro de las especies pertenecientes a este género se aisló Pasteurella multocida, conocido
agente del cólera aviar (Mutters y cols., 2005) sin embargo la frecuencia de aislamiento de esta
especie fue menor al 1%, la mayoría de la cepas encontradas en esta trabajo mostraron patrones

69
compatibles con el genero Pasteurella spp sin embargo los patrones encontrados no fueron los
suficientemente similares con los patrones descritos para cada especie por tanto únicamente se
reportó como perteneciente al género, de acuerdo con la reciente edición del manual Bergey hay
varias especies que, aunque reciben el genero Pasteurella, no comparten algunas de las
características de las verdaderas Pasteurellas, de tal manera que se espera que posteriormente
dichas especies cambien de ubicación. Los resultados presentados en este trabajo muestran el
aislamiento de varias cepas identificadas como Pasteurella spp, sin embargo será necesario
determinar posteriormente si son verdaderas Pasteurellas o bien cepas pertenecen a este grupo de
bacterias momentáneamente incluidas dentro de este género. En insectos este genero se ha
encontrado en garrapatas (Stojek y Dutkiewicz 2004) En este trabajo se aisló tanto
frecuentemente, cuadro 6, así como en alta carga microbiana >108ufc/mariposa figura 20, es
posible que este género microbiano pueda jugar un papel importante en el ciclo de la mariposa
monarca. Estudios previos han mostrado que el consumo de Asclepias syrica por parte de la
monarca en fase larvaria confiere a esta de un potente cardiotóxico que la protege de la
depredación por parte de las aves (Brower y cols. 1972; Ericsson 1973), del mismo modo si las
cepas identificadas como Pasteurella spp, presentasen los factores de virulencia propios de las
bacterias pertenecientes a este género, el mantener en el abdomen altas cantidades de
microorganismos patógenos de aves, tal vez podría conferir de una protección adicional a la
mariposa contra depredadores susceptibles a esta bacteria, sin embargo actualmente se conocen
tres especies de aves que consumen frecuentemente el abdomen de estas mariposas (Calvert y
cols., 1979).
Corynebacterium spp previamente ha sido encontrado en mariposas monarca crecidas en
laboratorio bajo condiciones ambientales controladas, encontrándose presente en las 5 fases
larvarias así como en el adulto (Kingsley 1972) en este trabajo se encontró en mas del 35% de las
mariposas analizadas figura 20, el hecho de encontrar este genero en un gran número de
individuos, además de que se ha encontrado en fase larvaria puede indicar que dicho
microorganismo puede realizar alguna función importante en el ciclo de vida de la mariposa.
Staphylococcus spp es un grupo bacteriano presente principalmente en mucosas y epitelio de
animales de sangre caliente, sin embargo también se ha encontrado en suelo, polvo, arena, o agua

70
(Kloos and Schleifer 1984), en este estudio se encontró este género en mas del 30% de las
mariposas (n=29), principalmente fue encontrado en abdomen, pero también se encontró en alas
cuadro 7, en otros estudios bacteriológicos realizados en insectos se ha identificado en el intestino
tanto de larvas de lepidópteros (Broderick y cols., 2004) como de dípteros adultos (Zayed y cols.,
2004; Kuzina y cols., 2004) o himenópteros adultos (Gilliam and Taber 1991), en relación a la
mariposa monarca, los estudios previos reportaron la presencia de la familia Micrococcaceae
tanto en los 5 estados larvarios como en la pupa y el adulto (Kingsley 1972); es importante
señalar en la fecha de publicación del estudio anterior el género Staphylococcus se ubicaba
dentro de la familia Micrococcacea (Breed y cols., 1957), aunque los resultados solamente
indican la presencia de esta familia, es probable que gran parte de este grupo hallan sido
Staphylococcus spp.
Kuthia spp es un bacilo gram positivo que se ha encontrado de diversas fuentes, entre ellas
destacan las heces de animales de granja, aunque también se ha encontrado en suelo, agua y aire
(Keddie and Shaw 1984) los resultados mostrados en el cuadro 7 muestran una frecuencia de
aislamiento del 13% únicamente en el abdomen de la monarca, debido a que las cepas aisladas no
mostraron un patrón característico con los reportados en el manual Bergey, los aislados
únicamente se reportaron como Kurthia spp, será necesario ejecutar otro tipo de pruebas para
identificar las especies de este género.
En este estudio también se analizó la microbiota presente en las alas de la mariposa monarca,
algunos de los microorganismos encontrados en esta región anatómica, tales como
Staphylococcus spp y Enterobacter spp, se han podido recuperar de fuentes ambientales, tales
como aire suelo o agua (Kloos and Schleifer 1984; Grimont and Grimont, 2005) lo que supone
que el suelo pueden ser una fuente de este tipo de microorganismos, que al ser arrastrados por el
aire entran en contacto con la mariposa monarca, principalmente en superficies expuestas como
las alas. 11 géneros bacterianos fueron aislados en frecuencia mayor al 5% en alas de la
mariposa, sin embargo únicamente Eikenella corrodens, y Klebsiella terrigena mostraron
frecuencias de aislamiento en alas estadísticamente mayores a los otros dos sitios anatómicos, lo
que hace suponer que estas especies aparentemente están relacionadas con las alas de la mariposa
monarca (p<0.05): En estudios previos se ha logrado recuperar a partir del intestino de termitas, a

71
Stenoxybacter acetivorans, una nueva especie propuesta, la cual esta filogenéticamente
relacionada con Eikenella corrodens (Wertz y Breznak 2007) en este estudio, el análisis de las
características fenotípicas de los aislados identificados como Eikenella corrodens permitió
agruparlos mediante el método UPGMA en una rama junto a Chryseobacterium spp (grupo
fenotípico H), figura 16, sin embargo será necesario realizar pruebas adicionales para corroborar
sí se trata de esta especie o de la especie recientemente propuesta. K. terrigena ha sido
encontrada en neurópteros (Woolfolk y Inglis 2003), pero también se ha encontrado asociada
muestras ambientales tales como el agua y el suelo (Grimont y Grimont, 2005), considerando que
los hallazgos han mostrado mayor frecuencia de recuperación en alas que en otros sitios
anatómicos, es posible que esta bacteria tenga contacto con las alas a través de las corrientes del
aire, las cuales pueden acarrear pequeñas partículas del suelo.
En este estudio se dividió a la mariposa monarca en abdomen, alas y cefalotórax en este ultimo
comprende la cabeza, las estructuras bucales así como el tórax, así mismo, dentro del cefalotórax
se encuentra una parte importante del aparato digestivo de las mariposas el cual es llamado parte
anterior “foregut”, este inicia desde la probóscide y continúa con la faringe y esófago hasta una
estructura utilizada como reserva alimenticia denominada “crop” (buche), todas estas estructuras
participan en la obtención procesamiento y almacenaje de alimento (Richards y Davies 1977;
Horn 1978), así mismo todas estas estructuras albergan microorganismos provenientes de la dieta
de la mariposa. En el caso de la mariposa monarca se ha visto que durante el tiempo de
hibernación, ésta sólo consume agua y néctar de las flores, las cuales prácticamente desaparecen
a medida que entra la temporada invernal, de tal manera que prácticamente sólo sobrevive con las
reservas energéticas obtenidas al alimentarse con Asclépias durante la fase larvaria (Calvert and
Brower 1986). Los hallazgos de microorganismo encontrados en el cefalotórax representan
aquellas especies que pueden ser obtenidas a partir del agua consumida pudiendo ser consideras
transitorias. Pero también es posible que algunas de estas especies se hayan adquirido desde antes
de la migración ya sea a través del consumo de plantas o del contacto con el ambiente, y que
hallan establecido alguna simbiosis con la mariposa monarca, estableciéndose en la parte anterior
del aparato digestivo de la mariposa. Los resultados encontrados muestran que especies como
Chromobacterium violaceum, Chryseobacterium meningosépticum, Enterobacter aerogenes,
Micrococcus christinae y Turicella otitidis mostraron frecuencia de aislamiento mayores en

72
cefalotorax que en los otros sitios anatómicos (p<0.05), por lo que aparentemente estas especies
están relacionadas con dicho sitio anatómico.
Chromobacterium violaceum es una bacteria que puede recuperarse de fuentes ambientales como
el agua o el suelo (Gillis and Logan 2005), en estudios bacteriológicos realizados en insectos se
ha podido recuperar de garrapatas (Stojek y Dutkiewicz 2004) por otra parte, estudios previos
muestran que esta especie puede ser patógena para algunos coleópteros (Martin y cols., 2007a),
heterópteros (Martin y cols., 2007b), en este estudio solo se logro aislar dicha especie en
cefalotórax en 8.7% de las mariposas (2/23), hasta el momento no hay estudios sobre toxicidad
en lepidópteros adultos, sin embargo se ha visto que C violaceum puede interferir en las
actividades fisiológicas llevadas a cabo en la fase larvaria de algunos lepidópteros (Funke y cols
2008), así mismo se ha determinado la capacidad de producir quitinasas, las cuales podrían
causar daño al exoesqueleto de los insectos (Gillis y Logan 2005), serán necesarios estudios
posteriores en las cepas aisladas a fin de determinar si pueden causar daño en adultos o larvas de
la mariposa monarca.
Chryseobacterium meningosepticum, Turicella otitidis y Micrococcus kristinae se aislaron en
cefalotórax en una frecuencia de 17.4% para las dos primeras especies y de 13% para M.
kristinae. C. meningosepticum se ha aislado principalmente en suelo, plantas y agua
(Schreckenberger y Graevenitz 1999), aunque esta no se ha reportado en insectos, la especie
Chryseobacterium indologenes se ha encontrado en cucaracha, en donde aparentemente mantiene
una relación simbiótica positiva con dicho insecto (Dugas y cols 2001). Por otra parte Turicella
otitidis, únicamente se ha encontrado en humanos como agente causal de otitis media (Funke y
cols., 1994; Renaud y cols., 1996), esta bacteria está estrechamente relacionada con
Corynebacterium, en este trabajo los patrones bioquímicos mostrados por las cepas aisladas
fueron congruentes con los descritos para Turicella otitidis, sin embargo podría tratarse de cepas
de Corynebacterium atípicas (Funke y cols., 1997), por tal motivo será necesario realizar
posteriormente estudios filogenéticos para corroborar dichos resultados. Finalmente el género
Micrococcus se ha podido aislar a partir de dípteros (Tóht y cols., 2005), coleópteros (Yilmaz y
cols., 2006), e himenópteros (Gilliam y Taber 1991). El agua y el suelo son fuentes ambientales
de este género, de tal manera es posible que la mariposa monarca adquiera transitoriamente esta

73
bacteria durante la ingesta de agua en los sitios de hibernación, sin embargo el papel que puede
jugar aun es desconocido, serán necesarios estudios adicionales para determinar sí este género es
transitorio en la mariposa monarca.
Varias especies y géneros bacterianos no presentaron variaciones tan grandes como para
considerar que la frecuencias de recuperación fue dependiente de algún sitio anatómico (p>0.05).
tal fue el caso de Pseudomonas spp, , Aeromonas spp, Acinetobacter spp, Vibrio spp, Klebsiella
spp, Plesimonas shigelloides Actinobacillus spp, así como los gram positivo, Bacillus spp,
Listeria spp y Streptococcus spp.
Pseudomonas aeroginosa, una especie bacteriana frecuentemente encontrada en este estudio,
cuadro 6, es otro microorganismo que ha mostrado ser entomopatógeno (Jander y cols., 2000),
Sin embargo también hay otros estudios en donde se ha encontrado como parte de la microbiota
en dípteros (Demaio y cols., 1996; Kuzina y cols., 2001) así como en larvas de lepidópteros
(Broderick y cols., 2004; Indiragandhi y cols., 2008). Uno de los factores involucrados en la
patogenicidad de este género es la capacidad de producir lipasas (Lonon y cols., 1988), aunque se
ha correlacionado la producción de lipasas con la virulencia de Pseudomonas aeroginosa en
lepidópteros es posible que esta bacteria pueda afectar a las mariposas al consumir sus reservas
de lípidos, sin embargo será indispensable realizar estudios posteriores enfocados a determinar el
efecto de este microorganismo en la mariposa monarca para poder establecer el papal que juega
este microorganismo.
Acinetobacter calcoaceticus fue encontrado en 5% de las mariposas, todos los aislamientos
fueron de abdomen, estudios previos han mostrados que esta bacteria es capaz de producir una
lipasa extracelular (Kok y cols., 1995), la cual podría ser nociva para la mariposa monarca si
dicho microorganismo es capaz de tener acceso y consumir sus reservas lipídicas. La
recuperación de este género en insectos es común en dipteros (Zayed y Bream 2004; Toth y cols.,
2004) y neurópteros (Dunn y Stabb 2005). Por otra parte Aeromonas spp también se ha
encontrado previamente asociada a insectos (Kuzina y cols., 2001; Stojek y Dutkiewicz 2004)
ambos géneros no han sido previamente encontrados en la mariposa monarca (Kingsley 1972), en
este trabajo aunque se encontró principalmente en abdomen y alas con un porcentaje de

74
aislamiento de (10.4%) y (4.2%) respectivamente, dicha frecuencia no fue estadísticamente
significativa (p>0.05), cuadro 6.
Los géneros Klebsiella spp, Vibrio spp, Actinobacillus spp y Plesiomonas shigelloides fueron
encontradas en baja proporción, Klebsiella spp, al igual que Enterobacter spp y Serratia spp, es
un genero ampliamente encontrado en fuentes ambientales como suelo, agua y plantas (Grimont
y Grimont 2005b) en insectos también se ha encontrado asociado a dípteros (Demaio y cols.,
1996; Kuzina y cols., 2001), Neuropteros (Dunn y Stabb, 2005), en la cuadro 6, se observa
también que Klebsiella terrigena mostró una frecuencia de recuperación aparentemente diferente
(p<0.05), encontrándose en mayor proporción en alas y cefalotórax sin embargo estos resultados
deben tomarse con reserva debido a bajo número de aislamientos 8.3% (N=2) y 4.2% (N=1)
respectivamente. Por otra parte Vibrio spp, un género halofilico, fue inesperadamente encontrado
principalmente en alas de la mariposa monarca, este hallazgo ha sido infrecuente en otros
trabajos, sin embargo en estudios bacteriológicos en insectos acuáticos se ha logrado recuperar
Vibrio cholerae, el agente responsable del cólera (Halpern y cols., 2006), en el caso de
lepidópteros no hay reportes previos, la identificación de las cepas agrupadas en el genero Vibrio
no mostraron una especie predominante (Vibrio alginolyticus N=2, Vibrio Cholerae No O1 N=2
Vibrio hollisae N=1, Vibrio spp atípico N=3), estudios posteriores serán necesarios para ayudar a
interpretar dichos resultados. Plesiomonas shigelloides se encontró tanto en abdomen como en
alas en frecuencias del 7.3% y 8.3% respectivamente tabla 6, en otros estudios bacteriológicos se
ha recuperado en dípteros (Lindh y cols., 2005).
Bacillus spp es un género caracterizado por la capacidad de producir endosporas muy resistentes
a la desecación, se encuentra ampliamente distribuido en fuentes ambientales como el suelo,
desde el cual puede ser acarreado a través del aire hacia el agua, plantas y otras fuentes
ambientales (Claus and Berkeley 1984) en este estudio fue encontrado en abdomen y alas en
frecuencia, ligeramente mayor al 10%, estudios microbiológicos en mariposas monarca crecidas
en laboratorio han mostrado que la familia Bacillaceae está presente en el adulto (Kingsley
1972), aunque en este estudio no se específica si se asilaron cepas de Bacillus, es muy probable,
ya que este género se ha encontrado como parte de la microbiota en varios insectos tales como:
lepidópteros (McLemore y cols., 2000; Broderick y cols., 2004), dípteros (Kuzina y cols., 2001;

75
Zayed and Bream 2004; Lindh y cols., 2005; Tóth y cols., 2005), himenópteros (Gilliam and
Prest, 1987; Gillian and Taber 1991; Jeyaprakash y cols., 2003) coleópteros (Moore, 1972;
Yilmaz y cols., 2006). Por otro lado también se han encontrado especies de Bacillus spp nocivas
para insectos, muchas de las cuales se emplean en el control biológico, tal es el caso de Bacillus
thuringiensis (Lacey y cols., 2001; Tamez-Guerra y cols., 2004;) debido la producción de
quitinasas así como toxinas que dañan el exoesqueleto de los insectos (Sampson y Gooday 1998),
en el caso de la mariposa monarca se ha observado susceptibilidad a la toxina principalmente en
el primer estadio larval (Hellmich y cols., 2001; Sears y cols., 2001), otras especies
entomopatógenas principalmente en fase larvaria son B. pumilus, B. megaterium, B. cereus
(Ertürk y cols., 2008) B. sphaericus (Myers and Yousten, 1978; Lacey y cols., 2001; Ertürk y
cols., 2008) y B. popilliae (Dutky, 1940; Lacey y cols., 2001). En este estudio se logró el
aislamiento de 4 cepas de B. popilliae, a partir de mariposas aparentemente sanas cuadro 7, se
requerirán estudios posteriores para evaluar las cepas aisladas pueden ser patógenas de la
mariposa monarca.
Los géneros Listeria spp y Streptococcus spp fueron encontrados en frecuencias inferiores al
10%, ambos grupos se ha podido recuperar de dípteros (Kuzina y cols., 2001), adicionalmente,
Streptococcus spp también se ha encontrado asociado con Termitas (Wertz y cols., 2007) y
coleopteros (Yilmaz y cols., 2006).
8.2.1 Diversidad microbiana en diferentes sitios y fases de hibernación.
La mariposa monarca tiene que realizar año con año la migración debido a las bajas temperaturas
registradas en los meses de Invierno en Canadá y EUA (Malcolm, 1987). En los bosques de
Oyamel situados en altitud mayor a los 3000 MSNM las condiciones climáticas permiten la sobre
vivencia de la mariposa monarca, la cual debe ser capaz de responder ante cambios en las
condiciones climáticas, en donde no solo se modifica la temperatura o humedad sino también
disponibilidad de flores para la obtención de néctar (Calvert and Brower 1986; Masters y cols.,
1988). Al analizar el comportamiento de la microbiota conforme avanza el período de
hibernación, cuadro 8, se observa que solo algunas microorganismos aparentemente muestran
disminución en la frecuencia de aislamiento, aunque esta disminución podría ser solo incidental,
también existe la posibilidad de que dicha variación este relacionada con algún cambio en las

76
condiciones de hibernación, de tal manera que la gran mayoría de las especies y géneros
mostrados en los cuadros 6 y 7 mantienen frecuencias de recuperación similares a lo largo de las
tres fases en que se dividió la hibernación.
En cuanto a las especies que sí mostraron variación estadísticamente significativa (p<0.05),
destaca la disminución en la frecuencia de recuperación tanto de Pasteurella bettyae como de
Serratía spp en el periodo inicial al intermedio e intermedio y final respectivamente de la
hibernación, por otra parte el genero Enterococcus spp y la especie Enterobacter taylorae
aparentemente muestran un aumento en la frecuencia de recuperación a medida que avanza el
periodo de hibernación, las razones de esta variación aun no son claras serán necesarios estudios
adicionales que permitan tanto determinar si esta variación en frecuencia es constante así como
esclarecer las causas de esta variación. Por otra parte las especies constantes y presentes en todo
el período de hibernación podrían representar las especies bacterianas bien adaptadas, las cuales
permanecen en la mariposa aun bajo fuertes cambios en las condiciones climáticas. Estos géneros
se encuentran en el cuadro 5 y destacan Enterobacter spp Pasteurella spp y Staphylococcus spp,
ya que todos estos mostraron frecuencias de recuperación en abdomen mayor al 20% durante las
tres temporadas de hibernación de la mariposa monarca. Aunque se espera que durante la
hibernación de la mariposa monarca, la microbiota no cambie sustancialmente, los cambios en las
poblaciones microbianas podrían ser consecuencia del cambio en las condiciones metabólicas del
hospedero encaminadas a cambiar su comportamiento para diversos fines como: el apareamiento
o reemigración.
A pesar de que los sitios de hibernación de la mariposa monarca presentan en común
características como la altura y tipo de vegetación, se sabe que existen cambios en cuanto a
temperaturas y humedad, además del impacto de las comunidades humanas tales como la tala.
Estudios previos han asociado mayor depredación de mariposas en sitios con menor densidad de
árboles (Alonso-Mejía y cols., 1998), a este respecto los cuatro sitios en los que se analizaron
diferencias en la microbiota (El Rosario La Mesa Cerro las Palomas y Sierra Chincua) comparten
la misma densidad de árboles, aunque, la tala ilegal aún es evidente en los dos últimos sitios
(WWF, 2004). Los resultados mostraron mínima variación en la frecuencia de aislamiento para la
mayoría de las especies y géneros. En Sierra chincua se observó una mayor frecuencia de

77
aislamiento de Klebsiella spp (p<0.05), cuadro 10, por otra parte en sitios con poco contacto con
las comunidades humanas tales como La mesa se encontró una mayor frecuencia de aislamiento
de Kingella spp, y Actinobacillus spp así como Serratia odorifera (p<0.05), para el caso de los
gram positivos, no se encontraron géneros o especies cuyas frecuencias de aislamiento fuesen
diferentes en cada sitio de hibernación cuadro 11.
Aunque aun no se han definido sí puede haber una microbiota en insectos y particularmente en
lepidópteros como la mariposa monarca, la mayoría de los trabajos que buscan identificar la
microflora asociada a ciertos insectos, dirigen sus estrategias a determinar que microorganismos
son los mas abundantes (Kingsley, 1972; Demaio y cols., 1996; Woolfolk and Inglis, 2003;
Broderick y cols., 2004; Zayed and Bream, 2004; Wertz and Breznak 2007), que
microorganismos son los mas frecuentemente detectados ya sea por métodos de cultivo o
independientes de este (Gilliam and Prest 1987; Kuzina y cols., 2001; Kittayapong y cols., 2002;
Woolfolk and Inglis, 2003; Stojek and Dutkiewicz, 2004; Zayed and Bream, 2004; Dunn and
Stabb, 2005) cuales microorganismos son mas persistentes frente a cambios morfológicos y
metabólicos en su hospedero (Kingsley, 1972; Lighthart, 1988; Demaio y cols., 1996; Dunn and
Stabb, 2005; Tóth y cols., 2005; Yilmaz y cols., 2006), así como, qué especies se presenten
asociados un insecto en particular independientemente del sitio geográfico donde habite
(González-Ceron y cols., 2003; Wertz and Breznak 2007). En este trabajo se consideraron los
criterios anteriores, tomando en conjunto todos los resultados presentados en este trabajo y
condensándolos en la tabla 15, se concluye que Enterobacter spp, Staphylococcus spp y
Pasteurella spp son probablemente parte de la microbiota asociada a la mariposa monarca
durante el periodo de hibernación en los bosque de oyamel.
8.2.2 Asociación entre O. elektroscirrha con grupos microbianos.
Una de las hipótesis es que la mortandad masiva de las mariposas es debida a cambios climáticos,
aunque se ha visto que el adulto infectado con O. elektroscirrha parece no ser susceptible (Leong
y cols., 1997; Altizer y cols., 1999) aún no se ha estudiado sí la infección de la mariposa con este
parasito le ocasiona vulnerabilidad para adquirir otros patógenos, en este punto se estudió la
microbiota asociada a la mariposa infectada tanto en mariposas vivas como en muertas y en tres
sitios anatómicos, aunque actualmente no se conoce cuales son las especies patógenas para la

78
mariposa monarca, estudios previos en algunos lepidópteros y otros insectos han concluido en el
reconocimiento de ciertas especies bacterianas como patógenas, éstas son: Serratia spp
(Kingsley,1972), Pseudomonas aeroginosa (Lonon y cols., 1988; Jander, y cols., 2000;), Bacillus
thuringiensis (Sampson., 1998; Tamez-Guerra y cols., 2004;), Bacillus popilliae (MacDonald y
cols., 1995), Bacillus sphaericus (Myers y cols., 1978) Acinetobacter calcoaceticus (Kok y cols.,
1995) los resultados generales mostraron el aislamiento de algunas de estas especies tales como
Serratia spp, y Bacillus spp, sin embargo estas especies no fueron aisladas en proporción mayor
en las mariposas parasitadas al compararlas con las no parasitadas (α=0.05) Figura 21. Una razón
que explique los resultados anteriores es que los microorganismos recuperados fueron aislados de
mariposas sanas, o al menos de mariposas que no presentaban daño aparente, tal como pérdida
del color en las alas, ruptura de las alas, patas o antenas etc. Aún falta determinar la proporción
de cepas lipolíticas y quitinolíticas entre los diferentes grupos de mariposas, estos resultados en
conjunto con los obtenidos de mariposas con signos de enfermedad, permitirán identificar las
cepas con potencialidad de ser patógenas para la mariposa monarca.
Cuadro 15. Géneros bacterianos propuestos para ser candidatos a la microbiota de abdomen en la mariposa monarca. x Representa aquellos géneros bacterianos que cumplen los criterios mencionados en la columna de la primer columna. √ Indican los géneros que cumplieron todos los criterios indicados en la primer columna y por lo cual son propuestos a ser parte de la microbiota asociada al abdomen de la mariposa monarca.
Los bacterias que mostraron una mayor frecuencia de aislamiento en mariposas parasitadas
comparadas con las mariposas no parasitadas fueron las especies de Enterobacter spp, Turicella
Acin
eto
bacte
r sp
p
Vib
rio
spp
Ae
rom
ona
s s
pp
En
tero
bacte
r sp
p
Kle
bsie
lla s
pp
Ba
cill
us s
pp
Ku
rth
ia s
pp
Sta
ph
ylo
co
ccu
s s
pp
Mic
rococcus s
pp
Pa
ste
ure
lla s
pp
Se
rra
tia
spp
Co
ryn
eb
acte
rium
spp
Ple
sio
mon
as s
p
Pseud
om
ona
s s
pp
Lis
teri
a s
pp
Ch
ryse
ob
acte
rium
spp
Str
ep
tococcus s
pp
√ √ √
Generos encontrados
Criterios considerados para la propuesta de microbiota asociada a la mariposa monarca
Géneros candidatos a ser parte de la microbiota
xGéneros presentes en el abdomen de la mariposa
monarca en alta carga microbiana > 107 UFC/mariposaGéneros que mostraron alta frecuencia de recuperación en abdomen (mayor al 10%)Géneros que no presentaron variación en la frecuencia de aislamiento con la fase de hibernaciónGéneros cuya variación no fue significativa entre los diferentes sitios de hibernación
x x x x x x x x x x x x x
x x x x x x x x x x
x x x x x x x x x
x x x x x

79
spp, y Eikenella spp. Aun no se ha encontrado antecedentes que asocien la presencia de especies
de estos géneros como patógenos de insectos o lepidópteros, de tal manera que estas cepas serán
investigadas para determinar si producen o no las enzimas hidrolíticas que característicamente
producen los entomopatógenos.
8.2.3 Diferencias entre la microbiota presente en las mariposas vivas, mariposas muertas y
muestras ambientales.
El objetivo de incluir el análisis de la microbiota bacteriana en mariposas muertas fue el obtener
datos respecto a los microorganismos que usualmente podrían encontrarse en mariposas muertas,
dichos microorganismos se asume que pueden ser propios de la mariposa, o bien ser de fuentes
ambientales como el suelo. Ya que cuando un organismo muere, queda a merced de la
colonización como diferentes clases de bacterias y hongos. Ahora considerando que la mariposa
monarca al igual que otros organismos no esta exenta de sufrir infecciones que causen muertes
masivas, es indispensable conocer los microorganismos que usualmente están presentes en
mariposas muertas, de tal manera que en futuras contingencias que ocasionen mortandades
masivas en la mariposa monarca, sea factible inferir cuales bacterias presentes en las mariposas
muertas podrían ser de fuentes no infecciosas y cuales podrías ser microorganismos no usuales,
candidatos a ser la causa de la gran mortandad. En los resultados mostrados en la figura 22 solo
hubo 3 géneros que mostraron diferencia en la frecuencia de aislamiento entre mariposas vivas y
muertas, estos fueron Staphylococcus spp y Enterobacter spp, que se encontraron en mayor
proporción en mariposas vivas, dicha diferencia podría explicarse considerando que estas cepas
puedan ser importantes en el ciclo de la mariposa monarca de tal manera que al morir la
mariposa, esta pierda ciertos factores necesarios para el crecimiento y mantenimiento de estas
cepas, esta hipótesis podría tener mas fuerza para Enterobacter spp, ya que como se observa en el
cuadro 15, dicha bacteria es candidata a ser parte de la microbiota normal de la mariposa.
Por otra parte Cedacea spp fue encontrada prácticamente en mariposas muertas, esta bacteria se
ha encontrado aislado de fuentes ambientales, principalmente en agua en insectos (Grimont y
cols., 2005), de tal manera que es posible que las mariposa muertas sean colonizadas por esta
bacteria presente en el agua del suelo o en el rocío de las plantas.

80
Los resultados del estudio ambiental, tabla 12 muestran que el suelo es la fuente ambiental más
abundante de microorganismos cultivables, seguidos por el agua, corteza y aire, estos resultados
eran esperados ya que en el suelo se encuentra la mayor disponibilidad de nutrientes, por otra
parte en aire se aislaron mas cepas muchas de las cuales resultaros ser de la misma especie.
La identificación de las cepas aisladas aun deberá completarse, sin embargo en la tabla 13 se
observa que en Aire las bacterias mas frecuentemente encontradas corresponden a los géneros
Pseudomonas spp, Klebsiella spp, Enterobacter spp así como Bacillus spp. Estos resultados
concuerdan con lo descrito en trabajos previos (Claus and Berkeley 1984; Garrity y cols., 2005;
Grimont and Grimont, 2005a; Grimont and Grimont, 2005b) por otra parte, dado que en el aire es
frecuente encontrar estos microorganismos, el hecho de encontrar estos microorganismos en
frecuencia alta sobre las alas de la mariposa monarca, (cuadro 6 y 7) puede ser resultado del
contacto de las mariposas con el aire.
En el suelo los principales géneros encontrados corresponden a: Alcaligenes spp, Pseudomonas
spp, Aeromonas spp, Enterobacter spp, Klebsiella spp, Serratia spp, así como Bacillus spp, de
igual manera estos resultados muestran bacterias que usualmente suelen encontrarse en fuentes
ambientales (Claus and Berkeley 1984; Garrity y cols., 2005; Grimont and Grimont, 2005a;
Grimont and Grimont, 2005b) la mayoría de estas bacterias fue recuperada de mariposas tanto
vivas como muertas, figuras 17 y 18, esta ultimas podrían adquirir esta microbiota al ser
colonizadas en el suelo, por otra parte el suelo puede ser arrastrado por el viento y llegar hasta la
mariposa monarca, de tal manera que puedan entrar en contacto con las alas explicando de ese
modo los frecuentes aislamientos de estos microorganismos sobre este sitio anatómico.
En el agua los géneros mas frecuentemente aislados muestra que Aeromonas spp y Enterobacter
spp ambas deberían de ser bacterias frecuentemente recuperadas del abdomen de la mariposa
monarca, ya que en los sitios de hibernación, la mariposa monarca prácticamente lo único que
ingiere es agua, los resultados experimentales concuerdan con lo esperado, en el cuadro 6 se
puede observar que Aeromonas spp se encuentra en al menos 10% de las mariposas así mismo
Enterobacter spp se aisló en frecuencia mayor al 25% de las mariposas, de tal manera que el agua

81
podría ser la fuente de estos bacterias, estudios previos han mostrado que ambas bacterias pueden
ser aisladas de agua (Grimont and Grimont, 2005a).
En el caso de la corteza, no se encontró bacterias que mostraran una frecuencia tan alta como en
las otras fuentes ambientales, es importante mencionar que usualmente el cultivo de
microorganismos de fuentes ambientales es infructoso debido a que mas del 95% de las bacterias
de este tipo de muestras no son cultivables, sin embargo se realizaron los cultivos para poder
correlacionar los hallazgos bacterianos en la mariposa monarca. Así conjuntando todos los
resultados, aparentemente géneros como Seratia spp, Bacillus spp, y Klebsiella spp, podrían ser
parte de la microbiota transitoria, esto debido a que son frecuentemente encontrados en el habitad
de la mariposa monarca, además de que en este lepidóptero pueden aislarse relativamente con
frecuencia, aunque no al grado de Enterobacter spp, Staphylococcus spp o Pasteurella spp,
(cuadro 15).
8.3 Análisis micológico.
Los hongos son un grupo de microorganismos muy abundante en todos los hábitat, en este trabajo
se analizaron e identificaron parcialmente los hongos recuperados del abdomen de la mariposa
monarca, al comparar la cantidad relativa de cepas aisladas se puede observar que el grupo de los
hongos y levaduras, muestran tasas bajas de aislamiento, lo anterior quizá pueda explicarse
debido a hecho de que hay varios hongos cuyo crecimiento es tan acelerado que invaden las
placas y tubos de cultivo, tal es el caso de los pertenecientes a la familia Mucoraceae (Ainsworth,
1973).
Los resultados presentados en el cuadro 14 muestran que aun falta realizar pruebas adicionales en
los hongos y levaduras aisladas, ya que muchos de ellos han mostrado patrones bioquímicos y
morfológicos atípicos, por lo tanto será necesario en un trabajo posterior, emplear pruebas de
biología molecular, basadas en la secuenciación del gen de rRNA18s para lograr la identificación
de estos hongos.
Uno de los pocos hongos completamente identificado es Cladosporium sp (Ainsworth, 1973),
este hongo ha sido encontrado directamente en las improntas de mariposa monarca

82
aparentemente este hongo se encuentra asociado con O. elektroscirrha ya que muchas de las
improntas que presentan este hongo, también presentan el parásito, sin embargo aún no hay datos
suficientes para reforzar esta hipótesis, de tal manera que serán necesarios estudios posteriores.

83
9. Conclusiones
La prevalencia de Ophryocystis elektroscirrha en adultos de Danaus plexippus, ha aumentado de
2.5 a 9 % del 2003 al 2008, dicha prevalencia es homogenea en las colonias y no está relacionada
con el sexo o con el sitio de hibernación.
Es probable que la mortandad natural de la mariposa monarca en condiciones climáticas normales
no está relacionada con la infección con Ophryocystis elektroscirrha, por el contrario los cambios
climáticos, particularmente nevadas, ocasionan mayor muerte entre mariposas infectadas con O.
elektroscirrha que las no infectadas.
La microbiota bacteriana asociada al interior del abdomen de la Mariposa Monarca es mayor
tanto en cantidad como en diversidad a la mostrada en alas y cefalotórax.
Existen especies bacterianas que permanecen constantes tanto en sitios como etapas del periodo
de hibernación, estas especies son Enterobacter spp, Pasteurella spp, y Staphylococcus spp. Estas
especies son candidatas a formar parte de la microbiota de la mariposa monarca.
Especies como Seratia spp, Bacillus spp, y Klebsiella spp podrían ser parte de la microbiota
transitoria en la mariposa monarca ya que se encontraron sistemáticamente en fuentes
ambientales dentro del hábitat de la mariposa monarca.
No se encontró diferencia en la cantidad de especies reconocidas como entomopatógenas,
recuperadas de mariposas parasitadas sin infección con Ophryocystis elektroscirrha, sin embargo
se encontró diferencia para otras bacterias tales como Listeria spp, Pasteurella spp, y
Corynebacterium spp, Eikenella spp, Enterobacter spp y Turicella spp.
Se encontró frecuentemente asociada a la mariposa monarca el hongo Cladosporium spp.

84
10. Bibliografía
• Ainsworth, G. C., F. K. Sparrow and A. S. Jossman. 1973. The fungi an advanced
treatise IVA taxonomic review with keys. Academic Press. 436-453.
• Akhtar, M. W., A. Q. Mirza and I. D. Chughtai. 1980. Lipase Induction in Mucor
hiemalis, Appl. Environ. Microbiol. 40:257-263.
• Alonso-Mejia, A., A. Arellano-Guillermo, and L. P. Brower. 1992. Influence of
Temperature, Surface Body Moisture and Height Aboveground on Survival of Monarch
Butterflies Overwintering in México. Biotropica 24:415-419.
• Alonso-Mejia, A., E. Montesinos-Patiño, E. Rendón-Salinas, L. P. Broker, and K.
Oyama. 1997. Influence of forest Canopo closure on rates of bird predation on
overwintering Monarch butterflies Danaus plexippus. Biol. Conserv. 85: 151-159.
• Altizer, S. M and K. S. Oberhauser 1999 Effects on the Protozoan Parasite
Ophryocystis elektroscirrha on the Fitness of Monarch Butterflies (Danaus plexippus).J.
Inver. Pathol. 74:76-88.
• Altizer, S. M. 2001. Migratory behaviour and host-parasite co-evolution in natural
populations of monarch butterflies infected with a protozoan parasite. Evol. Ecol. Res.,
3:611-632
• Altizer, S. M., S. K. Oberhauser and L. P. Brower. 2000 Association between host
migration and the prevalence of a protozoan parasite in natural populations of adult
monarch butterflies Ecol Entom. 25:125-139
• Arnott, H. J., K. M. Smith, and S. L. Fullilove. 1968. Ultrastructure of a Cytoplasmic
Polyhedrosis Virus Affecting the Monarch Butterfly, Danaus plexippus. J. Ultrastruct.
Res. 24:479-507.
• Bahar A. A., and Z. Demirbag. 2007. Isolation of pathogenic bacteria from Oberea
linearis (Coleoptera: Cerambycidae). Biología. Bratislava. 62:13-18.
• Bradley, C. A., and S. Altizer. 2005, Parasites hinder monarch butterfly flight:
implications for disease spread in migratory hosts. Ecol. Let. 8:290-300.
• Breed, R. S., E. G. D. Murray, and N. R. Smith (ed.), 1957. Bergey’s Manual of
Determinative Bacteriology, 7th ed. Williams and Wilkins, Baltimore MD

85
• Brenner, D. J., N. R. Krieg, J. R. Staley, and G. M. Garrity (ed.), Bergey's Manual of
Systematic Bacteriology, 2da Ed., vol. 2. Springer, New York, NY.
• Broderick, N. A., K. F. Rafia, R. M. Goodman, and J. Handelsman. 2004. Census of
the Bacterial Community of the Gypsy Moth Larval Midgut by Using Culture and
Culture-Independent Methods. Appl. Environ. Microbiol. 70:293-300.
• Brower, L. P., D. R. Kust, E. Rendon-Salinas, E. García-Serrano, K. R. Kust, J.
Miller, C. Fernandez del Rey and K. Pape. 2004. Catastrophic winter storm mortality
of monarch butterflies in Mexico during January 2002. p. 151-166. In K. S. Oberhauser
and M. J. Solensky (ed.), Monarch butterfly biology and conservation. Cornell University
Press, Ithaca, NY.
• Brower, L. P., G. Castilleja, A. Peralta, J. López-García, L. Bojorquez-Tapia, S.
Díaz, D. Melgarejo, and M. Missrie. 2002. Quantitative Changes in Forest Quality in a
Principal Overwintering Area of the Monarch Butterfly in México, 1971-1999.
Conservation in Practice 16:346-359.
• Brower, L. P., P. B. McEvoy, K. L. Williamson, and M. A. Flannery. 1972. Variation
in cardiac glycoside content of monarch butterflies from natural populationes in Eastern
North America. Science. 117:426-428
• Brown, J. J., and G. M. Chippendale. 1974. Migration of the Monarch Butterfly,
Danaus plexippus: Energy Sources. J. Insect. Physiol. 20:1117-1130.
• Calvert, W. H. and L. P. Brower 1986 The localization of monarch butterfly (Danaus
Plexippus L.) overwintering colonies in México in relation to topography and climate.
Journal of the Lepidopterists Society, 40:164-187.
• Calvert, W. H., L. E. Hedrick, and L. P. Brower. 1976. Mortality of the Monarch
Butterfly (Danaus plexippus L.):Avian Predation at Five Overwintering Sites in México.
Science. 204:847-851
• Cazemier, A. E., J. H. P. Hackstein, H. J. M. Op den Camp, J. Rosenberg, and C.
van der Drift. 1997. Bacteria in the Intestinal Tract of Different Species of Arthropods.
Microb. Ecol. 33:189-197.
• CEC. 2008. North American Monarch Conservation Plan. Commission for Environmental
Cooperation

86
• CEC, 2008a. Pérdida y degradación del hábitat de hibernación. In CEC. North American
Monarch Conservation Plan. Commission for Environmental Cooperation. p.27-28.
• CEC, 2008b. Situación legal, manejo y acciones. In CEC. North American Monarch
Conservation Plan. Commission for Environmental Cooperation. p.31-38.
• Cenedella, R. J. 1971. The lipids of the Female Monarch Butterfly, Danaus plexippus,
During Fall Migration. Insect Biochem. 1:244-247.
• Chadwick, J. S., S. S. Caldwell, and P. Chadwick. 1990. Adherence Patterns and
Virulence for Galleria mellonella Larvae of Isolates of Serratia marcescens. J. Invert.
Pathol. 55:133-134.
• Claus, D., and R. C. W. Berkeley. 1984. Genus Bacillus Cohn 1872, 174AL p. 1105-
1139. In N. R. Krieg and J. G. Holt (ed.), Bergey’s manual of systematic bacteriology.
Williams & Wilkins, Baltimore, MD.
• Daniel W. 2006 Bioestadística: Base para el análisis de las ciencias de la salud. 4ª. Ed.
Limusa Wiley. México.
• Dasch, G. A., E. Weiss, and K. P. Chang. 1984. Endosymbionts of insects, p. 811-833.
In N. R. Krieg and J. G. Holt (ed.), Bergey’s manual of systematic bacteriology. Williams
& Wilkins, Baltimore, MD.
• De Roode, J., A. J. Yates, and S. Altizer. 2008. Virulence-transmission trade-offs and
population divergence in virulence in a naturally occurring butterfly parasite. Proc. Natl.
Acad. Sci. 105:7489-7494.
• Demaio, J., C. B. Pumpuni, M. Kent, and J. C. Beier 1996. The midgut bacterial flora
of wild Aedes, Culex pipiens and Psorophora columbiae mosquitoes. Am. J. Trop. Med.
Hyg. 54:219-223.
• Dillon, R. J., and V. M. Dillon. 2004. The Gut Bacteria of Insects: Nonpathogenic
Interactions. Annu. Rev. Entomol. 49:71-92.
• Dugas, J. E., L. Zurek, B. J. Paster, and B. A. Keddie. 2001. Isolation and
characterization of a Chryseobacterium strain from the gut of American cockroach,
Periplaneta americana. Arch. Microbiol. 175:259-262.
• Dunn, A. K., and E. V. Stabb. 2005. Culture-Independent Characterization of the
Microbiota of the Ant Lion Myrmelon mobilis (Neuroptera: Myrmeleontidae). Appl.
Environ. Microbiol. 71:8784-8794

87
• Dutky, S. R. 1940. Two new spore-forming bacteria causing milky diseases of Japanese
beetle larvae. J. Agric. Res. 61:57-68
• Erickson, J. M. 1973. The utilization of various asclepias species by larvae of the
monarch butterfly, Danaus plexippus. Psyche 230-244
• Ertürk, Ö., M. Yaman, and I. Aslan. 2008. Effects of four Bacillus spp. of soil origin
on the Colorado potato beetle Leptinotarsa decemiline (Say). Entomological Research
38:135-138
• Fu, Y., A. S. Ibrahim, W. Fonzi, X. Zhou, C. F. Ramos, and M. A. Ghannoum. 1997.
Cloning and characterization of a gene (LIP1) which encodes a lipase from the pathogenic
yeast Candida albicans. Microbiology. 143:331-340.
• Fukatsu, T., T. Tsuchida, N. Nikoh, and R. Koga. 2001. Spiroplasma Symbiont of the
Pea Aphid, Acyrthosiphon pisum (Insecta: Homoptera). Appl. Environ. Microbiol.
67:1284-1291
• Funke, G., A. von Graeventiz, J. E. Clarridge, and K. A. Bernard. 1997. Clinical
Microbiology of Coryneform Bacteria. Clin. Microbiol. Rev. 10:125-159.
• Funke, G., S. Stubbs, M. Altwegg, A. Carlotti, and M. D. Collins. 1994. Turicella
otitidis gen. nov., sp. nov., a coryneform bactrium isolated from patients with otitis media.
Int. J. Syst. Bacteriol. 44:270-273.
• Funke, M., R. Büchler, V. Mahobia, A. Schneeberg, M. Ramm, and W. Boland.
2008. Rapid hydrolysis of quorum-sensing molecules in the gut of lepidopteran larvae.
Chembiochem. 9:1953-1959
• Galindo-Leal C. y E. Rendón Salinas 2000 Danaidas: Las Maravillosas Mariposas
Monarca. WWF Mexico Telcel Publicación Especial No. 1
• Garrity G. M., J. A. Bell and T. Lilburn. 2005. Genus I Pseudomonasr Migula
1894,237AL. Nom. Cons. Opin. 5 of the Jud. Comm. 1952, 121 p 323-379 In D. J.
Brenner, N. R. Krieg, J. R.Staley and G. M. Garrity (ed.), Bergey's Manual of Systematic
Bacteriology, 2da Ed., vol. 2. part B Springer, New York, NY.
• Gatehouse, H. S., S. D. G. Marshall, R. M. Simpson, L. N. Gatehouse, T. A. Jackson,
and J. T. Christeller. 2008. Serratia entomophila inoculation causes a defect in
exocytosis in Costelytra zealandica larvae. Insect Molecular Biology 17:375-385

88
• Gilliam, M., and D. B. Prest. 1987. Microbiology of Feces of the Larval Honey Bee,
Apis mellifera. J. Invert. Pathol. 49:70-75
• Gilliam, M., and S. Taber. 1991. Diseases, Pests, and Normal Microflora of Honeybees,
Apis mellifera, from Feral Colonies. J. Invert. Pathol. 58:286-289.
• Gillis, M., and N. A. Logan. 2005. Genus IV. Chromobacterium Bergonzoni 1881,
153AL p.824-827. In D. J. Brenner, N. R. Krieg, J. R.Staley and G. M. Garrity (ed.),
Bergey's Manual of Systematic Bacteriology, 2da Ed., vol. 2. part C Springer, New York,
NY
• Glare, T. R., G. E. Corbett, and T. J. Sadler. 1993. Association of a Large Plasmid
with Amber Disease of the New Zeland Grass Grub, Costelytra zealandica, Caused by
Serratia entomophila and Serratia proteamaculans. J. Invert. Pathol. 62:165-170.
• Glendinning, J. I., A. Alonso-Mejia, and L. P. Brower. 1988. Behavioral and
ecological interactions of foraging mice (Peromyscus melanotis) with overwintering
monarch butterflies (Danaus plexippus) in México. Oecologia. 75:222-227.
• González-Ceron, L., F. Santillán, M. H. Rodríguez, D. Méndez, and J. E. Hernández-
Ávila. 2003. Bacteria in midguts of field-collected Anopheles albimanus block
Plasmodium vivax sporogonic development. J. Med. Entomol. 30:371-374.
• Greenberg, B., and M. Klowden. 1972. Enteric bacterial interactions in insects. Am. J.
Clin. Nutr. 25:1459-1466.
• Grimont, P. A. D., F. Grimont, Farmer, and Asbury 2005. Genus IX Cedacea
Grimont, Grimont, Farmer and Asbury 1881a, 325VP (Enteric Group 15 Farmer, Grimont,
Grimont and Asbury 1980b 295). p. 648-651. In D. J. Brenner, N. R. Krieg, J. R.Staley
and G. M. Garrity (ed.), Bergey's Manual of Systematic Bacteriology, 2da Ed., vol. 2. part
B Springer, New York, NY.
• Grimont, P. A. D., and F. Grimont. 2005a. Genus XII Enterobacter Hormaeche and
Edwars 1960b, 72AL. Nom. Cons. Opin. 28, Jud. Comm. 1963, 38 p 661-669 In D. J.
Brenner, N. R. Krieg, J. R.Staley and G. M. Garrity (ed.), Bergey's Manual of Systematic
Bacteriology, 2da Ed., vol. 2. part B Springer, New York, NY.
• Grimont, P. A. D., and F. Grimont. 2005b. Genus XVI Klebsiella Trevisan 1885, 105AL
emend. Drancourt, Bollet, Carta and Roussselier 2001, 930. p. 685-693. In D. J. Brenner,

89
N. R. Krieg, J. R.Staley and G. M. Garrity (ed.), Bergey's Manual of Systematic
Bacteriology, 2da Ed., vol. 2. part B Springer, New York, NY.
• Hajek, A. E. 1999. Pathology and Epizootiology of Entomophaga maimaiga Infections in
forest Lepidoptera. Microbiol. Mol. Biol. Rev. 63:814-835.
• Halpern, M., and O. Landsberg, D. Raats, and E. Rosenberg. 2006. Culturable and
VBNC Vibrio cholerae: Interactions with Chironomid Egg Masses and Their Bacterial
Population. Microbial Ecology. 53:285-293.
• Hellmich, R. L., B. D. Siegfried, M. K. Sears, D. E. Stanley-Horn, M. J. Daniels, H.
R. Mattila, T. Spencer, K. G. Bidne, and L. C. Lewis. 2001. Monarch larvae sensitivity
to Bacillus thuringiensis-purified proteins and pollen. Proc. Natl. Acad. Sci. 98:11925-
11930.
• Herniou, E. A., J. A. Olszewski, D. R. O’Reilly, and J. S. Cory. 2004. Ancient
Coevoution of Baculoviruses nad Their Insect Hosts. J. Virol. 78:3244-3251.
• Holder-Francklin, M. A., A. Thorpe, and L. Wuest. 1992. Evaluatión of test employed
in the numerical taxonomy of river bacteria. Journal of Microbiological Methods 15:263-
277.
• Horn, D. J. 1978. Biology of Insects. 1st ed. W. B. Saunders Company Philadelphia PA.
p. 171-193.
• Indiragandhi, P., R. Anandham, M. Madhaiyan, and T. M. Sa. 2008. Characterization
of Plant Growth-Promoting Traits of Bacteria Isolated from Larval Guts of Diamondback
Moth Plutella xylostella (Lepidoptera: Plutellidae). Curr. Microbiol. 56:327-333
• Jackson, T. A., D. G. Boucias, and J. O. Thaler. 2001. Pathobiology of Amber Disease,
Caused by Serratia spp., in the New Zealand Grass Grub, Costelytra zealandica. J. Invert.
Pathol. 78:232-243.
• Janda J. M. 2005. Genus XXVII. Plesiomonas Habs and Schubert 1962, 324AL. p 740-
744 In D. J. Brenner, N. R. Krieg, J. R.Staley and G. M. Garrity (ed.), Bergey's Manual of
Systematic Bacteriology, 2da Ed., vol. 2. part B Springer, New York, NY.
• Jander, G., L. G. Rahme, and F. M. Ausubel. 2000. Positive Correlation between
Virulence of Pseudomonas aeroginosa Mutants in Mice and Insects. J. Bacteriol.
182:3843-3845.

90
• Jeyaprakash, A., M. A. Hoy, and M. H. Allsopp. 2003. Bacterial diversity in worker
adults of Apis mellifera capensis and Apis mellifera scutellata (Insecta: Himenoptera)
assessed using 16S rRNA sequences. J. Invert. Pathol. 84:96-103.
• Jiggins, F. M., G. D. Hurst, C. D. Jiggins, J. H. v d Schulenburg, and M. E. Majerus.
The butterfly Danaus chrysippus by male-killing Spiroplasma. Parasitology. 120:439-446
• Keddie M. R., and S. Shaw. 1984. Genus Kurthia Trevisan 1885, 92AL Nom. cons.
Opin. 13 Jud. Comm. 1954, 152. p. 1255-1258 In N. R. Krieg and J. G. Holt (ed.),
Bergey’s manual of systematic bacteriology. Williams & Wilkins, Baltimore, MD.
• Kingsley, V. V. 1972. Persistence of Intestinal Bacteria in the Developmental Stages of
the Monarch Butterfly (Danaus plexippus) J.Invert. Pathol. 20:51-58
• Kirsh, D., 1934. Factors influencing the Activity of Fungus Lipase. J. Biol. Chem. 421-
430.
• Kittayapong, P., V. Baimai, and S. O’Neill. 2002. Field prevalence of Wolbachia in the
mosquito vector Aedes albopictus. Am. J. Trop.Med. Hyg. 66, 108-111.
• Kloos, W. E., and K. H. Schleifer. 1984. Genus IV Staphylococcus Rosenbach 1884,
18AL p. 1013-1035. In N. R. Krieg and J. G. Holt (ed.), Bergey’s manual of systematic
bacteriology. Williams & Wilkins, Baltimore, MD.
• Koch, R. L., W. D. Hutchison, R. C. Venette, and G. E. 2003. Susceptibility of
immature monarch butterfly, Danaus plexippus (Lepidoptera: Nymphalidae Danainae), to
predation by Harmonia axyridis (Coleoptera: Coccinellidae). Biological Control. 28:265-
270.
• Kok, R.G., J. J. van Thor, I. M. Nugteren-Roodzant, M. B. Brouwer, M. R. Egmond,
C. B. Nudel, B. Vosman, and K. J. Hellingwerf. 1995. Characterization of the
extracellular lipase, LipA, of Acinetobacter calcoaceticus BD413 and sequence analysis
of the cloned structural gene. Mol. Microbiol. 5:803-818.
• Krieg, N. R., and J. G. Holt (ed.). 1984, Bergey’s manual of systematic bacteriology.
vol 1. Williams & Wilkins, Baltimore, MD.
• Kuzina, L. V., J. J. Peloquin, D. C. Vacek, and T. A. Miller. 2001. Isolation and
Identification of Bacteria Associated with Adult Laboratory Mexican Fruit Flies,
Anastrepha ludens (Diptera: Tephritidae). Curr. Microbiol. 42:290-294.

91
• Lacey, L. A., R. Frutos, H. K. Kaya, and P. Vail. 2001. Insect Pathogens as Biological
Control Agents: Do They Have a Future? Biological Control. 21:230-248.
• Leong, K. L. H., H. K Kaya, M. A. Yoshimura, and D. F. Frey. 1992. The occurrence
and effect of a protozoan parasite, Ophryocystis electroscirrha (Neogregarinida:
Ophryocystidae) on overwintering monarch butterflies, Danaus plexippus (Lepidoptera:
Danaidae) from two California winter sites Ecol. Entomol. 17:338-342.
• Leong, K. L. H., M. A. Yoshimura, H, K. Kaya and H. Williams 1997. Instar
Susceptibility of the Monarch Butterfly. J. Invertebr. Pathol. 69:79-83.
• Lighthart, B. 1988. Some Changes in Gut Bacterial Flora of Field-Grown Peridroma
saucia (Lepidoptera: Noctuidae) When Brought into the Laboratory. Appl. Environ.
Microbiol. 54:1896-1898
• Lindh, J. M., O. Terenius, and I. Faye. 2005. 16S rRNA Gene-Based Identification of
Midgut Bacteria from Field-Caught Anopheles gambiae Sensu Lato and A. funestus
Mosquitoes Reveals New Species Related to Know Insect Symbionts. Appl. Environ.
Microbiol. 71:7217-7223.
• Lonon, M. K., E. W. Donald, and D. C. Straus. 1988. Production of Lipase by Isolates
of Pseudomonas cepacia. J. Clin. Microbiol. 26:979-984.
• Lysenko, O. 1976. Chitinase of Serratia marcescens and Its Toxicity to Insects. J.Invert.
Pathol. 27:385-386
• Mac Faddin, J. F.1985. Media for Isolation cultivation-identification-Maintenance of
Medical Bacteria. Vol1 Williams & Wilkins, Baltimore, MD.
• Malcolm, S. B. 1987. Monarch Butterfly Migration in North America: Controversy and
Conservation. Tree 2:135-138
• Martin, P. A., D. Gundersen-Rindal, M. Blackburn, and J. Buyer. 2007a.
Chromobacterium subtsugae sp.nov., a betaproteobacterium toxic to Colorado potato
beetle and other insect pests. Int. J. Syst. Evol. Microbiol. 57:993-1009.
• Martin, P. A., E. Hirose, and J. R. Aldrich. 2007b. Toxicity of Chromobacterium
subtsugae to southern green stink bug (Heteroptera: Pentatomidae) and corn rootworm
(coleoptera: Chrysomelidae). J. Econ. Entomol. 100:680-684.

92
• Martínez M. F., 2007. Informe Final de monitoreo de mariposa monarca en México,
Durante la temporada de hibernación 2007-07. Reserva de la Biosfera Mariposa Monarca.
CONANP México. D.F.
• Masters, A. R., S. B. Malcolm, and L. P. Brower. 1988. Monarch Butterfly (Danaus
plexippus) thermoregulatory behavior and Adaptations for overwintering in Mexico.
Ecology. 69:458-467
• McLaughlin R. E. and J. Meyers 1970 Ophryocystis elektroscirrha sp.n., a
neogragarine pathogen of the monarch butterfly Danaus Plexippus (L.) and the Florida
queen butterfly D. gilippus berenice Cramer. J. of Protozoology, 17:300-305.
• McLemore, J. B., F. H. Tainter, and S. D. McElreath. 2000. Bacteria associated with
butterfly stain of Chilean tepa. Curr. Microbiol. 40:86-90.
• Moore G. E., 1972. Microflora from the Alimentary Tract of Healthy Southern Pine
Beatles, Dendroctonus frontalis (Scolytidae), and Their Posible Ralationship to
patogenicity. J. Invert. Pathol.19:72-75.
• Mutters, R., H. Christensen, and M. Bisgaard. 2005 Genus I Pasteurella Trevisan
1887, 94AL Nom. Cons. Opin. 13, Jud. Comm. 1954b, 153. p. 857-866. In D. J. Brenner,
N. R. Krieg, J. R.Staley and G. M. Garrity (ed.), Bergey's Manual of Systematic
Bacteriology, 2da Ed., vol. 2. part B Springer, New York, NY.
• Myers, P., and A. A. Yousten. 1978. Toxic Activity of Bacillus sphaericus SSII-1 for
Mosquito Larvae. Infect. Immun. 19:1047-1053.
• O’Callaghan, M., M. L. Garnham, T. L. Nelson, D. Baird, and T. A. Jackson. 1996.
The Pathogenicity of Serratia Strains to Lucilia sericata (Diptera: Calliophoridea). J.
Invert. Pathol. 66:22-27
• Prado, B., A. Jara, A. D. Morral, and E. Sánchez. 2001. Numerical Taxonomy of
Microorganisms Isolated from Goat Cheese Made in Chile. Curr. Microbiol. 43:396-399.
• Renaud F. N. R., A. Grégory, C. Barreau, D. Aubel, and J. Frenay. 1996.
Identification of Turicella otitidis Isolated from a Patient with Otorrhea Associated with
Surgery: Differentiation from Corynebacterium afermentans and Corynebacterium auris.
J. Clin. Microbiol. 34:2625-2627.

93
• Rendón-Salinas, E., S. Rodríguez-Mejía, M. Cruz-Piña, and C. Galindo-Leal. 2006.
Monitoreo de las colonias de hibernación de Mariposa Monarca: Superficie Forestal de
Ocupación en diciembre de 2005 Reporte WWF 2006.
• Reyes, V. F. 2004. Generalidades y potencialidad en biocontrol de las gregarinas
entomoparasitas. Ciencia UANL. 7:355-360.
• Richards O. W., and R. G. Davies. 1977. Order 27 Lepidoptera (Butterflies and Moths).
p. 1072-1099. In O. W. Richards and R. G. Davies. General textbook of entomology. 10th
ed., vol. 2. Chapman and Hall, London.
• Rohlf F.J., and R. R. Sokal. 1981. Comparing numerical taxonomic studies. Syst. Zool.
30:459-490
• Sampson, M. N., and G. W. Gooday. 1998. Involvement of chitinases of Bacillus
thuringiensis during pathogenesis in insects. Microbiology. 144:2189-2194.
• Savage, D. C. 1977. Microbial Ecology of the Gastrointestinal Tract. Ann. Rev.
Microbiol. 31:107-133
• Schreckenberger, P. C., and A. Graevenitz. 1999. Acinetobacter, Achromobacter,
Alcaligenes, Moraxella, Methylobacterium, and Other Nonfermentative Gram-Negative
Rods. p. 539-560. In P. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover and R. H.
Yolken. (ed.), Manual of Clinical Microbiology 7th ed. ASM Press, Washington, DC.
• Sears, M. K., R. L. Hellmich, D. E. Stanley-Horn, K. S. Oberhauser, J. M. Pleasants,
H. R. Mattila, B. D. Siegfried, and G. P. Dively. 2001. Impact of Bt corn pollen on
monarch butterfly populations: A risk assessment. Proc. Natl. Acad. Sci. 98:11937-11942.
• SEMARNAT, 2001. Programa de Manejo Reserva de la Biosfera Mariposa Monarca.
CONANP. México. D. F.
• Sikorowski, P. P., and A. M. Lawrence. 1998. Transmission of Serratia marcescens
(Enterobacteriaceae) in Adult Heliothis virescens (Lepidoptera: Noctuidae) Laboratory
Colonies. Biological control 12:50-55.
• Sneath, P. H. A. 2005. Numerical Taxonomy. p.39-42. In D. J. Brenner, N. R. Krieg, J.
R.Staley and G. M. Garrity (ed.), Bergey's Manual of Systematic Bacteriology, 2da Ed.,
vol. 2. part A Springer, New York, NY.
• Sokal, R. R. 1962. The principles and practice of Numerical Taxonomy. Contribution
1192 from the Department of Entomology. University of Kansas Lawrence Kansas.

94
• Stojek, N. M., and J. Dutkiewicz. 2004. Studies on the occurrence of Gram-negative
bacteria in ticks:Ixodes ricinus as a potential vector of Pasteurella. Ann. Agric. Environ.
Med. 11:319–322.
• Tamez-Guera P., M. M. Iracheta, B. Pereyra-Alferez, L. J. Galan-Wong, R. Gomez-
Flores, R. S. Tames Guerra and C. Rodríguez-Padilla. 2004. Caracterization of
Mexican Bacillus thuringiensis strain toxic for lepidopteran and coleopteran larvae J.
Invert. Pathol. 86:7-18.
• Thaler J., B. Duvic, A. Givaudan, and N. Boemare. 1998. Isolation and Entomotoxic
Propieties of the Xenorhabdus nematophilus F1 Lecithinase. Appl. Environ. Microbiol.
64:2367-2373.
• Toth, E. M., E. Hell, G. Kovács, A. K. Borsodi, and K. Márialigeti. 2005. Bacteria
Isolated from the Different Developmental Stages and Larval Organs of the Obligate
Parasitic Fly, Wohlfahrtia magnifica (Díptera: Sarcophagidae). Microbial Ecology 51:13-
21
• Urquhart, F. A. 1966. Virus-Caused Epizootic as a Factor in Population Fluctuations of
the Monarch Butterfly. J. Invertebr. Pathol. 8, 492-495
• Wassenaar L. I., K. A. Hobson 1998 Natal origins of migratory monarch butterflies at
wintering colonies in Mexico: New isotopic evidence. Proc. Natl. Acad.Sci USA
95:15436-15439.
• Wertz, J. T., and J. A. Breznak. 2007. Stenoxybacter acetivorans gen. nov., sp. nov., an
Acetate-Oxidizing Obligate Microaerophile among Diverse O2-Consuming Bacteria from
Termite Guts. Appl. Emviron. Microbiol. 73:6819-6828.
• Woolfolk, S. W., and G. D. Inglis. 2003. Microorganisms associated with field-collected
Chrysoperla rufilabris (Neuroptera: Chrysopidae) adults with emphasis on yeast
symbionts. Biological control 29:155-168.
• WWF, 2004. La tala ilegal y su impacto en la Reserva de la Biosfera Mariposa Monarca.
WWF-Mexico.
• Yilmaz, H., K. Sezen, H. Kati, and Z. Demirbag. 2006. The first study on the bacterial
flora of the European spruce bark beetle, Dendroctonus micans (Coleoptera: Scolytidae)
Biología Bratislava 61:679-686.

95
• Zayed, M. E., and A. S. Bream. 2004. Biodiversity of the microbial flora associated with
two strains of Culex pipiens (Diptera: Culidae). Commun. Agric. Appl. Biol. Sci. 69:229-
234.
• Zepeda-Lopez H. M., A. L. Sandoval, J. Bravo, A. Márquez, D. Ramírez, G. Jiménez
D. Soriano, S. Rodríguez, R. Mondragón, M. Bernal, E. Rendón, and A. A. Aguirre.
2005. Mortality of Wintering Monarch Butterflies: Are Emerging Pathogens the Cause of
Declining Populations?, abstr. Z-005, p.611-612. Abstr. 105th Gen. Meet. Am. Soc.
Microbiol. American Society for Microbiology, Washington, DC.

96
Apéndice A. Tabla A. Total de bacterias encontradas en abdomen, alas y cefalotórax de mariposas monarca Continúa...
Cefalotorax
Bacteria 96B ( 100.0% )C 24 ( 100.0% ) 23 ( 100.0% )Proteobacteria 90 ( 93.8% ) 20 ( 83.3% ) 20 ( 87.0% )
Betaproteobacteria 5 ( 5.2% ) 4 ( 16.7% ) 2 ( 8.7% )Burkholderiales 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )
Alcaligenaceae 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Alcaligenes spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Alcaligenes faecalis 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Bordetella spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Bordetella otras especies 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Comamonadaceae 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Comamonas spp 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Comamonas acidovorans 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Neisseriales 3 ( 3.1% ) 3 ( 12.5% ) 2 ( 8.7% )Neisseriaceae 3 ( 3.1% ) 3 ( 12.5% ) 2 ( 8.7% )
Eikenella spp 2 ( 2.1% ) 3 ( 12.5% ) 0 ( 0.0% )Eikenella corrodens 2 ( 2.1% ) 3 ( 12.5% ) 0 ( 0.0% )
Kingella spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Kingella otras especies 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Chromobacterium spp 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 8.7% )Chromobacterium violaceum 0 ( 0.0% ) 0 ( 0.0% ) 2 ( 8.7% )
Gammaproteobacteria 86 ( 89.6% ) 20 ( 83.3% ) 18 ( 78.3% )Xantomonadales 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Xantomonadaceae 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Stenotrophomonas spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Stenotrophomonas maltophilia 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Pseudomonadales 20 ( 20.8% ) 5 ( 20.8% ) 1 ( 4.3% )
Pseudomonadaceae 9 ( 9.4% ) 5 ( 20.8% ) 1 ( 4.3% )Pseudomonas spp 9 ( 9.4% ) 5 ( 20.8% ) 1 ( 4.3% )
Pseudomonas aeroginosa 6 ( 6.3% ) 3 ( 12.5% ) 1 ( 4.3% )Pseudomonas fluorescens 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Pseudomonas stutzeri 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Pseudomonas otras especies 1 ( 1.0% ) 2 ( 8.3% ) 0 ( 0.0% )
Moraxelaceae 11 ( 11.5% ) 1 ( 4.2% ) 0 ( 0.0% )Acinetobacter spp 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )
Acinetobacter baumannii 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Acinetobacter otras especies 5 ( 5.2% ) 0 ( 0.0% ) 0 ( 0.0% )Acinetobacter calcoaceticus 5 ( 5.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Moraxella spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Moraxella otras especies 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Alteromonadales 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Alteromonadaceae 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Shewanella spp 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Shewanella putrefaciens 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Vibrionales 4 ( 4.2% ) 3 ( 12.5% ) 1 ( 4.3% )Vibrionaceae 4 ( 4.2% ) 3 ( 12.5% ) 1 ( 4.3% )
Vibrio spp 4 ( 4.2% ) 3 ( 12.5% ) 1 ( 4.3% )Vibrio alginolyticus 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Vibrio alginolyticus atípico 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Vibrio cholerae 0 ( 0.0% ) 1 ( 4.2% ) 1 ( 4.3% )Vibrio hollisae 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Vibrio otras especies 3 ( 3.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Aeromonadales 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )Aeromonadaceae 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )
Aeromonas spp 10 ( 10.4% ) 1 ( 4.2% ) 0 ( 0.0% )Aeromonas hydrophila 7 ( 7.3% ) 0 ( 0.0% ) 0 ( 0.0% )Aeromonas hydrophila grupo 2 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )Aeromonas salmonicida ssp salmonicida 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterobacteriales 62 ( 64.6% ) 16 ( 66.7% ) 17 ( 73.9% )Enterobacteriaceae 62 ( 64.6% ) 16 ( 66.7% ) 17 ( 73.9% )
Cedacea spp 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )Cedacea sp 3 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Citrobacter spp 3 ( 3.1% ) 0 ( 0.0% ) 0 ( 0.0% )Citrobacter otras especies 3 ( 3.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Edwarsiella spp 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Edwarsiella hoshinae 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Enteric Group 69 5 ( 5.2% ) 1 ( 4.2% ) 0 ( 0.0% )
N=23Bacterias aisladas
Abdomen N=96 A
Alas N=24

97
Continúa...
Cefalotorax
Enterobacter spp 25 ( 26.0% ) 9 ( 37.5% ) 11 ( 47.8% )Enterobacter aerogenes 10 ( 10.4% ) 8 ( 33.3% ) 10 ( 43.5% )Enterobacter cancerogenus 3 ( 3.1% ) 1 ( 4.2% ) 0 ( 0.0% )Enterobacter cloacae 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )Enterobacter dissolvens 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Enterobacter sakazakii 4 ( 4.2% ) 0 ( 0.0% ) 1 ( 4.3% )Enterobacter taylorae 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Enterobacter otras especies 6 ( 6.3% ) 0 ( 0.0% ) 0 ( 0.0% )
Klebsiella spp 8 ( 8.3% ) 2 ( 8.3% ) 2 ( 8.7% )Klebsiella ornithinolytica 1 ( 1.0% ) 1 ( 4.2% ) 1 ( 4.3% )Klebsiella ozaenae 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Klebsiella planticola 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Klebsiella pneumoniae 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Klebsiella terrigena 0 ( 0.0% ) 2 ( 8.3% ) 1 ( 4.3% )Klebsiella otras especies 4 ( 4.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Kluyvera spp 2 ( 2.1% ) 0 ( 0.0% ) 1 ( 4.3% )Kluyvera ascorbata 2 ( 2.1% ) 0 ( 0.0% ) 1 ( 4.3% )
Moellerella spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Moellerella wisconsensis 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Obesumbacterium spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Obesumbacterium proteus biogrupo 2 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Plesimonas sp 7 ( 7.3% ) 2 ( 8.3% ) 0 ( 0.0% )Plesiomonas shigelloides 7 ( 7.3% ) 2 ( 8.3% ) 0 ( 0.0% )
Rahnella spp 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Rahnella acuatilis 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )
Salmonalla spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Salmonella choleraesuis sub gpo 3b arizonae atípica 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Serratia spp 24 ( 25.0% ) 3 ( 12.5% ) 4 ( 17.4% )Serratia entomophila 4 ( 4.2% ) 1 ( 4.2% ) 2 ( 8.7% )Serratia fonticola 3 ( 3.1% ) 1 ( 4.2% ) 0 ( 0.0% )Serratia liquefaciens 4 ( 4.2% ) 0 ( 0.0% ) 1 ( 4.3% )Serratia marcescens 10 ( 10.4% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia marcescens atípica 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia odorifera 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia odorifera atípica 2 ( 2.1% ) 0 ( 0.0% ) 1 ( 4.3% )Serratia plymuthica 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Serratia rubidaea 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )Serratia otras especies 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Yokenella spp 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Yokenella regensburgei 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Pasteurellaes 31 ( 32.3% ) 3 ( 12.5% ) 0 ( 0.0% )Pasteurellaceae 31 ( 32.3% ) 0 ( 0.0% ) 0 ( 0.0% )
Actinobacillus spp 6 ( 6.3% ) 0 ( 0.0% ) 0 ( 0.0% )Actinobacillus otras especies 6 ( 6.3% ) 0 ( 0.0% ) 0 ( 0.0% )
Pasteurella spp 29 ( 30.2% ) 3 ( 12.5% ) 0 ( 0.0% )Pasteurella bettyae 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Pasteurella multocida 1 3 ( 3.1% ) 1 ( 4.2% ) 0 ( 0.0% )Pasteurella multocida 2 1 ( 1.0% ) 2 ( 8.3% ) 0 ( 0.0% )Pasteurella otras especies 23 ( 24.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Firmicutes 55 ( 57.3% ) 9 ( 37.5% ) 6 ( 26.1% )Bacilli 55 ( 57.3% ) 9 ( 37.5% ) 6 ( 26.1% )
Bacillales 54 ( 56.3% ) 9 ( 37.5% ) 8 ( 34.8% )Bacillaceae 11 ( 11.5% ) 3 ( 12.5% ) 4 ( 17.4% )
Bacillus spp 11 ( 11.5% ) 3 ( 12.5% ) 0 ( 0.0% )Bacillus cereus 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Bacillus fastidiosus 2 ( 2.1% ) 1 ( 4.2% ) 1 ( 4.3% )Bacillus licheniformis 2 ( 2.1% ) 0 ( 0.0% ) 1 ( 4.3% )Bacillus popilliae 1 ( 1.0% ) 2 ( 8.3% ) 1 ( 4.3% )Bacillus schlegelii 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Bacillus otras especies 4 ( 4.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Planococcaceae 13 ( 13.5% ) 0 ( 0.0% ) 0 ( 0.0% )Kurthia spp 13 ( 13.5% ) 0 ( 0.0% ) 0 ( 0.0% )
Kurthia otras especies 13 ( 13.5% ) 0 ( 0.0% ) 0 ( 0.0% )Listeriaceae 1 ( 1.0% ) 2 ( 8.3% ) 2 ( 8.7% )
Listeria spp 1 ( 1.0% ) 2 ( 8.3% ) 2 ( 8.7% )Listeria denitrificans 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Listeria welshimeri 1 ( 1.0% ) 1 ( 4.2% ) 2 ( 8.7% )
Bacterias aisladas Abdomen Alas
N=96 A N=24 N=23

98
A Cantidad total de mariposas analizadas B número de aislamientos C frecuencia en porcentaje.
Cefalotorax
Staphylococcaceae 29 ( 30.2% ) 5 ( 20.8% ) 0 ( 0.0% )Staphylococcus spp 29 ( 30.2% ) 5 ( 20.8% ) 0 ( 0.0% )
Staphylococcus auricularis 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus camosus 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus cohnii ssp. Cohnii 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Staphylococcus haemolyticus 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus intermedius 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus lentus 8 ( 8.3% ) 0 ( 0.0% ) 0 ( 0.0% )Staphylococcus lugdunensis 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Staphylococcus sciuri 0 ( 0.0% ) 1 ( 4.2% ) 0 ( 0.0% )Staphylococcus warneri 1 ( 1.0% ) 2 ( 8.3% ) 0 ( 0.0% )Staphylococcus otras especies 15 ( 15.6% ) 0 ( 0.0% ) 0 ( 0.0% )
Lactobacillales 16 ( 16.7% ) 2 ( 8.3% ) 2 ( 8.7% )Lactobacillaceae 3 ( 3.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Lactobacillus spp 3 ( 3.1% ) 0 ( 0.0% ) 0 ( 0.0% )Lactobacillus confusus 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Lactobacillus confusus atípica 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Lactobacillus minor 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterococcaceae 4 ( 4.2% ) 0 ( 0.0% ) 0 ( 0.0% )Enterococcus spp 4 ( 4.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Enterococcus dispar 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterococcus durans 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterococcus faecalis 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Enterococcus otras especies 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Streptococcaceae 9 ( 9.4% ) 2 ( 8.3% ) 2 ( 8.7% )Streptococcus spp 9 ( 9.4% ) 0 ( 0.0% ) 2 ( 8.7% )
Streptococcus bovis 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )Streptococcus equi 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Streptococcus mitis 1 ( 1.0% ) 1 ( 4.2% ) 0 ( 0.0% )Streptococcus salivarius 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Streptococcus sanguis biotipo 2 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Streptococcus otras especies 5 ( 5.2% ) 0 ( 0.0% ) 0 ( 0.0% )
Actinobacteria 43 ( 44.8% ) 2 ( 8.3% ) 8 ( 34.8% )Actinobacteria 43 ( 44.8% ) 2 ( 8.3% ) 8 ( 34.8% )
Actinomycetales 43 ( 44.8% ) 2 ( 8.3% ) 8 ( 34.8% )Actinomycetaceae 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )
Micrococcaceae 10 ( 10.4% ) 0 ( 0.0% ) 5 ( 21.7% )Micrococcus spp 10 ( 10.4% ) 0 ( 0.0% ) 5 ( 21.7% )Micrococcus agilis 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Micrococcus kristinae 3 ( 3.1% ) 0 ( 0.0% ) 3 ( 13.0% )Micrococcus luteus 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )Micrococcus roseus 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Micrococcus otras especies 5 ( 5.2% ) 0 ( 0.0% ) 0 ( 0.0% )Microbacteriaceae 2 ( 2.1% ) 2 ( 8.3% ) 2 ( 8.7% )
Agromyces spp 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )Agromyces ramosus 2 ( 2.1% ) 1 ( 4.2% ) 0 ( 0.0% )
Curtobacterium spp 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Curtobacterium citreum 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )
Microbacterium spp 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Microbacterium leavaniformans 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )
Corynebacteriaceae 34 ( 35.4% ) 1 ( 4.2% ) 2 ( 8.7% )Corynebacterium spp 34 ( 35.4% ) 1 ( 4.2% ) 2 ( 8.7% )
Corynebacterium afermentans subsp afermentans 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Corynebacterium kroppenstedtii 1 ( 1.0% ) 0 ( 0.0% ) 0 ( 0.0% )Corynebacterium paurometabolum 0 ( 0.0% ) 0 ( 0.0% ) 1 ( 4.3% )Corynebacterium pseudodiphtheriticum 2 ( 2.1% ) 1 ( 4.2% ) 1 ( 4.3% )Corynebacterium otras especies 30 ( 31.3% ) 0 ( 0.0% ) 0 ( 0.0% )
Nocardiaceae 3 ( 3.1% ) 0 ( 0.0% ) 1 ( 4.3% )Nocardia spp 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )
Nocardia otras especies 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )Rhodococcus spp 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )
Rhodococcus otras especies 2 ( 2.1% ) 0 ( 0.0% ) 0 ( 0.0% )Bifidobacteriales 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )
Afiliación desconocida 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )Turicella spp 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )
Turicella otitidis 1 ( 1.0% ) 0 ( 0.0% ) 4 ( 17.4% )Bacterioidetes 1 ( 1.0% ) 1 ( 4.2% ) 5 ( 21.7% )
Flavobacteria 1 ( 1.0% ) 1 ( 4.2% ) 5 ( 21.7% )Flavobacteriales 1 ( 1.0% ) 1 ( 4.2% ) 5 ( 21.7% )
Flavobacteriaceae 1 ( 1.0% ) 1 ( 4.2% ) 5 ( 21.7% )Chryseobacterium spp 1 ( 1.0% ) 1 ( 4.2% ) 5 ( 21.7% )
Chryseobacterium indologenes 1 ( 1.0% ) 0 ( 0.0% ) 1 ( 4.3% )Chryseobacterium meningosepticum 0 ( 0.0% ) 1 ( 4.2% ) 4 ( 17.4% )
N=23Bacterias aisladas
Abdomen Alas N=96 A N=24







