

genetická diverzita člověka
na úrovni SNP je nízká: asi 0.1% cca. 90% variace je uvnitř populací
cca. 10% mezi populacemi (kontinenty)
bottleneck
Nature
Science
variabilita genomu

personal genomes
PGP-10 George Church George Church, professor of genetics, Harvard Medical School.
Misha Angrist, scientific editor at the Duke Institute for Genome Sciences and Policy in Durham.
Keith Batchelder, CEO at Genomic Healthcare Strategies.
Esther Dyson, investor and adviser to multiple technology firms.
Rosalynn Gill-Garrison, chief science officer at Sciona.
John Halamka, chief information officer at Harvard Medical School.
Stanley Lapidus, chief executive officer of Helicos.
Kirk Maxey, president of Cayman Chemical.
James Sherley, stem cell researcher and associate professor, formerly of MIT.
individual #10
James Watson
Craig Venter
Marjolein Kriek
Afričané, Číňané, Korejci, Japonci, genomy nádorů
1000 Genomes (~2500) Wellcome Trust Sanger Institute in Hinxton, England,
Beijing Genomics Institute Shenzhen in China,
NIH National Human Genome Research Institute (NHGRI)
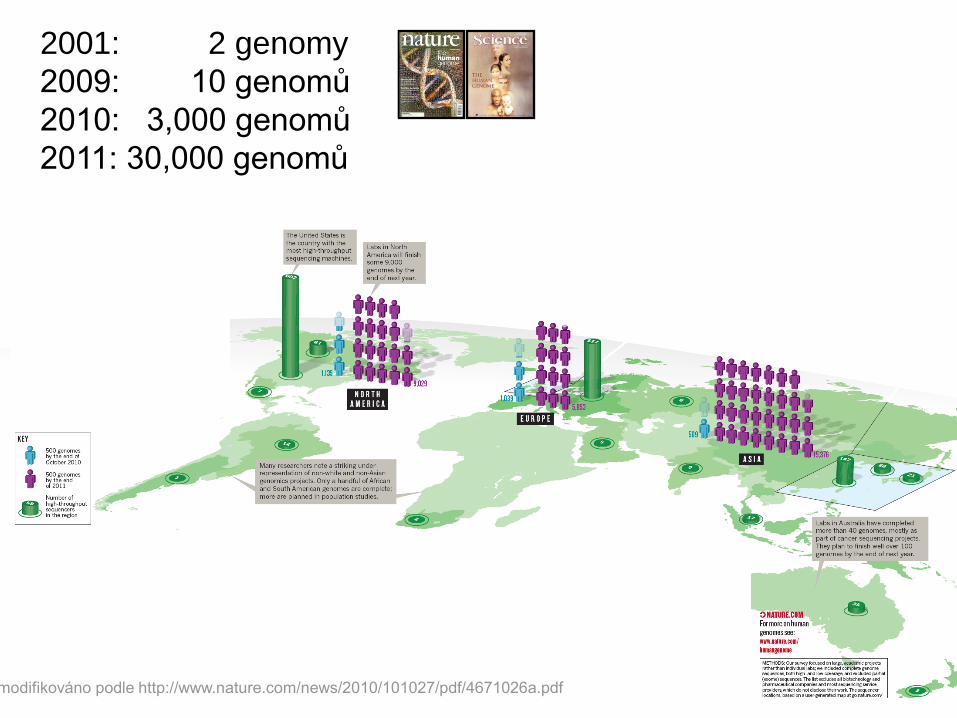
2001: 2 genomy
2009: 10 genomů
2010: 3,000 genomů
2011: 30,000 genomů
modifikováno podle http://www.nature.com/news/2010/101027/pdf/4671026a.pdf
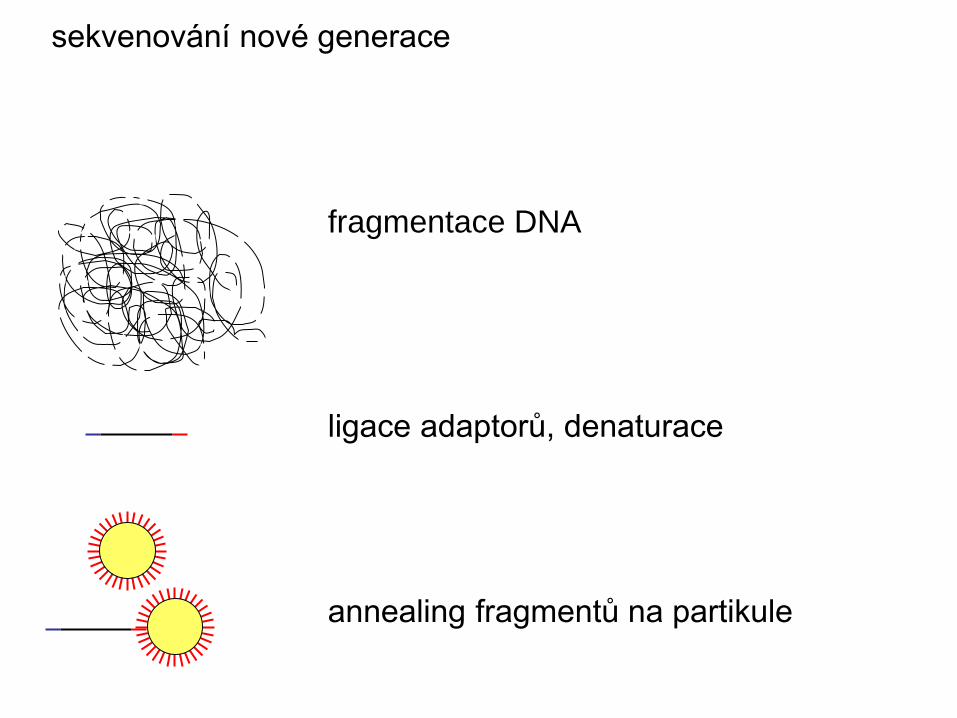
fragmentace DNA
ligace adaptorů, denaturace
annealing fragmentů na partikule
sekvenování nové generace

vytvoření emulze
emulzní PCR, denaturace

zachycení partikulí v pikotitrační desce

pyrosekvenace a detekce záblesků

pyrosekvenace a detekce záblesků
DNA(n) + dNTP (polymeráza) DNA(n+1) + PPi
APS + PPi (sulfuryláza) ATP
ATP + luciferin (luciferáza) oxyluciferin +
dNTP, ATP (apyráza) dNDP, dNMP, ADP, AMP, Pi
A T G C A T G C A T G C
T G AA T GG A G

454/Roche http://www.454.com/
Solexa/Illumina http://www.illumina.com/
SOLiD/ABI http://www.appliedbiosystems.com/
Helicos http://www.helicosbio.com/
Pacific Biosciences http://www.pacificbiosciences.com/
Complete Genomics http://www.completegenomicsinc.com/
sekvenování exomů - po "exon capture"
2nd generation: s amplifikací DNA
3rd generation: sekvenace jednotlivých molekul DNA
http://www.youtube.com/watch?v=77r5p8IBwJk
sekvenování nové generace
http://www.youtube.com/watch?v=kYAGFrbGl6E&NR=1

genomy nádorů
RNA seq
genová exprese, malé RNA (miRNA)
chromatin immunoprecipitation (ChIP) seq
metagenomika, mikrobiomy
fetální DNA v mateřské krvi
další využití sekvenování nové generace

ATG ACC CAG CAG CCA ATG AAA normální sekvence
Met Thr Gln Gln Pro Met Lys čtecí rámec je označen mezerami
▼
ATG CCC CAG CAG CCA ATG AAA bodová substituce typu missense
Met Pro Gln Gln Pro Met Lys (threonin je nahrazen prolinem)
▼
ATG ACC TAG CAG CCA ATG AAA bodová substituce typu nonsense
Met Thr STOP - - - - (předčasné ukončení syntézy proteinu)
▼
ATG ACA CAG CAG CCA ATG AAA tichá substituce
Met Thr Gln Gln Pro Met Lys (threonin je kódován jiným kodonem)
▼▼▼
ATG --- CAG CAG CCA ATG AAA delece bez posunu čtecího rámce
Met - Gln Gln Pro Met Lys (chybí jedna aminokyselina)
▼
ATG -CCC AGC AGC CAA TGA AA delece s posunem čtecího rámce
Met Pro Ser Ser Gln STOP - (jiné aminokyseliny + předčasná terminace)
▼▼▼ ▼▼▼ ▼▼▼
ATG ACC CAG CAG CAG CAG CAG CCA ATG AAA expanze trinukleotidové repetice
Met Thr Gln Gln Gln Gln Gln Pro Met Lys (vložen polyglutaminový úsek)
dynamické mutace
základní typy mutací

▼
ATGCCCCAGCAGCCAAT chromozóm (jedinec) A
základní typy polymorfismů DNA
▼
ATGCCCTAGCAGCCAAT chromozóm (jedinec) B
tři možné genotypy
polymorfismus typu SNP
(Single Nucleotide Polymorphism)
▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACAGAAA alela A
▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACACAGAAA alela B
polymorfismus typu STR
(Short Tandem Repeat)
▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACACACACAGAAA alela C
mnoho možných genotypů

bodové substituce: SNP, bodové mutace
malé delece a inserce
krátké tandemové repetice polymorfismy typu STR, VNTR
dynamické mutace (zejm. expanze trinukleotidů)
polymorfní inserce retroelementů
velké strukturální varianty: delece, duplikace, inverze
(SV - Structural Variants, CNV - Copy Number Variants) většinou submikroskopické
přes 10 tis. lokusů, medián několik kb
mikrodelečním syndromy
často v oblastech segmentálních duplikací
referenční sekvence genomu = ?
klinická interpretace CNV = ?
typy variability

velké fragmenty
střední fragmenty
malé fragmenty


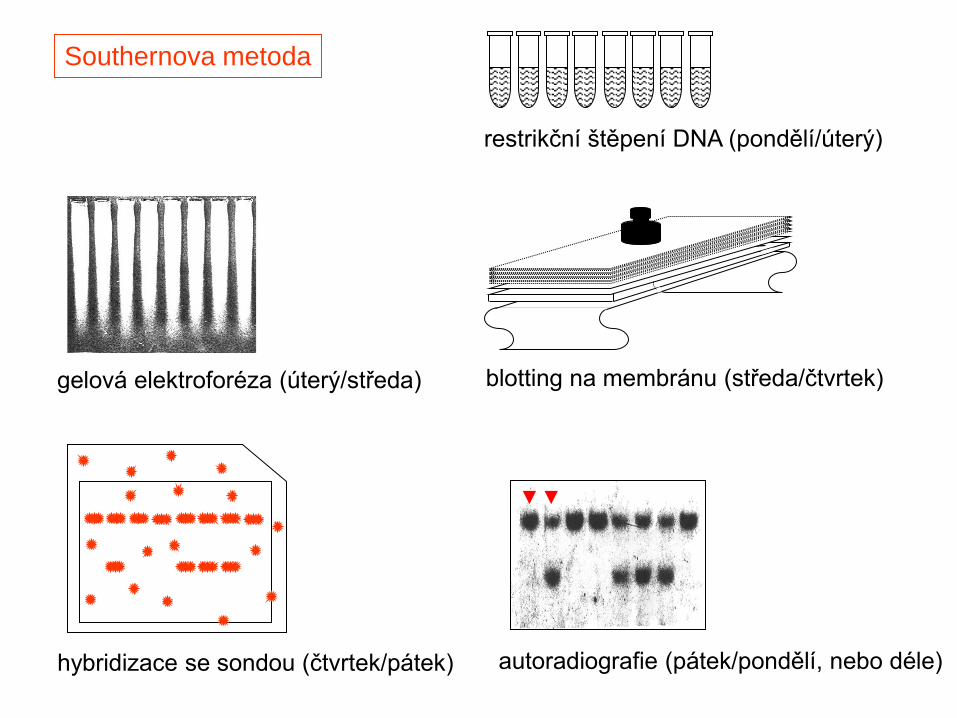
gelová elektroforéza (úterý/středa) blotting na membránu (středa/čtvrtek)
hybridizace se sondou (čtvrtek/pátek)
restrikční štěpení DNA (pondělí/úterý)
Southernova metoda
autoradiografie (pátek/pondělí, nebo déle)

Southern
1975
Northern
Western
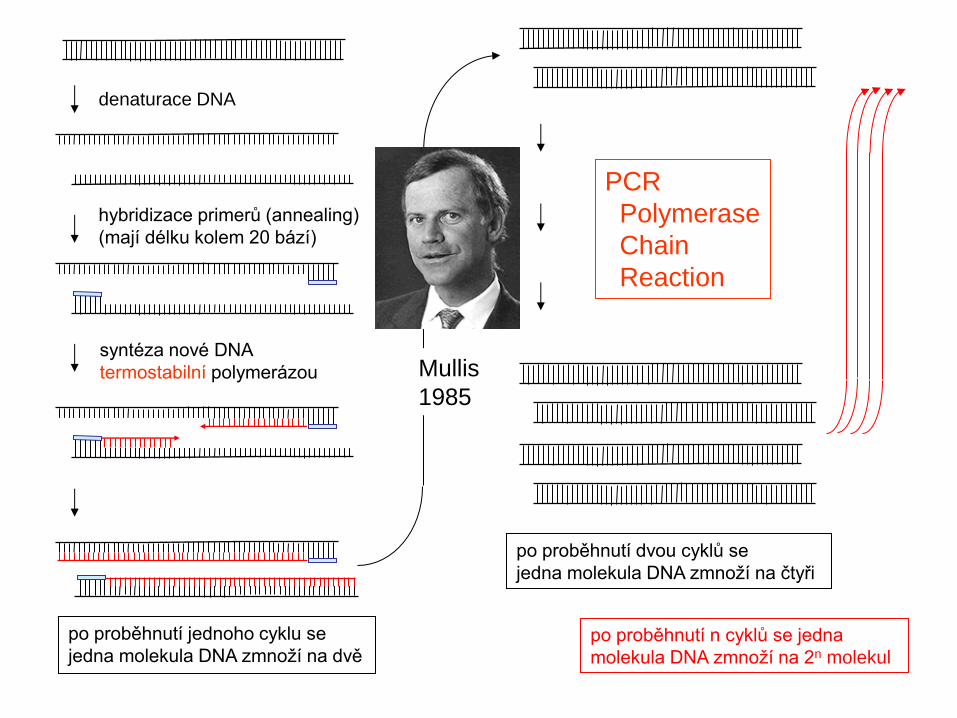
denaturace DNA
hybridizace primerů (annealing)
(mají délku kolem 20 bází)
syntéza nové DNA
termostabilní polymerázou
po proběhnutí jednoho cyklu se
jedna molekula DNA zmnoží na dvě
vstup
do dalšího
cyklu
po proběhnutí dvou cyklů se
jedna molekula DNA zmnoží na čtyři
po proběhnutí n cyklů se jedna
molekula DNA zmnoží na 2n molekul
PCR
Polymerase
Chain
Reaction
Mullis
1985


▼
ATGCCCCAGCAGCCAAT chromozóm (jedinec) A
základní typy polymorfismů DNA
▼
ATGCCCTAGCAGCCAAT chromozóm (jedinec) B
tři možné genotypy
polymorfismus typu SNP
(Single Nucleotide Polymorphism)
▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACAGAAA alela A
▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACACAGAAA alela B
polymorfismus typu STR
(Short Tandem Repeat)
▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼ ▼
ATGCCCCACACACACACACACACACAGAAA alela C
mnoho možných genotypů

1/1
1/2
1/3
2/3
3/3
3/4
800
100
200
300 238
168
308
378 4
3
2
1
M
identifikace
osob
4
3
2
1
? ? M
paternita
jednolokusové sondy

jednolokusové sondy
multilokusové sondy

multilokusové sondy
Pemberton & Amos Trends Genet 1990

polymorfismus x mutace ~ varianta definice: polymorfismus nad 1% x mutace pod 1%
98
95
ale DF508 ~ 1.5% (nosiči CF 1/25, mutace CF 1/50, z toho DF508 70%)

polymorfismus x mutace ~ varianta neutralita x nepříznivý vliv pro nositele
výhoda heterozygotů
závislost na genotypu (heterozygot, homozygot)
závislost na prostředí (černoši v Americe)
ale projev alely (polymorfismu či mutace) může záviset
na prostředí, na genotypu
a frekvence polymorfismu/mutace může odrážet selekční tlaky
v minulosti, které se dnes již nemusí projevovat

polymorfismus x mutace ~ varianta alela původní (ancestrální) x alela nově vzniklá (“mutací”)
nelze se opřít o dnešní frekvenci, ancestrální alela může být z populace odstraněna
persistence laktázy x intolerance laktózy
i kulturní prostředí může určovat, který genotyp je výhodný a který ne
persistence laktázy
Jobling, Hurles, Tyler-Smith: Human Evolutionary Genetics

sestřihové mutace
GT AG
1 2 3 9 5 6 8 7 4
GT AT
1 2 3 9 5 6 8 7 4
syndrom sousedících genů (contiguous gene syndrome)

hot-spots pro variabilitu
dinukleotid CG (CpG): meCG mutuje na TG
ATG ACG CAG CAG CCC CCT ATG AAA
Met Thr Gln Gln Pro Pro Met Lys
▼
ATG ATG CAG CAG CCC CCT ATG AAA
Met Met Gln Gln Pro Pro Met Lys
▼
ATG ACG CAG CAG CCC CTA TGA AA
Met Thr Gln Gln Pro Leu STOP -
tandemové repetice: sklouznutí při replikaci (slippage) rozptýlené repeaty: rekombinace mezi nealelními kopiemi
▼▼▼ ▼▼▼ ▼▼▼
ATG ACG CAG CAG CAG CAG CAG CCC CCT ATG AAA
Met Thr Gln Gln Gln Gln Gln Pro Pro Met Lys

1 2 3 4 5
1 2 3 4 5
vzorky 1-4:
úplná komplementarita,
sonda hybridizuje
vzorek 5:
neúplná komplementarita,
sonda nehybridizuje
alelově specifická
hybridizace
hybridizace s alelově
specifickými oligonukleotidy
(ASO)
DNA čipové technologie: paralelní hybridizace s mnoha oligonukleotidy
(nebo delšími klonovanými sondami) rozmístěnými hustě na pevném podkladu

GCGGCATGAACCGTAGGCCCATC
GCCGTACTTGGAATCCGG GCCGTACTTGGCATCCGG GCCGTACTTGGGATCCGG GCCGTACTTGGTATCCGG GCCGTACTTGG-ATCCGGG
CCGTACTTGGCATCCGGG CCGTACTTGGCCTCCGGG CCGTACTTGGCGTCCGGG CCGTACTTGGCTTCCGGG CCGTACTTGGC-TCCGGGT
5´ 3´
3´ 5´
DNA čipové technologie: resekvenace

5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´
3´CCGCGTGTCTCCTTCTCT 5´
3´CCGCGTGTCTCATTCTCT 5´
3´CCGCGTGTCTCGTTCTCT 5´
3´CCGCGTGTCTCTTTCTCT 5´
3´CCGCGTGTCTC-TTCTCTT 5´
A C
G T
del
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´

3´CCGCGTGTCTCCTTCTCT 5´
3´CCGCGTGTCTCATTCTCT 5´
3´CCGCGTGTCTCGTTCTCT 5´
3´CCGCGTGTCTCTTTCTCT 5´
3´CCGCGTGTCTC-TTCTCTT 5´
A C
G T
del

5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´
3´CCGCGTGTCTCCTTCTCT 5´
3´CCGCGTGTCTCATTCTCT 5´
3´CCGCGTGTCTCGTTCTCT 5´
3´CCGCGTGTCTCTTTCTCT 5´
3´CCGCGTGTCTC-TTCTCTT 5´
A C
G T
del
5´CGGCGCACAGAGAAAGAGAATCTCCGCAAG 3´

3´CCGCGTGTCTCCTTCTCT 5´
3´CCGCGTGTCTCATTCTCT 5´
3´CCGCGTGTCTCGTTCTCT 5´
3´CCGCGTGTCTCTTTCTCT 5´
3´CCGCGTGTCTC-TTCTCTT 5´
A C
G T
del
5´GCGCACAGAGAAAGAGAATC 3´
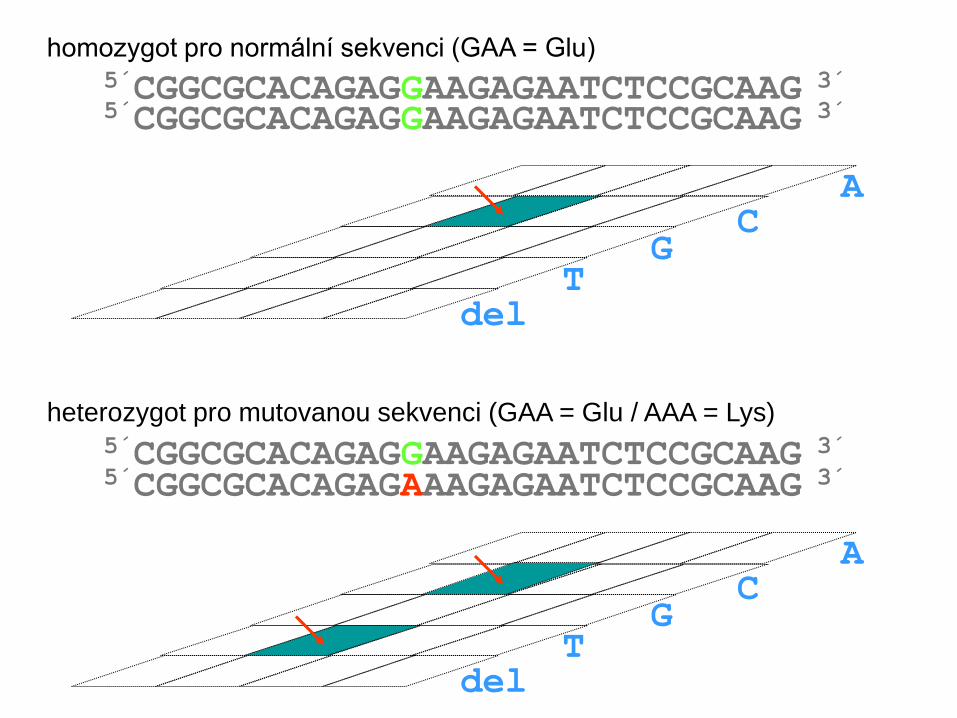
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´ homozygot pro normální sekvenci (GAA = Glu)
A C
G T
del
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´
5´CGGCGCACAGAGAAAGAGAATCTCCGCAAG 3´
heterozygot pro mutovanou sekvenci (GAA = Glu / AAA = Lys)
A C
G T
del
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´

www.
affymetrix.com

A C G T
del
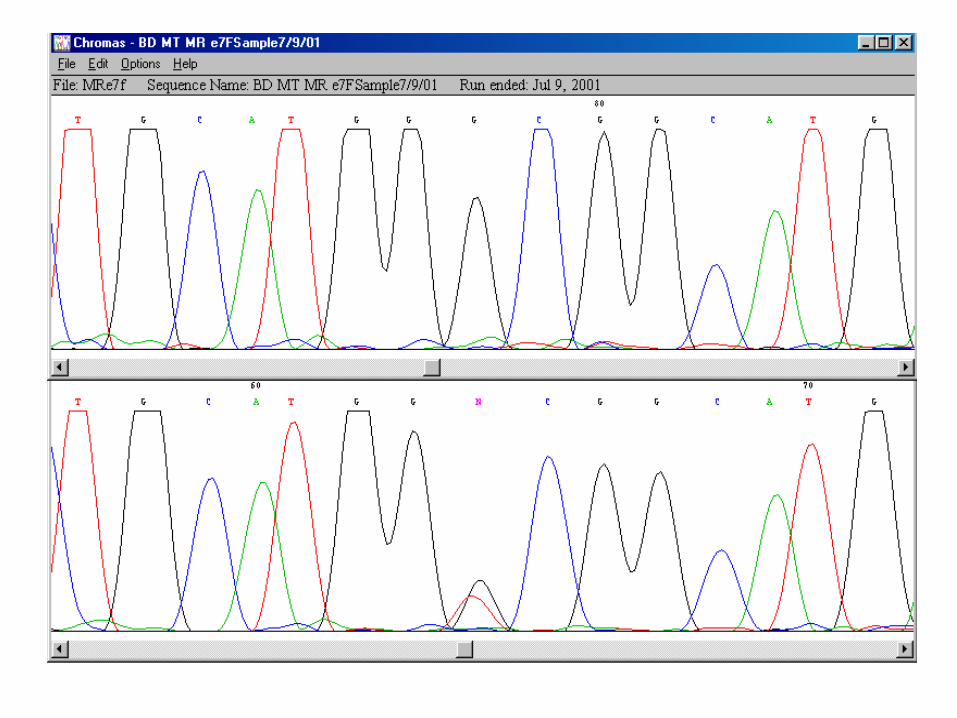
G C A C A G A G G A A A G A G A
heterozygot pro mutovanou sekvenci TP53 (kodon 286: GAA = Glu / AAA = Lys)

3´ TTCTCTTAGAGGCGTTCTT 5´
neznačená
testovaná
DNA
T
extenze oligonukleotidu
zakotveného v buňce čipu
o jeden značený dideoxynukleotid
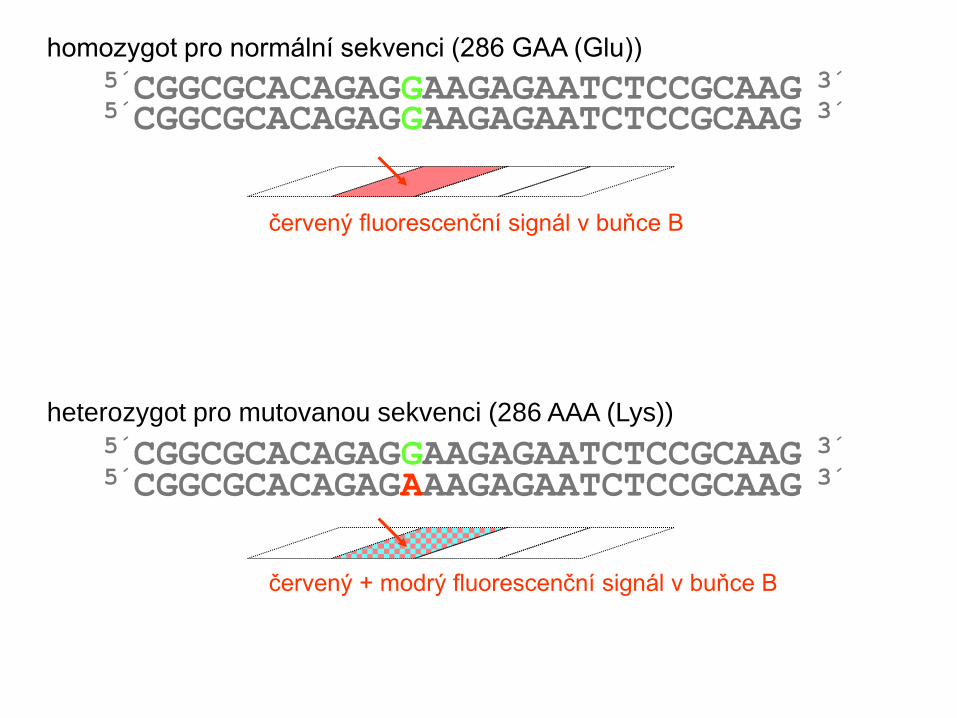
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´
červený fluorescenční signál v buňce B
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´ homozygot pro normální sekvenci (286 GAA (Glu))
heterozygot pro mutovanou sekvenci (286 AAA (Lys))
5´CGGCGCACAGAGGAAGAGAATCTCCGCAAG 3´
červený + modrý fluorescenční signál v buňce B
5´CGGCGCACAGAGAAAGAGAATCTCCGCAAG 3´
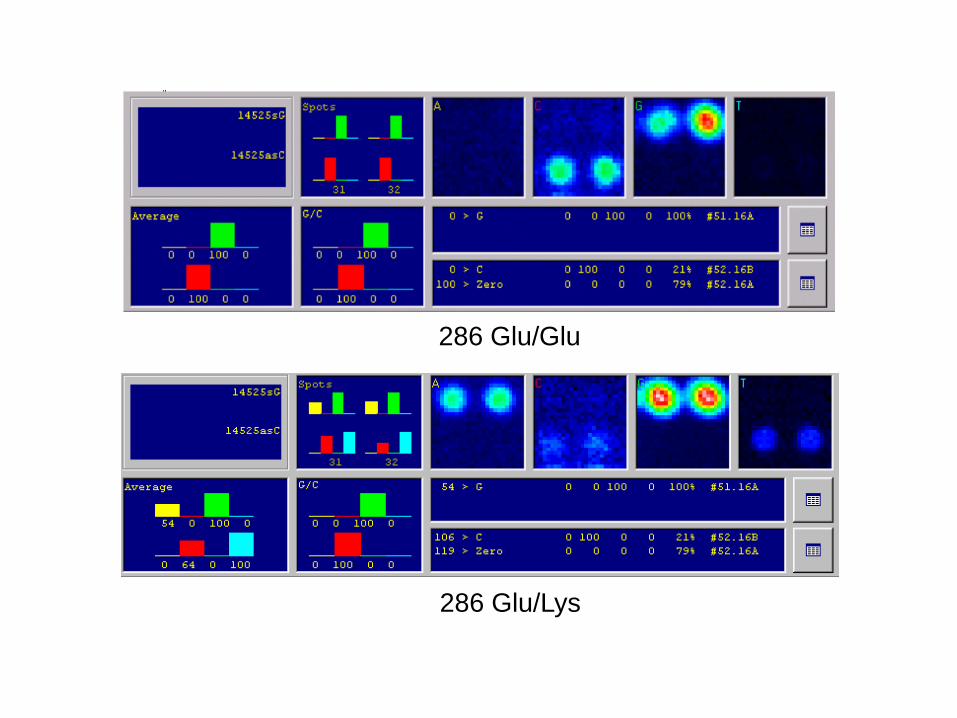
286 Glu/Lys
286 Glu/Glu

I.
II.
III.
1 2
1
1
2
2
pancreatic
cancer
(47y)
thyroid
cancer
(18y)
*
breast
cancer
(50y)
rhabdomyosarcoma
(3y)
non-Hodgkin lymphoma B
(7y)
*
*
G A G A A A G A G
Glu Glu Glu Lys
wild
type
III.1
blood
285 286 287
G A G G A G
G
G A A
Glu Glu Glu 285 286 287
GERMLINE p53 MUTATION
IN FAMILY 7
Glu286Lys (exon 8)
FAMILY 7
* proven carrier
* proven non-carrier
childhood cancer
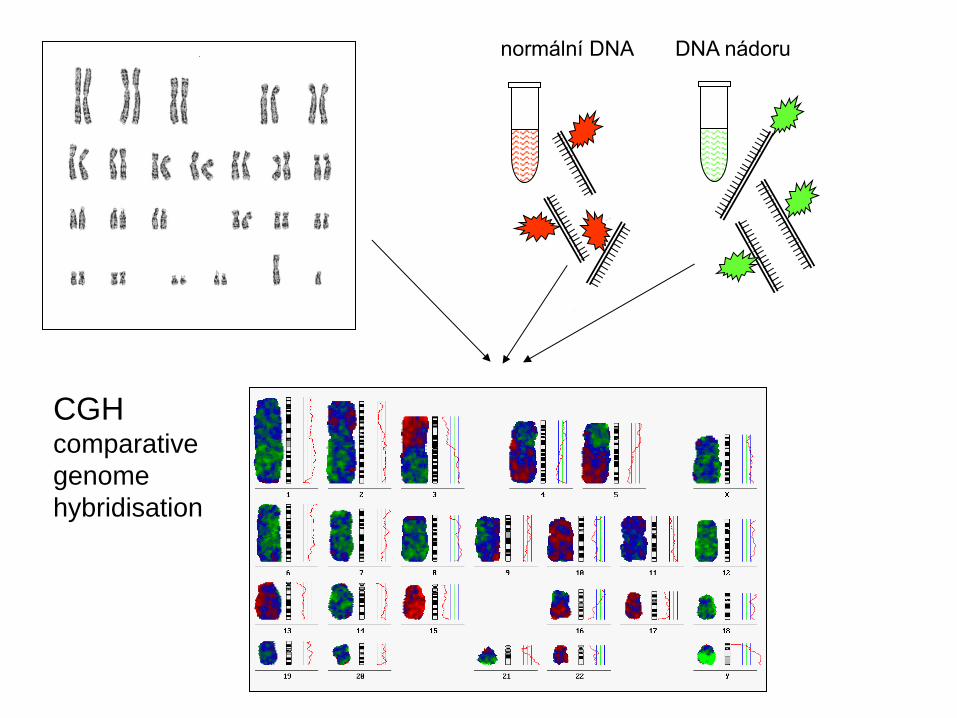
normální DNA DNA nádoru
CGH comparative
genome
hybridisation
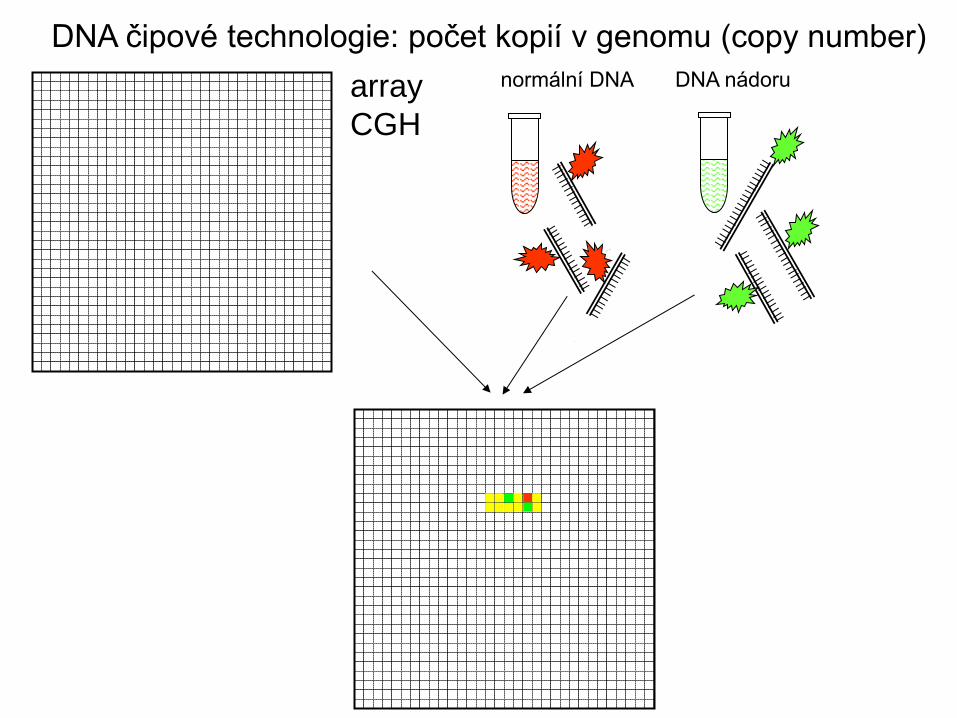
normální DNA DNA nádoru array
CGH
DNA čipové technologie: počet kopií v genomu (copy number)

G C A C A G C G A A T G G G A C G G T G A A C C A C G
17p 17q

fluorescenční in situ hybridizace (FISH)







