

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
MECHANISMS OF ENZYME ACTION
Introduction to enzymes
1. Substratspezifität
2. Coenzyme
3. Regulation enzymatischer
Aktivität
4. Enzym Nomenklatur
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Was sind Enzyme ? biologische Katalysatoren
Unterschied zu chemischen Katalysatoren?
1. Höhere Reaktionsrate.
2. Mildere Reaktionsbedingungen
unter 100°C, neutraler pH, Normaldruck.
3. Grössere Spezifität
selten Nebenprodukte, z.B ribosomale polypeptidsynthese -> keine
Fehler -> über 1000 AS. Dagegen chemische Synthese von Polypeptiden ->
Nebenprodukte -> nicht mehr als 100 AS
4. Regulationsmöglichkeit
allosterische Kontrolle, kovalente Modifikationen
-> Wie funktionieren Enzyme
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
An enzyme–substrate complex illustrating both the geometric and the physical
complementarity between enzymes and substrates.
Page 4
60
1. Substratspezifität
Kräfte welche Substrat und Enzym zusammen-
halten -> ähnlich denen die Proteinkonformation
beeinflussen
van der Waals, elektrostatische, H-Brücken,
hydrophobe Interaktionen.
Enzym -> vorgeformt für Substrat (Schlüssel-
Schloss)
aber auch verformbar-> induced fit.
komplementäre Ladungsverteilung
spezifische AS können interagieren und Bindung
stabilisieren.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Prochirale Differenzierung
Page 4
61
A. Stereospezifität
Enzyme sind hochspezifisch in Bindung
chiraler Substrate als auch in der Katalyse
der Reaktion.
Stereospzifität da Enzyme inherent chiral
sind.
Proteine -> nur aus L-Aminosäuren ->
assymmetrische Aktive Stellen.
z.B. Trypsin spaltet nur Polypeptide aus
L-Aminosäuren aber nicht aus D-AS.
Glucosemetabolsimus Enzyme spezifisch
für D-Glucose-Reste.
Beispiel Hefe Alkohol Dehydrogenase->
in Nachmittagvorlesung für Biochemiker.
CH3CH2OH + NAD+ <-> CH3CH + NADH + H+
ethanol acetaldehyd
O
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
The structures and reaction of nicotinamide-adenine dinucleotide (NAD+) and
nicotinamide adenine dinucleotide phosphate (NADP+).
Page 4
61
U. Albrecht BC1
Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
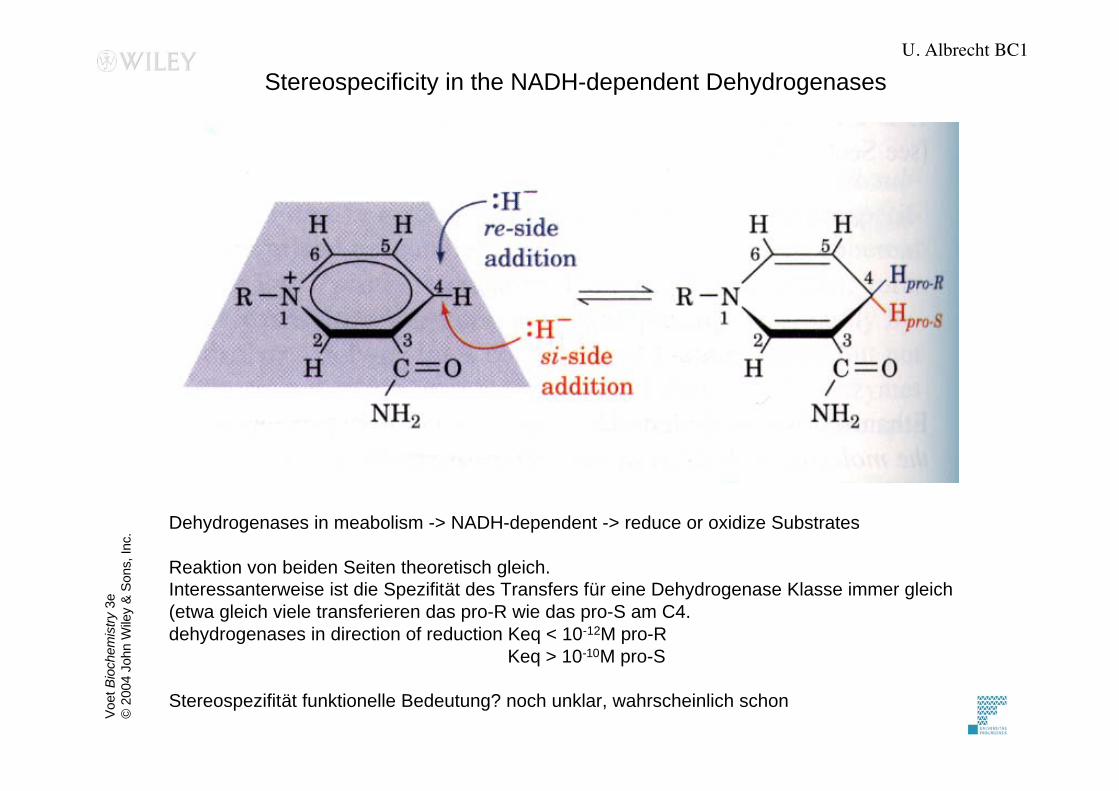
Stereospecificity in the NADH-dependent Dehydrogenases
Dehydrogenases in meabolism -> NADH-dependent -> reduce or oxidize Substrates
Reaktion von beiden Seiten theoretisch gleich.
Interessanterweise ist die Spezifität des Transfers für eine Dehydrogenase Klasse immer gleich
(etwa gleich viele transferieren das pro-R wie das pro-S am C4.
dehydrogenases in direction of reduction Keq < 10-12M pro-R
Keq > 10-10M pro-S
Stereospezifität funktionelle Bedeutung? noch unklar, wahrscheinlich schon
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
B. Geometrische Spezifität
Zusätzlich zur Stereospezifität sind viele Enzyme selektiv bezüglich der Identität der chemischen
Gruppen ihres Substrates.
Enzyme variieren im Ausmass dieser geometrischen Spezifität. Einige spezifisch für nur ein Substrat,
andere reagieren auch mit ähnlichen Substraten.
z.B. Hefe Alkoholdehydrogenase setzt am besten Ethanol um, kann aber auch entsprechende Aldehyde
und Ketone als auch Methanol und Isopropanol umsetzen.
Verdauungsenzyme sind sehr permissiv. Spezifitäten verschiedener Exopeptidasen
Allerdings variiert die Reaktionsrate mit der Identität der AS in der Umgebung des C-terms
Einige Enzyme sind sehr unspezifisch wie z.B. Chymotrypsin
Permissive Enzyme sind allerdings eher die
Ausnahme als die Regel.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Spezifität verschiedener Endopeptidasen
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
3. Coenzyme
Enzyme können alleine schlecht oxidations-reduktionsreaktionen katalysieren.
Cofaktoren -> ‘chemische Zähne’ der Enzyme
Cofaktoren:
1) Metal ionen, z.B. Zn2+ in carboanhydrase
CO2 + H2O -> HCO3- + H+
2) organische Moleküle, Coenzyme, wie z.B. NAD+,NADP+ , FAD+ -> transiente Assozation
Ferredoxin - NADP - reduktase
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
3) prosthetische Gruppen, z.B. Häm in Hämoglobin -> permanent Assoziiert
Coenzyme werden chemisch verändert -> muss wieder in Ursprungszustand
Für prosthetische Gruppen in einer separaten Phase der enzymat. Reaktion.
Für transient gebudnene wie NAD+ -> Regeneration durch anderes Enzym möglich.
Apoenzyme (inaktiv) + Cofaktor <-> Holoenzym (aktiv)
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
64
The Common Coenzymes.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Viele Vitamine sind Coenzym Vorläufer
Viele Organismen können gewisse Vorläufer für essentielle Cofaktoren nicht synthetisieren ->
sie müssen über die Nahrung aufgenomen werden.
= VITAMINE
Viele Coenzyme wurden über Heilung von Nähstoffmangelkrankheiten gefunden.
z.B.
Diese Moleküle heilen die Nährstoffmangelkrankheit Pellagra (Durchfall, Dermatitis, Demenz)
Nicotinamid kann aus Tryptophan synthetisiert werden. Mais basierte Ernährung hat wenig
Tryptophan und das Nicotinamid im Mais muss zuerst in verwertbare From umgewandelt werden
über eine leichte basische Behandlung (Mex. Indianer durchtränken das Maismehl mit lime Wasser,
das leicht basisch ist bevor sie Tortillas machen).
Alle Vitamine in menschlicher Ernährung die Coenzym Vorläufer sind, sind Wasserlöslich -> Tabelle
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
64
Vitamine die Coenzym Vorläufer sind
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Interessanterweise sind fettlösliche Vitamine wie Vit A und Vit D nicht
Komponenten von Coenzymen obwohl sie in Spuren auch wichtig sind
in der Nahrung höherer Organismen (Vit A -> Auge, Vit D -> Knochenmineralisierung)
Unsere Vorfahren konnten möglicherweise verschiedene Vitamine synthetisieren.
Da viele Vitamine im Ueberfluss in Nahrung vorhanden sind, oder durch die
Bakterien im Verdauungstrakt synthetisiert werden, konnte durch weglassen der
Vitaminsynthese Energie gespart werden und deshalb ist Vitaminsynthese in
unserer Evolution verlorengegangen.
The image shows a mixture of two pure cultures
of Lactobacillus intestinalis (the green elongated
elements) and Bifidobacterium longum (pinkish'grains') as seen through a microscope. In some
cases, an increased presence of these
microorganisms in the intestinal flora can have
probiotic effects which help strengthen immunity
against many pathologies of bacterial origin.
-> Probiotische Nahrungsmittel -> z.B. modifizierte Bakterien die vermehrt Vitamine oder andere
Substanzen produzieren und sich im Darm aufhalten. (LC1 Yoghurt) = funktionelle Lebensmittel
isotonische Getränke (Isostar).
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
4. Regulation enzymatischer Aktivität
Um metabolische Prozesse zu koordinieren muss enzymatische Aktivität regulierbar sein.
Dies ermöglicht es auf Umweltveränderungen zu antworten und zu wachsen, damit alles in
geordenter Weise abläuft.
2 Möglichkeiten wie man enzymatische Aktivität kontrollieren kann:
1. Kontrolle der Verfügbarkeit des Enzyms
Die Menge eines Enzyms in der Zelle Hängt von der Rate der Synthese und der Rate
der Degradation ab.
z.B. E.coli mit Medium ohne Lactose -> exprimieren das Abbauenzym für Lactose nicht
Wenn Lactose gegeben wird -> innerhalb von Minuten wird Enzym gemacht.
In verschiedene Geweben von Säugern unterschiedliche Enzyme exprimiert, obwohl
genetische Information in allen Zellen gleich (siehe später, expression und transmission
genetischer Information).
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
2. Kontrolle der Enzymaktivität
Die Enzymaktivität kann durch konformationelle und strukturelle Veränderungen be-
einflusst werden.
Aktivität direkt proportional zur Konzentration des Enzym-Substrat Komplexes. Diese
ist abhängig von konzentrationen von Enzym, Substrat und der Substratbindunsaffinität.
z.B. Hämoglobin -> allosterische Regulation -> Bindung kleiner Moleküle (=homotrope und
heterotrope Effektoren) O2, CO2, H+, BPG.
Beispiel: Allosterische Kontrolle der Aspartate Transcarboamylase (ACTase)
Erster Schritt der Pyrimidinbiosynthese
-> Komponenten der Nukleinsäuren.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
The rate of the reaction catalyzed by ATCase as a function of aspartate concentration.
Page 4
65
Rückkoplungsinhibition von ATCase reguliert die Pyrimidin Biosynthese
positiv homotrope cooperative Bindung von Aspartat und Carbamoyl phosphat.
heterotrope Inhibition durch cytidine triphosphate (CTP=pyrimidin)
heterotrope Aktivierung durch adenosin triphosphate (ATP=purin).
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Schematic representation of the pyrimidine biosynthesis pathway.
Page 4
66
Rückkopplungsinhibition
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Allosterische Aenderungen beeinflussen die Substratbindungsstelle der ATCase
T inaktiv (CTP) R aktiv (ATP)
Ansicht von oben
Ansicht von seite
c3
c3
r2
r2
r2
r2r2
c3
300kD
r = regultorische
Untereinhiet
c = catalytische
Untereinheit
c alleine höhere
katalytische Rate,
nicht beeinflusst
durch CTP oder ATP
r kann diese alleine
binden aber kann
c nur im Komplex
beeinflussen.
240s loop
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
67
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
67
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
67
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
67
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
68
Strukturelle Basis der allosterischen Regulation von ATCase
katalytisches monomer
carbamoylphosphat
aspartat
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
69
Glu 239
Wenn Glu 239 -> Gln 239 -> keine homo- und heterotropen Efekte mehr->
bleibt im mittleren Zustand (b)
Schematisches Diagramm welches die Konformationsänderungen in 2 vertikal miteinander
interagierenden katalytischen ATCase Untereinheiten illustriert.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Page 4
70
5. Enzymnomenklatur
Enzym Substratname + Endung -ase oder
Enzym katalyt. Aktion + Endung -ase
Für lange Zeit keine systemat. Regeln -> manchaml 2 Namen für das selbe Enzym
manchmal sagt Name nichts über die Aktion, z.B Catalase
(dismutation von H2O2 zu H2O und O2)
Enzyme werden klassifiziert und benannt gemäss der Reaktion die sie katalysieren
6 Hauptklassen
Subklassen
Sub- subklassen
-> jedes Enzym bekommt 2 Namen und eine Nummer aus 4 Zahlen.
Empfohlener Name -> Name für täglichen Gebrauch -> alter Name, Trivialname
Systematischer Name -> keine Ambiguität -> Name seiner Substrat(e) mit Endung -ase welcher
eine der 6 Hauptklassen spezifiziert.
z. B. carboxypeptidase A -> peptidyl-L-aminosäure hydrolase EC 3.4.17.1
Enzym Comission Hauptklasse 3 Subklasse 4 Sub-subklasse 17 Seriennummer (arbiträr)
hydrolase an peptidbindg metallocarboxypeptidasen
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
anderes Beispiel:
alcohol dehydrogenase -> alkohol:NAD+ oxidoreduktase EC 1.1.1.1.
Enzym Nomenklatur Datenbase: http://expasy.org/enzyme
http://www.chem.qmw.ac.uk/iubmb/enzyme/
http://www.brenda.uni-koeln.de
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
EC 2 Transferases
EC 2.1 Transferring one-carbon groupsEC 2.2 Transferring aldehyde or ketonic groupsEC 2.3 AcyltransferasesEC 2.4 GlycosyltransferasesEC 2.5 Transferring alkyl or aryl groups, other than methyl grouEC 2.6 Transferring nitrogenous groupsEC 2.7 Transferring phosphorus-containing groupsEC 2.8 Transferring sulfur-containing groupsEC 2.9 Transferring selenium-containing groups
EC 2.3 Acyltransferases
EC 2.3.1 Transferring groups other than amino-acyl groups
EC 2.3.2 Aminoacyltransferases
EC 2.3.3 Acyl groups converted into alkyl on transfer
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
EC 2.3.2 Aminoacyltransferases
EC 2.3.2.1 D-glutamyltransferaseEC 2.3.2.2 -glutamyltransferaseEC 2.3.2.3 lysyltransferaseEC 2.3.2.4 -glutamylcyclotransferaseEC 2.3.2.5 glutaminyl-peptide cyclotransferaseEC 2.3.2.6 leucyltransferaseEC 2.3.2.7 aspartyltransferaseEC 2.3.2.8 arginyltransferaseEC 2.3.2.9 agaritine -glutamyltransferaseEC 2.3.2.10 UDP-N-acetylmuramoylpentapeptide-lysine N6-
alanyltransferaseEC 2.3.2.11 alanylphosphatidylglycerol synthaseEC 2.3.2.12 peptidyltransferaseEC 2.3.2.13 protein-glutamine -glutamyltransferaseEC 2.3.2.14 D-alanine -glutamyltransferaseEC 2.3.2.15 glutathione -glutamylcysteinyltransferaseroups
converted into alkyl on transfer
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
EC 2.3.2.7Common name: aspartyltransferase
Reaction: L-asparagine + hydroxylamine = NH3 + L-aspartylhydroxamate
Other name(s): -aspartyl transferase; aspartotransferase
Systematic name: L-asparagine:hydroxylamine -aspartyltransferase
Links to other databases: BRENDA, EXPASY, KEGG,ERGO, CAS registry number: 37257-23-1
References:
1. Jayaram, H.N., Ramakrishnan, T. and Vaidyanathan, C.S.Aspartotransferase from Mycobacterium tuberculosisH37Ra. Indian J. Biochem. 6 (1969) 106-110.
U. Albrecht BC1

Voet B
iochem
istr
y 3
e
© 2
004 J
ohn W
iley &
Sons, In
c.
Enzym unter-
scheidet pro-S
und pro-R
und si , re Seiten
vom Nicotinamid
Ring
pro-S pro-R
pro-S
U. Albrecht BC1







