

1. Introduction
2. Animal models of behavioral
despair
3. Polyphenols in animal models
of behavioral despair
4. Expert opinion
Review
Natural polyphenols in themanagement of major depressionLokesh Pathak, Yadvendra Agrawal & Ashish Dhir††Gujarat Forensic Sciences University, Institute of Research & Development, Gujarat, India
Introduction: Natural polyphenols, the non-essential micronutrients, found in
array of plant products, are known to affect various physiological and bio-
chemical functions in the body. Studies have shown the protective effect of
these polyphenols in different neurological and mental disorders. These poly-
phenols modulate monoaminergic neurotransmission in the brain and thus
possess antidepressant-like activity at least in animal models of depression.
Areas covered: The present review discusses the use of these natural polyphe-
nols in the treatment of major depression. The review article discusses the
antidepressant potential of some important polyphenols such as amentofla-
vone, apigenin, chlorogenic acid, curcumin, ferulic acid, hesperidin, rutin,
quercetin, naringenin, resveratrol, ellagic acid, nobiletin and proanthocyani-
dins. The mechanism of action of these polyphenols in the treatment of major
depression is also discussed in detail.
Expert opinion: There is an exciting prospect in the discovery of natural
polyphenols as therapeutic agents in the treatment of major depression.
Keywords: antidepressants, antioxidant, monoaminergic neurotransmission, polyphenols
Expert Opin. Investig. Drugs [Early Online]
1. Introduction
‘Polyphenols’ is a broad category of natural agents comprising tannins, flavonols,flavanols, flavones, anthocyanins, proanthocyanidins, phenolic acids, hydroxyben-zoic acids, hydroxycinnamic acids and their structural variants [1,2]. These are natu-rally occurring substances that are present in fruits and beverages (fruit juice, wine,tea, coffee, chocolate and beer) and, to a lesser extent in vegetables, dry legumes andcereals [3]. Polyphenols have shown several beneficial effects illustrating their prom-ising role as therapeutic agents in various acute and chronic conditions of the bodysuch as cardiovascular and metabolic disorders [4,5], inflammation and cancer [6],oxidative stress [7,8], cerebral ischemia [9] and obesity [10,11]. Numerous studieshave shown that natural polyphenols play an important role in proper functioningof the brain [12-14]. These micronutrients, for example, are involved in learningand memory process, prevent neuroinflammation and possess antioxidantpotential [15-17]. These polyphenols are known to modulate various neurotransmittersystems in the brain. For example, curcumin found in Curcuma longa modulatesserotoninergic and dopaminergic neurotransmission in the brain and thereforecould be useful as an antidepressant agent [16]. The fact that these polyphenols alterdifferent neurotransmitters has prompted researchers to explore their usefulness inthe treatment of major depression.
Major depression is a serious medical illness that affects both developed anddeveloping countries [18,19], and has an estimated lifetime prevalence of about15 -- 20% [20-22]. It has been estimated that by the year 2020, depression will reachin second place in the ranking of disability-adjusted life-years [23]. The monoaminetheory of major depression has been well-accepted by researchers. According to thistheory, there is a decrease in the levels of various monoaminergic neurotransmitterssuch as norepinephrine, serotonin and dopamine in brains of depressed patients.
10.1517/13543784.2013.794783 © 2013 Informa UK, Ltd. ISSN 1354-3784, e-ISSN 1744-7658 1All rights reserved: reproduction in whole or in part not permitted
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

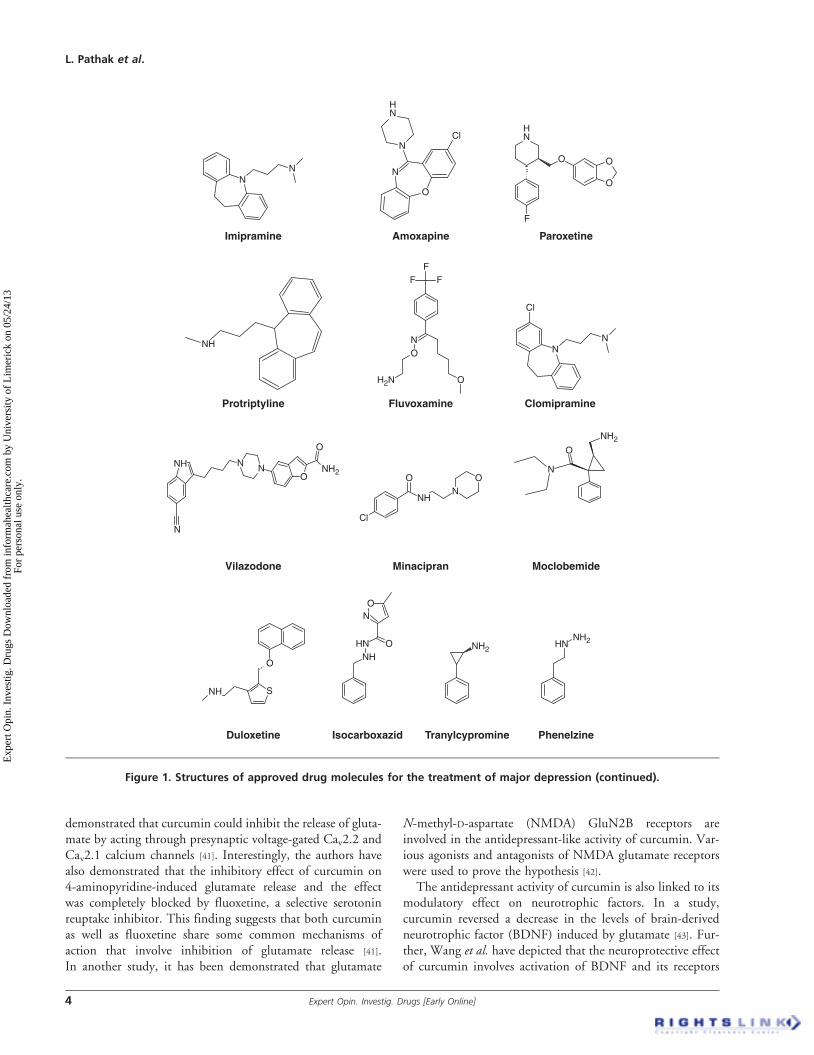
Molecules that reverse this deficiency are known to possessantidepressant activity [24]. Various new antidepressants suchas venlafaxine and triple reuptake inhibitors have beendiscovered based on this theory [25].More than 25 antidepressant agents are available commer-
cially (Figure 1) that has allowed treatment of depressedpatients in an efficient manner. However, acute or chronicadministration of these antidepressants could lead to seriousside effects, adverse drug reactions and drug--drug and food--drug interactions [26]. Unfortunately, about 10 -- 30% ofdepressed patient population still continues to have depressiveepisodes besides the availability of these wonderful drugs [27].All these factors justify a need for newer antidepressant agentspossessing better efficacy and minimal side effects [24,28].With this background, the present review attempts to dis-
cuss some of the polyphenols that are being tested extensivelyin animal models of major depression. The review alsoincludes the mechanism of action of these polyphenols fortheir antidepressant activity. Some of the polyphenols thathave been tested for their antidepressant activity are illustratedin Table 1 with their chemical structures representedin Figure 2. Based on the preclinical data, it is hypothesizedthat these polyphenols might be useful as an adjuvant therapywith already established antidepressant agents for the treatmentof major depression.
2. Animal models of behavioral despair
The discovery of various new molecular entities (NMEs) asantidepressants requires appropriate animal models withexcellent face, construct and predictive validity [29]. These ani-mal models should mimic human condition of major depres-sion. Various animal models have been proposed so far toevaluate the antidepressant-like activity of new molecules.Out of these models, forced swim test (FST) and tail-suspension test (TST) are majorly used. Although these mod-els do not mimic the human condition of major depression,however, antidepressants acting through modulating mono-aminergic neurotransmission are active in these test mod-els [30]. In a FST, animals are forced to swim inside a jar(with specific dimensions) filled with water (maintained at25�C) up to a certain height. During an initial 1 -- 2 min,animal struggles to get out of the water and remain mobile.After this struggling phase has passed, animal gives up to the
situation, and remain floating (immobile) inside the jar withvery little limb movements. This condition correlates withthe state of behavioral despair and various antidepressantsare known to reverse this immobility period [24]. Similarly,in a TST, an animal (mice or rat) is suspended by its tail ata certain height and for a definite period of time and itsimmobility period is assessed. Although both of these modelsmeasure immobility period in animals, however, they differeach other on neurochemical basis [31]. NMEs that reversethis immobility period in these two tests are studied furtherfor their antidepressant action. Some of the other modelsthat are routinely used for testing antidepressant-like activityof new molecules include olfactory bulbectomy, sucrose pref-erence, intracranial self-stimulation task and others. In thesebehavioral paradigms of despair, locomotor activity is alsoassessed simultaneously with the antidepressant activity. Ithas been seen that amphetamine-like psychostimulants arealso active in these animal models [32]. Therefore, researchersoften times use locomotor activity as a criterion to distinguishantidepressant and psychostimulatory activities in thesebehavioral paradigms of despair.
It has been found that natural polyphenols are active inthese animal models of despair. Neurochemical estimationhas shown that some of these polyphenols modulate variousneurotransmitters in the brain that are involved in the patho-physiology of various neuropsychiatric and neurodegenerativedisorders [33]. The details describing the antidepressant activ-ity of some of the natural polyphenols in these behavioralmodels of despair are explained in separate sub-headings.
3. Polyphenols in animal models ofbehavioral despair
The present section discusses the antidepressant-like activityof various natural polyphenols. These polyphenols could beused as an adjuvant therapy in the treatment of major depres-sion. However, it is strongly recommended that clinical trialsdescribing the efficacy and safety profile of these moleculesshould be carried out before these drugs are considered to beuseful in the treatment of major depression.
3.1 CurcuminCurcumin, a polyphenol, is the major active ingredient ofCurcuma longa [24]. Curcumin is known for its diverse biolog-ical actions and widely used in Indian system of medicine [34].A recent review on curcumin has elaborately described theeffect of curcumin in various neurodegenerative disorders [35].Curcumin is known to modulate the activity of various neuro-transmitter systems in the body via regulating the levels ofmonoamine oxidase-A (MAO-A) and monoamine oxidase-B (MAO-B) enzymes [16]. Xu et al. in 2007 showed thatcurcumin enhanced hippocampal neurogenesis in chronicallystressed mice and the effect was similar to imipramine, a stan-dard tricyclic antidepressant agent [36]. Curcumin showedantidepressant-like action in FST in both mice and rats.
Article highlights.
. Natural polyphenols in major depression
. Preclinical studies of natural polyphenols in animalmodels of depression
. Mechanism of action of polyphenols as anantidepressants
. Side effects of natural polyphenols
This box summarizes key points contained in the article.
L. Pathak et al.
2 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

Xu et al. in 2005 showed that curcumin at doses of 5 and10 mg/kg., p.o. was active in mouse FST and TST [37].Further, it has been shown that there is an increased level ofnoradrenaline and serotonin in frontal cortex and hippocam-pal brain regions following curcumin administration at a doseof 10 mg/kg., p.o. compared with control group [37]. Inanother similar kind of study, it has been shown that curcu-min is antidepressant in olfactory bulbectomy animal modelof major depression [38]. Curcumin totally reversed olfactorybulbectomy-induced reduced levels of serotonin, noradrena-line and increased levels of 5-hydroxyindoleacetic acid and4-dihydroxyphenylacetic acid in the hippocampus region ofmouse brain in this animal model [38].
The antidepressant activity of curcumin according toWang et al. is linked to modulation of serotoninergic system [39].
It has been shown that there is an involvement of 5-HT1A/1B
and 5-HT2C serotonin receptors in the antidepressant-likeactivity of curcumin [39]. Serotoninergic receptors producephysiological response via adenyl cyclase/cyclic adenosinemonophosphate (AC/cAMP) pathway. Li et al. have shownthat curcumin modulates AC/cAMP pathway and cAMPresponse element binding protein and the effect is mediatedthrough suppressing central 5-HT1A/1B/7 receptors [40].
Besides serotoninergic receptors, other receptor systems arealso involved in the antidepressant-like effect of curcumin.Glutamate, an excitatory neurotransmitter, is known to playan important role in the antidepressant-like action ofcurcumin [41]. Lin et al. have shown that curcumin couldinhibit the release of glutamate induced by 4-aminopyridine(a potassium channel blocker) [41]. Further, it has been
NH
ON
O
O
O NH
H
Cl
Cl
NH
H
Nortriptyline Venlafaxine Doxepin Raboxetine Sertraline
O
N
N
F
Escitalopram
O
N
N
F
Citalopram
N
Amitriptyline
Desipramine Fluoxetine Mirtazapine Trimipramine Mianserin
Cl
ONH
NO
NH NN
N
NN
N N
Cl
O
Bupropion Minaprine Trazodone
Maprotiline
NH
N
NH
NH O
F
F
F
NN
N
NN
N
N
N
O
OH
Figure 1. Structures of approved drug molecules for the treatment of major depression.
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 3
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.
demonstrated that curcumin could inhibit the release of gluta-mate by acting through presynaptic voltage-gated Cav2.2 andCav2.1 calcium channels [41]. Interestingly, the authors havealso demonstrated that the inhibitory effect of curcumin on4-aminopyridine-induced glutamate release and the effectwas completely blocked by fluoxetine, a selective serotoninreuptake inhibitor. This finding suggests that both curcuminas well as fluoxetine share some common mechanisms ofaction that involve inhibition of glutamate release [41].In another study, it has been demonstrated that glutamate
N-methyl-D-aspartate (NMDA) GluN2B receptors areinvolved in the antidepressant-like activity of curcumin. Var-ious agonists and antagonists of NMDA glutamate receptorswere used to prove the hypothesis [42].
The antidepressant activity of curcumin is also linked to itsmodulatory effect on neurotrophic factors. In a study,curcumin reversed a decrease in the levels of brain-derivedneurotrophic factor (BDNF) induced by glutamate [43]. Fur-ther, Wang et al. have depicted that the neuroprotective effectof curcumin involves activation of BDNF and its receptors
NN N
O
ClN
HN
HN
F
O O
O
Imipramine
NH
FFF
O
NO
H2N
NN
Cl
Clomipramine
N
NH N NO
O
NH2
NH2
N
O
Cl
O
NHN
O
Moclobemide
O
SNH
HNNH2
Amoxapine Paroxetine
Protriptyline Fluvoxamine
Vilazodone
Duloxetine
NHHN O
NO
NH2
Isocarboxazid Tranylcypromine Phenelzine
Minacipran
Figure 1. Structures of approved drug molecules for the treatment of major depression (continued).
L. Pathak et al.
4 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

TrkB-dependent MAP kinase (mitogen-activated proteinkinase) and PI3K (phosphatidylinositol 3-kinase) cascadewhen checked in rodent cortical neurons [44].
In one of the studies, curcumin has shown to increase thelevels of serotonin and dopamine neurotransmitters andinhibits the activity of MAO isoenzymes. Further, curcumindecreases immobility period in mouse FST [16]. It has beendemonstrated that curcumin could enhance the antidepressant-like activity of various standard antidepressants such as fluoxetine,venlafaxine or bupropion but not imipramine [16].
Huang et al. developed a novel model of depression. Theauthors in the present study administered corticosterone exog-enously chronically for 3 months. As a result, rats developeddepression-like state. In these rats, a decrease in the levels ofBDNF was found. Pretreatment of curcumin reversed thisdepression-like pattern in this animal model [45]. In a veryrecent study, the antidepressant-like activity of curcumin wasstudied in WKY rat model of depression. Curcumin waseffective both in acute and chronic dosing profiles and theresults were correlated with an increase in the levels of BDNFfollowing curcumin administration in this animal model [46].
3.2 ApigeninApigenin is one of the bioflavonoids abundantly found in cit-rus fruits [47]. The molecule has shown promising activity in
central nervous system (CNS) disorders such as anxiety andinsomnia [48,49]. Apigenin is also protective in cerebral ische-mia reperfusion injury in rats [50] suggesting that the moleculecould easily cross blood--brain barrier. Apigenin has shownanti-inflammatory activity in cultured microglia cells andtherefore the molecule is hypothesized to display neuroprotec-tive/disease-modifying properties in various neurodegenera-tive disorders including Alzheimer’s disease [51]. A recentstudy has shown the protective effect of apigenin in amy-loid-b25-35-induced toxicity in mice [52] suggesting that apige-nin could be useful in the treatment of Alzheimer’s disease.Apigenin has also shown antioxidant effect in kainate modelof excitotoxicity in mice [53]. In this study, apigenin preventedkainic acid-induced excitotoxicity via quenching reactiveoxygen species as well as inhibiting glutathione depletion inhippocampal neurons [53].
Antidepressant-like effect of apigenin was first shown byNakazawa et al. in various behavioral despair test models [54].Further, the central noradrenergic, dopaminergic and seroto-ninergic modulatory activities of apigenin was demonstratedin mice [54]. Apigenin when administered via intraperitonealroute in mice at doses of 12.5 and 25 mg/kg displayed signif-icant anti-immobility action in the FST. Further, at a higherdose of 100 mg/kg, the immobility period in apigenin-treated mice was similar to that of control or vehicle-treated
Table 1. Details of various polyphenols known for their antidepressant activity along with their biological source
and mechanisms of action.
S. no. Polyphenol Biological source Possible mechanism of
antidepressant action
Refs.
1 Curcumin Curcuma longa MAO inhibitory action, modulating BDNF,acting through glutamate receptors,antioxidative effect
[16,36-39]
2 Apigenin Citrus family, leavesof Perilla frutescens
Central noradrenergic, dopaminergic andserotoninergic activity, MAO-A inhibition,acting through hypothalamic pituitary adrenal axis
[56-58]
3 Chlorogenic acid Artemisia capillaris Thunb. Opioidergic pathway [13,67]
4 Amentoflavone Cnestis ferruginea MAO inhibitory action, modulating BDNF,acting via glutamate receptors, antioxidative effect,modulation of GABA-A receptors
[60]
5 Ellagic acid Raspberries, the stem and barkof eucalyptus species and nuts
Adrenergic, serotoninergic and/or NO system [70]
6 FA Ubiquitous plants Serotoninergic pathway [83,84]
7 Fisetin Strawberries and otherfruits and vegetables
Selective MAO-A inhibition,serotoninergic/noradrenergic
[92]
8 Hesperidin Citrus fruits Modulation of kappa opioidergic receptors [120,121]
9 Naringenin Citrus fruits Serotoninergic, noradrenergic [143,144]
10 Nobiletin Citrus fruits Serotoninergic, noradrenergic or dopaminergic [164,165]
11 Proanthocyanidins Grape seed extract,Hibiscus rosa-sinensis
Serotoninergic, noradrenergic and dopaminergic [154]
12 Cis/trans-resveratrol Polygonum cuspidatum, wine MAO-A inhibition [109,110]
13 Rutin Schinus molle Serotoninergic, noradrenergic pathways [132,125]
14 Quercetin St. John’s wort, Gingko biloba Serotoninergic, MAO-A inhibition, modulation ofBDNF-TrkB-PI3K/Akt pathways
[99-101]
BDNF: Brain-derived neurotrophic factor; FA: Ferulic acid; GABA-A: Gamma-aminobutyric acid type A; MAO: Monoamine oxidase; NO: Nitric oxide.
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 5
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

animals. The authors of this study have also measured neuro-transmitters turnover in mouse brain after 40 min stressinduced by forced swimming and its modulation by apigenin.It was found that apigenin at a dose of 25 mg/kg reversed adecrease in dopamine turnover in the amygdale region andincrease in dopamine turnover in the hypothalamus regionof the mouse brain [54]. This study signifies that there is arole of dopamine in the antidepressant-like action of apigenin.Further, the antidepressant-like action of apigenin wasreversed by haloperidol, a dopamine D2 receptor blocker.The antidepressant-like activity of apigenin is not completelyunderstood, however, the molecule has shown to inhibit
MAO enzyme activity with an IC50 value of 6.5 µM. Apige-nin is more effective in inhibiting MAO-A as comparedwith MAO-B activity [55]. MAO inhibitors are effective anti-depressant agents; however, the use of this class of drugs islimited due to their adverse food--drug interactions [24].Besides FST, apigenin was also evaluated in other animalmodels of depression such as chronic mild stress (CMS) [56].Apigenin in this study was administered via oral route. Ithas been found that chronic administration of apigeninreversed CMS-induced decrease in sucrose intake as comparedwith control animals [56]. Furthermore, it was shown thatCMS raised the levels of corticosterone in Wistar rats which
O
OHO
HO
OHO
HO
OHO
OH
O
OOO
OO
O
O
O
OHO
OH
OH
OH
OH
OH
OH
OH
HO
OH
OH
OH
OH
HO O
O
OH
OHHO
HOO
OH
O
O
OHOH
O
O
O
OH
OHO
HO
HO
O
HO
O O
OH
O
O
HO
OH
OHO O
O
OH
OH
OH
Amentoflavone
cis-Resveratrol Apigenin Chlorogenic acid Curcumin
Ferulic acidEllagic acid Fisetin Hesperidin
Rutin Quercetin Gallic acid
Naringenin
Nobiletin Proanthocyanidins Trans-Resveratrol
O
OH
O
OHO
O
OH
OO
O
HO
HO
HO
HO
HO
OH
OH O
O
OH O
OHO
OH
OH
OH
O
O
OH
OH
OHOO
OH
OHHO
O
OH
OOH
HO
OH
OHO OH
HO
OH
OH
Figure 2. Structures of natural polyphenols known for their antidepressant activity.
L. Pathak et al.
6 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

was reversed by chronic administration of apigenin [56]. Thestudy concluded that there are multiple mechanisms withregard to the antidepressant-like activity of apigenin [56].
3.3 AmentoflavoneAmentoflavone is a unique bioflavonoid consisting of apige-nin dimer. It has been shown that amentoflavone could easilycross blood--brain barrier when tested in vitro in primary cellcultures of porcine brain capillary endothelial cells [57]. How-ever, when tested in vivo in a mouse model, amentoflavonehas shown poor blood--brain barrier penetrability [58]. Limitedconcentration of amentoflavone was detected only after intra-peritoneal administration of the molecule at 10 mg/kg. Anin vitro study has suggested that amentoflavone extractedfrom different Hypericum species could inhibit flumazenilbinding to rat brain benzodiazepine binding sites at GABAA
receptor with an IC50 value of 14.9 ± 1.9 nM [59]. The studyfurther demonstrated that amentoflavone binds to the benzo-diazepine site of GABAA receptors [59]. A recent study hasevaluated the antidepressant-like action of amentoflavone.Amentoflavone isolated from root decoction of Cnestis ferrugi-nea (a traditional African herbal medicine often prescribed forpsychiatric disorders) was evaluated for its antidepressant-like activity in mice [60]. The antidepressant-like effect wasstudied using behavioral despair models such as FST andTST in 8-week-old naıve Swiss albino mice. Amentoflavone(6.25 -- 50 mg/kg, p.o.) or imipramine (20 mg/kg, p.o.) wasadministered to mice 1 h prior to the FST. It was foundthat the methanolic extract of C. ferruginea at 12.5 mg/kg,reduced immobility period and simultaneously enhancedswimming behavior in mice subjected to FST [60]. Similarly,the effect of oral administration of amentoflavone in FST,starting from 6.25 mg/kg was found to be more pronouncedand potent in reducing immobility period when comparedwith imipramine (20 mg/kg), a tricyclic antidepressant [60].It is suggested that attempts should be made to enhance theblood--brain barrier penetrability of amentoflavone.
3.4 Chlorogenic acidChlorogenic acid, a polyphenol, has been reported as an anti-oxidant and neuroprotectant in a number of previous stud-ies [61,62]. Chlorogenic acid is abundantly found incoffee [63]. A study showing anxiolytic and antioxidant proper-ties of chlorogenic acid at a dose level of 20 mg/kg has beenreported; the anti-anxiety effect was blocked by flumazenilsuggesting that the anti-anxiety effect of chlorogenic acid isdependent on its activity on GABAA-benzodiazepine recep-tors [62]. Various studies have shown protective effect ofchlorogenic acid in animal models of Alzheimer’s diseaseand cerebral ischemia [63-66]. In one study, chlorogenic acidwas found to inhibit the activity of acetylcholinesteraseenzyme (an enzyme involved in the degradation of acetylcho-line) with an IC50 value of 98.17 µg/ml [63]. Further, themolecule possesses free radical scavenging property with anIC50 value of 3.09 µg/ml [63].
There are not enough studies depicting the antidepressant-like profile of chlorogenic acid. Interestingly, there is a clinicalstudy demonstrating that decaffeinated coffee rich inchlorogenic acid could elevate mood in 39 healthy volunteersdepicting that chlorogenic acid could be useful as an antide-pressant/mood-elevator agent [67]. In one study, chlorogenicacid isolated from Artemisia capillaris Thunb. has shownantidepressant-like activity in chronically restrained mice(2 h/day for 14 days and rest for 14 days without any stress)when tested in mouse FST and TST. The mechanism ofantidepressant action of chlorogenic acid is not clear buthypothesized to act through opioidergic pathway [13].
3.5 Ellagic acidEllagic acid is a polyphenolic compound that occurs in plantssuch as raspberries, pomegranate, the stem and bark of euca-lyptus species and nuts [68]. This bioflavonoid has shown anti-oxidant, antifibrotic, anti-inflammatory, cardioprotective andanticancer properties [68,69]. Few studies have been carried outdemonstrating the antidepressant-like profile of ellagic acid.Girish et al. have shown that ellagic acid at doses of25 -- 100 mg/kg., p.o. when administered both in an acuteand chronic fashion reduces immobility period in both FSTand TST [70]. The effect of ellagic acid in these animal modelswas comparable with fluoxetine, a selective serotonin reuptakeinhibitor [70]. Further, the authors have shown the involve-ment of monoaminergic neurotransmitter receptors (bothserotonin and adrenergic) in the antidepressant-like actionof ellagic acid [70]. The authors further confirmed that thereis no role of opioidergic receptors in the antidepressant-like action of ellagic acid [70]. In another study carried byDhingra and Chhillar, the antidepressant-like activity of ella-gic acid was confirmed in immobilization-induced stress inmice. The authors in the present study induced stress inmice by immobilizing them once for a total of 150 min. Itwas shown that ellagic acid at 17.5 and 35 mg/kg reducedthe immobility period in stressed mice when checked inboth FST and TST animal models. The authors concludedthat there might be involvement of adrenergic, serotoninergicand/or nitric oxide (NO) system in the antidepressant-like action of ellagic acid in these animal models [71]. Thereare no clinical studies to prove the antidepressant-like effectof ellagic acid in major depression.
3.6 Ferulic acidFerulic acid (4-hydroxy-3-methoxycinnamic acid; FA) is aplant constituent known to scavenge free radicals in thebody and thus act as a powerful antioxidant agent [72]. Themolecule has been known to possess multiple pharmacologicaleffects including anti-inflammatory, anticancer, antidiabeticand anti-atherogenic and neuroprotective [73-76]. FA improveslearning and memory in rat model of vascular dementia [77].The authors have concluded three mechanisms of action inthe protective effect of FA in dementia. These mechanismsinclude i) antioxidative potential of FA, ii) improvement of
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 7
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

cholinergic system by FA and iii) antagonizing the action ofexcitatory amino acid [77]. It has been shown that long-term administration of FA prevents brain damage frombeta-amyloid protein and thus could be useful to preventthe occurrence of Alzheimer’s disease [78]. Sodium ferulate, asodium salt of FA is also known to possess sedative activity [79],and the plant extract containing this molecule has found itsusefulness in the treatment of headache and irritability, apo-plexy, stasis, etc. [80,81]. A very recent study has shown theeffect of FA on the proliferation of neural progenitor cellboth in vitro and in vivo [82]. When administered orally inmice, FA enhances the number of newly generated cells inthe dentate gyrus of the hippocampus of corticosterone-treated mice [82]. FA has been shown to possess antidepressantand prokinetic properties [83]. The molecule reduces immobil-ity period in rat FST, its anti-immobility effect was preventedby PCPA (p-chlorophenylalanine; serotonin depleter),WAY-100635 (selective serotonin 5-HT1A antagonist), ketan-serin (selective serotonin 5-HT2 antagonist), sulpiride (adopamine D2 receptor antagonist), SCH233390 (a dopamineD1 receptor antagonist), haloperidol (dopamine D2 receptorantagonist) and yohimbine (selective adrenergic alpha-2antagonist), respectively. In another recent study, FA(0.01 -- 10 mg/kg, p.o.) is active in behavioral despair modelsincluding FST and TST. It has been found that theantidepressant-like activity of this molecule is not dependenton changes in the locomotor activity. Some of the conven-tional antidepressants such as fluoxetine, paroxetine, sertraline(all selective serotonin reuptake inhibitors) enhance theantidepressant-like action of FA, thus demonstrating animportant role of serotonin in the antidepressant actionof this molecule. Further, it is confirmed that bothWAY-100635 (0.1 mg/kg, s.c., a selective 5-HT1A receptorantagonist) or ketanserin (5 mg/kg, i.p., a 5-HT2A receptorantagonist) reverse the antidepressant-like effect of FA inthis animal model [84].
3.7 FisetinFisetin (3,3¢,4¢,7-tetrahydroxyflavone) is a naturally occurringflavonoid and is found abundantly in strawberries and otherfruits and vegetables. Fisetin has a strong anti-inflammatoryproperty in brain microglia cells [85]. In a study carried outby Zheng et al., it was shown that fisetin suppressed the levelsof various inflammatory markers such as TNF-a, NO andprostaglandin E2 (PGE2) in lipopolysaccharide-stimulatedBV-2 microglia cells or primary microglia cultures [85]. Fisetinplays an important role in regulating brain functions [86-91].Antidepressant-like effect of fisetin in behavioral models ofdespair was recently investigated and the results suggestedthat fisetin (10 and 20 mg/kg, p.o.) dose dependently inhib-ited the immobility period in FST and TST without affectingthe locomotor activity [92]. Further, fisetin reversed hypother-mia induced by reserpine (a neurotransmitter-depletingagent). Neurochemical analysis revealed that fisetin couldenhance the levels of noradrenaline and serotonin in the
frontal cortex and hippocampal regions of mouse brain [92].Further, fisetin inhibits the activity of MAO-A enzyme with-out affecting the activity of MAO-B. This suggests that thecompound acts by modulating levels of both noradrenalineand serotonin and possesses no action toward dopaminelevels [92].
3.8 QuercetinQuercetin (3,3¢,4¢,5,7-pentahydroxyflavone) is a bioflavonoidwidely distributed in plant products and exists as glyco-sides [93]. There are numerous findings depicting the antide-pressant activity of food rich in quercetin and its glycosidessince last decade [94-98]. Dixon Clarke and Ramsay havereported that diet rich in quercetin could inhibit MAO activ-ity in brain [99]. This could further prevent neurotransmitterbreakdown [99]. The first study demonstrating theantidepressant-like profile of quercetin was conducted byAnjaneyulu et al. [100]. In this study, quercetin per se wasineffective in mouse FST. However, quercetin showedantidepressant-like action in streptozotocin-induced diabeticmice when tested in FST. The antidepressant-like effect ofquercetin (50 and 100 mg/kg) was comparable with fluoxe-tine (5 mg/kg., i.p.) or imipramine (15 mg/kg., i.p.) instreptozotocin-induced diabetic mice [100]. A similar kind ofstudy was published in 2007 by the same group demonstrat-ing that quercetin could induce depression-like phase andthis property involves the participation of alpha-2 adrenergicreceptors in its mechanism of action [101]. Yohimibine, analpha-2 adrenergic receptor blocker inhibited the behavioraldespair induced by quercetin [101].
Recently, an interesting study has been published depictingthe antistress effect of quercetin. Animals in this study were sub-jected to water immersion-restraint stress [102]. Animals werefound to have activated hypothalamic--pituitary--adrenal (HPAaxis) that was normalized with the treatment of quercetin [102].Quercetin in this study significantly suppressed elevated levelsof plasma corticosterone and adrenocorticotropic hormone.Results have shown that quercetin suppresses the expression ofcorticotrophin-releasing factor (CRF) mRNA [102]. The findingshave been confirmed by another group where CRF was injectedin animals to induce anxiety and depression-like stage and thepretreatment of quercetin reversed the effect of exogenouslyadministered CRF [103]. Quercetin is an interesting compoundto study in various brain disorders. Quercetin has shown to bea weak inhibitor of MAO-A enzyme in brain; however, thecompound does not affect MAO-A activity in mouse intestinalmitochondria. This study reflects that various drug--foodinteractions (e.g., hypertensive crisis observed with tyramine-containing food) that are commonly seen with the use ofcommercially available MAO inhibitors are not seen with theuse of quercetin [104]. Similarly, Yoshino et al. have shownthat quercetin inhibits MAO-A enzyme in mouse brainmitochondria and thus inhibits the breakdown of serotoninneurotransmitter [105]. Quercetin metabolite, quercetin 3-glucu-ronide, has powerful antioxidant activity as shown by
L. Pathak et al.
8 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

Yoshino et al. [105]. Beside MAO inhibiting and antioxidantproperties of quercetin, other factors may also contribute tothe antidepressant-like activity of quercetin. Quercetin is neuro-protective in focal cerebral ischemia model via activation ofBDNF-TrkB-PI3K/Akt signaling pathway. Therefore, querce-tin antidepressant-like activity relates to its ability to modulatelevels of BDNFs [105]. Thus, quercetin seems to be a viablecandidate to be included in antidepressant drug therapy.
3.9 ResveratrolResveratrol (3,4¢,5-trihydroxystilbene), is a natural phenoliccompound present in peanuts, red grapes and wine [106]. Itexists as cis- or trans-isomers [107,108]. It has been found thatresveratrol possesses an inhibitory effect on the uptake of[3H] NA (noradrenaine) and [3H]5-HT (serotonin) in ratsynaptosomes, prevents the uptake of [3H]5-HT in humanplatelets and inhibits MAO isoform activity [109]. The firststudy to demonstrate the antidepressant-like effect of trans-resveratrol in behavioral despair model was carried out byXu et al. [110]. The authors have shown that trans-resveratrol(20 -- 80 mg/kg, p.o.) significantly decreased the immobilityperiod in mouse models of behavioral despair without affect-ing the locomotor activity. Neurochemical analysis has shownthat trans-resveratrol at doses of 40 and 80 mg/kg produces asignificant increase in the levels of noradrenaline and seroto-nin. The mechanism is yet not clear, however, the authorshave demonstrated that trans-resveratrol could significantlyinhibit the activity of MAO-A enzyme [110]. Resveratrol isknown to upregulate the production of astroglia-derivedneurotrophic factors via activation of extracellular signal-regulated kinase 1/2 (ERK1/2) and cAMP responsiveelement-binding protein (CREB) [111]. By contrast, it hasbeen shown recently that resveratrol inhibits hippocampalneurogenesis when tested in healthy adult mice [112]. Also, ithas been shown that resveratrol inhibits the proliferation andsurvival of cultured neural progenitor cells [112]. Therefore,the exact mechanism regarding the antidepressant-like actionof resveratrol is not clear. Moreover, it was demonstrated byDi Liberto et al. that the antidepressant action of resveratrolis sex specific. Resveratrol structure is similar to diethylstilbes-trol, a synthetic estrogen. It was shown that resveratrol signif-icantly increased dopamine transporters in striatal region offemale mice and not in male mice [113]. It is yet not clearhow this will influence the activity of resveratrol in treatingpatients suffering from major depression.
3.10 HesperidinHesperidin is a flavanone glycoside abundantly found in thecitrus fruits [114]. Hesperidin decreases capillary permeabilityand fragility: it is also known as vitamin P [115]. There are sev-eral pharmacological reports on the action of hesperidin thatinclude its sedative potential [116,117], antioxidant ability [118],anti-inflammatory and analgesic properties [119]. Souza et al.demonstrated the antidepressant-like activity of hesperidinin mouse FST and TST [120]. In this study, it was observed
that hesperedin (0.1 -- 1 mg/kg., i.p.) reduced the immobilityperiod in both the tests and the antidepressant-like activitywas independent of alterations in locomotor activity [120]. Ithas been found that hesperidin is effective in TST at a subef-fective dose of 0.01 mg/kg [120]. Further, authors concludedthat the antidepressant-like effect of hesperidin in TST wasdependent on its interaction with the serotoninergic 5-HT1A
receptors. Another study has shown the involvement of kappaopioidergic receptors in the antidepressant-like effect of hesper-idin [121]. The antidepressant-like effect of hesperidin in FSTwas prevented by pretreatment of naloxone (a non-selectiveopioidergic receptor antagonist) and 2-(3,4-dichlorophenyl)-N-methyl-N-[(1S)-1-(3-isothiocyanatophenyl)-2-(1-pyrrolidinyl)ethyl]acetamide (DIPPA (1 mg/kg), a selective k-opioidreceptor antagonist) suggesting the involvement of kappa recep-tors in its antidepressant-like action [121]. Other actions of hes-peridin might also contribute toward its antidepressant-like activity. For example, hesperidin has shown anti-inflammatory and antioxidant activity in an experimental modelof stroke [122]. Interestingly, hesperidin reduced the levels ofinflammatory biomarkers such as TNF-a, IL-1b levels, induc-ible nitric oxide synthase (iNOS) expression and glial fibrillaryacidic protein (GFAP) in middle cerebral artery occluded ratsas compared with their respective controls [122]. In one of thestudies, hersperidin has been shown to enhance neuronal popu-lation when neuronal progenitor cells were treated with hesper-idin. There was an 80% increase in the number of b-tubulin IIIcells in this study [123]. Hesperidin seems to be a viable candidatefor the treatment of major depression.
3.11 RutinRutin, a diglycoside flavonol, is extensively found in manyplants including Hypericum perforatum. St. John’s wort, anextract of H. perforatum, is a traditional remedy for the treat-ment of depression since ages [124]. It has been found thatrutin is an important constituent of the Hypericum extractand is essential for its antidepressant activity [125]. This flavo-nol has been shown to exert several other biological activitiesincluding antimicrobial [126] anti-inflammatory [127], antioxi-dant [128], neuroprotective [129] and antiulcerogenic [130]
potentials. Rutin is observed to be absorbed slowly and lessefficiently as compared with quercetin on the basis of areaunder the plasma concentration--time curve values and rela-tive urinary excretions [131]. In order to study theantidepressant-like profile of rutin, Machado et al. carriedout a study exploring the effect of ethanolic extract of aerialpart of Schinus molle using FST and TST in mice [132]. Theauthors found that administration of rutin reduced the immo-bility period in the TST (0.3 -- 3 mg/kg, p.o.), but not in theFST. Further, pretreatment of mice with PCPA (inhibitor ofserotonin synthesis) or AMPT (a-methyl-p-tyrosine; inhibitorof noradrenaline synthesis) prevented the anti-immobilityeffect of rutin suggesting the involvement of neurotransmit-ters in its antidepressant-like action [132]. Rutin enhanced spa-tial memory in aged rats and further augmented the levels of
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 9
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

noradrenaline and dopamine neurotransmitters in hippocam-pus and hypothalamus regions of aged rat brains, respec-tively [133]. Similar to hesperidin and other polyphenols,rutin has also shown antioxidant and anti-inflammatoryactivities when tested in rats [134]. In streptozotocin-inducedinflammatory rat model, rutin prevented the overexpressionof inflammatory biomarkers such as cyclooxygenase-2(COX-2), GFAP, IL-8, iNOS, nuclear factor-kappaB(NF-kB) [134]. In repeated cerebral ischemic rats, rutin pre-vented spatial memory impairment and neuronal death inhippocampal CA1 region [135]. In contrast to these studies,Silva et al. have shown that rutin has a capacity to induceTNF-a and NO in primary glial cell culture [136]. Also,Boyle et al. conducted a 6-week randomized single-blind placebo controlled trial on 18 healthy female volunteersfor studying the pharmacokinetics of rutin (quercetin-3-O-b-rutinoside [137]. The authors in this study concluded that after6-week administration of rutin, there was no significant anti-oxidant effect observed in the plasma [137]. This might berelated to bioavailability issues associated with rutin. Morestudies are required to establish the use of rutin in braindisorders including its use in major depression.
3.12 NaringeninNaringenin (4,5,7-trihydroxyflavanone) is one of the mostabundant flavanones in citrus fruits [138]. Neuroinflammationhas been speculated to play a major role in the pathophysiol-ogy of major depression. Neuroinflammatory signalingpathways have emerged as new targets for antidepressantdrugs [139]. The fact that naringenin is known to reduceneuroinflammatory injuries in glial cells via inhibiting pro-inflammatory mediators such as p38 signaling cascades andthe STAT-1 transcription factor, it was hypothesized that nar-ingenin might be useful in the treatment of major depres-sion [140]. Like other flavonoids, naringenin also possessesantioxidant and anti-inflammatory properties [141]. Naringe-nin suppressed NF-kB signaling pathway in ischemic strokemodel, suggesting anti-inflammatory property [141]. Naringe-nin has a good oral bioavailability [142]. Naringenin has shownto inhibit the activity of MAO enzyme [143]. Antidepressant-like action of naringenin was investigated by Yi et al. Theauthors in this article have demonstrated that naringenin(10 -- 50 mg/kg) treatment could significantly decrease theimmobility period in the mouse TST but not in the FST.Moreover, pretreatment with PCPA or AMPT significantlyinhibited the anti-immobility effect of naringenin in theTST suggesting the role of serotoninergic and noradrenergicsystems in its antidepressant action [144].
3.13 ProanthocyanidinsProanthocyanidins polyphenols are the major component ofgrape seed extract [145,146]. Proanthocyanidins have demon-strated various pharmacological actions such as antioxi-dant [147], antinociceptive [148] and cardioprotective [149].Lu et al. found that proanthocyanidins-rich grape seed extract
improved the condition of diabetic neuropathy in rats viaacting through cellular signaling pathways such as advancedglycation product (AGEs), nuclear factor-kappa B p65(NF-kB65) [150]. Wang et al. demonstrated that metabolitesof monomeric proanthocyanidins could selectively reach brainand improve cognition by modulating synaptic plasticity inthe brain [151]. When tested in rat hippocampal cells, grapeseed proanthocyanidins inhibited glutamate-induced celldeath via inhibiting calcium signals and NO formation [152].In earlier findings, proanthocyanidins-rich dietary food con-stituents inhibited the activity of MAO enzyme in C6 glialcells [153]. However, the antidepressant-like activity of theproanthocyanidins in animal models was first demonstratedby Xu et al. [154]. The authors demonstrated effectiveness ofproanthocyanidins in animal models of behavioral despairafter oral administration in mice at dose range of 25 and50 mg/kg for 7 days. The antidepressant-like activity in thisanimal model is devoid of alterations in the locomotor activityof mice [154]. Further, neurochemical evidences revealedthat proanthocyanidins increases the levels of serotonin,noradrenaline and dopamine in various regions of the brainsuggesting the role of central monoaminergic system in itsantidepressant-like action [154].
3.14 NobiletinNobiletin is a polymethoxylated flavone abundantly found incitrus fruits [155,156]. Nobiletin has vast pharmacologicalactions including its applications in various clinical conditionssuch as cataract [157], obesity [158], metastasis [159], neuroin-flammation [160] and hyperglycemia [161]. Nobiletin has alsoshown activity in various animal models of CNS disorders.In one of the studies, nobiletin exerted neuroprotectiveeffects on ischemia-induced neuronal cell death in hippocam-pal CA1 region of the brain [162]. In another study,Matsuzaki et al. demonstrated neurotrophic property of nobi-letin in murine hippocampus [163]. The antidepressant-like activity of the nobiletin was demonstrated for the firsttime in animals by Yi et al. [164]. The authors in this studyadministered nobiletin to mice and observed a decrease inimmobility period in both FST and TST at a dose range of25 -- 100 mg/kg per oral. The antidepressant-like action ofnobiletin in this study was devoid of alterations in thelocomotor activity of mice as assessed by open-field test [164].Further, it has been shown that pretreatment of mice withWAY-100635 (a serotonin 5-HT1A receptor antagonist),cyproheptadine (a serotonin 5-HT2 receptor antagonist), pra-zosin (an a1-adrenoceptor antagonist), SCH23390 (a dopa-mine D1 receptor antagonist) or sulpiride (a dopamineD2 receptor antagonist) prevented the anti-immobility effectof nobiletin in TST. By contrast, pretreatment of animalswith yohimbine (an a2-adrenoceptor antagonist) or proprano-lol (a b-adrenoceptor antagonist) did not affect theantidepressant-like action of nobiletin in this animal model [164].The study confirmed the involvement of serotoninergic, norad-renergic or dopaminergic systems in antidepressant action of
L. Pathak et al.
10 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

nobiletin [164]. Nobiletin (oral administration for 11 days) alsoreversed memory impairment in olfactory-bulbectomizedmice [165] suggesting its antidepressant potential.
Polyphenols which were investigated during last decade fortheir antidepressant and anxiolytic activities are summarizedin Table 1.
4. Expert opinion
Major depression is a serious psychiatric disorder that requirespharmacological intervention. Besides the availability of someof the wonderful antidepressant molecules, research is stillgoing on to explore novel antidepressant molecules withunique mechanism of action. These novel antidepressantsare expected to possess better efficacy and safety profiles com-pared with existing ones. As discussed in previous sections ofthis review article, polyphenols have shown antidepressant-like activity in various animal models of major depression.Various mechanisms have been put forward to justify the anti-depressant potential of these molecules. These include:i) modulation of various neurotransmitters such as norepi-nephrine, serotonin and dopamine, ii) antioxidant property,iii) anti-inflammatory potential, iv) ability to modulate the
activity of MAO enzyme and v) acting on neurotransmitterreceptor systems. These mechanisms are listed in Figure 3.
Despite an extensive research on the use of natural polyphe-nols in neurological and psychiatric disorders, we have not yetreached to a stage that could lead to prescription of these pol-yphenols for the treatment of major depression. Research inthis potential area is much slower and ineffective due to thefollowing reasons: i) none of the animal models mimic anexact state of human major depression. An animal model ofmajor depression should possess all the cardinal features ofthe disease condition; ii) the exact pathophysiology of majordepression is not yet known because of the complex natureof human brain and iii) there are not enough clinical eviden-ces demonstrating the antidepressant profile of these polyphe-nols. It is proposed that these polyphenols could be useful inthe treatment of major depression provided proper preclinicaland clinical studies are carried out to understand their phar-macokinetic and pharmacodynamic properties.
Curcumin is considered as a wonder molecule that is exten-sively studied for its use in various body disorders includingits effectiveness in major depression. Curcumin has showneffective antidepressant-like action in various animal modelsof depression. Researchers are preparing various analogs of
MAO inhibition
Modulating BDNF
Acting through glutamatereceptors
Anti-oxidative effect
Serotoninergic,nor-adrenergic ordopaminergic
Selective MAO-Ainhibition
Serotoninergic/nor-adrenergic
Serotoninergic,noradrenergicpathways
Serotoninergic
MAO-A inhibition
Modulation of BDNF-TrκB-Pi\I3K/Akt pathways
MAO-Ainhibition
Modulation of GABA-Areceptors
Anti-oxidative effect
Modulation of BDNF
MAO inhibition
Acting throughhypothalamic pituitaryadrenal axis
Modulation ofnitric oxide system
Serotoninergic
Adrenergic
Serotoninergicpathway
Serotoninergicnor-adrenergicordopaminergic
Serotoninergic, nor-adrenergic
Modulation ofκ opioidergicreceptors
Curcumin
Proanthocyanidins
Fisetin
Rutin
Quercetin Resveratrol
Opioidergicpathway
MAO-A inhibition
Central noradrenergic,dopaminergic andserotoninenergic activity
Amentoflavone
Apigenin
Ellagic acid
Ferulic acid
Chlorogenic acidHesperidin, Naringenin and Nobiletin
Naturalpolyphenols
Acting via glutamatereceptors
Figure 3. Mechanism of action of various polyphenols depicting antidepressant activity.
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 11
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

curcumin that could enhance its bioavailability and blood--brain barrier penetrability. The bioavailability of these moleculescould be enhanced by using different approaches such as makingwater-soluble analogs of aqueous insoluble polyphenols or viausing nanotechnological intervention. Another important issuewith the use of polyphenols is their short half-life. For example,green tea polyphenols have a short half-life of about 3 h. There-fore, it will be necessary to drink green tea at least 8 times in aday to have antidepressant action. It is very important toenhance the half-life of such polyphenols either by making novelanalogs or by preventing their metabolism. It is a common myththat natural compounds are very safe. Some of these polyphenolsat higher doses have potential side effects that are listedin Table 2. Therefore, it is mandatory to perform clinical studiesto extract out the side-effect profile of these natural polyphenols.
It is expected that these polyphenols might be prescribed asan adjuvant therapy with already existing antidepressant mol-ecules in the treatment of major depression. Alternatively,these molecules could be prescribed in pharmacoresistantpatients where 3 or > 3 drugs are found to be ineffective inrelieving the symptoms. In summary, polyphenols researchis an exciting field to study and there are lots of opportunitiesin this upcoming area of research.
Declaration of interest
A Dhir is a Fellow of Department of Biotechnology, NewDelhi. The authors state no conflict of interest and havereceived no payment in preparation of this manuscript.
Table 2. Potential side effects of natural polyphenols listed for antidepressant activity.
S. no. Polyphenol Side effects Refs.
1 Curcumin Cytotoxic and genotoxic potential on human peripheral lymphocytes athigh doses are observed
[166]
2 Apigenin - -3 Chlorogenic acid Inflammation at high doses when tested in rats [167]
4 Amentoflavone Mutagenic when tested in Salmonella typhimurium [168]
5 Ellagic acid - -6 Hesperidin - -7 FA - -8 Fisetin - -9 Naringenin Known to induce expression of CYP1A1 in the colon tissue and thus
could promote colorectal carcinomas[169]
10 Nobiletin - -11 Resveratrol Inhibit various metabolizing enzymes leading to food--drug interactions [170]
12 Proanthocyanidins Inhibits iron absorption and therefore toxic when given in iron deficiency anemia [171]
13 Quercetin Depletion of intracellular glutathione content, total RNA amount and cellregulating genes, induction of CYP1A1 in colon and liver microsomes
[169,172]
14 Rutin - -
CYP1A1: Cytochrome 1A1; FA: Ferulic acid.
L. Pathak et al.
12 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

BibliographyPapers of special note have been highlighted as
either of interest (�) or of considerable interest(��) to readers.
1. Perron NR, Brumaghim JL. A review of
the antioxidant mechanisms of polyphenol
compounds related to iron binding.
Cell Biochem Biophys 2009;53:75-100
2. Manach C, Williamson G, Morand C,
et al. Bioavailability and bioefficacy of
polyphenols in humans. I. Review of
97 bioavailability studies. Am J
Clin Nutr 2005;81:230S-42S. Extensive study demonstrating
bioavailability of food containing
polyphenols in humans.
3. Han XH, Hong SS, Hwang JS, et al.
Monoamine oxidase inhibitory
components from Cayratia japonica.
Arch Pharm Res 2007;30:13-17
4. Agouni A, Lagrue-Lak-Hal AH,
Mostefai HA, et al. Red wine
polyphenols prevent metabolic and
cardiovascular alterations associated with
obesity in Zucker fatty rats (Fa/Fa).
PLoS One 2009;4:e5557
5. Visioli F, Galli C. The effect of minor
constituents of olive oil on cardiovascular
disease: new findings. Nutr Rev
1998;56:142-7
6. Middleton E Jr, Kandaswami C,
Theoharides TC. The effects of plant
flavonoids on mammalian cells:
implications for inflammation, heart
disease, and cancer. Pharmacol Rev
2000;52:673-751
7. Urquiaga I, Leighton F. Plant polyphenol
antioxidants and oxidative stress. Biol Res
2000;33:55-64
8. Visioli F, Galli C. The role of
antioxidants in the Mediterranean diet.
Lipids 2001;36:S49-52
9. Simonyi A, Wang Q, Miller RL, et al.
Polyphenols in cerebral ischemia: novel
targets for neuroprotection.
Mol Neurobiol 2005;31:135-47
10. Scalbert A, Manach C, Morand C, et al.
Dietary polyphenols and the prevention
of diseases. Crit Rev Food Sci Nutr
2005;45:287-306
11. Tian WX. Inhibition of fatty acid
synthase by polyphenols.
Curr Med Chem 2006;13:967-77
12. Yoshimura Y, Nakazawa H,
Yamaguchi F. Evaluation of the NO
scavenging activity of procyanidin in
grape seed by use of the TMA-PTIO/
NOC 7 ESR system. J Agric Food Chem
2003;51:6409-12
13. Park S, Sima Y, Hanb P, et al.
Antidepressant-like effect of chlorogenic
acid isolated from Artemisia capillaris
Thunb. Animal Cells Syst 2010;14:253-9
14. Arora V, Kuhad A, Tiwari V, et al.
Curcumin ameliorates reserpine-induced
pain-depression dyad: behavioural,
biochemical, neurochemical and
molecular evidences.
Psychoneuroendocrinology
2011;36:1570-81
15. Mei X, Xu D, Xu S, et al.
Gastroprotective and antidepressant
effects of a new zinc(II)-curcumin
complex in rodent models of gastric ulcer
and depression induced by stresses.
Pharmacol Biochem Behav
2011;99:66-74
16. Kulkarni SK, Bhutani MK, Bishnoi M.
Antidepressant activity of curcumin:
involvement of serotonin and dopamine
system. Psychopharmacology (Berl)
2008;201:435-42
17. Sanmukhani J, Anovadiya A,
Tripathi CB. Evaluation of
antidepressant like activity of curcumin
and its combination with fluoxetine and
imipramine: an acute and chronic study.
Acta Pol Pharm 2011;68:769-75
18. Wong ML, Licinio J. Research and
treatment approaches to depression.
Nat Rev Neurosci 2001;2:343-51
19. Wong ML, Licinio J. From monoamines
to genomic targets: a paradigm shift for
drug discovery in depression. Nat Rev
Drug Discov 2004;3:136-51. Novel targets for antidepressants are
discussed in detail.
20. Kessler RC, Berglund P, Demler O,
et al. National Comorbidity Survey
Replication. The epidemiology of major
depressive disorder: results from the
National Comorbidity Survey Replication
(NCS-R). JAMA 2003;289:3095-4105
21. Greist JH, Mundt JC, Kobak K. Factors
contributing to failed trials of new
agents: can technology prevent some
problems? J Clin Psychiatry 2002;2:8-13
22. Artigas F, Romero L, De Montigny C,
et al. Acceleration of the effect of selected
antidepressant drugs in major depression
by 5-HT1A antagonists. Trends Neurosci
1996;19:378-83
23. Cryan JF, Leonard BE. Depression: from
psychopathology to pharmacotherapy.
Mod Trends Pharmacopsychiat
2010;27:1-19
24. Kulkarni SK, Dhir A. Current
investigational drugs for major
depression. Expert Opin Investig Drugs
2009;18:767-88.. Comprehensive review depicting
various therapeutic regimens for the
treatment of major depression.
25. Andrews JM, Ninan PT, Nemeroff CB.
Venlafaxine: a novel antidepressant that
has a dual mechanism of action.
Depression 1996;4:48-56
26. Al-Harbi KS. Treatment-resistant
depression: therapeutic trends, challenges,
and future directions.
Patient Prefer Adherence 2012;6:369-88
27. Joffe RT, Levitt AJ, Sokolov ST.
Augmentation strategies.
J Clin Psychiatry 1996;57:2-31
28. Montgomery SA. Why do we need new
and better antidepressants?
Int Clin Psychopharmacol
2006;21:S1-S10.. Loopholes in the available therapeutic
interventions are discussed in detail.
29. Petit-Demouliere B, Chenu F,
Bourin M. Forced swimming test in
mice: a review of antidepressant activity.
Psychopharmacology (Berl)
2005;177:245-55
30. Rupniak NM. Animal models of
depression: challenges from a drug
development perspective.
Behav Pharmacol 2003;14:385-90
31. Yan HC, Cao X, Das M, et al.
Behavioral animal models of depression.
Neurosci Bull 2010;26:327-37
32. Borsini F, Lecci A, Sessarego A, et al.
Discovery of antidepressant activity by
forced swimming test may depend on
pre-exposure of rats to a stressful
situation. Psychopharmacology (Berl)
1989;97:183-8
33. Ebrahimi A, Schluesener H. Natural
polyphenols against neurodegenerative
disorders: potentials and pitfalls.
Ageing Res Rev 2012;11:329-45
34. Kulkarni SK, Dhir A. An overview of
curcumin in neurological disorders.
Indian J Pharm Sci 2010;72:149-54
35. Darvesh AS, Carroll RT, Bishayee A,
et al. Curcumin and neurodegenerative
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 13
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

diseases: a perspective. Expert Opin
Investig Drugs 2012;21:1123-40. Pluripotent actions of curcumin on
various therapeutic targets in CNS are
discussed in detail.
36. Xu Y, Ku B, Cui L, et al. Curcumin
reverses impaired hippocampal
neurogenesis and increases serotonin
receptor 1A mRNA and brain-derived
neurotrophic factor expression in
chronically stressed rats. Brain Res
2007;1162:9-18
37. Xu Y, Ku BS, Yao HY, et al. The effects
of curcumin on depressive-like behaviors
in mice. Eur J Pharmacol 2005;518:40-6
38. Xu Y, Ku BS, Yao HY, et al.
Antidepressant effects of curcumin in the
forced swim test and olfactory
bulbectomy models of depression in rats.
Pharmacol Biochem Behav
2005;82:200-6
39. Wang R, Xu Y, Wu HL, et al. The
antidepressant effects of curcumin in the
forced swimming test involve 5-HT1 and
5-HT2 receptors. Eur J Pharmacol
2008;578:43-50
40. Li YC, Wang FM, Pan Y, et al.
Antidepressant like effects of curcumin
on serotonergic receptor-coupled AC-
cAMP pathway in chronic unpredictable
mild stress of rats.
Prog Neuropsychopharmacol
Biol Psychiatry 2009;33:435-49
41. Lin TY, Lu CW, Wang CC, et al.
Curcumin inhibits glutamate release in
nerve terminals from rat prefrontal
cortex: possible relevance to its
antidepressant mechanism.
Prog Neuropsychopharmacol
Biol Psychiatry 2011;35:1785-93
42. Zhang L, Xu T, Wang S, et al.
NMDA GluN2B receptors involved in
the antidepressant effects of curcumin in
the forced swim test.
Prog Neuropsychopharmacol
Biol Psychiatry 2013;40:12-17
43. Wang R, Li YB, Li YH, et al. Curcumin
protects against glutamate excitotoxicity
in rat cerebral cortical neurons by
increasing brain-derived neurotrophic
factor level and activating TrkB.
Brain Res 2008;1210:84-91
44. Wang R, Li YH, Xu Y, et al. Curcumin
produces neuroprotective effects via
activating brain-derived neurotrophic
factor/TrkB-dependent MAPK and
PI-3K cascades in rodent cortical
neurons. Prog Neuropsychopharmacol
Biol Psychiatry 2010;34:147-53
45. Huang Z, Zhong XM, Li ZY, et al.
Curcumin reverses corticosterone-induced
depressive-like behavior and decrease in
brain BDNF levels in rats. Neurosci Lett
2011;493:145-8
46. Hurley LL, Akinfiresoye L, Nwulia E,
et al. Antidepressant-like effects of
curcumin in WKY rat model of
depression is associated with an increase
in hippocampal BDNF. Behav Brain Res
2013;239:27-30
47. Johnson JL, Rupasinghe SG, Stefani F,
et al. Citrus flavonoids luteolin, apigenin,
and quercetin inhibit glycogen synthase
kinase-3b enzymatic activity by lowering
the interaction energy within the binding
cavity. J Med Food 2011;14:325-33
48. Avallone R, Zanoli P, Puia G, et al.
Pharmacological profile of apigenin, a
flavonoid isolated from Matricaria
chamomilla. Biochem Pharmacol
2000;59:1387-94
49. Zanoli P, Avallone R, Baraldi M.
Behavioral characterisation of the
flavonoids apigenin and chrysin.
Fitoterapia 2000;71:S117-23
50. Liu C, Tu FX, Chen X. Neuroprotective
effects of apigenin on acute transient
focal cerebral ischemia-reperfusion injury
in rats. Zhong Yao Cai 2008;31:870-3
51. Rezai-Zadeh K, Ehrhart J, Bai Y, et al.
Apigenin and luteolin modulate
microglial activation via inhibition of
STAT1-induced CD40 expression.
J Neuroinflammation 2008;5:41
52. Liu R, Zhang T, Yang H, et al. The
flavonoid apigenin protects brain
neurovascular coupling against
amyloid-b-induced toxicity in mice.
J Alzheimers Dis 2011;24:85-100
53. Han JY, Ahn SY, Kim CS, et al.
Protection of apigenin against
kainate-induced excitotoxicity by
anti-oxidative effects. Biol Pharm Bull
2012;35:1440-6
54. Nakazawa T, Yasuda T, Ueda J, et al.
Antidepressant-like effects of apigenin
and 2,4,5-trimethoxycinnamic acid from
Perilla frutescens in the forced swimming
test. Biol Pharm Bull 2003;26:474-80
55. Han XH, Hong SS, Hwang JS, et al.
Monoamine oxidase inhibitory
components from Cayratia japonica.
Arch Pharm Res 2007;30:13-17
56. Yi LT, Li JM, Li YC, et al.
Antidepressant-like behavioral and
neurochemical effects of the
citrus-associated chemical apigenin.
Life Sci 2008;82:741-51
57. Gutmann H, Bruggisser R, Schaffner W,
et al. Transport of amentoflavone across
the blood-brain barrier in vitro.
Planta Med 2002;68:804-7
58. Colovic M, Fracasso C, Caccia S.
Brain-to-plasma distribution ratio of the
biflavone amentoflavone in the mouse.
Drug Metab Lett 2008;2:90-4
59. Baureithel KH, Buter KB, Engesser A,
et al. Inhibition of benzodiazepine
binding in vitro by amentoflavone, a
constituent of various species of
Hypericum. Pharm Acta Helv
1997;72:153-7
60. Ishola IO, Chatterjee M, Tota S, et al.
Antidepressant and anxiolytic effects of
amentoflavone isolated from Cnestis
ferruginea in mice.
Pharmacol Biochem Behav
2012;103:322-31
61. Chu YF, Brown PH, Lyle BJ, et al.
Roasted coffees high in lipophilic
antioxidants and chlorogenic acid
lactones are more neuroprotective than
green coffees. J Agric Food Chem
2009;57:9801-8
62. Bouayed J, Rammal H, Dicko A, et al.
Chlorogenic acid, a polyphenol from
Prunus domestica (Mirabelle), with
coupled anxiolytic and antioxidant
effects. J Neurol Sci 2007;262:77-84
63. Kwon SH, Lee HK, Kim JA, et al.
Neuroprotective effects of chlorogenic
acid on scopolamine-induced amnesia via
anti-acetylcholinesterase and
anti-oxidative activities in mice.
Eur J Pharmacol 2010;649:210-17
64. Oboh G, Agunloye OM, Akinyemi AJ,
et al. Comparative study on the
inhibitory effect of caffeic and
chlorogenic acids on key enzymes linked
to Alzheimer’s disease and some
pro-oxidant induced oxidative stress in
rats’ brain-in vitro. Neurochem Res
2012;published online November 2012;
doi:10.1007/s11064-012-0935-6
65. Ahn EH, Kim DW, Shin MJ, et al.
Chlorogenic acid improves
neuroprotective effect of
PEP-1-ribosomal protein S3 against
ischemic insult. Exp Neurobiol
2011;20:169-75
L. Pathak et al.
14 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

66. Lapchak PA. The phenylpropanoid
micronutrient chlorogenic acid improves
clinical rating scores in rabbits following
multiple infarct ischemic strokes:
synergism with tissue plasminogen
activator. Exp Neurol 2007;205:407-13
67. Cropley V, Croft R, Silber B, et al. Does
coffee enriched with chlorogenic acids
improve mood and cognition after acute
administration in healthy elderly? A pilot
study. Psychopharmacology (Berl)
2012;219:737-49
68. Girish C, Pradhan SC. Drug
development for liver diseases: focus on
picroliv, ellagic acid and curcumin.
Fundam Clin Pharmacol 2008;22:623-32
69. Uzar E, Alp H, Cevik MU, et al. Ellagic
acid attenuates oxidative stress on brain
and sciatic nerve and improves
histopathology of brain in
streptozotocin-induced diabetic rats.
Neurol Sci 2012;33:567-74
70. Girish C, Raj V, Arya J, et al. Evidence
for the involvement of the
monoaminergic system, but not the
opioid system in the antidepressant-like
activity of ellagic acid in mice.
Eur J Pharmacol 2012;682:118-25
71. Dhingra D, Chhillar R.
Antidepressant-like activity of ellagic acid
in unstressed and acute
immobilization-induced stressed mice.
Pharmacol Rep 2012;64:796-807
72. Srinivasan M, Sudheer AR, Menon VP.
Ferulic acid: therapeutic potential
through its antioxidant property. J Clin
Biochem Nutr 2007;40:92-100
73. Kawabata K, Yamamoto T, Hara A,
et al. Modifying effects of ferulic acid on
azoxymethane-induced colon
carcinogenesis in F344 rats. Cancer Lett
2000;157:15-21
74. Balasubashini MS, Rukkumani R,
Viswanathan P, et al. Ferulic acid
alleviates lipid peroxidation in diabetic
rats. Phytother Res 2004;18:310-14
75. Sultana R, Ravagna A,
Mohmmad-Abdul H, et al. Ferulic acid
ethyl ester protects neurons against
amyloid beta- peptide(1-42)-induced
oxidative stress and neurotoxicity:
relationship to antioxidant activity.
J Neurochem 2005;92:749-58
76. Yogeeta SK, Hanumantra RB,
Gnanapragasam A, et al. Attenuation of
abnormalities in the lipid metabolism
during experimental myocardial
infarction induced by isoproterenol in
rats: beneficial effect of ferulic acid and
ascorbic acid. Basic Clin
Pharmacol Toxicol 2006;98:467-72
77. Luo Y, Zhao HP, Zhang J, et al. Effect
of ferulic acid on learning and memory
impairments of vascular dementia rats
and its mechanism of action. Yao Xue
Xue Bao 2012;47:256-60
78. Yan JJ, Cho JY, Kim HS, et al.
Protection against beta-amyloid peptide
toxicity in vivo with long-term
administration of ferulic acid.
Br J Pharmacol 2001;133:89-96
79. Zhang YP, Yu LJ, Ma RD. Sedative and
hypnotic effect of sodium ferulate in
mice. Nei Mongol J Trad Chinese Med
2008;27:9-11
80. Li S. Compendium of materia medica
(reprinted from 1590 Jinling
wood-engraved edition). People’s Health
Press; Beijing: 2004
81. Jiangsu college of new medicine,
dictionary of traditional Chinese and
herbal medicine, Shanghai:1986
82. Yabe T, Hirahara H, Harada N, et al.
Ferulic acid induces neural progenitor
cell proliferation in vitro and in vivo.
Neuroscience 2010;165:515-24
83. Zhang YJ, Huang X, Wang Y, et al.
Ferulic acid-induced anti-depression and
prokinetics similar to Chaihu-Shugan-San
via polypharmacology. Brain Res Bull
2011;86:222-8
84. Zeni AL, Zomkowski AD, Maraschin M,
et al. Ferulic acid exerts
antidepressant-like effect in the tail
suspension test in mice: evidence for the
involvement of the serotonergic system.
Eur J Pharmacol 2012;679:68-74
85. Zheng LT, Ock J, Kwon BM, et al.
Suppressive effects of flavonoid fisetin on
lipopolysaccharide-induced microglial
activation and neurotoxicity.
Int Immunopharmacol 2008;8:484-94
86. Maher P, Akaishi T, Abe K. Flavonoid
fisetin promotes ERK-dependent long-
term potentiation and enhances memory.
Proc Natl Acad Sci USA
2006;103:16568-73
87. Ishige K, Schubert D, Sagara Y.
Flavonoids protect neuronal cells from
oxidative stress by three distinct
mechanisms. Free Radic Biol Med
2001;30:433-46
88. Maher P. The flavonoid fisetin promotes
nerve cell survival from trophic factor
withdrawal by enhancement of
proteasome activity.
Arch Biochem Biophys 2008;476:139-44
89. Maher P. Modulation of multiple
pathways involved in the maintenance of
neuronal function during aging by
fisetin. Genes Nutr 2009;4:297-307
90. Maher P, Dargusch R, Bodai L, et al.
ERK activation by the polyphenols fisetin
and resveratrol provides neuroprotection
in multiple models of Huntington’s
disease. Hum Mol Genet
2011;20:261-70
91. Akaishi T, Morimoto T, Shibao M, et al.
Structural requirements for the flavonoid
fisetin in inhibiting fibril formation of
amyloid beta protein. Neurosci Lett
2008;444:280-5
92. Zhen L, Zhu J, Zhao X, et al. The
antidepressant-like effect of fisetin
involves the serotonergic and
noradrenergic system. Behav Brain Res
2012;228:359-66
93. Lam TK, Shao S, Zhao Y, et al.
Influence of quercetin-rich food intake
on microRNA expression in lung cancer
tissues. Cancer Epidemiol
Biomarkers Prev 2012;21:2176-84
94. Butterweck V, Jurgenliemk G,
Nahrstedt A, et al. Flavonoids from
Hypericum perforatum show
antidepressant activity in the forced
swimming test. Planta Med 2000;66:3-6
95. Sakakibara H, Yoshino S, Kawai Y, et al.
Antidepressant-like effect of onion
(Allium cepa L.) powder in a rat
behavioral model of depression.
Biosci Biotechnol Biochem
2008;72:94-100
96. Sakakibara H, Ishida K, Grundmann O,
et al. Antidepressant effect of extracts
from Ginkgo biloba leaves in behavioral
models. Biol Pharm Bull
2006;29:1767-70
97. Paulke A, Noldener M,
Schubert-Zsilavecz M, et al. St. John’s
wort flavonoids and their metabolites
show antidepressant activity and
accumulate in brain after multiple oral
doses. Pharmazie 2008;63:296-302
98. Haleagrahara N, Radhakrishnan A,
Lee N, et al. Flavonoid quercetin
protects against swimming stress-induced
changes in oxidative biomarkers in the
hypothalamus of rats. Eur J Pharmacol
2009;621:46-52
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 15
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

99. Dixon Clarke SE, Ramsay RR. Dietary
inhibitors of monoamine oxidase. A. J,
Neural Transm 2011;118:1031-41
100. Anjaneyulu M, Chopra K, Kaur I.
Antidepressant activity of quercetin, a
bioflavonoid, in streptozotocin-induced
diabetic mice. J Med Food 2003;6:391-5
101. Kaur R, Chopra K, Singh D. Role of
alpha2 receptors in quercetin-induced
behavioral despair in mice. J Med Food
2007;10:165-8
102. Kawabata K, Kawai Y, Terao J.
Suppressive effect of quercetin on acute
stress-induced
hypothalamic-pituitary-adrenal axis
response in Wistar rats. J Nutr Biochem
2010;21:374-80
103. Bhutada P, Mundhada Y, Bansod K,
et al. Reversal by quercetin of
corticotrophin releasing factor induced
anxiety- and depression-like effect in
mice. Prog Neuropsychopharmacol
Biol Psychiatry 2010;34:955-60
104. Bandaruk Y, Mukai R, Kawamura T,
et al. Evaluation of the inhibitory effects
of quercetin-related flavonoids and tea
catechins on the monoamine
oxidase-A reaction in mouse brain
mitochondria. J Agric Food Chem
2012;60:10270-7
105. Yoshino S, Hara A, Sakakibara H, et al.
Effect of quercetin and glucuronide
metabolites on the monoamine
oxidase-A reaction in mouse brain
mitochondria. Nutrition 2011;27:847-52
106. Park HR, Kong KH, Yu BP, et al.
Resveratrol inhibits the proliferation of
neural progenitor cells and hippocampal
neurogenesis. J Biol Chem
2012;287:42588-600
107. Aggarwal BB, Bhardwaj A, Aggarwal RS,
et al. Role of resveratrol in prevention
and therapy of cancer: preclinical and
clinical studies. Anticancer Res
2004;24:2783-840
108. Fremont L. Biological effects of
resveratrol. Life Sci 2000;66:663-73
109. Yanez M, Fraiz N, Cano E, et al.
Inhibitory effects of cis- and
trans-resveratrol on noradrenaline and
5-hydroxytryptamine uptake and on
monoamine oxidase activity.
Biochem Biophys Res Commun
2006;344:688-95
110. Xu Y, Wang Z, You W, et al.
Antidepressant-like effect of trans-
resveratrol: involvement of serotonin and
noradrenaline system.
Eur Neuropsychopharmacol
2010;20:405-13
111. Zhang F, Lu YF, Wu Q, et al.
Resveratrol promotes neurotrophic factor
release from astroglia. Exp Biol Med
(Maywood) 2012;237:943-8
112. Park HR, Kong KH, Yu BP, et al.
Resveratrol inhibits the proliferation of
neural progenitor cells and hippocampal
neurogenesis. J Biol Chem
2012;287:42588-600
113. Di Liberto V, Makela J, Korhonen L,
et al. Involvement of estrogen receptors
in the resveratrol-mediated increase in
dopamine transporter in human
dopaminergic neurons and in striatum of
female mice. Neuropharmacology
2012;62:1011-18
114. Wasowski C, Loscalzo LM, Higgs J,
et al. Chronic intraperitoneal and oral
treatments with hesperidin induce central
nervous system effects in mice.
Phytother Res 2012;26:308-12
115. Morii S. Research for vitamin-P.
J Biochem 1939;29:487-501
116. Fernandez SP, Wasowski C,
Paladini AC, et al. Synergistic interaction
between hesperidin, a natural flavonoid,
and diazepam. Eur J Pharmacol
2005;512:189-98
117. Guzman-Gutierrez SL, Navarrete A.
Pharmacological exploration of the
sedative mechanism of hesperidin
identified as the active principle of Citrus
sinensis flowers. Planta Med
2009;75:295-301
118. Hirata A, Murakami Y, Shoji M, et al.
Kinetics of radical-scavenging activity of
hesperetin and hesperidin and their
inhibitory activity on COX-2 expression.
Anticancer Res 2005;25:3367-74
119. Galati EM, Monforte MT,
Kirjavainen S, et al. Biological effects of
hesperidin, a citrus flavonoid. (note I):
anti-inflammatory and analgesic activity.
Farmaco 1994;49:709-12
120. Souza LC, de Gomes MG, Goes AT,
et al. Evidence for the involvement of the
serotonergic 5-HT(1A) receptors in the
antidepressant-like effect caused by
hesperidin in mice.
Prog Neuropsychopharmacol
Biol Psychiatry 2013;40:103-9
121. Filho CB, Fabbro LD, de Gomes MG,
et al. Kappa-opioid receptors mediate the
antidepressant-like activity of hesperidin
in the mouse forced swimming test.
Eur J Pharmacol 2013;698:286-91
122. Raza SS, Khan MM, Ahmad A, et al.
Hesperidin ameliorates functional and
histological outcome and reduces
neuroinflammation in experimental
stroke. Brain Res 2011;1420:93-105
123. Nones J, E Spohr TC, Gomes FC.
Hesperidin, a flavone glycoside, as
mediator of neuronal survival.
Neurochem Res 2011;36:1776-84
124. Wentworth JM, Agostini M, Love J,
et al. St John’s wort, a herbal
antidepressant, activates the steroid X
receptor. J Endocrinol 2000;166:R11-16
125. Noldner M, Sch€otz K. Rutin is essential
for the antidepressant activity of
Hypericum perforatum extracts in the
forced swimming test. Planta Med
2002;68:577-80.. Rutin has been shown as important
active constituent for antidepressant
action of H. perforatum.
126. Arima H, Ashida H, Danno G.
Rutin-enhanced antibacterial activities of
flavonoids against Bacillus cereus and
Salmonella enteritidis.
Biosci Biotechnol Biochem
2002;66:1009-14
127. Guardia T, Rotelli AE, Juarez AO, et al.
Anti-inflammatory properties of plant
flavonoids. Effects of rutin, quercetin and
hesperidin on adjuvant arthritis in rat.
Farmaco 2001;56:683-7
128. Ostrakhovitch EA, Afanas’ev IB.
Oxidative stress in rheumatoid arthritis
leukocytes: suppression by rutin and
other antioxidants and chelators.
Biochem Pharmacol 2001;62:743-6
129. Gupta R, Singh M, Sharma A.
Neuroprotective effect of antioxidants on
ischaemia and reperfusion-induced
cerebral injury. Pharmacol Res
2003;48:209-15
130. La Casa C, Villegas I, Alarcon de la
Lastra C, et al. Evidence for protective
and antioxidant properties of rutin, a
natural flavone, against ethanol induced
gastric lesions. J Ethnopharmacol
2000;71:45-53
131. Manach C, Morand C, Demigne C,
et al. Bioavailability of rutin and
quercetin in rats. FEBS Lett
1997;409:12-16
132. Machado DG, Bettio LE, Cunha MP,
et al. Antidepressant-like effect of rutin
isolated from the ethanolic extract from
L. Pathak et al.
16 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

Schinus molle L. in mice: evidence for
the involvement of the serotonergic and
noradrenergic systems. Eur J Pharmacol
2008;587:163-8
133. Pyrzanowska J, Piechal A,
Blecharz-Klin K, et al. Influence of
long-term administration of rutin on
spatial memory as well as the
concentration of brain neurotransmitters
in aged rats. Pharmacol Rep
2012;64:808-16
134. Javed H, Khan MM, Ahmad A, et al.
Rutin prevents cognitive impairments by
ameliorating oxidative stress and
neuroinflammation in rat model of
sporadic dementia of Alzheimer type.
Neuroscience 2012;210:340-52
135. Pu F, Mishima K, Irie K, et al.
Neuroprotective effects of quercetin and
rutin on spatial memory impairment in
an 8-arm radial maze task and neuronal
death induced by repeated cerebral
ischemia in rats. J Pharmacol Sci
2007;104:329-34
136. Silva AR, Pinheiro AM, Souza CS, et al.
The flavonoid rutin induces astrocyte
and microglia activation and regulates
TNF-alpha and NO release in primary
glial cell cultures. Cell Biol Toxicol
2008;24:75-86
137. Boyle SP, Dobson VL, Duthie SJ, et al.
Bioavailability and efficiency of rutin as
an antioxidant: a human
supplementation study. Eur J Clin Nutr
2000;54:774-82
138. Zygmunt K, Faubert B, MacNeil J, et al.
Naringenin, a citrus flavonoid, increases
muscle cell glucose uptake via AMPK.
Biochem Biophys Res Commun
2010;398:178-83
139. Maes M. The cytokine hypothesis of
depression: inflammation, oxidative &
nitrosative stress (IO&NS) and leaky gut
as new targets for adjunctive treatments
in depression. Neuro Endocrinol Lett
2008;29:287-91
140. Amaro MI, Rocha J, Vila-Real H, et al.
Anti-inflammatory activity of naringin
and the biosynthesised naringenin by
naringinase immobilized in
microstructured materials in a model of
DSS-induced colitis in mice.
Food Res Int 2009;42:1010-17
141. Raza SS, Khan MM, Ahmad A, et al.
Neuroprotective effect of naringenin is
mediated through suppression of NF-kBsignaling pathway in experimental stroke.
Neuroscience
2012;published online 26 October 2012;
doi: 10.1016/j.neuroscience.2012.10.041
142. Bugianesi R, Catasta G, Spigno P, et al.
Naringenin from cooked tomato paste is
bioavailable in men. J Nutr
2002;132:3349-52
143. Olsen HT, Stafford GI, van Staden J,
et al. Isolation of the MAO-inhibitor
naringenin from Mentha aquatica L.
J Ethnopharmacol 2008;117:500-2
144. Yi LT, Li CF, Zhan X, et al.
Involvement of monoaminergic system in
the antidepressant-like effect of the
flavonoid naringenin in mice.
Prog Neuropsychopharmacol
Biol Psychiatry 2010;34:1223-8
145. Pataki T, Bak I, Kovacs P, et al. Grape
seed proanthocyanidins improved cardiac
recovery during reperfusion after
ischemia in isolated rat hearts. Am J
Clin Nutr 2002;75:894-9
146. Yamakoshi J, Sano A, Tokutake S, et al.
Oral intake of proanthocyanidin-rich
extract from grape seeds improves
chloasma. Phytother Res 2004;18:895-9
147. Preuss HG, Wallerstedt D, Talpur N,
et al. Effects of niacin-bound chromium
and grape seed proanthocyanidin extract
on the lipid profile of
hypercholesterolemic subjects: a pilot
study. J Med 2000;31:227-46
148. Uchida S, Hirai K, Hatanaka J, et al.
Antinociceptive effects of St. John’s wort,
Harpagophytum procumbens extract and
Grape seed proanthocyanidins extract in
mice. Biol Pharm Bull 2008;31:240-5
149. Sato M, Bagchi D, Tosaki A, et al.
Grape seed proanthocyanidin reduces
cardiomyocyte apoptosis by inhibiting
ischemia/reperfusion-induced activation
of JNK-1 and C-JUN. Free Radic
Biol Med 2001;31:729-37
150. Lu M, Xu L, Li B, et al. Protective
effects of grape seed proanthocyanidin
extracts on cerebral cortex of
streptozotocin-induced diabetic rats
through modulating AGEs/RAGE/NF-
kappaB pathway. J Nutr Sci Vitaminol
2010;56:87-97
151. Wang J, Ferruzzi MG, Ho L, et al.
Brain-targeted proanthocyanidin
metabolites for Alzheimer’s disease
treatment. J Neurosci 2012;32:5144-50
152. Ahn SH, Kim HJ, Jeong I, et al. Grape
seed proanthocyanidin extract inhibits
glutamate-induced cell death through
inhibition of calcium signals and nitric
oxide formation in cultured rat
hippocampal neurons. BMC Neurosci
2011;published online 3 August 2011;
doi: 1186/1471-2202-12-78
153. Mazzio EA, Harris N, Soliman KF. Food
constituents attenuate oxidase activity
and peroxide levels in C6 cells.
Planta Med 1998;64:603-7
154. Xu Y, Li S, Chen R, et al.
Antidepressant-like effect of low
molecular proanthocyanidin in mice:
involvement of monoaminergic system.
Pharmacol Biochem Behav
2010;94:447-53
155. Onoue S, Uchida A, Takahashi H, et al.
Development of high-energy amorphous
solid dispersion of nanosized nobiletin, a
citrus polymethoxylated flavone, with
improved oral bioavailability.
J Pharm Sci 2011;100:3793-801
156. Nakajima A, Yamakuni T, Haraguchi M,
et al. Nobiletin, a citrus flavonoid that
improves memory impairment, rescues
bulbectomy-induced cholinergic
neurodegeneration in mice.
J Pharmacol Sci 2007;105:122-6
157. Miyata Y, Oshitari T, Okuyama Y, et al.
Polymethoxyflavones as agents that
prevent formation of cataract: nobiletin
congeners show potent growth inhibitory
effects in human lens epithelial cells.
Bioorg Med Chem Lett 2013;23:183-7
158. Lee YS, Cha BY, Choi SS, et al.
Nobiletin improves obesity and insulin
resistance in high-fat diet-induced obese
mice. J Nutr Biochem 2013;24:156-62
159. Baek SH, Kim SM, Nam D, et al.
Antimetastatic effect of nobiletin through
the down-regulation of CXC chemokine
receptor type 4 and matrix
metallopeptidase-9. Pharm Biol
2012;50:1210-18
160. Cui Y, Wu J, Jung SC, et al.
Anti-neuroinflammatory activity of
nobiletin on suppression of microglial
activation. Biol Pharm Bull
2010;33:1814-21
161. Lee YS, Cha BY, Saito K, et al.
Nobiletin improves hyperglycemia and
insulin resistance in obese diabetic ob/ob
mice. Biochem Pharmacol
2010;79:1674-83
162. Yamamoto Y, Shioda N, Han F, et al.
Nobiletin improves brain
ischemia-induced learning and memory
deficits through stimulation of CaMKII
Natural polyphenols in the management of major depression
Expert Opin. Investig. Drugs [Early Online] 17
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.

and CREB phosphorylation. Brain Res
2009;1295:218-29
163. Matsuzaki K, Miyazaki K, Sakai S, et al.
Nobiletin, a citrus flavonoid with
neurotrophic action, augments protein
kinase A-mediated phosphorylation of
the AMPA receptor subunit, GluR1, and
the postsynaptic receptor response to
glutamate in murine hippocampus.
Eur J Pharmacol 2008;578:194-200
164. Yi LT, Xu HL, Feng J, et al.
Involvement of monoaminergic systems
in the antidepressant-like effect of
nobiletin. Physiol Behav 2011;102:1-6
165. Nagase H, Omae N, Omori A, et al.
Nobiletin and its related flavonoids with
CRE-dependent transcription-stimulating
and neuritogenic activities.
Biochem Biophys Res Commun
2005;337:1330-6
166. Sebastia N, Soriano JM, Barquinero JF,
et al. In vitro cytogenetic and genotoxic
effects of curcumin on human peripheral
blood lymphocytes. Food Chem Toxicol
2012;50:3229-33
167. Du WY, Chang C, Zhang Y, et al.
High-dose chlorogenic acid induces
inflammation reactions and oxidative
stress injury in rats without implication
of mast cell degranulation.
J Ethnopharmacol
2013;published online March 7 2013;
doi: 10.1016/j.jep.2013.01.042
168. Cardoso CR, de Syllos Colus IM,
Bernardi CC, et al. Mutagenic activity
promoted by amentoflavone and
methanolic extract of Byrsonima crassa
Niedenzu. Toxicology 2006;225:55-63
169. Hodek P, Hanustiak P, Krızkova J, et al.
Toxicological aspects of flavonoid
interaction with biomacromolecules.
Neuro Endocrinol Lett
2006;27(Suppl 2):14-17
170. Detampel P, Beck M, Krahenbuhl S,
et al. Drug interaction potential of
resveratrol. Drug Metab Rev
2012;44:253-65
171. Yun S, Zhang T, Li M, et al.
Proanthocyanidins inhibit iron
absorption from soybean (Glycine max)
seed ferritin in rats with iron deficiency
anemia. Plant Foods Hum Nutr
2011;66:212-17
172. Jazvinscak Jembrek M,
Cipak Gasparovic A, Vukovic L, et al.
Quercetin supplementation: insight into
the potentially harmful outcomes of
neurodegenerative prevention.
Naunyn Schmiedebergs Arch Pharmacol
2012;385:1185-97
AffiliationLokesh Pathak1, Yadvendra Agrawal1 &
Ashish Dhir†1,2 MPharm PhD†Author for correspondence1Scientist D,
Gujarat Forensic Sciences University,
Institute of Research & Development,
DFS Headquarters,
Sector 18-A, Gandhinagar,
Gujarat-382007, India
Tel: +91 79 65735505 06;
E-mail: [email protected] of Biotechnology,
New Delhi, India
L. Pathak et al.
18 Expert Opin. Investig. Drugs [Early Online]
Exp
ert O
pin.
Inv
estig
. Dru
gs D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Lim
eric
k on
05/
24/1
3Fo
r pe
rson
al u
se o
nly.







