

Zooplankton inshore offshore Mexican Central Pacific 67
Lat Am J Aquat Res 45(1) 67-78 2017
DOI 103856vol45-issue1-fulltext-7
Research Article
Physical chemical and zooplankton biomass variability (inshore-offshore) of
Mexican Central Pacific during El Nintildeo-La Nintildea 2010
Gloria Pelayo-Martiacutenez12
Aramis Olivos-Ortiz2 Carmen Franco-Gordo
1
Sonia Quijano-Scheggia2 Juan Gavintildeo-Rodriacuteguez
2
Tadashi Kono-Martiacutenez3 amp
Faacutetima Castro-Ochoa
3
1Centro de Ecologiacutea Costera Universidad de Guadalajara Jalisco Meacutexico 2Centro Universitario de Investigaciones Oceanoloacutegicas Colima Meacutexico
3Facultad de Ciencias Marinas Universidad Autoacutenoma de Baja California Ensenada Baja California Meacutexico Corresponding autor Aramis Olivos-Ortiz (aolivosucolmx)
ABSTRACT The response of zooplankton phytoplankton biomass (Chl-a concentration) and nutrients to the
oceanographic variability in the Mexican Central Pacific was studied over three oceanographic surveys realized
in 2010 (January May and October) The highest zooplankton biomass (48 mL 1000 m-3) was registered in May
when nitrites nitrates phosphates and silicates were also highest and mean Chl-a was 067 mg m-3 The lowest
zooplankton biomass was registered in October with a mean value of 25 mL 1000 m-3 Twenty-nine taxonomic
groups of zooplankton were recorded of which copepods contributed 71 of total abundance followed by
chaetognaths (6) and euphausiids (5) Multivariate BEST test demonstrated that mixed layer depth and
temperature at 25 m depth can explain the distribution of major zooplankton groups The study area was
influenced by El Nintildeo conditions which caused a seasonal shift Coastal upwelling was evident until late
summer when geostrophic analysis showed a predominant SW component near the Jalisco coast We suggest a
possible top-down control of zooplankton grazing over phytoplankton and acclimatization of both plankton
groups to variability in environmental conditions induced by the transition from warm to cold ENSO phase
Keywords zooplankton Chl-a nutrients mesoscale El Nintildeo Mexican Central Pacific
INTRODUCTION
Mesoscale features in the ocean constitute distribution
mechanisms for inorganic nutrients and planktonic
organisms Marine zooplankton is an important link
between primary producers and higher trophic levels
its spatial and temporal variability responds to the
influence of physical features such as currents wind
oceanic fronts gyres upwelling and downwelling
(Riandey et al 2005 Yebra et al 2009) On a seasonal
scale zooplankton abundance responds to nutrient
availability and phytoplankton abundance (as Chl-a
concentrations) with a time lag of up to two months in
temperate latitudes (Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda 2006) However in tropical regions like the
Mexican Central Pacific (MCP) where seasonal
changes are not easily detected the variability of
zooplankton abundance is usually governed by the in-
_____________________
Corresponding editor Sergio Palma
fluence of oceanographic features (upwelling gyres)
the effect of the dominant wind in the region or inter-
annual features such as El Nintildeo (Loacutepez-Sandoval et al
2009) In the last few years the number of zooplankton
biomass studies in the MCP has increased Franco-
Gordo et al (2004) described the MCP as a region with
a negative correlation of zooplankton biomass and
temperature based on high values of biomass
associated with the presence of low temperature water
Low abundances were observed during the El Nintildeo
period coinciding with studies on specific taxonomic
groups including copepods euphausiids and amphi-
pods (Ambriz-Arreola et al 2012 Gasca et al 2012
Kozak et al 2014)
Modeled dynamics in the study area show the
influence of the Mexican Coastal Current which is
generated at the Gulf of Tehuantepec as a sub-
thermocline flow that reaches the surface between 17ordm
68 Latin American Journal of Aquatic Research
and 20ordmN to continue northward up to the Gulf of
California The confluence of this current with a branch
of California Current converts the MCP into a transition
zone where mesoscale eddies meanders and filaments
are generated (Zamudio et al 2007 Godiacutenez et al
2010 Pantoja et al 2012 Goacutemez-Valdivia et al
2015) In addition the presence of a thermocline dome
(106ordmW 18ordmN) that plays an important role on the
coastal seasonal dynamics off SW Mexico has recently
been reported by Goacutemez-Valdivia et al (2015) who
proposed calling it ldquoThe Thermocline Cabo Corrientes
Domerdquo Based on this information the objective of this
study is to provide an integrated perspective of the
Mexican Central Pacific that includes the response of
zooplankton phytoplankton biomass (Chl-a) and
nutrients to the spatial-temporal hydrological varia-
bility in the region
MATERIALS AND METHODS
Field methods
Zooplankton and hydrographic data were obtained
along a 100 nm fringe from Cabo Corrientes Jalisco to Maruata Michoacaacuten on board the ldquoBIP XIIrdquo vessel
during 15-27 January (PCM1001) 25 May-4 June
(PCM1005) and 18-29 October (PCM1010) of 2010 The oceanographic surveys covered a polygonal area
with 19 stations (Fig 1) Zooplankton was sampled day or night with oblique tows using a standard Bongo net
fitted with 505 microm mesh and 06 m mouth diameter A calibrated digital flow meter was attached to the mouth
of the net All hauls were made at 200 m or from the
bottom to the surface (Smith amp Richardson 1977) Samples were preserved in a 4 formalin solution
buffered with sodium borate (Griffiths et al 1976) Nutrient and Chl-a samples were taken with a 5 L
Niskin bottle at 0 10 25 50 75 100 150 175 and 200
m depths and frozen after collection for subsequent laboratory analysis Vertical temperature and salinity
profiles of the water column were taken at each station with a Seabird model SBE-19 plus
Laboratory analysis
A total of 57 samples were analyzed (Fig 1) Zooplank-
ton biomass was calculated using the displaced volume method (Beers 1976) excluding organisms larger than
3 cm High density samples were fractioned using a Folsom plankton splitter from frac12 up to frac14 depending on
the density of each sample Biomass values were standardized to mL 1000 m-3 of filtered seawater
Organisms in the samples were identified to dominant taxonomic group using key sheets of Newell amp Newell (1973) Smith amp Johnson (1996) and Todd et al (1996)
The number of organisms in each functional group was standardized to 1000 m-3 of seawater (ind 1000 m-3)
Density profiles were obtained from temperature and salinity in order to calculate the mixed layer depth (MLD) according to Kara et al (2000) Geostrophic velocity was calculated from dynamic height profiles as proposed by Talley et al (2011) both parameters were processed using MATLAB 810604 software (2009) Nutrients (NO2 NO3 NH4 PO4 and Si (OH)4) were determined in a Skalar San Plus II segmented flow auto analyzer and Chl-a using colorimetric techniques with a PerkinElmeracutes UVVis spectrophotometer following Strickland amp Parsons (1972) and Grasshoff et al (1983)
Data analysis
Biotic and abiotic parameters were classified according to the water column structure during stratified (El Nintildeo) and semi-stratified (La Nintildea) conditions Statistical analysis did not indicate normal distribution (Kolmorogov-Smirnov and Shapiro-Wilk) so non-parametrical analyses were applied Mean biomass values in every cruise were calculated with a 95 confidence interval Two matrices were generated environmental data were standardized (temperature salinity and Chl-a at 25 m depth MLD coastal upwelling index (CUI) nutrients and zooplankton biomass) and zooplankton abundance data were transformed with x = log (x+1) to reduce the difference of scale between units and results Differences between groups were tested with ANOVA analysis in the case of significant results post-hoc analysis defined the differences For multidimensional analysis (MDS) and dissimilarity test Euclidean distance index was applied to environmental and Bray-Curtis to abundance data Differences between cruises and distance to coast were evaluated with one-way analysis of similarities (ANOSIM) To identify the variables that possibly controlled the distribution of major zooplankton groups a biotic and environment matching test (BEST) was performed In addition Spearman-rank correlation test was made between zooplankton biomass functional groups Chl-a nutrient and abiotic variables Functional groups with less than of 1 of the total abundance were excluded from statistical analysis Multivariate analyses were performed using PRIMER 60 software and non-parametric correlation was performed with STATISTICA 7 software
RESULTS
Hydrological conditions
Sea surface temperature (SST) showed a positive gradient from north to south During January a range
Zooplankton inshore offshore Mexican Central Pacific 69
Figure 1 Position of sampling sites off the Mexican Central Pacific
of 255-282degC was registered May showed the lowest values of the surveyed period (245-285degC) while in October the temperature varied from 245 to 296degC (Figs 2a-2c) Although variation between cruises appeared to be small temperature at 25 m depth was significantly lower in May than during January and October (F = 28 P lt 0001) A pulse of cold water was identified in front of the Jalisco coast during the surveyed period with the highest intensity in May when surface temperatures of 167ordmC were detected This variation of temperature values across the continental shelf also showed significant differences between transects (F = 4 P = 0024)
Salinity was characterized by minimum values in
the southern region especially along the coastal and
intermediate transects covering the central and southern
regions in January During January values ranged from
338 to 346 while May and October registered
maximum salinity values (340-348) for the entire
study area The thermocline was found around 80-100
m in January reaching the shallowest depth in October
(40-70 m) Geostrophic circulation indicated the presen-
ce of a cyclonic gyre during January and a stronger one
in May covering the central zone of the study area (Fig
2) During October there was no apparent oceano-graphic feature that caused the thermocline to rise in
fact the presence of an anticyclonic gyre was detected
in the center of the intermediate transect (Figs 2c and
2f) The mixed layer depth also showed significant
changes between cruises (F = 3139 P = 005) in
January it was deepest (26-63 m) getting shallower
during May and October with values ranging from 19
to 47 m
Based on the ANOSIM test environmental varia-
bles showed significant differences between climatic
periods with an R = 0579 (P lt 001) presenting the
most important differences during El Nintildeo (January)
Distance from the coast showed no statistically significant influence (R = 0071 P = 014)
Nutrients and Chl-a
With the exception of ammonium inorganic nutrients
showed a similar distribution pattern highest values
near upwelling zones and cyclonic gyres Median
values of nitrate+nitrite (1467 microM) phosphate (181
microM) and silicic acid (2421 microM) concentrations were
highest in May when ammonium was at its lowest
values In January ammonium (324 microM) showed
maximum median concentration and nitrate+nitrite
(220 microM) phosphate (074 microM) and silicic acid (709
microM) concentrations were at a minimum (Table 1)
Chl-a concentrations ranged from 001 in June to 1291 mg m-3 in January Median concentrations were
January 218 mg m-3 June 067 mg m-3 and October 088 mg m-3 (Fig 3)
70 Latin American Journal of Aquatic Research
Figure 2 Spatial distribution of a b c) temperature and d f g) salinity in January May and October 2010 along the Central Mexican Pacific Vectors show geostrophic currents
Values were statistically highest in the coastal zone (F = 357 P = 003) of the northern and southern region Temporal distribution did not show significant diffe-rences (F = 060 P = 055)
Zooplankton biomass and abundance
Mean zooplankton biomass varied from 25 mL 1000 m-3
in October to 478 mL 1000 m-3 in May (Fig 4a) Mean
biomass in May was significantly higher than January
and October (F = 1106 P lt 005) Distribution showed
a tendency to decrease offshore but it was not
statistically significant (F = 181 P = 0170) because of
the presence of some high values along the oceanic
transect In general the highest values of biomass (56-
95 mL 1000 m-3) over the entire study period were
observed in front of northern Michoacaacuten on the coastal
transect During January coastal station E1 also
showed major biomass with a value of 3022 mL 1000
m-3 Zooplankton biomass in May was high in E6
(6649 mL 1000 m-3) In October zooplankton biomass
showed a decrease in all areas especially in the
northern region
The zooplankton community was composed of 29
functional groups which varied temporally Eight
groups accounted for 95 of total abundance both in
January (copepods chaetognaths euphausiids amphi-
pods polychaetes pteropods fish larvae and decapods)
and May (copepods amphipods chaetognaths
euphausiids decapods pteropods fish larvae and
gasteropods) By October ten taxonomic groups
contributed 95 of the zooplankton community
copepods chaetognaths euphausiids fish larvae
amphipods siphonophores cladocerans polychaetes medusas and ostracods (Fig 4)
The remaining 5 of the zooplankton community
was composed of cladocerans ostracods doliolids
polychaete larvae medusae fish eggs ctenophores salps cephalopod paralarvae bivalves isopods appen-
dicularians pyrosomes mysiids heteropods and echinoderms larvae stomatopods and cirripeds
Zooplankton inshore offshore Mexican Central Pacific 71
Distribution of major groups
Copepods chaetognaths euphausiids and amphipods
were the most abundant groups during the study period accounting for 87 of the total abundance (Fig 5b)
Maximum median value of copepod abundance of
10939 ind 1000 m-3 was recorded in May while the
minimum of 9297 ind 1000 m-3 was recorded in
October The January cruise registered a density of
9900 ind 1000 m-3 In October 20 of the stations located in the center of the study area were characterized
Table 1 Mean concentration and standard error of
nutrients in the Mexican Central Pacific during January
May-June and October 2010
Jan May-Jun Oct
NO3+NO2 220 plusmn 044 1467 plusmn 113 1102 plusmn 162
NH4 324 plusmn 018 227 plusmn 017 225 plusmn 014
PO4 072 plusmn 014 181 plusmn 027 117 plusmn 015
Si (OH)4 709 plusmn 091 2421 plusmn 229 929 plusmn 152
by the lowest values (0-5000 ind 1000 m-3) The largest
aggregation of this group during the entire surveyed
period was observed at the inshore southern region
particularly in station E14 where abundances up to 48711 ind 1000 m-3 were found (Fig 5)
The average abundance of chaetognaths over the
entire study period was 823 ind 1000 m-3 High
abundances of this group were observed in the southern
study area except in January when the northern zone
registered the highest abundance of 1150 ind 1000 m-3
During October cruise values inshore of the central and
northern zone were lower than the oceanic region (Fig 5)
Median abundance of euphausiids varied from 595
ind 1000 m-3 in January to 860 ind 1000 m3 in May
36 of stations during January showed values in the
range of 200-400 ind 1000 m-3 Spatial distribution
during January and May showed high abundance in the
southern inshore region and low in the northern while
in October high abundance was observed in the central-
oceanic zone (Fig 5) Except for October amphipod
abundance was high in the southern inshore zone in
May it was 14599 ind 1000 m-3 at station E17 but
typical values during this cruise were between 600 and
800 ind 1000 m-3 Cross shelf distribution was
characterized by high values near the coast that
decreased in the oceanic region except for oceanic
station E3 that registered high values with respect to
those near the coast during January and October (Fig
5) Average abundance of amphipods was 845 ind 1000
m-3 in October
No significant differences between samples from
different coastal or oceanic transects were found in
ANOSIM analyses of biotic variables (R = 0078 P =
008) while a marginally significant seasonality was
evident between each oceanographic cruise with an R =
0468 (P lt 001)
Zooplankton vs abiotic features
Biota and environment matching test (BEST) showed
minor influence of environmental variables on taxo-
nomic zooplankton groups Temperature and MLD
were the main variables explaining zooplankton group
distribution in the study area (Rho = 038 P = 001)
Spearman correlation analysis also showed low rank
values between abiotic variables and abundance of
zooplankton groups and biomass (Table 2)
Chl-a had no statistical correlation with zooplank-
ton biomass and was negatively correlated with
chaetognaths ctenophores and doliolids (-027 lt r lt
041) while bivalves and decapod larvae showed a
positive correlation (r = 029 030)
There was no significant correlation between
temperature and zooplankton biomass However
salinity and zooplankton biomass were positively
correlated (r = 027) Most of the major zooplankton
groups did not show any significant correlation with
temperature or salinity Minor groups like bivalves and
pyrosomes were positively related with temperature
and negatively with salinity Pteropods and gasteropods
showed positive correlation with temperature (r = 028
044) Ostracods and doliolids showed negative
correlation with salinity (r = -026 -031) while
siphonophores had a positive correlation (r = 028 P
lt005) Negative correlations were found between
temperature with fish larvae and medusae (Table 1)
The MLD was not significantly correlated with
zooplankton biomass but was positively related with 9
zooplankton groups including copepods amphipods
and chaetognaths (027 lt r lt 060) Other less abundant
groups (lt1) such as siphonophores medusae
cladocerans heteropods appendicularians and echino-
derm larvae showed a negative correlation (-064 lt r lt
-028 P lt 005)
Zooplankton biomass and doliolids were positively
correlated with nitrates+nitrites phosphates and sili-
cates however none of most abundant groups
(copepods chaetognaths amphipods euphausiids)
were significantly correlated with nutrients In spite of
this maps of spatial distribution showed concordance
of high abundance values of copepods with nitrate+ nitrite during January in the central zone of the oceanic
transect and during October in the southeast region
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

68 Latin American Journal of Aquatic Research
and 20ordmN to continue northward up to the Gulf of
California The confluence of this current with a branch
of California Current converts the MCP into a transition
zone where mesoscale eddies meanders and filaments
are generated (Zamudio et al 2007 Godiacutenez et al
2010 Pantoja et al 2012 Goacutemez-Valdivia et al
2015) In addition the presence of a thermocline dome
(106ordmW 18ordmN) that plays an important role on the
coastal seasonal dynamics off SW Mexico has recently
been reported by Goacutemez-Valdivia et al (2015) who
proposed calling it ldquoThe Thermocline Cabo Corrientes
Domerdquo Based on this information the objective of this
study is to provide an integrated perspective of the
Mexican Central Pacific that includes the response of
zooplankton phytoplankton biomass (Chl-a) and
nutrients to the spatial-temporal hydrological varia-
bility in the region
MATERIALS AND METHODS
Field methods
Zooplankton and hydrographic data were obtained
along a 100 nm fringe from Cabo Corrientes Jalisco to Maruata Michoacaacuten on board the ldquoBIP XIIrdquo vessel
during 15-27 January (PCM1001) 25 May-4 June
(PCM1005) and 18-29 October (PCM1010) of 2010 The oceanographic surveys covered a polygonal area
with 19 stations (Fig 1) Zooplankton was sampled day or night with oblique tows using a standard Bongo net
fitted with 505 microm mesh and 06 m mouth diameter A calibrated digital flow meter was attached to the mouth
of the net All hauls were made at 200 m or from the
bottom to the surface (Smith amp Richardson 1977) Samples were preserved in a 4 formalin solution
buffered with sodium borate (Griffiths et al 1976) Nutrient and Chl-a samples were taken with a 5 L
Niskin bottle at 0 10 25 50 75 100 150 175 and 200
m depths and frozen after collection for subsequent laboratory analysis Vertical temperature and salinity
profiles of the water column were taken at each station with a Seabird model SBE-19 plus
Laboratory analysis
A total of 57 samples were analyzed (Fig 1) Zooplank-
ton biomass was calculated using the displaced volume method (Beers 1976) excluding organisms larger than
3 cm High density samples were fractioned using a Folsom plankton splitter from frac12 up to frac14 depending on
the density of each sample Biomass values were standardized to mL 1000 m-3 of filtered seawater
Organisms in the samples were identified to dominant taxonomic group using key sheets of Newell amp Newell (1973) Smith amp Johnson (1996) and Todd et al (1996)
The number of organisms in each functional group was standardized to 1000 m-3 of seawater (ind 1000 m-3)
Density profiles were obtained from temperature and salinity in order to calculate the mixed layer depth (MLD) according to Kara et al (2000) Geostrophic velocity was calculated from dynamic height profiles as proposed by Talley et al (2011) both parameters were processed using MATLAB 810604 software (2009) Nutrients (NO2 NO3 NH4 PO4 and Si (OH)4) were determined in a Skalar San Plus II segmented flow auto analyzer and Chl-a using colorimetric techniques with a PerkinElmeracutes UVVis spectrophotometer following Strickland amp Parsons (1972) and Grasshoff et al (1983)
Data analysis
Biotic and abiotic parameters were classified according to the water column structure during stratified (El Nintildeo) and semi-stratified (La Nintildea) conditions Statistical analysis did not indicate normal distribution (Kolmorogov-Smirnov and Shapiro-Wilk) so non-parametrical analyses were applied Mean biomass values in every cruise were calculated with a 95 confidence interval Two matrices were generated environmental data were standardized (temperature salinity and Chl-a at 25 m depth MLD coastal upwelling index (CUI) nutrients and zooplankton biomass) and zooplankton abundance data were transformed with x = log (x+1) to reduce the difference of scale between units and results Differences between groups were tested with ANOVA analysis in the case of significant results post-hoc analysis defined the differences For multidimensional analysis (MDS) and dissimilarity test Euclidean distance index was applied to environmental and Bray-Curtis to abundance data Differences between cruises and distance to coast were evaluated with one-way analysis of similarities (ANOSIM) To identify the variables that possibly controlled the distribution of major zooplankton groups a biotic and environment matching test (BEST) was performed In addition Spearman-rank correlation test was made between zooplankton biomass functional groups Chl-a nutrient and abiotic variables Functional groups with less than of 1 of the total abundance were excluded from statistical analysis Multivariate analyses were performed using PRIMER 60 software and non-parametric correlation was performed with STATISTICA 7 software
RESULTS
Hydrological conditions
Sea surface temperature (SST) showed a positive gradient from north to south During January a range
Zooplankton inshore offshore Mexican Central Pacific 69
Figure 1 Position of sampling sites off the Mexican Central Pacific
of 255-282degC was registered May showed the lowest values of the surveyed period (245-285degC) while in October the temperature varied from 245 to 296degC (Figs 2a-2c) Although variation between cruises appeared to be small temperature at 25 m depth was significantly lower in May than during January and October (F = 28 P lt 0001) A pulse of cold water was identified in front of the Jalisco coast during the surveyed period with the highest intensity in May when surface temperatures of 167ordmC were detected This variation of temperature values across the continental shelf also showed significant differences between transects (F = 4 P = 0024)
Salinity was characterized by minimum values in
the southern region especially along the coastal and
intermediate transects covering the central and southern
regions in January During January values ranged from
338 to 346 while May and October registered
maximum salinity values (340-348) for the entire
study area The thermocline was found around 80-100
m in January reaching the shallowest depth in October
(40-70 m) Geostrophic circulation indicated the presen-
ce of a cyclonic gyre during January and a stronger one
in May covering the central zone of the study area (Fig
2) During October there was no apparent oceano-graphic feature that caused the thermocline to rise in
fact the presence of an anticyclonic gyre was detected
in the center of the intermediate transect (Figs 2c and
2f) The mixed layer depth also showed significant
changes between cruises (F = 3139 P = 005) in
January it was deepest (26-63 m) getting shallower
during May and October with values ranging from 19
to 47 m
Based on the ANOSIM test environmental varia-
bles showed significant differences between climatic
periods with an R = 0579 (P lt 001) presenting the
most important differences during El Nintildeo (January)
Distance from the coast showed no statistically significant influence (R = 0071 P = 014)
Nutrients and Chl-a
With the exception of ammonium inorganic nutrients
showed a similar distribution pattern highest values
near upwelling zones and cyclonic gyres Median
values of nitrate+nitrite (1467 microM) phosphate (181
microM) and silicic acid (2421 microM) concentrations were
highest in May when ammonium was at its lowest
values In January ammonium (324 microM) showed
maximum median concentration and nitrate+nitrite
(220 microM) phosphate (074 microM) and silicic acid (709
microM) concentrations were at a minimum (Table 1)
Chl-a concentrations ranged from 001 in June to 1291 mg m-3 in January Median concentrations were
January 218 mg m-3 June 067 mg m-3 and October 088 mg m-3 (Fig 3)
70 Latin American Journal of Aquatic Research
Figure 2 Spatial distribution of a b c) temperature and d f g) salinity in January May and October 2010 along the Central Mexican Pacific Vectors show geostrophic currents
Values were statistically highest in the coastal zone (F = 357 P = 003) of the northern and southern region Temporal distribution did not show significant diffe-rences (F = 060 P = 055)
Zooplankton biomass and abundance
Mean zooplankton biomass varied from 25 mL 1000 m-3
in October to 478 mL 1000 m-3 in May (Fig 4a) Mean
biomass in May was significantly higher than January
and October (F = 1106 P lt 005) Distribution showed
a tendency to decrease offshore but it was not
statistically significant (F = 181 P = 0170) because of
the presence of some high values along the oceanic
transect In general the highest values of biomass (56-
95 mL 1000 m-3) over the entire study period were
observed in front of northern Michoacaacuten on the coastal
transect During January coastal station E1 also
showed major biomass with a value of 3022 mL 1000
m-3 Zooplankton biomass in May was high in E6
(6649 mL 1000 m-3) In October zooplankton biomass
showed a decrease in all areas especially in the
northern region
The zooplankton community was composed of 29
functional groups which varied temporally Eight
groups accounted for 95 of total abundance both in
January (copepods chaetognaths euphausiids amphi-
pods polychaetes pteropods fish larvae and decapods)
and May (copepods amphipods chaetognaths
euphausiids decapods pteropods fish larvae and
gasteropods) By October ten taxonomic groups
contributed 95 of the zooplankton community
copepods chaetognaths euphausiids fish larvae
amphipods siphonophores cladocerans polychaetes medusas and ostracods (Fig 4)
The remaining 5 of the zooplankton community
was composed of cladocerans ostracods doliolids
polychaete larvae medusae fish eggs ctenophores salps cephalopod paralarvae bivalves isopods appen-
dicularians pyrosomes mysiids heteropods and echinoderms larvae stomatopods and cirripeds
Zooplankton inshore offshore Mexican Central Pacific 71
Distribution of major groups
Copepods chaetognaths euphausiids and amphipods
were the most abundant groups during the study period accounting for 87 of the total abundance (Fig 5b)
Maximum median value of copepod abundance of
10939 ind 1000 m-3 was recorded in May while the
minimum of 9297 ind 1000 m-3 was recorded in
October The January cruise registered a density of
9900 ind 1000 m-3 In October 20 of the stations located in the center of the study area were characterized
Table 1 Mean concentration and standard error of
nutrients in the Mexican Central Pacific during January
May-June and October 2010
Jan May-Jun Oct
NO3+NO2 220 plusmn 044 1467 plusmn 113 1102 plusmn 162
NH4 324 plusmn 018 227 plusmn 017 225 plusmn 014
PO4 072 plusmn 014 181 plusmn 027 117 plusmn 015
Si (OH)4 709 plusmn 091 2421 plusmn 229 929 plusmn 152
by the lowest values (0-5000 ind 1000 m-3) The largest
aggregation of this group during the entire surveyed
period was observed at the inshore southern region
particularly in station E14 where abundances up to 48711 ind 1000 m-3 were found (Fig 5)
The average abundance of chaetognaths over the
entire study period was 823 ind 1000 m-3 High
abundances of this group were observed in the southern
study area except in January when the northern zone
registered the highest abundance of 1150 ind 1000 m-3
During October cruise values inshore of the central and
northern zone were lower than the oceanic region (Fig 5)
Median abundance of euphausiids varied from 595
ind 1000 m-3 in January to 860 ind 1000 m3 in May
36 of stations during January showed values in the
range of 200-400 ind 1000 m-3 Spatial distribution
during January and May showed high abundance in the
southern inshore region and low in the northern while
in October high abundance was observed in the central-
oceanic zone (Fig 5) Except for October amphipod
abundance was high in the southern inshore zone in
May it was 14599 ind 1000 m-3 at station E17 but
typical values during this cruise were between 600 and
800 ind 1000 m-3 Cross shelf distribution was
characterized by high values near the coast that
decreased in the oceanic region except for oceanic
station E3 that registered high values with respect to
those near the coast during January and October (Fig
5) Average abundance of amphipods was 845 ind 1000
m-3 in October
No significant differences between samples from
different coastal or oceanic transects were found in
ANOSIM analyses of biotic variables (R = 0078 P =
008) while a marginally significant seasonality was
evident between each oceanographic cruise with an R =
0468 (P lt 001)
Zooplankton vs abiotic features
Biota and environment matching test (BEST) showed
minor influence of environmental variables on taxo-
nomic zooplankton groups Temperature and MLD
were the main variables explaining zooplankton group
distribution in the study area (Rho = 038 P = 001)
Spearman correlation analysis also showed low rank
values between abiotic variables and abundance of
zooplankton groups and biomass (Table 2)
Chl-a had no statistical correlation with zooplank-
ton biomass and was negatively correlated with
chaetognaths ctenophores and doliolids (-027 lt r lt
041) while bivalves and decapod larvae showed a
positive correlation (r = 029 030)
There was no significant correlation between
temperature and zooplankton biomass However
salinity and zooplankton biomass were positively
correlated (r = 027) Most of the major zooplankton
groups did not show any significant correlation with
temperature or salinity Minor groups like bivalves and
pyrosomes were positively related with temperature
and negatively with salinity Pteropods and gasteropods
showed positive correlation with temperature (r = 028
044) Ostracods and doliolids showed negative
correlation with salinity (r = -026 -031) while
siphonophores had a positive correlation (r = 028 P
lt005) Negative correlations were found between
temperature with fish larvae and medusae (Table 1)
The MLD was not significantly correlated with
zooplankton biomass but was positively related with 9
zooplankton groups including copepods amphipods
and chaetognaths (027 lt r lt 060) Other less abundant
groups (lt1) such as siphonophores medusae
cladocerans heteropods appendicularians and echino-
derm larvae showed a negative correlation (-064 lt r lt
-028 P lt 005)
Zooplankton biomass and doliolids were positively
correlated with nitrates+nitrites phosphates and sili-
cates however none of most abundant groups
(copepods chaetognaths amphipods euphausiids)
were significantly correlated with nutrients In spite of
this maps of spatial distribution showed concordance
of high abundance values of copepods with nitrate+ nitrite during January in the central zone of the oceanic
transect and during October in the southeast region
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

Zooplankton inshore offshore Mexican Central Pacific 69
Figure 1 Position of sampling sites off the Mexican Central Pacific
of 255-282degC was registered May showed the lowest values of the surveyed period (245-285degC) while in October the temperature varied from 245 to 296degC (Figs 2a-2c) Although variation between cruises appeared to be small temperature at 25 m depth was significantly lower in May than during January and October (F = 28 P lt 0001) A pulse of cold water was identified in front of the Jalisco coast during the surveyed period with the highest intensity in May when surface temperatures of 167ordmC were detected This variation of temperature values across the continental shelf also showed significant differences between transects (F = 4 P = 0024)
Salinity was characterized by minimum values in
the southern region especially along the coastal and
intermediate transects covering the central and southern
regions in January During January values ranged from
338 to 346 while May and October registered
maximum salinity values (340-348) for the entire
study area The thermocline was found around 80-100
m in January reaching the shallowest depth in October
(40-70 m) Geostrophic circulation indicated the presen-
ce of a cyclonic gyre during January and a stronger one
in May covering the central zone of the study area (Fig
2) During October there was no apparent oceano-graphic feature that caused the thermocline to rise in
fact the presence of an anticyclonic gyre was detected
in the center of the intermediate transect (Figs 2c and
2f) The mixed layer depth also showed significant
changes between cruises (F = 3139 P = 005) in
January it was deepest (26-63 m) getting shallower
during May and October with values ranging from 19
to 47 m
Based on the ANOSIM test environmental varia-
bles showed significant differences between climatic
periods with an R = 0579 (P lt 001) presenting the
most important differences during El Nintildeo (January)
Distance from the coast showed no statistically significant influence (R = 0071 P = 014)
Nutrients and Chl-a
With the exception of ammonium inorganic nutrients
showed a similar distribution pattern highest values
near upwelling zones and cyclonic gyres Median
values of nitrate+nitrite (1467 microM) phosphate (181
microM) and silicic acid (2421 microM) concentrations were
highest in May when ammonium was at its lowest
values In January ammonium (324 microM) showed
maximum median concentration and nitrate+nitrite
(220 microM) phosphate (074 microM) and silicic acid (709
microM) concentrations were at a minimum (Table 1)
Chl-a concentrations ranged from 001 in June to 1291 mg m-3 in January Median concentrations were
January 218 mg m-3 June 067 mg m-3 and October 088 mg m-3 (Fig 3)
70 Latin American Journal of Aquatic Research
Figure 2 Spatial distribution of a b c) temperature and d f g) salinity in January May and October 2010 along the Central Mexican Pacific Vectors show geostrophic currents
Values were statistically highest in the coastal zone (F = 357 P = 003) of the northern and southern region Temporal distribution did not show significant diffe-rences (F = 060 P = 055)
Zooplankton biomass and abundance
Mean zooplankton biomass varied from 25 mL 1000 m-3
in October to 478 mL 1000 m-3 in May (Fig 4a) Mean
biomass in May was significantly higher than January
and October (F = 1106 P lt 005) Distribution showed
a tendency to decrease offshore but it was not
statistically significant (F = 181 P = 0170) because of
the presence of some high values along the oceanic
transect In general the highest values of biomass (56-
95 mL 1000 m-3) over the entire study period were
observed in front of northern Michoacaacuten on the coastal
transect During January coastal station E1 also
showed major biomass with a value of 3022 mL 1000
m-3 Zooplankton biomass in May was high in E6
(6649 mL 1000 m-3) In October zooplankton biomass
showed a decrease in all areas especially in the
northern region
The zooplankton community was composed of 29
functional groups which varied temporally Eight
groups accounted for 95 of total abundance both in
January (copepods chaetognaths euphausiids amphi-
pods polychaetes pteropods fish larvae and decapods)
and May (copepods amphipods chaetognaths
euphausiids decapods pteropods fish larvae and
gasteropods) By October ten taxonomic groups
contributed 95 of the zooplankton community
copepods chaetognaths euphausiids fish larvae
amphipods siphonophores cladocerans polychaetes medusas and ostracods (Fig 4)
The remaining 5 of the zooplankton community
was composed of cladocerans ostracods doliolids
polychaete larvae medusae fish eggs ctenophores salps cephalopod paralarvae bivalves isopods appen-
dicularians pyrosomes mysiids heteropods and echinoderms larvae stomatopods and cirripeds
Zooplankton inshore offshore Mexican Central Pacific 71
Distribution of major groups
Copepods chaetognaths euphausiids and amphipods
were the most abundant groups during the study period accounting for 87 of the total abundance (Fig 5b)
Maximum median value of copepod abundance of
10939 ind 1000 m-3 was recorded in May while the
minimum of 9297 ind 1000 m-3 was recorded in
October The January cruise registered a density of
9900 ind 1000 m-3 In October 20 of the stations located in the center of the study area were characterized
Table 1 Mean concentration and standard error of
nutrients in the Mexican Central Pacific during January
May-June and October 2010
Jan May-Jun Oct
NO3+NO2 220 plusmn 044 1467 plusmn 113 1102 plusmn 162
NH4 324 plusmn 018 227 plusmn 017 225 plusmn 014
PO4 072 plusmn 014 181 plusmn 027 117 plusmn 015
Si (OH)4 709 plusmn 091 2421 plusmn 229 929 plusmn 152
by the lowest values (0-5000 ind 1000 m-3) The largest
aggregation of this group during the entire surveyed
period was observed at the inshore southern region
particularly in station E14 where abundances up to 48711 ind 1000 m-3 were found (Fig 5)
The average abundance of chaetognaths over the
entire study period was 823 ind 1000 m-3 High
abundances of this group were observed in the southern
study area except in January when the northern zone
registered the highest abundance of 1150 ind 1000 m-3
During October cruise values inshore of the central and
northern zone were lower than the oceanic region (Fig 5)
Median abundance of euphausiids varied from 595
ind 1000 m-3 in January to 860 ind 1000 m3 in May
36 of stations during January showed values in the
range of 200-400 ind 1000 m-3 Spatial distribution
during January and May showed high abundance in the
southern inshore region and low in the northern while
in October high abundance was observed in the central-
oceanic zone (Fig 5) Except for October amphipod
abundance was high in the southern inshore zone in
May it was 14599 ind 1000 m-3 at station E17 but
typical values during this cruise were between 600 and
800 ind 1000 m-3 Cross shelf distribution was
characterized by high values near the coast that
decreased in the oceanic region except for oceanic
station E3 that registered high values with respect to
those near the coast during January and October (Fig
5) Average abundance of amphipods was 845 ind 1000
m-3 in October
No significant differences between samples from
different coastal or oceanic transects were found in
ANOSIM analyses of biotic variables (R = 0078 P =
008) while a marginally significant seasonality was
evident between each oceanographic cruise with an R =
0468 (P lt 001)
Zooplankton vs abiotic features
Biota and environment matching test (BEST) showed
minor influence of environmental variables on taxo-
nomic zooplankton groups Temperature and MLD
were the main variables explaining zooplankton group
distribution in the study area (Rho = 038 P = 001)
Spearman correlation analysis also showed low rank
values between abiotic variables and abundance of
zooplankton groups and biomass (Table 2)
Chl-a had no statistical correlation with zooplank-
ton biomass and was negatively correlated with
chaetognaths ctenophores and doliolids (-027 lt r lt
041) while bivalves and decapod larvae showed a
positive correlation (r = 029 030)
There was no significant correlation between
temperature and zooplankton biomass However
salinity and zooplankton biomass were positively
correlated (r = 027) Most of the major zooplankton
groups did not show any significant correlation with
temperature or salinity Minor groups like bivalves and
pyrosomes were positively related with temperature
and negatively with salinity Pteropods and gasteropods
showed positive correlation with temperature (r = 028
044) Ostracods and doliolids showed negative
correlation with salinity (r = -026 -031) while
siphonophores had a positive correlation (r = 028 P
lt005) Negative correlations were found between
temperature with fish larvae and medusae (Table 1)
The MLD was not significantly correlated with
zooplankton biomass but was positively related with 9
zooplankton groups including copepods amphipods
and chaetognaths (027 lt r lt 060) Other less abundant
groups (lt1) such as siphonophores medusae
cladocerans heteropods appendicularians and echino-
derm larvae showed a negative correlation (-064 lt r lt
-028 P lt 005)
Zooplankton biomass and doliolids were positively
correlated with nitrates+nitrites phosphates and sili-
cates however none of most abundant groups
(copepods chaetognaths amphipods euphausiids)
were significantly correlated with nutrients In spite of
this maps of spatial distribution showed concordance
of high abundance values of copepods with nitrate+ nitrite during January in the central zone of the oceanic
transect and during October in the southeast region
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048
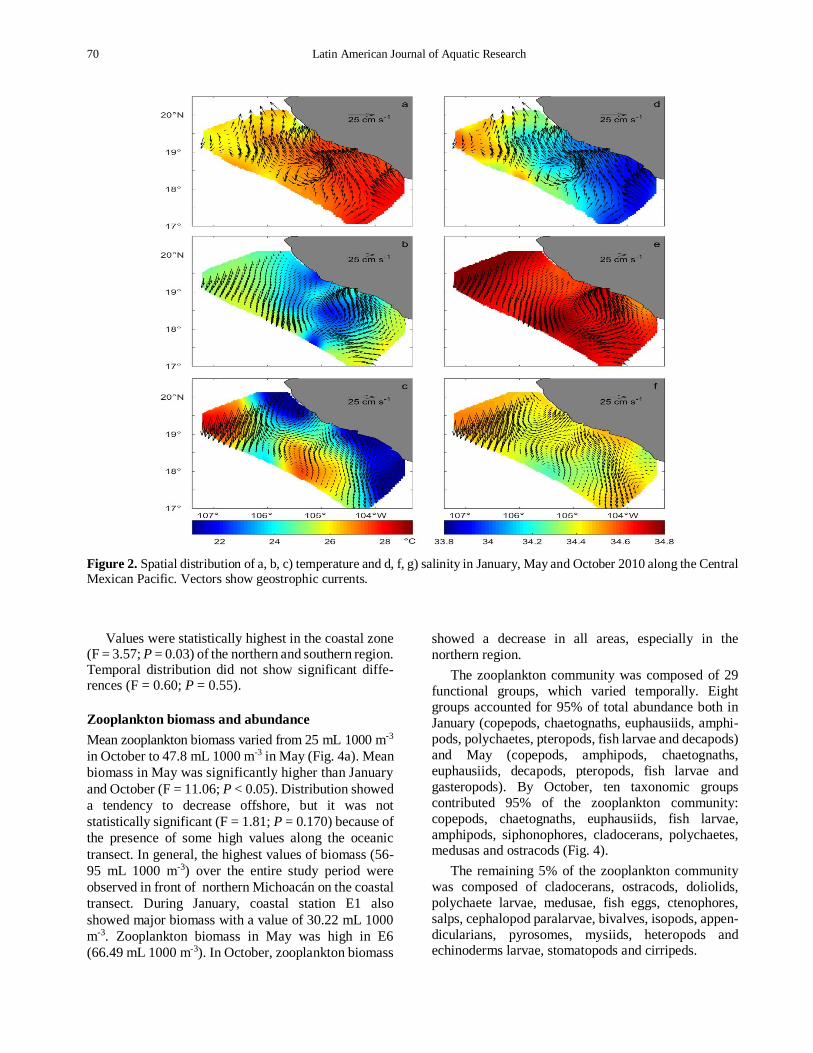
70 Latin American Journal of Aquatic Research
Figure 2 Spatial distribution of a b c) temperature and d f g) salinity in January May and October 2010 along the Central Mexican Pacific Vectors show geostrophic currents
Values were statistically highest in the coastal zone (F = 357 P = 003) of the northern and southern region Temporal distribution did not show significant diffe-rences (F = 060 P = 055)
Zooplankton biomass and abundance
Mean zooplankton biomass varied from 25 mL 1000 m-3
in October to 478 mL 1000 m-3 in May (Fig 4a) Mean
biomass in May was significantly higher than January
and October (F = 1106 P lt 005) Distribution showed
a tendency to decrease offshore but it was not
statistically significant (F = 181 P = 0170) because of
the presence of some high values along the oceanic
transect In general the highest values of biomass (56-
95 mL 1000 m-3) over the entire study period were
observed in front of northern Michoacaacuten on the coastal
transect During January coastal station E1 also
showed major biomass with a value of 3022 mL 1000
m-3 Zooplankton biomass in May was high in E6
(6649 mL 1000 m-3) In October zooplankton biomass
showed a decrease in all areas especially in the
northern region
The zooplankton community was composed of 29
functional groups which varied temporally Eight
groups accounted for 95 of total abundance both in
January (copepods chaetognaths euphausiids amphi-
pods polychaetes pteropods fish larvae and decapods)
and May (copepods amphipods chaetognaths
euphausiids decapods pteropods fish larvae and
gasteropods) By October ten taxonomic groups
contributed 95 of the zooplankton community
copepods chaetognaths euphausiids fish larvae
amphipods siphonophores cladocerans polychaetes medusas and ostracods (Fig 4)
The remaining 5 of the zooplankton community
was composed of cladocerans ostracods doliolids
polychaete larvae medusae fish eggs ctenophores salps cephalopod paralarvae bivalves isopods appen-
dicularians pyrosomes mysiids heteropods and echinoderms larvae stomatopods and cirripeds
Zooplankton inshore offshore Mexican Central Pacific 71
Distribution of major groups
Copepods chaetognaths euphausiids and amphipods
were the most abundant groups during the study period accounting for 87 of the total abundance (Fig 5b)
Maximum median value of copepod abundance of
10939 ind 1000 m-3 was recorded in May while the
minimum of 9297 ind 1000 m-3 was recorded in
October The January cruise registered a density of
9900 ind 1000 m-3 In October 20 of the stations located in the center of the study area were characterized
Table 1 Mean concentration and standard error of
nutrients in the Mexican Central Pacific during January
May-June and October 2010
Jan May-Jun Oct
NO3+NO2 220 plusmn 044 1467 plusmn 113 1102 plusmn 162
NH4 324 plusmn 018 227 plusmn 017 225 plusmn 014
PO4 072 plusmn 014 181 plusmn 027 117 plusmn 015
Si (OH)4 709 plusmn 091 2421 plusmn 229 929 plusmn 152
by the lowest values (0-5000 ind 1000 m-3) The largest
aggregation of this group during the entire surveyed
period was observed at the inshore southern region
particularly in station E14 where abundances up to 48711 ind 1000 m-3 were found (Fig 5)
The average abundance of chaetognaths over the
entire study period was 823 ind 1000 m-3 High
abundances of this group were observed in the southern
study area except in January when the northern zone
registered the highest abundance of 1150 ind 1000 m-3
During October cruise values inshore of the central and
northern zone were lower than the oceanic region (Fig 5)
Median abundance of euphausiids varied from 595
ind 1000 m-3 in January to 860 ind 1000 m3 in May
36 of stations during January showed values in the
range of 200-400 ind 1000 m-3 Spatial distribution
during January and May showed high abundance in the
southern inshore region and low in the northern while
in October high abundance was observed in the central-
oceanic zone (Fig 5) Except for October amphipod
abundance was high in the southern inshore zone in
May it was 14599 ind 1000 m-3 at station E17 but
typical values during this cruise were between 600 and
800 ind 1000 m-3 Cross shelf distribution was
characterized by high values near the coast that
decreased in the oceanic region except for oceanic
station E3 that registered high values with respect to
those near the coast during January and October (Fig
5) Average abundance of amphipods was 845 ind 1000
m-3 in October
No significant differences between samples from
different coastal or oceanic transects were found in
ANOSIM analyses of biotic variables (R = 0078 P =
008) while a marginally significant seasonality was
evident between each oceanographic cruise with an R =
0468 (P lt 001)
Zooplankton vs abiotic features
Biota and environment matching test (BEST) showed
minor influence of environmental variables on taxo-
nomic zooplankton groups Temperature and MLD
were the main variables explaining zooplankton group
distribution in the study area (Rho = 038 P = 001)
Spearman correlation analysis also showed low rank
values between abiotic variables and abundance of
zooplankton groups and biomass (Table 2)
Chl-a had no statistical correlation with zooplank-
ton biomass and was negatively correlated with
chaetognaths ctenophores and doliolids (-027 lt r lt
041) while bivalves and decapod larvae showed a
positive correlation (r = 029 030)
There was no significant correlation between
temperature and zooplankton biomass However
salinity and zooplankton biomass were positively
correlated (r = 027) Most of the major zooplankton
groups did not show any significant correlation with
temperature or salinity Minor groups like bivalves and
pyrosomes were positively related with temperature
and negatively with salinity Pteropods and gasteropods
showed positive correlation with temperature (r = 028
044) Ostracods and doliolids showed negative
correlation with salinity (r = -026 -031) while
siphonophores had a positive correlation (r = 028 P
lt005) Negative correlations were found between
temperature with fish larvae and medusae (Table 1)
The MLD was not significantly correlated with
zooplankton biomass but was positively related with 9
zooplankton groups including copepods amphipods
and chaetognaths (027 lt r lt 060) Other less abundant
groups (lt1) such as siphonophores medusae
cladocerans heteropods appendicularians and echino-
derm larvae showed a negative correlation (-064 lt r lt
-028 P lt 005)
Zooplankton biomass and doliolids were positively
correlated with nitrates+nitrites phosphates and sili-
cates however none of most abundant groups
(copepods chaetognaths amphipods euphausiids)
were significantly correlated with nutrients In spite of
this maps of spatial distribution showed concordance
of high abundance values of copepods with nitrate+ nitrite during January in the central zone of the oceanic
transect and during October in the southeast region
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

Zooplankton inshore offshore Mexican Central Pacific 71
Distribution of major groups
Copepods chaetognaths euphausiids and amphipods
were the most abundant groups during the study period accounting for 87 of the total abundance (Fig 5b)
Maximum median value of copepod abundance of
10939 ind 1000 m-3 was recorded in May while the
minimum of 9297 ind 1000 m-3 was recorded in
October The January cruise registered a density of
9900 ind 1000 m-3 In October 20 of the stations located in the center of the study area were characterized
Table 1 Mean concentration and standard error of
nutrients in the Mexican Central Pacific during January
May-June and October 2010
Jan May-Jun Oct
NO3+NO2 220 plusmn 044 1467 plusmn 113 1102 plusmn 162
NH4 324 plusmn 018 227 plusmn 017 225 plusmn 014
PO4 072 plusmn 014 181 plusmn 027 117 plusmn 015
Si (OH)4 709 plusmn 091 2421 plusmn 229 929 plusmn 152
by the lowest values (0-5000 ind 1000 m-3) The largest
aggregation of this group during the entire surveyed
period was observed at the inshore southern region
particularly in station E14 where abundances up to 48711 ind 1000 m-3 were found (Fig 5)
The average abundance of chaetognaths over the
entire study period was 823 ind 1000 m-3 High
abundances of this group were observed in the southern
study area except in January when the northern zone
registered the highest abundance of 1150 ind 1000 m-3
During October cruise values inshore of the central and
northern zone were lower than the oceanic region (Fig 5)
Median abundance of euphausiids varied from 595
ind 1000 m-3 in January to 860 ind 1000 m3 in May
36 of stations during January showed values in the
range of 200-400 ind 1000 m-3 Spatial distribution
during January and May showed high abundance in the
southern inshore region and low in the northern while
in October high abundance was observed in the central-
oceanic zone (Fig 5) Except for October amphipod
abundance was high in the southern inshore zone in
May it was 14599 ind 1000 m-3 at station E17 but
typical values during this cruise were between 600 and
800 ind 1000 m-3 Cross shelf distribution was
characterized by high values near the coast that
decreased in the oceanic region except for oceanic
station E3 that registered high values with respect to
those near the coast during January and October (Fig
5) Average abundance of amphipods was 845 ind 1000
m-3 in October
No significant differences between samples from
different coastal or oceanic transects were found in
ANOSIM analyses of biotic variables (R = 0078 P =
008) while a marginally significant seasonality was
evident between each oceanographic cruise with an R =
0468 (P lt 001)
Zooplankton vs abiotic features
Biota and environment matching test (BEST) showed
minor influence of environmental variables on taxo-
nomic zooplankton groups Temperature and MLD
were the main variables explaining zooplankton group
distribution in the study area (Rho = 038 P = 001)
Spearman correlation analysis also showed low rank
values between abiotic variables and abundance of
zooplankton groups and biomass (Table 2)
Chl-a had no statistical correlation with zooplank-
ton biomass and was negatively correlated with
chaetognaths ctenophores and doliolids (-027 lt r lt
041) while bivalves and decapod larvae showed a
positive correlation (r = 029 030)
There was no significant correlation between
temperature and zooplankton biomass However
salinity and zooplankton biomass were positively
correlated (r = 027) Most of the major zooplankton
groups did not show any significant correlation with
temperature or salinity Minor groups like bivalves and
pyrosomes were positively related with temperature
and negatively with salinity Pteropods and gasteropods
showed positive correlation with temperature (r = 028
044) Ostracods and doliolids showed negative
correlation with salinity (r = -026 -031) while
siphonophores had a positive correlation (r = 028 P
lt005) Negative correlations were found between
temperature with fish larvae and medusae (Table 1)
The MLD was not significantly correlated with
zooplankton biomass but was positively related with 9
zooplankton groups including copepods amphipods
and chaetognaths (027 lt r lt 060) Other less abundant
groups (lt1) such as siphonophores medusae
cladocerans heteropods appendicularians and echino-
derm larvae showed a negative correlation (-064 lt r lt
-028 P lt 005)
Zooplankton biomass and doliolids were positively
correlated with nitrates+nitrites phosphates and sili-
cates however none of most abundant groups
(copepods chaetognaths amphipods euphausiids)
were significantly correlated with nutrients In spite of
this maps of spatial distribution showed concordance
of high abundance values of copepods with nitrate+ nitrite during January in the central zone of the oceanic
transect and during October in the southeast region
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048
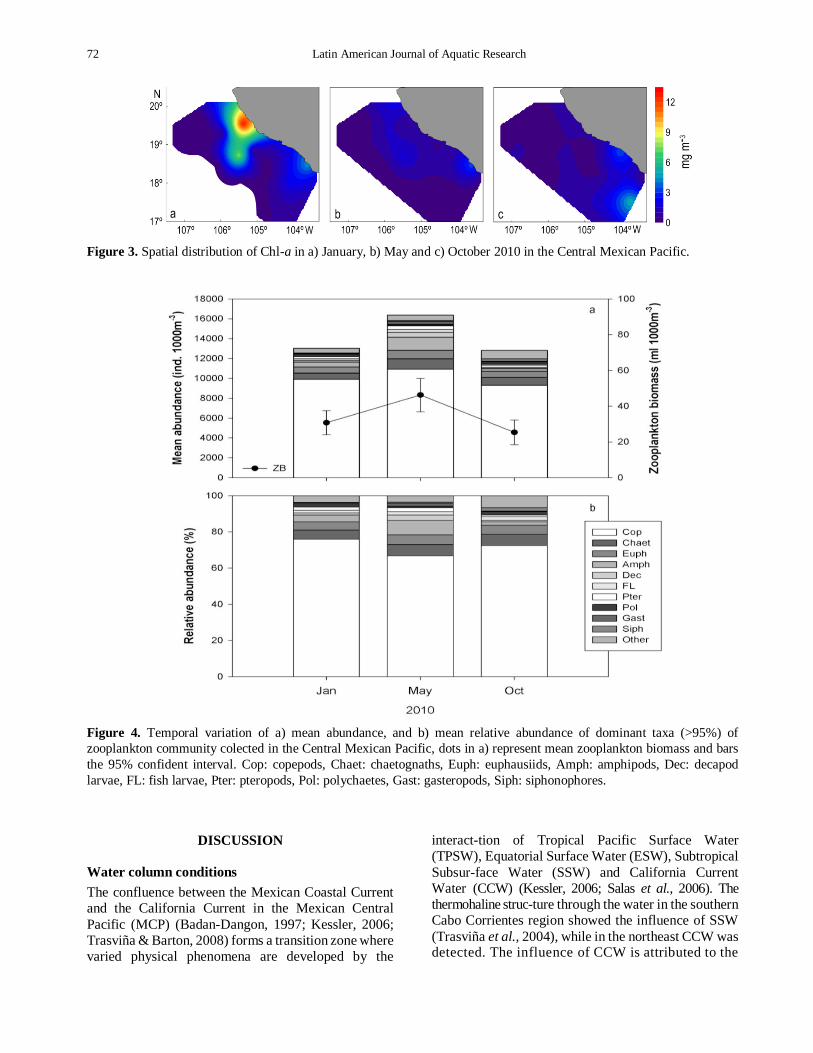
72 Latin American Journal of Aquatic Research
Figure 3 Spatial distribution of Chl-a in a) January b) May and c) October 2010 in the Central Mexican Pacific
Figure 4 Temporal variation of a) mean abundance and b) mean relative abundance of dominant taxa (gt95) of
zooplankton community colected in the Central Mexican Pacific dots in a) represent mean zooplankton biomass and bars
the 95 confident interval Cop copepods Chaet chaetognaths Euph euphausiids Amph amphipods Dec decapod
larvae FL fish larvae Pter pteropods Pol polychaetes Gast gasteropods Siph siphonophores
DISCUSSION
Water column conditions
The confluence between the Mexican Coastal Current
and the California Current in the Mexican Central
Pacific (MCP) (Badan-Dangon 1997 Kessler 2006
Trasvintildea amp Barton 2008) forms a transition zone where
varied physical phenomena are developed by the
interact-tion of Tropical Pacific Surface Water
(TPSW) Equatorial Surface Water (ESW) Subtropical
Subsur-face Water (SSW) and California Current
Water (CCW) (Kessler 2006 Salas et al 2006) The
thermohaline struc-ture through the water in the southern Cabo Corrientes region showed the influence of SSW
(Trasvintildea et al 2004) while in the northeast CCW was detected The influence of CCW is attributed to the
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048
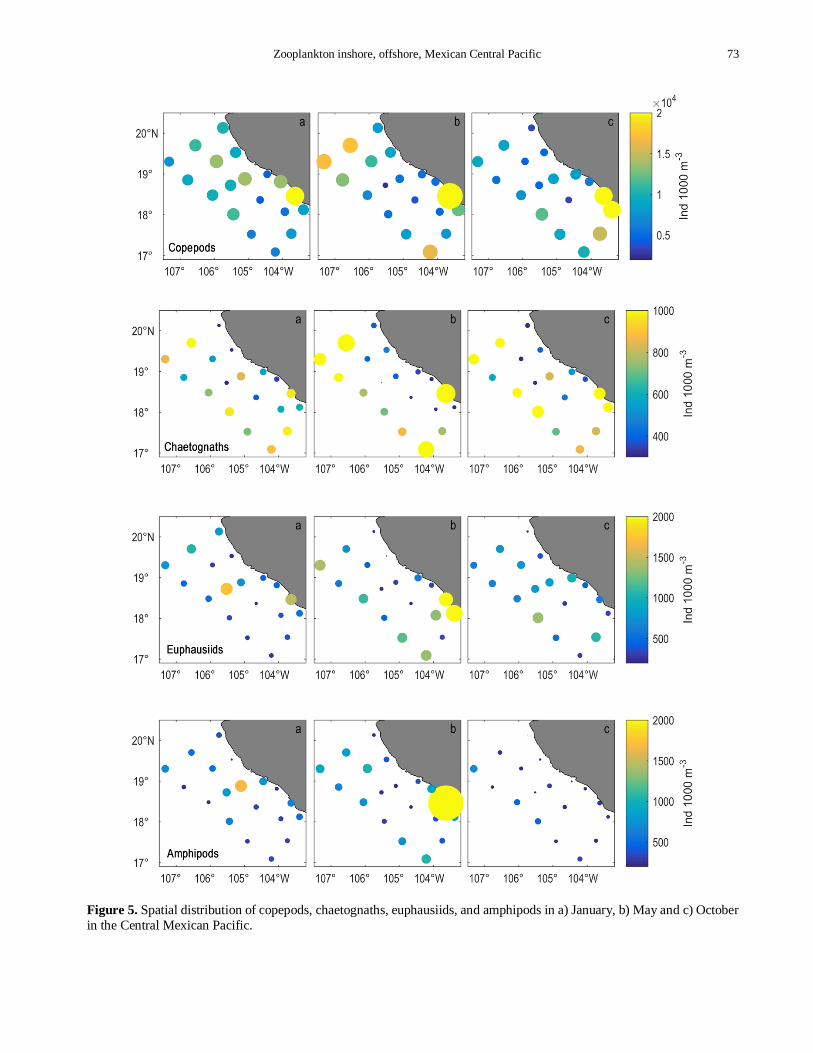
Zooplankton inshore offshore Mexican Central Pacific 73
Figure 5 Spatial distribution of copepods chaetognaths euphausiids and amphipods in a) January b) May and c) October
in the Central Mexican Pacific
74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

74 Latin American Journal of Aquatic Research
Ta
ble
2
Sp
earm
an r
ank c
orr
elat
ion
s bet
wee
n a
bio
tic
var
iable
s an
d m
ajor
dom
inan
t ta
xa
(gt9
5
) of
the
zoop
lan
kto
n c
om
munit
y C
orr
elat
ion v
alues
in b
old
are
sign
ific
ant
at P
= 0
05 C
UI
coas
tal
upw
elli
ng i
nd
ex M
LD
m
ixed
lay
er d
epth
T
te
mp
erat
ure
S
sa
lin
ity
Ch
l-a
ch
loro
ph
yll-
a N
O3
+N
O2
nit
rate
s+n
itri
tes
NH
4
amm
on
ium
P
O4
ph
osp
hat
e
SiO
2
sili
cate
s ac
id
ZB
zo
op
lan
kto
n b
iom
ass
Cop
co
pep
od
s A
mph
am
ph
ipods
Euph
euphau
siid
s D
P
dec
apod l
arvae
P
ol
poly
chae
tes
Pte
r p
tero
pods
Sip
h
siph
onoph
ores
C
hae
t c
hae
tog
nat
hs
FL
fi
sh l
arvae
an
d G
ast
gas
tero
pod
s
Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

Zooplankton inshore offshore Mexican Central Pacific 75
equatorward flow occurring mainly during winter-
spring months (Ambriz-Arreola et al 2012) which
may be upwelled due the presence of mesoscale
features originated by the steep bathymetry of the region (Salas et al 2006 Ambriz-Arreola et al 2012)
The thermohaline structure of the water column in
the study area has been described by Filonov amp
Tereshchenko (2010) and Ambriz-Arreola et al (2012)
who established three periods 1) mixed water column
from February to early June 2) a stratified period from
June to November and 3) a semi-mixed transitional
period commonly occurring in June and December
January In the context of our results the influence of
El Nintildeo conditions from January to March (Blunden et al 2011) shifted oceanographic and biological
conditions Geostrophic circulation in the northern zone
was dominant in the region and may have caused
deepening of the MLD Increased wind intensity in late
May to June (Bulgakov amp Martiacutenez-Zartarain 2006
Pantoja et al 2012) and the influence of La Nintildea
conditions from July to December (Diamond 2011)
promoted intense upwelling events which were
reflected in a shallow MLD in May and October these
processes lead to higher concentration of nitrites
nitrates phosphates and silicic acids and zooplankton biomass in the coastal zone
Chl-a showed an inverse behavior with respect to
nutrients and zooplankton biomass Mean concen-
tration of Chl-a in the study period was 124 mg m-3
Concentrations in the coastal zone were statistically
higher due to coastal upwelling promoted by the
dominant northwestern winds in the region especially
in January Considering phytoplankton as the major
food supply for zooplankton we expected concen-
trations of Chl-a to be positively correlated with the
high abundance of zooplankton biomass registered in
May but we found the inverse This result could be
explained by two possible reasons 1) a coastal
upwelling event prior to the survey period (15 days CUI
= 142 m3 s-1 100 m-1 coastline) caused phytoplankton to
bloom producing available particulate organic matter
compounds such as lipids carbohydrates and proteins
(not analyzed in this study) as food for zooplankton
during the survey period (Faumlrber-Lorda et al 2004a
2004b) and 2) the presence of small cells of marine
cyanobacteria such as Prochlorococcus in upwelling
regions and coastal zones of tropical oceans Although
we did not analyze the concentration of this
cyanobacteria it tends to have faster nutrient uptake
rates without contributing significantly to the
productivity of blooms (Ward et al 2012 2013)
Zooplankton grazing could be exerting a top-down
control over phytoplankton preventing the use of all
available nutrients and therefore affecting Chl-a This
has been previously reported in different areas of the
Central Mexican Pacific under warm conditions in
studies showing a clear association of primary
productivity with higher trophic levels (Torres-Orozco
et al 2005 Gaxiola-Castro et al 2008 Loacutepez-
Sandoval et al 2009 Ambriz-Arreola et al 2012)
Although El Nintildeo conditions registered in this study
ended in April transition into a ldquonormal periodrdquo could
have influenced phytoplankton cell acclimatization
(Lluch-Cota et al 1999) and induced lower Chl-a
production during May (Fig 3)
Zooplankton variability abundance and structure
In this study temporal variations of zooplankton
biomass appeared to be strongly influenced by
temperature considering that maximum values were
observed in the presence of low temperatures during
May Different authors have recognized the same
pattern in the MCP (Franco-Gordo et al 2004) and
other areas of Mexico (Hernaacutendez-Trujillo et al 2001)
which could be explained by the metabolism of these
organisms where high temperatures cause their
metabolic requirements to exceed available energy
(primary production) resulting in low biomass
development (Coyle et al 2008) Nevertheless low
zooplankton biomass values in this study probably
resulted from the broader oceanic sampling area (100
nm from coast) than the other studies (lt3 nm) and
therefore fertilization mechanisms were less intense
Our zooplankton values were also lower in the presence
of high values of nutrients and Chl-a during the January
and October cruises which could be due to the time lag
between the transference from primary producers to
zooplankter organisms Fernaacutendez-Aacutelamo amp Faumlrber-
Lorda (2006) reported a time lag of up to two months
in temperate regions while tropical latitudes could vary
temporally due the presence of local hydrographic
conditions This was the case of the May cruise where
high biomass values recorded in the oceanic region
were likely due to the presence of a cyclonic gyre in the inshore region (Fig 2)
Negative correlation of zooplankton biomass and
SST has been reported by other authors (Franco-Gordo et al 2002 2004 Fernaacutendez-Aacutelamo amp Faumlrber-Lorda
2006) The presence of the ENSO phenomenon during
January could have affected the zooplankton commu-
nity as the lowest biomass values were recorded during
that period The influence of abnormally high tempe-
ratures could lead to lower upwelling intensity lower
concentration of nutrients and the establishment of phytoplankton species not included in the zooplankton
diet which has been reported to cause disturbances in
the food chain including uptake and utilizations and
76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

76 Latin American Journal of Aquatic Research
competitivepredatory interactions (Barber amp Chavez 1983 Sebastian et al 2012)
Zooplankton structure in the all three cruises was dominated by copepods (66-76) they are cosmopolite organisms with a great capacity of adaptation to varied
environmental conditions in the same region (Hernaacutendez-Trujillo et al 2001 Lo et al 2004 Lavaniegos et al 2012) Chaetognaths were the only carnivorous zooplankton that significantly contributed to total abundance with 5-8 in agreement with Hossfeld (1996) and Aacutelvarez-Cadena et al (2008) who
reported that this zooplankton group accounted for 5-15 of total zooplankton abundance
During January crustacean herbivores such as
euphausiids and amphipods were dominants and other
groups including polychaetes pteropods and some
groups of meroplankton such as fish and decapod
larvae were also present During the May cruise relative
abundance of polychaetes diminished to lt1 while
gasteropods represented gt1 Lack of significant
correlation between temperature and polychaete abun-
dance in this study demonstrates the great species
diversity and affinity that can be found in the MCP
since it has been demonstrated that some species are
abundant in upwelling zones while other species could
be indicators of warm tropical water masses (Fernaacutendez-
Aacutelamo amp Sanvicente-Anorve 2005 Fernaacutendez-Aacutelamo amp
Faumlrber-Lorda 2006)
Only doliolids and pyrosomes registered a positive
correlation with temperature and negative with salinity
this could be explained by the influence of Tropical
Pacific Surface Water (TPSW) characterized by high
values of temperature low salinity and low nutrient availability (Fiedler amp Talley 2006)
There was a correlation between the abundance of
fish larvae and the relatively low SST high
productivity and high zooplankton biomass found
along the coastal zone in front of Michoacaacuten and
southern Colima This may be the influence of coastal
upwelling during the May and October cruises where
the input of subsurface nutrients promotes the
production of certain species of phytoplankton that
serve as food for fish larvae Leoacuten-Chaacutevez et al (2010)
studied the influence of circulation on larval fish
assemblages in the Eastern Tropical Pacific during
2010 and proposed that distribution is controlled by
large scale and mesoscale hydrodynamic processes in
the Eastern Tropical Pacific Although in this study fish
larvae were not identified to species level many
individuals of this group appear to be carried by a
northward current in January and May whereas in October the abundance of fish larvae coincide with an
upwelling zone in the Cabo Corrientes region and a
southward current in the coastal region of Michoacaacuten (distribution map not shown here)
The positive correlation of nitrites nitrates phosphates and silicates with zooplankton biomass and nitrate+nitrite with cladocerans medusae and siphono-phores leads us to suggest that upwelled waters could favor the abundance of these groups through primary production enhancement Although Richardson et al (2012) consider that the influence of high nutrients leads to the presence of large crustaceans and carnivo-rous zooplankton the energy transfer throughout trophic levels needs to be studied in order to establish the influence of inorganic compounds on specific zooplankton groups There are few studies focused on the marine environment because of its omnivorous nature which comprises a mixture of trophic types species size-classes and detritus (Gentleman et al 2003) This latter could explain why contrary to expectations and findings from previous studies (Franco-Gordo et al 2001 Lavaniegos et al 2002) in this study there was no statistical evidence of a correlation between Chl-a zooplankton biomass and the principal herbivorous groups (copepods euphau-siids ostracods amphipods salps) It is also important consider the dynamic nature of the marine environ-ment where there are no absolute distributional boundaries particularly in a transition region such as the MCP as well as the time lag of more or less one month for zooplankton abundance to reflect food intake (Faumlrber-Lorda et al 2004b Fernaacutendez-Aacutelamo amp Faumlrber-Lorda 2006) This study can be considered an approach to help understand the hydroclimatic influence on zooplankton groups of the Mexican Central Pacific
ACKNOWLEDGEMENTS
The Federal Commission of Electricity (CFE) and
University of Colima supported this study through the project ldquoMarine mammals and turtles monitoring in
Mexican Central Pacificrdquo during 2010 We want to thank all the people who assisted during the
oceanographic surveys on board the BIP-XII vessel
REFERENCES
Aacutelvarez-Cadena JN AR Almaral-Mendivil U
Ordoacutentildeez-Loacutepez amp A Uicab-Sabido 2008 Compo-
sicioacuten abundancia y distribucioacuten de las especies de
quetognatos del litoral norte del Caribe de Meacutexico
Hidrobiologica 18(1) 37-48
Ambriz-Arreola I J Goacutemez-Gutieacuterrez C Franco-
Gordo BE Lavaniegos amp E Godiacutenez-Domiacutenguez
2012 Influence of coastal upwelling-downwelling
variability on tropical euphausiid abundance and
Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

Zooplankton inshore offshore Mexican Central Pacific 77
community structure in the inshore Mexican Central
Pacific Mar Ecol Prog Ser 451 119-136
Badan-Dangon A 1997 La corriente costera de Costa
Rica en el Paciacutefico Mexicano In MF Laviacuten (ed)
Contribuciones a la oceanografiacutea fiacutesica en Meacutexico
Monografiacutea 3 Unioacuten Geofiacutesica Mexicana Meacutexico
pp 99-112
Barber RT amp FP Chavez 1983 Biological conse-
quences of El Nintildeo Science 222 1203-1210
Beers JR 1976 Determination of zooplankton biomass
In HF Steedman (ed) Zooplankton fixation and
preservation UNESCO Press Pariacutes pp 35-86
Blunden J D Arndt amp M Baringer 2011 State of the
climate in 2010 Bull Am Meteorol Soc 92(6) S1-S266
Bulgakov SN amp A Martiacutenez-Zartarain 2006 Surgencia
y vientos favorables en la costa oriental del Paciacutefico
mexicano In MC Jimeacutenez-Quiroz amp E Espino-Barr
(eds) Los recursos pesqueros y acuiacutecolas de Jalisco Colima y Michoacaacuten Vol 1 INP-SAGARPA
Manzanillo Colima pp 29-40
Coyle KO AI Pinchuk LB Eisner amp JM Napp
2008 Zooplankton species composition abundance
and biomass on the eastern Bering Sea shelf during
summer the potential role of water-column stability and nutrients in structuring the zooplankton
community Deep-Sea Res II 55 1775-1791
Diamond AJ 2011 Tropics In J Blundent DS Arndt
amp MO Baringer (eds) State of the climate in 2010
Bull Am Meteorol Soc 92(Suppl 6) S109-S138
Faumlrber-Lorda J M Laviacuten amp M Guerrero-Ruiz 2004a
Effects of wind forcing on the trophic conditions
zooplankton biomass and krill biochemical
composition in the Gulf of Tehuantepec Deep-Sea
Res II 51(6) 601-614
Faumlrber-Lorda J A Trasvintildea amp PC Verdiacuten 2004b
Trophic conditions and zooplankton distribution in the
entrance of the Sea of Corteacutes during summer Deep-
Sea Res II 51(6) 615-627
Fernaacutendez-Aacutelamo MA amp J Faumlrber-Lorda 2006
Zooplankton and the oceanography of the eastern
tropical Pacific a review Prog Oceanogr 69(2) 318-
359
Fernandez-Alamo MA amp L Sanvicente-Anorve 2005
Holoplanktonic polychaetes from the Gulf of
Tehantepec Mexico Cah Biol Mar 46(3) 227-239
Fiedler PC amp LD Talley 2006 Hydrography of the
eastern tropical Pacific a review Prog Oceanogr 69(2) 143-180
Filonov A amp I Tereshchenko 2010 El reacutegimen
termodinaacutemico en la costa de los estados de Jalisco y
Colima In E Godiacutenez-Domiacutenguez MdC Franco-
Gordo JA Rojo-Vaacutezquez FA Silva-Baacutetiz amp G
Gonzaacutelez-Sansoacuten (eds) Ecosistemas marinos de la
costa sur de Jalisco y Colima Universidad de
Guadalajara Jalisco pp 29-71
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2001 Zooplankton biomass variability in the
Mexican Eastern Tropical Pacific Pac Sci 55(2)
191-202
Franco-Gordo C E Godiacutenez-Domiacutenguez amp E Suaacuterez-
Morales 2002 Larval fish assemblages in waters off
the central Pacific coast of Mexico J Plankton Res
24(8) 775-784
Franco-Gordo C E Godiacutenez-Domiacutenguez AE Filonov
IE Tereshchenko amp J Freire 2004 Plankton biomass
and larval fish abundance prior to and during the El
Nintildeo period of 1997-1998 along the central Pacific
coast of Mexico Prog Oceanogr 63(3) 99-123
Gasca R C Franco-Gordo E Godiacutenez-Domiacutenguez amp E
Suaacuterez-Morales 2012 Hyperiid amphipod communi-
ty in the Eastern Tropical Pacific before during and
after El Nintildeo 1997-1998 Mar Ecol Prog Ser 455
123-139
Gaxiola-Castro G R Durazo B Lavaniegos M De la
Cruz-Orozco E Millaacuten-Nuacutentildeez L Soto-Mardones amp
J Cepeda-Morales 2008 Pelagic ecosystem response
to interannual variability off Baja California Cienc
Mar 34(2) 263-270
Gentleman W A Leising B Frost S Strom amp J
Murray 2003 Functional response for zooplankton
feeding on multiple resources a review of assumptions
and biological dynamics Deep-Sea Res II 50(22)
2847-2875
Godiacutenez VM E Beier M Laviacuten amp J Kurczyn 2010
Circulation at the entrance of the Gulf of California
from satellite altimeter and hydrographic observations
J Geophys Res 115(C4) 2156-2202
Goacutemez-Valdivia F A Pareacutes-Sierra amp A Flores-Morales
2015 The Mexican Coastal Current a subsurface
seasonal bridge that connects the tropical and
subtropical northeastern Pacific Cont Shelf Res 110
100-107
Grasshoff K K Kremling amp M Ehrhardt (eds) 1983
Methods of seawater analysis Verlag Chemie Florida
419 pp
Griffiths FB A Fleminger B Kimor amp M Vannucci
1976 Shipboard and curating techniques In HF
Steedman (ed) Zooplankton fixation and preser-
vation UNESCO Press Pariacutes pp 17-34
Hernaacutendez-Trujillo S F Goacutemez-Ochoa amp G Verdugo-
Diacuteaz 2001 Dinaacutemica del plancton en la regioacuten sur de
la Corriente de California Rev Biol Trop 49(1) 15-
30
Hossfeld B 1996 Distribution and biomass of arrow
worms (Chaetognatha) in Golfo de Nicoya and Golfo
Dulce Costa Rica Rev Biol Trop 44 157-172
78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048

78 Latin American Journal of Aquatic Research
Kara AB PA Rochford amp HE Hurlburt 2000 An
optimal definition for ocean mixed layer depth J Geophys Res 105(C7) 16803-16821
Kessler WS 2006 The circulation of the eastern tropical
Pacific a review Prog Oceanogr 69(2) 181-217
Kozak ER C Franco-Gordo E Suaacuterez-Morales amp R
Palomares-Garciacutea 2014 Seasonal and interannual
variability of the calanoid copepod community
structure in shelf waters of the Eastern Tropical
Pacific Mar Ecol Prog Ser 507 95-110
Lavaniegos B G Heckel amp P Ladroacuten de Guevara 2012
Variabilidad estacional de copeacutepodos y cladoacuteceros de
bahiacutea de Los Aacutengeles (Golfo de California) e
importancia de Acartia clausi como alimento del
tiburoacuten ballena Cienc Mar 38(1A) 11-30
Lavaniegos BE LC Jimeacutenez-Peacuterez amp G Gaxiola-
Castro 2002 Plankton response to El Nintildeo 1997-1998
and La Nintildea 1999 in the southern region of the
California Current Prog Oceanogr 54 33-58
Leoacuten-Chaacutevez CA L Saacutenchez-Velasco E Beier MF
Laviacuten VM Godiacutenez amp J Faumlrber-Lorda 2010 Larval
fish assemblages and circulation in the Eastern
Tropical Pacific in autumn and winter J Plankton
Res 32(4) 397-410
Lo W-T C-L Chung amp C-T Shih 2004 Seasonal distribution of copepods in Tapong Bay southwestern
Taiwan Zool Stud 43(2) 464-474
Loacutepez-Sandoval D J Lara-Lara M Laviacuten S Aacutelvarez-
Borrego amp G Gaxiola-Castro 2009 Productividad
primaria en el Paciacutefico oriental tropical adyacente a
Cabo Corrientes Meacutexico Cienc Mar 35(2) 169-182
Lluch-Cota D D Lluch-Belda S Lluch-Cota J Loacutepez-
Martiacutenez 1999 Las pesqueriacuteas y El Nintildeo In Los
impactos de El Nintildeo en Meacutexico Sep-Conacyt-IAI
Meacutexico DF pp 137-178
Newell GE amp RC Newell 1973 Marine plankton a
practical guide Hutchinson Educational London 221
pp
Pantoja D S Marinone A Pareacutes-Sierra amp F Goacutemez-
Valdivia 2012 Modelacioacuten numeacuterica de la hidro-
grafiacutea y circulacioacuten estacional y de mesoescala en el
Paciacutefico central mexicano Cienc Mar 38(2) 363-
379
Riandey V G Champalbert F Carlotti I Taupier-
Letage amp D Thibault-Botha 2005 Zooplankton
distribution related to the hydrodynamic features in the
Algerian Basin (western Mediterranean Sea) in
summer 1997 Deep-Sea Res I 52(11) 2029-2048
Received 5 March 2016 Accepted 12 September 2016
Richardson AJ D McKinnon amp KM Swadling 2012
Zooplankton In E Poloczanska A Hobday amp A Richardson (eds) A marine climate change impacts
and adaptation report card for Australia 2012
[httpwwwoceanclimatechangeorau] Reviewed 5
July 2015
Salas J D Gomis A Olivos-Ortiz amp G Garcia-Uribe
2006 Seasonal hydrodynamical features on the continental shelf of Colima (west coast of Mexico)
Sci Mar 70(4) 719-726
Sebastian P H Stibor S Berger amp S Diehl 2012
Effects of water temperature and mixed layer depth on
zooplankton body size Mar Biol 159(11) 2431-
2440
Smith DBL amp KB Johnson 1996 A guide to marine
coastal plankton and marine invertebrate larvae
KendallHunt Publishing Company Iowa 221 pp
Strickland JDH amp TR Parsons 1972 A practical
handbook of sea water analysis Fish Res Bd Can
Bull 167 310 pp
Talley LD GL Pickard WJ Emery amp JH Swift
2011 Descriptive physical oceanography an intro-
duction Elsevier Boston 560 pp
Todd CD MS Laverack amp G Boxshall 1996 Coastal
marine zooplankton A practical manual for students Cambridge University Press Cambridge 116 pp
Torres-Orozco E A Trasvintildea A Muhlia-Melo amp S
Ortega-Garciacutea 2005 Mesoscale dynamics and
yellowfin tuna catches in the Mexican Pacific Cienc
Mar 31(4) 671-683
Trasvintildea A amp ED Barton 2008 Summer circulation in
the Mexican tropical Pacific Deep-Sea Res I 55(5)
587-607
Trasvintildea A D Luch-Cota AE Filonov amp A Gallegos
2004 Oceanografiacutea y El Nintildeo In V Magantildea-Rueda
(ed) Los impactos del Nintildeo en Meacutexico Universidad
Autoacutenoma de Meacutexico Secretariacutea de Gobernacioacuten
Meacutexico pp 69-101
Ward B S Dutkiewicz O Jahn amp M Follows 2012 A
size-structured food-web model for the global ocean
Limnol Oceanogr 57(6) 1877-1891
Ward BA S Dutkiewicz amp MJ Follows 2013 Modelling spatial and temporal patterns in size-
structured marine plankton communities top-down
and bottom-up controls J Plankton Res 36(1) 31-47
Yebra L RP Harris EJH Head I Yashayaev LR
Harris amp AG Hirst 2009 Mesoscale physical
variability affects zooplankton production in the Labrador Sea Deep-Sea Res I 56(5) 703-715
Zamudio L HE Hurlburt EJ Metzger amp CE Tilburg
2007 Tropical wave induced oceanic eddies at Cabo
Corrientes and the Mariacutea Islands Mexico J Geophys
Res 112 C05048







