

Protein synthesis 30.4 Ribosome Structure and
Assembly 30.5 Mechanics of Protein Synthesis

30.4 Ribosome Structure and Assembly
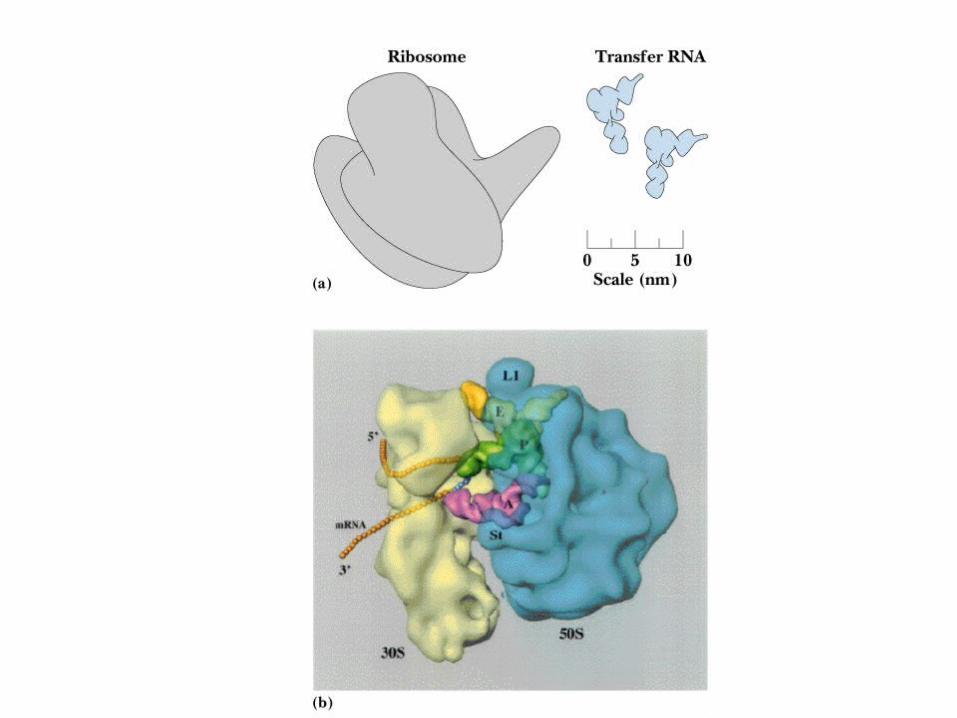
E. coli ribosome is 25 nm diameter, 2520 kD in mass, and consists of two unequal subunits that dissociate at < 1mM Mg2+
30S subunit is 930 kD with 21 proteins and a 16S rRNA
50S subunit is 1590 kD with 31 proteins and two rRNAs: 23S rRNA and 5S rRNA
These ribosomes and others are roughly 2/3 RNA 20,000 ribosomes in a cell, 20% of cell's mass

Ribosomal RNA 3 rRNA molecules
23S, 16S, 5S Derived from a single 30S rRNA
precursor transcript Extensive intrachain H-bonding 2/3 rRNA is helical

Ribosomal Proteins One of each per ribosome, except
L7/L12 with 4 L7/L12 identical except for extent of
acetylation at N-terminus Only one protein is common to
large and small subunits: S20 = L26 Variety of structures, still being
characterized

Ribosome Assembly/Structure
If individual proteins and rRNAs are mixed, functional ribosomes will assemble
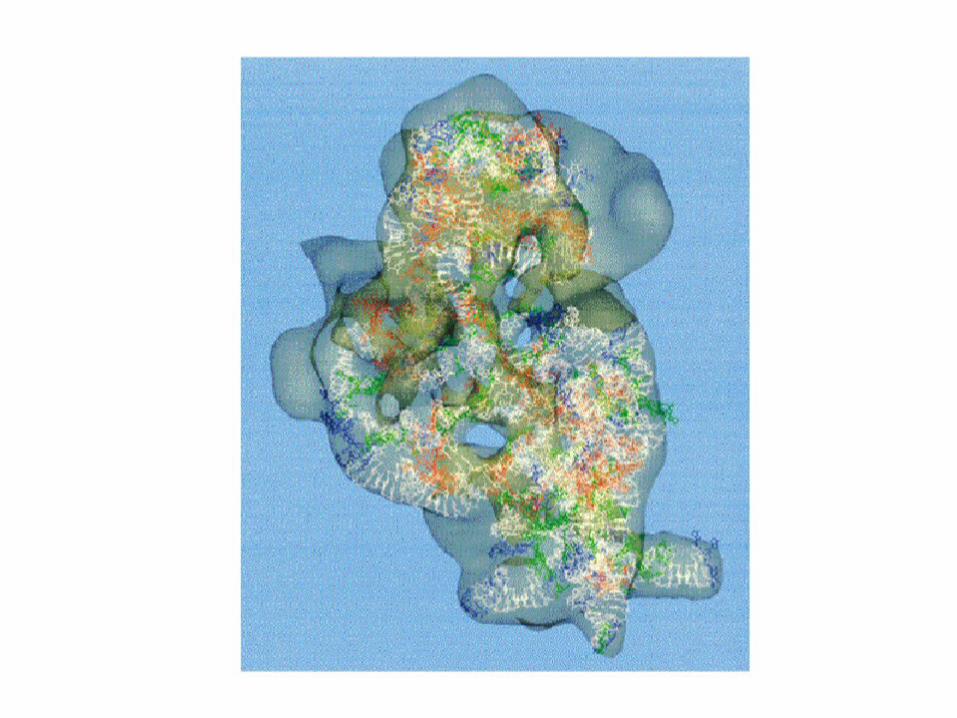
Gross structures of large and small subunits are known - see Figure 30.12
A tunnel runs through the large subunit Growing peptide chain is thought to thread
through the tunnel during protein synthesis


Eukaryotic Ribosomes
Mitochondrial and chloroplast ribosomes are quite similar to prokaryotic ribosomes, reflecting their supposed prokaryotic origin
Cytoplasmic ribosomes are larger and more complex, but many of the structural and functional properties are similar
See Table 30.6 for properties

30.5 Mechanics of Protein Synthesis
All protein synthesis involves three phases: initiation, elongation, termination
Initiation involves binding of mRNA and initiator aminoacyl-tRNA to small subunit, followed by binding of large subunit
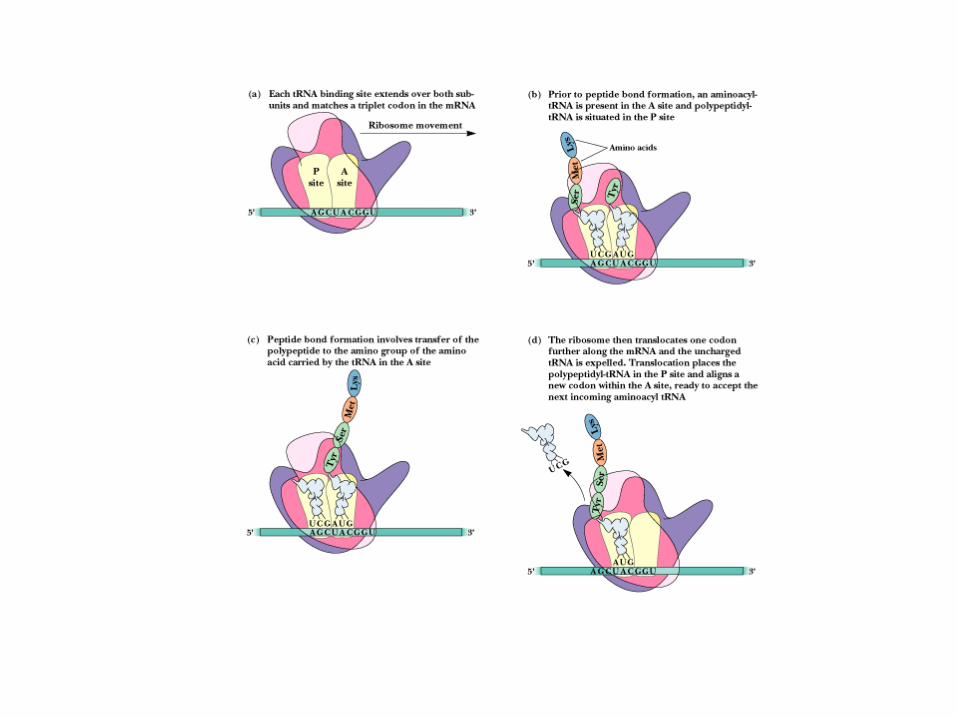
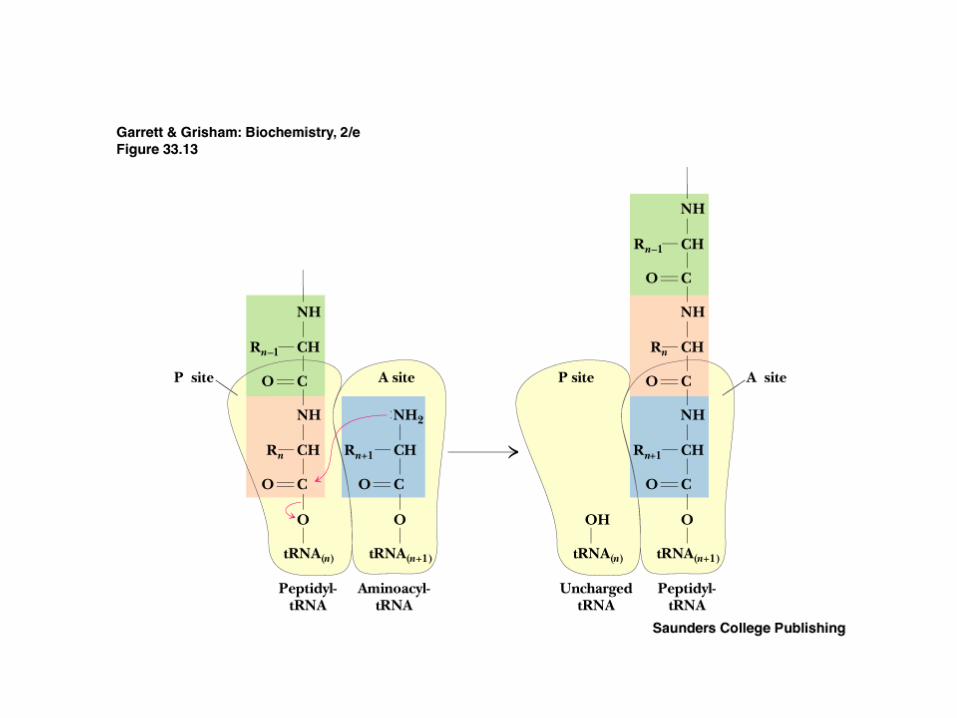
Elongation: synthesis of all peptide bonds - with tRNAs bound to acceptor (A) and peptidyl (P) sites. See Figure 30.13
Termination occurs when "stop codon" reached


Prokaryotic Initiation The initiator tRNA is one with a formylated
methionine: f-Met-tRNAfMet
It is only used for initiation, and regular Met-tRNAm
Met is used instead for Met addition
N-formyl methionine is first aa of all E.coli proteins, but this is cleaved in about half
A formyl transferase adds the formyl group (see Figure 30.15)


More Initiation Correct registration of mRNA on ribosome
requires alignment of a pyrimidine-rich sequence on 3'-end of 16S RNA with a purine-rich part of 5'-end of mRNA
The purine-rich segment - the ribosome-binding site - is known as the Shine-Dalgarno sequence (see Figure 30.17)
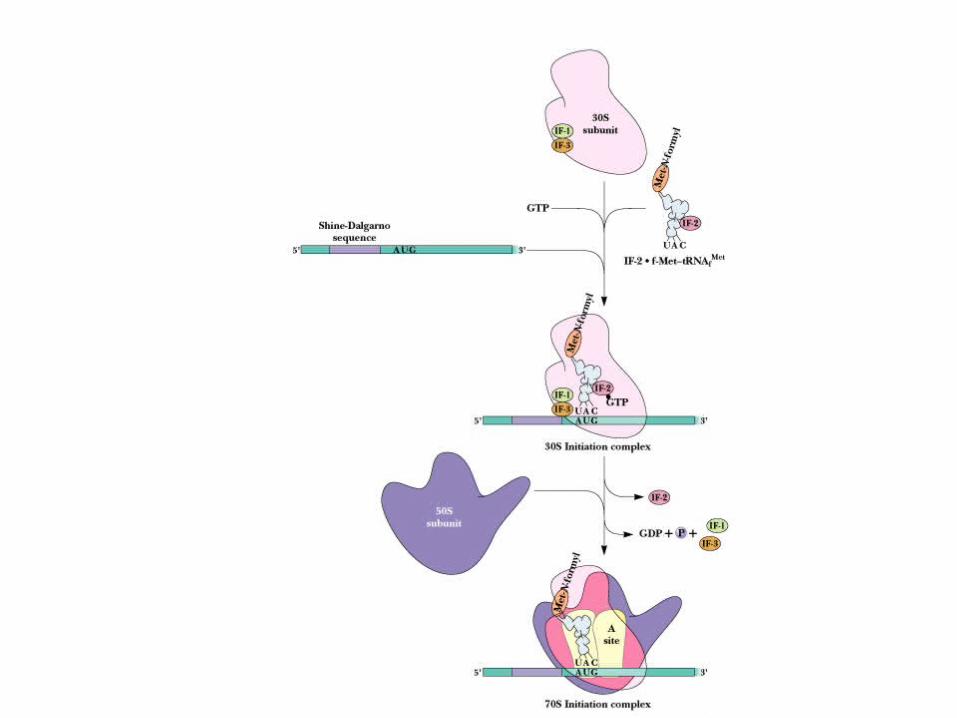
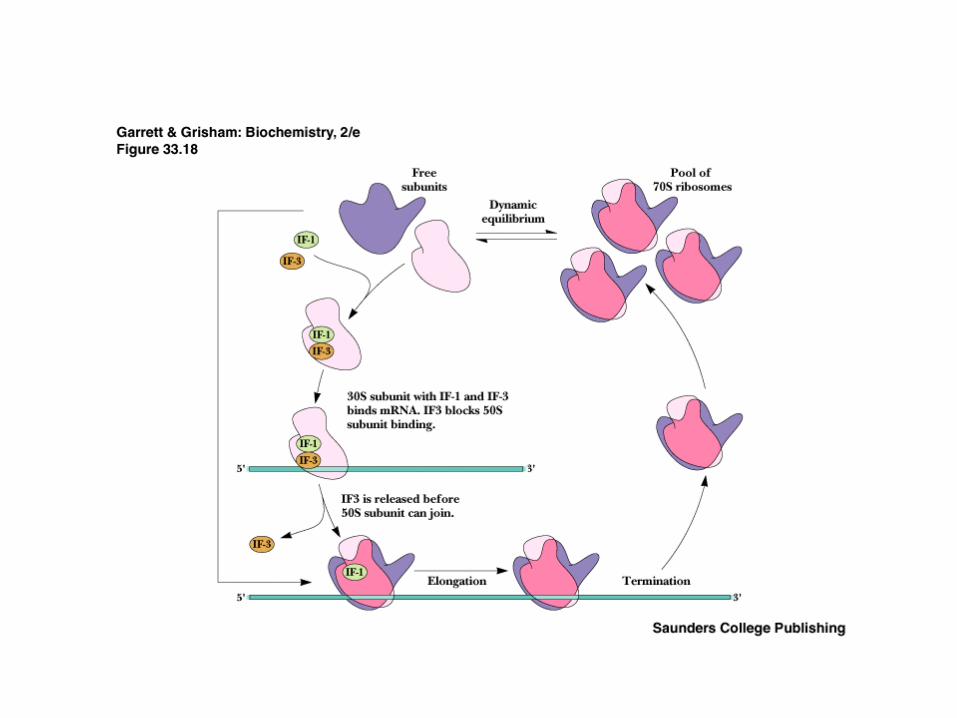
Initiation factor proteins, GTP, N-formyl-Met- tRNAfMet, mRNA and 30S ribosome form the 30S initiation complex


Events of Initiation 30S subunit with IF-1 and IF-3 binds mRNA,
IF-2, GTP and f-Met-tRNAfMet (Figure 30.18)
IF-2 delivers the initiator tRNA in a GTP-dependent process
Loss of the initiation factors leads to binding of 50S subunit
Note that the "acceptor site" is now poised to accept an incoming aminoacyl-tRNA


The Elongation Cycle Elongation factor Tu will bring each aa-tRNA into
the A site Decoding center of 16S rRNA makes sure the proper aa
tRNA is in the A site by direct surveillance
Peptide bond formation occurs by direct transfer of the peptidyl chain from the tRNA bearing it to the NH2 group of the new amino acid
Translocation of the one-residue-longer peptidyl tRNA to the P site to make room for the next incoming aa-tRNA at the A site.

EF-Tu-EF-Ts cycleEF-Tu-EF-Ts cycle
The The elongation factorselongation factors are vital to cell are vital to cell function, so they are present in significant function, so they are present in significant quantities (EF-Tu is 5% of total protein in quantities (EF-Tu is 5% of total protein in E. E. colicoli (Table 30.8) (Table 30.8)
EF-Tu binds aminoacyl-tRNA and GTP EF-Tu binds aminoacyl-tRNA and GTP Aminoacyl-tRNA binds to A site of ribosome Aminoacyl-tRNA binds to A site of ribosome
as a complex with EF-Tu and GTP as a complex with EF-Tu and GTP GTP is then hydrolyzed and EF-Tu:GDP GTP is then hydrolyzed and EF-Tu:GDP
complex dissociatescomplex dissociates EF-Ts recycles EF-Tu by exchanging GTP for EF-Ts recycles EF-Tu by exchanging GTP for
GDPGDP
The The elongation factorselongation factors are vital to cell are vital to cell function, so they are present in significant function, so they are present in significant quantities (EF-Tu is 5% of total protein in quantities (EF-Tu is 5% of total protein in E. E. colicoli (Table 30.8) (Table 30.8)
EF-Tu binds aminoacyl-tRNA and GTP EF-Tu binds aminoacyl-tRNA and GTP Aminoacyl-tRNA binds to A site of ribosome Aminoacyl-tRNA binds to A site of ribosome
as a complex with EF-Tu and GTP as a complex with EF-Tu and GTP GTP is then hydrolyzed and EF-Tu:GDP GTP is then hydrolyzed and EF-Tu:GDP
complex dissociatescomplex dissociates EF-Ts recycles EF-Tu by exchanging GTP for EF-Ts recycles EF-Tu by exchanging GTP for
GDPGDP


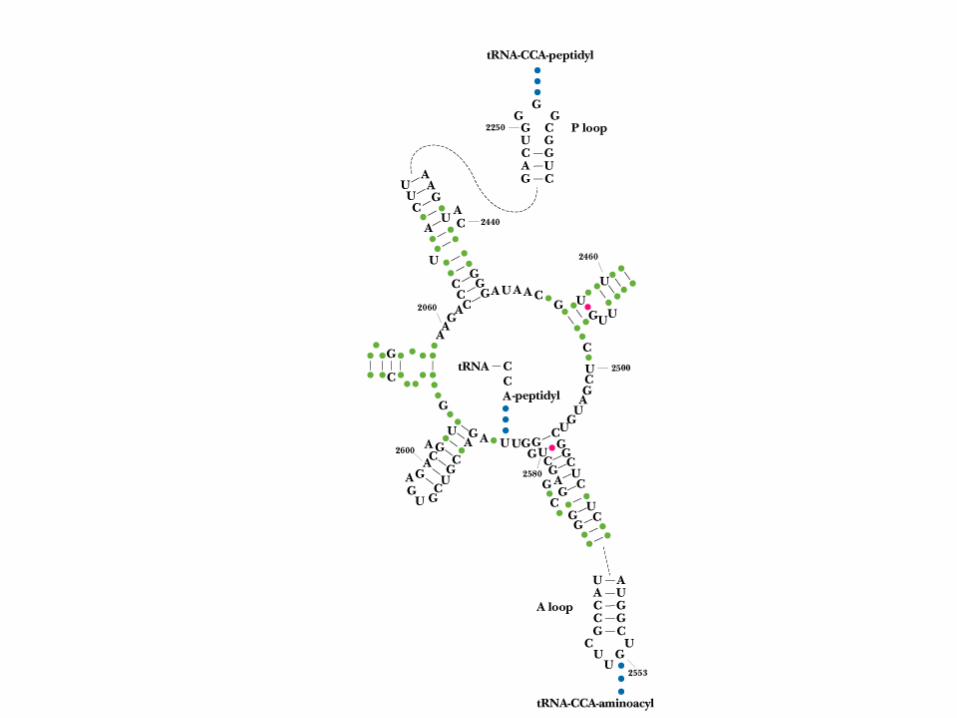
Peptidyl Transferase This is the central reaction of protein synthesis 23S rRNA is the peptidyl transferase! The "reaction center" of 23S rRNA is shown in
Figure 30.22 - these bases are among the most highly conserved in all of biology.
Translocation of peptidyl-tRNA from the A site to the P site follows (see Figures 30.19 & 30.21 ) catalyzed by EF-G.


Peptide Chain Termination
Proteins known as "release factors" recognize the stop codon at the A site
Presence of release factors with a nonsense codon at A site transforms the peptidyl transferase into a hydrolase, which cleaves the peptidyl chain from the tRNA carrier


The Role of GTP Hydrolysis IF-2, EF-Tu, EF-G, RF-3 are all GTP-
binding proteins Part of the G protein superfamily
Hydrolysis drives essential conformation changes
IF-2, EF-Tu, EF-G, RF-3 interact with the same site on the 50S subunit, the factor binding center

PolysomesPolysomes
mRNA with several ribosomes Polyribosomes
All protein synthesis occurs on polysomes
Procaryotes have around 10, eucaryotes fewer than 10 ribosomes
mRNA with several ribosomes Polyribosomes
All protein synthesis occurs on polysomes
Procaryotes have around 10, eucaryotes fewer than 10 ribosomes







