

RECRUITMENT OF MOTOR UNITS DURING LENGTHENING
CONTRACTIONS OF HUMAN FLEXOR CARPI RADIALIS
Paula J. Foht
BSc, University of Alberta, 1996
BKin, University of Calgary, 1998
THESIS SUBMïiTED IN PARTIAL FüLFiLLMENT OF
THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
(Neuroph ysiolog y)
in the School of Kinesiology
O Paula J. Foht. 1999
SiMON FRASER UNNERSlTY
December, 1999
Al1 tights feserved. This work may not be
teproduced in whoIe or in part, by photocopy
or other means, without permission of the author.

The author has grantted a non- L ' a m a accordé une licence non exchuive licence allowhg the exchuive permettant la National L i i of Canada to Bibliothèque nationale du Canada de reproduce, loan, distriiute or seU reproduire, prêter, distn'buer ou copies of this thesis in m i c m f q vendre des copies de cette thèse sous papa or electronic fonnats. La forme de microfiche/iih, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la proprieté du copynighî in this thesis. Neither the droit d'auteur qui protège cette thése. thesis nor substantial extracts h m it Ni ia thèse ni des extraits substantiels may be printed or otherwjse de ceiie-ci ne doivent êûe imprimbs reproduced without the author's ou autrement reproduits sans son permission. autorisation.

The orderly recruitment of motor units according to the size principle has been
show to apply to many different physiological conditions during muscle contractions.
However, exceptions to an orderly recmitment of motor units have been claimed during
muscle lengthening (eccentric) contractions by Nardone et al. (1989) and Howell et ai.
(1995) and have gained acceptance in the literature (Binder et al., 1996). According to these
authors, lengthening contractions involve a selective recruitment of large, fast-twitch motor
units with derecruitment of smail slow-twitch motot units. Careful examination of their data,
leads to questions about their interpretation. Therefore, the goal of this project was to study
the recmitment order of motor units in flexor carpi radialis during lengthening contraction of
wrist flexors. The subject contracted the wrist flexors isometrically against a background
load and recruited at least one discemible active motor unit. A lengthening contraction
followed this isometric phase. In the first paradigm a lengthening contraction was produced
when the subject yielded CO a constant Ioad and allowed the wrist to be extended. The second
paradigrn involved an additional ramp increase in load superimposed on the isometric
contraction producing a lengthening contraction or a step increase in load applied to the wrist
producing a very fast lengthening contraction resuiting in a stretch reflex. The results
showed a variety of motor unit recruitment and firing patterns during the lengthening phases
of contraction. However, no systematic selective recniiûnent was observed. The tonic lower
threshold motor units either changed their firing rate or were derecmited. If an additional
motor unit(s) was recruited during the lengthening contraction, it was the next higher
threshold unit and not a selectively activated large, high threshold unit. Our study further
supports the universality of the size principle of motor unit reccuitment.

DEDICATION
To my parents who give me al1 the encouragement, support and love a daughter needs.

1 would like to thank Dr. Parveen Bawa, first of al1 for the incredible
opponunity to do this research, as well as for her continuous patience and support dong the
way. I would also like to thank Dr. John Dickinson, Dr. Wade Parkhouse and Dr. Charles
Krieger for their assistance on this thesis. Thanks to my family for always king just a phone
cal1 away when 1 needed them. Finally, 1 would like to thank James, but words cannot
express how much 1 cherish him for his encouraging inspiration, unselfish support and most
of al1 for his endless love during this graduate degree.


Angular Displacement and Velocity ............................................................. 36
Angular Displacements and Mean Angular Velocities ........................................... 40 Constant Load hngthening Contractions .............................................................. 45
............................................................................................................ Preloads 45 . . Tension and Position ........................................................................................ 46 Single Motor Unit Responses .................................................................... .A6 Surface EMG ................................................................................................... 52 Unusual Recruitment and Firing of Single Motor Units ................................. 54 Cornparison of two types of Lengthening Contractions .................................. 55 Oscillatory Behaviour ...................................................................................... 56
Im@ Ramp Load Lengthening Contractions .................................................... 56 Preloads and Imposed Ramp Loads ................ .. ............... .. .... -56 . . Tension and Position ........................................................................................ 56 Single Motor Unit Responses ......................................................................... -57
................................................................................................... Surface EMG 62 Unusual RecnWtment and Firing of Single Motor Units ................................. 63
................ ..................*........................................*.... OsciHatory Behaviour .. 66
Im@ Step L o d Lengthening Contractions ................................................... 66
EMG Tremor ............................................................................................................. 68
............................................................. Summary d Motor Unit Firing Behaviour 68
............................................................................... Limitations OP the Sîudy. 1
Angular Displacement and Velocity ......................................................................... 73 Single Motor Unit Recruitment and Firing Pattern .............................................. 75
Comparison of the Two Types of Constant Load Lengthening Contractions .. 75 ................................................................................................. Constant Lods 76
Tmposed Ramp Loads ..................................................... .. 77 ............................ ....................................................................................... Imposed Step Loads -78
Surface EMG .............................................................................................................. 78
Input to the Motor Uniîs ........................................................................................... 82 .................................................................................................................. Conclusion 83
vii

LIST OF TABLES AND FIGURES
Table 1: Summary of the parameten generated for each motor unit and each individual ............................................................................................. leagthening contraction 34
............................... Table 2: Mean surface EMG for constant load lengthening contractions 53
Table 3: Mean surface EMG for imposed load lengthening contractions ............................... 62
................................................................................ Figure 1: Apparatus set-up 23
.............. Figure 2: Front view of gravitational load attached to the handle of the apparatus 24
....... Figure 3(A-C): Set-up for experimental recording of SMU. flexor and extensor EMG 25
Figure 3(D-G): Set-up for experimental recording of position. tension. toque load. preload. ....................................................................................... and trigger 27
Figure 4: (A) Experimentai set-up used to produce the position template (B) Example of ................................ position template of camp profile that subjects followed 29
Figure 5: Experimental set-up for generation of step and rarnp increase imposed load ................................. lengthening contractions produced by the torque motor 30
Figure 6: Set-up for data ................................................................................. 33
Figure 7: Drawing of exarnples of constant and imposed load lengthening contractions .......................................................... preceded by isometric contraction 37
Figure 8: Frequency distribution of angular displacement during constant load lengthening ..................................................................................... contractions 41
Figure 9: Frequency distribution of angular displacement during imposed load lengthening ...................................................................................... contractions 41
Figure 10: Frequency distribution of mean angular velocity during constant load lengthening .................................................................................... contractions 43
Figure 1 1: Frequency distribution of mean angular velocity during imposed load lengthening .................................................................................... contractions 43
.................................. Figure 12: Frequency distribution of total angular displacement 44
......................................... Figure 13: Frequency distribution of totai angular velocity 45
Figure 14: Tension and position traces of constant gravitational load lengthening contraction ......... iiîustrating biphasic pattern of tension during fast lengthening contractions 47

Figure 15: Typical resdts from constant load lengthening contractions paradigm showing ........................................................ silencing of tonic single motor units 48
Figure 16: Typicai results h m constant load lengthening contractions paradigm showing .................................................. continuation of single motor unit activity 50
Figure 17: Typicai results h m constant load lengthening contractions paradigm showing re- ..................................................... recruitment of tonic single motor units 51
........... Figure 18: Typical example of EMG during constant load lengthening contraction 54
Figure 19: Example of unusual pattern of recruitment during constant load lengthening ..................................................................................... contractions 55
......... Figure 20: Tension and position traces during imposed load lengthening contraction 57
Figure 21: Typicai results from imposed load lengthening contractions paradigm showing ................................................................ silencing of tonic motor units 59
Figure 22: Typicai results from imposed load lengthening contractions paradigm showing .................................................. continuation of single motor unit activity 60
Figure 23: Typicai results from imposed load lengthening contractions paradigm showing continuation of single motor unit activity over two different speeds ................... 61
Figure 2 4 Typical resuits h m imposed load lengthening contractions paradigrn showing ................................................ rec~itment of additional single motor units 62
........... Figure 25: Typicai exarnple of EMG during imposed load lengthening contractions 65
Figure 26: Example of recruitment of larger threshold unit during imposed load lengthening ....................................................................................... contraction 65
..... Figure 27: Typicai resuiis from step torque pulse imposed load Iengîhening contraction 67
..................... Figure 28: Example of accentuated tremor during lengthening contractions 69
..................... Figure 29: ExampIe of tremor during imposed load lengthening contraction 70

INTRODUCTION
Neucomusdar Control
Motor commands fiom the centrat nervous system are expressed through
changes in the magnitude of neural excitation of skeletal muscle. These changes in muscle
excitation generate force and movement, the magnitude and direction of which depends on
the properties of the muscle tissue and the mechanical loads experienced by the muscle.
Force control in a skeletal muscle is governed ùoeh by descending and reflex inputs
converging on motoneurons. Motoneurons integrate dl inputs (excitatory and inhibitory)
and, in turn, excite the muscle.
The totd force output of a siceletal muscle can be graded in two ways. First,
different numbers of motor units cari be remit& or derecniited. The second mode of force
regdation is achieved by changing the rate of discharge of individual motoneurons by rate
coding. Rate coding alter5 the force output of individual single motor units by using their
capacity to produce a partially fus& tetanus, in which the mean force output becomes
progressiveIy greater with increasing rate of motoneuron discharge. The total force of an
individual motor unit depends on the size of the motor unit and the rate of discharge. In
general, the relative contributions of rate coding and recruitment to muscle force output vary
widely from one muscle to another and depend on tbe types of motor units comprising the
muscle (Freund, 1983).

Mutor Unit a d the Motoneuron P d
A motor unit includes an alpha motoneuron and al1 ihe muscle fibres
innervatsd by it. M e n an alpha motoneuron discharges under normal circumstances, al1
muscie fibres of the motor unit contract, and a particular alpha motoneuron may participate in
a variety of reflexive and voluntary movements. Synaptic input h m a variety of sources
converge on the a-motoneuron where it semes as the last stage of integration to shape a
motor output to the muscle.
A given skeletal muscle is innervated by a set of alpha motoneurons that
comprise the motoneuron pool of the muscle. In the spinal cord, the somata of the
motoneurons constituting a motoneuron pool are arranged in longitudinai columns extending
rostrocaudally in Rexed's lamina IX of the ventral hom of the grey matter. The location of
different motoneuron pools is somatotopicaily relateci to that of their muscles, and
motoneuron pooh that innervate axial musculature tend to be located mediaily while those
innervating limb musculature are located laterdly (Brown, 198 1).
Alpha motoneurons of a pool Vary in size, biophysical properties including
input resistance, axon diameter, conduction velacity, and depth and duration of after-
hyperpolarization (Burke, 198 1; Henneman and Mendell, 1981). The relationship between
electrophysiological propetties of rnotoneurons and contractile responses of the muscle
fibres, which has k e n reported in several muscles throughout the body for hurnans and other
species, is described below and has implications for tecniitment patterns of motor units
(Burke, 198 1; He~eman and Mencieu, 1981). Srnalier, slower conducting motoneurons with
low rheobase values generaily innervate type 1 (SOG) muscle fibres. (The rheobase value is
the minimum current input to the motoneUron needed to elicit an action potential.) These

muscle fibres produce s m d tetanic forces and are fatigue cesistant. Small motonemns have
longer afterhyperpolarization which Limit these motonemns to fm at lower rates. These
motor uni& with long contraction times need very low rates to fuse, hence they are innervated
by motoneurons with longer afterhyperpolarizations. Medium diameter motonewons
innervate type IIA (Fûû) muscle fibres, which have medium to large tetanic force. Larger,
fast conducting motoneurons with high rheobase values tend to innervate type IIB (FG),
which produce very large tetanic force. Fast contracting motor units require higher firing
rates for fusion and are appropriately innervated by motoneurons with short duration
afterhypolarizations. The electrophysiologicai characteristics of the motoneurons that supply
slow and fast twitch muscle fibres differ and are, in part, responsible for the specific firing
patterns of these cells.
In order to classify motor units, Burke et ai. (1973) used the technique
introduced by Edstfim and Kugelberg (1969). The latter authors showed that by electrically
stimulating single motoneurons, it is possible to elicit tetanic contractions of muscle fibres
belonging to the same motor units. Prolonged tetanic activity of muscle fibres of a motor unit
depletes it of glycogen. Such fibres are distinguished from the rest of the muscle fibres by
the absence of glycogen in stained cross-sections of the muscle. Initidy, Burke (1967)
divided motor units into two groups: type S motor units are those that contain slow type 1
(SO) muscle fibres and type F motor units are those that contain fast contracting motor units
having type iI muscle fibres. Subsequently, the fast contracting motor units were
subdivided into two categones on the basis of theh susceptibility to fatigue: fast fatigue
resistant (FR) and fast fatiguable (FF) motor units, whose muscle fibres were found to be type
IIa (FOG) and type IIb (FG), respectively (Burke et ai., 1973).

The physiological and biochemical properties of motor can be divided
into distinct types, and the differentiation of motor units into types is helpfd in understanding
the neural control of motor units. However, in reality, motor unit properties show a
continuous variation in properties such as maximal tetanic force, isometric twitch contraction
time and amplitude, and fatigue tesistance (Freund, 1983), and a continuous spectrum of a-
motoneurons properties ranging from S to FF type characteristics exists (Henneman, et al.,
i974). The studies involving cat motor units and other animal models have provideci
fundamental information in regard to motor units. Using the pnnciples that are well
established in the cat motor units, information can be extrapolated to human rnotor units. Al1
properties do not transfer directly, but ovedl, it is fair to conclude that the organization of
motor unit populations in humans is basically the same as found in rnany mammalian
muscles (Burke, 198 1).
Muscle Activation
The central nervous system controls and monitors movement by transmitting
to and receiving signals h m muscle through an extensive neural network. Every cell of a
motoneuron pool is activated through a large number of spinal and supraspinal neural inputs.
Spinal inputs affecting motoneurons originate fiom muscle spindles, golgi tendon organs,
joint and cutaneous receptors, and spinal intemeurons while supraspinal pathways that input
ont0 the motoneurons include the corticospinal tract originating h m the cerebrai cortex, the
rubrospinal tract originating h m the red nucleus, the vesti%ulospinal tract originating h m
the vestibular nuclei and the reticuiospinal tract originating fiom the reticuiar formation.

Patterns of single motor unit activity in animals and humaas have provided
insight into the neuromuscular control of muscle. One such study led to the 'size principle*
of motor unit tecruitment which enunciated that to achieve a smooth graduai increase in
force, smail motor units that are normally composed of slow twitch fatigue cesistant fibres,
are recruited before large motor units that are usually composed of fast twitch, fatiguable
muscle fibres. Because most muscles are composed of motor units of heterogeneous
properties, the precise order of motor unit recmitment will greatly affect the output.
Recruitment of Motor Units
ûrderly Recruitment according to the Size tinciple
The 'size principle' proposed by Henneman and his CO-workers (Henneman,
1957; Henneman and Olson, 1965; Henneman et al., 1965, 1974) is the orderly recruitment of
motor units according to their size, i.e. small motor units have lower thresholds than, and are
thus recruited before, large motor units. Using recordings obtained h m smali numbers of
motor axons in fine filaments of ventral mot of decerebrate cats, the authors found that
motoneurons with srnail amplitude axonal action potentials were recmited fmt in response to
muscle süetch (Henneman et al., 1965). Muscle stretch increases excitatory spindle afferent
input to motoneurons. As the amount of stretch was increased, these small motoneumns
increased their firing rate as other motoneurons with larger amplitude action potentiais were
recniited. When the amount of stretch was decreased, motor axons showing the lowest-
amplitude action potentials wece the last to be derecruited.
Recruitment was found to be ordered according to the amplitude of the motor
axon action potential where motoneurons with smali action potentials were recniited before

motoneurons with large amplitudes. Since the amplitude of recorded axonal action potential
is directly related to the diameter of its axon, it can be postulateci that motoneurons are
recniited in order of increasing axonal diameter (Clamann and Henneman, 1976). Further,
axonai diameter is proportional to the toiai surface area of the motoneuron (Clamann and
Henneman, 1976), so it follows that motoneurons are recruited by excitatory inputs in order
of increasing size of the motoneunin. As weU, the axon diameter that varies with
motoneuron size, influences the velocity at which action potentials are propagated. Thus,
rnotor axonal conduction velocity was also found to be related to motoneuron size
(Henneman and Mendell, 1981; Bawa et al., 1984).
Additional studies revealed that such recruitment could be observed under a
variety of different activation conditions (Henneman et al., 1965). Excitatory and inhibitory
inputs to motoneurons from the peripheral and supraspinal systems were produced to test the
recruitment pattern of motor units. For a net excitatory input to a motoneuron pool, a
characteristic order of recniitment was always obsewed, and for an overall inhibitory input, a
systernatic deremitment of motor units was observed (Henneman and Mendeil, 198 1). The
size of the motoneuron, and thus the motor unit, is important in detennining its response to
synaptic influences. "As a result of many of the inherent properties of motoneurons, and
various inputs to the pools, orderly rec~iîment of motoneurons of skeletal muscles is very
robust" (Binder et ai., 1996).
Motor Unit Recruümeni in Humans
Human studies do not permit investigations into the synaptic and cellular
mechanisms of the size principle, but they allow one to approach the matter h m a functional
6

viewpoint. In this case, one monitors whether motor unit behaviow follows this principle
duriag natural muscle contraction under different voluntary conditions for different motor
tasks. Numerous experiments have been conducted on animal and human muscles, during
both voluntary and reflex contractions, to ascertain whether the size principle of orderly
recniitment applies. It has been shown to prevail with only a few possible physiologically
relevant exceptions reported to date (See Reviews: Henneman and Mendell, 198 1; Calancie
and Bawa, 1990; Cope and Clark, 1995; Binder et al., 1996).
Isometric voluntary contractions
The orderly pattern of recniitrnent was first shown to apply to humans during
isometric voluntary contractions of first dorsal interosseous muscle by Milner-Brown et al.
(1973b). The contractile twitch force generated by individual motor units was computed by
recording the motor unit potential with a selective needle electrode inserted into the first
dorsal interosseous muscle and recording the tension produced by the muscle. Spike
tciggered averaging was used to extract the twitch force that was correlatexi with each
discharge of the single motor unit. A nearly linear relationship was obsewed between the
twitch tension generated by the rnotor unit and threshold voluntary force level at which the
motor unit became active. This force thteshold is d i i t l y proportional to the total input to
the motoneuron pooi which in turn, is related to the net synaptic input arriving at each
motoneuron.
Similar recmitment patterns have been shown during slow voluntacy isometric
contractions of many human muscles including the extensor digitonun cornmunis (Mooster
and Chan, 1977; Riek and Bawa, 1992), the masseter and the temporaki (Yemm, 1977), the

flexor carpi radialis (Calancie and Bawa, 198!%), the flexor carpi ulnaris (Jones et al., 1993),
the abductor pollicis brevis (Thomas et al., 1987), and the extensor carpi radialis (Riek and
Bawa, 1992).
Direction-dependent motor unit recniiûmnt patterns have been reported when
a muscle is perforrning different voluntary tasks. Muscles including the first dorsal
interosseous (Desmedt and Godaux, 198 i; Thomas et al., 1987), the abductor pollicis brevis
(Thomas et al., 1987). and the biceps brachii (Ter Haar Romeny et al., 1984) have shown the
task group pattern of motor unit recruitment. These studies suggests that motor units tended
to be preferentially recmited for one task over another task. However, muscles with
identified task groups in which al1 the motor units contribute to a specific contraction, are not
exceptions to an orderly rec~itment of motor units. Using the extensor digitomm conununis,
recniitment of motor units within each individual task group was found to be according to the
size principle (Riek and Bawa, 1992). Extensions of the middle and ring finger were
periormed separately and together, as welj as extension of the wrist, and for each subgroup or
task group of the EDC motoneuron pool, motor unit recruitment thresholds were plotted
against their twitch tension and as with other isometric motor unit recruitment studies, an
orderly recruitment was seen for each task group (Riek and Bawa, 1992). A muscle once
thought to possess task groups, the flexor carpi ulnaris muscle was studied during isometric
flexion and ulnar deviation, CO-contraction of wrist flexors and extensors and non-isometric
flexion of the wrist (Jones et al., 1993). No task group was observed in FCU, all motor uaits
contributed to each of the four tasks, and orderly reccuitment was observecl in each of the four
tasks.
Grimby and Hannerz (1968) studied the recruitment order of motor units on

voluntary contraction of human tibia1 muscle using needle electrodes. They claimed that the
recruitment order of motor units is different in tonic and phasic activity. A rapid contraction
was said to be initiated by large unit and then sustained by a smdler unit that sustained
isometric contraction, They conc~udtxî that a human could selectively activate appropriate
motor units depending on the task (Grimby and Hannen, 1973). Desmedt and Godaux
(1977) did not find such selective recruitment during a more thorough investigation of tonic
and phasic voluntary activity. These authors studied pairs of motor units of the tibialis
anterior during fast baiiistic contractions as well as slow ramp contractions. The recruitment
threshold of motor units during ballistic contractions was show to be dependent on the rate
of change in muscle force. This phenomenon of high rates of change in force decreasing the
recmitment threshold for the motor units has been well documented (Tanji and Kato, 1973;
Desmedt and Godaux, 1977; 198 1). Due to difficulties in establishing a recruitment
threshold of units during ballistic contractions, the ballistic threshold of motor unit was
determined to be the mean force at which the motor unit always fired. The rank order of
pairs of motor units was established with this method and motor units were found to follow
an orderly recmitment. Reversais of recruitment were seen in about 11% of the trials, but
largely between units with similar recmitment thresholds.
Reflexve contractions
During slow voluntary activation, motor units are recruited and discharge
tonicaiiy. During reflexes produced by aimost syncbnous affecent inputs, a motor unit
responds phasicaliy, wirh genedy only one spike in response to the input. The single motor
unit activity during fast stretch reflexes imposed by toque motors was examined and the

orderly recruitment of motor units was obse~ed during the süetch reflexes in humans
(Calancie and Bawa, 1985a). Through observance of the surface EMG in the upper limb
muscles, a sudden stretch to a muscle results in two distinct periods of muscle activity: a
short latency response occumng approximately 25 to 50 rns after onset of stretch and a long
latency response occumng approximately 50 to 90 ms &et the onset of the stretch (Calancie
and Bawa, 1985a). The short latency response is thought to be mediated by the muscle
spindles via the spinal stretch reflex pathway wMe the long latency response is thought to be
mediated through suprapinal pathways. Both responses precede the voluntary response that
occurs after approximately 100 ms. Due to ihe probabilistic nature of motoneuron response
to phasic input, reflex responses are averaged over a large number of inputs. The magnitude
of the response is measured in terms of the number of discharges per input, which represents
the response probability (Calancie and Bawa, 1985a). It was shown that a motor unit's
response probability during both the short and long latency reflex periods was higher for the
lower threshold units than for motor units of higher recruitment thresholds. This suggested
that descending motor pathways and afferent fibres have a similar organization in terms of
inputs to the motoneuron pool. The probability of response was found to be dependent on
many properties including the level of facilitation of the motoneunin pool (background
activity), the strength of the imposed perturbation, and the preparatory set of the subject, yet
the recniitment order remaineci unchanged (Calancie and Bawa, 1985a).
Voluntary dynamic contractions
Voluntaq dynamic contractions incIude shortenhg (concentric) contractions
during which an active muscle shortens during movement and lengthening (eccentric)
10

contractions during which an active muscle is lengthened dwing movement. In comparison
to isometric contractions, there are relatively fewer studies of the motor unit rec~itment and
firing pattern during dynamic contractions. During dynamic contractions, variation in the
shape and amplitude of the motor unit action potential due to the relative movement of the
intramuscular electroâe with respect to the motor units that are recorded can increase the
difficulty in motor unit identification. It can also be difficult to ascertain whether the
observance of the onset of an action potential is a result of recmitment of a new unit or due to
delayed recording of an already active unit because of movement of the electrode during the
change in muscle length.
The orderly recruitment of motor units according to the size principle has ken
shown to occur during human voluntary dynamic movements. Thomas et al. (1987) observed
28 motor unit pairs from the first dorsal interosseous and 9 motor unit pairs from the
abductor pollicis brevis during a repetitive opening and closing of scissors movement. The
relative rank order recruitment thresholds for pairs of motor units observed during the
dynamic movements was determined through cemitment during voluntary isometric
contraction of the units. Motor uni& had recruitment thresholds up to approximately 50%
maximum voluntary contraction. Motor unit recruitment order, according to increasing
twitch tension amplitude, was preserved during repetitive dynarnic movements. Four pairs of
rnotor units (14%) from first dorsal interosseous and two pairs (22%) from abductor pollicis
brevis, showed recruiîment reversais for more than 50% of scissor movements. However,
these unit pairs showed similar recruitment thresholds. Therefore, motor unit recniitment
was predominantly ordered by size during repetitive scissor movements, with signincant
number of reversais for units of simiiar thresholds.

in addition to firing of motor units of similar thresholds, hi& rate of reversals
have been attributed to the pbasic nature of contractions l i e the scissor movement performed
in the study by Thomas et ai. (1987). Phasic contractions that involve a fast and large input
to the motor unit and result in a brief discharge of a small number of action potentiais can
lead to revenais because of the prababilistic nature of response, In a later study using the
fmt dorsal intemsseous muscle during tonic functional tasks, Jones et ai. (1994) observed
less than 10% of recruitment reversals. An orderly recruitment of motor units was show to
occur during these tonic contractions involving abduction of the index finger, rotation of a
knob and pincer grip of a clip. Tonic contractions where the increasing torque over the
different functional task contractions was canied out over relatively longer periods showed
significantly less reversals. When an input to the motoneuron is phasic, reversals have a
higher probability of occurring due to noisy input to the motoneuron. Because of this noise,
reversals are more likely between motoneurons of similar threshold, and less probable
between units with a large difference in thresholds. A third factor which can contribute to
reversals between motor units of different sizes is the axonai conduction velocity. Desmedt
and Godaux (1977) attributed the reversals they found during ballistic contractions to
differences in conduction velocities of units with similar recniitment thresholds.
Using intrmuscular EMG recordings of the biceps brachii, Moritani et al.
(1988) observed the activity of motor units during shortenhg and lengthening contractions.
Both types of contractions involved 120 degrees of movement, h m 30 and 150 degrees for
shortenhg and lengthening contractions, respectively, with full extension at O degrees in 5
seconds agaiast a 3û% maximum voluntary contraction quivalent load, Greater
motor unit activity (as estimated h m population EMG) and the presence of motor units with
12

relatively large spike amplitudes were seen during shortening contractions in comparison to
lengthening contractions at three separate positions (45,90, 135 degrees). The data showed
that lengthening contractions were associated with less motor unit activity as demonstrated by
recniitment of relatively smail spike amplitude motor units and less marked rate modulation
compareci to shortening contractions. Smaller spike amplitudes and lower spike frequencies
were seen at ail three angles, although only 2 out of 3 joint angles of each property were
significant. The recruitment pattern of the motor units indicated that different threshold
motor units were usually recmited in order of increasing spike amplitude even at higher
levels of muscle contractions.
Sogaard et al. (1996) and Kossev and Christova (1998) studied specifically the
motor unit recmitment patterns of the biceps brachii during shortening and lengthening
dynamic contractions. Sogaard et al, (1996) used loads of approximately 10% MVC. A total
of 1 19 motor units were identified during dynamic contractions at mean angular velocities of
10 degrees per second and were examined for recruitment pattern. Several contractions of 20
degrees per second and 40 degrees per second were perfonned with 30 and 9 identified motor
units, respectively. ûver 60% of the identified motor units were found to be active in both
shortening and lengthening contractions for each velocity. Further, no significant difference
in amplitude, duration or area of the motor units were found between the set of motor units
that were active only during the shortening phase, lengthening phase or both phases. The rate
modulation was deemed the important difference between shortening and lengihening
contractions and not the selective m i t m e n t of motor units during each typ of contraction.
Mean fiting rate of the motor units decreased ~ign~cantly when the contraction changed
h m a shortening to a lengthening contraction, and this lower mean firing rate during
13

lengthening contractions has also been found in other studies on decerebrate cats (Cordo and
Rymer, 1982) and in humans (Tax et al., 1990).
Using the biceps brachii muscle, Kossev and Christova (1998) identified 45
motor units during shortening and lengthening contractions. The subjects performed
contractions of slow (4.94 I 5.3 Nmis) and fast (9.20 110.05 Nmls) velocities with a mean of
24.96 14.30 % maximum voluntary contraction and a high limit of 52 % MVC for the
relative workloads. Al1 but one of the motor units that they investigated were active during
both the shortening and lengtbening contraction phases as weil as during isometric
contractions. The one motor unit that was active only during the shortening and isometric
contractions was derecruited pcior to the lengthening contraction. An orderly recruitment and
detecruitment of motor units with respect to their isometric remitment threshold was seen
although they observed thai the recruitment threshold during the shortening phase was found
to be lower than the detecruitment threshold during the lengthening phase of particular motor
units. Generally, the derecruitment of motor units during a lengthening contraction was the
reverse of the recruitment during the shortening.
AUernative Patterns of Motor Unit Recruitnient
While some reversais in recruitment order are seen in nearly al1 experirnents
focused on the recmitment of motor units, orderfy recruitment of motor units has been
generally observed (Henneman and Mendell, 1981; Cope and Clark, 1995) in a variety of
different studies. However, some sudies suggest deviations k m the size-ordered
remitment under specific conditions. These studies include electricd stimulation of
cutaneous afferents (Kanda et ai., 1977; Garnett and Stepheas, 1980,198 1; Masakado et ai.,
14

L991) and of muscle afferents (Davies et ai., 1993). As well, a few studies have claimed
lengthening contractions as possible exceptions to the orderly recruitment of motor units
(Nardone et ai., 1989; Howell et al., 1995).
Studies using cutaneous input to the motoneuron have proposed exceptions to
the size principle. in decerebrate cab, it was shown that cutaneous input from the sural nerve
superimposed on stretch reflexes or tendon vibration refiexes tended to decrease the firing
rate of relatively low threshold medial gasûacnemius motoneurons while increasing the
excitation of the higher threshold motoneurons (Kanda et al., 1977). The larger motoneurons
were said to receive a pa ter amount of excitatory input while the smaller motoneurons
received a proportionally greater amount of inhibition. With this activation pattern, large
motoneurons would be recniited in preference to the small motoneurons, and such a
recruitment scheme could be considered an exception to the size principle.
Clark et ai. (1993) attempted to conficm the findings under similar conditions
in the decerebrate cat. The study found a smdl percentage of medial gastrocnemius motor
units with slow axonal conduction velocity exhibited a slowing of their firing rates during
cutaneous input produced by skin pinch. However, when pairs of motor units were recniited
purely by tendon vibration, or skin pinch or elacîrical stimulation of the sural nerve,
recruitment order was the same and the lower tbreshold unit had the slower conduction
velocity in 90% of cases. Thus the size principle recmitment scheme was used during both
peripheral Ia and cutaneous input to the medial gastrocnemius motoneuron pool.
In humans, cutaneous stimulation has also been used to demonstrate the
possibility of exceptions to the size principle (Stephens et al., 1978; Gamett and Stephens,
1980,1981). in this case electncai stimulation of the digital nerves of the index figer
15

demonsûated tendencies toward inhibition of f ~ n g of lower threshold units and facilitation
in higher threshold units. The physiological relevance of electrical stimulation bas been
questioned because normal tactile stimulation of the index finger did not produce disorderly
m i n n e n t of motor units (Cope and Clark, 1995). Fwthermore, since this change was
observed after 5 minutes of continuous firing of motor unit, the question of fatigue may
complicate the explanation of observed pattern.
Lengthening Conîmcîions as Possible Excepiions to Orderly Recruitment
Contrary to the comprehensive evidence suggesting that motor unit
recruitment occurs according to the size principle, possible exceptions have been reported.
As previously mentioned, some investigators have implicated lengthening contractions to
involve an alternative recruitment pattern of motor units where large fast-twitch motor units
are recruited selectively instead of small slow-twitch motor units.
In 1989, Nardone et al. recorded the single motor unit activity of the soleus, medial
and lateral gastrocnemii muscles, as well as the position signal during isotonic contractions.
Using 15-20% maximum voluntary contraction loads, the subject either maintained an
isometric contraction, performed a shortening contractions in which the ankie was extended
through shortening of ankle extensors, or performed a lengthening contractions in which the
ankle was flexed through lengthening of active ankie extensors. The velocities of the
dynamic contractions ranged h m 10 to 70 degrees per second. Approxirnately 70% of the
identified motor units were reported to be active exclusively in either the shortening or the
lengthening phases. The motor units active exclusively during Iengthening contractions bad a
higher mean isameiric recniitmeat threshold (55.2 I 6.1 Nm) compared to the motor uniîs
16

that were recruited exclusively during shortening contractions (34.8 I22.6 Nm) and the motor
units tbat were active during both the shortening and lengthening phases (27.2 I Nm). The
authors claimed that during the lengthening contractions, high threshold, fast twitch large
motor units were recruited selectively as low threshold, slow twitch small motor units were
preferentially derecmited.
Nardone et al. also reported an increase in EMG activity during the
lengthening contractions. They argued that the larger magnitude of surface EMG during
lengthening contractions was due to recruitment of very large motor uni& wfiich not only
compensated for the derecruitment of small motor units, but resulted in an overail increase in
EMG activity during the lengthening contractions. This was used as additionai support for
their theory of selective recruitment of large units.
The relaxation times of the motor units were presented as a plausible argument
to support the reported selective recruitment of fast motor units over the slow motor units
during rapid lengthening. They hypothesized that lengthening contractions perfonned by the
derecruitment of the fast conducting-fast relaxing large motor units of the agonist muscle
would be more advantageous than the derecruitment of slow conducting-slow relaxing motor
units, since the force developed by the large motor units would terminate faster. They also
suggested that the use of fast-twitch units with their fast relaxation times is necessary for
control of fast lengthening contractions (Nardone et al., 1989). However, tbis rationale seems
questionable for constant load lengthening contractions, for in order to yield to a lad, a
decrease in the force generated by a muscle is required. It would seem inefficient for the
centrai nervous system to first recruit large motor units which generate large amounts of force
when the goai of the muscle is to obtain a decrease in force.

Investigation by Howeii et d.(1995) of the single motor unit activity of the
fmt dorsal interosseous was petformeci during sinusoidal shortening and lengthening
contractions. They reprted selective recniitment chuing lengthening contractions of 3 out of
21 (14%) motor units studied. From this 14 % pattern, they concwred with the hypothesis of
Nardone et al. (1989) that there is a selective reçruitment of motor units during lengthening
contractions. The 3 motor units were said to be recruited when other units were seen to be
decreasing their discharge rate or were king derecniited. However, their results do not
clearly show a pattern of selective recruitment. First, they observed an alternative
recruitment strategy in only 14% of their units and secondly, these units had relatively low
thresholds with respect to the motor unit population. Secondly, their figures illustrate that
two of the three motor units fimd with smaller threshold units fung in the background.
Thecefore, only one of the motor units had an unusual firing pattern. Furthemore, the
lengthening contractions of their subjects were not smooth, but dispiayed imgularities in
their position traces. The position traces showed clear oscillations during the lengthening
contraction, which implied that the lengthening contractions were superimposed by ballistic
shortening contractions. The ballistic contractions of the subjects could have resulted in
bursts of additional higber threshold motor unit activity during the lengthening contraction.
Importance of Lengthening Contractions
iengthening contractions occur when a load is applied to an active muscle,
which is greater than the force generated by the muscle. The source of the load developing
the extemal force could be gravity, the muscle action of an antagonist muscle p u p or an
externaüy applied load. General functions of lengthening contractions include deceleration
18

of body segments, increased mechanical advantage, spring-lie functioas, and shock
absorption. Decelention happens in throwing, where deceleration of the forearm by a
lengthening contraction of the elbow flexors is essential to prevent elbow injury; and durhg
running in which a lengthening contraction of the hamstrings decelerates the swinging leg.
Shock absorption is achieved by the lengthening quadriceps femoris at foot strike during
w u n g and running or when landing from a jump. Spring-like functions include the storage
of elastic energy during the lengthening contraction and subsequently transmitting it to a
shoriening contraction dunng a jump such as when the hamsûings lengthen to lower the thigh
in flexion in preparation to jump and then the hamstrings shorten to raise the thigh in
extension. In conclusion, lengthening contractions are relevant physiologicaliy and hence an
understanding of the types of motor unit firing during this phase is essential.
Objectives of the Thesis
Conflicting evidence exists in studies involving the f i n g pattern of motor
units during lengthening contractions. Some studies have shown an orderly recniitment of
motot units fmm smail to large during lengthening contractions (Thomas et ai, 1987;
Moritani et al., 1988; Sogaard et al., 1996; Kossev and Christova, 1998) while studies by
Nardone et ai. (1989) and Howell et d. (1995) suggest selective recmiûnent of large fast-
twitch motor units accompankd by dececruitment of small slow-twitch motor units. This
'evidence'bas gained acceptance in the litetature (Bin&r et al., 1996). The data in the studies
of Nardone et al. (1989) and Howell et al. (1995) are not convincing as îhere data can be
interpreted differenily.
Therefore, tfie goal of this pmject was to examine motor unit d t m e n t and

firing pattern during lengthening conttactions. A comprehensive investigation involving
lengthening contractions of varying background activity, velocities and loads was perfomed
with the human flexor carpi radialis in order to characterize the motor unit recmitment and
firing pattern during lengthening contractions. Our hypothesis was that motor unit
recruitment during lengthening contractions would follow the six principle of orderly
recmitment. The size pnnciple has been shown in virtually al1 types of muscle contractions
and the evidence for selective recmitment during lengthening contractions is not convincing.
To constitute a systematic pattern by the central nervous system for selective recmitment of
large motor units, clear and consistent evidence would have to be obtained during the
lengthening contractions. This evidence would have to include repeated trials with
unarnbiguous selective mmitment of a high threshold unit(s) with distinct derecmitment of a
low threshold motor unit(s).

Generai Overview
To examine the recruitment of motor units during lengthening contractions,
subjects performed initial isomeûic contractions followed by lengthening contractions of
wrist flexors at various velocities. The first paradigm involved constant load lengthening
contractions either with constant gravitational loads produced by attaching weights or with
constant cunent loads produced by a toque motor. The subject conuacted the wrist flexors
isometrically against a background load then decreased the tension generated by the muscles
so that the constant load extended the wrist joint, thus producing a lengthening contraction of
the active muscles. The second paradigm involved imposed load lengthening contractions
which included camp increases and step increases in load. Again, the subject began by
contracting the wrist flexors isometrically against a background load. An additional ramp
increase in load at various velocities or a step increase in load was applied to the wrist joint
while the subject tried to maintain aconstant contraction in the muscle. The additional load
extended the wrist producing a lengthening contraction.
Subjects
The experiments were conducted on the wrist flexors of six volunteers (2 male
and 4 female). Subjects were right-handed with no history of neurological disease and ranged
in age ftom 22 to 55 years old. The experiments were approved by the Ethics Cornmittee for
Human Experiments at Simon Fraser University, and each subject signed the subject consent
fom.

A ~ p a i 9 -
The subjecîs were seated with their cight foreatm rested on a padded
horizontal plaîform. A vertical handle secured against the metacarpophalangeai joint of the
subject's palm was positioned 8-10 cm away from the rotational axis of the wrist. Secwing
the subject's hand to the handle was done to ensure relaxeci fingers during wrist extension. A
horizontal bar equipped with strain gages connected the handle to a vertical shaft of a
precision torque motor (Aeroflex TQ 82W), and h e axis of wrist rotation was coaxial with
that of the toque motor shaft. To prevent lateral movements of the wrist, two padded clamps
were positioned proKimal to the wrist. Figure 1 illustrates the apparatus from a lateral view.
To achieve a constant gravitational load, weights were attached to the handle by way of a
pulley. Figure 2 illustrates the apparatus from the front with the gravitational load set-up.
Data Recordhg
Single motor unit (SMU) electrical activity was recorded from the flexor carpi
radialis (KR) muscle using bipolac intramuscular microelectrodes. These microelectrodes,
which were consmcted in the laboratory, consisted of two 30-50 pm HML coated stainless
steel wire (California Fine Wire Company) inserted into the shaft of a 25 gauge needle and
held in place with epoxy. The microelectrodes were sterilized with rubbing alcohol and
inserted into the FCR. The SMU activity was preampmed (Grass Pl5 AC) with a gain of
100 and filtered with a band pas of 100 Hz - 10 kHz, then filtered with a AC conditionhg
amplifier (100 Hz high pas). The experimentd set-up for the FCR SMU is shown in Figure
3A. The S M ü activity was monitored on an oscilloscope, and an audio speaker for subject
audio-visual feedback. It was recorded on a video cassette recorder (VCR) tape using a

;train Gauges for Tension
Potentiometer - for Position
FExtensor EMG
Torque Motor
Figure 1: The appam~s set-up for aording the FCR SMU activity, the flexor surface EMG, the extensor surface EMG, the tension, and the position.

e Figure 2: Front view of gravitational load attachcd to the handle of the apparatu during constant load lengthening contractions.
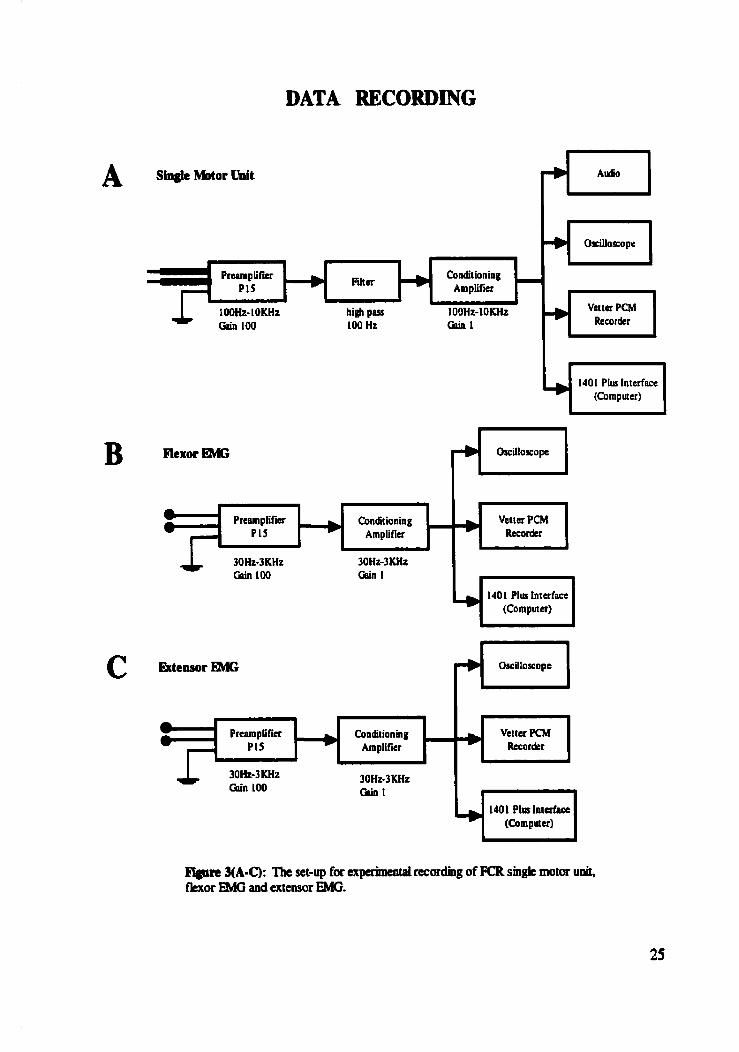
DATA RECORDING
30Hz-3KHz Gain LOO
Conditioning Amplifier
- 1401 Plia Interface
{Computer)
Oscilloicopc
I,
-
30Hz-3 KHz Gain1
1401 Plus tnterface
Orilloscopr?
I ~ H z - ~ O W 100 Hz GPinl
Conditioning Amplifier
Rgure %A-C): Tht set-up for expetimentai recordhg of FCR sin& motor unit, k o r FMG and extensor EMG.
Veiter PCM Record#
Vetia PeM Recorder
Prenmpiitier PIS
I,
Conditionhg Amplifier .
Vetlet P a Recordcr

Vetter mode1 4000A pulse code modulator 0 recorder.
Surface electromyographic (EMG) activity was recorded fmm wrist flexon
and extensors (extensor carpi radialis). The skin was prepared by shaving a patch of skin
over the muscle and cleaning it with alcohol. Two 9 mm disc Ag-AgCI electrodes with
electrode cream (Grass EC2) were then secured approximately 2 cm apart on ttie skin over
the muscle ôelly. The signals were fed to a Grass PL5 AC preamplifier where it was
amplified and band passed filtered at 30 Hz - 3 kHz and recorded on a VCR with Vetter PCM
recorder. It wlis viewed simultaneously on an oscilloscope and on the cornputer screen using
a 1401 PLUS Cambridge Electronic Design (CED) interface. The EMG of the wrist extensors
was monitored to ensure the movement of the wrist was accomplished by only the stretch of
the active wrist flexor muscles, and not by activation of the extensors. Voluntary activation
of the extensors would recipmally inhibit flexors and prevent any new recruitment. Figures
3B and 3C illustrate the experimental set-up for flexor and extensor EMG, respectively. A
ground electrode ;vas secured to the right upper arm of the subject.
The angular displacement of the wrist was transduced by a potentiometer
attached to the shaft of the torque motor. The wrist position signal was sent io an amplifier
and then to the Vetter PCM interface for recording, and to an oscilloscope for viewing online.
Figure 3D illustrates the experimental set-up for position. To provide position feedback for
the subject during lengthening contractions, the wrist position was displayed on an
oscilloscope at a slow sweep speed, and the subject was asked to follow the position trace
template provided on the oscilloscope. Three different position templates were coostructed
using a fiitered square pulse generated by a Grass Sa8 stimulator to produce slow, medium

DATA RECORDING (conti)
E Tension
Orillorope for Subject Position
Potentiometer
,
Vetter PCM Recorder
Oscilloscope P--'
DC
Conditioning Ampüfur
I )
- 1401 Interface
amputer
Vetter PCM Recorder
1401 Interface amputer
Bridgc Amplifier
Fi* WG): The sa-up for recordhg of position, teasion, torque bad p W and uigger.
Dc
Vetter PCM Recordrr
ûasr 918 Simulntor
Gnrr 918 3irnuintor
- II)
Conditionhg , Amplifut
Couditionhg - Vettet PCM A m p ü f i kcordcr
Conditioning A m p l f ~ r
DC
v

ami fast velocities. Subjects were not required to trace the template exactly, but were asked
to use it as a guide to produce lengthening contractions of different velocities. The
experimental set-up and an example for the position trace template are shown in figure 4.
To mesure tension produced by the wrist flexors, strain gauges (EA-13-
250MQ-350) were mounted on the horizontal bar connecting the handle to the toque motor.
The tension signal from strain gauges was fed to a bridge amplifier (Vishay Instrument, DC - 1000 Hz band pass) then displayed on an oscilloscope and recorded on a VCR (Vetter PCM
recorder). Figure 3E illustrates the experimental set-up for tension.
The precision torque motor driven by a servo-amplifier was used to provide
resistance to wrist flexion through a background load (preload) during the isomeüic
contractions preceding the constant current load and imposed load lengthening contractions.
The prebad produced by the torque motor was used to control the strength of contraction of
the FCR by activating one or several tonic firing of motor units. Figure 3F illustrates the
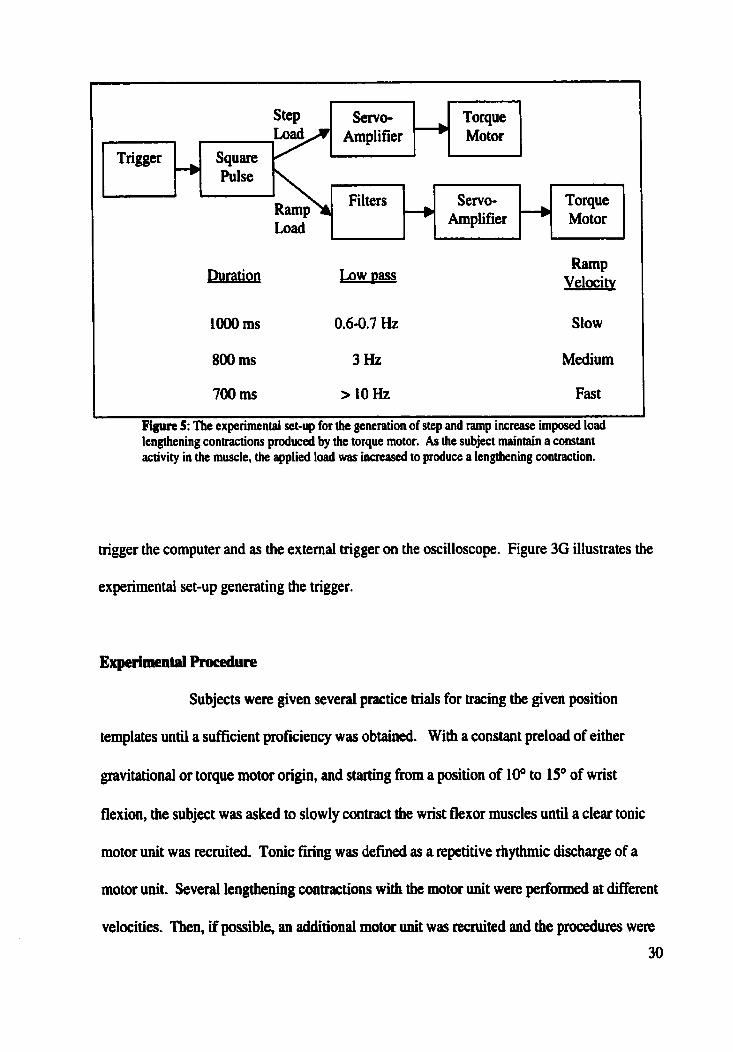
experimental set-up for producing torque load. The torque motor also provided the ramp and
step increase in load for the imposed load lengthening contractions and step pulse imposed
load lengthening contractions, respectively. The ramp and step increases in load were
achieved through a triggered square pulse generated by a Grass S88 stimulator that drove the
servo-amplifier of the torque motor. The square pulse was sent directly to the setvo-ampiüïer
for the step load and was low pass filtered at different ftequencies to obtain different rates of
ramp increases in toque load. The details of the experimental set-up for producing ramp and
step increase in loads are show in figure 5. A trigger generated by the Grass S88 stimulator
was used to initiate the ramp and step increase in load of the imposed load lengthening, to

Velociîy
Slow
Medium
Fast
Square Puise
Low uass
0.7 Hz
1.1 Hz
3.0 Hz
Figure 4: (A) The experimental set-up used to m u c e Ihe position template. (B) An example of a position kmplate of a ramp profite that subjects followed voluntarily during constant load lengthening conaactions. The solid line reprcsents the continuous position template that the subjects followed and the dotted line npresents the subject's position signal. The oscilloscope divisions were 200ms horizontally and 05 volts vertically.
,, Filter Position Trace Profle on Oscilloscope
S tep Servo- Torque Amplifier Motor
Trigger Square + Puise
Filters Servo- Torque -b Amplifiei Motor
Duration b w pass Rame Velociiy
1000 ms 0.64.7 Hz Slow
I 800 ms 3 H z Medium
I 700 ms > 10Hz Fast L
Fipn 5: The experimenial set-up for hc generation of step and ramp increase imposed load lengthening contractions produced by the torque motor. As the subject maintain a constant activity in the muscle, the applied load was increased to produce a lengthening contraction.
trigger the computer and as the extemal trigger on the oscilloscope. Figure 3G illustrates the
expecimental set-up generating the trigger.
Experimental Procedure
Subjects were given several practice trials for tracing the given position
templates until a sufficient proficiency was obtained. With a constant preload of either
gravitational or torque motor origin, and siarting h m a position of 10" to 15' of wtist
flexion, the subject was asked to slowIy contract the wrist flexor muscles until a clear tonic
motor unit was recruited. Tonic firing was &fined as a repetitive rhythmic discharge of a
rnotor unit, Several lengthening contractions with the motor unit were perfonned at different
velocities. Then, if possible, an additional motor unit was recruited and the procedures were
30

repeated. The strength of the preload and of the ramp loads was adjusted for clacity in the
motor unit responses. Frequent rest periods were given to the subject to reduce the
probability of fatigue.
The details of experimental pmdigms performed with audio-visual feedback
of the FCR SMU activity and visual feedback of position are:
1. Constant load lengthening contractions. The subject was asked to hold an
isometric contraction with wrist flexors against a preload. When the motor unit firing was
steady, a position profile template was pmented on the oscilloscope. Attempting to match
the position template, the subject stopped resisting the load such that the load extended the
wrist at the approximate velocity of the position template. Tbree different velocities of
lengthening contractions were performed with approximately ten trials of each condition.
The constant load that was held by the subject was either provided by gravitationai weights or
by a constant current fed to the torque motor. In the latter case, even though the current was
constant, the load during wrist extension was not. For these experiments, genenlly, quite
high threshold units were studied. Because of friction in the system, a small preload would
not extend the wrist. High velocities of lengthening contractions could be achieved only with
large preioads, and hence, higher threshold units.
2. Impmed load lengthening contractions. The subject performed an isomeûic
contraction against a constant background load provided by the torque motor. Then an
additionai ramp load was imposed on the background load. During the imposed load, the
subject w;is asked to try to maintain a constant level of motor unit activity in the muscle
throughout the lengthening conttaction. Because the increased load was not matched by
increased muscle contraction, the increase in load impsed a lengthening contraction on the
31

subject. The dope of the tamp load was changed which led to a range of corresponding
angular velocity of lengthening contractions. A step Uicrease in load produced a very fast
lengthening contraction by stretching the active muscle very quickly. Step loads used for
these experiments were very small, and hence produçed very small wrist extensions. Small
step loads were essential for recording discemible motor unit activity.
Data Analysis
Dara analysis were dune off-line on a Pentium 200 PC computer. The data
recorded on VCR cassette was transferred to ihe computer wich the Cambridge Electronic
Design (CED) 1401 Plus computer interface and Spike2 software. The SMU activity, the
position, the tension, the flexor EMG and the extensor EMG were filtered, arnplified and
outputted to tfie computer to be captured on separate analog (waveform) channels. Motor
unit activity, discriminateri by two in series BAK window discriminators, produced transitor-
transitor logic (TïL) puises corresponding to targeted motor units. The 'ZTL pulse for the
targeted SMU and the trigger for the computer were recorded on wparate event (+) channels.
Figure 6 illustrates tbe set-up for data analysis. The SMU analog and the SMU-TTL data
were compared to ensure correct discrimination of the action potentid. TIL pulses from
action potentials that were missed by ihe discriminator were added and extraneous TTLs
were deleted h m the event market record.
In order to examine the behaviour of rnotor unit firing during kngthening
contractions, several parameters of the motor uaits, lengthening contractions and paradigms
were generated using scripts written for the SPKE2 software. Table 1 surnmarizes the
parameters that are subsequently described in the text.

DATA ANALYSIS
Oscilloscope a
Y 1401 Plus Interface 30Hz-3 KHz Cornputer I
1401 Plus Interface Cornputsr
Oscilloscope
2 BAK
Vetter PChl Recorder
Fienor W I) Oscilloscope
Pbsition ~sciiloscope
Vetter PCM Recorder Amplifier
1401 Plus lnterface Computer
Conditionhg Amplifier
-
k
Vettw PCM Recorder
IOOHz-IOKHz
üi i iminntors
Figure 6: The set-up for dûta analysk of single motor unit, SMU T lL extensor EMG, position and tension.
-
1401 Plia Interface 30Hz-3KHz Cornputer
-
Tension I) Osciiioscope
140 1 Plus tnterface Cornputer
Vetter PCM Recorder
Conditioning Amplifier
- h
1401 Plus Interface DC Cornputer
- 4 Conditionhg Amplifier

Table 1: Summary of the parameters generated for each motor unit and for each individual lengthening contraction.
. .
MUTA Motor unit isomeûic recruiûnent threshold
1 Isometnc relative recniitment order I
1 1 Mean angular velocity 1
Lcn*ning contraction
Motor Unit Triggered Average: During movement, the geornetric relationships between the
muscle fibres and the needle electrode can change and, thus, affect the waveform of a motor
unit (Desmedt, 1% 1). SMU action potential shape and size were not always completely
reliable in consistently tracking and identifying a particular SMU. In order to track a motor
unit throughout the lengthening contraction and distinguished it from other moior units which
may have a similar shape, a motor unit triggered average (MUTA) was generated using spike
triggered averaging between SMU-ITL pulses and the unrectified surface flexor EMG. The
MUTA of an SMU stays constant while the shape of rnicmlectrode recorded SMU potential
frequently changed with movement. This aided in identiwng the single motor unit
throughout the lengthening contraction.
Motor unit ncniitment and firing pattern Mean rectifiai surface EMG activity
, ~otai cimges in tension Angular diplacement
Isometric Recruitment Ordec The absolute isometric recniihnent threshold for a motor unit
was determined using isometric voluntary contraction. Starting with a relaxed muscle, the
subject recruited single motor units by slowly increasing the isometric tension. The level of
tension at which a motor unit dischargeci tonically was defined as its isometric cemitment
threshoid. The absolute recruitment thresholds values were calculaîed as tension values and
34

as a percent of the maximal voluntq contraction (5% MVC). The absolute isometric
recmitment threshold was unable to be determined for many rnotor units. In sucb cases the
relative isometric reccuitment order was determined for a pair (or group) of motor units. This
included classiQing the motor units as lower or higher threshold unÎts according to the
relative rank order in which they were recruited during the isometric contraction.
Motor unit recnritntent andfiring pttern: Numerous experiments have established the
orderly recruitrnent of motor units according to the size pn'nciple during isometric
contractions (Milner-Brown et al. 1973b; Henneman and Mendell, 198 1). The corollary is
that if recniitment (or derecruitment) of motor units during lengthening contractions mimics
that of isometric contraction, Iengthening contractions can also be said to foIlow an orderly
recruitment according to the size principle. Establishing the isometric tecruitment order
allowed for cornparison to the lengtheaing contraction recruitment, and thus characterization
of the motor unit recruitment and firing patterns during the lengthening contraction. When
the absolute nor relative isometn'c recruitment order of the motor units could not be
established, the lengthening contractions could not be used because the recruitrnent strategy
for lengthening contractions could consequently not be detennined.
Surface EMG: Quantification of the surface EMG was performed to d o w for cornparisons
of population fiexor EMGs between the isometric and tengthening contractions. The flexor
surface EMG activity was quantifiai by rectifying the EMG signal and dekrmining the mean
integrated EMG values of 200 ms before and 200 ms after the start of the lengthening
contraction. For each motor unit king snidied, correspondhg mean integrated EMG values
35

were averaged for one type of lengthening contraction. Cornparison between tbe mean
integtated EMG values for the two intervals were made using a two-tailed, paired t-test with
a significance ciiffierence reported at a level of p 5 0.05.
Total changes in tension: D u h g lengthening contractions, there was a change in tension
values. Figwe 7 shows the tension trace for a typicai example of both the constant Lod and
imposed load lengthening contractions and illustrates the direction of change in tension
during constant load and imposed load Iengthening contractions. The totai change in tension
started at To and ended at Tm~.
Angular displacement und velocify: Figure 7 illustrates the position values used for angular
displacement and velocity for the constant load and imposed load lengthening contraction.
The angular position values at the start of lengthening contraction (Po), after 100 ms (Piw),
200 ms (PZoo), 500 ms (Pm), and at the end of the lengthening contraction (PEND) were
obtained to calculate the angular displacements at respective time interval. For constant load
lengthening contractions, the start of initial decrease in tension signifîed the start of the
lengthening contraction, and for imposed load lengthening contraction, it was the start of the
increase in tension. A total lengthening contraction was defineci h m the initiai change in
tension until the last change in position. The mean angular velociity of tbe wrist was
calculated using the angulu displacerrient of the wrist divided by change in time. To
compare lengthening contractions of different duraiions and Iengths, the angular velocities
were cdculated after 100 ms, 200 ms, 500 ms and for the totai Iengthening contraction. The

I I
Tension I I 1 t I I I I I 1 I
Position I I I
I Tension I
I
I I 1 I I I
Position I I I
PO! ey x
I
! Extension
Flexion
Extension
Flexion
Eîgarc 7 Drawing of typical examples of consiant a d im@ Ioad lengthening contiactions pceceded by an isomeaic contraction. The position values at the times (t) indicated are used CO caldate ihe angular displacement after lOOms (PIm- Po), 2lXlms (Pm- Pd, 500ms (Pm- Po), and the total position change of the lengthening contraction (Pw> - Po). They are a h used ta calculate the mean angular velocities a i k 100nis ((Piao-P0)Ço.l~) 200ais ((Pm- Pm.2s), HlOms ((Pm- Po}105s), as well as the mcan angular velacity of the total lengthening conaacricm ((PEND-Po)/(tm- ta}). Tbe iod change in tension is detennined by Tw - TO.

range of the velocities of lengthenhg contractions is important in establishing cornparisons of
velocities to other studies.

Ail lengthening contractions were preceded by an isometric contraction. At
least 1 tonicaily firing motor unit, and up to as many as 5 discemible single motor units, were
held f h g in the background during the isometric phase before the start of the lengthening
contraction phase. For each set of lengthening cantractions, one motor unit action potential
was targeted and tracked throughout the set by discrimination to a Tmt pulse. However, the
recruitment and firing pattern of al1 discemible motor units recorded dong with the tracked
motor units was followed in order to establish the recruitment order of the motor units. The
subject received audio feedback only on the targeted motor unit.
Data are presented for 70 tracked single motor units recorded h m tbe flexor
carpi radialis of six subjects. Absolute isometric recruitment thresholds were obtained for 10
out of 70 motor units establishing a range from 110 g up to 646 g (2% to 11% MW).
Absolute isometric recruitment thresholds of other units were not available, however the
preloads used for recruitment of other units were similar, leading to a comparable range of
motor unit thresholds. Furthemore, motor units with relatively high recmitment thresholds
were studied for constant load lengthening contractions because in or&r to elicit high
velocity Lengthening contractions, a large preload was necessary to achieve a strong initial
isometric contraction.
With a targeted motor unit firing in the background, the subjects performed a
mean of 20 (range h m 3 to 88) lengthening contractions. At any one template velocity or
level of background activity, 3 to 12 lengthening contractions were completed for a tracked
motor unit. A total of 500 constant load lengthening contractions were perfonned with 35

tracked single motor units wbich were isolated during isometric contractions. Of these 35
units, 20 tracked single motor units were studied with gravitationdy loaded lengthening
contractions, and 15 were studied with constant current loaded lengthening contractions.
556 imposed load lengthening contractions were executed with 36 different tracked single
motor units, and 33 1 step pulse load lengthening contractions were observed with 9 different
cr;tcked single motor units. Frequently, there were additional motor units recorded
simultaneously with the targeted motor units mentioned above. The presence of these other
motor units was essential to draw important conciusions about motor unit behaviour during
lengthening contractions because they aiiow for a recruitment order to be establisbed between
different motor units.
Angular Diplacements and Mean Anplar Velocities
Resuits of recruitment order and firing pattern of motor uni& depend on the
velocity of the lengthening contraction (Nrudone et al., 1989; Kossev and Chnstova, 1998).
The experiments were designed to cover a wide range of velocities. The angular
displacement (change of wrist position) after 100 ms, 200 ms, and 5ûû ms h m the start of
the lengthening contraction (defined as the initiai change in tension) were computed. Figure
8 shows the fiequency disiribution of angular displacement of the wrist represented by the
degrees of wrist extension afcer 100 ms, 200 ms, and 500 ms for the constant Ioad
lengthening contractions including both gravitational and constant current loads. Figure 9
shows the same for the imposed load lengthening contractions. Both figures iiiustrate similar
ranges of anguiar displacemeots. 1t is important to note that even though a subject was
ptesented with t h position templates to foliow, subjects found template tracing dift?cult,
40

*~ngular Displacement aïter 200ms
*Angular Displacement alter SOOms
I 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42 44 46 48 5 Angulir Displitement (degremm)
- .-.
Figure 8: The frequency distribution of the mgular displacement during consant lond lengthening contractions. Change in wrist position after Iûûms, 200ms, and 500rns is displayed.
80 -
* ~ n ~ u l a r Dlrplacemanl alter 2OOmr
+Angular Dlsplacrmrnl alter zooms 60
Angulir Displrcement (drgraes)
Figure 9: The frequency distribution of the angular displacement during imposed load lengthening contractions. Change in position d e r 100ms, 200ms, and 500ms is displayed.

particulady during constant load lengchening contractions. This led to a wide range of
velocities of lengthening contractions.
The mean angular velocity was derived from the total change in angular
displacement over the t h e period. Because of enormous diffecences in background loads,
and sensitivity of the potentiometer, onset of displacement vacied considerably. Therefore,
average displacement over 100 ms, 200ms, and 500 ms per unit time were taken as estimates
of velocity of lengthening. The mean angular velocities obtained from the position data in
degrees per second during the initial 100 ms, 200 ms, and 500 ms of the constant load and
imposed load lengthening contractions are depicted in figures 10 and 1 1, respectively.
Angular velocities by differentiating the position trace were not calculated because the rate of
change of angular displacement was very non-lineu, thiit is lengthening did not occur at a
constant velocity.
-r-FhlAnglarvqaikgfiiriiaMnr
-tMoenAnglarv~&IirignR1SOOrs
5 10 15 P 2 5 3036 40 4 5 5 0 5 5 a6570 75 80 85 90 96lCû106110 m m k g u l r i h b d l y ( ~
Figure 10: The frequency distribution ofthe mean angular velocity during constant load lengthening contractions. Mean angular velocity during the Grst IOOms, 2OOms. 500ms is displayed.
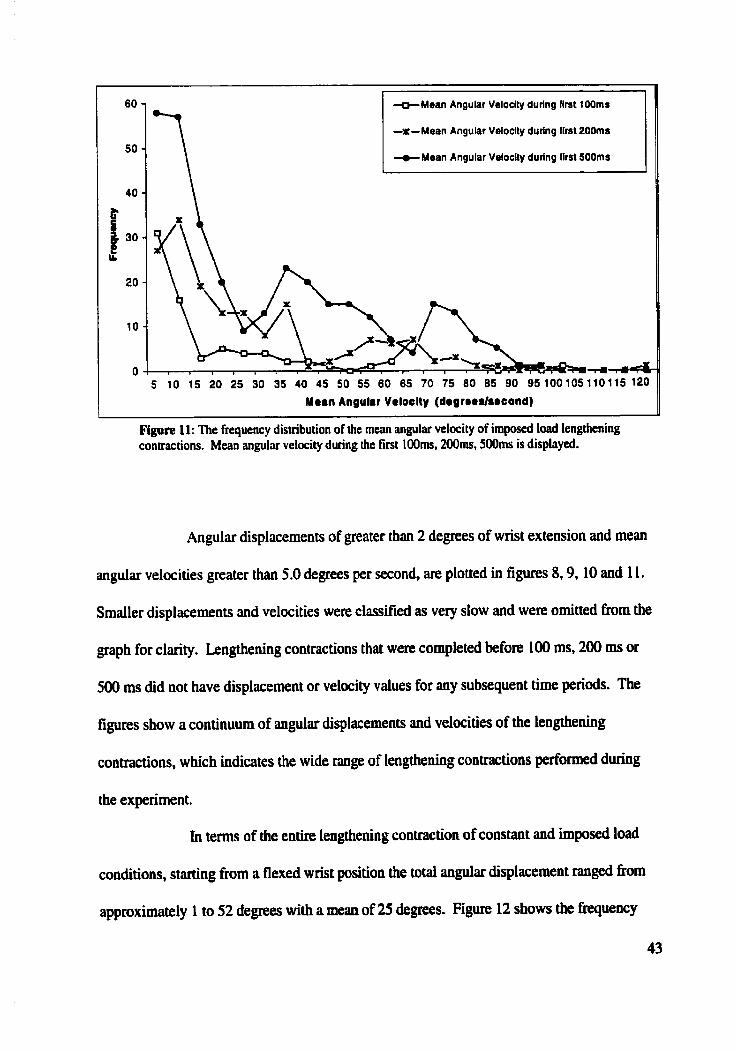
+Mean Angular Velocity during lirst 100ms
-x-Mean Angular Velocity during first 200ms
+Mean Angular Velocity during first SOOms
M i i n Angulir Viloclty (drgr~irlsicond)
Figure 11: The frequency distribution ofthe mtrui angular velocity of imposcd load lengthening contractions. Mean angular velocity dwing ihe first IOOrns, 200ms. 500ms is displayed.
Angular displacements ofgreater tban 2 degrees of wrist extension and mean
angular velocities greater than 5.0 degrees per second, are plotted in figures 8,9, 10 and 1 1.
Smaller displacements and velocities were ciassifieci as very slow and were omitted from the
graph for clmity. Lengthening contractions that were completed before LOO ms, 200 ms or
500 ms did not have displacement or velocity values for any subsequent tirne priods. The
figures show a continuum of angular dispiacements and velocities of the lengthening
contractions, which indicates the wide range of lengthening contractions performed during
the expriment.
In terms of the entire lengthening contraction of constant and imposed load
conditions, starting from a flexed wrist position the total angular displacement ranged h m
appmximately 1 to 52 degrees wiih a mean of 25 &gcees. Figure 12 shows the frequency
43

O 2 4 6 8 10 121416 1 8 2 0 2 2 2 4 2 6 2 8 3 0 3 2 3 4 3 6 3 8 4 0 4 2 4 4 4 6 4 8 5 0 5 2 toid -a Dkpla#mnt (ckqom)
Figure 12: The frequency distribution of the toial mgulru dispiiiccment of the lengthening conmctions.
distribution of the total angular displacement during the iengthening contraction. Figure 13
shows the frequency distributions of the mean angular velocities for al1 of the lengthening
contractions including the constant load and the imposed load lengthening contractions. The
mean angular velocities for the total lengthening contractions ranged from approximately 0.2
to 128 degrees per second. This was caiculated fmm the total position change divided by the
total time of completion of the lengthening contraction, Constant load lengthening
contractions had a range h m 0.2 to 124 degrees per second. imposed load lengthening
contractions had a range ftom 0.3 to 128 degrees per second. The velocities for the total
lengthening contraction are usefiil for description of the lengthening contraction as weii as
cornparison purposes to the velocities used in other studies. Lengthening contractions were
divided into slow and fast lengtfiening velocities. The classifications consist of slow
44

&Constant Load *lmposld lood
1 Total Angular Vmlocity (dmgrmsshmcond) Fi yn 13: The frequency distribution of the total angular velocity for the lengthening contractions.
lengthening contractions included velocities below approximately 35 degrees per second and
fast lengthening contraction included velocities pa te r than 50 degrees per second, and in
between they were considered medium velocity lengthening contractions. The classification
is not discrete nor absolute, but simply an approximate range of velocities in whicfi
lengthening contractions exhiiited similar characteristics.
Constant Load Lengthening Contractions
P r e l d
The preloads during the gravitational loaded lengthening contractions ranged
from 130 g to 1000 g correspondhg to approximately 2% to 17% of the maximum voluntary

contraction, The prehads imposed by the toque motor during a constant current lengthening
contraction cange h m 125 g to 750 g (2% to 13% MW). Through variation of the I d , a
range of muscle forces inciuduig different amounts of motor units could be æcruited.
Tension and Position
The start of ii constant Load lengthening contraction was characterised by a
distinct decrease in the tension exerted by tbe subject. Dunng a typicd slow lengthening
contraction, the dec~ase in tension continued slowly and graduaily throughout of the
lengthening contraction. However, during the fast lengthening contractions (generally p a t e r
thsin Wh), a biphasic shape in the tension trace was observed. Fit, an initial decrease in
tension occurred to allow for wtist extension followed by an increase in tension presumably
to slow and stop the lengtbening contraction. Even though the position trace showed a
rnonotonic extension of the wrist, the tension trace showed a biphasic shape. The upper trace
in Figure 14 shows a typicd example of this shape. The relationship of position and tension
is important in understanding the firing patterns of motor units during lengthening
contractions. The tension pattern gives insight into the motor unit recniitment and ring
pattern during lengthening.
Single Motor Unit Responses
M e n the subject with a tonicaily ïJring motor unit(s) dowed the background
load to extend the wrist, three generai patterns of motor unit discharge were observed during
repeated lengthening conu~~:tions: (1)the toaic motor uni@) silenceci during ihe lengtheoing
contraction, (2) the tonic motor unit(s) contiaued to fire during the lengthening conuaction,
46

1
Figure 14: Tension and position traces of a constant gnvitational load lengthening conîriwion illusîrating the biphasic pattern of tht tension trace during fast lengthening contractions. The angular velocity of the lengthening conuaction is approximately 8501s (pb405).
and (3) initially the tonic motor unit(s) is silenced, then midway through the lengthening
contraction, the same unit(s) tlred.
The predominant firing pattern of single motor units during constant load
lengthening contractions showed a silencing of the tonically firing motor unit(s) in the
background. Figure 15 shows a typical example of a constant load lengthening contraction
opposed by a gravitational load (A) and a constant load lengthening contraction opposed by a
torque motor load (B). Out of the 35 motor units studied during the constant load
lengthening contractions, 28 motor units exhibited the pattern of complete silencing of motor
units during the lengthening phase. 37% (186 out of 500) con lengthening contractions, 186
or 37% exhibited this behaviour. The time of occurrence of the silencing vacied, it could start
right at the onset of the initial decrease in tension or any time during the lengthening

Figure 15: Typical results h m constant load lengihening conûactions paradigm showing the silencing of the tonic single motor uni& (A) This figure shows 3 tonic motor units (1 large and 2 smali plus additional very small units) king silenceci during a gravitationd load lengthening contraction. The large, higher threshold unit was the tint unit to be demiteci. Note the large oscillations in the tension trace and the pasition trace during the slow contraction of 36" in -1.6s (22Smfs) (pb5OS). (B) This figure shows 3 tonic motor units (1 large and 2 smali) king silenccd during a constant cumnt load lengthening contraction. Again, oscillations, though l e s proncninced, are seen in the tension and position traces of this slow contraction of LCP in - 1.4s ('7%) (mg 105). Traces ftom top to bonom, flexor EMG, tension. position, SMU are displayed, An upwards change in the tension trace indicate-s an increase in tension and a downwards change indicates a demûse Extension is shown by an upwards change in the position trace aiad flexion is shown by a downwards change. The vertical bar indicates the beginnjng of îhe lengthcning conimtion.

contraction. S o m lengtfiening contractions showed an abrupt silencing or derecniitment of
alL tonic motor units whiie others showed a progressive orderly derecruitment of motor units.
When more than one unit fïred during the isometric contraction, the large motos units were
derecruited before small motor units as seen in Ggwe 15.
The second pattern of lengthening contractions with constant I d showed a
continued firing of the motor unit(s) in ihe background. Continued Firing of the motor units
included variation in the number of units that sustained activity. In some cases al1 tonic
motor units continued to Fm and in ohers higher threshold motor units were derecruited
while the small motor units continued as is iiiustrated in figure 16. The pattern of continued
firing motor units was seen in 20 out of 35 motor units. and uccurred during 30% (150 out of
500) of the constant load lengthening contractions. Generally, if a motor unit continued to
fre throughout the entire constant load lengthening contraction, the mean firing rate of the
single motor unit would decrease until the singie motor unit cesised to F i or the lengthening
contraction ended.
While many of the constant load lengthening contractions involved e i h r a
complete or partial silencing of the single motor units, there were many lengthening
contractions that exhibited a temporary silencing of single motor unit(@ and then ph& re-
recruitment of the same mit(~). This ttiitd pattern of single motor unit firing o c c d with
21 motor units during 32% (158 out of HIO) of the constant load lengthening contractions.
Figure 17 illustrates atypical example. The recruitment after silencing of the motor units
during the lengthening contraction occurred at various times ranging h m approximate1y 150
to 300 ms. The phasic recniitment of the motor unit@) was associated with an inctease in
tension which was decelerathg the lengthening phase. Also, during this phasic firing of
49

Figure 16: Typical results from constant load lengthening contractions paradigm showing the continuation of the single motor unit açtivity, (A) This figure shows 2 tonic motor uni& (1 large lower thnshold and 1 small higher thrcshold) continuing throughout a gravitational load lengthening contraction. Note the oscillations in the tension and the position traces in this slow contraction of 375" in -2.5s (15%) (pb712). (B) This t7gure shows 4 tonic motor units (1 large higher thnshold and 3 srnall lower thrcshold) of which the large unit is cieredted early in the lengthcning contraction while the small motor unie continue to tire until the end of the constant c m n c load lengthening contraction. The contraction is 6" in -15s long (4%) (kj 14). Traces €rom top to bottom, fiexor EMG, tension, position. $MU are displayed. An upwards change in the tension trace indicaies an increase in tension and adownwards change indicsites a decrease- Extension is shown by an upwards change in the position trace and flexion is show by a downwards change. The vertical dashed bar indicates the beginning of the lengthening contraction.
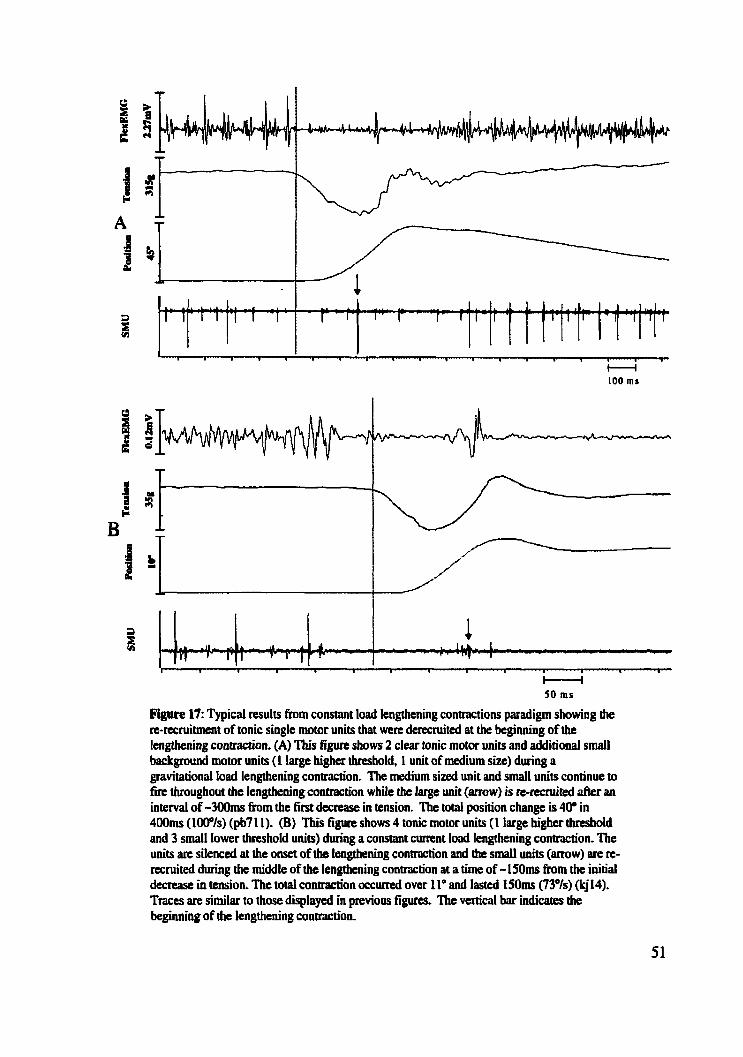
i?ipre 17: Typical results from constant load lengthening contractions paradigm showing the re-recruitment of tonic single motor units that were dereccuited at the beginning of the Iengthening contraction. (A) This figure shows 2 clear tonic motor uni& and additional small background motor units (1 large higher ihreshold, 1 unit of medium size) during a gravitational load lengthening contraction, The medium sized unit and small uni& continue to tire throughout the lengthening contraction while the large unit (mow) is te-ncnùted a k r an interval of -3OOms h m the îùst dec- in tension. The total position change is 40" in 400111s (10Ws) (pb72 1)- (B) This figure shows 4 tonic motor units (1 large higher thnshold and 3 small lower threshold units) d k n g a constant cunent Ioad lengthening contraction. The units are silenced at îhe onset of the lengthening contraction and the small units ( m w ) are rc- m i t e d during the middle of thc lengthening contraction at a time of -150ms from the initiai derrease in tension. The total contraction o c c d over 11" and lasted 150ms (73'1s) (kj141. Traces are similar to those displaycd in previous figures. The vertical bar indicates the beginning of the lengthening contraction.

motor units during lengthening contraction, a phasic burst of surface EMG was observed
which presumably contributed to the increase in tension and bcaking of the wrist extension.
The important points to note are that the unit firing phasically during lengthening in figure
17A is ihe same unit which was firing in the background. Secondly, while the large unit
derecmited, the lower threshold, smaller contractions units continued to fire during the
lengthening contraction. In figure 17B, only the smailer, lower threshold units fired
phasicaiiy, and the large one did not. No additional units were recruited.
Surface EMG
in the literature, it has been suggested that due to selective recruitment
of larger units, EMG during lengthening contractions is higher than during isometric
contractions (Nardone et al., 1989). To evaluate this ünding, the surface EMG
recorded fmm wrist flexors during the controlled isometric contraction was cornpared
to the lengthening contraction for each set of units recorded during constant load
conditions. The vaiues for the integrated EMG for al l the different lengthening
contractions are included in Table 2. The table Iists the mean surface EMG for 200
ms before and after the onset of the lengthening contraction for 35 motor units.
Paired t-test showed a statistically signïficant decrease in mean integrated EMG
during the lengthening contraction. Figure 18 shows a typical example of the EMG
activity. During the isometnc contraction for period of 200 ms before the lengthening
contraction, a mean EMG activity of 83.65 fl was observed and during the fkst 200
ms during Iengthening contraction that value dropped to 6.36 p.
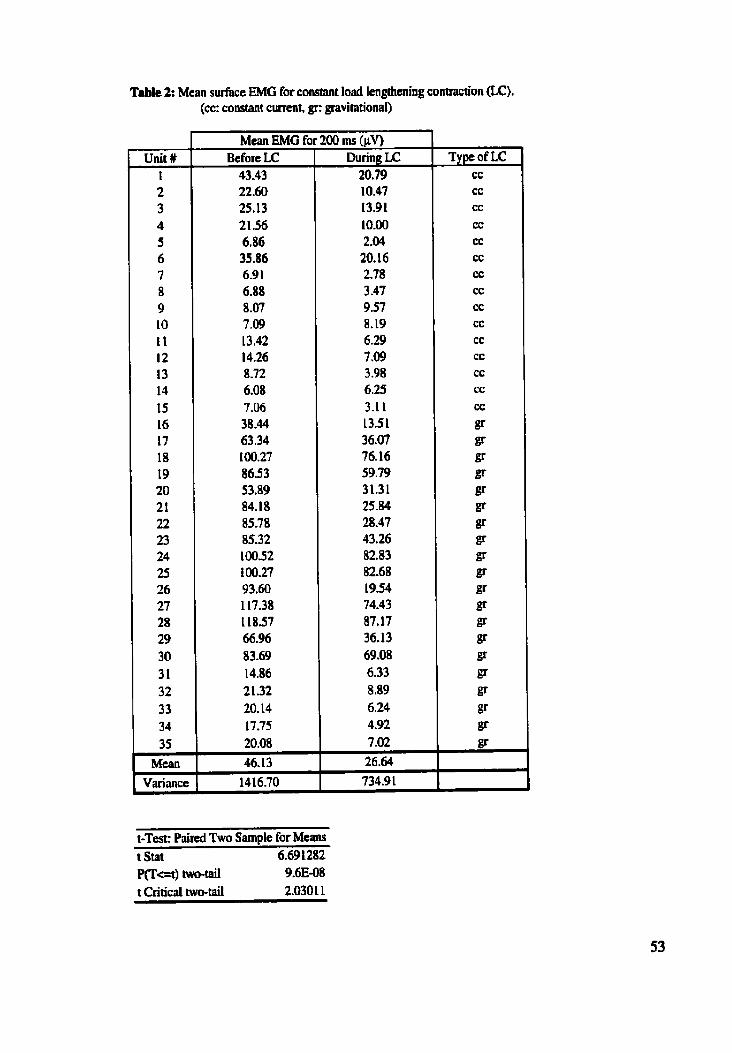
Table 2: Mean surface EMG for constant load lengthening contraction (LC).
Unit # 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 1 8 19 20 2 1 22 23 24 25 26 27 28 29 30 3 1 32 33 34 35 M m
Variance
(cc: constant curent, gr: gravitational)
M u n EMG fa Before LC
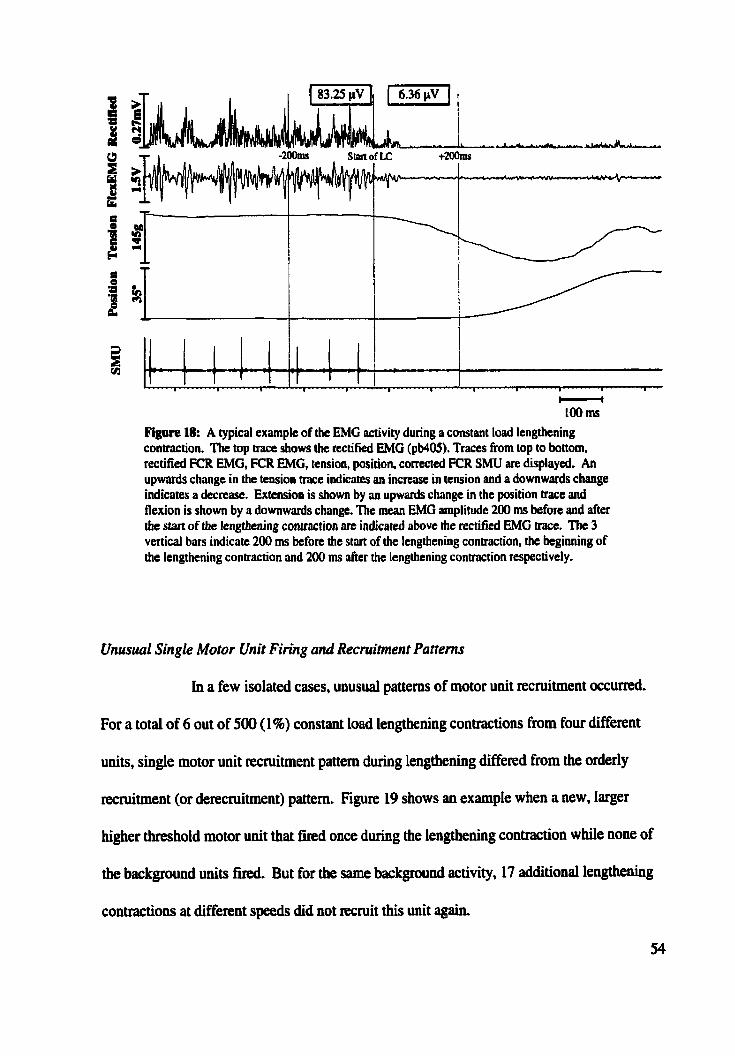
Figure 18: A typical example of the EMG activity during a constant load lengthening contraction. The top tnce shows the rectifiai EMG (pb405). Traces from top to bottom, rectified FCR EMG, FCR EMG, tension, position. comcted FCR SMU are displayed. An upwards change in the tension trace indiates an inm;ise in tension and a downwards change indicates a decrcasc. Extension is show by an upwiuds change in the position trace and flexion is shown by a downwards change. The mean EMG amplitude 200 ms before and after the start of the lcnglhcning convacrion are indicated above the rectifiai EMG trace. The 3 vertical bars indicate 200 ITE before thc s t u t of the Iengthening contraction, the beginning of the lengthening contraction and 200 ms &et the lengihening contraction respectively.
Unusual Single Motor Unit Firing and Recruiîment Patterns
In a few isolated cases, unusual patterns of motor unit recruitment occurred.
For a total of 6 out of 500 (1%) constant load lengthening contractions from four different
units, single motor unit rec~itrnent pattern during lengthening differed from the orderly
recmitment (or derecniitment) pattern. Figure 19 shows an example when a new, Iarger
higher threshold motor unit that h d once duRng the lengthening contraction whiie none of
the background units Wd. But for the same background activity, 17 additional lengthening
contractions at different speeds did not recniit tbis unit again.

Figure 19: An example of an unusual panern of m i t m e n t during the constant load lengthening contractions pandigm. The figure shows 4 tonic motor units (2 large higher ihnshold and 2 small lower thceshold units). A new large unit is n c ~ i t c d during the lengihening contraction (pbl6). The totai contraction occurred over 15" and lasted 1.8 s (8 "Is). Traces h m top to bottom, FCR EMG, tension, position, FCR SMU are displayed. An upwards change in the tension trace indicates an increase in tension and a downwards change indicates a demase. Extension is shown by an upwards change in the position trace and flexion is shown by a downwards change. The vertical bar indicates the beginning of the lengthening contraction. In a t o d of 18 lengthening contractions npeated on these motor units, thii pattern was seen only once.

Oscillatory behiour
Imgularities in the form of oscillations in the position and tension traces were
evident dwing the lengthening contractions. The amplitude of the oscillations varied and
were generally dependent on the velocity of the lengthening contraction. Large oscillations
were observed in the tension traces for slower, pmlonged lengthening contractions. Figures
15A and 16A illustrate the pmnounced slow oscillations in tension seen during the decrease
in tension. Oscillations are noted in figure 15B although they are of quite smaller amplitude.
The oscillations are seen to be relatively absent for fast lengthening contractions. For
example, figures 17A and 17B show only very small oscillations in the tension and position
record. Frequencies of the oscillations ranged from 3 Hz in figure 16A to 8 Hz in figure 19.
Imposed Ramp h d Lengthening Contractions
Preloarls and Imposed Ramp Loads
The preloads applied by the torque motor varied from 125 g to 750 g (2% to
13% MVC) at 10 cm. The ramp load that was superimposed on the preload was aiso
ptoduced by the torque motor and vacied in the camp increase of speed and magnitude.
These ranged fiom 250g to 750 g. The total preload plus the load never exceeded 1000 g
(iimited by capacity of the torque motor),
Tension and Position
The start of an imposed load lengthening contraction was characterised by an
increase in net tension. The tension trace continued to increase during the lengthening
contraction as the load imposed on the subject increased. Slow irnposed load lengthening
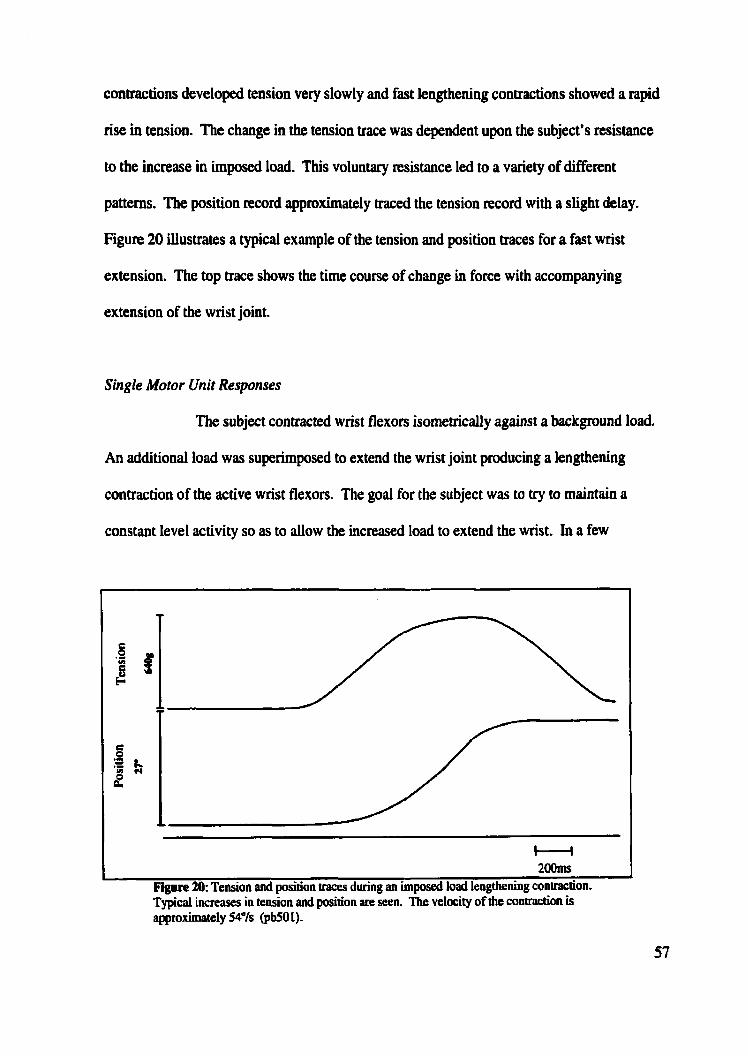
contractions developed tension very slowly and fast lengthening contractions showed a rapid
rise in tension. The change in the tension trace was dependent upon the subject's resistance
to the increase in imposed load. This voluntary resistance led to a variety of different
patterns. The position record appmximately traced the tension record with a slight delay.
Figure 20 illustrates a typical example of the tension and position traces for a fast wcist
extension. The top trace shows the tirne course of change in force with accompanying
extension of the wrist joint.
Single Motor Unit Responses
The subject contracted wrist flexors isometrically against a background load.
An additional load was superirnposed to extend the wrist joint producing a lengthening
contraction of the active wrist flexors. The goal for the subject was to try to maintah a
constant level activity so as to allow the increased load to extend the wrist. In a few
I Figure 20: Tension and position traces during an imposed load lengthening contraction. ~Gical incre;ises in ten&n and position are An. The velocity of the contraction is appmximately 54"/s (pM0 1).

experiments step pulse loads were also superimposed to lengthen the actively contracting
wrist flexors. However, only a litnited number of trials were perfonned for two reasons.
First, such a study has been previously done in detail (Calancie and Bawa, 198Sa) and,
second, step loads that produced comparable wrist extensions as in the other conditions
produced very strong reflexes. During such strong reflex activity clear distinguisbable
motor units could not be recorded and thus the firing and rec~itment pattern could not be
detemiined.
The imposed load lengthening contractions showed large variability in the
single motor unit responses, some were similar to patterns observeci wiih constant load
lengthening contractions, the magnitude of the load and the camp speed of the load. The
patterns were largely dependent upon the subject's voluntary response to the imposed loads.
The three typical firing patterns during imposed load lengthening contractions included (1) a
decreased firing rate andlot silencing of the tonic motor unit(s), (2) a continuation andor an
increase in the firing rate of the motor unit(s), and (3) recniitment of additionai higher
threshold unit(s).
The first pattern of a constant load lengthening contraction showed a decrease
in firing rate andior a silencing of the firing of the motor unit@) in the background. Figure
2 1 is a typical example of the silencing of motor units during such conditions. The pattern
of decreasing rate or silencing of the background firing motor units was seen in 6 out of 36
motor units and occurred during 10% (56 out of 556) of the imposed load lengthening
contractions. In the example shown in figure 21, it cm be seen that during lengthening a l l
motor units are derecniited, the larger before the mal1 ones which would be expected
according to the sue principle.

Figure 21: Typical resulu from imposed load lengthening conuactions paradigm showing the silencing of the tonic motor units. This figure shows 4 tonic motor units (1 large and 3 small) plus additional very smail uni& k ing silenceci midway through a lengthening contraction. The large, higher thnshold unit was the first unit to be derecruited. Note the small oscillations in the tension trace during the contraction of 36" in -800111s (45%) (pb409). Traces of FCR EMG, tension, position, FCR SMU are displayed respectively from top to bonom. An upwards change in the tension trace indicates an increase in tension and a downwards change indicates r demase. Extension is shown by an upwards change in the position trace and flexion is shown by ûdownwiuds change. The vertical bar indicates the beginning of the lengthening contraction.
The predominant f ~ n g pattern of single motor units during imposed load
lengthening contractions was a continuation of the r i n g of the tonic motor unit(s) in the
background according to the instruction given to the subject. Figure 22 is an example of a
recording in which the subject maintained some motor unit activity. The large higher
îhreshold unit was derecniited, but the small lower threshold units continued to fire
throughout the lengthening contraction. Figure 23 provides two additional examples of the
continuation of motor units with slightly different patterns during lengthening contraction.
Ali background units continued to tire thughout with slight lengthening of interspike
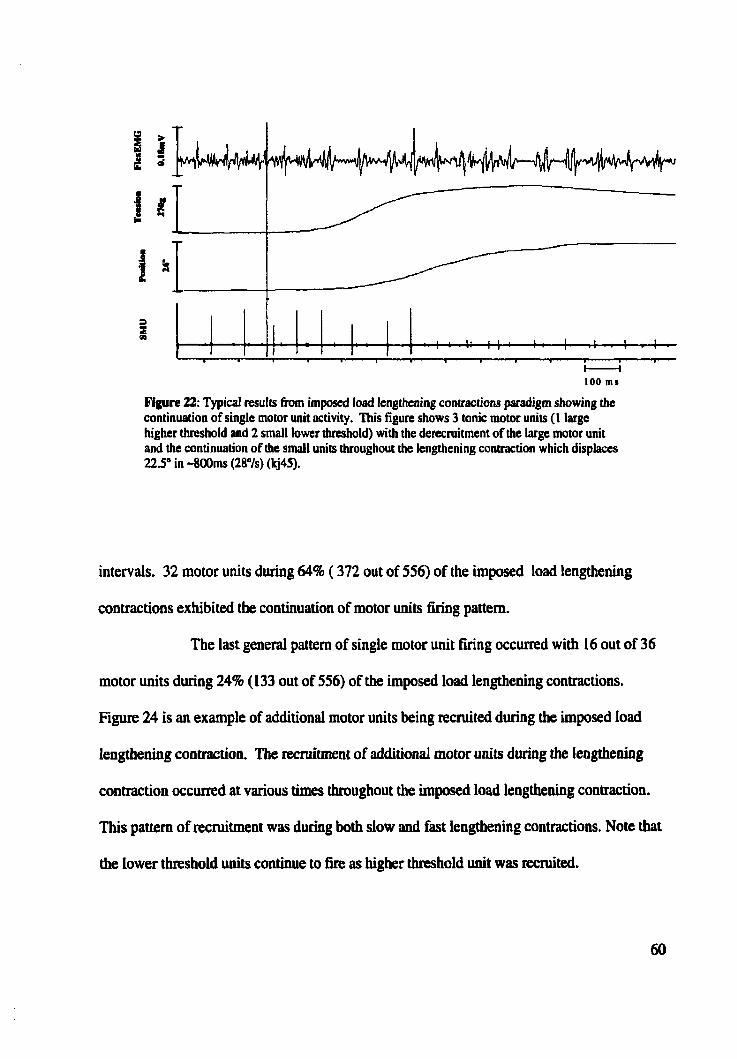
Figure 22: Typicd resutts fiom imposed load lengthening contractions paradigm showing the continuation of single motor unit activity. This figure shows 3 tonic motor uni& (1 large higher threshold and 2 small lower threshold) with the derdtment of the Iiuge motor unit and the continuation of the smdl units throughout the lengthening contraction which displaces 22.5" in -800ms (28%) (kj45).
intervals. 32 motor units during 64% ( 372 out of 556) of the imposed load lengthening
contractions exhibited the continuation of motor units nring pattern.
The last generai pattern of single motor unit firing occurred with 16 out of 36
motor units during 24% (133 out of 556) of the imposed load lengthening contractions.
Figure 24 is an example of additional motor units king recruited during the imposed load
lengthening contraction. The recniitment of additional motor uni& during the lengthening
contraction occurred at various rimes throughout the imposed load lengthening contraction.
This pattern of recruiûnent was during both slow and fast Iengthening contractions. Note that
the lower threshoId uni& continue to 6re as higher threshold unit was recruited.
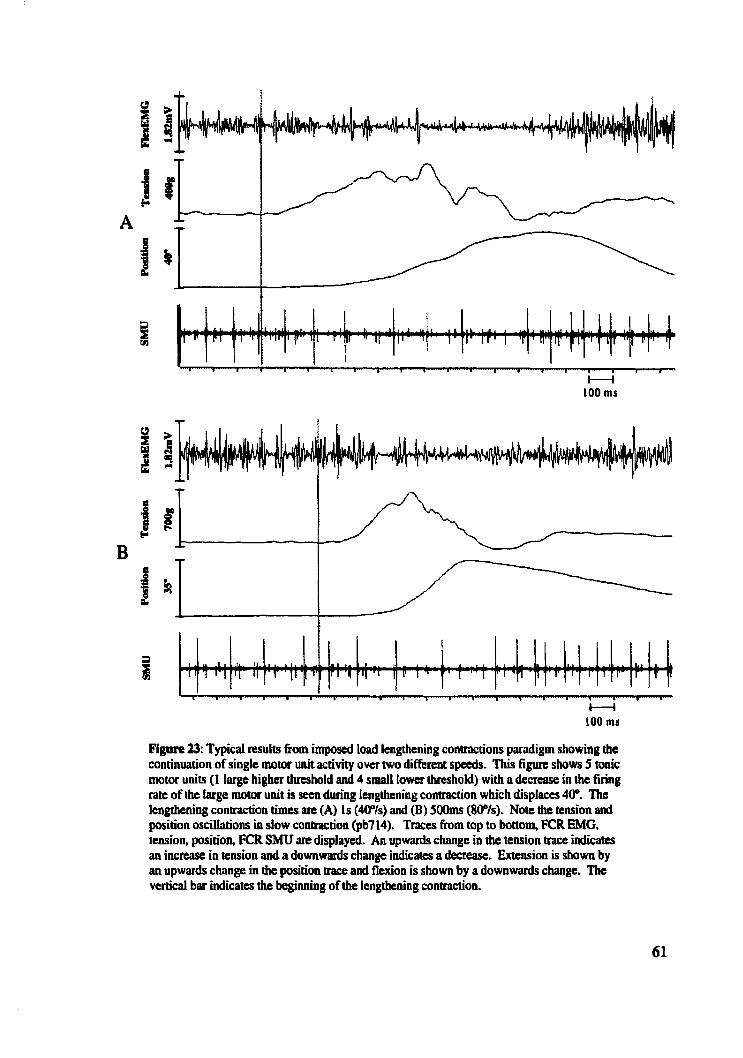
Figure 23: Typical results from imposai load lengthening contractions paradigm showing the continuation of single motor unit activicy over two difirent specds. This figure shows 5 tonic motor units (1 large highcr thrcshold and 4 small tower thmhold) with a demase in the firing rate of the large motor unit is seen during Iengthening contraction which displaces 400. The lengthening contraction times are (A) 1s (40'1s) and (B) 500ms (8001s). Note the tension and position osciilations in slow contraction (pb714). Traces from top to bottom. FCR EMG, tension, position, FCR SMU are displaycd. An upwards change in the tension tnce indicates an inctease in tension and a downwards change indicaies a decteosc. Extension is shown by an u p w d s change in the position Irace and flexion is show by adownwards change. The vertical bar indicates the beginnhg ofthe lengtkning conaction.

Figure 24: Typical results from imposed Ioad lengthening contractions pandigm showing the recniitment of additional single motor units during h e lengthening contraction. This figure shows 1 small lower threshold tonic motor unit with the additional larger threshold motor units appearing mid-way through the Iengthening contraction. The total position change is 12" in -1.1s (1 1'1s) (mg103). Traces ftom top to bonom, FCR EMG, tension, position, FCR SMU are displayed. An upwards change in the tension trace indicates an increiise in tension and û downwards change indicates a dwrease. Extension is shown by an upwards change in the position trace and flexion is shown by a downwards change. The vertical bar indicates the beginning of the lengthening contraction.
Su#ace EMG
During imposed load lengthening contractions, if the subject tried to maintain
the same activity before and during the lengthening contraction, surface EMG should be
comparable during the two periods. Table 3 îists the mean integrated surface EMG for 28
units 200 rns before and after the onset of the hposed load lengthenhg contraction. Using a
paired t-test, the mean EMG values of ail mirs 200ms before and d e r the start of the
62

lengthening contraction were compared. Paired t-test showed a statisticaüy signincant
decrease in mean integrated EMG during the lengthening contraction. However, note much
more variability was seen, as 10 units showed an increase in activity during the lengthening
contraction and 26 units showed a decrease. Figure 25 illustrates an example of the integrated
EMG of an imposed load lengthening contraction. During the isometric contraction for
period of 200 ms before the lengthening contraction, a mean integrated EMG activity of
46.04 pV was obse~ed and during the first 200 rns during lengthening contraction that value
was 50.00 pV. Note that the mean integrated EMG value over the first 400 ms of the
lengthening contraction did increase to 86.34 pV.
ilnusual Single Motor Unit Finng and Recruitment Patterns
As was seen during the constant load lengthening contractions, unusual
patterns of motor unit recruitment occurred during imposed load lengthening contractions. in
9 out of 556 (2%) imposed ioad lengthening contractions of five motor units exhibited
patterns of recruitment that differed h m the orderly recruitment (or derecruitment) patterns.
Figure 26 shows an example of a large bigher threshold motor unit f i n g during the
lengthening contraction and the smalier motor unit ceased to Fm. Twelve lengthening
contractions were perfonned with this set of motor units at this approxirnate velocity and 2
out of the 12 lengthening contractions showed this type of phasic recniitment. Since phasic
recruitment is probabilistic, 2 out of 12 daes not qualify for reversed recruitment. Most
imponantly, the large unit was not a vecy large twitch motor unit caused by the selective
recruitment of large units. It was just the next higher threshold unit as shown in figure 24. It
just happened to 6re only once.

Table 3: Mean surface EMG for imposed rmp load lengthening contractions (LC).
Unit # 1 2 3 4 5 6 7 8 9 10 1 1 12 13 14 15 16 17 18 19 20 21 22 23 24 2s 26 27 28 29 30 3 1 32 33 34 35 36
Mean Variance
Before LC 16.77 21.51 67.20
. 52.51 57.25 57.90 92.75 68.87 73.3 1 90.79 78.70 62.77 72.02 169.43 15 1.69 40.10 34.60 40.74 33.01 1 1.79 2 1.69 39.04 24.46 18 -43 30.12 12-77 11-70 8.98 4.05 8.25 11.64 10.66 7.86 7.87 6.45 6.50 40.88 1604.79
NOTE: 10 units showed an increase in activity during the lengthening contraction. 26 showcd a decrcase
Mean EMG for
I
[-Test: Paired Two Sample for Means t Stat 323 PVc=t) two-tail 0.00256 t Critical two-tail 203
200 ms (pV) Dunng LC
23.92
II
I -

u 100 ms
Figure 25: A typical example of the EMG activity during an imposed load lengthening contraction. The mcan ampütude of EMG for a typical i m p d toad 200 ms bcfore and 200 ms after the start of the lengthening contraction determinui fÎom the f i t decnasc in tension are shown in the boxes (pb803). Nom. This pwticular example wouid pmbably show a significant increasc in the mean iEMG if the time uscd for integration was expanded,
C---(
100 ms Figure 26: An example of recruinnent of a largcrthrshold unit during an imposed load lengthening contraction. The smallet, Iowa thrrstiold unit ceases ta tire and a large unit was recruitcd. Howevcr. very srnaIl uaiis do contiaue to fm in background. Fmm the EMG trace, it is evident that a large burst of muscle activity o c c d at the time of the large motor unit firing and the smaller unit may be cefractory and ihus unable to fire (pb6û6).

Oscillatory Behaviour
As was seen in constant load lengthening contractions, oscillations were
evident in the tension and position traces during imposed load lengthening contractions.
Slow lengthening contractions, such as in figure 23A. showed distinct oscillations in both the
tension and position trace. Faster lengthening contractions showed significantly smaller
osciliations as seen in figure 24.
Imposed Step Load Lengthening Contractions
Square puises, 150 ms in duration fed to the torque motor, extended the wrist
joint producing small fast camp lengthening contraction of wrist flexors. These lengthening
contractions had small angular displacements of 1.5 to 6 degrees. These displacements are
small, although similar to some lengthening contractions that were completed in the otber
two conditions. These displacements are not comparable to displacements used in the
Literature. However, the muscle is king actively lengthened. Such fast stretches elicit strong
stretch reflexes. As shown previously by Calancie and Bawa (1985a), a size-ordered reflex
recruitment during the rapid stretch was found. The tonic background single motor units
tended to respond during both the shoa latency reflex (30-55ms) and the long latency reflex
(55-100ms) periods and additionai larger units which were the next higher threshold units to
be recruited through voluntary effort. Figure 27A shows background tonic units firing dwing
the stretch reflex with an additionai larger unit responded during the long latency reflex
period. This large unit is just the next unit to be recruited voluntarily as shown in figure 27B.
The unit was recniited to fire tonicaüy and the loads were irnposed to examine aU tonic
rnotor units.

Figure m. Typical results h m ttsestep iuque pulse imposed load lengthening contraction. (A) This figure shows 4 tonic motor units (1 large and 3 small). During the step toque pulse lengthening contraction, a new h g e higher thrrshoId motor unit is recruited -65111s after the beginning of the toque pulse. The other background tonic units also fire concumntiy (pb14). (B) In the same expcriment, ihe large moior unit which fired phasically in A, is recniited to be tonic to increase the total to 5 tonic uni& (2 large and 3 small). Now. during the step torque pulse, a decreasc in the inter-spike inmval is secn with the large motor unit fuing again at -60111s aftcr the application of îhc toque pulse(pb 14). Traces, ftom top to bottom, comsponding to FCR EMG, tension, pition, FCR SMU are diiplayed. An upwards change in the tension trace indic- an i~xease in tension and a downwards change indicates a decrease. Extension is show by an upwards cbange in the position trace and flexion is shown by a dowawards change. The vertical bar indicates the beginning of the lengthening contraction.

EMG Tremor
Thmugh a visuai inspection of the data, a noticeable trernor in the surface
EMG was observed before and during many lengthening contractions. Figure 28A and 28B
show typicai examples of this phenomena. The slight tremor was seen in selected recordings
of aimost al1 the subjects across al1 threshold motor unit(s). Subjects experienced muscle
tremor for brief periods while they were holding a constant force and/or d u ~ g the
leagthening contraction. The trernor is depicted as bursting of EMG activity made
increasingly evident with rectification as in figure 29, In many cases, the tremor was
accompanied by brief periods of synchronous finng of single motor unit. It was found that
the tremor was increasingly evident during later sets of lengthening contractions or usuaily
during or following lengthening contractions with large preloads probably resulting in fatigue
of the muscle.
Summary of Motor Unit Firing Behaviour
Dunng the lengthening contractions of the human FCR, an orderiy
recruitment or derecruitment of motor units according to their relative voluntaty isometric
threshold was seen during approximately 98% of the lengthening contractions. There was no
evidence to support systematic selective recruitment of large, fast-twitch motor units with a
derecniitment of the smail, slow twitch units. Constant and imposed load lengthening
contractions of different velocities gave a variety of different patterns, but the large majority
of units exhiiited recniitrnent and 6ring patterns bat were in accordance with the size
principle of orderly recruitment. When Iarge motor uni& were recruited, they were generdy
the next highest threshold motor unit to be recruited with isometric voluntary effort.

Figure UI: An example of the accentuoted amor during a lengthening contraction. (A) Bursts are seen in the surface EMG and are emphasized in the rcctified signal. The single motor unit activity shows king bursts of single motot units during the lengthening conuaction. Synchronization ofunits may possibly be occwring which is evident from the random finng of motor units during the initiai isometric periad, and bwsts of xtivity during the lengthening contraction. Nok that the large motor uni& do not firr done, but they accompanied by smder units. Traces, h m top io bottom, corccsponding to FCR EMG, tension, position, FCR W aie displayed, An upwards change in the tension trace indicates an increase in tension and a downwards change indicales a demase. Extension is shown by an upwards change in the position trace and flexion is shown by a downwards change types of lengthening contractions. The vatical bar indicates ihe beginning of the lengthening conaaction.

Figure 29: An example of amor during an imposed load lengthening contraction (pb408). The rectified FCR EMG makes the tremor quite apparent. Traces, fiom top to bottom, correspondhg to rectified FCR EMG, FCR EMG, tension, position, FCR SMU are displayed. An upwards change in the tension m e indicates an increase in tension and a downwards change indicates a decrease. Extension is shown by an upwards change in the position trace and flexion is show by a downwiuds change, The vertical bar indicates the beginning of the lengthening contraction.

DISCUSSION
The purpose of this study was to characterize the recniitment and firing
pattern of motor units during lengthening contractions. To accomplish this the isometric
recruitment order of al1 discernible motor units was determined, with the lower and higher
threshold units defined according to either their isometric reccuitment threshold or theu
relative recniitment order during the isometric contraction. In generai, the recniitrnent of
motor uni& during lengthening contractions of the wrist flexon was shown to follow the
recmitment of motor units during isometric contractions. The absence of selective
recniitment of large, higher th~shold motor units during the lengthening contractions of the
flexor carpi radialis strengthens support for the size principle of m i t m e n t of motor units.
Limitations of the study
The success of single motor unit experiments is dependent on correctly
tracking individual motor units with the a h of determining their firing and recniitment
pattern. Several trials did not meet the discrimination criteria and were discarded during
recording or were not included in the analysis. The exact success rate of the experiments was
unavailable because on-line analysis revealed the motor units during several lengthening
contractions were indistinguishable and thus were not recordai for M e r off-line anaiysis.
In total 70 motor units were tracked during a total of 1056 constant and imposeci load
lengthening contractions which we believe gave us an adequate sample for detennining the
recruitment pattern of motor units.
Perfonning single motor unit experiments is not a simple task. It requires
7 1

subjects who are able to conirol the intensity of their muscle contractions so that individual
motor units can be identified wMe performing the necessary experimental procedure.
Therefore, skilled subjects are required which results in a small number of subjects during
single motor unit experiments. In our study, only 6 subjects were involved. Yet, aii the
subjects in our study did produce the sarne generai patterns of firing of motor units and we do
not believe this produce any biases in the data. In addition to the number of lengthening
The establishment of the motor unit recruitment during lengthening
contractions relies on the validity of the cornparison between the recruitment of motor units
during isometric and lengthening contractions. We believe this to be true because an orderly
recruitment of motor units according to the size principle has clearly been determined during
voluntary isometric contractions of muscles (Milner-Brown et al., 1973b; Monster and Chan,
1977; Yemm, 1977; Calancie and Bawa, 198% Thomas et al,, 1987; Riek and Bawa, 1992;
Jones et al., 1993). Since the order of recniitment for isometric contractions was considered
as the control, only those sets of single motor uni& where the isometric recruitment order was
established were included in the study for Iengthening contractions.
Recording of motor unit activity during movements can result in the variation
of the shape and amplitude of the action potentials due to the movement of the electrode with
respect to the muscle fibres. Because of variations, careful visual inspection of the motor
units or generation of MUTA was necessary to correctly identify the motor units. Even
though, the change in size and sbape of motor units often occurred during the lengthening
movement, the firing of the unit couid be foUowed h m one motor unit action potential to the
next because the change was usually gradual through the lengthening contraction. In many
instances, the original size and shape returned wkn the wtist was retunied to the original
72

position. However, some data was unable to be inclukd in the study because movement of
the needle made it impossible to establish identity of the unit under investigation.
The delimitation of the study rats on the generalizability of ihe flexor carpi
radialis to other human muscles. Oae might question whether certain muscles, wbich might
experience rapid lengthening contractions frequently during behavioural movements, are
more prone to the selective recruitment of large motor units. Although lengthening
contractions of the flexor carpi radialis do not readily occur, the biceps brachii does
reoccuringly experience rapid lengthening contractions and in three studies using this muscle,
no selective recruitment was observed. We believe that with a more thorough study, like the
one presented hete with the flexor carpi radialis, the triceps surae will be shown to follow an
orderly recmitment according to the size pcinciple. However, in an attempt to compare motor
unit recruitment strategies across muscles, it is important that the muscle possess sirnilar
input ihrough neural connections. The flexor carpi radialis and the triceps surae muscle do
possess similar central nervous system connections, including Renshaw cells and
heteronymous connections. It is also difficult to compare parameters such as contraction
velocities across different muscles because of different muscle properties including muscle
length, architecm and fibre type composition. We attempted to control for the ciifferences
between muscles in order to compare between muscles, but acknowledge that comparisons do
have restrictions.
Anguiar Disphcement and Velocity
Comparison of the velocities and position changes obtained in this siudy to
other studies reveals that a sufficient range of angular displacements and anguiar velocities
73

were performed. The relative relaxation times of motor units in the individual muscles were
considered. The slowest relaxation tirne of motor units of the triceps surae occurs in the
soleus muscle which has a relaxation time of 500 ms (ref). In the study by Nardone et al.,
ankle extensor lengthening contractions were performed at approximately 25 degrees of ankie
flexion in 1 second, which is approximately twice the relaxation time of the slow soleus
motor unit. The flexor carpi radialis has a fast relaxation time of 100 ms. Therefore, an
analogous velocity is 25 degrees in 200 ms for a final velocity of approximately 100 degree
per second. Approximately 30 lengthening contractions were performed at velocities above
100 degrees per second. Therefore, we believe comparable velocities were performed.
However, as described above it is dificuit to compare contraction velocities across different
muscles so we attempted to cover a large range in order to include any velocities where
selective recruitment in the flexor carpi radialis may occur.
It must be mentioned that comparison of angular displacements, angular
velocities and the percent of maximum voluntary contraction between studies using different
muscles is limited due to many factors. For example, one could postulate whether the
excursions of the wrist flexors in this study is comparable to that of the uiceps surae in the
study of Nardone et al. (1989). Normai range of motion at joints could be compared, but that
still does not take into account the differences in original muscle lengths, changes in muscle-
tendon ratios or muscle moment arms.
It is dificuit to compare magnitudes of angular velocities because of different
muscle properties, including lengths of the muscles being studied. As weii, the other studies
used the statt of the change in position as the start of the lengthening contraction, where in
our study the start of the change in tension was defmed as the start of the lengthening

contraction. Therefote, surne disctepancies in the exact velocities may occur* To overcome
the potentiai restriction, the ideal strategy was to cover a wide range of velocities. In
addition, if accordhg to Nardone et al. (1989), selective d t m e n t occurred only in a
certain range of velocities then it was necessary to test the entire range of velocities. in our
study a large range of velocities were completed. Specifically, the range of velocities
comparable to that useci during other studies we covered was from and no velocities showed
a tendency for large motor units to be recruited selectively.
Single Motor Unit RQCtlljtment and Rring Pattern
Our results showed no evidence of s y s t e d c selective mniitment of large
higher threshold motor units with derecruitment of small lower threshold units. The vast
rnajority (98%) of the discrirninated motor units was recruited in order of tecruitment
increasing size. Subjects performed two types of lengthening contractions of the wrist
flexors. During the different lengthening contractions, many variations of firing patterns
were observed. The firing pattern during the lengthening contraction usually involved a wide
variety of recmitment, derecruitment and increasing or decreasing firing rates of motor units,
but none of the Iengthening contractions violated the size principle of motor unit recruitment.
Comporison of the Ibo opes of Consîant Load Lengthenhg Conhacnions
The gravitationaily loaded and the constant current l d e d lengthening
contraction revealed no significant difference in the pattern of their lengthening contractions.
Traces for each type of lengtheaing contraction showed the same gened patterns, which is
evident in the similar figures. Both types also have a similar range of velocities of

contraction. The central nervous system does not distinguish the type of load (wbether
gravitational or peoduced by toque motor) that stretched the active muscle, only the effect of
the applied load is apparent.
Constani Lm&
During some slow lengthening contractions, the angular displacement of the
wrist extension was smooth with derecruitment of background units. The derec~itment
occurred at various times during the lengthening contraction, but no additional remitment
was seen dunng this derecruitment. However, often when a subject petformed a slow
lengthening contraction, the angular displacement was not smooth as it was superimposed by
slow oscillations as shown in figure 21. The same phenomena can be seen in the figures of
the studies by Howell et al. (1995) and Nardone et al. (1989). Perhaps îhese slow oscillations
which include very small, fast shortening or isometric contractions akin to ballistic
contractions would recruit a burst of low and higher threshold motor units. The tension
traces in our study frequently showcd small increases in tension similar to small
superimposed ballistic shortening contractions during which some recruitment of motor units
were seen. Most of the motor units were the same motor units that had been tonically firing
in the background. Additional rec~ited units were those detennined to be the next highest
threshold and not selected large, higher threshold units. However, the Nardone et al. (1989)
study did not report any tension records for their lengthening contractions and thus
cornparison is unavailable.
During fast lengthening contractions with constant load, most of the tnds
showed a silencing of aü the motor activity during the lengthening contraction. At the start
76

of lengthening contraction, tension decreased with simultaneous decrease in motor unit
activity. This was foUowed by a fast increase in tension, while the wrist was still extending.
The biphasic shape of tension, illustrateci in figure 21, may explain the sudden re-recruitment
of same motor units such that in or&r to stop the lengthening contraction as it nears the end
of the required displacement, the muscle contracts and recmits motor units to consequently
end the lengthening contraction. Therefore, fast lengthening contractions repeated saw an
orderly recruitment of the same lower threshold motor units ihat were noted during the
isometric contraction. This is contradictory to the suggestion by Nardone et al. (1989) that
large motor units would be recmited in order to provide a fast relaxation time during the fast
lengthening contraction. The motor units that were recruited in our study had a sufficiently
short relaxation time to produce the rapid lengthening contractions
Imposed Ramp LwdP
When an active muscle was lengthened by an additional ramp, again an
orderly recruitment was obsewed with no systernatic selective recruitment of large, higher
threshold motor units over small, lower threshold units. During lengthening contraction with
slow imposed ramp loads, background units generally continued to fire with no additional
units recruited. The tension increase was usually very slow as weU. With fast imposed ramp
loads, additional units were fquently recruited, though not at the expense of derecruitment
of lower threshold units that were tonically firiag in the background. These additional
recruited units were the next highest threshold units recniited ituough voluntary recniitment.
The tension inctease during the lengthening contractions was due to the resistance of the
imposed ramp load, ibis was also generally associated with increased fîring of the motor unit.
77

The imposeci load displayed more variability in the recruitrnent pattern of its units due to the
higher possibility of response of ihe subject. It depends on a balance between how much a
subject lets their wrist be extended versus how much they resisted voluntarily.
With slow ramp increase, we did not observe recruitment of additional units.
With faster camps very frequently we observed pbasic firing of additional units. These
additional units, again, were the next higher threshold units tested either with voluntary slow
or ballistic contractions. It may also be noted that the burst of additional units was most
probably due to fast voluntary reaction of the subject. The motor units fired at approximately
200 to 250 ms after the onset of the imposed ramp load. This time is much longer than the
time for reflexes (25-100 ms). With fast imposed ramp loads, additional units were
frequently recmited, though not at the expense of derecruitment of lower threshold units that
were tonically firing in the background. These additional recmited units were the next
highest threshold units recniited through voluntary recmitment (Figure 27).
Imposed Step Loadr
The lengthening contractions in our study that were produced with step ramp
loads showed an orderly recruitment pattern. This corresponds to results established during
the thorough study by Calancie and Bawa (1985a) on motor unit recruitment during imposed
step loads on the wrist flexors tbat demonstrated that recruitment proceeds in an orderly
fashion according to the size principle through the production of the stretch reflex.
Surfaœ EMG
During lengthening contractions with constant loads, the surface EMG of the
78

wrist flexors showed a decrease in activity in our study. This is contrary to the increase in
surface EMG during lengthening contractions reported by Nardone et al. (1989). However,
in accordance with our results, Moritani et al. (1988) and Kossev and Christova (1998) did
not observe an increased surface EMG during the lengthening contraction. Both reported a
decrease in EMG activity pointing to tlse efficacy of lengthening contractions due to the
mechanical stiffness of the muscle. During lengthening contractions with irnposed loads, in
our study the surface EMG was generally lower. The instruction given to the subject was to
maintain a constant level of activity in the muscle, but variability in the response of the
subject to the irnposed load causes fluctuations in the surface EMG activity, but overall,
EMG was significantly lower during lengthening contractions. This confinns the efficacy of
lengthening contractions that has been reportecl in the literature such that under comparable
work loads, the electrical activity recorded involved in lengthening contractions is less than
that involved in the shortening of muscles (Bigland and Lippold, 1954; Basmajin, 1967;
Bigland-Ritchie and Wood, 1976). This lower recorded electrical activity at any given force
is achieved with either fewer fibres actïvated or lower frequencies of discharge or a
combination of both (Stauber, 1989).
Bursts in surface EMG activity accompanied by synchronous firing of motor
units occurred during several trials. The oscillations observed in the EMG of our study
developed either during trials involving relatively high loads like those incurred during the
gravitational loaded lengthening contractions or following these bigh load trials whüe
performing irnposed load lengthening contractions. This is consistent with involuntary
physiological tremor. Synchtonization of discharge of motor units can occur when an input
to the motoneuron pool is sufficiently large @ietz,et al., 1976), which occurs with

contractions that required large amounts of force (Jessop and Lippold, 1977). With increasing
fatigue and especially tremor brought on by fatigue, motor units tend to fire synchronously
(Milner-Brown et al., 1973a). The EMG bursting and the synchronization of motor units
were not quantifiai due to the limited and sporadic occurrences of the phenomena But
whatever the cause of the supposed trcmor, îhe recruitment and firing pattern of the motor
units did not violate the size principle.
Random Reversals of Recruitment Order
In our study, 2% of lengthening contractions showed a reversal of the
remitrnent order in motor units. These reversals occurred between motor units that were
rec~ited successiveiy during voluntary isometcic contractions and, thus, were sirnilar in their
recruitment thresholds. Reversals can d l y occur with motor units of similar conduction
velocities or twitch contraction times. Nomally in motor recruiünent studies, reversals occur
for less than approximately 10% to 15% of trials (Henneman and Mendell, 198 1). These
reversals do not have serious functional consequences due to the evidence that reversals
occur between motor units with similar properties (Senn et al,, 1997), and thus will not
interfere with gradual force production. Howeli et al. (1995) suggested a systematic selective
recruitrnent order with a 14% (3 out of 21) rate of reversal of motor units. Two of the three
units in their study could be better interpreted as recniitment revetsals between motor units of
similar thresholds. The one other motoc unit might betîer be explained through the
probabilistic f ' n g of motor units (Jones et al., 1994). Furthemore, this probabilistic firing
does not provide evidence for a theory of selective m i t m e n t of motor units during
lengthening contractions, but is a random phenornenon.

Selective Recruitment
Selective recruiûnent of motor units implies more than the random
recruitment reversai of motor units wiih similar thresholds like that seen in the HoweU et al.
(1995) or a random firing of a motor unit, but it involves the deactivation of the lower
threshold smail motor unit dong with recmibnent of a normally higher threshold large motor
unit. This was show by Kanda et al. (1977) wiih electrical stimulation of sural nerve in the
cat. Surai nerve stimulation inhibits slow S type motoneurons and excites F type
motoneurons. With a smng synchronws stimulation of sural nerve selective recruitment
was seen in triceps surae muscles.
In our study, large motor units were not selectively activated during
recruitment. Nardone et al. reported rnotor units that would fire during the lengthening of
active muscles and were silent during shortening and isornetric contractions. We believe that
the data from Nardone et ai. (1989) can be explained by Zengthening contractions with
irregularities in their position traces caused by small ballistic contractions occurring during
the lengthening phase. Therefore, the units that were said to be recruited only during the
lengthening phase were actually tecruited during mal1 isornetric or shortening contractions.
Even though we argue against selective m i t m e n t within the same muscles,
shifting of activity between different muscles is possible and observed. The synergistic
activity between muscles performing a task is not in question. For example, a shift of
recnlltment of motor units from soIeus (a slow, single joint muscle) to gastrocnemius (a fast,
two joint muscle) may mcur between hese spergistic muscles during a specifîc task such as
walking or nuining. This shifting of activity feom soleus to gaswnemius has been noted in
8 1

the literatuce (Nardone and Schieppati, f 988). However, shifting of activity from one muscle
to the next does not support a selective tecIuitment pattem because it is the recmitment order
of motor units within a muscle that is important, This shifting between different muscles
with different properties could not be confimed in the forelimb muscle because properties
(including speed of contraction) of all forearm muscle are quite similar.
Input to the Motor Udîs
The muscle activation pattem for isometric and dynarnic voluntary
contractions originates frorn commands of the central nervous system. if isornetric,
shortening and lengthening contractions are controlled through different command inputs to
the motoneurons as some would suggest (Nardone et al., 1989; Enoka, 1996), then an active
muscle producing one type of muscular contraction, for example, an isornetric contraction,
which changed to another contraction, for example, a lengthening contraction, should display
a change in the motor units recruited. However, our results showed the same motor units
recmited consistently during both the isometric and lengthening contractions, although with
different firing patterns. This suggests similar, but different levels of excitatory input to the
motoneurons depending on the specific type and level of contraction.
Possible mechanisms complicating the performance of lengthening
contractions and the variation of position, tension and motor unit activity during the
lengthening contractions may be expIained by additional input h m muscle afferents,
including muscle spindles, due to muscle Iengîh changes. Sensory output h m muscle
spindles can be generated by stretching the muscle. During lengtheniag contractions, small
itregular changes in angular displacements associated with uninteaded acceleration of a
82

lengthening movement can cause an i n c r e d discharge h m spindle endings (Burke et al.,
1978). These spindle responses can be associated with adjustments in the discharge pattern
of the voluntarily activated mucor units at latencies corresponding to the operation of spinal
reflex mechanism (Burke et al., 1978).
From a functional point of view, the size principle is a logical and economical
way for the central nervous system to control force production (Henneman and Mendell,
198 1). The size principle of orderly recniitment simplifies the neuromuscular control by
reducing the degrees of fieedom of the newomuscuhr system (Cope and Clark, 1995). For a
given level of signal descending to the spinal cord, an appropriate number of motoneurons
are recruited with suitable properties to carry out the desired movement. At higher tensions,
because the motor units are generally composed of larger muscle fibres and larger number of
muscle fibres, their tension increments are still appmximately the sarne proportional increase
in the total tension already present at the tirne of their recruitment (Henneman and Olson,
1965; Binder et al., 1996). Derecruitment from large to small motor units then would also
lead to an incremental decrease in force and as a result fine control can be possible at al1
tension levels.
Conclusion
The purpose of the study was to gain better understanding of the recruitment
of motor units during lengthening contractions. Our results show that during lengthening
contractions, flexor carpi radialis motor utiits are d t e d and detecniited in an orderly
manner according to the size principle and selective recruitment of large, higher threshold
motor units during lengthening contractions does not occur. Although cornparisons are doue
83

between muscles, further study needs to be done with the triceps surae muscles in order to be
able to directly investigate lengthening contractions that are comparable to those performed
during the study by Nardone et al., 1989. We suggest h m our results that the studies which
have ciaimed selective recruitment during lengthening contractions may have rnisinterpreted
their data. Lengthening contractions in Our study did not show selective recruitment of
motor units suggested by Nardone et al. (1989) and Howell et al. (1995). Our results support
the recruitment patterns reported by the previous results of Moriani et al. (1988), Sogaard et
al. (1996) and Kossev and Christova (1998).
Since the motor unit is the final conunon pathway for motor cornrnands, the
recruitment and firing pattern of the motor unit can provide insight into the complex
organization of the neural control of movement. Knowledge of how the human
newomuscular system controls the activation of a muscle during lengthening contraction has
applications in many areas. Particularly, due to muscle king actively lengthened in many
normal activities illustrated by the presence of lengthening contractions in movements such
as walking, running, jumping and throwing, just to name a few.

Bawa P, Binder MD, Ruenzel P, Henneman E. bruitment order of motor neurons in stretch reflexes is highly comlated with their axonal conduction velocity. J Neurophysiol, 52,410420,1984
Bigland B, Lippold OCJ. The relation between force, velocity, and integrated ekctrical activity in human muscles. 3 Physr'oi, 123,2 14-224, 1954.
Bigland-Ritchie B, Woods JJ. Integrated electromyogtam and oxygen uptake during positive and negative work. J Physiol, 260,267477, 1976.
Binder MD, Heckman CJ, Powers RK. The physiological control of motoneuron activity. In Brookhaa JM, Mountcastle VB (eds), HandboUk of Physiology. Z?te Netvous System, Motor Cuntrol. Bethesda, Amencan Physiologicai Society, pp. 345-422, 19%.
Brown AG. Organization in the Spinal Cord. Springer-Verag, Berlin, 198 1.
Burke RE. Motor unit types of cat triceps surae muscle. J Physiol, 193, 141-160, 1967.
Burke RE. Motor units: anatomy, physiology and functional organization. In Brookhart JM, Mountcastle VB (ais), Handbook of Physiology. The Nervous Systent. Motor Control. Bethesda, Amecican Physiological Society, pp. 345-422, 198 1,
Burke RE, Levine DN, Tsairis P and Zajac FE, Physiological types and histochemical profiles in motor units of the cat gastcocnemius. 3 Physiol, 234,723-748, 1973.
Burke D, Hagbarth KE, Lofstedt L. Musde spindfe activity in man during shortenhg and lengthening contractions. J Physiol, 277, 1 3 1-42, 1978.
Calancie B, Bawa P. Firing patterns of hurnan flexor carpi radialis motor units during the stretch reflex. J Neuropriysiol, 53 (S), 1179-1 193, 1985a.
Calancie B, Bawa P. Voluntary and retiexive recruinnent of flexor carpi radialis motor units in humans. J Neuruphysiol, 53 (9,1194-1200,1985b.
Caiancie B, Bawa P. Motor cemitment in humans. in Binder MD, Mendell LM, The Segmental Moror Sysrm. New York, Oxford University Press, pp. 75-95,1990.
Clamaun HP, Henneman E. EIecuical measwement of axon diameter and its use in relating motoneumn s h to criticai firing level. JNeurophysiol, 39,844851, 1976,
Cl& BD, Dacko SM, Cope TC. Cutaneous stimulation fails to alter motor unit &taient in the decerehate cat. J Neumphysiol, 70, 1433-1439, 1993.

Cope TC, Clark BD. Are there important exceptions to the size principle of alpha motoneurone recniitment? Alpha and Gamnta Motor System Symposium, London, Juiy 1995.
Cordo PJ, and Rymer WZ. Motor-unit activation pattem in lengthening and isometric contractions of hindlimb extensor muscles in the decerebrate cat. J Neurophysiol, 47, 282-296,1982.
Davies L, Wiegner AW, Young RR. Variation in firing order of human soleus motonewons during voluntary and reflex activation. Brain Res, 602, 104-1 IO, 1993.
Desmedt JE, Godaux E. Ballistic contractions in man: characteristic recmitment pattem of single motor units of the tibialis anterior muscle. J Physiol, 264,673-693, 1977.
Desmedt JE, Godaux E. Spinal motoneuron recmitment in man: Deordering with direction, but not with speed of voluntary movement. Science, 214,933-936, f 981.
Dietz V, Bischofberger E, Wita C, Freund HJ. Correlation between the dischanges of two simultaneously recorded motor units and physiological tremor. Elecfroencephalogr Clin Neurophysiol, 4û,97-105,1976.
Edstrom L, Kugelberg E. Histochernical composition, distribution of fibres and fatiguability of single motor units. Anterior tibia1 muscle of the rat. J Neurol Neurosurg Psychiat, 43, 424-433, 1968.
Enoka RM. Eccentric contractions requke unique activation strategies by the nervous system. J Appl Physiol, 8 1,2339-2346, 1996.
Freund W. Motor unit and muscle activity in voluntary motor control. Physiol Rev, 63,387- 436,1983.
Garland SJ, Cooke JD, Miller KJ, Ohtsuki T, Ivanova T. Motor unit activity during human single joint movements. JNeurophysiol, 76,1982-1990,1996.
Garnett R, Stephens JA. Changes in the recruitment thresholds of motor units producecl by cutaneous stimulation in man. J P hysiol, 3 1 1,463-473,198 1.
Garnett R, Stephens JA. The reflex responses of single motors units in human k t dorsal intemsseous muscle foUowing cutaneous afferent stimulation. J Physiol, 303,35 1-364, 1980.
Gcimby L, Hannen J. Recnùtment order of motor unit5 on voluntary contraction: changes induced by proprioceptive afferent activity. J Neurol Neurosurg Psychiahy, 3 1,565-73, 1968.

Grimby L, Hmerz J. Differences in mmitmcnt order and discharge pattern of motor units in the eady and late flexion reflex components in man. Acta Physiol S c d , 90,555-564, 1973.
Henneman E. Relation between size of neurons and iheir susceptibility ta discharge. Science, 126,1345- 1347,1957.
Henneman E, Clamann VD, Gilles VD, Skinner RD. Rank order of rnotrneurons within a pool: law of combination. J Neurophysioï, 373,1338- 1349, 1974.
Henneman E, Mendel1 LM. Functiond organization of motoneuron pool and its inputs. In Brooldiart JM, Mountcasrle V5 (eds), H d o o k of Physiology. The Nervous System. Motor Control. Bethesda, American Physiological Society, pp. 423407,198 1 .
Henneman E, Olsen CB. Relations between structure and function in the design of skeletal muscles. J Neurophysiol, 28,58 1-598,1965.
Henneman E, Somjen G, Carpenier Dû. Functional significance of cell size in spinal motorneurT1nes. J Neurophysiol, 28,560-580,1965.
Howeil JN, Fuglevand AG, Walsh ML, Bigland-Ritchie B. Motor unit activity during isomeüic and concentric-eccenüic contractions of the human first dorsal interosseus muscle. J Neurophysiol, 74,901-904, 1995.
Huxley AF, Simmons RM. Proposed mechanisrn of force generation in striated muscle. Nature, 235,533538, 197 1.
Jessop J, Lippold OC. Altered synchro~zation of motor unit firing as a mechanism for long- lasting increases in the tremor of human hand muscles following brief, strong effort [pmeedings]. J Physiol, 269,29P-30P, 1977.
Jones KE, Bawa P, McMiilan AS. Recmitment of motor units in buman flexor carpi ulnaris. Brain Res, 6û2,354-6, 1993.
Jones KE, Lyons M, Bawa P, Lemon RN. Recruitment order of motoneurons during functional tasks. Exp Bmin Res, 100,503-508, 1994.
Kanda K, Burke RE, WalmsIey B. Different control of fast and slow twitch motor units in the decerebraie cat. &p Brain Res, 29.57-74,1977.
Kossev A, Chnstova P. Discharge pattern of human motor uni& during dynamic concentric and eccentric contractions. Elechoencephdgr Clin NeurUphysiol, 1109,245-255, 1998.
Loeb GE. The conûol and responses of mammalian muscle spindles during normdy

executed motor tasks. Exerc Sport Sci Rev, 12,157-204, 1984.
Masakado Y, Karnen G, De Luca Ci. Effect of percutamous stimulation of motor unit firing behavior in man. Exp Brain Res, 86,426-432, 199 1.
Milner-Brown HS, Stein RB, Yemm R. The contractile properties of human motor units during voluntary isometric contractions. J Physiol, 228,285-306,1973a
Milner-Brown HS, Stein RD, Yemm R. The orderly recruitment of human motor units during voluntary isomeüic contractions. J Physiol, 230,359-370,1973b.
Milner-Brown HS, Stein RD, Yemm R. Changes in the firing rate of human motor units during linearly changing voluntary contraction. J Physiol, 230,371-390, 1973c.
Monster AW, Chan H. Isometric force production by motor units of extensor digitorum cornmunis muscle in man. J Neurophysiol, 40, 1432-1443,1977.
Moritani T, Muramats0 S, Muro M. Activity of motor units during concenûic and eccentric contractions. Am J Phys Med, 66,338-350, 1988.
Nardone A, Romano C, Schieppati M. Selective recniitrnent of hi&-threshold human motor units during voluntary isotonic lengthening of active muscles. J Physiol, 409,451-471, 1989.
Nardone A, Schieppati M. Shift of activity from slow ta fast muscle during voluntary lengthening contractions of the triceps surae muscles in humans. J Physiol, 395,363-381, 1988.
Riek S, Bawa P. Recruifment of motor units in human foram extensors. J Neurophysiol, 68, 100-108, 1992.
Senn W, Wyler K, Clamann HP, Kleinle J, Luscher Hi€, Muller L. Size principle and information theory. Bi01 Cyber, 76,ll-22, 1997.
Sogaard K, Christeasen H, Jensen BR, Finsen L, Sjogaard G. Motor control and kinetics during low level concentric and eccentric contractions in man. Electroencephalgr Clin Neurophysiol, 101,453-460,1996.
Stauber WT. Eccentric Action of Muscles: Physiology, Injury, and Adaptation. f ietc Sport Sci Rev, 17, 157-185, 1989.
Stephens JA, Garnett R, Buller NP. Reversal of recruitment order of single motor units producd by cutaneous stimulation during voluntary muscie contraction in man. Nature, 272,362-364,1978.

Tanji J, Kato M. Discharge of single motor units at voluntary contraction of abductor digiti minimi muscle in man. Exp Neurol, 40,771-783,1973.
Tax AA, Denier van der Gon JJ, Gielen CC, Kleyne M. Differences in centrai contcol of m. biceps brachii in movement tasks and force tasks. Exp Bmin Res, 79,138-42,1990.
Ter Haar Romeny BM, Denier van der Gon JJ, Gielen CCAM. Relation between location of a motor unit in the human biceps brachii and its critical finng levels for different tasks. Exp Neurol, 85,63 l-6SO,l984.
Thomas CK, Ross BH, Calancie B. Human motor-unit recniitment d u h g isometric contractions and repeated dynamic movements. J Neurophysiol, 57,3 1 1-324, 1987.
Yemm R. The orderly recniitment of motor units of the masseter and temporal muscles during voluntary isometric contraction in man. J Physioi, 265, 163-174, 1977.







