

Title:
Molecular cloning and characterization of coclaurine N-
methyltransferase from cultured cells of Coptis japonica*
Authors:
Kum-Boo Choi‡, Takashi Morishige‡, Nobukazu Shitan‡, Kazufumi
Yazaki‡, §, and Fumihiko Sato‡, §,**
Addresses:
‡, Division of Applied Life Sciences, Graduate School of Agriculture,
Kyoto University, Kyoto 606-8502, Japan
§, Division of Integrated Life Science, Graduate School of Biostudies,
Kyoto University, Kyoto 606-8502, Japan
**, To whom all correspondence should be addressed:
Dr. Fumihiko Sato
Tel:+81-(75)753-6380,
Fax:+81-(75)753-6398.
E-mail: [email protected]
Running title:
Coptis coclaurine N-methyltransferase
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on October 26, 2001 as Manuscript M106405200 by guest on July 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Summary
S-Adenosyl-L-methionine: coclaurine N-methyltransferase (CNMT)
converts coclaurine to N-methylcoclaurine in isoquinoline alkaloid
biosynthesis. The N-terminal amino acid sequence of Coptis CNMT was
used to amplify the corresponding cDNA fragment, and later to isolate
full-length cDNA using 5’- and 3’-RACE. The nucleotide sequence and
predicted amino acid sequence showed that the cDNA encoded 358
amino acids which contained a putative S-adenosyl-L-methionine
binding domain and showed relatively high homology to tomato
phosphoethanolamine-N-methyltransferase. Recombinant protein was
expressed in E. coli and its CNMT activity was confirmed. Recombinant
CNMT was purified to homogeneity and enzymological characterization
confirmed that Coptis CNMT has quite broad substrate specificity; i.e.
not only for 6-O-methylnorlaudanosoline and norreticuline but also for
6,7-dimethoxy-1,2,3,4-tetrahydroisoquinoline. The evolution of N-
methyltransferases in secondary metabolism is discussed based on
sequence similarity.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Introduction:
S-Adenosyl-L-methionine: coclaurine N-methyltransferase
(CNMT) (1-3) catalyzes the transfer of a methyl group from S-adenosyl-
L-methionine to the amino group of the tetrahydrobenzylisoquinoline
alkaloid coclaurine. This is a unique N-methyltransferase in the
biosynthesis of benzylisoquinoline alkaloids (Fig. 1). This enzyme is
thought to be important because N-methylation of coclaurine strongly
enhances the 4’-O-methylation activity of 3’-hydroxy-N-
methylcoclaurine 4’-O-methyltransferase and enables the sequential
metabolic conversion of substrates (4, 5). Furthermore, the enzymatic
activity at this important step is rather low relative to the entire
biosynthetic pathway (1, 6-8). Thus, we purified this CNMT and
characterized its properties. Previous studies have clearly indicated that
CNMT is non-stereospecific and has broad substrate specificity; Coptis
enzyme methylated even simple dihydroxyisoquinoline alkaloids.
Whereas several O-methyltransferases have been characterized at the
molecular level (4, 9-12), there have been very few molecular studies of
N-methyltransferases in secondary metabolite biosynthesis in plants (13,
14). The differences in the primary structures, including the S-adenosyl
methionine binding site, of NMTs and OMTs reported so far suggest
that molecular isolation of CNMT based on the structural similarity of
methyltransferases would be very difficult. Thus, we adapted the
conventional strategy to isolate cDNA based on the amino acid sequence
of purified enzyme. Whereas our purified CNMT fraction still contained
two protein bands of about 45 kD, careful inspection of the
chromatographic behavior of the proteins and enzyme activity suggested
that the slow-moving 45 kD polypeptide would encode CNMT (1).
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Based on this observation, we determined the N-terminal amino acids,
isolated the corresponding cDNA fragment, and finally the full-length
cDNA. The nucleotide sequence of cDNA suggested that the deduced
amino acid sequence showed some similarity to known NMTs such as
phosphoethanolamine NMT. Further characterization of recombinant
protein heterologously expressed in Escherichia coli confirmed that this
isolated cDNA encoded CNMT. Recombinant CNMT was purified to
homogeneity and its enzymological properties were characterized. The
evolution of CNMT involved in secondary metabolism is discussed
based on the sequence similarity of this novel NMT.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Experimental Procedures:
Plant material- Cultured Coptis japonica cells with high berberine
productivity were maintained as described previously (15). Ten-day-old
cultured cells were harvested and used for the extraction of mRNA.
Chemicals- Anabasine, (R,S)-1-[(3,4-dihydroxyphenyl)methyl]-
1,2,3,4-tetrahydro-6,7-isoquinolinediol (norlaudanosoline), 6,7-
dimethoxy-1,2,3,4-tetrahydroisoquinoline hydrochloride, 1,2,3,4-
tetrahydroisoquinoline, 1-methyl-6,7-dihydroxy-1,2,3,4-
tetrahydroisoquinolinehydrobromide, 1,2,3,4-tetrahydro-3-isoquinoline
carboxylic acid hydrochloride and (+)-emetine dihydrochroride hydrate
were purchased from Aldrich. (R)-Coclaurine and (S)-coclaurine were
gifts from Dr. N. Nagakura, Kobe Women’s College of Pharmacy. Other
alkaloids were gifts from Mitsui Petrochemical Industries Ltd. All other
chemicals used were of the highest purity available.
Amino acid sequence analysis- To determine N-terminal amino acid
sequences, 100 µg of a preparation of purified CNMT (1) was
electrotransferred on a polyvinylidene difluoride (PVDF) membrane
(Nihon Milipore, Yonezawa, Japan) from a gel after sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as described
elsewhere (9). A protein band of 45 kDa with slower mobility than an
accompanying band was cut from the PVDF membrane and the amino
acid sequence of the peptide was determined with a protein sequencer
(model 477A/120A; Applied Biosystems).
cDNA library construction of Coptis japonica- Total RNA was
prepared from 10 g of 10-day-old cultured Coptis cells and poly(A)+
RNA was isolated from 880 µg of total RNA using oligotex (dT)30-
Super (TaKaRa) according to the conventional protocol (16). A cDNA
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

library was constructed according to the method of Gubler and Hoffman
(17). In brief, the first strand of cDNA was synthesized using 3 µg of
poly(A)+ RNA, oligo (dT) linker-primer, RAV-2 reverse transcriptase
(Stratagene), and SuperScript II reverse transcriptase (Stratagene). After
the second strand of cDNA was synthesized, blunt-end adaptor
containing the EcoR I-Not I-BamH I restriction site was ligated into
synthesized cDNA, and the cDNA fragments were then ligated into
pDR196 vector (18).
Amplification of the cDNA fragment for the N-terminal sequence-
Degenerated primers were designed from the N-terminal amino acid
sequence determined and the codon usage estimated from known Coptis
sequences (4, 9). The sense primer USP was 5’-
GCIGTIGARGCIAARCARAC-3’ to AVEAKQT and the antisense
primer UAP was 5’-TCRACRAHTAYAGYAGYAT-3’ to YDDIKQL;
I= inosine, R=A or G, H=A or T or C, Y=C or T. The N-terminal cDNA
fragment was amplified for 30 cycles: 30 sec at 94 ºC, 30 sec at 50 ºC,
and 45 sec at 72 ºC. The PCR product was subcloned into pGEM-T
vector (Promega) and its nucleotide sequence was determined. To isolate
the 3’ fragment of CNMT cDNA, primers were designed based on the
nucleotide sequence of the amplified cDNA fragment and vector
sequence. The sense primer UFSP and the antisense primer UFAP were
5’-GCTGTGGAAGCAAAGCAAACAAAGAAGGCAGC-3’ and 5’-
ACGACTCACTATAGGGCGAATTGG-3’, respectively. Rapid
amplification of the 5’ end of cDNA (5’RACE) was carried out by the
procedure of Frohman (19) with a modification to obtain full-length
cDNA. The sense primer 5RSP for 5’ RACE was designed for a PMA1
promoter in the pDR196 vector (18). The antisense primers, 5RAP1 and
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

5RAP2, were designed for 5’RACE based on the cDNA sequence as
follows; 5’-CAATTGTGAATCCAAGGTCTCAACCTCTGTTGC-3’
and 5’-CTCTCGCAGTATAAATCCAGCATCGC-3’, respectively.
PCR was performed for 30 cycles using a step program; 30 sec at 94 ºC,
at 30 sec 52 ºC, and 30 sec at 72 ºC. PCR fragments were isolated and
subcloned into pGEM-T, and their nucleotide sequences were
determined.
Heterologous expression of CNMT- To express CNMT in E. coli,
full-length and partially truncated cDNAs were amplified by PCR using
30 cycles of 30 sec at 94 ºC, 90 sec at 60 ºC, and 30 sec at 72 ºC. Sense
primers (UESP: 5’-
GTTGCCATGGCTGTGGAAGCAAAGCAAACAAAGAAGGC-3’;
UESP-45N: 5’-
GTTGCCATGGACTTGTTAAAACAGTTGGAGCTGGGC-3’) were
designed to construct full-length and truncated cDNA and to introduce
an Nco I site (underlined). An antisense primer (UEAP; 5’-
GCGGAATTCACGACTCACTATAGGGCGAATTGG-3’) was used to
introduce an EcoR I site (underlined). The full-length cDNA was cloned
into the pET-21d vector (Novagen, WI) and its nucleotide sequence was
confirmed by DNA sequencing. Both expression vectors were
introduced in E. coli BL21 (DE3). Recombinant protein production was
induced in E. coli grown at 37 ºC with shaking in LB medium by adding
1 mM isopropylthiogalactoside, and E. coli was then incubated at 30 ºC
for an additional 3 h. Recombinant protein was extracted from E. coli in
100 mM Tris-HCl buffer (pH 7.5) containing 10 mM ascorbate and 20
mM 2-mercaptoethanol and used for the enzyme assay after desalting
through an NAP-5 column (Amersham Pharmacia Biotech).
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA sequence analysis- DNA sequencing was performed in both
directions in a DSQ-2000L automated sequencer (Shimadzu, Kyoto,
Japan) with a Thermosequence fluorescent-labeled primer cycle
sequencing kit (Amersham Pharmacia Biotech).
CNMT assay- CNMT activity was measured in 100 mM potassium
phosphate (pH 7.0), 25 mM sodium ascorbate, 0.1 mM norreticuline, 1
mM S-adenosyl-L-methionine (AdoMet), and the enzyme preparation at
30 ºC for 30 min. Although (S)-coclaurine is the true intermediate in
berberine biosynthesis, norreticuline was used as the substrate for
routine assays of CNMT due to its ready availability and similar product
formation in the reaction, as described previously (1). After the enzyme
reaction, proteins were denatured with methanol and the reaction
product was analyzed by HPLC (mobile phase, 35 % acetonitrile and
1 % acetic acid; column, LiChrospher 100 RP-18 (4 x 250 mm; Cica-
Merck); flow rate, 0.5 ml/min; detection, absorbance measurement at
280 nm). Product formation was confirmed by liquid chromatography-
mass spectroscopy (LCMS-2010, Shimadzu, Kyoto, Japan). To
determine the substrate specificity of CNMT, transfer of the 3H-labeled
methyl group of S-adenosyl-L-[methyl 3H]-methionine (NEN Life
Science Products, radioactivity; 1.5 MBq/µmol) to the product was
measured with 1 mM substrate as described previously. The activity was
shown as a relative incorporation value with 1 mM norreticuline as a
reference substance (1).
Purification of recombinant CNMT from E. coli lysate- Large
amounts of recombinant CNMT were produced in 600 ml culture of E.
coli. Unless otherwise noted, CNMT was purified as described
previously (1). In brief, CNMT was purified by Phenyl Sepharose, Q-
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Sepharose, and Mono P chromatography. All of the buffers used
contained 20 mM 2-mercaptoethanol and 10 % glycerol. The purified
enzyme was kept at -20 ºC in the presence of ca 40 % glycerol until use.
Other methods- The subunit molecular mass of the enzyme was
estimated by SDS-PAGE (10 % polyacrylamide), and the molecular
mass of the native enzyme was determined by gel filtration
chromatography on a Superose 12 column (Amersham Pharmacia
Biotech) in fast protein liquid chromatography. Protein concentration
was determined according to Bradford (20) with bovine serum albumin
as the standard.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Results:
Isolation of cDNA for CNMT- In our previous report (1), we purified
CNMT 340-fold from cultured Coptis cells, but this highly purified
fraction still contained two protein bands. Based on a detailed analysis
of the chromatographic elution profile and enzyme activity, we
estimated that the slow-moving polypeptide should correspond to
CNMT. To obtain molecular information on CNMT, purified CNMT
was blotted on PVDF membranes and its N-terminal amino acid
sequence was determined. These 33 amino acids were determined to be
as shown in Fig. 2.
Since several attempts to isolate full-length cDNA with degenerated
primers designed for this N-terminal sequence failed, we tried to amplify
the corresponding cDNA for this N-terminal amino acid sequence.
Primers were designed based on the codon usage predicted from known
Coptis genes (4, 9) and cDNA was amplified under conditions of rather
low stringency. cDNA fragments of a corresponding length (ca 100 bp)
were amplified, subcloned and sequenced. The nucleotide sequences of
these amplified fragments indicated that a predicted amino acid
sequence of a clone corresponded to the determined N-terminal
sequence of CNMT (data not shown).
Based on the nucleotide sequence determined in a cloned PCR
fragment, strict complementary primers were designed to obtain almost
a full-length 3’-fragment of CNMT. The amplified fragment (1.3 kb)
was subcloned into pGEM-T vector and sequenced. The amino acid
sequence predicted from the determined nucleotide sequence showed the
presence of a putative AdoMet binding domain, which indicated that the
correct sequence was amplified. A 5’-fragment of CNMT was obtained
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

by 5’-RACE using an internal nucleotide sequence in the 3’-fragment of
CNMT. After the sequence of this 5’-fragment was determined, full-
length cDNA was re-cloned from the cDNA library using UESP and
UEAP primers, and the nucleotide sequence was confirmed.
Nucleotide sequence and predicted amino acid sequences- Isolated
cDNA clone pCJCNMT1 contained 1,274 nucleotides with an open
reading frame that encoded 358 amino acids with a deduced Mr of
41,733 (DDBJ/GenBank/EMBL accession number AB061863) (Fig. 2).
Using the deduced amino acid sequence, we searched for homologous
sequences in the DDBJ/GenBank database using the BLAST search
program (http://blast.genome-ad.jp). BLAST search showed that the
deduced amino acid sequence had a relatively high similarity to
cyclopropane-fatty-acyl-phospholipid synthase of Mesorhizobium loti
(36 % identity; 108 aa/293 aa) and phosphoethanolamine N-
methyltransferase of Lycopersicon esculentum (28 % identity; 35 aa/124
aa), whereas the highest similarity (about 50 % identity; 152 aa/299 aa)
was found for a hypothetical protein of Arabidopsis thaliana (T05192).
O-Methyltransferases identified in berberine biosynthesis showed much
lower homology (about 10 % identity; 42 aa/358 aa). Phylogenic
analysis clearly indicated that OMTs from Coptis belong to a different
branch than CNMT (Fig. 3).
Expression of the recombinant polypeptide in E. coli and its CNMT
activity- Its amino acid sequence suggested that isolated cDNA would
encode CNMT of Coptis. To confirm the identity of this clone,
recombinant protein was produced in E. coli and enzyme activity was
determined. Enzyme assay with the crude E. coli lysate clearly showed
that the enzyme extract prepared from E. coli expressing pCJCNMT1
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

had methylation activity for norreticuline (data not shown), whereas the
extract of E. coli carrying the control pET-21d vector showed no
enzymatic activity. Further LC-MS analysis confirmed that recombinant
CNMT formed reticuline from norreticuline in the presence of AdoMet
(Fig. 4). These results indicated that the isolated clone correctly encodes
Coptis CNMT. Partially truncated CNMT was also produced to examine
the importance of the N-terminal sequence. This 15-amino acid deletion
did not change the enzyme activity, whereas the protein was not purified
and the specific activity was not determined.
Purification of recombinant CNMT and its characterization- Since
CNMT purified from cultured Coptis cells was not homogeneous, we
purified recombinant CNMT to homogeneity and characterized its
enzymological properties. Recombinant CNMT was successfully
purified to homogeneity from E. coli lysate only after 3 column
chromatographies but at a 5-fold higher yield than from cultured Coptis
cells (Table 1, Fig. 5). Finally, purified recombinant CNMT showed
much higher specific activity than native CNMT purified from Coptis
cells, probably due to the high expression in transformed E. coli and the
following rapid purification. Full-length CNMT was hydrophilic, as
predicted from a hydropathy plot (data not shown).
Since the specific activity was considerably different, we re-
examined the enzymological properties in comparison with those of
native CNMT purified from cultured Coptis cells. Enzyme assays at
various pH values indicated that the optimum pH for recombinant
CNMT activity was about 7.0. Half-maximal activity was found at pH
6.5 or 8.0. Recombinant CNMT, like native CNMT, did not require
divalent cations for activity. The addition of Cu2+, Co2+, or Mn2+ at 5
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

mM severely inhibited the activity by 52 %, 37 %, and 52 %
respectively. Other cations (Ca2+, Mg2+, and Fe2+) had no effect on
CNMT activity.
Substrate-saturation kinetics of purified recombinant CNMT for
norreticuline and AdoMet were the typical Michaelis-Menten. Therefore,
kinetic parameters were estimated from double-reciprocal plots of the
initial velocity versus the substrate concentration (data not shown). By
varying the concentration of norreticuline and [methyl-3H] AdoMet in
the range of 0.125 - 1 mM, a set of apparent Km and Vmax values could
be calculated and replotted to determine the actual Km and Vmax. The
respective Km values of recombinant CNMT for norreticuline and
AdoMet were 0.16 and 0.39 mM. The pattern of primary reciprocal plots
was representative of a sequential substrate binding mechanism. These
findings for recombinant CNMT indicated that the previous data
obtained for purified native CNMT (1) were reliable even though the
specific activity was considerably low. Our data also indicated that
recombinant CNMT should be quite useful for biotransformation of the
intermediates of alkaloid biosynthesis to N-methyltransfered products.
Substrate specificity of recombinant CNMT- Previously, we found
that Coptis CNMT had relatively broad substrate specificity. Since
heterologous expression in E. coli provided a sufficient amount of
recombinant enzyme, we examined the substrate specificity. The
incorporation of radioactivity from S-adenosyl-L-[methyl-3H]
methionine to the products was used to determine substrate specificity.
When norreticuline was the control substrate (i.e. relative incorporation
100 %), the respective relative activities with (R)-coclaurine and (S)-
coclaurine were 240 % and 153 %, which were comparable to values
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

reported previously. Recombinant CNMT methylated 6,7-dimethoxy-
1,2,3,4-tetrahydroisoquinoline as much as coclaurine and the
methylation product was confirmed by LC-MS (data not shown, Table
2). However, the methylation of 1,2,3,4-tetrahydroisoquinoline,
tryptamine, and purine was not detected either by measuring
radioactivity incorporation or by HPLC analysis (data not shown).
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Discussion:
Based on the N-terminal amino acid sequence of purified CNMT and
information on codon usage in Coptis genes, a full-length cDNA
encoding CNMT was successfully cloned and its enzymatic activity was
confirmed in a heterologous expression system. Recombinant CNMT
purified from transgenic E. coli showed a significantly higher activity
than that of CNMT purified from cultured Coptis cells. CNMT deduced
from the cDNA sequence was 358 amino acids long, with a calculated
molecular mass of 41 kDa, which was slightly lower than the apparent
molecular mass (45 kDa) of purified CNMT determined by SDS-PAGE.
SDS-PAGE analysis of purified recombinant CNMT revealed a similar
mobility for CNMT purified from Coptis cells, indicating that this
discrepancy between calculated molecular mass and apparent molecular
mass was not due to post-translational modification, but rather to
experimental variation, probably associated with the surface charge or
conformation of CNMT.
Since this CNMT sequence was the first reported NMT sequence in
isoquinoline alkaloid biosynthesis and the third reported NMT in
alkaloid biosynthesis in plants (13, 14), the sequences of these plant
alkaloid NMTs and other methyltransferases were compared to get more
information about the diversity of methyltransferases. Whereas several
plant O-methyltransferases have relatively high sequence homology,
ranging from 32 to 71 %, the diversity of N-methyltransferases is quite
high (Fig. 3); the sequence identity among NMTs is about 5-15 %. This
means that each NMT may have evolved from an independent origin.
CNMT showed relatively high homology to a hypothetical Arabidopsis
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

protein (T05192) and cyclopropane-fatty-acyl-phospholipid synthase of
Mesorhizobium (21), whereas the functional similarity is not clear.
The recent determination of the 3-dimensional structures of plant O-
methyltransferases suggested the structural conservation of an AdoMet
binding site among methyltransferases (22). As Chandrashekhar and
Vincent (23) have deduced from 56 plant enzyme sequences, motif A
(V/I/L)(V/L)(D/K)(V/I)GGXX(G/A) is present in all plant O-
methyltransferases with 0-2 mismatches. Motifs B and C,
(V/I/F)(A/P/E)X(A/P/G)DAXXXK(W/Y/F) and
(A/P/G/S)(L/I/V)(A/P/G/S)XX(A/P/G/S)(K/R)(V/I)(E/I)(L/I/V)
respectively, (X is any amino acid), are conserved 98 % with 0-3
mismatches. On the other hand, the AdoMet recognition domain was not
as highly conserved in NMTs, even though motif A was found among
almost all OMTs. In CNMT, only motif A is present with 2 mismatches
(Fig. 6). The lower conservation of the AdoMet binding domain in NMT
might be due to structural differences between NMTs and OMTs.
Whereas many OMTs have an AdoMet binding site in their C-terminal
half, motif A in CNMT and other NMTs is located at the N-terminal end
(24-26). Interestingly, a putative Arabidopsis protein (T05192) also has
this motif A, and cyclopropane-fatty-acyl-phospholipid synthase also
shows a similar identity in this region. Some enzymes in secondary
metabolism such as putrescine N-methyltransferase and tropinone
reductase have been postulated to have evolved from enzymes in
primary metabolism such as spermine synthase or the short-chain
dehydrogenase gene family (13, 27). Further characterization of the
enzyme activity of Arabidopsis T05192 and site-directed mutagenesis of
CNMT should provide clues for understanding the evolution of CNMT.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgments:
We greatly appreciate Dr. Fang-Sik Che of Nara Institute of Science
and Technology for N-terminal amino acid sequencing of CNMT. We
also thank Dr. N. Nagakura and Mitsui PetroChemical Industries Ltd. for
their generous gifts of the alkaloids. We are grateful to Dr. W. Frommer
of the University of Tuebingen for the gift of pDR196.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

References:
1. Choi, K.B., Morishige, T., and Sato, F. (2001) Phytochemistry 56,
649-655
2. Frenzel, T., and Zenk, M.H. (1990) Phytochemistry 29, 3491-3497
3. Wat, C.K., Steffens, P., and Zenk, M.H. (1986) Z. Naturforschung
41c, 126-134
4. Morishige, T., Tsujita, T., Yamada, Y., and Sato, F. (2000) J. Biol.
Chem. 275, 23398-23405
5. Stadler, R., and Zenk, M.H. (1993) J. Biol. Chem. 268, 823-831
6. Sato, F., Takeshita, N., Fitchen J.H., Fujiwara, H., and Yamada, Y.
(1993) Phytochemistry 32, 659-664
7. Sato, F., Tsujita, T., Katagiri, Y., Yoshida, S., and Yamada, Y.
(1994) Eur. J. Biochem. 225, 125-131
8. Yamada, Y., and Okada, N. (1985) Phytochemistry 24, 63-65
9. Takeshita, N., Fujiwara, H., Mimura, H., Fitchen, J.H., Yamada, Y.,
and Sato, F. (1995) Plant Cell Physiol. 36, 29-36
10. Ibrahim, R.K. (1997) Trends Plant Sci. 2, 249-250
11. Ibrahim R.K., Bruneau, A., and Bantignies, B (1998) Plant Mol. Biol.
36, 1-10
12. Frick, S., and Kutchan, T.M. (1999) Plant J. 17, 329-339
13. Hibi, N., Higashiguchi, S., Hashimoto, T., and Yamada, Y. (1994)
Plant Cell 6, 723-735
14. Kato, M., Mizuno, K., Crozier, A. Fujimura, T., and Ashihara, H.
(2000) Nature 406, 956-7
15. Sato, F., and Yamada, Y. (1984) Phytochemistry 23, 281-285
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

16. Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecular
Cloning: A Laboratory Manual (2nd ed.), Cold Spring Harbor
Laboratory, New York
17. Gubler, U., and Hoffman, B.J. (1983) Gene 25, 263-269
18. Rentsch, D., Laloi, M., Rouhara, I., Schmelzer, E., Delrot, S., and
Frommer, W.B. (1995) FEBS Lett. 370, 264-268
19. Frohman, M.A. (1990) PCR Protocols: a Guide to Methods and
Applications. (Gelfand, D.H., Sninsky, J., and White, T.J. eds) pp.28-38,
Academic Press, San Diego
20. Bradford, M.M. (1976) Anal. Biochem. 72, 248-254
21. Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,
Sasamoto, S., Watanabe, A., Idesawa, K., Ishikawa, A., Kawashima, K.,
Kimura, T., Kishida, Y., Kiyokawa, C., Kohara, M., Matsumoto, M.,
Matsuno, A., Mochizuki, Y., Nakayama, S., Nakazaki, N., Shimpo, S.,
Sugimoto, M., Takeuchi, C., Yamada, M., and Tabata, S. (2000) DNA
Res. 7, 331-338
22. Zubieta, C., He X.Z., Dixon, R.A., and Noel, J.P. (2001) Nature
Struct. Biol. 8, 271-279
23. Chandrashekhar, P.J., and Vincent, L.C. (1998) Plant Mol. Biol. 37,
663-674
24. Quaife, C.J., Hoyle, G.W., Froelick, G.J., Findley, S.D., Baetge, E.E.,
Behringer, R.R., Hammang, J.P., Brinster, R.L., and Palmiter, R.D.
(1994) Transgenic Res. 3, 388-400
25. Ying, Z., Mulligan, R.M., Janney, N., and Houtz, R.L. (1999) J. Biol.
Chem. 274, 36750-36756
26. Scoilas, A., Black, M.H., Talieri, M., and Diamandis, E.P. (2000)
Biochem. Biophys. Res. Commun. 278, 349-359
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

27. Nakajima K., Hashimoto, T., and Yamada, Y. (1993) Proc. Natl.
Acad. Sci. U.S.A. 90, 9591-9595
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Footnotes:
*This research was supported in part by a Grant-in-Aid B (08456172) for
Scientific Research from the Ministry of Education, Science, Culture and
Sports (Japan) and a Research for the Future Program grant (JSPS-RFTF
00L01607) from the Japan Society for Promotion of Science (F.S.).
1Abbreviations: AdoMet, S-adenosyl-L-methionine; CNMT,
coclaurine N-methyltransferase; LC-MS, liquid chromatography-mass
spectrometry; NMT, N-methyltransferase; OMT, O-methyltransferase;
PCR, polymerase chain reaction; 5’RACE, 5’ rapid amplification of
cDNA end; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure legends:
Fig. 1
Schematic biosynthetic pathway of berberine and the structure of
isoquinoline alkaloid tested as CNMT substrate (squared). 6OMT;
norcoclaurine 6-O-methyltransferase, CNMT; coclaurine N-
methyltransferase, 4’OMT; 3’-hydroxy-N-methylcoclaurine 4’-O-
methyltransferase, SMT; scoulerine 9-O-methyltransferase
Fig. 2
Nucleotide and deduced amino acid sequences of S-adenosyl-L-
methionine: coclaurine N-methyltransferase. The N-terminal amino
acid sequence determined from purified CNMT is boxed. Bold arrows
(USP, UAP) above the nucleotides indicate the sense and antisense
primers for N-terminal cloning of 100 bp. Thin arrows (UFSP, UESP-
45N) with dotted lines are PCR primers for the C-terminal fragment.
Thin arrows (5RAP1, 5RAP2) indicate the sense primers for 5’RACE.
Asterisk shows the stop codon. Motif A, a conserved sequence motif in
plant AdoMet-dependent methyltransferase, is shadowed.
Fig. 3
Phylogenic tree of N- and O-methyltransferases. Amino acid sequences
obtained from GenBankTM were used for tree building. Nine sequences were
aligned by the multisequence alignment program in GENETYX-MAX Ver.11
(Software Development Inc., Japan) using the UPGMA (unweighted pair
group maximum average) method.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Fig. 4
LC-MS analysis of recombinant CNMT reaction product (a), authentic
reticuline (b), and authentic norreticuline (c).
Fig. 5
SDS-PAGE analysis of purified CNMT.
(a), (b), and (c) show the analysis of crude extract of transformed E. coli,
purified recombinant CNMT, and native CNMT purified from Coptis cells,
respectively. Fractions purified on a Mono P column were analyzed by SDS-
PAGE in 10 % polyacrylamide gel and stained with Coomassie Brilliant Blue.
Fig. 6
Amino acid sequence alignment of the motif A domains of the N- and O-
methyltransferases in Coptis japonica, a hypothetical protein in
Arabidopsis thaliana, and cyclopropane-fatty-acyl-phospholipid synthase
in Mesorhizobium loti.
Motifs A is conserved sequence motif in plant S-adenosyl-L-methionine-
dependent methyltransferases. Amino acids conserved in all of the sequences
are boxed and similar ones are shadowed. CNMT; coclaurine N-
methyltransferase, cyclopropane; cyclopropane-fatty-acyl-phospholipid
synthase, 4’OMT; 3’-hydroxy-N-methylcoclaurine 4’-O-methyltransferase,
6OMT; norcoclaurine 6-O-methyltransferase, SMT; scoulerine 9-O-
methyltransferase
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from
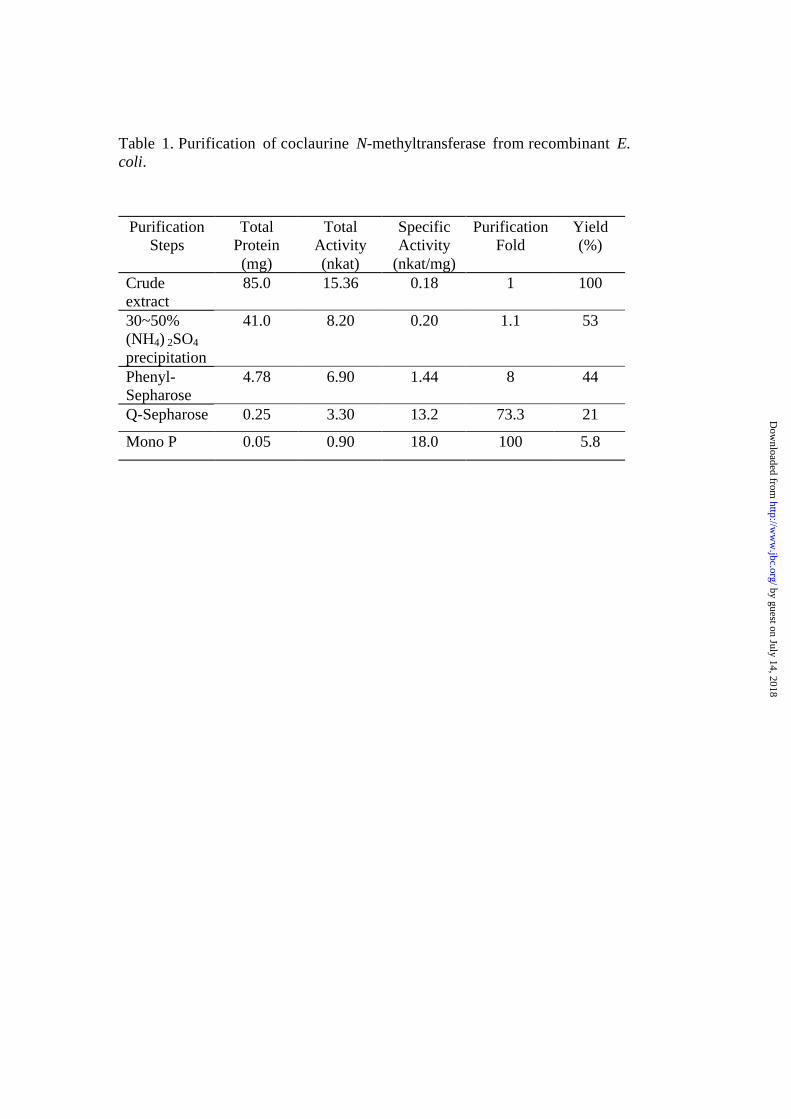
Table 1. Purification of coclaurine N-methyltransferase from recombinant E.coli.
PurificationSteps
TotalProtein(mg)
TotalActivity(nkat)
SpecificActivity
(nkat/mg)
PurificationFold
Yield(%)
Crudeextract
85.0 15.36 0.18 1 100
30~50%(NH4) 2SO4
precipitation
41.0 8.20 0.20 1.1 53
Phenyl-Sepharose
4.78 6.90 1.44 8 44
Q-Sepharose 0.25 3.30 13.2 73.3 21
Mono P 0.05 0.90 18.0 100 5.8
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Table 2. Substrate specificity of purified recombinant coclaurine N- methyltransferasea.
(R)-Coclaurine
(S)-Coclaurine
Relative
activity R1b R2 R3 R4
240 OMe OH H OH
153 OMe OH H OH
OMe OH OH OMe
OH OH OH OH
OMe OH OH OH
(R,S)-Norreticuline
(R,S)-Norlaudanosoline
(R,S)-6-O-
Methylnorlaudanosoline
(R,S)-Scoulerine
6,7-Dimethoxy-1,2,3,4-
tetrahydroisoquinoline
1-Methyl-6,7-dihydroxy-
1,2,3,4-tetrahydroisoquinoline
1,2,3,4-Tetrahydroisoquinoline
1,2,3,4-Tetrahydro-3-
isoquinoline carboxylic acid
(+)-Emetine
100
49
69
0
180
15
0
0
0
aThe enzyme reaction mixture was incubated for 10 min at 30 ˚C; total volume 50
µl containing 100 mM potassium phosphate (pH 7.0), 2.5 mM sodium ascorbate,
1 mM 3H-AdoMet and 20 µl of the purified Mono P fraction (ca. 0.2 µg protein).bSee Fig.1 for the numbering system.
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

NHHO
HO
OH
NHH3CO
HO
OH
NCH3
H3CO
HO
OH
NCH3
H3CO
HO
OH
NCH3
H3CO
HO
OCH3
NH3CO
HOH
H H H
H HOH
NH3CO
HO NR1
R2
OH OH
H
R4
OCH3
OCH3
OCH3
HR3
Berberine
(S )-Scoulerine
SMT
(S )-Tetrahydrocolumbamine
(S )-N-Methylcoclaurine
(S )-3'-Hydroxy-N-methylcoclaurine
4'OMT
(S )-Reticuline
CNMT
(S )-Coclaurine
6OMT
(S )-Norcoclaurine
L-Tyrosine
Fig. 1
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

M A V E A K Q T K K A A I V E L L
K Q L E L G L V P Y D D I K Q L I
R R E L A R R L Q W G Y K P T Y
E E Q I A E I Q N L T H S L R Q M
K I A T E V E T L D S Q L Y E I P
I E F L K I M N G S N L K G S C
C Y F K E D S T T L D E A E I A M
L D L Y C E R A Q I Q D G Q S V L
D L G C G Q G A L T L H V A Q K
Y K N C R V T A V T N S V S Q K E
Y I E E E S R R R N L L N V E V K
L A D I T T H E M A E T Y D R I
L V I E L F E H M K N Y E L L L R
K I S E W I S K D G L L F L E H I
C H K T F A Y H Y E P L D D D D
W F T E Y V F P A G T M I I P S A
S F F L Y F Q D D V S V V N H W T
L S G K H F S R T N E E W L K R
L D A N L D V I K P M F E T L M G
N E E E A V K L I N Y W R G F C L
S G M E M F G Y N N G E E W M A
S H V L F K K K *
ATGGCTGTGG AAGCAAAGCA AACAAAGAAG GCAGCCATAG TAGAGTTGTT
AAAACAGTTG GAGCTGGGCT TGGTTCCATA TGATGATATT AAGCAGCTCA
TAAGGAGGGA ACTGGCAAGG CGCCTGCAAT GGGGTTATAA ACCTACTTAT
GAAGAACAAA TAGCTGAAAT CCAAAACTTA ACTCATTCTC TGCGACAAAT
GAAAATTGCA ACAGAGGTTG AGACCTTGGA TTCACAATTG TACGAGATTC
CTATTGAGTT TCTAAAGATT ATGAATGGAA GTAACTTAAA AGGAAGTTGT
TGCTACTTCA AAGAAGATTC AACAACATTA GATGAAGCTG AGATAGCGAT
GCTGGATTTA TACTGCGAGA GAGCTCAAAT CCAAGATGGA CAGAGTGTTC
TTGATCTTGG ATGTGGGCAA GGAGCTCTTA CATTACATGT TGCACAGAAA
TATAAAAACT GTCGCGTAAC AGCAGTAACA AATTCAGTTT CACAAAAAGA
GTACATTGAA GAAGAATCAA GGAGACGTAA TTTGTTGAAT GTGGAAGTCA
AATTGGCAGA CATAACCACA CATGAGATGG CTGAGACATA CGATCGTATT
TTGGTAATAG AGTTGTTTGA GCACATGAAG AACTATGAAC TTCTCCTGAG
GAAAATCTCA GAGTGGATAT CGAAAGATGG GCTTCTCTTT CTAGAGCACA
TATGCCACAA GACCTTTGCT TACCACTATG AGCCTCTAGA CGACGACGAT
TGGTTTACAG AGTACGTGTT TCCTGCTGGG ACTATGATCA TACCATCTGC
ATCGTTCTTT TTGTATTTCC AGGATGACGT TTCGGTTGTG AACCATTGGA
CTCTTAGTGG GAAGCACTTT TCGCGTACCA ATGAGGAATG GTTGAAGAGA
TTGGACGCAA ACCTTGATGT TATTAAACCA ATGTTTGAGA CTTTAATGGG
AAATGAGGAA GAGGCAGTGA AGTTGATTAA CTATTGGAGA GGATTTTGTT
TATCTGGAAT GGAAATGTTT GGATATAACA ATGGTGAAGA ATGGATGGCA
AGTCATGTTC TGTTCAAGAA AAAATGA
USP
UFSP UESP-45N
UAP
5RAP1
5RAP2
Motif A
50
100
150
200
250
300
350
400
450
500
550
600
650
700
750
800
850
900
950
1000
1050
1077
Fig. 2
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

4'OMT(D29812;Coptis japonica)
putrescine NMT(AB004324;Nicotiana sylvestris)
phosphoethnolamine NMT(AAG59874;Lycopersicon esculentum)
hypothetic protein(T05192;Arabidopsis thaliana)
CNMT(AB061863;This work)
SMT(D29809;Coptis japonica)
6OMT(D29811;Coptis japonica)
caffeine synthase(AB031280;Camellia sinensis)
phenylethanolamine NMT(X52730-1;Human)
0.3308
0.3308
0.2562
0.5870
0.5740
0.1033
0.9199
0.9199
0.4216
1.1192
1.1192
0.1451
0.0772
0.8349
0.3261
0.3261
Figure 3.
0.3308
1.1192
0.1451
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

192299
330
(a)
330
299192
(b)
N
H3CO
HO
OH
OCH3
CH3
Reticuline m/z=330
N
H3CO
HOCH3
+
m/z=192
178299
316
(c)
N
H3CO
HO
OH
OCH3
HN
H3CO
HOH
+
Norreticuline m/z=316
m/z=178
Fig. 4
m/z
m/z
m/z
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

kDa
111
73
48
33
28
22
(a) (b) (c)
Fig. 5
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

hypothetical(T05192) VLDIGCGWGCNMT(AB061863) VLDLGCGQGSMT(D29809) LVDVGGGIG6OMT(D29811) LVDVGGGTG4'OMT(D29812) LVDVGGGNG
cyclopropane(BAB53730) ILELGCGWG
Fig. 6
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from

SatoKum-Boo Choi, Takashi Morishige, Nobukazu Shitan, Kazufumi Yazaki and Fumihiko
cultured cells of Coptis japonicaMolecular cloning and characterization of coclaurine N-methyltransferase from
published online October 26, 2001J. Biol. Chem.
10.1074/jbc.M106405200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 14, 2018http://w
ww
.jbc.org/D
ownloaded from







