jou rn al hom epage: www.elsev ier .com/ locate /bbr
esearch report
ex differences in diazepam effects and parvalbumin-positive GABAeurons in trait anxiety Long Evans rats
ebecca Ravenellea, Nichole M. Neugebauerb, Timothy Niedzielakc,. Tiffany Donaldsonc,∗
Department of Biological Sciences, Fordham University, Rose Hill Campus, Bronx, NY 10458, USADepartment of Psychiatry and Behavioral Sciences, Northwestern University, Feinberg School of Medicine, 303 E Chicago Avenue, #12-104, Chicago,
L 60611, USADevelopmental and Brain Sciences, Department of Psychology, University of Massachusetts Boston, 100 Morrissey Boulevard, Boston, MA 02125, USA
i g h l i g h t s
Female Long Evans rats bred for low anxiety phenotypes show less anxiety than males.Male animals were more sensitive to the anxiolytic effects of acute diazepam.Male rats show greater parvalbumin immunoreactivity (PV-IR) in the central amygdala.Females show greater PV-IR in the caudate putamen compared to males.
r t i c l e i n f o
rticle history:eceived 4 January 2014eceived in revised form 25 April 2014ccepted 28 April 2014vailable online 6 May 2014
eywords:nxietyABAiazepamlevated plus mazetriatummygdala
a b s t r a c t
In clinical populations, prevalence rates for a number of anxiety disorders differ between males andfemales and gonadal hormones are thought to contribute to these differences. While these hormoneshave been shown to modulate the anxiolytic effects of the benzodiazepine agonist diazepam in somemodels, findings are inconsistent. Here, we tested for sex differences in response to anxiogenic stimulifollowing a 30-min diazepam (1.0 mg/kg) pre-treatment in male and female rats showing high (HAn) andlow (LAn) anxiety-like behavior on the elevated plus maze. Acute diazepam administration resulted indecreased anxiety-like behavior only in HAn males as demonstrated by a significant increase in percentopen arm time in the elevated plus maze (EPM). Immunohistochemical analysis for parvalbumin (PV; acalcium-binding protein that selectively stains GABAergic neurons) in central amygdala (CeA), caudateputamen (CPu) and the hippocampus indicated the number of GABAergic interneurons in these areasdiffered across sex and anxiety trait. In the CPu, females had significantly more PV-immunoreactive (IR)cells than males, and LAn females had greater PV-IR neurons than HAn females. In the CeA, males displayed
an increased number of PV-IR neurons compared to females, with no differences found between LAn andHAn. Further, trait differences were evident in the CA2 region of the hippocampus, regardless of sex.Taken together, these data suggest that gonadal hormones and trait anxiety may influence the sensitivityto the anti-anxiety effects of diazepam and these differences may be due in part to the distribution of
urons
GABA-containing interne
. Introduction
It is estimated that 15% of the world’s population is sufferingrom an anxiety disorder [1] with lifetime prevalence rates at 19%
for males and 31% for females [2]. Pre-clinical models in whichanimals are selectively bred for enhanced anxiety reactions are ahelpful tool used in neurobiological studies of anxiety-like traits[3,4]. Although alterations in �-aminobutyric acid (GABA) trans-mission have been implicated, the exact mechanisms underlyinganxiety disorders remain elusive [5].
Classic benzodiazepines such as diazepam are commonly used
to treat anxiety disorders. Diazepam facilitates action at GABAAreceptors and these receptors are found throughout limbic brainregions, including the caudate putamen (CPu), the central nucleusof the amygdala (CeA) and the CA1 and CA2 regions of the
ippocampus [5,6]. Previous studies have illustrated that benzo-iazepines also modulate neuroendocrine function [7,8], however,he exact effects of diazepam on the HPA axis are not yet fullynderstood. Selectively bred male rats classified by high or lownxiety-like behavior show differential anxiolytic reactions toiazepam (1 mg/kg). While high anxiety-like behavior animalsxhibit a diazepam-induced decrease in anxious behavior as indi-ated by a 20-fold increase in percent open arm time on thelevated plus maze (EPM), the diazepam-induced decrease in anx-ous behavior in the low anxiety-like behavior animals is 2.5-fold9]. Although both anxiety lines exhibit elevated corticosteroneCORT) and adrenocorticotrophic hormone (ACTH) levels post EPMxposure, this increase is more pronounced in high anxiety-likeehavior animals [9].
Sex differences have been observed in anxiety disorders withemales exhibiting up to three times the prevalence rate for gen-ral anxiety disorder [10]. Animal models have also illustratedex differences in anxiety on several measures including the EPM,pen field and social interaction tests [11] suggesting a possiblenfluence of hormones on anxiety states. Both estrogen receptorER) forms alpha and beta have been implicated in GABAergicell regulation and amygdala GABAA cell densities are associatedith general activity on the EPM [12]. Female ERalpha knock-
ut (KO) mice exhibit increases in spontaneous activity whileRbeta KO mice spend less time on the open arms of the EPMndicating loss of ERbeta subtype increases anxiety-like behav-or [13,14]. Circulating gonadal hormones are also believed tonfluence sensitivity to benzodiazepines in humans [15] althoughn rats, previous work has found that intact females and malesoth show an increase in open arm time and entries follow-
ng acute and chronic diazepam administration [16]. Estrogenas been found to rapidly alter dopaminergic neurotransmis-ion through membrane-bound receptors on GABAergic neuronsn the CPu (for review, see [17]), an area that has been impli-ated in anxiety states in both clinical and animal models18,19].
Still other brain regions have been shown to contribute tohe behavioral expression of anxiety. Immediate early gene neu-al mapping of anxiogenic stimuli has revealed that in anxietyaradigms fear and stress areas often overlap [16] and acuteiazepam administration increases c-fos-immuonreactive (IR) cells
n the hypothalamic parventricular nucleus (PVN), CeA and theupraoptic nucleus (SON) [20]. High anxiety behavior animalshow a blunted c-fos IR response in the medial prefrontal cor-ex (mPFC) after diazepam administration compared to their lownxiety behavior counterparts [21]. The amygdala, an importantart of the limbic system and regulatory region for HPA axis acti-ation, shows differential c-fos-IR in trait anxiety animals withnhanced reactivity in high anxiety lines [22–25]. The hippocam-us has also been implicated in the regulation of the HPA axiss chronic stress induces dendritic atrophy and decreased glu-ocorticoid receptor expression in this region [26]. Moreover,ocal GABA neuron modulation has been described in many ofhe brain regions that show dysregulation in animals with traitnxiety, such as hippocampus [27] amygdala [28] and the CPu29].
In the present study, we set out to determine how sex andrait anxiety might interact to influence the anxiolytic actionf diazepam, and whether differences in the number of GABAnterneurons in the CeA, CPu or hippocampus might correlate withaseline and/or drug responses. We used females and males fromhe fourth filial generation of selectively outbred high (HAn) and
ow (LAn) anxiety Long Evans rats, tested in the EPM after bothehicle and diazepam administration and then measured the distri-ution of GABA neurons within regions implicated in the regulationf anxiety.
n Research 270 (2014) 68–74 69
2. Materials and methods
2.1. Experimental animals
Sixteen female (250–350 g) and sixteen male (500–700 g) adultLong Evans rats were obtained from pre-existing extreme trait anx-iety lines bred at the University of Massachusetts Boston. The traitanxiety lines were phenotyped on the elevated plus maze, we choseanimals that were in the lower quartile for percent time on openarms (OA) as high anxiety (HAn) and those in the upper quartilefor most %OA time as low anxiety (LAn). HAn and LAn females(n = 16 per group) were mated with unrelated same phenotypedmales. For the current study, all animals were procured from filialfour of the trait anxiety lines and were housed in standard Plex-iglas (48 cm × 25 cm × 22 cm) laboratory cages 2–4 rats per cageaccording to trait anxiety and sex resulting in four groups (n = 8 pergroup): HAn/female, HAn/male, LAn/female and LAn/male. Animalswere group housed according to weight guidelines outlined in theGuide for the Care and Use of Laboratory Animals and all protocolswere approved by the IACUC of the University of MassachusettsBoston.
2.2. Elevated plus maze
The EPM is a plus-shaped maze constructed of black Plex-iglas (Med Associates, VT) with two opposing open arms(50 cm × 6.5 cm) without walls and two opposing arms(50 cm × 6.5 cm) with high-enclosed walls (50 cm × 6.5 cm ×15 cm) connected via a central platform (6.5 cm × 6.5 cm). Themaze was set at an elevation of 70 cm above the floor. Animalswere placed at the central platform and allowed to explorethe maze for 5 min. Percent arm entries and percent open armtime were dependent measures of anxiety-like behavior and thetotal number of closed arm entries was used as an indicator oflocomotor activity. All data were recorded via computer usingautomated Med Associates software. Animals were tested on EPMthree separate times with at least three weeks between eachtesting session and in a novel room. This inter-trial interval (ITI)and novel maze placement was chosen based findings suggestingthat this ITI is sufficient to remove the one-trial tolerance [30].Testing sessions consisted of a baseline test where no drug wasadministered (Baseline) and two sessions where either diazepam(1 mg/kg) or vehicle (VEH) was administered 30 min prior to EPMtesting. Testing began at adulthood (PND 100) and continued atPND 145 and PND 166 for remaining EPM + VEH/Drug sessions.
2.3. Drug treatment
Prior to placement in the EPM (30 min) animals were admin-istered either diazepam (1 mg/kg/ml, IP) suspended in 1% Tween20/20 ml distilled water or equal volume vehicle injections of1% Tween-20/dH2O IP (VEH) using a within-subjects design. Thediazepam dose was chosen based on previously reported findings ofthe anxiolytic effects on EPM [5]. Animals were randomly assignedto drug condition in a counter-balanced manner to avoid order bias.On each test day, animals were transferred to the testing area andallowed to habituate for 15 min and then weighed before injection.
2.4. Transcardial perfusion
At the termination of the behavioral testing, all animalswere administered a lethal dose of Fatal-Plus (approximately
0.075–0.2 ml i.p.). Animals were transcardially perfused with 0.9%isotonic saline followed by 4% paraformaldehyde to fix the brain tis-sue. Brains were extracted, blocked and stored in a 20% sucrose–4%paraformaldehyde and then a 10% sucrose–4% paraformaldehyde
7 l Brain Research 270 (2014) 68–74
st4
2
fbpMlitafoLrat5cutrR
2
uiA(tfmwaWbawP
3
3
abietbpTas%wiO
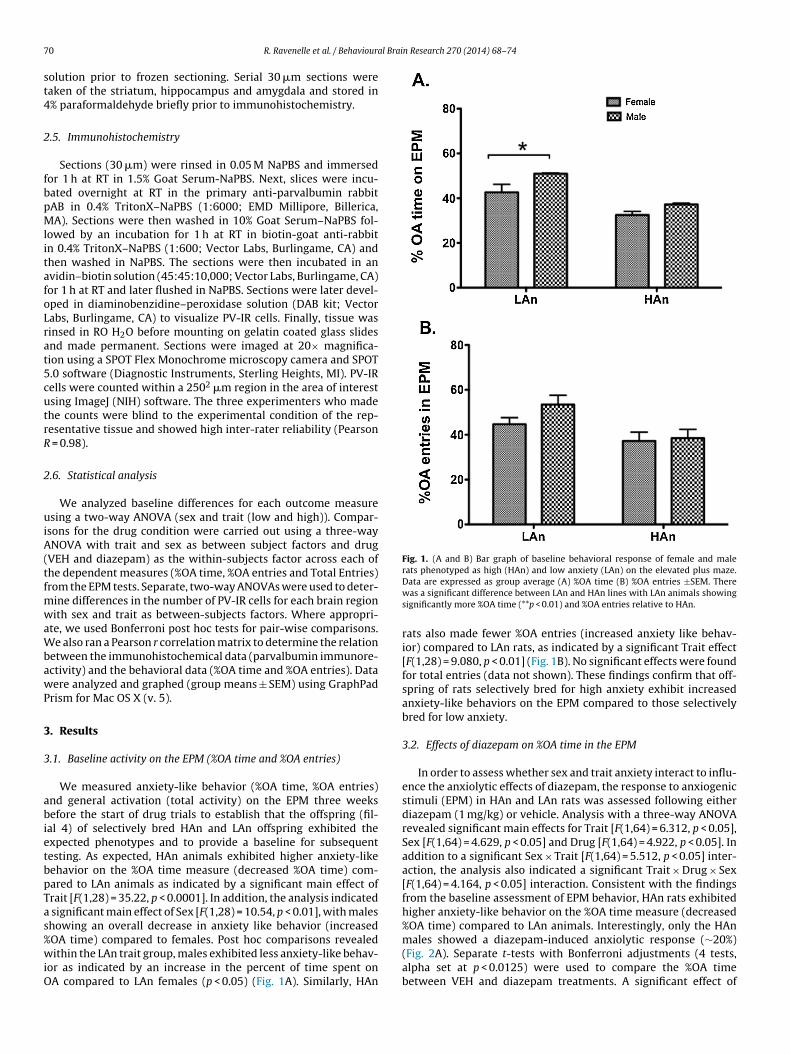
Fig. 1. (A and B) Bar graph of baseline behavioral response of female and malerats phenotyped as high (HAn) and low anxiety (LAn) on the elevated plus maze.
0 R. Ravenelle et al. / Behavioura
olution prior to frozen sectioning. Serial 30 �m sections wereaken of the striatum, hippocampus and amygdala and stored in% paraformaldehyde briefly prior to immunohistochemistry.
.5. Immunohistochemistry
Sections (30 �m) were rinsed in 0.05 M NaPBS and immersedor 1 h at RT in 1.5% Goat Serum-NaPBS. Next, slices were incu-ated overnight at RT in the primary anti-parvalbumin rabbitAB in 0.4% TritonX–NaPBS (1:6000; EMD Millipore, Billerica,A). Sections were then washed in 10% Goat Serum–NaPBS fol-
owed by an incubation for 1 h at RT in biotin-goat anti-rabbitn 0.4% TritonX–NaPBS (1:600; Vector Labs, Burlingame, CA) andhen washed in NaPBS. The sections were then incubated in anvidin–biotin solution (45:45:10,000; Vector Labs, Burlingame, CA)or 1 h at RT and later flushed in NaPBS. Sections were later devel-ped in diaminobenzidine–peroxidase solution (DAB kit; Vectorabs, Burlingame, CA) to visualize PV-IR cells. Finally, tissue wasinsed in RO H2O before mounting on gelatin coated glass slidesnd made permanent. Sections were imaged at 20× magnifica-ion using a SPOT Flex Monochrome microscopy camera and SPOT.0 software (Diagnostic Instruments, Sterling Heights, MI). PV-IRells were counted within a 2502 �m region in the area of interestsing ImageJ (NIH) software. The three experimenters who madehe counts were blind to the experimental condition of the rep-esentative tissue and showed high inter-rater reliability (Pearson
= 0.98).
.6. Statistical analysis
We analyzed baseline differences for each outcome measuresing a two-way ANOVA (sex and trait (low and high)). Compar-
sons for the drug condition were carried out using a three-wayNOVA with trait and sex as between subject factors and drug
VEH and diazepam) as the within-subjects factor across each ofhe dependent measures (%OA time, %OA entries and Total Entries)rom the EPM tests. Separate, two-way ANOVAs were used to deter-
ine differences in the number of PV-IR cells for each brain regionith sex and trait as between-subjects factors. Where appropri-
te, we used Bonferroni post hoc tests for pair-wise comparisons.e also ran a Pearson r correlation matrix to determine the relation
etween the immunohistochemical data (parvalbumin immunore-ctivity) and the behavioral data (%OA time and %OA entries). Dataere analyzed and graphed (group means ± SEM) using GraphPad
rism for Mac OS X (v. 5).
. Results
.1. Baseline activity on the EPM (%OA time and %OA entries)
We measured anxiety-like behavior (%OA time, %OA entries)nd general activation (total activity) on the EPM three weeksefore the start of drug trials to establish that the offspring (fil-
al 4) of selectively bred HAn and LAn offspring exhibited thexpected phenotypes and to provide a baseline for subsequentesting. As expected, HAn animals exhibited higher anxiety-likeehavior on the %OA time measure (decreased %OA time) com-ared to LAn animals as indicated by a significant main effect ofrait [F(1,28) = 35.22, p < 0.0001]. In addition, the analysis indicated
significant main effect of Sex [F(1,28) = 10.54, p < 0.01], with maleshowing an overall decrease in anxiety like behavior (increased
OA time) compared to females. Post hoc comparisons revealedithin the LAn trait group, males exhibited less anxiety-like behav-
or as indicated by an increase in the percent of time spent onA compared to LAn females (p < 0.05) (Fig. 1A). Similarly, HAn
Data are expressed as group average (A) %OA time (B) %OA entries ±SEM. Therewas a significant difference between LAn and HAn lines with LAn animals showingsignificantly more %OA time (**p < 0.01) and %OA entries relative to HAn.
rats also made fewer %OA entries (increased anxiety like behav-ior) compared to LAn rats, as indicated by a significant Trait effect[F(1,28) = 9.080, p < 0.01] (Fig. 1B). No significant effects were foundfor total entries (data not shown). These findings confirm that off-spring of rats selectively bred for high anxiety exhibit increasedanxiety-like behaviors on the EPM compared to those selectivelybred for low anxiety.
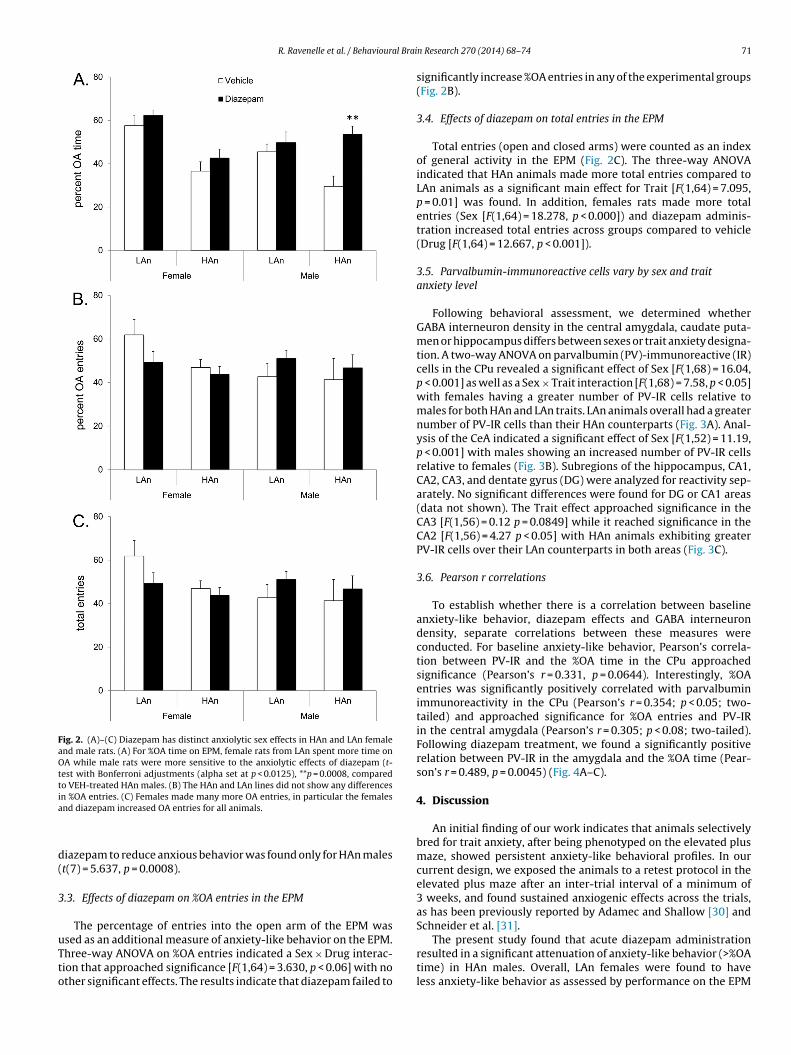
3.2. Effects of diazepam on %OA time in the EPM
In order to assess whether sex and trait anxiety interact to influ-ence the anxiolytic effects of diazepam, the response to anxiogenicstimuli (EPM) in HAn and LAn rats was assessed following eitherdiazepam (1 mg/kg) or vehicle. Analysis with a three-way ANOVArevealed significant main effects for Trait [F(1,64) = 6.312, p < 0.05],Sex [F(1,64) = 4.629, p < 0.05] and Drug [F(1,64) = 4.922, p < 0.05]. Inaddition to a significant Sex × Trait [F(1,64) = 5.512, p < 0.05] inter-action, the analysis also indicated a significant Trait × Drug × Sex[F(1,64) = 4.164, p < 0.05] interaction. Consistent with the findingsfrom the baseline assessment of EPM behavior, HAn rats exhibitedhigher anxiety-like behavior on the %OA time measure (decreased%OA time) compared to LAn animals. Interestingly, only the HAn
males showed a diazepam-induced anxiolytic response (∼20%)(Fig. 2A). Separate t-tests with Bonferroni adjustments (4 tests,alpha set at p < 0.0125) were used to compare the %OA timebetween VEH and diazepam treatments. A significant effect of
R. Ravenelle et al. / Behavioural Brai
Fig. 2. (A)–(C) Diazepam has distinct anxiolytic sex effects in HAn and LAn femaleand male rats. (A) For %OA time on EPM, female rats from LAn spent more time onOA while male rats were more sensitive to the anxiolytic effects of diazepam (t-test with Bonferroni adjustments (alpha set at p < 0.0125), **p = 0.0008, comparedto VEH-treated HAn males. (B) The HAn and LAn lines did not show any differencesia
d(
3
uTto
n %OA entries. (C) Females made many more OA entries, in particular the femalesnd diazepam increased OA entries for all animals.
iazepam to reduce anxious behavior was found only for HAn malest(7) = 5.637, p = 0.0008).
.3. Effects of diazepam on %OA entries in the EPM
The percentage of entries into the open arm of the EPM was
sed as an additional measure of anxiety-like behavior on the EPM.hree-way ANOVA on %OA entries indicated a Sex × Drug interac-ion that approached significance [F(1,64) = 3.630, p < 0.06] with nother significant effects. The results indicate that diazepam failed to
n Research 270 (2014) 68–74 71
significantly increase %OA entries in any of the experimental groups(Fig. 2B).
3.4. Effects of diazepam on total entries in the EPM
Total entries (open and closed arms) were counted as an indexof general activity in the EPM (Fig. 2C). The three-way ANOVAindicated that HAn animals made more total entries compared toLAn animals as a significant main effect for Trait [F(1,64) = 7.095,p = 0.01] was found. In addition, females rats made more totalentries (Sex [F(1,64) = 18.278, p < 0.000]) and diazepam adminis-tration increased total entries across groups compared to vehicle(Drug [F(1,64) = 12.667, p < 0.001]).
3.5. Parvalbumin-immunoreactive cells vary by sex and traitanxiety level
Following behavioral assessment, we determined whetherGABA interneuron density in the central amygdala, caudate puta-men or hippocampus differs between sexes or trait anxiety designa-tion. A two-way ANOVA on parvalbumin (PV)-immunoreactive (IR)cells in the CPu revealed a significant effect of Sex [F(1,68) = 16.04,p < 0.001] as well as a Sex × Trait interaction [F(1,68) = 7.58, p < 0.05]with females having a greater number of PV-IR cells relative tomales for both HAn and LAn traits. LAn animals overall had a greaternumber of PV-IR cells than their HAn counterparts (Fig. 3A). Anal-ysis of the CeA indicated a significant effect of Sex [F(1,52) = 11.19,p < 0.001] with males showing an increased number of PV-IR cellsrelative to females (Fig. 3B). Subregions of the hippocampus, CA1,CA2, CA3, and dentate gyrus (DG) were analyzed for reactivity sep-arately. No significant differences were found for DG or CA1 areas(data not shown). The Trait effect approached significance in theCA3 [F(1,56) = 0.12 p = 0.0849] while it reached significance in theCA2 [F(1,56) = 4.27 p < 0.05] with HAn animals exhibiting greaterPV-IR cells over their LAn counterparts in both areas (Fig. 3C).
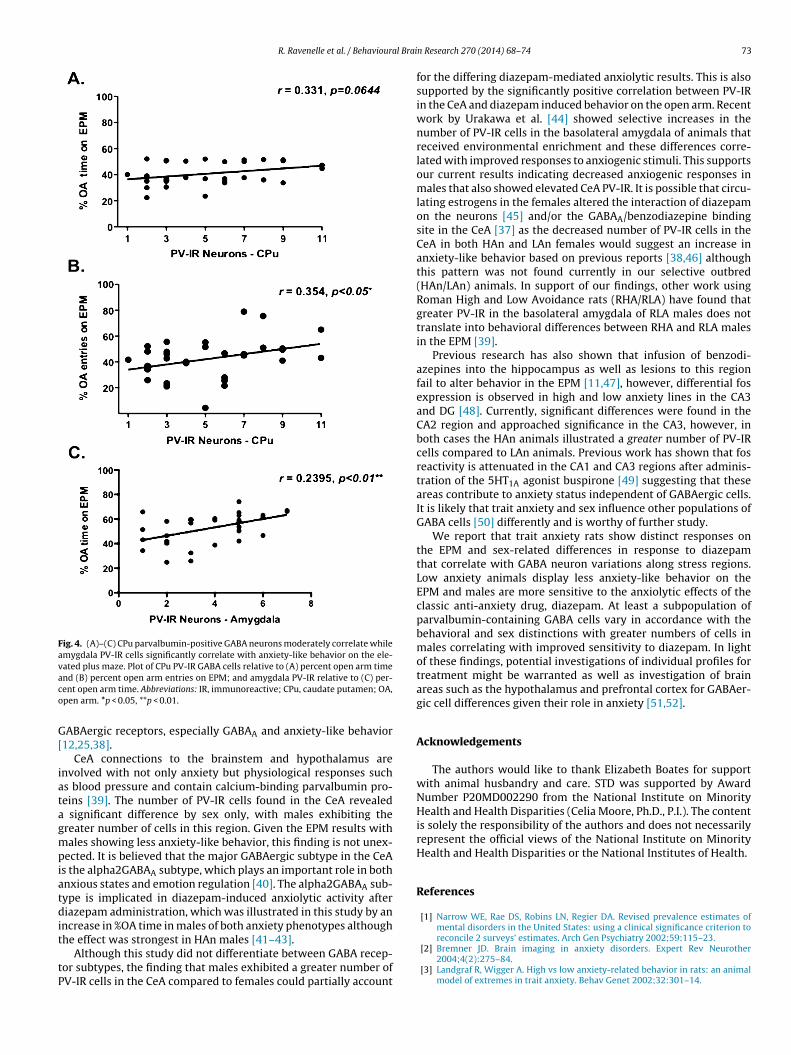
3.6. Pearson r correlations
To establish whether there is a correlation between baselineanxiety-like behavior, diazepam effects and GABA interneurondensity, separate correlations between these measures wereconducted. For baseline anxiety-like behavior, Pearson’s correla-tion between PV-IR and the %OA time in the CPu approachedsignificance (Pearson’s r = 0.331, p = 0.0644). Interestingly, %OAentries was significantly positively correlated with parvalbuminimmunoreactivity in the CPu (Pearson’s r = 0.354; p < 0.05; two-tailed) and approached significance for %OA entries and PV-IRin the central amygdala (Pearson’s r = 0.305; p < 0.08; two-tailed).Following diazepam treatment, we found a significantly positiverelation between PV-IR in the amygdala and the %OA time (Pear-son’s r = 0.489, p = 0.0045) (Fig. 4A–C).
4. Discussion
An initial finding of our work indicates that animals selectivelybred for trait anxiety, after being phenotyped on the elevated plusmaze, showed persistent anxiety-like behavioral profiles. In ourcurrent design, we exposed the animals to a retest protocol in theelevated plus maze after an inter-trial interval of a minimum of3 weeks, and found sustained anxiogenic effects across the trials,as has been previously reported by Adamec and Shallow [30] andSchneider et al. [31].
The present study found that acute diazepam administrationresulted in a significant attenuation of anxiety-like behavior (>%OAtime) in HAn males. Overall, LAn females were found to haveless anxiety-like behavior as assessed by performance on the EPM
72 R. Ravenelle et al. / Behavioural Brain Research 270 (2014) 68–74
Fig. 3. (A)–(C) GABA neurons containing the calcium binding protein parvalbumin (PV) were counted in several brain areas implicated in anxiety. (A) Females showed ag istentm campaC
(pgammatd
acmbHsfsd
opn
reater number of PV-IR in the CPu, regardless of trait, #p < 0.05. LAn animals consales had the most PV-IR cells. (C) HAn PV-IR counts were greater than LAn in hippo
eA, central amygdala.
>%OA time, >%OA entries) than HAn females, however, diazepamretreatment did not reduce anxious behavior in either femaleroup. These findings are supported by previous work showing
1.0 mg/kg dose of diazepam decreased anxiety-like behavior inales but not females [32]. While it is possible that the acute treat-ent in this study (single 1 mg/kg injection) was too low to elicit an
nxiolytic response in females, our work is consistent with a reporthat found no changes on EPM in females administered an acuteose of diazepam (single 2.5 mg/kg injection) [16].
As expected, LAn animals showed less anxiety-like behavior rel-tive to HAn animals and this effect was more pronounced in malesompared to females. Following the diazepam challenge, LAn ani-als in general spent more time on OA and made more OA entries
ut the only significant anxiolytic effect was found in HAn males.owever, analysis of total entries revealed that HAn females made
ignificantly more total entries than LAn females even though LAnemales had greater %OA entries. This suggests that HAn and LAntatus is likely reflecting differences in anxiety level as opposed toifferences in exploration or locomotor activity in the EPM.
The effects of the estrous cycle on anxiety could explain thebserved sex differences as previous work illustrates that duringroestrous (i.e., elevated levels of progestin and plasma allopreg-anolone) females show decreased anxiety-like behavior on the
ly showed greater PV-IR relative to HAn, ***p < 0.001. (B) In the central amygdala,l regions (CA2 and CA3). Abbreviations: IR, immunoreactive; CPu, caudate putamen;
EPM [33]. Allopregnanolone is a direct GABAA receptor modulatorthat acts as an anxiolytic with rebound anxiety observed dur-ing withdrawal from the treatment [34]. Although we found anincrease in total entries after diazepam administration in females,it failed to act as an anxiolytic. Gonadal hormones may have playedan important role as estradiol and progesterone have been found tomodify the number of GABA receptors in the brain and possibly alterGABA–benzodiazepine binding sites [35,36]. While fluctuations inhormone levels were not a focus of the current work, it is a limita-tion that we did not measure estrogen levels since they have beenshown to interact with the effects of diazepam on avoidance con-ditioning and locomotor activity in rats [37]. Analysis of PV-IR cells(PV targets a subpopulation of GABAergic cells that contain this cal-cium binding protein) in the CPu revealed a significant difference bysex with females exhibiting a greater number of PV-IR cells in bothHAn and LAn lines relative to males. It is possible that sex differ-ences in the CPu can be attributed to estradiol acting on GABAergicneurons [17]. In the current study, LAn animals were found tohave an increased number of PV-IR cells compared to HAn animals
in the CPu. Consistent with this finding, less anxiety-like behav-ior was positively correlated with increased GABA neuron densityin this region. This finding is in agreement with previous workthat has suggested a negative correlation between the number of
R. Ravenelle et al. / Behavioural Brai
Fig. 4. (A)–(C) CPu parvalbumin-positive GABA neurons moderately correlate whileamygdala PV-IR cells significantly correlate with anxiety-like behavior on the ele-vated plus maze. Plot of CPu PV-IR GABA cells relative to (A) percent open arm timeaco
G[
iatagmpiatdit
tP
nd (B) percent open arm entries on EPM; and amygdala PV-IR relative to (C) per-ent open arm time. Abbreviations: IR, immunoreactive; CPu, caudate putamen; OA,pen arm. *p < 0.05, **p < 0.01.
ABAergic receptors, especially GABAA and anxiety-like behavior12,25,38].
CeA connections to the brainstem and hypothalamus arenvolved with not only anxiety but physiological responses suchs blood pressure and contain calcium-binding parvalbumin pro-eins [39]. The number of PV-IR cells found in the CeA revealed
significant difference by sex only, with males exhibiting thereater number of cells in this region. Given the EPM results withales showing less anxiety-like behavior, this finding is not unex-
ected. It is believed that the major GABAergic subtype in the CeAs the alpha2GABAA subtype, which plays an important role in bothnxious states and emotion regulation [40]. The alpha2GABAA sub-ype is implicated in diazepam-induced anxiolytic activity afteriazepam administration, which was illustrated in this study by an
ncrease in %OA time in males of both anxiety phenotypes although
he effect was strongest in HAn males [41–43].
Although this study did not differentiate between GABA recep-or subtypes, the finding that males exhibited a greater number ofV-IR cells in the CeA compared to females could partially account
n Research 270 (2014) 68–74 73
for the differing diazepam-mediated anxiolytic results. This is alsosupported by the significantly positive correlation between PV-IRin the CeA and diazepam induced behavior on the open arm. Recentwork by Urakawa et al. [44] showed selective increases in thenumber of PV-IR cells in the basolateral amygdala of animals thatreceived environmental enrichment and these differences corre-lated with improved responses to anxiogenic stimuli. This supportsour current results indicating decreased anxiogenic responses inmales that also showed elevated CeA PV-IR. It is possible that circu-lating estrogens in the females altered the interaction of diazepamon the neurons [45] and/or the GABAA/benzodiazepine bindingsite in the CeA [37] as the decreased number of PV-IR cells in theCeA in both HAn and LAn females would suggest an increase inanxiety-like behavior based on previous reports [38,46] althoughthis pattern was not found currently in our selective outbred(HAn/LAn) animals. In support of our findings, other work usingRoman High and Low Avoidance rats (RHA/RLA) have found thatgreater PV-IR in the basolateral amygdala of RLA males does nottranslate into behavioral differences between RHA and RLA malesin the EPM [39].
Previous research has also shown that infusion of benzodi-azepines into the hippocampus as well as lesions to this regionfail to alter behavior in the EPM [11,47], however, differential fosexpression is observed in high and low anxiety lines in the CA3and DG [48]. Currently, significant differences were found in theCA2 region and approached significance in the CA3, however, inboth cases the HAn animals illustrated a greater number of PV-IRcells compared to LAn animals. Previous work has shown that fosreactivity is attenuated in the CA1 and CA3 regions after adminis-tration of the 5HT1A agonist buspirone [49] suggesting that theseareas contribute to anxiety status independent of GABAergic cells.It is likely that trait anxiety and sex influence other populations ofGABA cells [50] differently and is worthy of further study.
We report that trait anxiety rats show distinct responses onthe EPM and sex-related differences in response to diazepamthat correlate with GABA neuron variations along stress regions.Low anxiety animals display less anxiety-like behavior on theEPM and males are more sensitive to the anxiolytic effects of theclassic anti-anxiety drug, diazepam. At least a subpopulation ofparvalbumin-containing GABA cells vary in accordance with thebehavioral and sex distinctions with greater numbers of cells inmales correlating with improved sensitivity to diazepam. In lightof these findings, potential investigations of individual profiles fortreatment might be warranted as well as investigation of brainareas such as the hypothalamus and prefrontal cortex for GABAer-gic cell differences given their role in anxiety [51,52].
Acknowledgements
The authors would like to thank Elizabeth Boates for supportwith animal husbandry and care. STD was supported by AwardNumber P20MD002290 from the National Institute on MinorityHealth and Health Disparities (Celia Moore, Ph.D., P.I.). The contentis solely the responsibility of the authors and does not necessarilyrepresent the official views of the National Institute on MinorityHealth and Health Disparities or the National Institutes of Health.
References
[1] Narrow WE, Rae DS, Robins LN, Regier DA. Revised prevalence estimates ofmental disorders in the United States: using a clinical significance criterion to
reconcile 2 surveys’ estimates. Arch Gen Psychiatry 2002;59:115–23.
[anxiety and hyporeactivity to stress of the dorsomedial prefrontal cortex: acombined phMRI and Fos study in rats. Neuroimage 2004;23:382–91.
4 R. Ravenelle et al. / Behavioura
[4] Landgraf R, Wigger A. Born to be anxious: neuroendocrine and genetic corre-lates of trait anxiety in HAB rats. Stress 2003;6:111–9.
[5] Heidt SA, Mou L, Ressler KJ. In vivo knockdown of GAD67 in the amygdala dis-rupts fear extinction and the anxiolytic-like effect of diazepam in mice. TranslPsychiatry 2012;13(2), http://dx.doi.org/10.1038/tp.2012.101.
[6] Bristow DR, Martin IL. GABA preincubation of rat brain sections increases[3H]GABA binding to the GABAA receptor and compromises the modulatoryinteractions. Eur J Pharmacol 1988;148:283–8.
[7] De Souza EB. Neuroendocrine effects of benzodiazepines. J Psychiatr Res1990;24:111–9.
[8] Giordano R, Grottoli S, Brossa P, Pellegrino M, Destefanis S, Lanfranco F,et al. Alprazolam (a benzodiazepine activating GABA receptor) reduces neu-roendocrine responses to insulin-induced hypoglycaema in humans. ClinEndocrinol (Oxf) 2003;59:314–20.
[9] Liebsch G, Linthorst ACE, Neumann ID, Reul JMHM, Holsboer F, Landgraf R.Behavioral, physiological, and neuroendocrine stress responses and differentialsensitivity to diazepam in two Wistar rat lines selectively bred for high and lowanxiety related behavior. Neuropsychopharmacology 1998;19:381–96.
10] Yonkers KA, Ellison JM. Anxiety disorders in women and their pharmacologicaltreatment. In: Jensvold MF, Halbreich U, Hamilton JA, editors. Psychophar-macology and women. Washington, DC: American Psychiatric Press; 1996. p.261–85.
11] Gonzalez LE, Ouagazzal AM, File SE. Stimulation of benzodiazepine recep-tors in the dorsal hippocampus and median raphe reveals differentialGABAergic control in two animal tests of anxiety. Eur J Neurosci 1998;10:3673–780.
12] Yilmazer-Hanke DM, Roskoden T, Zilles K, Schwegler H. Anxiety related behav-ior and densities of glutamate GABAa, acetylcholine and serotonin receptors inthe amygdala of seven inbred mouse strains. Behav Brain Res 2003;145:145–59.
13] Krezel W, Dupont S, Krust A, Chambon P, Chapman PF. Increased anxiety andsynaptic plasticity in estrogen receptor �-deficient mice. Proc Natl Acad Sci US A 2001;98:12278–82.
14] Ogawa S, Chan J, Gustafsson JA, Korach KS, Pfaff DW. Estrogen increases loco-motor activity in mice through estrogen receptor alpha: specificity for the typeof activity. Endocrinology 2003;144:230–9.
15] Sundström I, Ashbrook D, Bäckström T. Reduced benzodiazepine sensitivity inpatients with premenstrual syndrome: a pilot study. Psychoneuroendocrinol-ogy 1997;22:25–38.
16] Singewald N. Altered brain activity processing in high-anxiety rodents revealedby challenge paradigms and functional mapping. Neurosci Biobehav Rev2007;31:18–40.
17] Becker JB. Gender differences in dopaminergic function in striatum and nucleusaccumbens. Pharmacol Biochem Behav 1999;64:803–12.
18] Favilla C, Abel T, Kelly MP. Chronic G(alpha)s signaling in the striatumincreases anxiety-related behaviors independent of development. J Neurosci2008;28:13952–6.
19] Hasler G, Fromm S, Alvarez RP, Luckenbaugh DA, Drevets WC, Grillon C. Cerebralblood flow in immediate and sustained anxiety. J Neurosci 2007;27:6313–9.
20] Rodgers RJ, Dalvi A. Anxiety, defence and the elevated plus-maze. NeurosciBiobehav Rev 1997;21:801–10.
21] Herman JP, Ostrander MM, Mueller NK, Figueredo H. Limbic system mecha-nisms of stress regulation: hypothalamic–pituitary–adrenocortical axis. ProgrNeuropscyhopharmacol Biol Psychiatry 2005;29:1201–13.
22] Anand A, Shekhar A. Brain imaging studies in mood and anxiety disor-ders: a special emphasis on the amygdala. Ann N Y Acad Sci 2003;985:370–88.
23] Blanchard DC, Canteras NS, Markham CM, Pentkowski NS, Blanchard RJ. Lesionsof structures showing FOS expression to cat presentation: effect on respon-siveness to a cat, cat odor, and nonpredator threat. Neurosci Biobehav Rev2005;29:1243–53.
24] Frank E, Salchner P, Aldag JM, Salomé N, Singewald N, Landgraf R, et al. Geneticpredisposition to anxiety-related behavior determines coping style, neuroen-docrine response, and neuronal activation during social defeat. Behav Neurosci2006;120:60–71.
25] Kalueff AV, Nutt DJ. Role of GABA in anxiety and depression. Depress Anxiety2007;24:495–517.
26] Ulrich-Lai YM, Herman JP. Neural regulation of endocrine and autonomic stressresponses. Nat Neurosci 2009;10:397–409.
27] Klausberger T, Magill PJ, Márton LF, Roberts JDB, Cobden PM, Buzsáki G, et al.Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo.Nat Neurosci 2003;421:844–8.
28] Marowsky A, Yanagawa Y, Obata K, Vogt KE. A specialized subclass of interneu-rons mediates dopaminergic function. Neuron 2005;48:1025–37.
[
n Research 270 (2014) 68–74
29] Teper JM, Bolam P. Functional diversity and specificity of neostriatal interneu-rons. Curr Opin Neurobiol 2004;14:685–92.
30] Adamec R, Shallow T. Effects of baseline anxiety on response to kindling of theright medial amygdala. Physiol Behav 2000;70:67–80.
31] Schneider P, Ho Y-J, Spanagel R, Pawlak CR. A novel elevated plus-mazeprocedure to avoid the one-trial tolerance problem. Front Neurosci 2011;5:1–8.
32] Wilson MA, Burghardt PR, Ford KA, Wilkinson MB, Stefany D. Anxiolytic effectsof diazepam and ethanol in two behavioral models: comparison of males andfemales. Pharmacol Biochem Behav 2004;78:455–8.
33] Frye CA, Petralia SM, Rhodes ME. Estrous cycle and sex differences in perfor-mance on anxiety tasks coincide with increases in hippocampal progesteroneand 3alpha, 5alpha-THP. Pharmacol Biochem Behav 2000;67:587–96.
34] Smith SS. Withdrawal properties of a neuroactive steroid: implications forGABAa receptor gene regulation in the brain and anxiety behavior. Steroids2002;67:519–28.
35] Maggi A, Perez J. Progesterone and estrogens in the rat brain: modulation ofGABA receptor activity. Eur J Pharmacol 1984;103:165–8.
36] Perez J, Zucchi I, Maggi A. Estrogen modulation of the gamma-aminobutyricacid receptor complex in the central nervous system of the rat. J PharmacolExp Ther 1988;244:1005–10.
37] Díaz-Véliz G, Butrón S, Venavides MS, Dussaubat N, Mora S. Gender, estrouscycle, ovariectomy, and ovarian hormones influence the effects of diazepam onavoidance conditioning in rats. Pharmacol Biochem Behav 2000;66:887–92.
38] Rägo L, Kiivet RA, Harro J, Pôld M. Behavioral differences in an elevated plus-maze: correlation between anxiety and a decreased number of GABA andbenzodiazepine receptors in mouse cerebral cortex. Naunyn Schmiedeberg’sArch Pharmacol 1988;337:675–8.
39] Yilmazer-Hanke DM, Faber-Zuschratter H, Linke R, Schwegler H. Contributionof amygdala neurons containing peptides and calcium-binding proteins to fear-potentiated startle and exploration-related anxiety in inbred Roman high andlow-avoidance rats. Eur J Neurosci 2002;15:1206–18.
40] Fritschy JM, Möhler H. GABAERGIC-receptor heterogeneity in the adult ratbrain: differential regional and cellular distribution of seven major subunits.J Comp Neurol 1995;359:154–94.
41] Crestani F, Keist R, Fritschy JM, Benke D, Vogt K, Prut L, et al. Trace fear condi-tioning involves hippocampal alpha5 GABA(A) receptors. Proc Natl Acad Sci US A 2002;99:8980–5.
42] Löw K, Crestani F, Keist R, Benke D, Brünig I, Benson JA, et al. Molecu-lar and neuronal substrate for the selective attenuation of anxiety. Science2000;290:131–4.
43] McKernan RM, Rosahl TW, Reynolds DS, Sur C, Wafford KA, Atack JR, et al.Sedative but not anxiolytic properties of benzodiazepines are mediated by theGABA(A) receptor alpha1 subtype. Nat Neurosci 2000;3:587–92.
44] Urakawa S, Takamoto K, Hori E, Sakai N, Ono T, Nishijo H. Rearing in enrichedenvironment increases parvalbumin positive small neurons in the amygdalaand decreases anxiety like behavior of male rats. BMC Neurosci 2013;14:1–11.
45] Rewal M, Wen Y, Wilson A, Simpkins JW, Jung ME. Role of parvalbumin inestrogen protection from ethanol withdrawal syndrome. Alcohol Clin Exp Res2005;29:1837–44.
46] Crestani F, Lorez M, Baer K, Essrich C, Benke D, Laurent JP, et al. DecreasedGABA-A receptor clustering results in enhanced anxiety and a bias for threatcues. Nat Neurosci 1999;2:833–9.
47] Treit D, Menard J. Dissociations among the anxiolytic effects of septal, hip-pocampal and amygdaloid lesions. Behav Neurosci 1997;111:653–8.
48] Salomé N, Salchner P, Viltart O, Sequeira H, Wiggger A, Landgraf R, et al.Neurobiological correlates of high (HAB) versus low anxiety-related behav-ior (LAB): differential fos expression in HAB and LAB rats. Biol Psychiatry2004;55:715–23.
49] Wislowska-Stanek A, Zienowicz M, Lehner M, Taracha E, Bidzinski A, MaciejakP, et al. Buspirone attenuates conditioned fear-induced c-Fos expression in therat hippocampus. Neurosci Lett 2005;389:115–20.
50] Gulyás AI, Tóth K, Dános P, Freund TF. Subpopulations of GABAergic neuronscontaining parvalbumin, calbindin, D28k, and cholecystokinin in the rat hip-pocampus. J Comp Neurol 1991;312:371–8.
51] Kalisch R, Salomé N, Platzer S, Wigger A, Czisch M, Sommer W, et al. High trait
52] Singewald N, Salchner P, Sharp T. Induction of c-Fos expression in specificareas of the fear circuitry in rat forebrain by anxiogenic drugs. Biol Psychiatry2003;53:275–83.