Page 1
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
57
1Efecto inductor del colorante amarillo azo sobre la expresión de genes y actividad de
oxidasas de Pleurotus ostreatus crecido en fermentación sumergida.
Induction effect of azo yellow dye on gene expression and activity of oxidases of
Pleurotus ostreatus grown in submerged fermentation.
1José Camacho Valenzuela, 2Soley Berenice Nava Galicia, 3Rubén Díaz Godínez, 1Saúl Tlecuitl
Beristain, 2Verónica Garrido Bazán, 2Martha Dolores Bibbins Martínez.
1Universidad Politécnica de Tlaxcala, Tepeyanco, Tlaxcala, México. Tel: +(52)012464651300
2Centro de Investigación en Biotecnología Aplicada IPN, Tepetitla, Tlaxcala, México. Tel:
+(52)2484870765
3Centro de Investigación en Ciencias Biológicas, Universidad Autónoma de Tlaxcala, México. Tel:
+(52)2484815482
RESUMEN. El hongo Pleurotus ostreatus es un basidiomiceto empleado para el desarrollo
de métodos de biorremediación de compuestos xenobióticos y recalcitrantes, debido a su
sistema enzimático de carácter no especifico, capaz de catalizar la oxidación y/ó hidrólisis
de una gran diversidad de compuestos xenobióticos, entre ellos, colorantes. El objetivo del
proyecto fue estudiar los patrones de expresión y producción de las enzimas ligninolíticas
que posee este hongo (lacasas, manganeso peroxidasas, versátil peroxidasas y dye
peroxidasas) en presencia del colorante amarillo azo (AYG) adicionado al inicio de la fase
exponencial de crecimiento (168 h) a concentraciones de 50 ppm y 500 ppm.
Con respecto a la expresión génica, la condición AYG a 50 ppm y la condición AYG a 500
ppm generaron un perfil de expresión diferencial respecto al tiempo sobre los genes poxa1b
y 1podyp. Sin embargo, los niveles de ARNm de las condiciones AYG a 50 ppm y AYG a
500 ppm disminuyeron en comparación con los de la fermentación basal. De igual forma,
no se detectó expresión para los genes mnp y vp. Así mismo, con respecto a la actividad
enzimática, la fermentación con el colorante AYG a 50 ppm mostró una actividad similar a
la condición basal. No obstante, el punto máximo de actividad en la condición AYG a 500
ppm muestra diferencias significativas al punto máximo de actividad en la fermentación
basal (4.57 veces mayor para lacasas, 4.24 veces mayor para DyP, 6.7 veces mayor para
Recibido: Abril, 2015.
Aprobado: Junio, 2015
Page 2
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
58
manganeso peroxidasas y 2.02 veces mayor para versátil peroxidasas), situación que
sugiere un notorio efecto de inducción por parte del colorante amarillo azo sobre las
actividades en mención.
La disminución de los niveles de ARNm de los genes poxa1b y 1podyp de la condición
AYG a 500 ppm con respecto a la fermentación basal sugiere que el colorante podría estar
induciendo la expresión de otros genes de isoformas que pudiesen estar involucradas en las
elevadas actividades enzimáticas observadas para lacasas y DyP debido a un mayor
requerimiento por parte del hongo de su participación que de aquellas enzimas codificadas
por los genes poxa1b y 1podyp para oxidar el colorante.
ABSTRACT. Pleurotus ostreatus is a basidiomycete fungi used for the development of
bioremediation methods of xenobiotics and recalcitrant compounds, due to its non-specific
enzyme system that is able of catalyzing the oxidation and/or hydrolysis in a wide variety
of compounds, including dyes.
The project's objective was to study the expression and production patterns of ligninolytic
enzymes of Pleurotus ostreatus (laccases, manganese peroxidases, versatile peroxidases
and dye peroxidases) in the presence of azo yellow dye (AYG) added at the beginning of
the exponential growth phase of this microorganism (168h) at concentrations of 50 ppm
and 500 ppm.
The condition of AYG at 50 ppm and AYG at 500 ppm generated a differential profile
expression with respect to time over poxa1b and 1podyp genes. However, the RNAm levels
of this conditions decreased in comparison with the non-added dye fermentation. For the
case of vp and mnp genes, expression was not detected. Likewise, the condition of AYG at
50 ppm showed a similar activity to the non-dye fermentation. Otherwise, the highest
activity in condition of AYG at 500 ppm showed significant differences in comparison of
the highest activity in non-dye fermentation (4.57 times bigger for laccases, 4.24 times
bigger for DyP, 6.7 times bigger for MnP and 2.02 times bigger for VP). This suggests a
strong effect of induction by the azo yellow dye over the enzymatic activities mentioned.
The decrease of transcriptional levels in poxa1b and 1podyp genes in the fermentation with
500 ppm of AYG with respect to non-dye fermentation suggest that the azo yellow dye
might induced the expression of others isoforms genes that could be involved in the high
enzymatic activities observed for laccases and DyP due to a bigger requirement by the
fungus of their participation than the ones that are encoded by poxa1b and 1podyp genes for
the oxidation of the dye under study.
Page 3
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
59
Palabras clave: Biorremediación, Pleurotus ostreatus, oxidasas, expresión diferencial
Key words: Bioremediation, Pleurotus ostreatus, oxidases, differential expression
INTRODUCCIÓN
La contaminación de ríos en Tlaxcala por una cierta gama de colorantes y compuestos
fenólicos provenientes de los efluentes de las industrias textiles, papeleras y químicas que
no reciben algún tipo de tratamiento previo, yace como una problemática que aqueja
algunas regiones del estado debido a la naturaleza xenobiótica de los contaminantes en
cuestión.
Actualmente existen algunas técnicas de carácter físico, químico y biológico que pueden
eliminar este tipo de sustancias, sin embargo, su utilización presenta algunas desventajas
importantes, siendo los altos costos de operación, el uso de grandes cantidades de químicos
y la formación de productos secundarios tóxicos los más representativos. Debido a ello se
ha vuelto preciso el desarrollo de nuevos métodos alternativos que contribuyan en mejor
manera a la solución del problema en mención.
Se ha reportado la capacidad de algunos hongos de pudrición blanca de degradar
biológicamente compuestos recalcitrantes de naturaleza diversa. De igual forma, estos
organismos han sido ampliamente estudiados debido a su habilidad para degradar lignina y
compuestos fenólicos, lo anterior como resultado de la producción de hidrolasas y oxidasas
para llevar a cabo la ruptura de los polímeros estructurales de la lignocelulosa hasta
compuestos simples y fácilmente asimilables para el crecimiento (Evans y col., 1994).
Teniendo en cuenta que la lignina es un polímero no específico, el sistema involucrado en
su descomposición también funciona de manera no específica. Ésto permite que el sistema
ligninolítico de los hongos de pudrición blanca reduzca una gran variedad de contaminantes
aromáticos (Bumpus y col., 1987). De igual forma, este grupo de hongos ha mostrado ser
efectivo para remover y detoxificar efluentes industriales (Kumar y col., 2007). En vista de
lo anterior, estos hongos se emplean en la actualidad para desarrollar técnicas de
biorremediación de compuestos xenobióticos. Las principales enzimas ligninoliticas que
intervienen en estos procesos son hemoproteínas extracelulares denominadas enzimas
peroxidasas, conocidas como lignino peroxidasa (LiP, EC1.11.1.14), manganeso peroxidasa
(MnP, EC 1.11.1.13) y versátil peroxidasa (VP, EC 1.11.1.16), peroxidasa decolorante de
tintes (DyP, EC 1.11.1.19), además de la oxidasa lacasa (EC 1.10.3.2).
Page 4
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
60
Es por esto que el desarrollo de metodologías eficientes y económicamente factibles a partir
del uso de organismos como los hongos basidiomicetos, representa una oportunidad
potencial de lograr la biorremediación sobre ríos y otros sitios afectados por la
contaminación con compuestos recalcitrantes generados por diversas industrias.
Por lo anterior el objetivo del presente trabajo fue determinar el patrón de expresión de las
enzimas lacasas, manganeso peroxidasas, dye peroxidasas y versátil peroxidasas producidas
por Pleurotus ostreatus en fermentación sumergida en presencia del colorante amarillo azo
inoculado al inicio de la fase exponencial de crecimiento del hongo (168h).
METODOLOGÍA
Fermentaciones
Se establecieron tres fermentaciones sumergidas, siendo dos de ellas en presencia de
colorante amarillo azo a una concentración de 500 y 50 ppm así como de una basal como
referencia, todo esto en matraces de 125 mL que contenían 50 mL de medio, mismos que se
mantuvieron a una temperatura de 25 °C a 120 rpm durante 23 días. La inoculación del
micelio se llevó a cabo con 3 pellets de 4 mm de diámetro para cada matraz, tomado de la
periferia de una colonia de Pleurotus ostreatus (cepa ATCC 32783). El colorante se agregó
al inicio de la fase exponencial (168h). El pH del medio se ajustó a 6.5. La composición del
medio de cultivo para todos los casos fue el reportado por Téllez-Téllez en el 2008.
Se realizaron muestreos para las 3 fermentaciones de los siguientes puntos: 168h, 175h,
199h, 223h, 247h, 384h y 499h. Por cada punto de muestreo, se filtró la biomasa de cada
matraz con el equipo de filtración al vacío WELCH INSUL CLASS B (MODEL NO.
2546B-01) para ser secada después en horno a 60 ºC durante 24 h. La biomasa se reportó en
gramos de biomasa seca por litro de medio (gL-1). De los mismos puntos de muestreo,
también se recuperó micelio y se almacenó a -80 °C para su posterior extracción de ARN.
Así mismo, el sobrenadante de cada matraz se recuperó en tubos eppendorf y se almacenó a
su vez a -20 °C para medir posteriormente la actividad enzimática.
Parámetros cinéticos de crecimiento
Page 5
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
61
Para determinar los parámetros de crecimiento se consideró la ecuación logística de
crecimiento microbiano, utilizando además la herramienta “Solver” de la hoja electrónica
de Excel (Microsoft) para la minimización del error cuadrático (Díaz-Godínez y col, 2001).
Medición de consumo de fuente de carbono
Para cuantificar el consumo de la fuente de carbono (glucosa), se adaptó el método del
DNS de Miller (1959). La mezcla de reacción contenía 20 µL de extracto crudo enzimático,
2 mL de DNS y 980 µL de H2O desionizada. Las muestras a leer se incubaron en baño
maría en tubos de ensayo a temperatura de ebullición durante 5 minutos.
Una vez transcurrido el tiempo de ebullición, la reacción se detuvo sumergiendo los tubos
en agua con hielo. Por otro lado el blanco contenía los mismos volúmenes, sin embargo,
éste no fue incubado como las muestras medidas.
Se utilizó un blanco para cada punto evaluado. La absorbancia fue leída a 575 nm. Se
realizó una curva patrón de glucosa.
Extracción de ARN total
El aislamiento de ARN total de los micelios obtenidos de la fermentación a 50 ppm se llevó
a cabo empleando el método de fenol-cloroformo con un búffer NTES. De igual forma, el
aislamiento de ARN total de los micelios obtenidos de la fermentación basal y a 500 ppm
se llevó a cabo con el método del TRIZOL (Invitrogen).
Los puntos seleccionados para la realización de los análisis moleculares fueron los
siguientes: 168h, 175h, 199h, 223h, 27h, 384h y 499h.
Electroforesis en geles con formaldehído
Una vez hechas las extracciones, se corrieron geles de agarosa al 1.5% en electroforesis
para evaluar la integridad del ARN obtenido. El gel se preparó con 0.6 g de agarosa, 4 mL
de MOPS 10X, 6 mL de formaldehído, 30 mL de agua DEPC y 1.5 µL de bromuro de
Page 6
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
62
etidio. Después, se cuantificó el ARN extraído mediante espectrofotometría a una longitud
de onda de 260 nm. Se ajustó la concentración a 300 ngµL-1.
Se corrió un gel de agarosa con formaldehído para visualizar la integridad de los productos
ajustados. Posteriormente se les dio un tratamiento con ADNasa y se dejaron en incubación
a -20ºC.
RT-PCR
La síntesis de ADNc se realizó utilizando primers dT y la transcriptasa reversa Moloney
murine Leukemia virus (M-MuLV-RT; Fermentas).
PCR
Para las 3 fermentaciones, se llevaron a cabo las amplificaciones de los genes bajo estudio
(poxa1b, 1podyp, mnp y vp) con la técnica de PCR. Las condiciones de ciclado se muestran
en la siguiente tabla (Tabla 1):
Tabla 1. Programa de ciclado
Etapa Temperatura Tiempo Ciclos
1° Desnaturalización inicial 94°C 3 minutos 1
2° Desnaturalización
Alineamiento
Extensión
94°C
56°C
72°C
50 segundos
50 segundos
50 segundos
40
3° Extensión final 72°C 10 minutos 1
4° 4°C ∞ 1
Electroforesis en geles con búffer TAE 1X
Page 7
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
63
Se corrieron geles de agarosa al 2% en electroforesis para visualizar la amplificación de los
genes de cada uno de los puntos seleccionados. El gel se preparó con 0.7 g de agarosa, 35
mL de búffer TAE 1X y 1.5 µL de bromuro de etidio.
Actividad enzimática de lacasas
La actividad de lacasa (EC: 1.10.3.2) de los tiempos monitoreados se determinó utilizando
como sustrato 2,6-dimetoxifenol (DMP) (Díaz-Godínez, 2009). La mezcla de reacción se
realizó con 950 μL de DMP 2 mM en búffer de fosfatos 0.1 M a pH de 6.5, y 50 μL de
EEE. La actividad se determinó por incremento en la absorbancia a 468 nm (468= 10000
M-1 cm-1) de la mezcla de reacción en una celda Peltier a 40 °C durante un minuto (UV/Vis
Jenway 6405). Se empleó como blanco toda la mezcla de reacción sustituyendo los 50 μL
de EEE por 50 μL de agua desionizada. Una unidad de actividad de lacasa (UI) se
consideró como la cantidad de enzima que se requiere para oxidar un µmol de sustrato por
minuto en un litro de reacción.
Actividad enzimática de dye peroxidasa
La actividad de DyP (EC: 1.11.1.19) se midió utilizando ABTS como sustrato en presencia
de H2O2. (Salvachúa, 2013). La mezcla de reacción se realizó con 980 µL de ABTS 2.5
mM en búffer de tartratos 0.1 M a pH de 3 con H2O2 0.01 mM y 20 µL de EEE. La
actividad se determinó por aumento en la absorbancia a 436 nm (=29300 M-1 cm-1) de la
mezcla de reacción en una celda Peltier a 40°C durante un minuto (UV/Vis Jenway 6405).
Se empleó como blanco toda la mezcla de reacción antes de incubar. Una unidad de
actividad de DyP (UI) se consideró como la cantidad de enzima que se requiere para oxidar
un µmol de sustrato por minuto en un litro de reacción.
Actividad enzimática de manganeso peroxidasa
La actividad de MnP (EC: 1.11.1.13) se midió utilizando MnSO4 como sustrato en
presencia de H2O2 (Giardina, 2000). La mezcla de reacción se realizó con 950 µL de
MnSO4 .5 mM en búffer de malonatos 50 mM a pH de 4.5, 30 µL de H2O2 .05 mM y 20 µL
de EEE. La actividad se determinó por aumento en la absorbancia a 270 nm (270=11590
M-1cm-1) de la mezcla de reacción en una celda Peltier a 25°C durante un minuto (UV/Vis
Page 8
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
64
Jenway 6405). Se empleó como blanco toda la mezcla de reacción excepto los 30 µL de
H2O2. Una unidad de actividad de VP (UI) se consideró como la cantidad de enzima que se
requiere para oxidar un µmol de sustrato por minuto en un litro de reacción.
Actividad enzimática de versátil peroxidasa
La actividad de VP (EC: 1.11.1.16) se midió utilizando MnSO4 como sustrato en presencia
de H2O2 (Boada, 2005). La mezcla de reacción se realizó con 950 µL de MnSO4 .1 mM en
búffer de tartratos .1 M a pH de 5, 30 µL de H2O2 .1 mM y 20 µL de EEE. La actividad se
determinó por aumento en la absorbancia a 238 nm (=6500 M-1 cm-1) de la mezcla de
reacción en una celda Peltier a 40°C durante un minuto (UV/Vis Jenway 6405). Se empleó
como blanco toda la mezcla de reacción excepto los 30 µL de H2O2.
Una unidad de actividad de VP (UI) se consideró como la cantidad de enzima que se
requiere para oxidar un µmol de sustrato por minuto en un litro de reacción.
RESULTADOS Y DISCUSIÓN
Cinética de crecimiento y consumo de fuente de carbono
La fermentación que presentó la biomasa máxima (Xmáx) más baja fue aquella con presencia
de AYG a 50 ppm con 6.69 gL-1, seguida de la fermentación basal (8.4 gL-1) y de la
fermentación con presencia de AYG a 500 ppm (12.37 gL-1).
Las velocidades específicas de crecimiento (µ) siguieron el mismo orden, siendo 0.055 h-1
para la basal y 0.041 h-1 y 0.089 h-1 para las fermentaciones con 50 y 500 ppm del colorante
respectivamente (Tabla 2).
Tabla 2. Parámetros cinéticos de crecimiento (Xmáx y µ) de las fermentaciones de
Pleurotus ostreatus
Parámetro Basal AYG 50 ppm AYG 500 ppm
Page 9
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
65
Xmax (gL-1) 8.4 6.69 12.37
µ (h-1) 0.055 0.041 0.089
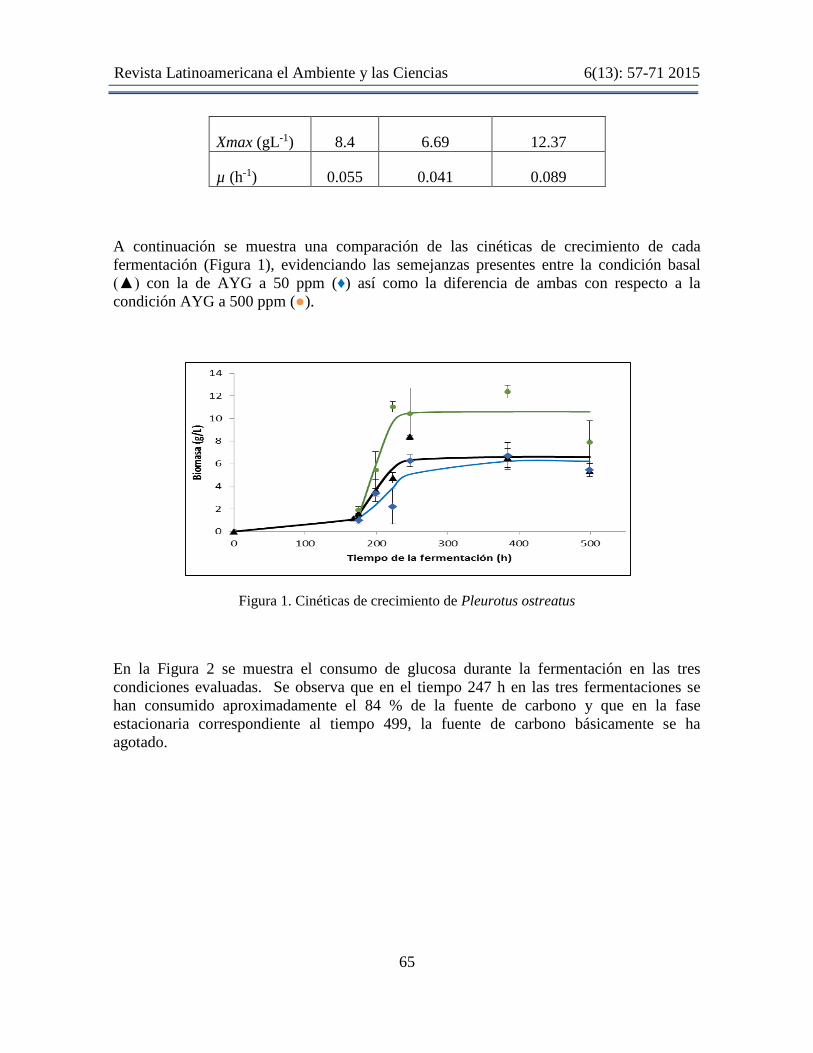
A continuación se muestra una comparación de las cinéticas de crecimiento de cada
fermentación (Figura 1), evidenciando las semejanzas presentes entre la condición basal
(▲) con la de AYG a 50 ppm (♦) así como la diferencia de ambas con respecto a la
condición AYG a 500 ppm (●).
Figura 1. Cinéticas de crecimiento de Pleurotus ostreatus
En la Figura 2 se muestra el consumo de glucosa durante la fermentación en las tres
condiciones evaluadas. Se observa que en el tiempo 247 h en las tres fermentaciones se
han consumido aproximadamente el 84 % de la fuente de carbono y que en la fase
estacionaria correspondiente al tiempo 499, la fuente de carbono básicamente se ha
agotado.
Page 10
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
66
Figura 2. Consumo de fuente de carbono de las 3 fermentaciones evaluadas
Determinación de actividad enzimática
La actividad de DyP mostró un notorio incremento en presencia de AYG a 500 ppm siendo
el pico máximo de actividad de 3594.709 UI/L a las 499h en comparación con la condición
AYG a 50 ppm donde el pico máximo de actividad fue de 538.39 UI/L a las 499h y con la
fermentación basal el pico máximo de actividad fue de 846.84 UI/L a las 247h (Figura 3).
Figura 3. Comparación de las actividades enzimáticas de DyP en las 3 fermentaciones
0.00
2.00
4.00
6.00
8.00
10.00
12.00
0 100 200 300 400 500
Co
nce
ntr
ació
n d
e g
luco
sa (
g/L
)
Tiempo de la fermentación (h)
F. basal
Page 11
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
67
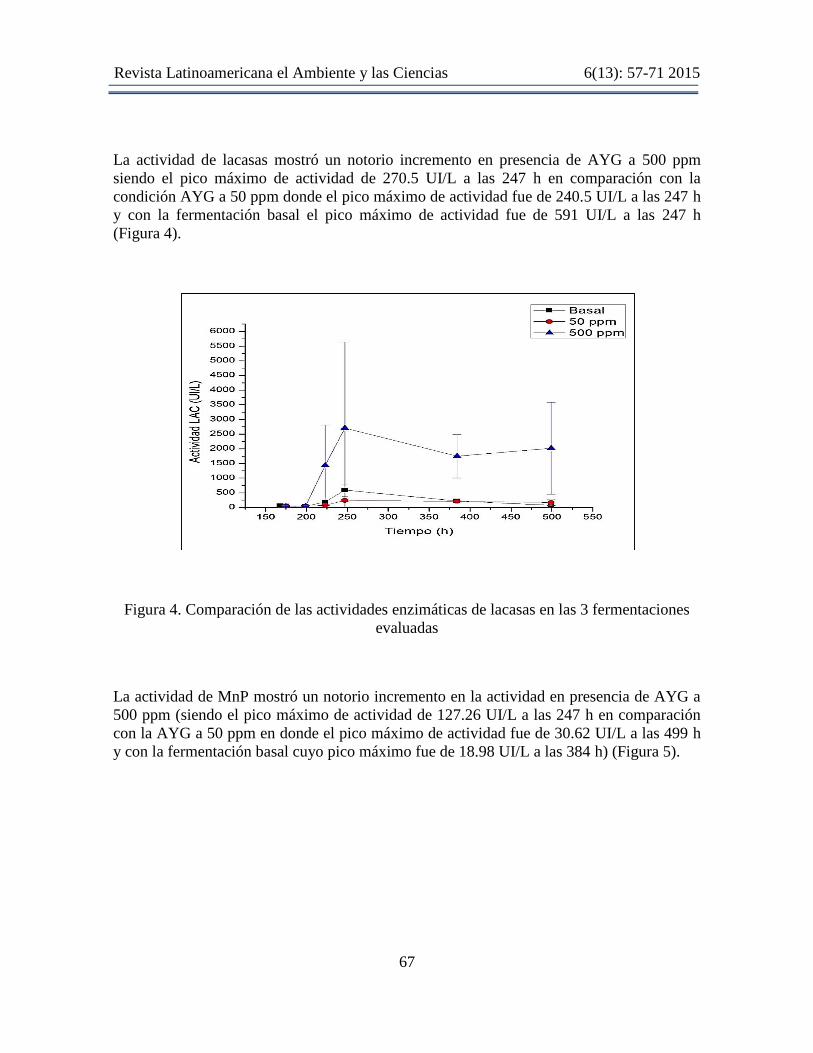
La actividad de lacasas mostró un notorio incremento en presencia de AYG a 500 ppm
siendo el pico máximo de actividad de 270.5 UI/L a las 247 h en comparación con la
condición AYG a 50 ppm donde el pico máximo de actividad fue de 240.5 UI/L a las 247 h
y con la fermentación basal el pico máximo de actividad fue de 591 UI/L a las 247 h
(Figura 4).
Figura 4. Comparación de las actividades enzimáticas de lacasas en las 3 fermentaciones
evaluadas
La actividad de MnP mostró un notorio incremento en la actividad en presencia de AYG a
500 ppm (siendo el pico máximo de actividad de 127.26 UI/L a las 247 h en comparación
con la AYG a 50 ppm en donde el pico máximo de actividad fue de 30.62 UI/L a las 499 h
y con la fermentación basal cuyo pico máximo fue de 18.98 UI/L a las 384 h) (Figura 5).
Page 12
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
68
Figura 5. Comparación de las actividades enzimáticas de MnP en las 3 fermentaciones evaluadas
La actividad de VP mostró un notorio incremento en la actividad en presencia de AYG a
500 ppm, siendo el pico máximo de actividad de 865.38 UI/L a las 499 h en comparación
con la condición AYG a 50 ppm en donde el pico máximo de actividad fue de 134.61 UI/L
a las 384 h y con la fermentación basal en la cual el pico máximo de actividad fue de
426.92 UI/L (Figura 6).
Figura 6. Comparación de las actividades enzimáticas de VP en las 3 fermentaciones
Page 13
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
69
Perfil de expresión génica
En la siguiente figura 7 se muestra el perfil de expresión de los genes poxa1b, 1podyp, mnp
y vp. El gen gpd fue utilizado como control interno. En las tres fermentaciones evaluadas se
puede apreciar que no se detectó expresión para vp y mnp. En primera instancia se esperaría
observar una mayor expresión en la fermentación con AYG a 500 ppm en comparación con
la condición basal dados los resultados obtenidos en la medición de la actividad enzimática
de DyP y lacasas, sin embargo, se puede apreciar una expresión mayor en la fermentación
basal a diferencia de las condiciones AYG a 500 y 50 ppm para los genes de lacasa
(poxa1b) y dye peroxidasa (1podyp). Lo anterior sugiere que el colorante podría estar
induciendo la expresión de otros genes de isoformas que pudiesen estar involucradas en las
elevadas actividades enzimáticas visualizadas debido a un mayor requerimiento por parte
del hongo de su participación que de aquellas codificadas por los genes poxa1b y 1podyp
para oxidar el colorante.
Figura 7. Perfil de expresión transcripcional de los genes evaluados en las 3 fermentaciones
descritas
CONCLUSIONES
El colorante AYG a una concentración de 500 ppm generó un claro efecto de inducción
sobre las actividades enzimáticas evaluadas en comparación a la condición basal en sus
Page 14
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
70
respectivos puntos de actividad máxima (4.24 veces mayor en DyP, 4.57 veces mayor en
lacasas, 6.7 veces mayor en MnP y 2.02 veces mayor en VP) mientras que la fermentación
con AYG a 50 ppm presentó una actividad similar a la condición basal.
Así mismo, la condición AYG a 50 ppm y la condición AYG a 500 ppm generaron un
perfil de expresión diferencial de los genes poxa1b y 1podyp respecto al tiempo. Sin
embargo, los niveles de ARNm de las fermentaciones AYG a 50 ppm y AYG a 500 ppm
disminuyeron en comparación con aquellos correspondientes a la condición basal.
AGRADECIMIENTOS
Al CONACYT, beca No. 21700, proyecto CB.134348.
BIBLIOGRAFÍA
Bumpus, J. A.; Aust, S. D. (1987). Biodegradation of environmental pollutants by the white
rot fungus Phanerochaete chrysosporium: involvement of the lignin degrading system. BioEssays, 6(4), 166-170.
Díaz-Godínez, G.; Soriano-Santos, J.; Augur, C.; Viniegra-González, G. (2001).
Exopectinases produced by Aspergillus niger in solid-state and submerged fermentation: a
comparative study. Journal of Industrial Microbiology and Biotechnology, 26(5), 271-275.
Díaz-Godínez, R. 2009. Efecto del pH inicial de desarrollo de pleurotus ostreatus en
fermentación sumergida sobre su actividad de lacasas. Tesis profesional. UAM. Tlaxcala.
México.
Evans, C. S.; Dutton, M. V.; Guillén, F.; Veness, R. G. (1994). Enzymes and small
molecular mass agents involved with lignocellulose degradation. FEMS Microbiology
Reviews, 13(2-3), 235-239.
Page 15
Revista Latinoamericana el Ambiente y las Ciencias 6(13): 57-71 2015
71
Giardina P.; Palmieri G.; Fontanella B.; Rivieccio V.; Sannia G. (2000). Manganese
Peroxidase Isoenzymes Produced by Pleurotus ostreatus Grown on Wood Sawdust.
Archives of biochemistry and biophysics, 376(1), 171-179.
Kumar, K.; Devi, S. S.; Krishnamurthi, K.; Dutta, D.; Chakrabarti, T. (2007).
Decolorisation and detoxification of Direct Blue-15 by a bacterial consortium. Bioresource
technology, 98(16), 3168-3171.
Miller, G. L. (1959). Use of dinitrosalicylic acid reagent for determination of reducing
sugar. Analytical chemistry, 31(3), 426-428.
Perez-Boada, M.; Ruiz-Duenas, F. J.; Pogni, R.; Basosi, R.; Choinowski, T.; Martínez, M.
J.; Martínez, A. T. (2005). Versatile peroxidase oxidation of high redox potential aromatic
compounds: site-directed mutagenesis, spectroscopic and crystallographic investigation of
three long-range electron transfer pathways. Journal of molecular biology, 354(2), 385-
402.
Salvachúa, D.; Prieto, A.; Martínez, Á. T.; Martínez, M. J. (2013). Characterization of a
novel dye-decolorizing peroxidase (DyP)-type enzyme from Irpex lacteus and its
application in enzymatic hydrolysis of wheat straw. Applied and environmental
microbiology, 79(14), 4316-4324.
Téllez-Téllez M.; Fernández, F. J.; Montiel-González A. M.; Sánchez, C.; Díaz-Godínez,
G.; 2008. Growth and laccase production by Pleurotus ostreatus in submerged and solid-
state fermentation. Applied Microbiology and Biotechnology, 81:675-679.