1
Phylogenetic Diversity among Low Virulence Newcastle Disease 1
Viruses from Waterfowl and Shorebirds and Comparison of 2
Genotype Distributions to Poultry-Origin Isolates 3
4
L. Mia Kim,1 Daniel J. King,1 Phillip E. Curry,1a David L. Suarez,1 David E. Swayne,1 5
David E. Stallknecht,2 Richard D. Slemons,3 Janice C. Pedersen,4 Dennis A. Senne,4 6
Kevin Winker,5 Claudio L. Afonso1* 7
USDA ARS Southeast Poultry Research Laboratory, 934 College Station Rd, Athens, GA 30605,1 8
Department of Population Health, The University of Georgia, Athens, GA 30602,2 Department of 9
Veterinary Preventive Medicine, The Ohio State University, Columbus, Ohio 43210,3 USDA 10
APHIS VS, National Veterinary Services Laboratories, Diagnostic Virology Laboratory-Avian 11
Section, Ames, IA 50010,4 University of Alaska Museum, 907 Yukon Drive 12
Fairbanks, Alaska 99775 5 13
14
Correspondent: 15
USDA ARS 16
Southeast Poultry Research Laboratory 17
934 College Station Rd 18
Athens, GA 30605 19
706-546-3642 20
706-546-3161 fax 21
[email protected] 22
a SC Department of Health and Environmental Control, Bureau of Laboratories Molecular Microbiology Division, 8231 Parklane Rd., Columbia, SC 29223
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.00843-07 JVI Accepts, published online ahead of print on 12 September 2007
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
2
Running title: Phylogenetic diversity of low virulence NDV 1
Abstract: 250 words 2
Text: 5600 words 3
4
Abstract 5
Low virulence Newcastle disease viruses (loNDV) are frequently recovered from wild 6
bird species, but little is known about their distribution, genetic diversity, or potential to cause 7
disease in poultry. NDV isolates recovered from cloacal samples of apparently healthy waterfowl 8
and shorebirds (WS) in the U.S. during 1986 to 2005 were examined for genomic diversity and 9
their potential for virulence (n = 249). In addition 19 loNDV isolates from U.S. live bird markets 10
(LBMs) were analyzed and found to be genetically distinct from NDV used in live vaccines, but 11
related to WS-origin NDV. Phylogenetic analysis of the fusion protein identified nine novel 12
genotypes among the class I NDV and new genomic subgroups were identified among genotypes 13
I and II of the class II viruses. The WS-origin viruses exhibited broad genetic and antigenic 14
diversity and some WS genotypes displayed a closer phylogenetic relationship to LBM-origin 15
NDV. All NDV were predicted to be lentogenic based upon sequencing of the fusion cleavage 16
site, intracerebral pathogenicity index, or mean death time in embryo assays. The USDA real-17
time RT-PCR (RRT-PCR) assay that targets the matrix gene identified nearly all of the class II 18
NDV viruses tested, but failed to detect class I viruses from both LBM and WS. The close 19
phylogenetic proximity of some WS and LBM loNDV suggests that viral transmission may 20
occur among wild birds and poultry; however, these events may occur unnoticed due to the broad 21
genetic diversity of loNDV, the lentogenic presentation in birds, and the limitations of current 22
rapid diagnostic tools. 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
3
Introduction 1
Viruses of the avian paramyxovirus serotype-1 (APMV-1), synonymous with Newcastle 2
disease virus (NDV) belong to the genus Avulavirus in the family Paramyxoviridae (34) and 3
encompass a diverse group of single stranded, negative sense, non-segmented RNA viruses of 4
approximately 15.2 Kb that have a broad host range in bird species. The virulent forms of the 5
virus, which exhibit an intracerebral pathogenicity index ≥0.7, are the cause of Newcastle disease 6
(ND), a notifiable avian disease that must be reported to the World Organization for Animal 7
Health (OIE). Classically these viruses have been grouped by virulence phenotype, with 8
lentogenic, mesogenic, and velogenic strains reflecting increasing levels of virulence. Those 9
viruses categorized either as mesogens or velogens are currently defined as virulent Newcastle 10
disease viruses (vNDV) and lentogens are viruses of low virulence (loNDV). Newcastle disease 11
viruses can cause clinical signs ranging from subclinical infections to 100% morbidity and/or 12
mortality depending on the virulence of the virus and the susceptibility of the host. Lentogenic 13
viruses typically cause subclinical infections or mild respiratory disease. Mesogens are of 14
intermediate virulence, usually resulting in moderate respiratory disease with occasional nervous 15
signs. Velogens, the most virulent viruses, are those that may cause extensive hemorrhagic 16
lesions, particularly in the gastrointestinal tract (viscerotropic) and/or a predominance of nervous 17
signs (neurotropic) (4). Virulent NDV are considered exotic to United States (U.S.) poultry; 18
however, the lentogenic strains are common among domestic poultry (27,32,36,45) and wild bird 19
populations (25,27,43,50). The occurrence of vNDV infections in birds is a notifiable event and 20
when these infections occur in poultry trade restrictions are imposed. Virulent NDV infections 21
present a significant threat to the U.S. poultry industry as evidenced by the 2002-03 California 22
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
4
outbreak (41) which resulted in the destruction of 3.3 million birds and cost nearly $200 million 1
dollars to eradicate (53). 2
Newcastle disease viruses have historically been grouped into either genotypes (9) or 3
genetic lineages (2) under one serotype (APMV-1). Recent analysis of the genome size and 4
sequence analysis of the F and L genes has revealed 2 distinct clades within the APMV serotype-5
1 viruses: class I and II (14). While there are few published genomic sequences of class I 6
isolates, which have been primarily recovered from waterfowl of the order Anatidae and samples 7
from U.S. live bird markets (LBMs), the class II viruses comprise the vast majority of sequenced 8
NDV and include isolates recovered from poultry (gallinaceous birds), pet and wild bird viruses 9
(2,45). The class II NDV are further categorized into genotypes I to IX with the genomic 10
sequences of commonly used vaccine strains resembling virulent NDV isolated during the 1940s 11
(class II, genotype II). 12
The natural ecology of NDV is not fully understood and research has mainly focused on 13
tracing either the origin of specific viruses or the spread of virulent viruses during ND outbreaks 14
in poultry (5-7,12,15,21,41,44). Of the few studies characterizing NDV infections in wild bird 15
populations, some suggest that waterfowl provide a natural reservoir for NDV and 16
epidemiological links have been hypothesized between outbreak isolates recovered from poultry 17
and those isolates found in wild bird populations (18,25-27,42,50). For example, phylogenetic 18
analyses have identified virulent NDV isolates recovered from pigeons and migrating cormorants 19
as the likely source of some NDV outbreaks in poultry (1,10,22,31,37,51). 20
Although recent data have implicated U.S. LBMs as the source of respiratory pathogens 21
such as avian influenza virus (AIV) and NDV that cause disease in domestic poultry 22
(11,32,40,45,46); there is a paucity of data on the relationship between NDV circulating in wild 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
5
waterfowl and shorebirds and NDV causing infections in poultry from LBMs. Earlier reports on 1
the genomic analysis of NDV recovered in U.S. LBMs showed that these loNDV isolates were 2
related to class II NDV strains which have been used for live vaccines in commercial poultry 3
(32), but more recently three U.S. LBM isolates recovered during 2001 and 2002 are 4
representative of class I viruses (45). Additionally, the recently reported LBM-origin isolates 5
from Hong Kong were also found to be class I viruses (28). 6
Understanding the epidemiology of loNDV infections in waterfowl and LBM poultry has 7
been hampered by the fact that these viruses typically produce no clinical signs and that class I 8
isolates are often not detected by rapid screening methods such as reverse transcription PCR 9
(RT-PCR). Sensitive rapid diagnostic assays exist to detect class II NDV such as the USDA-10
validated real time RT-PCR which targets the matrix gene (M-gene assay); however, due to the 11
heterogeneous genetic nature of this virus, class I viruses tested often fail to be detected by the 12
M-gene assay (28). Previous evaluation of the nucleotide sequence alignment of the M-gene 13
assay probe site of class I and II viruses revealed a high degree of mismatches between the two 14
clades, and this is likely the reason that the class I viruses escape detection by real time RT-PCR 15
(28,29). 16
Here, Newcastle disease viruses from U.S. LBMs (n = 19) that demonstrated NDV-17
specific hemagglutination-inhibition (HI) activity with polyclonal antiserum, but which tested 18
negative for NDV using the M-gene assay, were partially sequenced and found to be genetically 19
distinct and phylogenetically distant from vaccine and virulent viruses reported worldwide. In 20
addition, 249 NDV isolates recovered from cloacal samples collected during 1986 to 2005 from 21
apparently healthy waterfowl and shorebirds (WS) in the U.S. were analyzed and select isolates 22
were tested using the M-gene assay and a panel of monoclonal antibodies. We established the 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
6
phylogenetic relationship between the LBM- and WS-origin NDV isolates, determined the 1
geographic distribution and broad genetic diversity among the WS-origin viruses, and evaluated 2
the potential for virulence of U.S. WS-origin viruses. 3
4
Materials and Methods 5
Hemagglutination (HA) and hemagglutination-inhibition (HI) assays. The HA and 6
HI assays were completed by microtiter methods. The HA assay of allantoic fluids harvested 7
from inoculated embryonating eggs was used to identify NDV-positive embryos. Confirmation 8
of NDV-positive fluids and antigenic characterization of virus isolates was conducted by HI 9
using microtiter methods as previously described (30). Four HA units of viral test antigen were 10
used in completing the HI assay with monoclonal antibodies and polyclonal antiserum. 11
Isolates and sequence data. Newcastle disease viruses were obtained from the Southeast 12
Poultry Research Laboratory (SEPRL) repository or collected during a multi-institutional 13
cooperative WS virus monitoring project between University of Georgia Southeastern 14
Cooperative Wildlife Disease Study, the University of Alaska Museum, and The Ohio State 15
University Department of Veterinary Preventive Medicine and USDA-SEPRL (47). Viruses 16
from wild-captured WS were isolated from cloacal swab material by standard virus isolation 17
methods in embryonating chicken eggs (3,49). Two-hundred-forty-nine U.S. WS isolates (Table 18
1) were obtained from monitoring samples for shorebirds of the family Scolopacidae in DE (n = 19
17) and NJ (n = 8); and from duck and geese sampling in MD (n = 121), MN (n = 43), OH (n = 20
15), LA (n = 16), TX (n = 26), and AK (n = 3; includes one previously published isolate; 21
Northern Pintail/US(AK)/196/1998, Genbank accession EF027165). The 19 U.S. LBM 22
environmental and poultry isolates were obtained from surveillance samples submitted to the 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
7
USDA APHIS National Veterinary Services Laboratory from MA (n = 1), NJ (n = 10), NY (n = 1
7), and RI (n = 1). In addition, previously described samples from the Agriculture, Fisheries and 2
Conservation Department of Hong Kong SAR (n = 21) (28) and Shelduck/France/MC-110/1977 3
(GenBank accession AF003726) were included totaling 290 isolates (Supplemental Table S1). 4
For comparisons, all isolates were grouped into four major categories based on the source: Anas 5
platyrhynchos (mallard and Anas rubripes or black duck; n = 169), Other Anatidae (other ducks 6
and geese; n = 56), Scolopacidae (shorebirds; n = 25), and Live bird market (LBM; 7
environmental and domestic poultry isolates; n = 40). 8
RNA was extracted from allantoic fluids using Trizol LS (Invitrogen, Carlsbad, CA) 9
according to manufacturer instructions. Briefly, 750µl of Trizol LS reagent was added to 250µl 10
of allantoic fluid, vortexed, and incubated at room temperature for 7 minutes. The RNA was 11
separated into the aqueous phase with the addition of 200µl of chloroform, precipitated with 12
isopropanol, and then centrifuged to pellet the RNA. After one wash with 70% ethanol, RNA 13
was dried and resuspended in RNAse-free water. PCR amplification of the RNA was performed 14
using the Qiagen One Step reverse transcription PCR (RT-PCR) kit (Qiagen, Valencia, CA). 15
Amplified products were separated on a 1% agarose gel, the bands excised and eluted using the 16
QIAquick Gel extraction kit (Qiagen), and the samples quantified using a standard 17
spectrophotometer. 18
All sequencing reactions were performed with fluorescent dideoxynucleotide terminators 19
in an automated sequencer (ABI 3700 automated sequencer; Applied Biosystems Inc., Foster 20
City, CA). Nucleotide sequence editing and analyses were conducted with the LaserGene 21
sequence analysis software package (LaserGene, version 5.07; DNAStar, Inc., Madison, WI). 22
Using the full-length genome positions from the NDV LaSota vaccine strain complete genome 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
8
(accession # AF077761), the homologous regions sequenced were as follows: a 374 bp partial F 1
gene (positions 4554 to 4917; n = 206), a 254 bp partial fragment spanning the M and F genes 2
(positions 4826 to 5066; n = 40), and the complete coding region for the F gene (positions 4544 3
to 6205; n = 21). 4
Phylogenetic analysis. Maximum likelihood (ML) phylogenetic analysis with bootstrap 5
values for n = 100 replicates was performed using Phyml under the general time reversible 6
(GTR) model of nucleotide substitutions, ML estimates of base frequencies, estimated 7
transition/transversion ratio and proportions of invariable sites with 4 categories of substitution 8
rates (19). For DNAML trees (16) default conditions were used, and distance based analysis 9
conducted with the Neighbor Joining method was done using the Tajima-Nei nucleotide 10
substitution model in the BioEdit Sequence Alignment Editor (Hall, T.A. Abstr. Nucl. Acids. 11
Symp. Ser. 41:95-98, 1999). The 374 bp region of the F gene, which has commonly been used 12
for phylogenetic analysis of NDV (2), was sequenced to compare the U.S. LBM isolates (n = 19) 13
to 209 WS viruses. For reference, previously published GenBank sequences from known NDV 14
clades (class I n = 13; class II n = 47), in addition to 21 class I LBM isolates from Hong Kong 15
(28) were included. The 254 bp region was used to compare the 40 remaining WS isolates (for a 16
total of 249) to 106 LBM and WS viruses previously analyzed in the 374 bp analysis (data not 17
shown). For the purpose of discussion and to maintain consistency with previous naming 18
conventions, the term “genotype” was used here to describe isolates that reproducibly group 19
together on a distinct branch of a phylogenetic tree. The class I genotypes were indicated using 20
Arabic numerals for ease of differentiating them from class II genotypes. 21
Pathogenicity assessment. The pathogenic potential for selected class I isolates was 22
evaluated using standard assay methods to determine the intracerebral pathogenicity index (ICPI) 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
9
in one-day-old chicks (3) and a modified procedure was used to determine mean death time 1
(MDT) in embryonating eggs. The MDT was performed by inoculating the allantoic cavity of 9-2
11 day old embryonating specific pathogen free eggs with serial 10-fold dilutions of a virus 3
isolate. The eggs were incubated at 37°C and candled twice daily (early morning and late 4
afternoon) for seven days, and the time of embryo mortality was recorded. The MDT of a 5
minimum lethal dose was interpreted as the mean time in hours for the embryo death in the 6
highest dilution at which all eggs died (3). 7
Real time reverse-transcription PCR. Select LBM and WS NDV isolates (n = 50) were 8
tested using the USDA-validated M-gene assay (53). 9
Monoclonal antibodies (mAbs) and antiserum. Nine mAbs with different NDV 10
specificities were used for differentiating isolates by the HI assay as previously described 11
(30,33). The mAbs obtained from USDA, APHIS, National Veterinary Services Laboratories 12
(Ames, Iowa) included B79, 15C4, 10D11 (35), AVS (48), and 617/161 (13). Additional mAbs 13
prepared at SEPRL included P15D7, P11C9, P3A11, and P10B8. The reactivity of the mAbs was 14
as follows: the AVS mAb inhibits many lentogens including B1 and LaSota strains; 10D11 15
inhibits neurotropic velogens and mesogens like the Roakin strain; 15C4 inhibits most APMV-1 16
except pigeon paramyxovirus-1 (PPMV-1); B79 inhibits most APMV-1 including most PPMV-17
1; P15D7, P11C9, P3A11, and P10B8 identify additional antigenic diversity among APMV-1; 18
and the 617/161 mAb inhibits only PPMV-1 within the APMV-1 group. A positive result was 19
defined as antibody-inhibited HA, and a negative result was defined as no HA inhibition. The 20
polyclonal chicken NDV antiserum was prepared at SEPRL by immunization of chickens with 21
inactivated NDV-LaSota. 22
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
10
The sequences for 267 U.S. isolates reported here have the following GenBank accession 1
numbers: 374 bp EF564874-EF565079; 254 bp EF564834-EF564873; F gene EF564813-2
EF564833 (refer to Supplemental Table S1a). Accession numbers for previously published 3
sequences used in the analyses are in Supplemental Table S1b. 4
5
Results 6
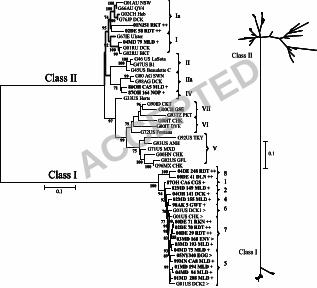
Newcastle disease virus isolates from U.S. LBMs were genetically distinct and 7
phylogenetically distant from vaccine and virulent strains of NDV. A 374 bp region of the 8
fusion protein from 19 U.S. LBM samples during 2005-2006 was sequenced and 9
phylogenetically compared to the corresponding regions of representative viruses from the NDV 10
class I and II clades (n = 81; Figure 1). Based upon analysis of this region all of the U.S. LBM 11
viruses corresponded to the class I clade and were therefore phylogenetically distant from class II 12
viruses such as the vaccine viruses in genotypes I and II and the frequently observed virulent 13
viruses (genotypes II to IX). The largest cluster of U.S. LBM-origin viruses (Figure 1; bolded 14
and designated with the symbol ‘>’) in this study was most closely related to previously reported 15
U.S. LBM-origin viruses from 2001 (G01US DCK1, -DCK2, -CHK; accessions AY626266-68) 16
(45). Recently identified NDV recovered from Hong Kong LBM samples (Figure 1; boxed with 17
asterisk) formed a separate cluster which lacked close association with other viruses. One of the 18
U.S. LBM-origin NDV isolates (05NJ329ENV) grouped together with a duck virus isolated in 19
Taiwan during 1995 (G95T DCK; accession AY135757) (2), and European isolates from 20
Germany (G01DEU DLN, G99DEU DCK; accessions DQ096595, DQ097393) (14) and 21
Denmark (G97DK CHK, -OST; accessions AY175733, AY75744) (2) formed a nearby cluster. 22
Another cluster near to the largest LBM cluster included WS viruses from the U.S. (G80US 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
11
COO; accession DQ096594) (14), New Zealand (G98NZ DCK2; accession AY175774) (2), and 1
Germany (G89DEU DCK; accession AY175732) (2). A virulent virus from Ireland that caused 2
an outbreak in poultry during 1990 (§ vG90IE CHK; accession AY972102) (5) was also found in 3
the class I clade (Figure 1). 4
5
Live bird market-origin isolates were found to be closely related to WS-origin 6
isolates. To investigate the source of the U.S. LBM loNDV they were compared to 249 WS 7
isolates obtained from healthy birds during 1986 to 2005 in eight different states (Table 1 and 8
Supplemental Table S1a). Phylogenetic analysis of the 374 bp region demonstrated that U.S. 9
LBM-origin viruses were related to class I WS-origin isolates and that of the nine class I 10
genotypes identified, four were represented by U.S. LBM viruses (Figure 2a). The largest cluster 11
of U.S. LBM viruses, which were isolated from environmental samples, eggs, chickens and 12
ducks in several northeastern U.S. states during 2005-06, corresponded to genotype 5 (15/19) 13
which also included a large proportion of the U.S. WS isolates (60/173). The WS isolates in 14
closest association with the U.S. LBM viruses were all from mallards (Anas platyrhynchos), the 15
most notable of which was a 1999 isolate from Minnesota (Figure 2a; arrow, black typeface, 16
bolded, 99MNCA8MLD, genotype 5) that is closely related to LBM viruses isolated in the 17
northeastern U.S. during 2005-06. Statistical analysis of the branches of this tree using ML 18
confirmed the significant probability (> 0.01) for the node connecting the 99MNCA8MLD 19
isolate to the cluster of genotype 5 LBM isolates. Interestingly, genotype 6 was found only 20
among the U.S. LBM isolates including previously described isolates from 2001 (G01US DCK1, 21
-CHK; blue typeface, bolded, accessions AY626266 and -68) (45). A single U.S. LBM isolate 22
from New York in 1995 (95NY263CHK; brown typeface, bolded) was classified as genotype 1 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
12
which comprised many of the oldest class I WS isolates identified herein (circa 1987) and it was 1
most similar to 88LACA4BWT (Figure 2a; black typeface, bolded). The only known virulent 2
class I virus appears on a separate branch (§ vG90IECHK; red typeface, bolded, accession 3
AY972102) (5) apparently basal or sister to genotypes 1-7 (Figure 2a; genotype 1), indicating 4
that it represents a separate genotype. The final U.S. LBM isolate (05NJ329ENV; brown 5
typeface, bolded) was categorized in genotype 4 and was most closely related to a 1995 Taiwan 6
duck isolate (G95TW DCK; blue typeface, bolded, accession AY135757) (2) and a U.S. WS 7
isolate from Alaska (98AK5GWT; black typeface, bolded). The LBM viruses from Hong Kong 8
were clearly separated from the other class I viruses and formed genotype 3 (Figure 2a; denoted 9
by asterisk). 10
11
Broad phylogenetic diversity was found among the U.S. WS isolates (n = 249) 12
representing class I and II viruses from nine different genotypes. Seven of the nine novel 13
class I genotypes were represented by U.S. WS isolates (Table 1 and Figure 2a). For the class I 14
WS viruses (Figure 2a; designated by ‘+’), those from Anatidae species (especially those from 15
mallards) were found to be related to the LBM viruses,(Figure 2a; designated by ‘>’ in brown 16
typeface) while the isolates from shorebirds (Figure 2a; designated by ‘++’ in green typeface) 17
tended to form distinct clusters within genotypes 7 and 8 that were distinct from either the 18
Anatidae or LBM viruses. Some temporal and geographic clustering was observed; however, this 19
could be due to the isolation of multiple related samples from the same location, and multiple 20
genotypes were often present in the same year and location (Table 2). 21
The class II WS viruses formed distinct clusters related to the previously described 22
genotypes I and II (Figure 2b); for the purpose of discussion these subgroups will be referred to 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
13
as Ia and IIa. A cluster of mallard isolates from Maryland during 2004 was closely related to 1
viruses in genotype I which are primarily lentogenic viruses such as the 1967 Ulster strain 2
(G67IE Ulster; accession AY562991). However, no viruses closely related to the widely used 3
LaSota and B1 live vaccines were isolated from any of the WS-origin samples. A separate cluster 4
of predominantly shorebird viruses formed a subgroup (genotype Ia; designated by ‘++’) with 5
the viruses endemic to Australia (vG98AU CHK, G98AU CHK, G66AUQV4; accessions 6
AY175722, AY175658, AF217084), New Zealand (G98NZ DCK1, accession AY175730), Asia 7
(G76JP DCK, G02CH Heb; accession M24705, AF217084), and the U.S. (G03CA TKY, G83US 8
LOO; accessions AY175642, AY175736). Of particular note was the presence of an isolate from 9
a domestic 12-week-old turkey from a farm in Ontario, Canada (G03CA TKY) within a closely 10
related cluster of shorebird isolates from the U.S. Anatidae viruses related to genotype II. This 11
cluster, which includes a diverse group of lentogens and mesogens in addition to many vaccine 12
strains, formed another subgroup (IIa) with previously published WS viruses from Argentina 13
(G98AG DCK, G00AG SWN; accessions AY727881, -82) (14) and a chicken isolate from Spain 14
(G88ES CHK, accession AY175642) (2). 15
Overall, the majority of WS-origin viruses were isolated from mallards, which 16
represented nearly 70% of the WS viruses sequenced (169/249; Table 1) and the most frequently 17
observed genotype among these viruses was class I genotype 5 (65/249). The Anatidae species 18
alone were host to genotypes 1, 2, 4, 5, 7, and 9 of class I, and I, Ia, and IIa of class II (Table 1), 19
with genotypes 5 and 7 being identified most often. The shorebird (Scolopacidae) isolates were 20
predominantly of genotype 7, with genotype 5 being conspicuously absent among these viruses. 21
Class II genotype IIa represented the third most common genotype (59/249; Table 1) with 22
mallards representing the most common host. 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
14
1
Diverse WS genotypes were distributed across eight states over the 20 year period. 2
Multiple genotypes circulated across the U.S. over time and within a region during a given year 3
(Table 2); for example, three different class I genotypes occurred in Alaska during 1998 (from 4
Anatidae sp.) and four class I genotypes occurred in Minnesota during 2000 (all from mallards). 5
In Maryland during 2004 this held true for both class I and II viruses as well. Multiple genotypes 6
were also found circulating within a specific source group (species) in the Maryland region 7
(Table 3). Mallards during 2001-2004 (n = 101) were infected by five of the nine class I 8
genotypes (1, 2, 4, 5, and 7). The data in Table 3 also demonstrated a change in the 9
predominating genotype from year to year. Whereas genotype 5 was most commonly observed in 10
this subset during 2001 (16/16), it was not identified again until 2004 (38/54). Interestingly, 11
genotype 5 was most prevalent in mallards in 2004 and in the U.S. LBM isolates during 2005-06 12
(15/17). 13
14
The broad phylogenetic diversity found in the 374 bp F gene analysis was confirmed 15
by analysis of the complete coding region of the F gene. Representative WS viruses (Figure 3; 16
Anatidae designated by ‘+’ and shorebirds designated by ‘++’) and LBM viruses (Figure 3; 17
designated by ‘>’) were analyzed using Phyml ML with bootstrap values for n = 100 replicates 18
(Figure 3; bootstrap values ≥ 70 shown for each major node), and was reproducible using both 19
DNAML and Neighbor Joining techniques (data not shown). These results confirmed the 20
phylogenetic diversity observed using the shorter F gene genomic regions, clearly distinguished 21
the LBM viruses from vaccine and virulent viruses, and validated the close phylogenetic 22
relationship between WS and LBM viruses. 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
15
1
Waterfowl and shorebird isolates from the U.S. possessed a highly attenuated 2
phenotype. Classical pathotyping using MDT, ICPI, and molecular classification based on the 3
deduced amino acid sequence of the fusion cleavage site were used to determine the pathogenic 4
potential of representative WS isolates (Table 4a and b). Eighty WS isolates representing various 5
genotypes among class I and II were pathotyped using the ICPI in day-old chicks (Table 4a). All 6
isolates tested (80/80) had low ICPI values of <0.2 (on a 0-2.0 scale). The MDT assay in eggs 7
was >90 hours (data not shown) for 58 of these 80 isolates, which is characteristic of lentogenic 8
viruses and indicates that the viruses tested have a low potential to cause significant disease in 9
poultry (3). 10
The deduced amino acid sequences confirmed the presence of lentogenic fusion cleavage 11
sites for all WS- and LBM-origin isolates characterized by the limited number of basic amino 12
acids at positions 112-116 and the presence of leucine at position 117 (Table 4b; n = 290). The 13
predominant motif among class I isolates, 110-GGERQERL-117, was found in 85.5% of isolates 14
(183/214) and is common among other reported lentogenic strains (45). This motif was also 15
found among U.S. (3/19) and Hong Kong (10/21) LBM isolates; however, the majority of the 16
U.S. LBM isolates had the single amino acid substitution aspartate for glutamate at position 115 17
(15/19). The remaining LBM isolates from Hong Kong shared two motifs that were unique to 18
that group (11/21). The predominant motif among class II isolates 110-GGEKQGRL-117 had the 19
amino acid substitution lysine for arginine at position 113 and glycine for glutamate at position 20
115. 21
22
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
16
Broad phylogenetic diversity may impact the efficacy of rapid detection methods for 1
class I WS and LBM viruses. To determine whether the WS isolates would be detected using 2
the USDA validated M-gene assay developed for class II NDV, a panel of 29 WS and 21 LBM 3
viruses (n = 19 U.S. and n = 2 Hong Kong) were tested (class I n = 42; II n = 8; Figure 4). Over 4
two thirds (30/42) of class I viruses were suspect or failed to be detected (cycle threshold [CT] 5
value ≥ 35 or zero) and the remainder (10/42) were weakly positive with high CT values (30-34). 6
Although the majority of class II viruses were accurately identified with the M-gene assay (6/8), 7
the remaining two viruses from ruddy turnstones in genotype Ia demonstrated suspect CT values 8
(02NJ60RDT, 02DE54RDT; CT values 34-38). All samples contained quantifiable RNA and 9
produced a product of expected size when amplified using F gene primers in a standard RT-PCR 10
assay, and selected class I viruses were positive (CT values < 30) with a previously developed L-11
gene targeted real time RT-PCR assay (28) (data not shown). 12
The 58 U.S. isolates from Anatidae species during 1986-1999 that were used for MDT 13
determination were also evaluated using a monoclonal antibody (mAb) binding assay with a 14
novel panel (Table 5). Isolates were classified into seven distinct groups based on the mAb 15
binding patterns as follows: group 1 (n = 33) was characterized by binding of mAbs AVS, 15C4, 16
B79, P15D7,P11C9, P3A11; group 1A (n = 3) included binding of mAb P10B8; group 2 17
exhibited binding of 15C4, B79, P15D7,P11C9, P3A11 (n = 1); whereas group 2A did not bind 18
mAb P11C9 (n = 1); the third group (n = 17) bound only to mAb B79; group 4 did not bind to 19
any of the mAbs (n = 2); and group 5 demonstrated binding to mAbs AVS, 15C4, B79, P15D7, 20
and P11C9 (n = 1). Monoclonal antibody groups 2, 3, and 4 correlated with class I viruses, with 21
group 3 representing genotypes 1, 2, 4, 5, 7, and 9 which are found in the Anatidae group. 22
Groups 1, 1A, 2A, and 5 corresponded to the class II viruses, with group I primarily composed of 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
17
genotypes II and IIa. Although differentiation between class I and II viruses was possible using 1
the mAb assay, only a few of the mAbs recognized class I viruses and the assay was not capable 2
of distinguishing differences between class I genotypes. 3
4
Discussion 5
The discovery of a large number of class I NDV viruses recovered from LBMs in the 6
northeastern U.S. and the existence of genetically related viruses found in WS suggest that 7
epidemiological connections may exist between them. Identification of an earlier LBM isolate 8
(Figure 2a; 95NY263CHK) in class I, genotype 1 which is related to the oldest WS-origin 9
viruses analyzed here, and a class I genotype 5 mallard isolate (99MNCA8MLD) that was 10
closely related to the largest cluster of 2005-06 LBM isolates suggest that transmission between 11
WS and birds in the LBM has occurred. While the mallard isolate (99MNCA8MLD) was 12
geographically removed from where the cluster of LBM viruses was isolated (northeastern U.S.), 13
the data for the geographic distribution combined with the natural mobility of wild birds 14
indicates that NDV strains are likely highly mobile. Another explanation for the similarity 15
between the WS- and LBM-origin viruses could be the existence of parallel reservoirs that are 16
either evolutionarily static or evolving in similar directions and at comparable rates; however this 17
would be an evolutionarily implausible scenario based on the rapid capacity for RNA viruses to 18
change, the history of evolution for other NDV (2,51,54), and the year to year changes in the 19
phylogenetic trees presented here. In this study, there were few cases where the class I LBM 20
viral genotypes lacked WS counterparts (e.g. Figure 2a; class I genotype 6 from the U.S. and 21
genotype 3 from Hong Kong) and the data are simply insufficient to determine a viral ancestor. 22
In each of these cases, increased sampling of at risk domestic and wild birds may potentially 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
18
reveal reservoirs of low virulence ND viruses, but the absence of such data does not discard the 1
possibility of viral transmission among these populations. 2
The identification of a class II genotype Ia virus from a 12-week old domestic turkey in 3
Ontario during 2003 (G03CA TKY, Figure 2b) that clustered closely with U.S. shorebird isolates 4
from New Jersey in 2001 (01NJRKN, RDT, SDL) suggests that class II loNDV may also 5
occasionally transmit between wild and domestic populations. It is noteworthy to mention that 6
while billions of doses of live NDV vaccine are used worldwide in poultry, the absence of class 7
II vaccine or vaccine-like viruses in wild birds suggests that transmission from vaccinated 8
domestic poultry into wild birds may not be a frequent occurrence. 9
The data herein support a complex viral ecology for loNDV infections in wild birds 10
because it presents evidence of circulation of more than one genotype within a species and 11
within a geographic location as well as circulation of more than one genotype during the same 12
year. The re-occurrence of genotypes over time presents the possibility of cyclic predominance 13
of various lineages. Cyclic patterns of subtypes have been reported for avian influenza virus 14
where isolation of an HA subtype may be followed one to two years later by reduced isolation 15
rates for the same subtype (39). 16
These data also provide substantial insight into the broad phylogenetic diversity of NDV 17
and emphasize the potential for more genotypes yet to be discovered; however, the study herein 18
is likely not sufficient to establish the precise genotypes that may be encountered. For example, 19
viruses from class I genotypes 1, 2 and 6 were delineated based upon distance and clustering, but 20
they may in fact represent additional genotypes. The ability to discern separate genotypes will 21
improve over time as more viruses are identified and sequenced. The diversity observed at the 22
phylogenetic level would be expected to follow at the antigenic and protein levels as well, as 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
19
suggested by the mAb results and the differences in the deduced F proteins (up to 17.4% amino 1
acid differences, data not shown). Here we have only explored avian WS species from the orders 2
Anseriformes and Charadriiformes, and while NDV has been documented to infect at least 241 3
different bird species (4) other non-domestic avian families in these orders and other non-4
domestic avian orders remain virtually unexplored for loNDV. 5
Although class I and II loNDV viruses in this study were predicted to exhibit lentogenic 6
phenotypes, concern exists regarding possible genetic changes to loNDV upon replication in 7
poultry. Virulent viruses of similar genotypes to those found in WS and LBM have previously 8
been recovered (5,18). The 1990 Ireland outbreak was caused by a virulent class I virus (vG90IE 9
CHK; accession AY972102), which in this study corresponds to genotype 1 or the progenitor-10
type viruses (Figure 2a); and the Australian outbreak of 1998-200 was caused by the mutation to 11
virulence of a class II genotype I virus, which are typically lentogenic. Genomic analysis of the 12
class II genotype I viruses (genotype Ia in the present study which included several species of 13
U.S. waterfowl) from the Australian outbreak provided evidence that lentogenic viruses have the 14
potential to become virulent over time (18), and that a change in the cleavage site of the fusion 15
protein of the native virus resulted in increased virulence (17). 16
The performance of two diagnostic tools commonly used for rapid identification of NDV 17
viruses is also of concern. Approximately 70% of WS-origin isolates identified corresponded to 18
class I viruses (174/249) that are poorly or not detected by the USDA M-gene assay (28,29). The 19
use of mAb assays for rapid characterization may also not be optimal for class I viruses as these 20
assays have predominantly been developed and optimized to recognize class II viruses. Because 21
mAbs are directed against single epitopes, their ability to detect a broad spectrum of viruses is 22
often limited. Preliminary testing using an L-gene targeted real time RT-PCR assay detected 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
20
class I LBM viruses from Hong Kong (28) and other select WS and LBM isolates (data not 1
shown); however, due to the phylogenetic diversity of loNDV, multiple real time RT-PCR tests 2
may be needed to detect all WS genotypes. 3
Active surveillance of LBMs focused on the detection of vNDV is laudable, but many 4
challenges still remain. Live bird markets promote the commingling of multiple bird species in 5
close quarters and provide opportunities for transmission of disease agents (38,40,41,46). In the 6
U.S., LBMs are part of a complex system that provides fresh poultry, typically in larger cities. 7
Because these markets receive birds from multiple sources, confine multiple bird species in close 8
quarters, and provide processing services onsite, they provide an environment in which reservoir 9
species such as waterfowl are closely housed with gallinaceous hosts. While the USDA has been 10
working to control low pathogenic avian influenza (LPAI) in U.S. LBMs since 1986, the 11
Uniform Standards were released in 2004 and the official program was instituted during 2006 12
which includes registration of LBM premises, documentation of test-negative status of birds 13
entering the market, and the testing of premises and examination of records (52). Over 15,000 14
samples were submitted to NVSL specifically for AIV and NDV surveillance in 2003 (Suarez, 15
D.L., Abstr. 108th U.S. Animal Health Association Annual Meeting, p.93, 2004). In 2002, more 16
than 15,000 specimens were tested at the NVSL for AIV and NDV surveillance; however, few of 17
the surveillance samples originated from backyard poultry. Since the 2002-03 outbreak of vNDV 18
in California, Nevada, and Arizona, which was epidemiologically linked to infections in 19
backyard poultry and game fowl, surveillance among this population has been significantly 20
intensified. For WS populations, NDV monitoring has been sporadic and often occurs in 21
conjunction with other monitoring programs such as AIV (8,20,23,24). Detection of vNDV in 22
wild bird populations often occurs after outbreaks in poultry (18,25-27,50). To date, hardly any 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
21
data exist regarding loNDV circulating in WS populations, and their highly attenuated phenotype 1
makes them difficult to track among poultry because they may circulate undetected due to lack 2
of clinical signs. 3
In summary, we have demonstrated the broad genetic diversity and lentogenic 4
presentation of loNDV from WS, and have identified a close phylogenetic proximity with LBM-5
origin loNDV suggesting that viral transmission may occur among wild birds and poultry. 6
Evidence that NDV monitoring in the U.S. using the USDA M-gene assay may not be adequate 7
to detect all NDV from class I and II emphasizes the need for new diagnostic assays to identify 8
circulating reservoirs of loNDV in wild bird populations and LBM poultry. Further 9
epidemiologic studies will be needed to clarify the prevalence and origin of loNDV in wild bird 10
populations and LBM poultry, in addition to the ongoing genetic characterization of new 11
isolates. 12
13
Acknowledgments 14
The authors would like to gratefully acknowledge Joan Beck, Scott Lee, Dawn Williams-15
Coplin, Ginger Goekjian, Britta Hanson, and Page Luttrell for technical assistance, the South 16
Atlantic Area Sequencing Facility for nucleotide sequencing, and Michael Wege for providing 17
samples for screening. 18
This work was funded by USDA CRIS project numbers 6612-32000-039-00D, 6612-19
32000-041-01S, 6612-32000-041-04S, 6612-32000-041-06S and 6612-32000-039-03S and 20
USPEA (SEPEA #332). Mention of trade names or commercial products in this manuscript is 21
solely for the purpose of providing specific information and does not imply recommendation or 22
endorsement by the U.S. Department of Agriculture. 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
22
Reference List 1
2
1. Aldous, E. W., C. M. Fuller, J. K. Mynn, and D. J. Alexander. 2004. A molecular 3
epidemiological investigation of isolates of the variant avian paramyxovirus type 1 virus 4
(PPMV-1) responsible for the 1978 to present panzootic in pigeons. Avian Pathol. 5
33:258-269. 6
2. Aldous, E. W., J. K. Mynn, J. Banks, and D. J. Alexander. 2003. A molecular 7
epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates 8
by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. 9
Avian Pathol. 32:239-256. 10
3. Alexander D.J. 1998. Newcastle disease virus and other avian paramyxoviruses, p. 156-11
163. In Swayne D.E. (ed.), A Laboratory Manual for the Isolation and Identification of 12
Avian Pathogens. The American Association of Avian Pathologists, Kennett Square, PA. 13
4. Alexander, D. J. 2003. Newcastle disease, p. 64-87. In Y. M. Saif, H. J. Barnes, J. R. 14
Glisson, A. M. Fadly, L. R. McDougald, and D. E. Swayne (eds.), Disease of Poultry. 15
Iowa State Press, Ames, IA. 16
5. Alexander, D. J., G. Campbell, R. J. Manvell, M. S. Collins, G. Parsons, and M. S. 17
McNulty. 1992. Characterisation of an antigenically unusual virus responsible for two 18
outbreaks of Newcastle disease in the Republic of Ireland in 1990. Vet.Rec. 130:65-68. 19
6. Alexander, D. J., H. T. Morris, W. J. Pollitt, C. E. Sharpe, R. L. Eckford, R. M. 20
Sainsbury, L. M. Mansley, R. E. Gough, and G. Parsons. 1998. Newcastle disease 21
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
23
outbreaks in domestic fowl and turkeys in Great Britain during 1997. Vet.Rec. 143:209-1
212. 2
7. Alexander, D. J., G. W. Wilson, P. H. Russell, S. A. Lister, and G. Parsons. 1985. 3
Newcastle disease outbreaks in fowl in Great Britain during 1984. Vet.Rec. 117:429-434. 4
8. Alfonso, C. P., B. S. Cowen, and H. van Campen. 1995. Influenza A viruses isolated 5
from waterfowl in two wildlife management areas of Pennsylvania. J.Wildl.Dis. 31:179-6
185. 7
9. Ballagi-Pordany, A., E. Wehmann, J. Herczeg, S. Belak, and B. Lomniczi. 1996. 8
Identification and grouping of Newcastle disease virus strains by restriction site analysis 9
of a region from the F gene. Arch.Virol. 141:243-261. 10
10. Banerjee, M., W. M. Reed, S. D. Fitzgerald, and B. Panigraphy. 1994. Neurotropic 11
velogenic Newcastle disease in cormorants in Michigan: pathology and virus 12
characterization. Avian Dis 38:873-878. 13
11. Bulaga, L. L., L. Garber, D. A. Senne, T. J. Myers, R. Good, S. Wainwright, S. 14
Trock, and D. L. Suarez. 2003. Epidemiologic and surveillance studies on avian 15
influenza in live-bird markets in New York and New Jersey, 2001. Avian Dis. 47:996-16
1001. 17
12. Capua, I., P. M. Dalla, F. Mutinelli, S. Marangon, and C. Terregino. 2002. Newcastle 18
disease outbreaks in Italy during 2000. Vet.Rec. 150:565-568. 19
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
24
13. Collins, M. S., D. J. Alexander, S. Brockman, P. A. Kemp, and R. J. Manvell. 1989. 1
Evaluation of monoclonal antibodies raised against an isolate of the variant avian 2
paramyxovirus type I responsible for the current panzootic in pigeons. Arch.Virol. 3
104:53-62. 4
14. Czegledi, A., D. Ujvari, E. Somogyi, E. Wehmann, O. Werner, and B. Lomniczi. 5
2006. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease 6
virus) and evolutionary implications. Virus Res. 120:36-48. 7
15. Farley, J. M., C. H. Romero, M. G. Spalding, M. L. Avery, and D. J. Forrester. 8
2001. Newcastle disease virus in double-crested cormorants in Alabama, Florida, and 9
Mississippi. J Wildl.Dis 37:808-812. 10
16. Felsenstein, J. 1981. Evolutionary trees from DNA sequences: a maximum likelihood 11
approach. J Mol.Evol. 17:368-376. 12
17. Gould, A. R., E. Hansson, K. Selleck, J. A. Kattenbelt, M. Mackenzie, and A. J. 13
Della-Porta. 2003. Newcastle disease virus fusion and haemagglutinin-neuraminidase 14
gene motifs as markers for viral lineage. Avian Pathol. 32:361-373. 15
18. Gould, A. R., J. A. Kattenbelt, P. Selleck, E. Hansson, A. la-Porta, and H. A. 16
Westbury. 2001. Virulent Newcastle disease in Australia: molecular epidemiological 17
analysis of viruses isolated prior to and during the outbreaks of 1998-2000. Virus Res. 18
77:51-60. 19
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
25
19. Guindon, S., F. Lethiec, P. Duroux, and O. Gascuel. 2005. PHYML Online--a web 1
server for fast maximum likelihood-based phylogenetic inference. Nucleic Acids Res. 2
33:W557-W559. 3
20. Hanson, B. A., D. E. Swayne, D. A. Senne, D. S. Lobpries, J. Hurst, and D. E. 4
Stallknecht. 2005. Avian influenza viruses and paramyxoviruses in wintering and 5
resident ducks in Texas. J Wildl.Dis 41:624-628. 6
21. Haruna, E. S., D. Shamaki, G. O. Echeonwu, K. A. Majiyagbe, Y. Shuaibu, and D. 7
R. Du. 1993. A natural outbreak of Newcastle disease in guinea-fowl (Numida meleagris 8
galeata) in Nigeria. Rev.Sci.Tech. 12:887-893. 9
22. Heckert, R. A., M. S. Collins, R. J. Manvell, I. Strong, J. E. Pearson, and D. J. 10
Alexander. 1996. Comparison of Newcastle disease viruses isolated from cormorants in 11
Canada and the USA in 1975, 1990 and 1992. Can.J.Vet.Res. 60:50-54. 12
23. Hlinak, A., R. U. Muhle, O. Werner, A. Globig, E. Starick, H. Schirrmeier, B. 13
Hoffmann, A. Engelhardt, D. Hubner, F. J. Conraths, D. Wallschlager, H. 14
Kruckenberg, and T. Muller. 2006. A virological survey in migrating waders and other 15
waterfowl in one of the most important resting sites of Germany. J Vet Med.B Infect.Dis 16
Vet Public Health 53:105-110. 17
24. Hlinak, A., T. Muller, M. Kramer, R. U. Muhle, H. Liebherr, and K. Ziedler. 1998. 18
Serological survey of viral pathogens in bean and white-fronted geese from Germany. J 19
Wildl.Dis 34:479-486. 20
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
26
25. Huovilainen, A., C. Ek-Kommone, R. Manvell, and L. Kinnunen. 2001. Phylogenetic 1
analysis of avian paramyxovirus 1 strains isolated in Finland. Arch.Virol. 146:1775-1785. 2
26. Jorgensen, P. H., K. J. Handberg, P. Ahrens, H. C. Hansen, R. J. Manvell, and D. J. 3
Alexander. 1999. An outbreak of Newcastle disease in free-living pheasants (Phasianus 4
colchicus). Zentralbl.Veterinarmed.B 46:381-387. 5
27. Jorgensen, P. H., K. J. Handberg, P. Ahrens, O. R. Therkildsen, R. J. Manvell, and 6
D. J. Alexander. 2004. Strains of avian paramyxovirus type 1 of low pathogenicity for 7
chickens isolated from poultry and wild birds in Denmark. Vet.Rec. 154:497-500. 8
28. Kim, L., D. King, D. Suarez, C. Wong, and C. Afonso. 2007. Characterization of Class 9
I Newcastle disease virus isolates from Hong Kong bird markets and detection using real-10
time reverse transcription PCR. J.Clin.Microbiol 45:1310-1314. 11
29. Kim, L., D. Suarez, and C. Afonso. 2007. Use of bioinformatics in improving detection 12
of Newcastle disease virus, p. 227-249. In M. Rogeria de Almeida, M. Pires Moraes, J. 13
Patarroyo, P. Vidigal, and A. Borem (eds.), Biotechnology and Animal Health 14
International Meeting. Federal University of Viscosa, Viscosa, Brazil. 15
30. King, D. J. 1996. Avian paramyxovirus type 1 from pigeons: isolate characterization and 16
pathogenicity after chicken or embryo passage of selected isolates. Avian Dis. 40:707-17
714. 18
31. King, D. J. 1996. Influence of chicken breed on pathogenicity evaluation of velogenic 19
neurotropic Newcastle disease virus isolates from cormorants and turkeys. Avian Dis. 20
40:210-217. 21
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
27
32. King, D. J. and B. S. Seal. 1997. Biological and molecular characterization of Newcastle 1
disease virus isolates from surveillance of live bird markets in the northeastern United 2
States. Avian Dis. 41:683-689. 3
33. Kommers, G. D., D. J. King, B. S. Seal, and C. C. Brown. 2001. Virulence of pigeon-4
origin Newcastle disease virus isolates for domestic chickens. Avian Dis. 45:906-921. 5
34. Lamb, R., P. L. Collins, D. Kolakofsky, J. A. Melero, Y. Nagai, M. B. A. Oldstone, 6
C. R. Pringle, and B. K. Rima. 2005. p. 655-668. In C. M. Fauquet, M. A. Mayo, J. 7
Maniloff, U. Desselberger, and L. A. Ball (eds.), Virus Taxonomy, Eighth Report of the 8
International Committee on Taxonomy of Viruses. Elsevier Academic Press, San Diego. 9
35. Lana, D. P., D. B. Snyder, D. J. King, and W. W. Marquardt. 1988. Characterization 10
of a battery of monoclonal antibodies for differentiation of Newcastle disease virus and 11
pigeon paramyxovirus-1 strains. Avian Dis 32:273-281. 12
36. Marin, M. C., P. Villegas, J. D. Bennett, and B. S. Seal. 1996. Virus characterization 13
and sequence of the fusion protein gene cleavage site of recent Newcastle disease virus 14
field isolates from the southeastern United States and Puerto Rico. Avian Dis. 40:382-15
390. 16
37. Meulemans, G., T. P. van den Berg, M. Decaesstecker, and M. Boschmans. 2002. 17
Evolution of pigeon Newcastle disease virus strains. Avian Pathol. 31:515-519. 18
38. Mullaney, R. 2003. Live-bird market closure activities in the northeastern United States. 19
Avian Dis 47:1096-1098. 20
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
28
39. Olsen, B., V. J. Munster, A. Wallensten, J. Waldenstrom, A. D. Osterhaus, and R. A. 1
Fouchier. 2006. Global patterns of influenza a virus in wild birds. Science 312:384-388. 2
40. Panigrahy, B., D. A. Senne, and J. C. Pedersen. 2002. Avian influenza virus subtypes 3
inside and outside the live bird markets, 1993-2000: a spatial and temporal relationship. 4
Avian Dis 46:298-307. 5
41. Pedersen, J. C., D. A. Senne, P. R. Woolcock, H. Kinde, D. J. King, M. G. Wise, B. 6
Panigrahy, and B. S. Seal. 2004. Phylogenetic relationships among virulent Newcastle 7
disease virus isolates from the 2002-2003 outbreak in California and other recent 8
outbreaks in North America. J.Clin.Microbiol. 42:2329-2334. 9
42. Pfitzer, S., D. J. Verwoerd, G. H. Gerdes, A. E. Labuschagne, A. Erasmus, R. J. 10
Manvell, and C. Grund. 2000. Newcastle disease and avian influenza A virus in wild 11
waterfowl in South Africa. Avian Dis 44:655-660. 12
43. Rosenberger, J. K., S. Klopp, and W. C. Krauss. 1975. Characterization of Newcastle 13
disease viruses isolated from migratory waterfowl in the Atlantic flyway. Avian Dis 14
19:142-149. 15
44. Seal, B. S., D. J. King, D. P. Locke, D. A. Senne, and M. W. Jackwood. 1998. 16
Phylogenetic relationships among highly virulent Newcastle disease virus isolates 17
obtained from exotic birds and poultry from 1989 to 1996. J.Clin.Microbiol. 36:1141-18
1145. 19
45. Seal, B. S., M. G. Wise, J. C. Pedersen, D. A. Senne, R. Alvarez, M. S. Scott, D. J. 20
King, Q. Yu, and D. R. Kapczynski. 2005. Genomic sequences of low-virulence avian 21
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
29
paramyxovirus-1 (Newcastle disease virus) isolates obtained from live-bird markets in 1
North America not related to commonly utilized commercial vaccine strains. 2
Vet.Microbiol. 106:7-16. 3
46. Senne, D. A., D. L. Suarez, J. C. Pedersen, and B. Panigrahy. 2003. Molecular and 4
biological characteristics of H5 and H7 avian influenza viruses in live-bird markets of the 5
northeastern United States, 1994-2001. Avian Dis 47:898-904. 6
47. Spackman, E., D. E. Stallknecht, R. D. Slemons, K. Winker, D. L. Suarez, M. Scott, 7
and D. E. Swayne. 2005. Phylogenetic analyses of type A influenza genes in natural 8
reservoir species in North America reveals genetic variation. Virus Res. 114:89-100. 9
48. Srinivasappa, G. B., D. B. Snyder, W. W. Marquardt, and D. J. King. 1986. Isolation 10
of a monoclonal antibody with specificity for commonly employed vaccine strains of 11
Newcastle disease virus. Avian Dis. 30:562-567. 12
49. Swayne D.E., Senne D.A., and Beard C.W. 1998. Avian influenza, p. 150-155. In 13
Swayne D.E. (ed.), A Laboratory Manual for the Isolation and Identification of Avian 14
Pathogens. The American Association of Avian Pathologists, Kennett Square, PA. 15
50. Takakuwa, H., T. Ito, A. Takada, K. Okazaki, and H. Kida. 1998. Potentially virulent 16
Newcastle disease viruses are maintained in migratory waterfowl populations. 17
Jpn.J.Vet.Res. 45:207-215. 18
51. Ujvari, D., E. Wehmann, E. F. Kaleta, O. Werner, V. Savic, E. Nagy, G. Czifra, and 19
B. Lomniczi. 2003. Phylogenetic analysis reveals extensive evolution of avian 20
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
30
paramyxovirus type 1 strains of pigeons (Columba livia) and suggests multiple species 1
transmission. Virus Res. 96:63-73. 2
52. USDA APHIS. 2004. Prevention and control of H5 and H7 low pathogenicity avian 3
influenza in the live bird marketing system, p. 1-21. USDA APHIS. 4
53. Wise, M. G., D. L. Suarez, B. S. Seal, J. C. Pedersen, D. A. Senne, D. J. King, D. R. 5
Kapczynski, and E. Spackman. 2004. Development of a real-time reverse-transcription 6
PCR for detection of Newcastle disease virus RNA in clinical samples. J.Clin.Microbiol. 7
42:329-338. 8
54. Yu, L., Z. Wang, Y. Jiang, L. Chang, and J. Kwang. 2001. Characterization of newly 9
emerging Newcastle disease virus isolates from the People's Republic of China and 10
Taiwan. J.Clin.Microbiol. 39:3512-3519. 11
12
13 ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
31
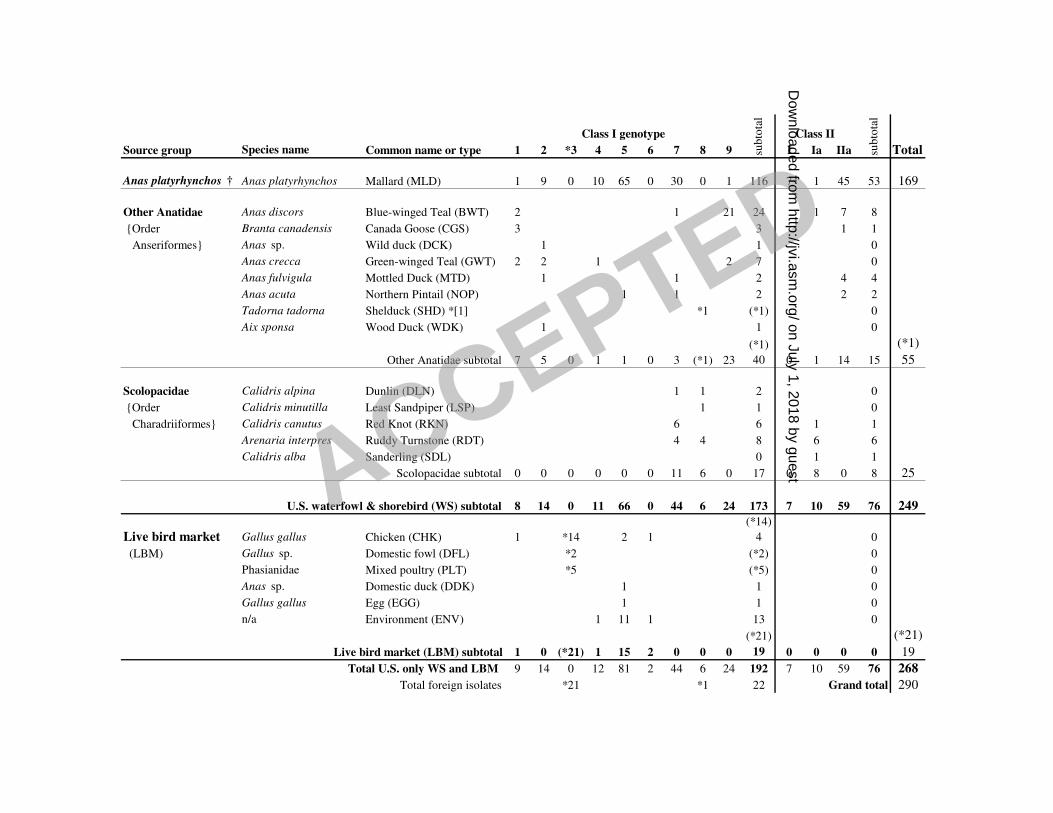
Table 1. Distribution of Newcastle disease isolates by source group, class, and genotype (n = 1
290). Asterisks indicate foreign isolates. The numbers in the body of the table indicate the 2
number of isolates. Order names provided in curved brackets where applicable. Abbreviations 3
for species used in the phylogenetic analysis are indicated in parentheses. 4
5
† includes 1 environmental mallard isolate and 2 Black Duck (Anas rubripes) isolates 6
7
Table 2. Distribution of U.S. Newcastle disease isolates from waterfowl and shorebirds (WS) 8
and live bird markets (LBM) by year, state of collection, class, and genotype (n = 268). The 9
numbers in the body of the table indicate the number of isolates. 10
Class I genotypes Class II
Source Year † State 1 2 4 5 6 7 8 9
sub
tota
l
I Ia IIa
sub
tota
l
Total
WS 1986 OH 1 1 1
1987 LA 2 1 3 1 6 7 10
OH 3 3 3 3 6
1988 LA 2 2 2 6 6
1998 AK 1 1 1 3 3
MN 1 3 4 6 6 10
1999 MN 4 4 21 21 25
2000 DE 7 3 10 10
MN 1 1 1 1 4 4 4 8
NJ 1 1 2 2
2001 DE 2 2 2
MD 16 16 5 5 21
NJ 1 1 4 4 5
OH 1 1 1
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
32
TX 1 1 4 4 5
2002 DE 1 1 3 3 4
MD 6 9 15 2 2 17
NJ 1 1 1
OH 1 1 1
TX 21 21 21
2003 MD 2 15 17 4 4 21
OH 3 1 4 4
2004 DE 1 1 1
MD 1 38 14 53 7 1 1 9 62
OH 1 1 1 1 2
WS Total 8 14 11 66 0 44 6 24 173 7 10 59 76 249
LBM 1995 NY 1 1 1
2005 MA 1 1 1
NJ 1 7 1 9 9
NY 5 1 6 6
RI 1 1 1
2006 NJ 1 1 1
LBM Total 1 0 1 15 2 0 0 0 19 0 0 0 0 19
Grand Total 9 14 12 81 2 44 6 24 192 7 10 59 76 268
† U.S. abbreviations as follows: Alaska = AK, Delaware = DE, Louisiana = LA, Massachusetts 1
= MA, Maryland = MD, Minnesota = MN, New Jersey = NJ, New York = NY, Ohio = OH, 2
Rhode Island = RI, Texas = TX. 3
4
Table 3. Distribution U.S. class I Newcastle disease isolates from resident, wild mallard ducks 5
(Anas platyrhynchos) collected in Maryland by year and genotype (n = 101). Only those 6
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
33
genotypes represented by isolates are shown (5 of 9 genotypes). The numbers in the body of the 1
table indicate the number of isolates. 2
Collection Class I genotype
Source group year 1 2 4 5 7 Total
Anas platyrhynchos 2001 16 16
2002 6 9 15
2003 2 15 17
2004 1 38 14 53
Total 1 8 9 54 29 101
3
Table 4. a) Results for intracerebral pathogenicity assay (ICPI) of Newcastle disease isolates by 4
class and genotype (n = 80). Viruses with an ICPI of < 0.7 are considered lentogens. b) Deduced 5
fusion cleavage site amino acid motifs, position 110-117, for Newcastle disease isolates from 6
waterfowl and live bird market samples; results by class and genotype (n = 290). Asterisks 7
denote foreign isolates. 8
a) 9
Class I genotype Class II
ICPI 1 2 4 5 7 8 9 I Ia IIa Total
0-0.1 8 5 1 8 7 2 3 1 5 33 73
>0.1-0.2 1 2 4 7
8 6 1 10 7 2 3 1 5 37 80
b) 10
Fusion cleavage Class I genotype Class II genotype
site motif 1 2 3 4 5 6 7 8 9 I Ia IIa Total
EGERQERL *5 5
GGEQQERL *6 6
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
34
GGERQDRL 16 1 17
GGERQERL 12 11 *10 13 59 1 46 6 (*1) 24 183
GGERQGRL 1 1
GGVRQERL 2 2
EGGKQGRL 4 4
GGEKQGRL 46 46
GGGKQGRL 8 5 8 21
GVEKQGRL 1 1
RGGKQGRL 4 4
Total 12 11 *21 13 75 2 49 6 (*1) 24 8 9 59 290
1
Table 5. Monoclonal antibody binding (mAb) patterns for class I (n = 20) and class II (n = 38) 2
Newcastle disease isolates obtained from Anatidae species during 1986-1999. Seven different 3
mAb patterns differentiated the viruses by class and distinguished some by genotype. 4
† mAb pattern
mAb Class I Class II
group
1 2 4 5 7 9 Ia IIa Total
2 --+++++-- 1 1
3 ---+----- 7 1 1 1 5 2 17
4 --------- 2 2
1 +-+++++-- 1 32 33
1A +-++++++- 3 3
2A --+++-+-- 1 1
5 +-+++---- 1 1
10 1 1 1 5 2 1 22 58
† (+) = positive binding, (-) = no binding. 5
AV
S
10D
11
15C
4
B79
P15D
7
P11C
9
P3A
11
P10B
8
617/1
6 ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
35
Figure 1. Phylogenetic comparison of a 374 bp region of the fusion gene from HI-positive, 1
matrix-real time RT-PCR-negative U.S. live bird market (LBM) viruses (bolded, designated by 2
‘>,’ n = 19; previously published designated with a G, n = 3) to recent Hong Kong LBM isolates 3
(boxed with asterisk; n = 21) and representative Newcastle disease viruses from all known 4
lineages (class I, n = 9; II, n = 29). The tree was constructed using Phyml Maximum Likelihood 5
with 100 bootstrap replicates. The symbol § at isolates in class I and in class II, genotype I, 6
represents atypical virulent viruses. Vaccine viruses are in genotypes I (G67IE Ulster, G66AU 7
QV4) and II. All known class II viruses in genotypes II to IX are virulent viruses. Inset displays 8
the distance between class I and II viruses with the large cluster of U.S. LBM isolates circled and 9
the Hong Kong LBM circled with an asterisk. Class II genotypes are denoted in brackets. Scale 10
indicates the branch length based on the number of nucleotide substitutions per site. 11
12
Virus designations represent an 8-10 character name containing the 2 digit year of collection, 13
location abbreviation, unique virus identification (1-3 characters), and species abbreviation (refer 14
to Supplemental Table S1a). Previously published GenBank sequences are denoted by the letter 15
G, the 2 digit year, location abbreviation, and either a species abbreviation or common name 16
(refer to Supplemental Table S1b). 17
18
Figure 2. a) Phylogenetic analysis of the 374 bp fusion gene fragment for class I waterfowl and 19
shorebird (waterfowl designated by ‘+’ in black typeface; shorebird designated by ‘++’ in green 20
typeface) and live bird market isolates (LBM designated by ‘>’ in brown typeface) with 21
previously published GenBank sequences for reference (designated with a G in blue typeface). 22
The tree was constructed using Phyml Maximum Likelihood with 100 bootstrap replicates. 23
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
36
Asterisk indicates LBM isolates from Hong Kong; arrow represents close relation to LBM 1
viruses; § indicates a virulent isolate, which as a single outlier was not assigned a genotype. Inset 2
displays the relative distance between genotypes, and demarcations show locations of LBM 3
isolates. Scale indicates the branch length based on the number of nucleotide substitutions per 4
site. 5
6
7
b) Phylogenetic analysis of the 374bp fusion gene fragment for class II waterfowl (designated by 8
‘+’) and shorebird isolates (designated by ‘++’) with previously published GenBank sequences 9
for reference (designated with a G). The tree was constructed using Phyml Maximum Likelihood 10
with 100 bootstrap replicates. Vaccine viruses are in genotypes I (G67IE Ulster, G66AU QV4) 11
and II (G46US LaSota, G89US VGGA). Inset displays the relative distance between genotypes. 12
Scale indicates the branch length based on the number of nucleotide substitutions per site. 13
14
15
Figure 3. Phylogenetic comparison of complete coding region of the fusion gene from U.S. live 16
bird market (LBM) viruses (designated by ‘>’; n = 2) and waterfowl (designated by ‘+’) and 17
shorebird (designated by ‘++’) isolates (WS; n = 21) with previously published Newcastle 18
disease viruses from other known genotypes as reference (all designated by a G: class I LBM 19
designated by ‘>,’ n = 3; class II, n = 25). The tree was constructed using Phyml Maximum 20
Likelihood with 100 bootstrap replicates. Genotypes for Class I and II are denoted in brackets; 21
arrow within genotype 5 represents close relation to LBM viruses. Inset displays the distance 22
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
37
between Class I and Class II viruses. Scale indicates the branch length based on the number of 1
nucleotide substitutions per site. 2
3
4
Figure 4. Distribution of cycle threshold values for 50 Newcastle disease isolates from 5
waterfowl and shorebirds (WS; n = 29) and live bird markets (LBM; n = 21) representing class I 6
(n = 42) and II (n = 8) using the USDA matrix gene-targeted real time RT-PCR assay. Class I 7
genotype 3 contains only isolates from LBM in Hong Kong. Cycle threshold (CT) value 8
interpretation: lower values indicate earlier detection; values ≥35 were considered suspect; a CT 9
value of zero is negative for the assay. 10
11
Supplemental Table S1. a) List of waterfowl (WS) and live bird market (LBM) isolates sorted 12
by class, genotype and species. Asterisk indicates isolates which had only the 254 bp fragment of 13
the matrix to fusion genes amplified. b) Previously published GenBank accessions used in 14
phylogenetic analysis. 15
16 ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Source group Species name Common name or type 1 2 *3 4 5 6 7 8 9 I Ia IIa Total
Anas platyrhynchos † Anas platyrhynchos Mallard (MLD) 1 9 0 10 65 0 30 0 1 116 7 1 45 53 169
Other Anatidae Anas discors Blue-winged Teal (BWT) 2 1 21 24 1 7 8
{Order Branta canadensis Canada Goose (CGS) 3 3 1 1
Anseriformes} Anas sp. Wild duck (DCK) 1 1 0
Anas crecca Green-winged Teal (GWT) 2 2 1 2 7 0
Anas fulvigula Mottled Duck (MTD) 1 1 2 4 4
Anas acuta Northern Pintail (NOP) 1 1 2 2 2
Tadorna tadorna Shelduck (SHD) *[1] *1 (*1) 0
Aix sponsa Wood Duck (WDK) 1 1 0
Other Anatidae subtotal 7 5 0 1 1 0 3 (*1) 23
(*1)
40 0 1 14 15
(*1)
55
Scolopacidae Calidris alpina Dunlin (DLN) 1 1 2 0
{Order Calidris minutilla Least Sandpiper (LSP) 1 1 0
Charadriiformes} Calidris canutus Red Knot (RKN) 6 6 1 1
Arenaria interpres Ruddy Turnstone (RDT) 4 4 8 6 6
Calidris alba Sanderling (SDL) 0 1 1
Scolopacidae subtotal 0 0 0 0 0 0 11 6 0 17 0 8 0 8 25
Live bird market Gallus gallus Chicken (CHK) 1 *14 2 1
(*14)
4 0
(LBM) Gallus sp. Domestic fowl (DFL) *2 (*2) 0
Phasianidae Mixed poultry (PLT) *5 (*5) 0
Anas sp. Domestic duck (DDK) 1 1 0
Gallus gallus Egg (EGG) 1 1 0
n/a Environment (ENV) 1 11 1 13 0
1 0 (*21) 1 15 2 0 0 0
(*21)
19 0 0 0 0
(*21)
19
Total U.S. only WS and LBM 9 14 0 12 81 2 44 6 24 192 7 10 59 76 268
Total foreign isolates *21 *1 22 Grand total 290
24914 76U.S. waterfowl & shorebird (WS) subtotal 5970
sub
tota
l
sub
tota
l
Live bird market (LBM) subtotal
Class II Class I genotype
66 08 11 1044 6 24 173ACCEPTED on July 1, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
*G05HK PLT >G05HK CHK >G05HK CHK >
G04HK DFLG04HK CHK >
G05HK64DFLG05HK365 PLT >
G05HK355 CHK >G03HK374 CHK >G03HK372PLT >G03HK371 CHK >
G03HK377PLT >G03HK373PLT >
G03HK378 CHK >G03HK375 CHK >
G05HK366 CHK >G05HK362 CHK >G05HK363 CHK >G05HK361 CHK >G05HK360 CHK >G05HK359 CHK >
G97DK OSTG97DK CHKG01DEU DLN
G99DEU DCK05NJ329 ENV >
G95TW DCK05NY342 CHK >
05NJ339 ENV >
G01US DCK1 >G01US CHK >
G01US DCK2 >05NJ341 ENV >
05RI337DCK
05NY336 ENV >
05NJ333 ENV >
05NJ344 ENV >
05NJ346 ENV >
05NY347 ENV >
05NJ348 ENV >
05NY331 ENV >
05NY334 CHK >
05MA330 ENV >
05NJ332 ENV >
06NJ351 ENV >
05NY340EGG
05NJ338 CHK >
95NY263 CHK >
G80US COOG98NZ DCK2G89DEU DCK
vG90IE CHK §G97AG PGN
G00IT PGNG72US Fontana
G93TZ PKTG90ID CKT
G00IT CHKG93US ANH
G02US GFLG70AG CHKG60MY CHK
G33US HertsG51JP CHK
G32AU CHK G85CH CHK G97CH GSE
G00AG SWNG98AG DCK
G88ES CHKG48USRoakinG45US Beaudette C
G01RU DCKG02RU BKT
G95IE DCKG67IE Ulster
G03CA TKYG83US LOO
G98NZ DCK1
vG98AU CHK §G66AU QV4
100
98
100
100
96
92
94
99
99
100
93
100
83
100
97
70
77
96
75
100
85
75
90
8984
100
0.2
Class II
Class I
IIIIX
II
I
VIIIIV
VII
V
VI
0.2
Class IIClass I
*
*G05HK PLT >G05HK CHK >G05HK CHK >
G04HK DFLG04HK CHK >
G05HK64DFLG05HK365 PLT >
G05HK355 CHK >G03HK374 CHK >G03HK372PLT >G03HK371 CHK >
G03HK377PLT >G03HK373PLT >
G03HK378 CHK >G03HK375 CHK >
G05HK366 CHK >G05HK362 CHK >G05HK363 CHK >G05HK361 CHK >G05HK360 CHK >G05HK359 CHK >
G97DK OSTG97DK CHKG01DEU DLN
G99DEU DCK05NJ329 ENV >
G95TW DCK05NY342 CHK >
05NJ339 ENV >
G01US DCK1 >G01US CHK >
G01US DCK2 >05NJ341 ENV >
05RI337DCK
05NY336 ENV >
05NJ333 ENV >
05NJ344 ENV >
05NJ346 ENV >
05NY347 ENV >
05NJ348 ENV >
05NY331 ENV >
05NY334 CHK >
05MA330 ENV >
05NJ332 ENV >
06NJ351 ENV >
05NY340EGG
05NJ338 CHK >
95NY263 CHK >
G80US COOG98NZ DCK2G89DEU DCK
vG90IE CHK §G97AG PGN
G00IT PGNG72US Fontana
G93TZ PKTG90ID CKT
G00IT CHKG93US ANH
G02US GFLG70AG CHKG60MY CHK
G33US HertsG51JP CHK
G32AU CHK G85CH CHK G97CH GSE
G00AG SWNG98AG DCK
G88ES CHKG48USRoakinG45US Beaudette C
G01RU DCKG02RU BKT
G95IE DCKG67IE Ulster
G03CA TKYG83US LOO
G98NZ DCK1
vG98AU CHK §G66AU QV4
100
98
100
100
96
92
94
99
99
100
93
100
83
100
97
70
77
96
75
100
85
75
90
8984
100
0.2
Class II
Class I
IIIIX
II
I
VIIIIV
VII
V
VI
*G05HK PLT >G05HK CHK >G05HK CHK >
G04HK DFLG04HK CHK >
G05HK64DFLG05HK365 PLT >
G05HK355 CHK >G03HK374 CHK >G03HK372PLT >G03HK371 CHK >
G03HK377PLT >G03HK373PLT >
G03HK378 CHK >G03HK375 CHK >
G05HK366 CHK >G05HK362 CHK >G05HK363 CHK >G05HK361 CHK >G05HK360 CHK >G05HK359 CHK >
G97DK OSTG97DK CHKG01DEU DLN
G99DEU DCK05NJ329 ENV >
G95TW DCK05NY342 CHK >
05NJ339 ENV >
G01US DCK1 >G01US CHK >
G01US DCK2 >05NJ341 ENV >
05RI337DCK
05NY336 ENV >
05NJ333 ENV >
05NJ344 ENV >
05NJ346 ENV >
05NY347 ENV >
05NJ348 ENV >
05NY331 ENV >
05NY334 CHK >
05MA330 ENV >
05NJ332 ENV >
06NJ351 ENV >
05NY340EGG
05NJ338 CHK >
95NY263 CHK >
G80US COOG98NZ DCK2G89DEU DCK
vG90IE CHK §G97AG PGN
G00IT PGNG72US Fontana
G93TZ PKTG90ID CKT
G00IT CHKG93US ANH
G02US GFLG70AG CHKG60MY CHK
G33US HertsG51JP CHK
G32AU CHK G85CH CHK G97CH GSE
G00AG SWNG98AG DCK
G88ES CHKG48USRoakinG45US Beaudette C
G01RU DCKG02RU BKT
G95IE DCKG67IE Ulster
G03CA TKYG83US LOO
G98NZ DCK1
vG98AU CHK §G66AU QV4
100
98
100
100
96
92
94
99
99
100
93
100
83
100
97
70
77
96
75
100
85
75
90
8984
100
0.20.2
Class II
Class I
IIIIXIX
IIII
II
VIIIIV
VII
V
VI
VII
V
VI
0.2
Class IIClass I
* 0.2
Class IIClass I
* 0.2
Class IIClass I
0.20.20.2
Class IIClass I
*
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
§
0.02
5
6
7
4
*3
1
9
8
2
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
0.02
*3
1
9
8
6
7
4
2
5
99MN CA8 MLD +
§
0.02
5
6
7
4
*3
1
9
8
2
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
0.02
*3
1
9
8
6
7
4
2
5
§
0.02
5
6
7
4
*3
1
9
8
2
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
0.02
§
0.02
5
6
7
4
*3
1
9
8
2
§
0.02
5
6
7
4
*3
1
9
8
2
§
0.02
5
6
7
4
*3
1
9
8
2
§
0.020.02
5
6
7
4
*3
1
9
8
2
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
0.02
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +
04MD 139 MLD +04MD 134 MLD +04MD 136 MLD +04MD 138 MLD +04MD 113 MLD +04MD 117 MLD +04MD 115 MLD +04MD 119 MLD +04MD 121 MLD +04MD 124 MLD +04MD 104 MLD +04MD 108 MLD +04MD 101 MLD +04MD 100 MLD +04MD 99 MLD +04MD 97 MLD +04MD 96 MLD +04MD 94 MLD +04MD 93 MLD +04MD 92 MLD +
04MD 128 MLD +04MD 126 MLD +04MD 110 MLD +
04MD 109 MLD +04MD 107 MLD +04MD 125 MLD +04MD 123 MLD +04MD 122 MLD +04MD 120 MLD +04MD 118 MLD +04MD 116 MLD +04MD 137 MLD +04MD 114 MLD +04MD 112 MLD +04MD 135 MLD +04MD 130 MLD +04MD 129 MLD +04MD 140 MLD +
03OH 189 MLD +03OH 188 MLD +03OH 186 MLD +01MD 196 BLK +
01MD 194 MLD +
05RI 337 DCK >05NJ 341 ENV >
06NJ 351 ENV >05NJ 348 ENV >05NY 347 ENV >05NJ 346 ENV >05NJ 344 ENV >05NY 336 ENV >05NY 340 EGG05NJ 338 CHK 05NJ 333 ENV >05NJ 332 ENV >05NY 331 ENV >
05NY 334 CHK > 05MA 330 ENV >G01US DCK2 >
G98AK6 NOP +00MN 272 MLD +
01MD 214 MLD +01MD 212 MLD +01MD 211 MLD +01MD 210 MLD +01MD 209 MLD +01MD 208 MLD +01MD 207 MLD +01MD 205 MLD +01MD 204 MLD +01MD 201 MLD +01MD 202 MLD +01MD 200 MLD +
01MD 199 MLD +01MD 198 MLD +
05NY 342 CHK > 05NJ 339 ENV >
G01US DCK1 >G01US CHK >
98AK9 NOP +04MD 80 MLD +04MD 81 MLD +
04MD 75 MLD +04MD 74 MLD +04MD 85 MLD +
00MN 290 MLD + 03MD 193 MLD +03OH 191 BWT +
04MD 89 MLD +04MD 91 MLD +04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
04MD 77 MLD +04MD 90 MLD +04MD 87 MLD +04MD 83 MLD +
04MD 88 MLD +01TX 274 MTD +
00DE 32 RDT ++00NJ 33 RKN ++00DE 29 RDT ++00DE 34 RDT ++00DE 36 RDT ++00DE 38 RKN ++00DE 39 DLN ++
04MD 144 MLD +00DE 71 RKN ++01DE 47 RKN ++01DE 46 RKN ++04MD 142 MLD +01NJ 44 RKN ++
03MD 182 MLD +03MD 183 MLD +03MD 181 MLD +03MD 179 MLD +03MD 178 MLD +03MD 177 MLD +03MD 176 MLD +03MD 175 MLD +03MD 173 MLD +03MD 172 MLD +03MD 171 MLD +03MD 168 ENV +
03MD 180 MLD +03MD 174 MLD +G95TW DCK
05NJ 329 ENV >98AK5 GWT +
00MN 289 MLD +02MD 161 MLD +02MD 162 MLD +02MD 160 MLD +02MD 159 MLD +02MD 155 MLD +02MD 158 MLD +02MD 154 MLD +
02MD 153 MLD +02MD 152 MLD +
02MD 148 MLD +02MD 147 MLD +02MD 151 MLD +
02MD 149 MLD +02MD 150 MLD +
98MN 266 MLD +02MD 157 MLD +
04OH 141 DCK02OH 156 WDK +03MD 185 MLD +03MD 184 MLD +G97DK OSTG97DK CHK
G01DEU DLN G99DEU DCKG05HK 366 CHK > G05HK 359 CHK > G05HK 360 CHK > G05HK 361 CHK > G05HK 362 CHK >G05HK 363 CHK >
G03HK 378 CHK > G03HK 375 CHK > G03HK 374 CHK >
G03HK 372PLT >G03HK 371 CHK >
G03HK 373PLT >G03HK 377PLT >
G05HK 365PLT >G05HK 355 CHK >
05HK 364DFL >04HK 367DFL >04HK 368 CHK >
G05HK 358 CHK > G05HK 357PLT >G05HK 356 CHK >
04MD 98 MLD +87OH CA7 CGS +87OH CA6 CGS +87LA CA3 GWT +87OH 264 CGS +
95NY 263 CHK > 88LA CA4 BWT +
88LA CA1 GWT +Gv90IE CHK §
04DE 248 RDT ++02DE 67 RDT ++
00DE 31 RDT ++00DE 41 DLN ++00NJ 40LSS ++00DE 35 RDT ++
G80US COOG98NZ DCK2G89DEU DCK
G77FR SHD88LA CA2 GWT +
00MN 271 MLD + 02TX 321 BWT +02TX 320 BWT +02TX 319 BWT +02TX 318 BWT +02TX 323 BWT +02TX 322 BWT +02TX 324 BWT +02TX 317 BWT +02TX 316 BWT +02TX 315 BWT +02TX 314 BWT +02TX 313 BWT +02TX 312 BWT +02TX 311 BWT +02TX 310 BWT +02TX 309 BWT +02TX 307 BWT +02TX 308 BWT +02TX 280 BWT +02TX 279 BWT +02TX 278 BWT +
95
91
99
73
100
75
100
80
86
71
78
9460
88
78
100
100
97
9371
90
96
99
91
100
97
94
100
100
100
100100
82
95
100
87100
100
100
0.020.02
*3
1
9
8
6
7
4
2
5
*3
1
9
8
*3*3
11
99
88
6
7
4
2
5
66
77
44
22
55
99MN CA8 MLD + 99MN CA8 MLD +
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
00MN 273 MLD +03MD 170 MLD +
98MN 267 MLD +00MN 269 MLD +
01MD 216 MLD +00MN 270 MLD +04OH 131 MLD +98MN 268 MLD +98MN 265 MLD +
00MN 297 MLD +99MN 284 NOP +
01MD 217 MLD +03MD 192 MLD +
03MD 190 MLD +01MD 219 MLD +99MN 165 MLD +99MN 283 MLD +99MN 163 MLD +
01MD 220 MLD +01MD 215 MLD +
02MD 146 MLD +02MD 145 MLD +
04MD 143 MLD +86OH CA5 MLD +03MD 167 MLD +
G00AG SWNG98AG DCK
G88ES CHK87OH 164 NOP +
01OH 197 BWT01TX 306 MTD +01TX 277 MTD +01TX 276 MTD +01TX 275 MTD +
G48US Roakin
G45US Beaudette CG46US LaSota
G00CH GSE
G89US VGGAG97CH UNK
04MD 79 MLD +04MD 78 MLD +04MD 73 MLD +04MD 72 MLD +04MD 133 MLD +04MD 86 MLD +04MD 84 MLD +
G01RU DCKG02RU BKT
G95IE DCKG67IE Ulster
vG98AU CHK §
G98AU CHK98NZ DCK1G66AU QV4
G76JP DCK
G02CH Heb 01NJ 51 RKN ++01NJ 50 RDT ++
01NJ 48 RDT ++G03CA TKY01NJ 45 SDL ++
G83US LOO02NJ 60 RDT ++
02DE 58 RDT ++04MD 95 MLD +
02DE 70 RDT ++
02DE 54 RDT ++G85CH CHKG97CH GSE
G33US HertsG51JP CHK
G32AU CHKG70AG CHK
G60MY CHKG02US GFL
G96MX CHKG00HN CHK
G93US ANHG71US MXD
G82IT PGNG99AG PGN
G97AG PGNG00IT PGNG90KY CHK
G72US FontanaG74RU CKN
G93TZ PKTG00IT CHK
G95TW DFLG90ID CKT
G00TW CHKG98CH CHK
G01CH GSE
100
100
70
92
99
97
96
93
97
100
85
100
100
78
92
74
100
100
100
74
96
87
99
99
100
83
90
100
99
98
70
91100
77
88
79
73
79
99
76100
0.05
III/IV
IX
VIII
IIa
II
Ia
I
V
VI
VII
0.05
IIa
II
I
Ia
IXIII/IV
VIII
VVI VII
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
0.1
Class II
Class I
G01AU NSWG66AU QV4
G02CH HebG76JP DCK
01NJ51 RKT ++
02DE 58 RDT ++
G67IE Ulster04MD 79 MLD +
G01RU DCKG02RU BKT
G46 US LaSotaG47US B1
G45US Beaudette C G00 AG SWNG98AG DCK
86OH CA5 MLD +
87OH 164 NOP +
G33US HertsG90ID CKT
G00CH GSEG93TZ PKT
G00IT CHKG00IT DVE
G72US FontanaG92US TKY
G93US ANHG71US MXD
G00HN CHKG02US GFL
G96MX CHK04DE 248 RDT ++
00DE 41 DLN ++
87OH CA6 CGS +
02MD 149 MLD +
04OH 141 DCK +
02MD 155 MLD +
98AK 5 GWT +
G01US DCK1 >
G01US CHK >00DE 71 RKN ++
02DE 70 RDT ++
00DE 29 RDT ++
03MD 168 ENV >
03MD 193 MLD +
04MD 75 MLD +
05NY340 EGG >
99MN CA8 MLD +
01MD 194 MLD +
04MD 94 MLD +
01MD 208 MLD +
G01US DCK2 >
100
100
100
74
99
100
92
95
100
100
99
71
96
79
100
100
75
99
97
93
100
99
98
98
99
89
70
83
92
77
94
100
100 8
1
2
4
6
7
5
0.1
Class II
Class I
Ia
I
II
IIa
IV
VII
VI
V0.1
Class II
Class I
0.10.1
Class II
Class I
G01AU NSWG66AU QV4
G02CH HebG76JP DCK
01NJ51 RKT ++
02DE 58 RDT ++
G67IE Ulster04MD 79 MLD +
G01RU DCKG02RU BKT
G46 US LaSotaG47US B1
G45US Beaudette C G00 AG SWNG98AG DCK
86OH CA5 MLD +
87OH 164 NOP +
G33US HertsG90ID CKT
G00CH GSEG93TZ PKT
G00IT CHKG00IT DVE
G72US FontanaG92US TKY
G93US ANHG71US MXD
G00HN CHKG02US GFL
G96MX CHK04DE 248 RDT ++
00DE 41 DLN ++
87OH CA6 CGS +
02MD 149 MLD +
04OH 141 DCK +
02MD 155 MLD +
98AK 5 GWT +
G01US DCK1 >
G01US CHK >00DE 71 RKN ++
02DE 70 RDT ++
00DE 29 RDT ++
03MD 168 ENV >
03MD 193 MLD +
04MD 75 MLD +
05NY340 EGG >
99MN CA8 MLD +
01MD 194 MLD +
04MD 94 MLD +
01MD 208 MLD +
G01US DCK2 >
100
100
100
74
99
100
92
95
100
100
99
71
96
79
100
100
75
99
97
93
100
99
98
98
99
89
70
83
92
77
94
100
100 8
1
2
4
6
7
5
0.1
Class II
Class I
Ia
I
II
IIa
IV
VII
VI
V
G01AU NSWG66AU QV4
G02CH HebG76JP DCK
01NJ51 RKT ++
02DE 58 RDT ++
G67IE Ulster04MD 79 MLD +
G01RU DCKG02RU BKT
G46 US LaSotaG47US B1
G45US Beaudette C G00 AG SWNG98AG DCK
86OH CA5 MLD +
87OH 164 NOP +
G33US HertsG90ID CKT
G00CH GSEG93TZ PKT
G00IT CHKG00IT DVE
G72US FontanaG92US TKY
G93US ANHG71US MXD
G00HN CHKG02US GFL
G96MX CHK04DE 248 RDT ++
00DE 41 DLN ++
87OH CA6 CGS +
02MD 149 MLD +
04OH 141 DCK +
02MD 155 MLD +
98AK 5 GWT +
G01US DCK1 >
G01US CHK >00DE 71 RKN ++
02DE 70 RDT ++
00DE 29 RDT ++
03MD 168 ENV >
03MD 193 MLD +
04MD 75 MLD +
05NY340 EGG >
99MN CA8 MLD +
01MD 194 MLD +
04MD 94 MLD +
01MD 208 MLD +
G01US DCK2 >
100
100
100
74
99
100
92
95
100
100
99
71
96
79
100
100
75
99
97
93
100
99
98
98
99
89
70
83
92
77
94
100
100
G01AU NSWG66AU QV4
G02CH HebG76JP DCK
01NJ51 RKT ++
02DE 58 RDT ++
G67IE Ulster04MD 79 MLD +
G01RU DCKG02RU BKT
G46 US LaSotaG47US B1
G45US Beaudette C G00 AG SWNG98AG DCK
86OH CA5 MLD +
87OH 164 NOP +
G33US HertsG90ID CKT
G00CH GSEG93TZ PKT
G00IT CHKG00IT DVE
G72US FontanaG92US TKY
G93US ANHG71US MXD
G00HN CHKG02US GFL
G96MX CHK04DE 248 RDT ++
00DE 41 DLN ++
87OH CA6 CGS +
02MD 149 MLD +
04OH 141 DCK +
02MD 155 MLD +
98AK 5 GWT +
G01US DCK1 >
G01US CHK >00DE 71 RKN ++
02DE 70 RDT ++
00DE 29 RDT ++
03MD 168 ENV >
03MD 193 MLD +
04MD 75 MLD +
05NY340 EGG >
99MN CA8 MLD +
01MD 194 MLD +
04MD 94 MLD +
01MD 208 MLD +
G01US DCK2 >
100
100
100
74
99
100
92
95
100
100
99
71
96
79
100
100
75
99
97
93
100
99
98
98
99
89
70
83
92
77
94
100
100 8
1
2
4
6
7
5
0.1
Class II
Class I
Ia
I
II
IIa
IV
VII
VI
V
8
1
2
4
6
7
5
8
1
2
4
6
7
5
8
1
2
4
6
7
5
0.1
Class II
Class I
Ia
I
II
IIa
IV
VII
VI
V
0.10.1
Class II
Class I
Ia
I
Ia
I
II
IIa
IV
II
IIa
IV
VII
VI
VII
VI
VV
ACCEPTED
on July 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
0
10
20
30
40
87LACA4BWT
87OHCA7CGS
87LACA3GWT
87LACA1GWT
95NY263CHK
04OH141WD
02MD148MLD
02MD151MLD
05HK359CHK
05HK363CHK
98AK5GWT
05NJ329ENV
99MNCA8MLD
04MD113MLD
04MD114MLD
98AK6NOP
05MA330ENV
05NY331ENV
05NJ332ENV
05NJ333ENV
05NY334CHK
05UN336ENV
05RI337ENV
05NJ338CHK
05NY340CHK
05NJ341ENV
05NJ344UNK
05NJ345UNK
05NJ346ENV
05NY347UNK
05NJ348ENV
05NJ339ENV
05NY342CHK
00DE39DLN
98AK9NOP
00DE29RDT
00DE36RDT
01DE47RKN
00DE38RKN
00DE41DLN
00NJ40LSP
87LACA2GWT
04MD72MLD
04MD73MLD
02NJ60RDT
02DE54RDT
01NJ50RDT
02DE58RDT
87OH164NOP
99MN163MLD
Ct value
Cla
ss I C
lass II
gen
oty
pe
1 2 *3 4 5 6 7 8 9 I Ia IIa
WS
LB
MACCEPTED on July 1, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from