STUDIES ON BEHAVIOR. II. THE EFFECTS OF PEXTOBARBITAL, METHAMPHETAMINE AND SCOPOLAMINE ON PERFORMANCES IN PIGEONS INVOLVING DISCRIMINATIONS’2 PETER B. DEWS Department of Pharmacology, Harvard Medical School, Boston, Massachusetts Received for publication June 27, 1955 The application of the Skinner box technique to study the behavioral effects of drugs has been described in a previous communication (Dews, 1955). The work was done on pigeons; the birds were intermittently rewarded with food for pecking a transilluminated plastic disc (the key). Two different schedules of reward were used; each gave rise to a characteristic pecking performance. The two different performances showed differential sensitivity to modification by pentobarbital. The schedule of reward was thus shown to be a relevant environ- mental variable in determining the behavioral effect of pentobarbital. It is a characteristic feature of this type of behavior (“operant behavior”- Skinner, 1938) that it may be brought under “stimulus control”. If a pigeon is rewarded according to an appropriate schedule for pecking the key when certain suitable environmental stimuli3 are present, but is never rewarded when other stimuli are present, then the pecking performance comes to differ according to which of the stimuli are present. The pigeon “discriminates” between the stimuli, and behaves more or less appropriately to each. Alternatively, it may be said that the stimuli “control” the behavior of the animal. In the present work, the effect of three drugs (pentobarbital, methamphetamine, and scopolamine) on some aspects of such discriminatory behavior has been studied. It has been shown that these drugs, in doses up to those abolishing all pecking behavior under the conditions of study, did not interfere with the differential performance to a simple pair of stimuli. On the other hand, when a more complicated set of stimuli was used, it was found that pentobarbital, and methamphetamine but not scopolamine reduced the usual difference between the performance when the stimuli were appropriate for rewards and the performance when the stimuli were inappropriate for rewards. MATERIALS AND METHODS. Apparatus. The Skinner pigeon box previously described (Dews, 1955) was modified for the present work. Four different colored bulbs (G.E. C 7 1/2) were arranged so that the key could be transilluminated by any one of them, giving four possible “key colors”. In addition, a 12 watt source of white light was put in the roof of the pigeon compartment at the opposite end to the key; this light (the “house light”), when on, brightly illuminated the whole pigeon compartment. Hence 8 different combina- 1 This work was supported in part by funds received from the William F. Milton Fund of Harvard University, and in part by funds received from The Roche Anniversary Foun- dation of Hoffmann-La Roche Inc. 2 A preliminary report appears in Fed. Proc., 14: 322, 1955. ‘The term “stimulus” is used here in its usual physiological sense of an environmental influence which causes, or tends to cause, a change in a living system. 380
Transcript
STUDIES ON BEHAVIOR. II. THE EFFECTS OF PEXTOBARBITAL,
METHAMPHETAMINE AND SCOPOLAMINE ON PERFORMANCES
IN PIGEONS INVOLVING DISCRIMINATIONS’2
PETER B. DEWS
Department of Pharmacology, Harvard Medical School, Boston, Massachusetts
Received for publication June 27, 1955
The application of the Skinner box technique to study the behavioral effects
of drugs has been described in a previous communication (Dews, 1955). The
work was done on pigeons; the birds were intermittently rewarded with food
for pecking a transilluminated plastic disc (the key). Two different schedules of
reward were used; each gave rise to a characteristic pecking performance. The
two different performances showed differential sensitivity to modification by
pentobarbital. The schedule of reward was thus shown to be a relevant environ-
mental variable in determining the behavioral effect of pentobarbital.
It is a characteristic feature of this type of behavior (“operant behavior”-
Skinner, 1938) that it may be brought under “stimulus control”. If a pigeon is
rewarded according to an appropriate schedule for pecking the key when certain
suitable environmental stimuli3 are present, but is never rewarded when other
stimuli are present, then the pecking performance comes to differ according to
which of the stimuli are present. The pigeon “discriminates” between the stimuli,
and behaves more or less appropriately to each. Alternatively, it may be said
that the stimuli “control” the behavior of the animal. In the present work, the
effect of three drugs (pentobarbital, methamphetamine, and scopolamine) on
some aspects of such discriminatory behavior has been studied. It has been
shown that these drugs, in doses up to those abolishing all pecking behavior
under the conditions of study, did not interfere with the differential performance
to a simple pair of stimuli. On the other hand, when a more complicated set of
stimuli was used, it was found that pentobarbital, and methamphetamine but
not scopolamine reduced the usual difference between the performance when the
stimuli were appropriate for rewards and the performance when the stimuli were
inappropriate for rewards.
MATERIALS AND METHODS. Apparatus. The Skinner pigeon box previously described
(Dews, 1955) was modified for the present work. Four different colored bulbs (G.E. C 71/2) were arranged so that the key could be transilluminated by any one of them, giving
four possible “key colors”. In addition, a 12 watt source of white light was put in the roofof the pigeon compartment at the opposite end to the key; this light (the “house light”),when on, brightly illuminated the whole pigeon compartment. Hence 8 different combina-
1 This work was supported in part by funds received from the William F. Milton Fundof Harvard University, and in part by funds received from The Roche Anniversary Foun-
dation of Hoffmann-La Roche Inc.
2 A preliminary report appears in Fed. Proc., 14: 322, 1955.‘The term “stimulus” is used here in its usual physiological sense of an environmental
influence which causes, or tends to cause, a change in a living system.
380
The symbols S+ and S- as here defined correspond to the symbols SD and S� respec-
tively used in the psychological literature in this field. S+ and S- have been used for
typographical convenience and because they suggest their meaning more obviously.
STUDIES ON BEHAVIOR 381
DIAGRAMMATIC REPRESENTATIONOF SCHEDULES
DRUG
CONTROL AFTER DRUG
SCHEDULE SERIES 5,j, SERIES
1
2
3
4
R+ � R+� � R+� R+� R+
B �R+� W+W%� B�R+�W+�
0 30 45 75
TIME IN MINS
NO LIGHTS REWARDS
+ HOUSE LIGHTS ON �J NO REWARDS
FIG. 1. Diagram showing sequence of stimuli in the various schedules used. R, B, Y and
W indicate that the red, blue, yellow and white lights respectively were on behind the key.
tions of stimuli were possible, viz. 4 different key colors each with or without the house
light; of these, 6 combinations were actually used. In the presence of certain of these com-
binations, the pigeon was intermittently rewarded with access to food for 4-5 seconds for
pecking the key; these combinations of stimuli will he referred to as S+. In the presence
of other combinations, the pigeon was never rewarded; these combinations of stimuli will
be referred to as S-i. Responses made in the presence of S+ and S- were counted sepa-
rately on digital counters; in addition a permanent time-cumulative response record was
taken in all experiments, as previously described.
Schedules. In the presence of S+ rewards were arranged as follows. For a predetermined
period of time following a reward (or the start of the S+ period) no peck was rewarded;
when this time had elapsed, the first peck made was rewarded. The minimum interval
which necessarily intervened between successive rewards was not constant; in the present
work 15 different intervals were used, varying from 0 (two consecutive pecks rewarded)
to 2 minutes, with an arithmetic mean of 1 minute. The sequence of the intervals was
randomly determined in the first instance but then kept the same through all the experi-
ments. This scheme is an example of a “variable interval” schedule of reward in the ter-
minology of Skinner (1953). In the presence of S-, by definition, the pigeon never received
any food.
In definitive experiments, S+ and S- were presented alternately, each for 5 minutes.
The changeover from one to the other was abrupt. Three alternations, giving a total pres-
entation of S+ and S- of 15 minutes each, comprised a “series”. Each experiment started
382 PETER B. DEWS
with the bird waiting 15 minutes in the completely darkened box, followed by a control
series. The pigeon was then removed from the box, injected with the appropriate dose of
drug (or saline alone) and immediately replaced in the darkened box. The after drug seriesstarted 15 minutes later.
Four schedules, which differed according to the combinations of key color and presence
or absence of house light constituting S+ and 5-, were programmed. The arrangements
are shown diagramatically in figure 1. It will be seen that in schedules 1, 2, and 3, S+ and
5- differed in a single constant feature, viz., either key color (schedule 1) or presence orabsence of the house light (schedules 2 and 3). On the other hand, in schedule 4, not only
were there 3 different 5+ and 3 different 5- but, in the first four periods of a series, thecontingency associated with each individual stimulus was conditional; neither the key
color alone nor the presence or absence of the house light alone characterized the stimuli
as S+ or 5-; e.g. the presence of the house light was part of an S+ combination when thekey was blue; but was part of an 5- combination when the key was red. In contrast, inthe last two periods of a schedule 4 series the yellow and white key colors uniquely charac-
terized the stimuli as S+ and 5- respectively exactly as did the red and blue key colorsof schedule 1.
Procedure. Male white pigeons of between 400 and 500 grams weight were maintained
in a standardized state of food deprivation (but with constant free access to water) and
were trained to work in the Skinner box, as previously described (Dews, 1955). They werethen assigned to the various schedules just described and run once daily, most days, until
the completion of the experiments. The programming circuits were arranged so that eachpeck in the presence of 5- could be made to reset the timer which was timing the 5 minute
period of presentation of S-. Thus, when this “5- renewing” device was in operation,the 5- periods continued until the pigeon had not pecked for 5 minutes. In general, 5-renewing was used only during training of the birds; it was never in operation during de-finitive experiments. Stable performance developed much more slowly on schedule 4 (2-3
weeks) than on schedules 1, 2, 3 (4-5 days). Three pigeons were studied on schedule 1, one
each on schedules 2 and 3 and two on schedule 4.Pentobarbital sodium,’ methamphetamine hydrochloride’ and scopolamine hydro-
bromide7 were dissolved in 0.9 per cent sodium chloride solution and were injected intra-muscularly. Doses are stated in terms of these salts and give the total dose given to thebirds. The various doses of each drug were given in random order; each dose was given on
two occasions to the same pigeon. In general, where more than one drug was studied in thesame pigeon, the observations on one drug were completed before starting observations onanother drug.
Measurements of drug effects. Two indices have been used. 1) The output ratio (O.R.)
was defined as the ratio of the average rate of response in the presence of S+ in the after
drug series to the average rate of response in the presence of S+ in the control series inthe same experiment. It is thus a measure of the effect of drugs on performance in thepresence of 8+. 2) The discriminative ratio (D.R.) was defined as the complement of theratio of the average rate of response in the presence of 5- to the average rate of response
in the presence of 5+ in the same series. Hence if the pigeon does not respond in the pres-ence of 5- but responds in the presence of 5+ in the same series, the D.R. is 1; while if
the rates of response are identical in the presence of both 5+ and 5- the D.R. is zero.
RESULTS. The 1 minute variable interval schedule of reward, in operation
during all the S+ periods, gave rise to a relatively constant rate of responsethroughout the time of presentation of S+ (see figures 2 and 3). While the
average rate varied amongst the pigeons from less than 60/mm. to more than
‘Generously donated by Abbott Laboratories.‘Generously donated by Burroughs, Wellcome & Co.
Generously donated by Merck & Co.
RESPCNS�
RtSPONS(S
1000
500
3 MGPB
STUDIES ON BEHAVIOR 383
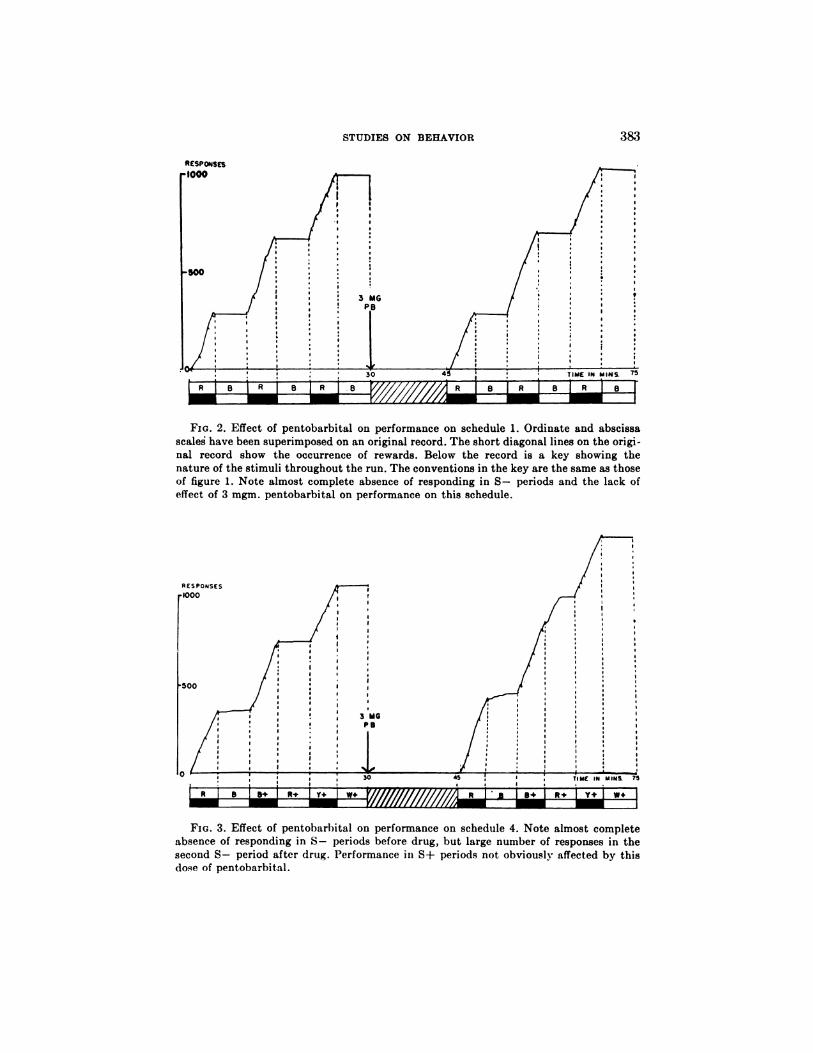
FIG. 2. Effect of pentobarbital on performance on schedule 1. Ordinate and abscissa
scaled have been superimposed on an original record. The short diagonal lines on the origi-nal record show the occurrence of rewards. Below the record is a key showing the
nature of the stimuli throughout the run. The conventions in the key are the same as thoseof figure 1. Note almost complete absence of responding in 5- periods and the lack ofeffect of 3 mgm. pentobarbital on performance on this schedule.
0
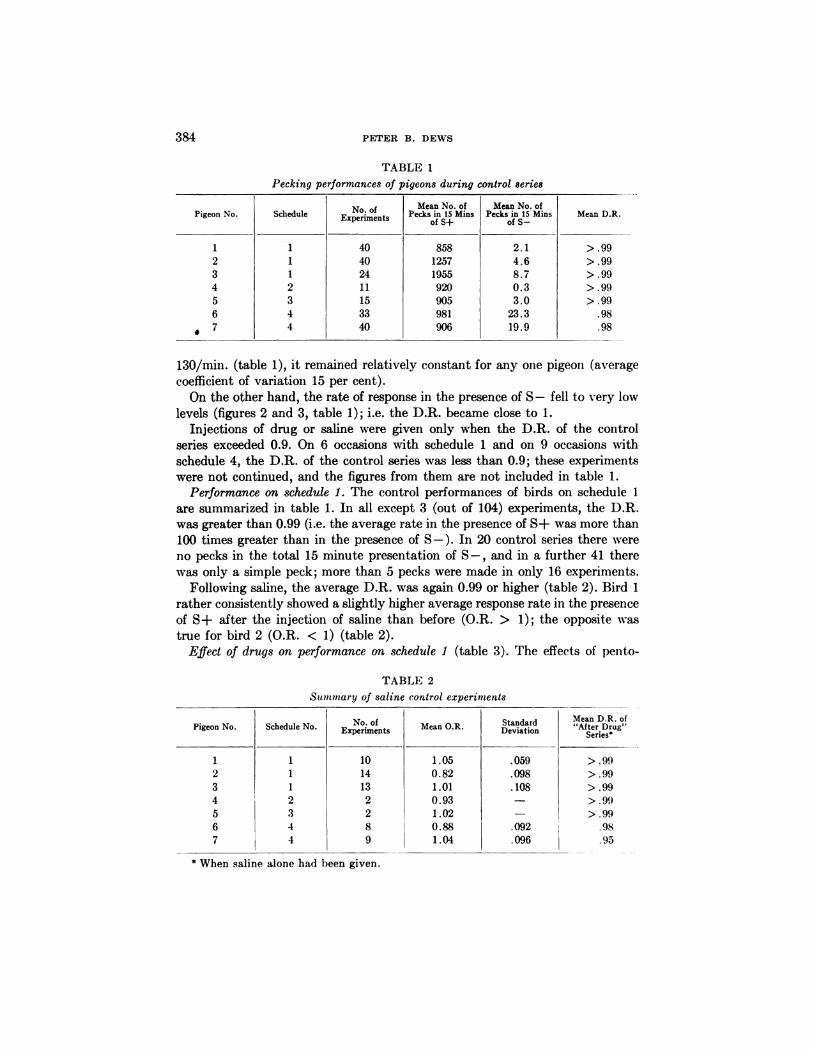
FIG. 3. Effect of pentobarbital on performance on schedule 4. Note almost complete
absence of responding in S- periods before drug, but large number of responses in thesecond S- period after drug. Performance in S+ periods not obviously affected by thisdose of pentobarbital.
384 PETER B. DEWS
* When saline alone had been given.
TABLE 1
Pecking performances of pigeons during control series
Pigeon No. Schedule No ofExperiments
Mean No. ofPecks in 15 Mins
ofS+
Mean No. ofPecks in 15 Mins
ofS-Mean DR.
1
2
3
45
67
1
1
12
3
4
4
40
40
2411
15
3340
858
1257
1955
920
905
981906
2.1
4.6
8.70.33.0
23.319.9
>99
>.99
>99>99
>99.98.98
130/mm. (table 1), it remained relatively constant for any one pigeon (average
coefficient of variation 15 per cent).
On the other hand, the rate of response in the presence of S - fell to very low
levels (figures 2 and 3, table 1); i.e. the D.R. became close to 1.
Injections of drug or saline were given only when the D.R. of the control
series exceeded 0.9. On 6 occasions with schedule 1 and on 9 occasions withschedule 4, the D.R. of the control series was less than 0.9; these experiments
were not continued, and the figures from them are not included in table 1.
Performance on schedule 1. The control performances of birds on schedule 1
are summarized in table 1. In all except 3 (out of 104) experiments, the D.R.
was greater than 0.99 (i.e. the average rate in the presence of S+ was more than
100 times greater than in the presence of S -). In 20 control series there were
no pecks in the total 15 minute presentation of S -‘ and in a further 41 there
was only a simple peck; more than 5 pecks were made in only 16 experiments.
Following saline, the average D.R. was again 0.99 or higher (table 2). Bird 1
rather consistently showed a slightly higher average response rate in the presence
of S+ after the injection of saline than before (O.R. > 1); the opposite was
true for bird 2 (O.R. < 1) (table 2).
Effect of drugs on performance on schedule 1 (table 3). The effects of pento-
TABLE 2
Summary of saline control experiments
Pigeon No. Schedule No. No. ofExperiments Mean O.R. Standard
Deviation
Mean D.R. of
“After Drug”Series’
1 1 10 1.05 .059 > .99
2 1 14 0.82 .098 >99
3 1 13 1.01 .108 >99
4 2 2 0.93 - >99
5 3 2 1.02 - >99
6 4 8 0.88 .092 .98
7 4 9 1.04 .096 .95
‘SE. = .063. ‘SE = .025.
STUDIES ON BEHAVIOR 385
TABLE 3
Effect of pentobarbital, methamphetamine and scopolamine on performance on schedule I