

STUDIES ON BEHAVIOR. II. THE EFFECTS OF PEXTOBARBITAL,
METHAMPHETAMINE AND SCOPOLAMINE ON PERFORMANCES
IN PIGEONS INVOLVING DISCRIMINATIONS’2
PETER B. DEWS
Department of Pharmacology, Harvard Medical School, Boston, Massachusetts
Received for publication June 27, 1955
The application of the Skinner box technique to study the behavioral effects
of drugs has been described in a previous communication (Dews, 1955). The
work was done on pigeons; the birds were intermittently rewarded with food
for pecking a transilluminated plastic disc (the key). Two different schedules of
reward were used; each gave rise to a characteristic pecking performance. The
two different performances showed differential sensitivity to modification by
pentobarbital. The schedule of reward was thus shown to be a relevant environ-
mental variable in determining the behavioral effect of pentobarbital.
It is a characteristic feature of this type of behavior (“operant behavior”-
Skinner, 1938) that it may be brought under “stimulus control”. If a pigeon is
rewarded according to an appropriate schedule for pecking the key when certain
suitable environmental stimuli3 are present, but is never rewarded when other
stimuli are present, then the pecking performance comes to differ according to
which of the stimuli are present. The pigeon “discriminates” between the stimuli,
and behaves more or less appropriately to each. Alternatively, it may be said
that the stimuli “control” the behavior of the animal. In the present work, the
effect of three drugs (pentobarbital, methamphetamine, and scopolamine) on
some aspects of such discriminatory behavior has been studied. It has been
shown that these drugs, in doses up to those abolishing all pecking behavior
under the conditions of study, did not interfere with the differential performance
to a simple pair of stimuli. On the other hand, when a more complicated set of
stimuli was used, it was found that pentobarbital, and methamphetamine but
not scopolamine reduced the usual difference between the performance when the
stimuli were appropriate for rewards and the performance when the stimuli were
inappropriate for rewards.
MATERIALS AND METHODS. Apparatus. The Skinner pigeon box previously described
(Dews, 1955) was modified for the present work. Four different colored bulbs (G.E. C 71/2) were arranged so that the key could be transilluminated by any one of them, giving
four possible “key colors”. In addition, a 12 watt source of white light was put in the roofof the pigeon compartment at the opposite end to the key; this light (the “house light”),when on, brightly illuminated the whole pigeon compartment. Hence 8 different combina-
1 This work was supported in part by funds received from the William F. Milton Fundof Harvard University, and in part by funds received from The Roche Anniversary Foun-
dation of Hoffmann-La Roche Inc.
2 A preliminary report appears in Fed. Proc., 14: 322, 1955.‘The term “stimulus” is used here in its usual physiological sense of an environmental
influence which causes, or tends to cause, a change in a living system.
380

The symbols S+ and S- as here defined correspond to the symbols SD and S� respec-
tively used in the psychological literature in this field. S+ and S- have been used for
typographical convenience and because they suggest their meaning more obviously.
STUDIES ON BEHAVIOR 381
DIAGRAMMATIC REPRESENTATIONOF SCHEDULES
DRUG
CONTROL AFTER DRUG
SCHEDULE SERIES 5,j, SERIES
1
2
3
4
R+ � R+� � R+� R+� R+
B �R+� W+W%� B�R+�W+�
0 30 45 75
TIME IN MINS
NO LIGHTS REWARDS
+ HOUSE LIGHTS ON �J NO REWARDS
FIG. 1. Diagram showing sequence of stimuli in the various schedules used. R, B, Y and
W indicate that the red, blue, yellow and white lights respectively were on behind the key.
tions of stimuli were possible, viz. 4 different key colors each with or without the house
light; of these, 6 combinations were actually used. In the presence of certain of these com-
binations, the pigeon was intermittently rewarded with access to food for 4-5 seconds for
pecking the key; these combinations of stimuli will he referred to as S+. In the presence
of other combinations, the pigeon was never rewarded; these combinations of stimuli will
be referred to as S-i. Responses made in the presence of S+ and S- were counted sepa-
rately on digital counters; in addition a permanent time-cumulative response record was
taken in all experiments, as previously described.
Schedules. In the presence of S+ rewards were arranged as follows. For a predetermined
period of time following a reward (or the start of the S+ period) no peck was rewarded;
when this time had elapsed, the first peck made was rewarded. The minimum interval
which necessarily intervened between successive rewards was not constant; in the present
work 15 different intervals were used, varying from 0 (two consecutive pecks rewarded)
to 2 minutes, with an arithmetic mean of 1 minute. The sequence of the intervals was
randomly determined in the first instance but then kept the same through all the experi-
ments. This scheme is an example of a “variable interval” schedule of reward in the ter-
minology of Skinner (1953). In the presence of S-, by definition, the pigeon never received
any food.
In definitive experiments, S+ and S- were presented alternately, each for 5 minutes.
The changeover from one to the other was abrupt. Three alternations, giving a total pres-
entation of S+ and S- of 15 minutes each, comprised a “series”. Each experiment started

382 PETER B. DEWS
with the bird waiting 15 minutes in the completely darkened box, followed by a control
series. The pigeon was then removed from the box, injected with the appropriate dose of
drug (or saline alone) and immediately replaced in the darkened box. The after drug seriesstarted 15 minutes later.
Four schedules, which differed according to the combinations of key color and presence
or absence of house light constituting S+ and 5-, were programmed. The arrangements
are shown diagramatically in figure 1. It will be seen that in schedules 1, 2, and 3, S+ and
5- differed in a single constant feature, viz., either key color (schedule 1) or presence orabsence of the house light (schedules 2 and 3). On the other hand, in schedule 4, not only
were there 3 different 5+ and 3 different 5- but, in the first four periods of a series, thecontingency associated with each individual stimulus was conditional; neither the key
color alone nor the presence or absence of the house light alone characterized the stimuli
as S+ or 5-; e.g. the presence of the house light was part of an S+ combination when thekey was blue; but was part of an 5- combination when the key was red. In contrast, inthe last two periods of a schedule 4 series the yellow and white key colors uniquely charac-
terized the stimuli as S+ and 5- respectively exactly as did the red and blue key colorsof schedule 1.
Procedure. Male white pigeons of between 400 and 500 grams weight were maintained
in a standardized state of food deprivation (but with constant free access to water) and
were trained to work in the Skinner box, as previously described (Dews, 1955). They werethen assigned to the various schedules just described and run once daily, most days, until
the completion of the experiments. The programming circuits were arranged so that eachpeck in the presence of 5- could be made to reset the timer which was timing the 5 minute
period of presentation of S-. Thus, when this “5- renewing” device was in operation,the 5- periods continued until the pigeon had not pecked for 5 minutes. In general, 5-renewing was used only during training of the birds; it was never in operation during de-finitive experiments. Stable performance developed much more slowly on schedule 4 (2-3
weeks) than on schedules 1, 2, 3 (4-5 days). Three pigeons were studied on schedule 1, one
each on schedules 2 and 3 and two on schedule 4.Pentobarbital sodium,’ methamphetamine hydrochloride’ and scopolamine hydro-
bromide7 were dissolved in 0.9 per cent sodium chloride solution and were injected intra-muscularly. Doses are stated in terms of these salts and give the total dose given to thebirds. The various doses of each drug were given in random order; each dose was given on
two occasions to the same pigeon. In general, where more than one drug was studied in thesame pigeon, the observations on one drug were completed before starting observations onanother drug.
Measurements of drug effects. Two indices have been used. 1) The output ratio (O.R.)
was defined as the ratio of the average rate of response in the presence of S+ in the after
drug series to the average rate of response in the presence of S+ in the control series inthe same experiment. It is thus a measure of the effect of drugs on performance in thepresence of 8+. 2) The discriminative ratio (D.R.) was defined as the complement of theratio of the average rate of response in the presence of 5- to the average rate of response
in the presence of 5+ in the same series. Hence if the pigeon does not respond in the pres-ence of 5- but responds in the presence of 5+ in the same series, the D.R. is 1; while if
the rates of response are identical in the presence of both 5+ and 5- the D.R. is zero.
RESULTS. The 1 minute variable interval schedule of reward, in operation
during all the S+ periods, gave rise to a relatively constant rate of responsethroughout the time of presentation of S+ (see figures 2 and 3). While the
average rate varied amongst the pigeons from less than 60/mm. to more than
‘Generously donated by Abbott Laboratories.‘Generously donated by Burroughs, Wellcome & Co.
Generously donated by Merck & Co.
RESPCNS�
RtSPONS(S
1000
500
3 MGPB
STUDIES ON BEHAVIOR 383
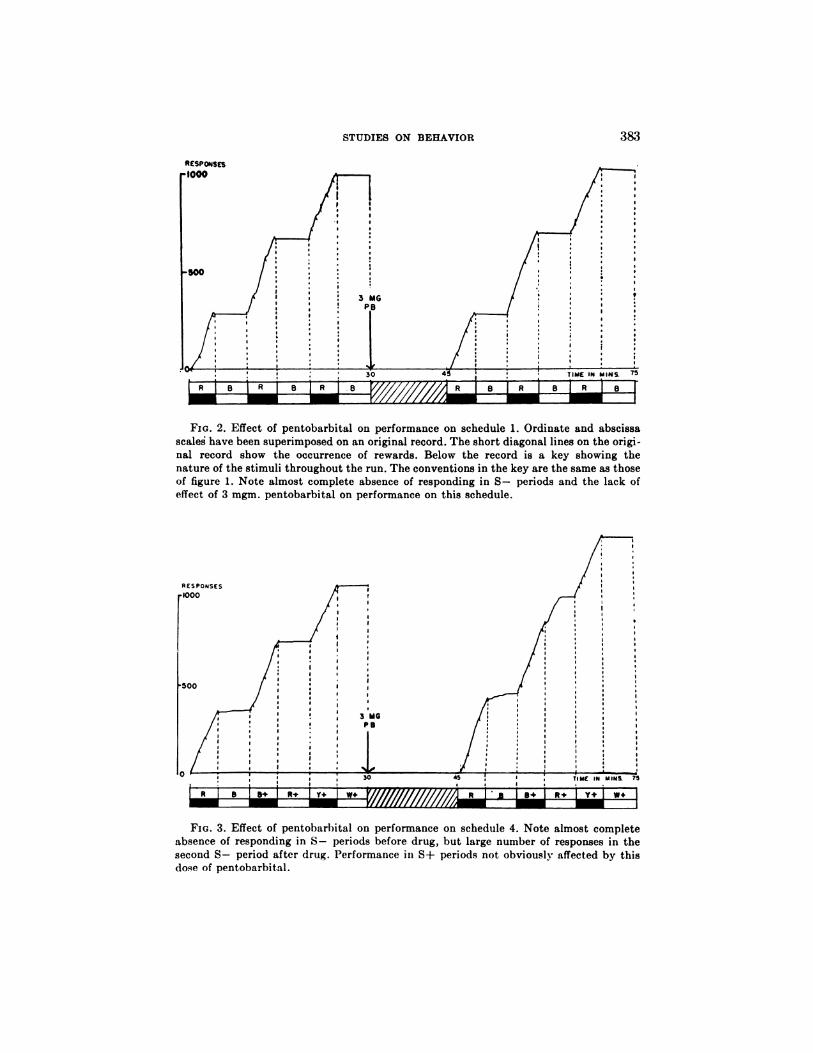
FIG. 2. Effect of pentobarbital on performance on schedule 1. Ordinate and abscissa
scaled have been superimposed on an original record. The short diagonal lines on the origi-nal record show the occurrence of rewards. Below the record is a key showing the
nature of the stimuli throughout the run. The conventions in the key are the same as thoseof figure 1. Note almost complete absence of responding in 5- periods and the lack ofeffect of 3 mgm. pentobarbital on performance on this schedule.
0
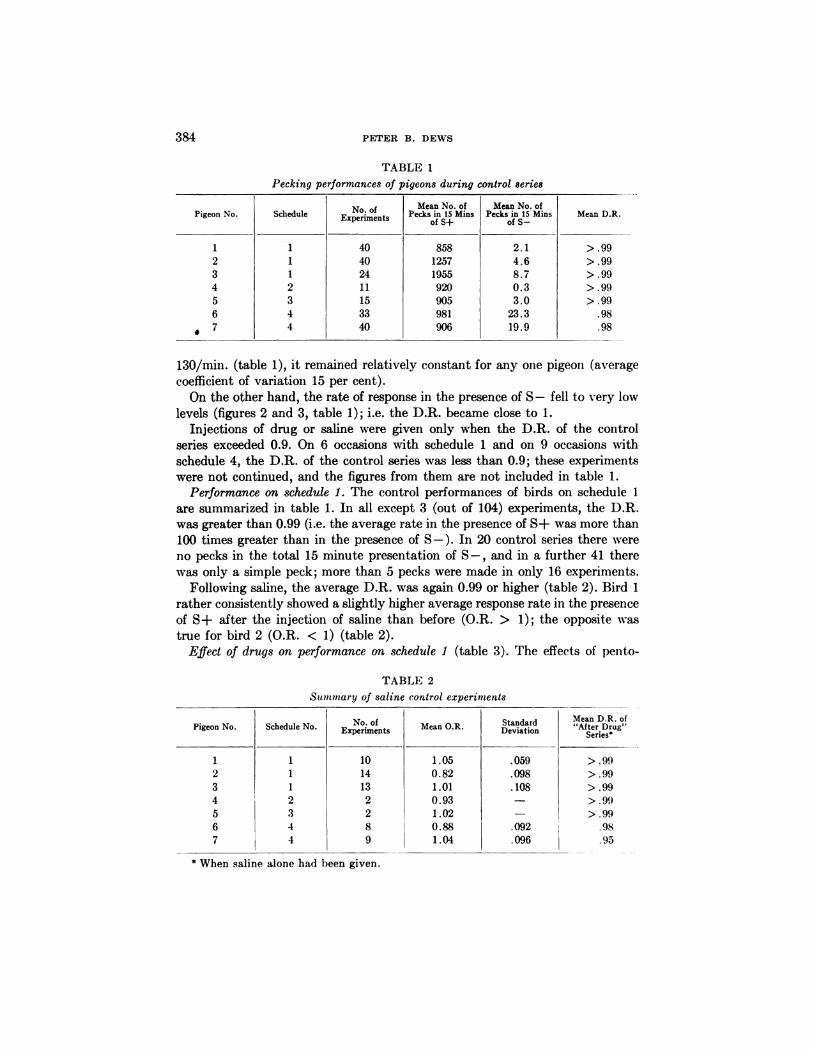
FIG. 3. Effect of pentobarbital on performance on schedule 4. Note almost complete
absence of responding in S- periods before drug, but large number of responses in thesecond S- period after drug. Performance in S+ periods not obviously affected by thisdose of pentobarbital.
384 PETER B. DEWS
* When saline alone had been given.
TABLE 1
Pecking performances of pigeons during control series
Pigeon No. Schedule No ofExperiments
Mean No. ofPecks in 15 Mins
ofS+
Mean No. ofPecks in 15 Mins
ofS-Mean DR.
1
2
3
45
67
1
1
12
3
4
4
40
40
2411
15
3340
858
1257
1955
920
905
981906
2.1
4.6
8.70.33.0
23.319.9
>99
>.99
>99>99
>99.98.98
130/mm. (table 1), it remained relatively constant for any one pigeon (average
coefficient of variation 15 per cent).
On the other hand, the rate of response in the presence of S - fell to very low
levels (figures 2 and 3, table 1); i.e. the D.R. became close to 1.
Injections of drug or saline were given only when the D.R. of the control
series exceeded 0.9. On 6 occasions with schedule 1 and on 9 occasions withschedule 4, the D.R. of the control series was less than 0.9; these experiments
were not continued, and the figures from them are not included in table 1.
Performance on schedule 1. The control performances of birds on schedule 1
are summarized in table 1. In all except 3 (out of 104) experiments, the D.R.
was greater than 0.99 (i.e. the average rate in the presence of S+ was more than
100 times greater than in the presence of S -). In 20 control series there were
no pecks in the total 15 minute presentation of S -‘ and in a further 41 there
was only a simple peck; more than 5 pecks were made in only 16 experiments.
Following saline, the average D.R. was again 0.99 or higher (table 2). Bird 1
rather consistently showed a slightly higher average response rate in the presence
of S+ after the injection of saline than before (O.R. > 1); the opposite was
true for bird 2 (O.R. < 1) (table 2).
Effect of drugs on performance on schedule 1 (table 3). The effects of pento-
TABLE 2
Summary of saline control experiments
Pigeon No. Schedule No. No. ofExperiments Mean O.R. Standard
Deviation
Mean D.R. of
“After Drug”Series’
1 1 10 1.05 .059 > .99
2 1 14 0.82 .098 >99
3 1 13 1.01 .108 >99
4 2 2 0.93 - >99
5 3 2 1.02 - >99
6 4 8 0.88 .092 .98
7 4 9 1.04 .096 .95

‘SE. = .063. ‘SE = .025.
STUDIES ON BEHAVIOR 385
TABLE 3
Effect of pentobarbital, methamphetamine and scopolamine on performance on schedule I
Pento-barbitalDosage(mgm.)
Mean’OR.
Mean’D.R.
Metham-phetamine
Dosage(mgm.)
Mean’OR.
Mean’DR.
Sco�olamineosage
(m�.)
Mean’OR.
Mean’DR.
04 0.90’ >99 0� 0.926 >.99 0� 1.05� >991.0 0.91 >.99 0.1 0.98 >.99 0.001 1.02 >.993.0 1.00 >99 0.3 0.78 >99 0.003 0.87 >.99
4.0 0.22 >99 1.0 0.27 >99 0.01 0.76 >995.2 0.01 >99 1.7
3.0
0.22
0.01>.99>99
0.0170.03
0.64
0.14>99>99
0.0520.1
0.080.02
>99>99
‘Mean of birds 2 and 3. ‘Mean of birds 1 and 2. ‘Bird 1 only. 4Saline alone.
‘SE. = .074. ‘SE. = .067. 7S.E. = .059.
barbital were studied in birds 2 and 3 (figure 2). Following doses of 4.0 mgm.
or 5.2 mgm., the average rate of response in the presence of S+ was greatly
reduced; but at no dose level did the average D.R. fall below 0.99.
Similar results were obtained with methamphetamine on birds 1 and 2. At
no dose level did the birds show any increase in the number of responses in the
presence of S -, although doses of 0.3 mgm. or more caused pronounced be-
havioral effects, as shown by the fall in the average rate of response in the
presence of S+.
Scopolamine was studied in bird 1 only. Again, even doses sufficient to affect
performance grossly in the presence of S + caused no breakdown of discrimina-
tive behavior; the D.R. never fell even to 0.99.
Performance on schedule 4. The average D.R. in the control series of birds 6
and 7 working on schedule 4 was 0.98 (table 1, figure 3) indicating an approxi-
mately 50 fold difference in rates of response in the presence of S+ and S -.
Both the O.R. and D.R. were well maintained in experiments in which saline
alone was injected (table 2).
TABLE 4
Effect of pentobarbital, methamphetamine and scopolamine on performance on schedule 4 ofbirds 6 and 7
Pentobarbital Methamphetamine Scopolamine
Dose MeanOR.
MeanD.R.
Dose MeanOR.
MeanD.R.
D�eI
Mean MeanOR. . DR.
0
1
3
5.2
0.95’
0.97
0.97
0.36
0.95’0.93
0.82
0.06
0
0.1
0.3
0.52
1.0
0.951.00
1.07
1.10
0.71
0.950.960.56
0.89
0.76
00.0030.01
0.03
0.1
0.95 0.950.86 0.97
0.50 0.98
0.30 >0.99
0.02 >0.99

386 PETER B. DEWS
* S.E. = .037.
Effect of drugs on performance on schedule 4 (table 4). The effects of all three
drugs were studied in both pigeon 6 and pigeon 7.
Following 3 mgm. of pentobarbital consistently more responses were made
in the presence of S - than following saline alone, although the average rate of
response in the presence of S+ was not much affected. This is illustrated in
figure 3. It will be noted that in this particular experiment, most of the responses
in S - occurred in the first part of the second S - period after the drug. The re-
sponding in S - often took the form of a period of steady responding, as here.
In general, such periods of responding were rare and short in the third S -
period (see below), but there was no obvious tendency for them to occur in
any particular part of the first or second S - periods. In spite of this variability
in the time of occurrence of the pecks during S -, the total number made through-
out the presentation of the various S - was reasonably constant. Following
5.2 mgm. of pentobarbital the average rate of response in the presence of S -
was almost as high as that in the presence of S+.
Methamphetamine in appropriate doses also caused a fall in D.R. to values
below those seen following saline, even when the rate of response in the presence
of S+ was well sustained or even increased. The extent of fall of D.R. was
variable, and over a considerable range bore no obvious relation to dosage.
However, since the D.R. following saline showed great consistency (coefficient
of variation 2.5 per cent) there can be no reasonable doubt that the lowering of
the D.R. by methamphetamine was a real phenomenon.
Injection of scopolamine led to no fall in the D.R.; on the contrary, discrim-
inative performance even improved following doses causing a fall in response
rate in the presence of S+.
As explained earlier (see Schedules) a schedule 4 series may be divided into
2 parts: an initial 4 periods in which the implication of the stimuli was condi-
tional; and a final 2 periods in which the implication of the key color was un-
ambiguous, as in schedule 1. The D.R. of each of these parts has been calcu-
lated separately for the after drug series following saline, pentobarbital and
methamphetamine. Following saline alone, the D.R. for the conditional part
was slightly, though consistently, lower than for the simple part (table 5).
Following the various doses of pentobarbital and methamphetamine studied
TABLE 5
Differential effect of pentobarbital and methamphetamine on performance o�f birds 6 and 7 in
“conditional” and “simple” discrimination situations of schedule 4
Pentobarbital Methamphetamine
Dose DR DR(mgm.) -� (conditional) -� (simple)
Dose(mgm.)
DR DR(conditional) (simple) -
0 095* >.99 0 095* >.991 0.79 >99 0.3 0.55 0.76
3 0.20 >.99 0.52 0.88 0.98
5.2 <0 0.85 1.0 0.57� 0.85

STUI)IES ON BEhAVIOR 387
TABLE 6
Effect of pentobarbital and methasnphetamine on performance on schedules 2 and S of
birds S and 4
Pentobarbital Methamphetamine
Dose OR DR � Dose OR -� DR
0 1.04 >0.99 0 1.04 >0.99
3 0.96 >0.99 0.3 0.81 >0.99
5.2 0.29 0.99 0.52 0.56 >0.99
the D.R. for the conditional part was always less than that for the simple part;
the difference was usually large as can be seen from the averages shown in table
5. It should be noted that the D.R. for the simple part did, on some occasions,
fall to values lower than any seen with schedule 1.
Performance on schedule 2 and 3 and effect of drugs. Performance on schedules
2 and 3 did not differ in any significant way from performance on schedule 1
(tables 1 and 2). Doses of pentobarbital and methamphetamine which caused a
fall in the D.R. of birds working on schedule 4 had no effect on the D.R. of birds
working on schedules 2 and 3 (table 6).
DISCUSSION. It has been shown that the discriminative performance of the
pigeons based on either a difference in key color or the presence or absence of
the house light was not disrupted by pentobarbital, methamphetamine or
scopolamine in doses up to those causing complete cessation of pecking. On the
other hand, pentobarbital and methamphetamine, but not scopolamine, inter-
fered with the discriminative performance on schedule 4, a schedule which in-
volved six different stimulus combinations and a conditional situation. These
findings are clearly in agreement with the widely held view that “more complex”
performance is more susceptible to drug effects than “simple” performance.
However, the terms “complex” and “simple” are not easy to define in this
context, and it would appear preferable to describe the differential sensitivity
to drugs in terms of the operational difference between the schedules.
The analysis summarized iii table 5 makes it appear that it is the conditional
feature of schedule 4 rather than simply the increased number of stimulus com-
binations involved, which determines this difference. However, the D.R. of the
simple part of schedule 4 did fall on occasions, indicating that it was not al-
together comparable to the completely simple schedules 1, 2, and 3. Moreover,
the constant sequence of stimuli means that the effect of the conditional and
simple parts of schedule 4 is confounded with time after drug. Further experi-
mental analysis is required.
Since the S+ and S- periods alternated, a change in stimuli always implied
a change in contingencies. Hence the changes of stimuli, rather than the nature
of the stimuli themselves may have played a part in the sudden changes in
pecking rate shown in figures 2 and 3. It should be emphasized that schedule
4 did not differ from the other schedules in this regard.
No attempt was made to identify the specific feature or features of the stimuli

388 PETER B. DEWS
on which the discriminations were based. The differences between the stimuli
were made as gross as convenient ; but it was not relevant to the present work
to determine whether, for example, the discrimination of schedule 1 was based
Oil the qualities of redness and blueness of the stimuli. There was, in fact, a
distinct difference in brightness between the red and blue lights, and no attempt
was made to balance them in this regard.
The absence of effect of scopolamine on the discriminations of schedule 4 has
no obvious explanation. The pigeons ate freely when offered grain following doses
of scopolamine abolishing pecking behavior in the Skinner box. Even a dose as
large as 10 mgm. scopolamine (i.e. 100 times larger than the dose almost abolish-
ing pecking behavior) caused no grossly observable change in behavior; 15 mgm.
caused vomiting; 45 mgm. still caused only vomiting with no other unequivocal
signs. From the point of view of the present work, the effects of scopolamine are
of interest in showing that the effects of pentobarbital and methamphetamine
on the I).R. were not entirely nonspecific.
At the appropriate dose level, both methamphetamine and pentobarbital
caused a slight increase in the O.R. The very modest “stimulating” effect of
methamphetamine seen in the present experiments is believed to be due to the
nature of the schedule of reward. The rate of pecking is of the order of 1/sec.
while rewards are set up only 1/mm. on the average. A doubling in rate would
mean that the rewards would be received at most only 0.5 sees. earlier, while
120 pecks/reward instead of 60 pecks/reward would be made. This decrease in
“efficiency” with increase in rate probably tends to minimize any increase in
rate above its already “inefficiently” high value.
The “S - renewing” procedure was used during training to prevent the
change from S - to S + (a rewarding event) from occurring shortly after a peck
made in the presence of S - ; such chance coincidences tend to maintain pecking
in the presence of S -‘ increasing the probability of further coincidences of peck
and change to S+ and thus starting a vicious circle.
SUMMARY
Pigeons were rewarded with occasional access to food for 4-S seconds for peck-
ing a translucent plastic disc when certain environmental stimuli were present,
but were never rewarded when other environmental stimuli were present. They
(Same to peck at a steady rate of the order of 60/mill. when the stimuli appro-
priate to rewards were present but at a rate of only about 1/mill. or less when
the stimuli appropriate to no rewards were present.
When the stimulus appropriate to rewards was a red light behind the traiis-
lucent disc and the stimulus appropriate to no rewards was a blue light, then
this differential performance was not disrupted by pentobarbital, methamphet-
amine or scopolamine, although as the dosage of any of the drugs was increased,
pecking in the presence of the red light was progressively reduced.
When the stimuli appropriate to rewards were a red light, a blue light plus a
differently located white light (the house light) or a yellow light plus the house
light, and the stimuli appropriate to no rewards were a blue light alone, or a red

STUDIES ON BEHAVIOR 38�)
or white light plus the house light, then the differential performance between the
sets of stimuli was reduced by pentobarbital and methamphetamine, but not
by scopolamine.
Preliminary analysis of the differences between the schedules determining
this differential sensitivity to drugs is presented.
ACKNOWLEDGMENT. The author wishes to thank Mr. Briah Connor for con-
scientious assistance throughout these experiments.
REFERENCES
DEws, P. B.: Tiiis JOURNAL, 113: 393, 1955.SKINNER, B. F.: The Behavior of Organisms, New York, App1eton-Ccntur�, p. 19, 1938.
SKINNER, B. F.: Science and human behavior. New York, Macmillan, P. 102, 1953.

MERCURIALS AND RENAL SH CONCENTRATIONS 399
TABLE 6
Effect of mersalyl on sulfhydryl concentrations in the cytoplasm of rat kidney cells (in vii ro)*
Incubation Time in0.02 .41 Mersalyl
Proximal Tubules(Terminal Portions)
Proximal Tubules(Convoluted Portions)
Distal ConvolutedTubules
EPer cent
ofcontrol
EPer cent
ofcontrol
EPer cent
ofcontrol
Saline (60 minutes)..1 minute
2 minutes
30 minutes60 minutes
.419 ± .012263 ± .009
246 ± .008
228 ± .008
203 ± .006
100
63
59
5548
.606 ± .009
.478 ± .009
.433 ± .008
.351 ± .007
.364 ± .010
100
79
71
58
60
.538 ± .008
.375 ± .011
.339 ± .011
.290 ± .005
.271 ± .007
100
70
6354
50
5 Each value represents the mean of 15 measurements in 15 different cells.
mechanism. It is possible that the pattern of sulfhydryl depression after mersalyl
administration may reflect the presence of two distinct reabsorptive mechanisms.
Although the collecting ducts generally have been considered to be passive
tubes which convey the formed urine to the renal pelvis, certain indications
that they play a role in reabsorption have appeared in the literature from time
to time. Schmidt-Nielson and Schmidt-Nielson (1952) cited the relatively
important role of collecting ducts of the kangaroo rat, Dipodomys, in water
reabsorption. Darmady (1954) correlated a resistance to Pitressin in patient.s
with diabetes insipidus and cases of death preceded by massive water diuresis
with marked atrophy of collecting duct epithelium. Wirz et al. (1951), on the
basis of freezing point depression studies, concluded that an osmotic drain of
water from collecting ducts to the hypertonic surroundings occurs in the rat
and hamster. Our data show an effect of mercurials on cells of the collecting
ducts and it may well be that mercurial diuresis is augmented by some inter-
ference in reabsorptive capacities of collecting ducts.
Fawaz and Fawaz (1954) found that the optimal diuretic dose in rats was
approximately 10-15 mgm. mersalyl/kgm. when given intravenously, and that
the height of water diuresis was reached in 2-4 hours. These results of Fawaz
and Fawaz have been confirmed in our laboratories. In a large series of experi-
ments, it was found that the maximum diuretic period following the intra-
muscular administration of 5-15 mgm. mersalyl/kgm. occurred at 2-5 hours.
Thus the pharmacological and biochemical events relating to mercurial diuresis
seem to occur simultaneously.
The effects of cysteine and BAL reported here are consistent with the pharma-
cological data obtained when they are used in conjunction with mercurials.
Earlier work by Handley (1949) and Handley and Lavik (1950) indicated
that mercurials inhibit the enzyme, succinic dehydrogenase. They reported
inhibition in kidney homogenates of rats treated with mercurials but Fawaz
and Fawaz (1951) were not able to confirm these reports.
While this work was in progress several reports appeared in the literature
suggesting that mercurials produce diuresis by inactivating succinic dehy-
drogenase. Mustakallio and Telkk#{228} (1953) injected rats with a mercurial diuretic







