Plant Physiol. (1989) 89, 743-752 0032-0889/89/89/0743/1 0/$01 .00/0 Received for publication August 18, 1988 and in revised form October 7, 1988 An Auxin-Regulated Gene of Arabidopsis thaliana Encodes a DNA-Binding Protein1 Thierry Alliotte, Christine Tire, Gilbert Engler, Johan Peleman, Allan Caplan, Marc Van Montagu*, and Dirk lnze Laboratorium voor Genetica, Rijksuniversiteit Gent, B-9000 Gent, Belgium ABSTRACT We have isolated a single-copy gene from the plantArabidopsis thaliana, called dbp, which encodes a lysine-rich, DNA-binding protein. The Dbp protein has a molecular weight and a composi- tion resembling histone HI. When the dbp gene was expressed in bacteria, the protein product bound DNA nonspecifically. The dbp gene is expressed constitutively in all parts of the plant but is induced five times above this basal level in apical zones. In vitro hormone-depletion experiments showed that the expression in the shoot apex could be induced by exogenous auxin. In situ hybridizations in the root apex indicated that the expression of dbp is enhanced in the region of cell division. Auxin is, arguably, the most intensively studied of the plant growth hormones. The natural and omnipresent auxin IAA plays important roles in the regulation of cell division, cell extension (or elongation), and cell differentiation (for a re- view, see Ref. 39). However, despite the vast literature on the physiology of auxins, little information is available on their mode of action at the molecular level, and it is only in the last few years that a number of auxin-regulated genes have been isolated (for reviews, see Refs. 19, 45). Auxins have been shown to regulate the transcription of an extensive set of genes. On excised soybean and pea hypocotyls, auxin stimulates immediate cell elongation, accompanied by the immediate induction (within 5-30 min) of several mRNA species (19, 45). Although the effect of auxin on cell division has been analyzed less intensively than its effect on cell elongation, the availability of tobacco cell suspension cultures dependent only on auxin for continuous growth has provided a convenient experimental system to investigate the role of this hormone in cell division processes. After a starvation period in hormone-free medium, such cell suspension cultures react quickly to auxin addition with renewed cell division in concert with the induction of specific mRNAs (47). In this system, cell division-associated RNAs start to accumulate within 30 min to 2 h after auxin addition. 'This work was supported by grants from the A.S.L.K. Kanker- fonds, the Fonds voor Geneeskundig Wetenschappelijk Onderzoek (FGWO 3.0001.82), and the Services of the Prime Minister (OOA 12.0561.84 and UIAP 120C0 187). T. A. is indebted to L'Air Liquide S.A. for financial support, C. T. is a Research Assistant, and D. I. a Senior Research Assistant of the National Fund for Scientific Re- search (Belgium). Besides these extensively studied primary effects of auxins on gene expression, a number of more long-term responses have been characterized at the molecular level. For example, it is well documented that 18S and 25S ribosomal RNAs accumulate after several hours of auxin treatment, due to a selective enhancement of RNA polymerase I activity (for a review, see Ref. 19). Several polypeptides that respond to long-term auxin exposure have been identified. A 2,4-D treat- ment of intact pea seedlings induces cellulase mRNAs after a lag period of approximately 24 h (49). In another study (15, 16), a 24-h auxin treatment of soybean hypocotyls was shown to cause an approximate eightfold increase in the abundance of ribosomal protein mRNAs. The similar induction kinetics for RNA polymerase I activity and ribosomal protein mRNA accumulation suggests that their synthesis might be regulated coordinately. In the present study, we have identified a single-copy gene, called dbp, with a high basal level of expression in all parts of mature Arabidopsis thaliana plants and elevated expression in the root and shoot apex. The expression of dbp in isolated peduncle segments can be enhanced by a 4-h exposure to exogenous auxin and thus is a member of the set of long-term auxin-responsive genes (19). Moreover, the Dbp polypeptide is lysine-rich and shares several biochemical properties with histone H1. Although it is biochemically and immunologi- cally distinct from histone H1, we show here that the Dbp polypeptide binds double-stranded DNA apparently in a sim- ilar fashion. MATERIALS AND METHODS Plant Material and Bacterial Strains Four- to 6-week-old Arabidopsis thaliana var Columbia, grown under greenhouse conditions, were used as plant ma- terial (see also Ref. 41). Except when specifically stated, all molecular clonings were performed with the Escherichia coli strain MC1061 (5). The X phage library was amplified using the E. coli strain NM538 (14). Experiments of bacterial expression were carried out with the strain SG4044 lon- (18). Construction and Screening of an A. thaliana cDNA Library The procedure for cDNA synthesis and cloning has been described elsewhere in detail (12). The cDNA was synthesized 743
Received for publication August 18, 1988and in revised form October 7, 1988
An Auxin-Regulated Gene of Arabidopsis thaliana Encodesa DNA-Binding Protein1
Thierry Alliotte, Christine Tire, Gilbert Engler, Johan Peleman, Allan Caplan,Marc Van Montagu*, and Dirk lnze
Laboratorium voor Genetica, Rijksuniversiteit Gent, B-9000 Gent, Belgium
ABSTRACT
We have isolated a single-copy gene from the plantArabidopsisthaliana, called dbp, which encodes a lysine-rich, DNA-bindingprotein. The Dbp protein has a molecular weight and a composi-tion resembling histone HI. When the dbp gene was expressedin bacteria, the protein product bound DNA nonspecifically. Thedbp gene is expressed constitutively in all parts of the plant butis induced five times above this basal level in apical zones. Invitro hormone-depletion experiments showed that the expressionin the shoot apex could be induced by exogenous auxin. In situhybridizations in the root apex indicated that the expression ofdbp is enhanced in the region of cell division.
Auxin is, arguably, the most intensively studied ofthe plantgrowth hormones. The natural and omnipresent auxin IAAplays important roles in the regulation of cell division, cellextension (or elongation), and cell differentiation (for a re-view, see Ref. 39). However, despite the vast literature on thephysiology of auxins, little information is available on theirmode of action at the molecular level, and it is only in thelast few years that a number of auxin-regulated genes havebeen isolated (for reviews, see Refs. 19, 45).Auxins have been shown to regulate the transcription of an
extensive set ofgenes. On excised soybean and pea hypocotyls,auxin stimulates immediate cell elongation, accompanied bythe immediate induction (within 5-30 min) of several mRNAspecies (19, 45). Although the effect of auxin on cell divisionhas been analyzed less intensively than its effect on cellelongation, the availability oftobacco cell suspension culturesdependent only on auxin for continuous growth has provideda convenient experimental system to investigate the role ofthis hormone in cell division processes. After a starvationperiod in hormone-free medium, such cell suspension culturesreact quickly to auxin addition with renewed cell division inconcert with the induction of specific mRNAs (47). In thissystem, cell division-associated RNAs start to accumulatewithin 30 min to 2 h after auxin addition.
'This work was supported by grants from the A.S.L.K. Kanker-fonds, the Fonds voor Geneeskundig Wetenschappelijk Onderzoek(FGWO 3.0001.82), and the Services of the Prime Minister (OOA12.0561.84 and UIAP 120C0 187). T. A. is indebted to L'Air LiquideS.A. for financial support, C. T. is a Research Assistant, and D. I. aSenior Research Assistant of the National Fund for Scientific Re-search (Belgium).
Besides these extensively studied primary effects of auxinson gene expression, a number of more long-term responseshave been characterized at the molecular level. For example,it is well documented that 18S and 25S ribosomal RNAsaccumulate after several hours of auxin treatment, due to aselective enhancement of RNA polymerase I activity (for areview, see Ref. 19). Several polypeptides that respond tolong-term auxin exposure have been identified. A 2,4-D treat-ment of intact pea seedlings induces cellulase mRNAs after alag period of approximately 24 h (49). In another study (15,16), a 24-h auxin treatment of soybean hypocotyls was shownto cause an approximate eightfold increase in the abundanceof ribosomal protein mRNAs. The similar induction kineticsfor RNA polymerase I activity and ribosomal protein mRNAaccumulation suggests that their synthesis might be regulatedcoordinately.
In the present study, we have identified a single-copy gene,called dbp, with a high basal level of expression in all parts ofmature Arabidopsis thaliana plants and elevated expressionin the root and shoot apex. The expression of dbp in isolatedpeduncle segments can be enhanced by a 4-h exposure toexogenous auxin and thus is a member ofthe set of long-termauxin-responsive genes (19). Moreover, the Dbp polypeptideis lysine-rich and shares several biochemical properties withhistone H1. Although it is biochemically and immunologi-cally distinct from histone H1, we show here that the Dbppolypeptide binds double-stranded DNA apparently in a sim-ilar fashion.
MATERIALS AND METHODS
Plant Material and Bacterial Strains
Four- to 6-week-old Arabidopsis thaliana var Columbia,grown under greenhouse conditions, were used as plant ma-terial (see also Ref. 41).
Except when specifically stated, all molecular clonings wereperformed with the Escherichia coli strain MC1061 (5). TheX phage library was amplified using the E. coli strain NM538(14). Experiments of bacterial expression were carried outwith the strain SG4044 lon- (18).
Construction and Screening of an A. thaliana cDNALibrary
The procedure for cDNA synthesis and cloning has beendescribed elsewhere in detail (12). The cDNA was synthesized
743
Plant Physiol. Vol. 89, 1989
from poly(A)+ RNA isolated from whole Arabidopsis plantsand was cloned into the PstI site of the plasmid pUC 18 (51).The library was screened for tissue-specific cDNAs follow-
ing a protocol described for screening genomic libraries (41).The clone pATClA was first characterized as hybridizing toRNA present in all organs of Arabidopsis.
RNA Preparation and Analysis
Total RNA was isolated from fresh Arabidopsis tissue es-sentially as described by Jones et al. (21) and poly(A)+ RNAwas selected by oligo(dT) cellulose affinity chromatographyas described by Slater (42). The RNA was quantified spectro-photometrically.For the detailed analysis in peduncle segments, we used the
rapid RNA extraction procedure ofLogemann el al. (25) withan additional step to remove contaminating polysaccharides.After the first precipitation following homogenization in ex-traction buffer and phenolization, the pellets were resus-pended in 0.1 M sodium acetate (pH 6.0), and polysaccharideswere precipitated by gently adding ethanol up to 10% finalvolume, and chilling on ice for 10 min. After centrifugation,the supernatants were collected and RNA was further purifiedby a second precipitation with 2.5 volumes of ethanol, at-200C.For Northern hybridizations, 10 ,ug of total RNA was
denatured in formamide and formaldehyde, electrophoresedon formaldehyde gels as described by Maniatis et al. (26), andtransferred by capillary blotting onto Hybond-N (Amersham)filters. To obtain highly specific probes, the cDNA insert ofpATClA was recloned in the plasmid pGem-2 (Promega),yielding pGATC1A, and single-stranded riboprobes were syn-thesized using SP6 polymerase (31). Northern hybridizationswere performed overnight at 680C in 50% formamide, 5 xSSPE, 0.25% nonfat dry milk, 0.5% SDS, 10% Dextransulfate, and 20 ,ug/mL denatured herring sperm DNA. Thehybridizations were washed at 68°C twice with 3 x SSC, 1%SDS, and twice with 1 x SSC, 1% SDS. The filters wereautoradiographed on Kodak XAR-5 films with intensifyingscreens at -70°C.
In Situ Hybridization
35S-Labeled riboprobes were prepared from plasmidpGATCIA using 1160 Ci/mmol "S-UTP (Amersham) togive a final specific activity of 4 x 108 dpm/,ug. Full-lengthtranscripts were reduced to an average size of 0.1 to 0.15 kb2by alkaline hydrolysis (10, 27).The size of both full-length and hydrolyzed transcripts was
checked on a 6% polyacrylamide gel and the amount ofsynthesized RNA was calculated.Root tips of 4-week-old A. thaliana plants were fixed for
30 min in 1% glutaraldehyde in 0.05% cacodylate buffer (pH7). This fixative was chosen because it permitted tissue mor-phology preservation and acceptable mRNA retention in thetissue.The tissue was then embedded in paraplast (7). Sections of
10 ,m were made with a Microtome 2050 Supercut (Reichert-Jung) and pretreated according to Cox et al. (10).
2Abbreviations: kb, kilobase pair; bp, base pair.
Hybridization of the probes to the slides was performedessentially as described by Barker et al. (2).
Autoradiography and developing were as described by An-gerer and Angerer (1).
Hormone Treatment on A. thaliana Peduncle Segments
Tests for hormone depletion and induction were performedon segments excised from Arabidopsis peduncles essentiallyaccording to a procedure for pea hypocotyls described byTheologis and Ray (46). Peduncles were excised from 4-week-old plants grown in the greenhouse, thoroughly washed insterile water, cut into 0.5-cm segments, and incubated insterile Murashige and Skoog (MS) liquid medium (34) with15 mm sucrose, 1 mM Mes (pH 6.0), and 50 ,ug/mL chlor-amphenicol at 28°C in the dark with 150 rpm shaking.Hormones were added after a 20-h washing in these con-
ditions and the different samples were further incubated for8 to 24 h. Aliquots of about 200 mg tissue were collected forRNA extraction.
Screening of a Genomic Library
An amplified subset of 5 x 104 plaques of the A. thalianagenomic library in X EMBL4 (37) was screened with a nick-translated probe of pATClA, according to standard tech-niques (26). Twelve positively hybridizing genomic cloneswere first observed, two of which remained positive whenrescreened. One clone was chosen for further analysis andwas called XATC1.
Bacterial Expression System and Protein Preparation
The bacterial expression system was based on translationalfusion with the N-terminal sequence of the X cro gene undercontrol of the X PR promoter for conditional expression inthe presence of a cIts allele. The expression vector was fromthe pLK70 series (3) and used in the presence of the plasmidpCI857 (38), in the E. coli strain SG4044.
Expression was induced by a temperature shift from 28 to42°C on a culture at A60 = 0.2, and by further incubation for4 h at 42°C. Cells were then collected by centrifugation andlyzed by osmotic shock with ice-cold water as described byKoshland and Botstein (22). The fusion polypeptide, presentin the insoluble material, was recovered in the pellet fractionafter centrifugation and resuspended in a small volume bysonication.The acid extraction test was carried out by adding 10
volumes of 0.4 N sulfuric acid (or 5% perchloric acid) to asmall volume ofconcentrated protein preparation. After mix-ing thoroughly, nonsoluble proteins were pelleted by centrif-ugation, supernatants were collected, and the extractions wererepeated& twice more. Acid-soluble proteins were recoveredfrom the pooled supernatant by precipitation with 18% (finalconcentration) TCA in the presence of 0.015% deoxycholateas carrier. The recovered pellets were washed once in acidifiedacetone (200 mL of acetone and 0.1 mL of concentratedHCI), three times in acetone, and shortly dried before resus-pension in SDS-PAGE loading buffer.
744 ALLIOTTE ET AL.
ARABIDOPSIS GENE ENCODING DNA-BINDING PROTEIN
R L PB IP F S TL TD
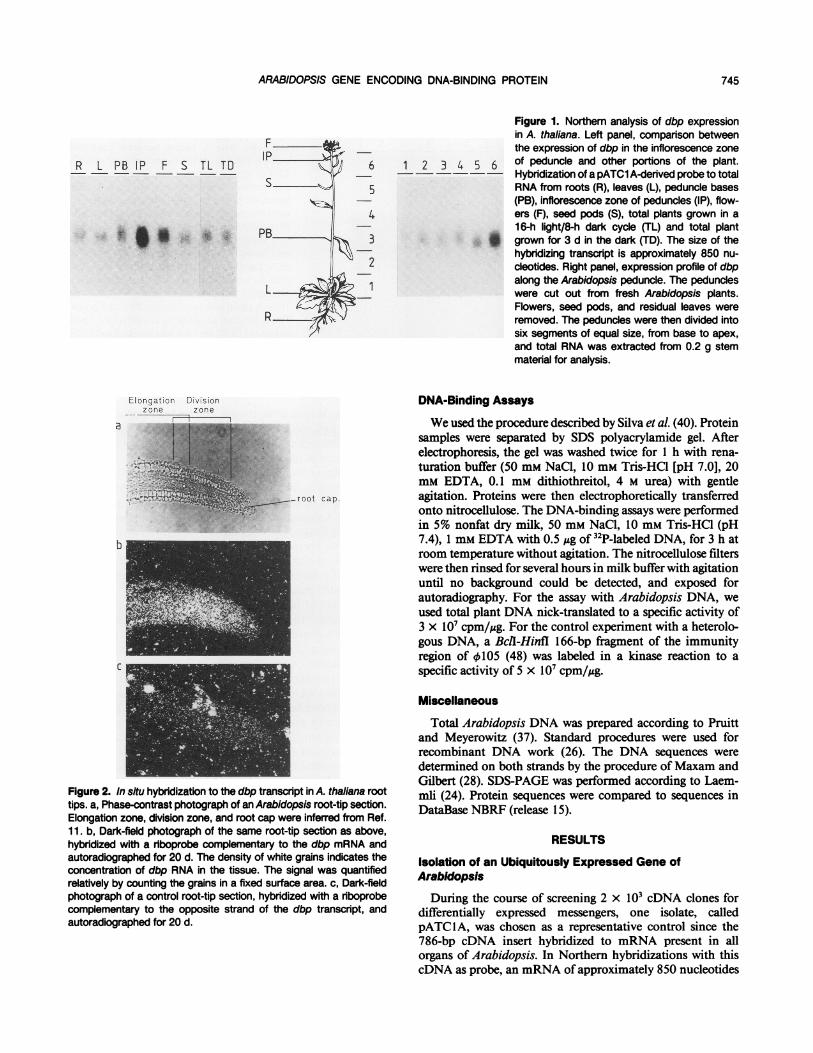
Figure 1. Northem analysis of dbp expressionin A. thaliana. Left panel, comparison betweenthe expression of dbp in the inflorescence zone
2 3 4 5 6 of peduncle and other portions of the plant.--~-- - Hybridization of a pATC1 A-derived probe to total
RNA from roots (R), leaves (L), peduncle bases(PB), inflorescence zone of peduncles (IP), flow-ers (F), seed pods (S), total plants grown in a16-h light/8-h dark cycle (TL) and total plant
* grown for 3 d in the dark (TD). The size of thehybridizing transcript is approximately 850 nu-cleotides. Right panel, expression profile of dbpalong the Arabidopsis peduncle. The peduncleswere cut out from fresh Arabidopsis plants.Flowers, seed pods, and residual leaves wereremoved. The peduncles were then divided intosix segments of equal size, from base to apex,and total RNA was extracted from 0.2 g stemmaterial for analysis.
ELongation Divisionzone zone~-i
a
-root cap.
b
c
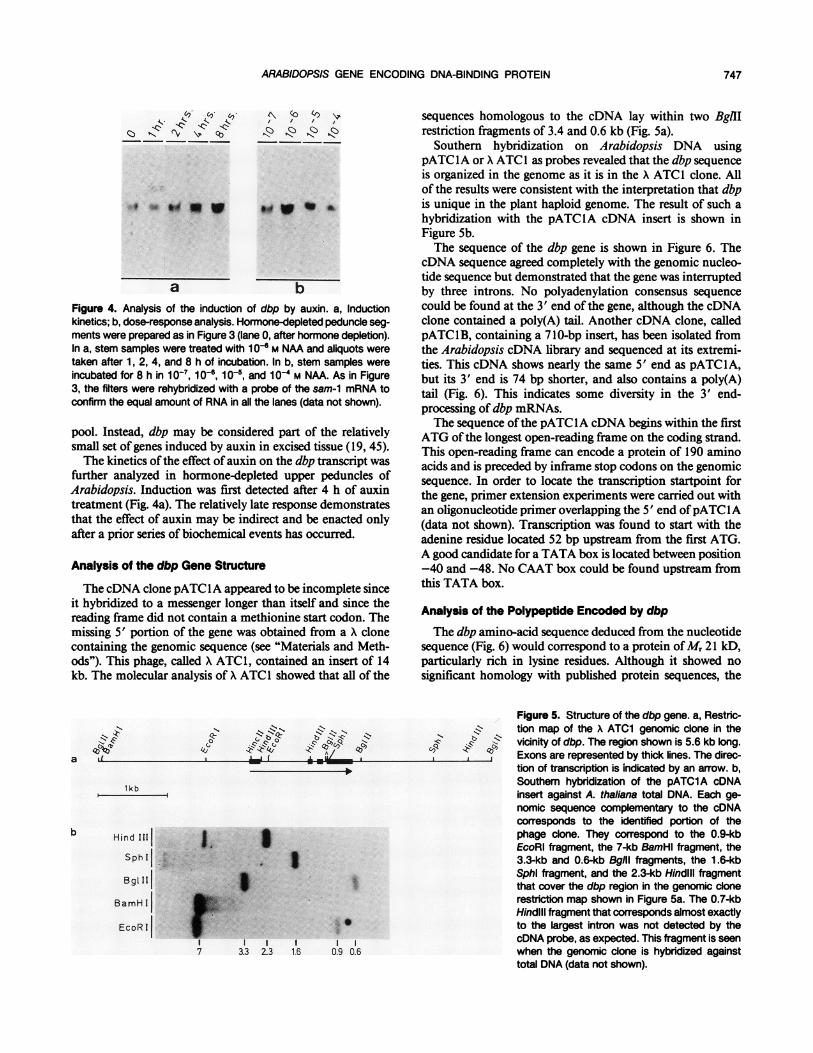
Figure 2. In situ hybridization to the dbp transcript in A. thaliana roottips. a, Phase-contrast photograph of an Arabidopsis root-tip section.Elongation zone, division zone, and root cap were inferred from Ref.11. b, Dark-field photograph of the same root-tip section as above,hybridized with a riboprobe complementary to the dbp mRNA andautoradiographed for 20 d. The density of white grains indicates theconcentration of dbp RNA in the tissue. The signal was quantifiedrelatively by counting the grains in a fixed surface area. c, Dark-fieldphotograph of a control root-tip section, hybridized with a riboprobecomplementary to the opposite strand of the dbp transcript, andautoradiographed for 20 d.
DNA-Binding Assays
We used the procedure described by Silva et al. (40). Proteinsamples were separated by SDS polyacrylamide gel. Afterelectrophoresis, the gel was washed twice for 1 h with rena-turation buffer (50 mM NaCl, 10 mm Tris-HCI [pH 7.0], 20mM EDTA, 0.1 mM dithiothreitol, 4 M urea) with gentleagitation. Proteins were then electrophoretically transferredonto nitrocellulose. The DNA-binding assays were performedin 5% nonfat dry milk, 50 mm NaCl, 10 mM Tris-HCI (pH7.4), 1 mM EDTA with 0.5 Ag of 32P-labeled DNA, for 3 h atroom temperature without agitation. The nitrocellulose filterswere then rinsed for several hours in milk buffer with agitationuntil no background could be detected, and exposed forautoradiography. For the assay with Arabidopsis DNA, weused total plant DNA nick-translated to a specific activity of3 x 107 cpm/,ug. For the control experiment with a heterolo-gous DNA, a Bcll-Hinfl 166-bp fragment of the immunityregion of 105 (48) was labeled in a kinase reaction to aspecific activity of 5 x 107 cpm/,ug.
Miscellaneous
Total Arabidopsis DNA was prepared according to Pruittand Meyerowitz (37). Standard procedures were used forrecombinant DNA work (26). The DNA sequences weredetermined on both strands by the procedure of Maxam andGilbert (28). SDS-PAGE was performed according to Laem-miu (24). Protein sequences were compared to sequences inDataBase NBRF (release 15).
RESULTS
Isolation of an Ubiquitously Expressed Gene ofArabidopsis
During the course of screening 2 x 103 cDNA clones fordifferentially expressed messengers, one isolate, calledpATC1A, was chosen as a representative control since the786-bp cDNA insert hybridized to mRNA present in allorgans of Arabidopsis. In Northern hybridizations with thiscDNA as probe, an mRNA ofapproximately 850 nucleotides
745
Plant Physiol. Vol. 89, 1989
NT
A B
was equally present in roots, leaves, peduncle bases, and seedpods, and was not affected by constant light or darkness (Fig.1). The gene encoded by this cDNA will be referred to as dbp,for reasons discussed later. In order to estimate the abundanceof this messenger, radioactively labeled cDNA prepared fromleaves was hybridized to nylon filters containing equimolaramounts of the pATClA insert and the coding region of theArabidopsis small subunit gene for ribulose-1,5-bisphosphatecarboxylase, ATS1 (23). The amount of radioactivity hybrid-izing to each gene demonstrated that the dbp messenger was
approximately 20-fold less abundant in leaves than themRNA of the small subunit family (data not shown).
Maximal Expression of dbp occurs in Apical Regions
Northern analysis showed that the dbp transcript was 4 to5 times more abundant in the uppermost part of the inflo-rescence zone on the peduncle than in the lower regions ofthe floral stems (Fig. 1). This region corresponds to a zone ofcell growth. In order to test this correlation between mRNAabundance and cell growth in organs other than the peduncle,in situ hybridizations were carried out on Arabidopsis roottips. Figure 2 shows a representative example of 24 in situhybridizations of root segments from two independent exper-iments. The signal obtained with a single-stranded riboprobe,complementary to the dbp mRNA, is stronger in the divisionzone of the root tip and approximately 4 times lower in theelongation zone and in the root cap. The concentration ofdbp mRNA in these regions may indicate either more dbpmessenger per cell, or because meristematic cells are smallerthan nondividing ones, more cells per unit area. In eithercase, the probes are clearly monitoring dbp mRNA sinceriboprobes complementary to the antisense strand of dbp,used as a control for nonspecific hybridization, showed a
uniform background labeling of 14 independently analyzedroot tips (Fig. 2c).
dbp Expression Responds to Exogenous Auxin
We investigated whether the expression ofdbp in Arabidop-sis apical regions was controlled by plant hormones. Follow-
Figure 3. Effect of plant hormones on theexpression of dbp. The upper third of A. thalianapeduncles (lane 1, before hormone depletion)was cut into 0.5-cm segments and incubated for20 h in liquid medium without hormones. Afterthis hormone-depleting treatment (lane 2, afterhormone depletion), the peduncle segmentswere incubated, respectively, in the presence of
13 00W N 6-benzylaminopurine (lanes 3 and 4), naphtha-lene acetic acid (lanes 5 and 6), and gibberellicacid (lanes 7 and 8) at a concentration of 1 -6 Meach. Samples of 0.2 g were taken for total RNAanalysis after 8 h (lanes 3, 5, and 7) and 24 hincubation with hormones (lanes 4, 6, and 8).The left panel was hybridized with a dbp probe.To confirm that all the lanes contained the sameamount of RNA, the filter was rehybridized in theright panel with a probe for the S-adenosylme-thionine synthetase (sam-1) mRNA (36), whichgave similar signals for all samples.
ing similar studies done on pea and soybean hypocotyls (forexample, Ref. 46), the upper third (the inflorescence zone andthe adjacent region, designated 5 and 6 in Fig. 1) of pedunclesfrom mature plants was cut into 0.5-cm segments, and incu-bated in liquid Murashige and Skoog medium (see "Materialsand Methods"). Analysis was carried out by Northern hybrid-ization on total RNA. As shown in Figure 3, the amount ofdbp mRNA decreased four times, down to the basal level, byincubating the peduncle segments overnight in medium con-taining no hormones. This suggested that the enhancedexpression of dbp in the shoot apex might be under positivecontrol of a labile or diffusible molecule that could be lostfrom isolated tissue segments.A number of plant growth hormones were applied to the
depleted tissue to study which restored dbp expression. Thedbp mRNA concentrations decreased in tissue with cytokinin(BAP) or no additional hormone (Fig. 3, lanes 3 and 4; anddata not shown), while gibberellic acid (GA3) stimulated geneexpression twofold over the course of 24 h. In marked con-trast, auxin (NAA) could induce maximal expression, about10 times above basal level, in less than 8 h (Fig. 3). Similardbp induction profiles were obtained with IAA and 2,4-D(data not shown). The maximal induction was obtained at aconcentration of 1o-6 M NAA: both lower and higher concen-trations resulted in weaker induction (Fig. 4b). Moreover, acombination of auxin and GA3 had the same effect as auxinalone (data not shown). Unlike the effects in upper peduncles,the expression of dbp in the lower part of Arabidopsis floralstems (Fig. 1, regions 1 and 2) remained at the basal level andshowed no response to the hormone treatment (data notshown).The specificity of the response of dbp to auxin was tested
by rehybridizing the same filter with a probe prepared from agene encoding S-adenosylmethionine synthase (sam-l) thathas been isolated from Arabidopsis (36). In contrast to thebehavior of dbp mRNA, sam- 1 transcripts did not decline invitro, nor change with any of the subsequent treatments (Fig.3). These observations suggest that the changes in dbp RNAwere not reflections of uniform fluctuations in the mRNA
746 ALLIOTTE ET AL.
ARABIDOPSIS GENE ENCODING DNA-BINDING PROTEIN
4-.D~X &)
%o4f \P
-c0* *0 In to 64 p 49
a bFigure 4. Analysis of the induction of dbp by auxin. a, Inductionkinetics; b, dose-response analysis. Hormone-depleted peduncle seg-ments were prepared as in Figure 3 (lane 0, after hormone depletion).In a, stem samples were treated with 1 0 M NM and aliquots weretaken after 1, 2, 4, and 8 h of incubation. In b, stem samples wereincubated for 8 h in 10-7, 10¶, 10-5, and 104 M NAA. As in Figure3, the filters were rehybridized with a probe of the sam-1 mRNA toconfirm the equal amount of RNA in all the lanes (data not shown).
pool. Instead, dbp may be considered part of the relativelysmall set of genes induced by auxin in excised tissue (19, 45).The kinetics ofthe effect of auxin on the dbp transcript was
further analyzed in hormone-depleted upper peduncles ofArabidopsis. Induction was first detected after 4 h of auxintreatment (Fig. 4a). The relatively late response demonstratesthat the effect of auxin may be indirect and be enacted onlyafter a prior series of biochemical events has occurred.
Analysis of the dbp Gene Structure
The cDNA clone pATClA appeared to be incomplete sinceit hybridized to a messenger longer than itself and since thereading frame did not contain a methionine start codon. Themissing 5' portion of the gene was obtained from a A clonecontaining the genomic sequence (see "Materials and Meth-ods"). This phage, called X ATC1, contained an insert of 14kb. The molecular analysis of A ATC1 showed that all of the
sequences homologous to the cDNA lay within two BglIIrestriction fragments of 3.4 and 0.6 kb (Fig. 5a).
Southern hybridization on Arabidopsis DNA usingpATClA or X ATC1 as probes revealed that the dbp sequenceis organized in the genome as it is in the X ATC1 clone. Allof the results were consistent with the interpretation that dbpis unique in the plant haploid genome. The result of such ahybridization with the pATClA cDNA insert is shown inFigure 5b.The sequence of the dbp gene is shown in Figure 6. The
cDNA sequence agreed completely with the genomic nucleo-tide sequence but demonstrated that the gene was interruptedby three introns. No polyadenylation consensus sequencecould be found at the 3' end of the gene, although the cDNAclone contained a poly(A) tail. Another cDNA clone, calledpATClB, containing a 7 10-bp insert, has been isolated fromthe Arabidopsis cDNA library and sequenced at its extremi-ties. This cDNA shows nearly the same 5' end as pATClA,but its 3' end is 74 bp shorter, and also contains a poly(A)tail (Fig. 6). This indicates some diversity in the 3' end-processing of dbp mRNAs.The sequence ofthe pATCIA cDNA begins within the first
ATG of the longest open-reading frame on the coding strand.This open-reading frame can encode a protein of 190 aminoacids and is preceded by inframe stop codons on the genomicsequence. In order to locate the transcription startpoint forthe gene, primer extension experiments were carried out withan oligonucleotide primer overlapping the 5' end ofpATC IA(data not shown). Transcription was found to start with theadenine residue located 52 bp upstream from the first ATG.A good candidate for a TATA box is located between position-40 and -48. No CAAT box could be found upstream fromthis TATA box.
Analysis of the Polypeptide Encoded by dbpThe dbp amino-acid sequence deduced from the nucleotide
sequence (Fig. 6) would correspond to a protein ofMr 21 kD,particularly rich in lysine residues. Although it showed nosignificant homology with published protein sequences, the
N
.1a
N N 4Z
"
lkb
Hind I K
SphiI
BamH I
EcoR I
I 37 3.3
S
Figure 5. Structure of the dbp gene. a, Restric-.$S tion map of the X ATC1 genomic clone in the
vicinity of dbp. The region shown is 5.6 kb long., Exons are represented by thick lines. The direc-
tion of transcription is indicated by an arrow. b,Southern hybridization of the pATC1A cDNAinsert against A. thaliana total DNA. Each ge-nomic sequence complementary to the cDNAcorresponds to the identified portion of thephage clone. They correspond to the 0.9-kbEcoRI fragment, the 7-kb BamHI fragment, the3.3-kb and 0.6-kb Bgill fragments, the 1.6-kbSphl fragment, and the 2.3-kb HindIll fragmentthat cover the dbp region in the genomic clonerestriction map shown in Figure 5a. The 0.7-kbHindill fragment that corresponds almost exactlyto the largest intron was not detected by thecDNA probe, as expected. This fragment is seenwhen the genomic clone is hybridized againsttotal DNA (data not shown).
735 TTATTTACTATATGTAATTCCTTCT6TT66A6TTTC6TTTrATAAACTACTATTT6rAT6ACCTTATTAGCCAATTAGAGCAACAAAATTTAACTTA6THind III
SWTAATA^^ TTCT6TrCAA6CAAcATATrCCATT6TTTATATrArA^6TTGcTTTrTr6CcArACCATC6At6AGCATAACCTAA6AA'^^6cTrCATCrcK P I E E H T P K K A S S
M3 6rTTC66CCGATA6A+TCA6r6CrATAACAcACATTrrTTCTCTrcTrTCCArTTTCTTrA6rTACTTAGTTCAGTAATTTrACTTCAAAT6Tr6TrT6T6T666 S A D R
1036 GTTrTrCA^T6Tr.ATACTTr.Ccr.ACTT6rAAAAA^6A6AArAAAAc6rTCATTCATCAAA6CATGG6AuA6A6Ar.TAA6T6cAOA66cTA6^AcACATD V I L A D L E K E K K T S F I K A N E E S E K S K A E N a
1235 TT666^ArAAAACAA6AAAGCACGA6Ac6A6^CTCAACTTrAGAA6ATCGA66AAAAATTAA6AA^6^^^66AAAAcGCGTCrTAc66AAAATGAAGAACW E N S K K A A V E A O L R K I E E K L E K K K A O Y G E K 4 K N
1335 AAAGTAGCTGCAATCCACAAGTTAGCAGAA6AGAAGAGAGCAATGGTTGAA6CTA^AAAAAGGAGAAGACTTCTCGAAGCTGAAGAAATGGGG6CTAAGTK V A A I H K L A E E K R A M V E A K K G E E L L E A E E M G A K
1435 ATAGAGCCACTGGTGTAGTACCAAAGGCAACGTGGG6ATGTTTCTAAGCCTTrATTGAATTTGTArTCTTTGAACAATTCATCTCTGrTcTrcTTTcTTCTY R A T G V V P K A T C G C F
size and composition of the deduced protein (Table I) resem-bled pea and human HI histones (for a review, see Ref. 50).Dbp differs noticeably from HI in that it contains cysteineand tryptophan and has a low basic/acidic ratio (Table I).
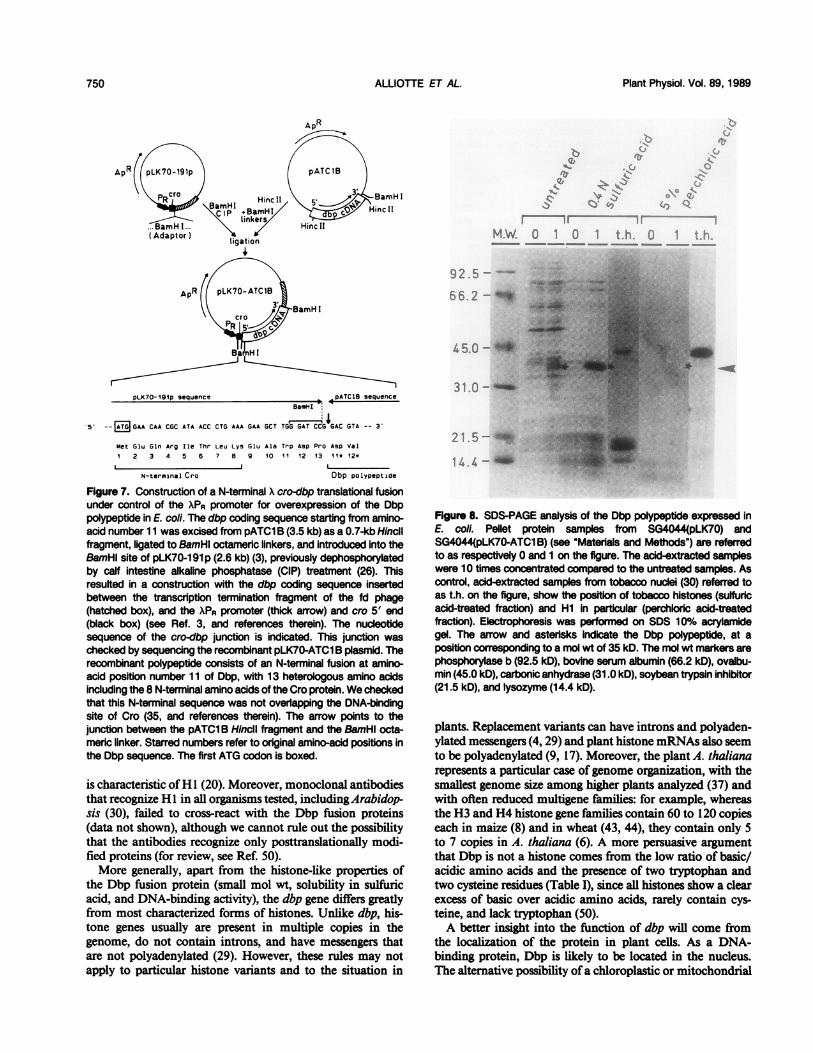
In order to further analyze the Dbp polypeptide, we intro-duced the coding sequence of the pATClB cDNA insert intoa bacterial expression vector, replacing the first 10 aminoacids with a short N-terminal translational fusion with the Xcro gene (Fig. 7). We checked carefully that the very short Crosequence present in the Dbp fusion protein did not overlapthe DNA-binding site of Cro (35, and references therein).As shown in Figure 8, expression of dbp in bacteria gives
rise to a polypeptide with an apparent Mr of 35 kD on SDSpolyacrylamide gels. The Dbp polypeptide is soluble in 0.4 Nsulfuric acid, a property of basic proteins that is used for thepurification of histones, but not in 5% perchloric acid, whichdistinguishes it from histone HI (20). Furthermore, the bac-
Figure 6. Sequence of the dbp gene. Thenucleotide sequence shown is 2356 bplong. Numbering refers to the transcrip-tion start, indicated by +1 above it. Acandidate TATA sequence is boxed. In-tron positions are shown by hooks. The5' and 3' ends of both pATClA andpATClB cDNA inserts are indicated byarrows. The 3' end of the pATClA andpATClB sequences are followed by 6and 10 adenine residues, respectively,which is the evidence of posttranscrip-tional polyadenylation. The correspondingamino acids are written under the nucleo-tide sequence in the one-letter code. Thestop codon is marked by an asterisk.Positions of the restriction sites shown inFigure 5a are indicated.
terial-made Dbp protein exhibits DNA-binding activity (Fig.9). The DNA-binding assay was performed on protein extractsseparated by PAGE, electroblotted into nitrocellulose filters,and incubated with 32P-labeled DNA in a buffer adjusted to50 mM NaCl and containing 5% nonfat dry milk. Under theseassay conditions, purified tobacco HI and the Dbp fusionprotein bound both Arabidopsis total DNA (Fig. 9b) and afragment of bacteriophage DNA. Bacterial extracts also con-tained a 38-kD protein that bound to Arabidopsis but not tobacteriophage DNA (Fig. 9b). The reaction conditions weresufficiently restrictive to exclude binding to any of the moreabundant proteins on the gel.
These experiments demonstrate that the DNA-bindingproperty of the Dbp fusion does not appear to be moresequence-specific than histone HI, or if it is, then the recog-nition sequence for dbp must be relatively common andaccessible in unfractionated DNA. Dbp binding, in fact, is less
ALLIOTTE ET AL.748
ARABIDOPSIS GENE ENCODING DNA-BINDING PROTEIN
Table I. Comparison between the Amino Acid Compositions and Mol Wt of the Dbp Polypeptide, theHuman H1 Histone (13), and the H1 Histone from Pea (17)
Amino Acid Distribution Human Hi Histone Pea Hi Histone Dbp
specific than the binding activity of the 38-kD E. coli proteinapparent in Figure 9b, but not in Figure 9c. These experimentsdo not exclude the possibility that dbp and H 1 exhibit distin-guishable DNA-binding properties in vivo.
DISCUSSION
We have isolated a single-copy gene from A. thaliana, dbp,encoding a DNA-binding protein. The dbp gene shows auniform basal level of expression in all parts of mature plantsand an enhanced level in the root and shoot apex. In roots,the region of enhanced expression was localized by in situhybridization and shown to correspond to the zone of celldivision in the apex.The enhanced level of expression of dbp in the shoot apex
coincides with a site ofauxin biosynthesis in plants (for review,see Ref. 39). Ifthis region is excised and depleted ofhormones,the concentration of dbp mRNA declines to levels foundelsewhere in the plant. The mRNA abundance is raised ifexcised tissue is treated with auxin. Thus, it is possible thatthe distribution of dbp RNA in intact plants partially reflectsauxin concentration. However, lower portions ofthe peduncledid not respond to the hormone treatments. The expressionof dbp, therefore, must be limited not only by the amount ofavailable hormone, but also by a second factor influencingtranscription or messenger processing or stability that couldbe more abundant in growing tissues than elsewhere.The regulation of the dbp gene is distinguishable from
auxin-induced genes that form part of the primary auxinresponse (19, 45, 47). The response of dbp to the auxintreatment is slow (between 4 and 8 h), and the dose-response
curve shows a maximum at physiological concentrations of106 M (for review, see Ref. 19) and inhibition at high auxinconcentrations. Such a dose-response curve is typical of theeffect of auxin on the growth of different plant organs (forreview, see Ref. 33) and similar to an auxin-regulated cellulaseof peas (49). The possibility to induce dbp expression aboveits basal level using gibberellic acid further supports the con-clusion that this enhanced expression is under multiple con-trols. Both auxin and gibberellic acid are known to stimulatestem elongation in a variety ofspecies (32) and may, therefore,regulate overlapping sets of genes under appropriate assayconditions. The induction of dbp by auxin is thus part of theindirect, long-term auxin response in the plant tissue (seeintroduction).The dbp amino-acid sequence shows a similarity with his-
tone H1, based on the size of the polypeptide and the highlysine content. Moreover, the strong ionic charge of thepolypeptide decreases its mobility on SDS-PAGE, leading toan apparently higher Mr (35 kD instead of 21 kD), in thesame manner as the purified tobacco H1 used as a control(Fig. 8). In addition, the Dbp polypeptide is extractable in 0.4N sulfuric acid, like histones. Above all, we could demonstratethat Dbp and H 1 share similar, apparently nonspecific, DNA-binding properties. The new gene was named dbp because itencodes a DNA-binding protein.However, despite the considerable variations in the H1
sequence between cell types and organisms (for a review, seeRef. 29), the Dbp protein can probably not be classified as avariant of histone HI. First and foremost, the Dbp polypep-tide is not soluble in 5% perchloric acid, whereas this solubility
749
Plant Physiol. Vol. 89, 1989
-BamH I
ApR
pLK70-191p sequence opATCLB sequence
BemHI,
5' -- Ag GAA CAA CGC ATA ACC CTG AAA GAA GCT TGG GAT CCG GAC GTA -- 3-
Met Glu Gln Arg Ile Thr Leu Lys Glu Ala Trp Asp Pro Asp Val
1 2 3 4 5 6 7 8 9 10 11 12 13 11* 12*
Dbp polypepticaeN-terminai Cro
Figure 7. Construction of a N-terminal X cro-dbp translational fusionunder control of the XPR promoter for overexpression of the Dbppolypeptide in E. coli. The dbp coding sequence starting from amino-acid number 11 was excised from pATC1 B (3.5 kb) as a 0.7-kb Hincilfragment, ligated to BamHl octameric linkers, and introduced into theBamHl site of pLK70-191 p (2.6 kb) (3), previously dephosphorylatedby calf intestine alkaline phosphatase (CIP) treatment (26). Thisresulted in a construction with the dbp coding sequence insertedbetween the transcription termination fragment of the fd phage(hatched box), and the XPR promoter (thick arrow) and cro 5' end(black box) (see Ref. 3, and references therein). The nucleotidesequence of the cro-dbp junction is indicated. This junction waschecked by sequencing the recombinant pLK70-ATC1 B plasmid. Therecombinant polypeptide consists of an N-terminal fusion at amino-acid position number 11 of Dbp, with 13 heterologous amino acidsinciuding the 8 N-terminal amino acids of the Cro protein. We checkedthat this N-terminal sequence was not overlapping the DNA-bindingsite of Cro (35, and references therein). The arrow points to thejunction between the pATC1 B Hincll fragment and the BamHl octa-meric linker. Starred numbers refer to original amino-acid positions inthe Dbp sequence. The first ATG codon is boxed.
is characteristic ofH 1 (20). Moreover, monoclonal antibodiesthat recognize Hl in all organisms tested, includingArabidop-sis (30), failed to cross-react with the Dbp fusion proteins(data not shown), although we cannot rule out the possibilitythat the antibodies recognize only posttranslationally modi-fied proteins (for review, see Ref. 50).More generally, apart from the histone-like properties of
the Dbp fusion protein (small mol wt, solubility in sulfuricacid, and DNA-binding activity), the dbp gene differs greatlyfrom most characterized forms of histones. Unlike dbp, his-tone genes usually are present in multiple copies in thegenome, do not contain introns, and have messengers thatare not polyadenylated (29). However, these rules may notapply to particular histone variants and to the situation in
r ---
M.W. 0 1 0 1 t.hh. u
92.5-
65.2- L
45.0-^ _
31.0-l
21 .5--
14 4 -
Figure 8. SDS-PAGE analysis of the Dbp polypeptide expressed inE. coli. Pellet protein samples from SG4044(pLK70) andSG4044(pLK70-ATC1 B) (see "Materials and Methods") are referredto as respectively 0 and 1 on the figure. The acid-extracted sampleswere 10 times concentrated compared to the untreated samples. Ascontrol, acid-extracted samples from tobacco nuclei (30) referred toas t.h. on the figure, show the position of tobacco histones (sulfuricacid-treated fraction) and Hi in particular (perchloric acid-treatedfraction). Eectrophoresis was performed on SDS 10% acrylamidegel. The arrow and asterisks indicate the Dbp polypeptide, at aposition corresponding to a mol wt of 35 kD. The mol wt markers arephosphorylase b (92.5 kD), bovine serum albumin (66.2 kD), ovalbu-min (45.0 kD), carbonic anhydrase (31.0 kD), soybean trypsin inhibitor(21.5 kD), and lysozyme (14.4 kD).
plants. Replacement variants can have introns and polyaden-ylated messengers (4, 29) and plant histone mRNAs also seemto be polyadenylated (9, 17). Moreover, the plant A. thalianarepresents a particular case ofgenome organization, with thesmallest genome size among higher plants analyzed (37) andwith often reduced multigene families: for example, whereasthe H3 and H4 histone gene families contain 60 to 120 copieseach in maize (8) and in wheat (43, 44), they contain only 5to 7 copies in A. thaliana (6). A more persuasive argumentthat Dbp is not a histone comes from the low ratio of basic/acidic amino acids and the presence of two tryptophan andtwo cysteine residues (Table I), since all histones show a clearexcess of basic over acidic amino acids, rarely contain cys-teine, and lack tryptophan (50).A better insight into the function of dbp will come from
the localization of the protein in plant cells. As a DNA-binding protein, Dbp is likely to be located in the nucleus.The alternative possibility ofa chloroplastic or mitochondrial
750 ALLIOTTE ET AL.
ARABIDOPSIS GENE ENCODING DNA-BINDING PROTEIN
t.h. 1 0
'..s,.
i& -a&i
435 Za ;Z
a_ .
Figure 9. DNA-binding assay on the Dbp pro-tein. a, Coomassie-stained control; b, autoradi-ogram of a DNA-binding assay with 32P-labeledArabidopsis DNA; c, autoradiogram of a DNA-binding assay with a 32P-labeled double-strandedDNA fragment from the Bacillus t105 phage. A
t. h. 1 a t. l 0preparation of purified tobacco Hi histone isindicated by t.h. Protein samples fromSG4044(pLK70) and SG4044(pLK70-ATC1 B)are referred to as 0 and 1, respectively, as inFigure 8. Total protein samples from E. coli
S _ . (independent from those used in Fig. 8) wereused in this experiment to check the specificityof the DNA-binding activity. Electrophoresis wasdone on SDS 10% acrylamide gel. The arrow
dhh_ and asterisks indicate the Dbp polypeptide. In b,the assay with Arabidopsis total DNA revealedsome E. coli DNA-binding protein, both in 0 and1. In c, the assay was performed with the Bcll-Hinfl 166-bp fragment of the immunity region of4105 phage (48). A strong DNA-binding activity(not shown) was also detected on the low-mo-
b c lecular wt bands of the bacterial preparationswhich migrate at the same position as the nu-cleosomal tobacco histones and which alsoshowed some enrichment after sulfuric acid ex-traction (Fig. 8). The apparent mol wt of tobaccohistone Hi (43 kD) and of the Dbp fusion protein(35 kD) is indicated.
DNA-binding protein is unlikely since there is no distinguish-able transit peptide in the amino acid sequence. Experimentsusing immuno-detection with antibodies raised against thefusion polypeptide will confirm the nuclear location andpossibly the association of the Dbp protein with chromatin.
ACKNOWLEDGMENTS
The authors are particularly grateful to Dr. L. Mazzolini fortobacco histone samples, to Dr. Kathleen Cox and Dr. Robert Gold-berg for communicating their method for in situ hybridization beforepublication and to Dr. Gary Drews for advice and technical help. Wethank Li-Huang Zhu, Jan Gielen, and Jeroen Coppieters for excellenttechnical assistance; Chris Bowler and Dr. Chumpol Borkird forcritical reading of the manuscript; Martine De Cock, Karel Spruyt,Stefaan Van Gijsegem, and Vera Vermaercke for help in preparingit.
LITERATURE CITED
1. Angerer LM, Angerer RC (1981) Detection of polyA+ RNA insea urchin eggs and embryos by quantitative in situ hybridi-zation. Nucleic Acids Res 9: 2819-2840
2. Barker SJ, Harada JJ, Goldberg RB (1988) Cellular localizationof soybean storage protein mRNA in transformed tobaccoseeds. Proc Natl Acad Sci USA 85: 458-462
3. Botterman J, ZabeauM (1987) A standardized vector system formanipulation and enhanced expression ofgenes in Escherichiacoli. DNA 6: 583-591
5. Casadaban MJ, Cohen SN (1980) Analysis ofgene control signalsby DNA fusion and cloning in Escherichia coli. J Mol Biol138:179-207
6. Chaboute M-E, Chaubet N, Philipps G, Ehling M, Gigot C
(1987) Genomic organization and nucleotide sequences oftwohistone H3 and two histone H4 genes ofArabidopsis thaliana.Plant Mol Biol 8: 179-191
7. Chadwick R, McGinnis W (1987) Temporal and spatial distri-bution of transcripts from the Deformed gene of Drosophila.EMBO J 6: 779-789
8. Chaubet N, Philipps G, Chaboute M-E, Ehling M, Gigot C(1986) Nucleotide sequences of two corn histone H3 genes.Genomic organization of the corn histone H3 and H4 genes.Plant Mol Biol 6: 253-263
9. Chaubet N, Chaboute M-E, Clement B, Ehling M, Philipps G,Gigot C (1988) The histone H3 and H4 mRNAs are polyade-nylated in maize. Nucleic Acids Res 16: 1295-1304
10. Cox K, DeLeon DV, Angerer LM, Angerer RC (1984) DetectionofmRNAs in sea urchin embryos by in situ hybridization usingasymmetric RNA probes. Dev Biol 101: 485-502
11. Cutter EG (1971) Plant Anatomy: Experiment and Interpreta-tion, Part 2: Organs (A series ofstudent texts in ContemporaryBiology). Edward Arnold, London, pp 5-44
12. De Loose M, Alliotte T, Gheysen G, Genetello C, Gielen J,Soetaert P, Van Montagu M, Inze D (1988) Primary structureof a hormonally regulated jB-glucanase of Nicotiana plumba-ginifolia. Gene 70: 13-23
13. Doenecke D, Tonjes R (1986) Differential distribution of lysineand arginine residues in the closely related histones HI' andH5. Analysis of a human HI' gene. J Mol Biol 187: 461-464
14. Frischauf A-M, Lehrach H, Poustka A, Murray N (1983)Lambda replacement vectors carrying polylinker sequences. JMol Biol 170: 827-842
15. Gantt JS, Key JL (1983) Auxin-induced changes in the level oftranslatable ribosomal protein messenger ribonucleic acids insoybean hypocotyl. Biochemistry 22: 4131-4139
16. Gantt JS, Key JL (1985) Coordinate expression of ribosomalprotein in RNAs following auxin treatment of soybean hypo-cotyls. J Biol Chem 260: 6175-6181
17. Gantt JS, Key JL (1987) Molecular cloning of a pea H 1 histonecDNA. Eur J Biochem 166: 119-125
18. Gottesman S, Gottesman M, Shaw J, PearsonML (1981) Protein
751
Plant Physiol. Vol. 89, 1989
degradation in E. coli: the Ion mutation and bacteriophagelambda N and cII protein stability. Cell 24: 225-233
20. Johns EW (1977) The isolation and purification of histones.Methods Cell Biol 16: 183-203
21. Jones JDG, Dunsmuir P, Bedbrook J (1985) High level expres-sion of introduced chimeric genes in regenerated transformedplants. EMBO J 4: 2411-2418
22. Koshland D, Botstein D (1980) Secretion of beta-lactamase re-quires the carboxy end of the protein. Cell 20: 749-760
23. Krebbers E, Seurinck J, Herdies L, Cashmore AR, Timko MP(1989) Four genes in two diverged subfamilies encode theribulose-1,5-bisphosphate carboxylase small subunit polypep-tides ofArabidopsis thaliana. Plant Mol Biol (in press)
24. Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680-685
25. Logemann J, Schell J, Willmitzer L (1987) Improved methodfor the isolation of RNA from plant tissues. Anal Biochem163: 16-20
27. Martineau B, Taylor WC (1986) Cell-specific photosyntheticgene expression in maize determined using cell separationtechniques and hybridization in situ. Plant Physiol 82: 613-618
28. Maxam AM, Gilbert W (1980) Sequencing end-labeled DNAwith base-specific chemical cleavages. Methods Enzymol 65:499-559
29. Maxson R, Cohn R, Kedes L, Mohun T (1983) Expression andorganization of histone genes. Annu Rev Genet 17: 239-277
30. Mazzolini L, Vaeck M, Van Montagu M (1989) Conservedepitopes on plant H1 histones recognized by monoclonal an-tibodies. Eur J Biochem (in press)
31. Melton DA, Krieg PA, Rebagliati MR, Maniatis T, Zinn K,Green MR (1984) Efficient in vitro synthesis of biologicallyactive RNA and RNA hybridization probes from plasmidscontaining a bacteriophage SP6 promoter. Nucleic Acids Res12: 7035-7056
32. Metraux J-P (1987) Gibberellins and plant cell elongation. In PJDavies, ed, Plant Hormones and Their Role in Plant Growthand Development. Martinus Nijhoff Publishers, Dordrecht, pp296-317
33. Moore TC (1979) Biochemistry and Physiology of Plant Hor-mones. Springer-Verlag, New York, pp 32-89
34. Murashige T, Skoog F (1962) A revised medium for rapid growthand bio assays with tobacco tissue cultures. Physiol Plant 15:473-497
35. Pakula AA, Young VB, Sauer RT (1986) Bacteriophage X cro
mutations: effects on activity and intracellular degradation.Proc Natl Acad Sci USA 83: 8829-8833
36. Peleman J, Boerjan W, Engler G, Seurinck J, Botterman J,Alliotte T, Van Montagu M, Inze D (1989) Strong cellularpreference in the expression of a housekeeping gene of Arabi-dopsis thaliana encoding S-adenosylmethionine synthetase.Plant Cell 1: 81-93
37. Pruitt RE, Meyerowitz EM (1986) Characterization of the ge-nome ofArabidopsis thaliana. J Mol Biol 187: 169-183
38. Remaut E, Tsao H, Fiers W (1983) Improved plasmid vectorswith a thermoinducible expression and temperature-regulatedrunaway replication. Gene 22: 103-113
39. Sembdner G, Gross D, Liebisch H-W, Schneider G (1980) Bio-synthesis and metabolism of plant hormones. In J MacMillan,ed, Hormonal regulation of development I. Encyclopedia ofPlant Physiology (New Series), Vol 9. Springer-Verlag, Berlin,pp 281-444
40. Silva CM, Tuily DB, Petch LA, Jewell CM, Cidlowski JA (1987)Application of a protein-blotting procedure to the study ofhuman glucocorticoid receptor interactions with DNA. ProcNatl Acad Sci USA 84: 1744-1748
41. Simoens CR, Peleman J, Valvekens D, Van Montagu M, Inze D(1988) Isolation of genes expressed in specific tissues of Ara-bidopsis thaliana by differential screening ofa genomic library.Gene 67: 1-11
42. Slater RJ (1984) The purification of poly(A)-containing RNA byaffinity chromatography. Methods Mol Biol 2: 117-120
43. Tabata T, Sasaki K, Iwabuchi M (1983) The structural organi-zation and DNA sequence ofa wheat histone H4 gene. NucleicAcids Res 11: 5865-5875
44. Tabata T, Fukasawa M, IwabuchiM (1984) Nucleotide sequenceand genomic organization of a wheat histone H3 gene. MolGen Genet 196: 397-400
45. Theologis A (1986) Rapid gene regulation by auxin. Annu RevPlant Physiol 37: 407-438
46. Theologis A, Ray PM (1982) Early auxin-regulated polyadeny-lated mRNA sequences in pea stem tissue. Proc Natl Acad SciUSA 79: 418-421
47. van der Zaal EJ, Memelink J, Mennes AM, Quint A, LibbengaKR (1987) Auxin-induced mRNA species in tobacco cell cul-tures. Plant Mol Biol 10: 145-157
48. Van Kaer L, Van Montagu M, Dhaese P (1987) Transcriptionalcontrol in the EcoRI-F immunity region of Bacillus subtilisphage 4105. Identification and unusual structure of the oper-ator. J Mol Biol 197: 55-67
49. Verma DPS, Maclachlan GA, Byrne H, Ewings D (1975) Regu-lation and in vitro translation of mRNA for cellulase fromauxin-treated pea epicotyls. J Biol Chem 250: 1019-1026