Activation of Adenovirus Early Promoters and Lytic Phase inDifferentiated Strata of Organotypic Cultures of
Human KeratinocytesFrancisco Noya,1 Cristina Balague,2† N. Sanjib Banerjee,1 David T. Curiel,2,3
Thomas R. Broker,1 and Louise T. Chow1*Department of Biochemistry and Molecular Genetics,1 Departments of Medicine, Pathology, and Surgery,
Division of Human Gene Therapy,3 and Gene Therapy Center,2 University of Alabamaat Birmingham, Birmingham, Alabama 35294-0005
Received 27 November 2002/Accepted 10 March 2003
Human oncolytic adenoviruses have been used in clinical trials targeting cancers of epithelial origin. To gaina better understanding of the infectious cycle of adenovirus in normal human squamous tissues, we examinedthe viral infection process in organotypic cultures of primary human keratinocytes. We show that for theinfection to occur, wounding of the epithelium is required. In addition, infection appears to initiate at the basalor parabasal cells that express the high-affinity coxsackievirus-adenovirus receptor, CAR, whereas the pro-ductive phase takes place in differentiated cells. This is due, at least in part, to the differentiation-dependentactivation of the E1A and E2A early promoters and E4 promoters. We also show that adenovirus infectiontriggers a response mediated by the abnormal accumulation of cyclin E and p21cip1 proteins similar to the onepreviously observed in human papillomavirus-infected tissues. However, the virus seems to be able to overcomeit, at least partially.
During the last decade, human adenoviruses have been ex-ploited as vectors for gene transfer and gene therapy of variousmonogenic and polygenic diseases, with limited success. Morerecently, the potential of employing replication-competent ad-enoviruses as oncolytic agents has been put to the test inclinics, especially with neoplasias of epithelial origin, such ashead-and-neck and prostate cancers (9, 23). The specificity ofthis new generation of anticancer agents relies on its prefer-ential and efficient replication in tumors (1). The systems tostudy oncolytic adenoviruses ordinarily employ primary ortransformed cell lines grown as monolayer cultures or as xeno-grafts in nude mice. Comparatively few efforts have been madeto expand our understanding of the viral life cycle in the nor-mal stratified human epithelia, such as those encountered dur-ing clinical gene transfer. Thus, important aspects of the biol-ogy of the virus as related to vector safety could have beenoverlooked. Furthermore, differences in the physical charac-teristics of the epithelial tissue and submerged monolayer cellscould also have important consequences for the infectivity ofthe virus in vivo. For instance, the protective cornified layer ofa stratified epithelium may act as an effective barrier againstadenovirus infection in addition to the possible heterogeneousdistribution of the coxsackievirus-adenovirus receptor (CAR)across the epithelium (14, 15).
One alternative culture system that faithfully recapitulatesthe morphological and physiological characteristics of the nor-
mal stratified squamous epithelium is the organotypic raft cul-ture of primary human keratinocytes (PHKs) (hereafter calledraft cultures). This system is composed of a dermal equivalentconsisting of fibroblasts embedded in a collagen matrix, on topof which neonatal foreskin keratinocytes are seeded. When theassembly is cultured at the air-medium interphase for 9 days,the keratinocytes proliferate, stratify, and differentiate, devel-oping into a three-dimensional epithelium that closely resem-bles the native tissue. This culture can be harvested and stud-ied by standard histological techniques (for a review, seereference 7). Hutchin et al. (15) have employed this systemwith nonreplicating adenoviruses for the study of gene transferto oropharyngeal epithelial cells. A similar system was recentlyused to show that the binding of the fiber to CAR facilitatesviral dissemination (37). On the other hand, we used the PHKraft culture system to screen and evaluate new conditionallyreplicating adenoviruses that would only proliferate in humanpapillomavirus (HPV)-positive cells on the basis of a conser-vation of adenovirus-HPV protein functions (described be-low). We have reported unexpected findings. For instance, anadenovirus type 5 E1A mutant, CB016, unable to bind pRb,p107, and p300, replicates in the fully stratified and differen-tiated squamous epithelium with delayed kinetics compared toreplication in HPV E6/E7-transduced raft cultures (3). Theseobservations indicate that adenoviruses have alternative meansof mobilizing cellular genes needed to support viral DNA rep-lication in differentiated keratinocytes, such as those requiredto supply deoxyribonucleoside triphosphates.
Indeed, many characteristics of human adenoviruses areshared by other small DNA tumor viruses, such as HPVs. Forexample, both virus families encode proteins that inactivatep53, which normally prevents DNA synthesis in the presence ofDNA damage or promotes apoptosis to maintain genome sta-
* Corresponding author. Mailing address: Department of Biochem-istry and Molecular Genetics, University of Alabama at Birmingham,510 McCallum Basic Health Sciences Bldg., 1918 University Blvd.,Birmingham, AL 35294-0005. Phone: (205) 975-8300. Fax: (205) 975-6075. E-mail: [email protected].
† Present address: Almirall Prodesfarma, Target Validation Unit,Barcelona Science Park, Barcelona 08028, Spain.
bility (29, 36). Both also inactivate the tumor suppressor reti-noblastoma susceptibility protein (pRb), which controls G1/Stransition (4). The differentiation-dependent expression of theoncogenic HPV E7 protein reestablishes an S phase milieu inotherwise differentiated squamous cells, in which productiveinfection takes place (5, 10). Thus, it is possible that adenovirusreplication is also differentiation dependent. As a matter offact, CB016-induced cytopathic effects (CPE) in PHK raft cul-tures occurred in the differentiated strata, although the kera-tinocytes had been infected prior to being raised to the air-medium interphase (3). These observations strongly suggestthat adenovirus infection is differentiation dependent in this
setting. Several early reports suggest the importance of thedifferentiation status of the cell in promoting the activation ofthe early genes required to initiate the amplification of adeno-virus (13, 18, 22, 34). The notion that the virus life cycle isaffected by cellular differentiation is also suggested by thepreferential expression of CAR, the high-affinity receptor ofadenovirus, in the less-differentiated cells of the epithelium(15, 37).
Here we report the use of the raft culture system to study theinfectious cycle of the wild-type adenovirus at high and lowmultiplicities of infection (MOI). Even though the adenovirusserotypes commonly used in virotherapy do not naturally infect
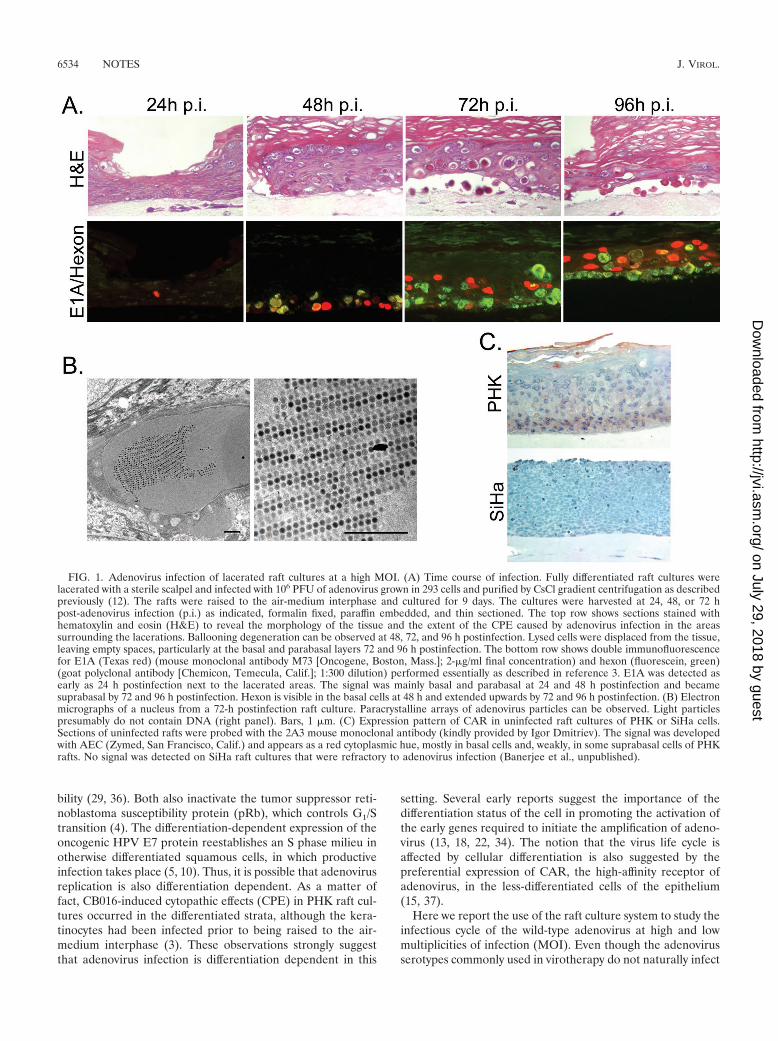
FIG. 1. Adenovirus infection of lacerated raft cultures at a high MOI. (A) Time course of infection. Fully differentiated raft cultures werelacerated with a sterile scalpel and infected with 106 PFU of adenovirus grown in 293 cells and purified by CsCl gradient centrifugation as describedpreviously (12). The rafts were raised to the air-medium interphase and cultured for 9 days. The cultures were harvested at 24, 48, or 72 hpost-adenovirus infection (p.i.) as indicated, formalin fixed, paraffin embedded, and thin sectioned. The top row shows sections stained withhematoxylin and eosin (H&E) to reveal the morphology of the tissue and the extent of the CPE caused by adenovirus infection in the areassurrounding the lacerations. Ballooning degeneration can be observed at 48, 72, and 96 h postinfection. Lysed cells were displaced from the tissue,leaving empty spaces, particularly at the basal and parabasal layers 72 and 96 h postinfection. The bottom row shows double immunofluorescencefor E1A (Texas red) (mouse monoclonal antibody M73 [Oncogene, Boston, Mass.]; 2-�g/ml final concentration) and hexon (fluorescein, green)(goat polyclonal antibody [Chemicon, Temecula, Calif.]; 1:300 dilution) performed essentially as described in reference 3. E1A was detected asearly as 24 h postinfection next to the lacerated areas. The signal was mainly basal and parabasal at 24 and 48 h postinfection and becamesuprabasal by 72 and 96 h postinfection. Hexon is visible in the basal cells at 48 h and extended upwards by 72 and 96 h postinfection. (B) Electronmicrographs of a nucleus from a 72-h postinfection raft culture. Paracrystalline arrays of adenovirus particles can be observed. Light particlespresumably do not contain DNA (right panel). Bars, 1 �m. (C) Expression pattern of CAR in uninfected raft cultures of PHK or SiHa cells.Sections of uninfected rafts were probed with the 2A3 mouse monoclonal antibody (kindly provided by Igor Dmitriev). The signal was developedwith AEC (Zymed, San Francisco, Calif.) and appears as a red cytoplasmic hue, mostly in basal cells and, weakly, in some suprabasal cells of PHKrafts. No signal was detected on SiHa raft cultures that were refractory to adenovirus infection (Banerjee et al., unpublished).
cornified stratified epithelium, we show that the system can stillprovide important information on the virus life cycle and virus-host interactions that are likely to occur in a clinical setting.We also examined the transcriptional activities of several ad-enoviral early promoters by using a reporter assay. We showthat in a productive infection, adenovirus has to have access tothe basal layers of the epithelium where CAR is more abun-dant. However, at a low MOI, the virus remains at a low copynumber until the infected cell or its daughter cell starts thedifferentiation program. Once the infected cell enters the mid-strata of the epithelium, the early promoters are activated,promoting the amplification of the viral genome. We also show
that virus infection triggers an antiviral response that involvesthe costabilization of p21cip1 and cyclin E, analogous to theone exhibited against HPV-induced unscheduled DNA synthe-sis in raft cultures and in benign HPV lesions (16, 25, 30).However, unlike HPVs, adenovirus seems to be able to over-come this response, at least in part.
Adenovirus infects only lacerated raft cultures. We firstexamined the infection process of human adenovirus type 5 ina fully stratified and differentiated squamous epithelium. Theassembly of PHKs and dermal equivalent was lifted to theair-medium interphase, cultured for 9 days, and harvested.Twenty-four, 48, 72, or 96 h before harvest, adenovirus was
FIG. 2. Differentiation-dependent adenovirus promoter activation and virus reproduction at a low MOI. (A) Time course of infection oflacerated PHK raft cultures. Fully differentiated raft cultures were lacerated and infected with 104 PFU of adenovirus. The rafts were harvested24, 48, and 72 h postinfection. (Top) The sections were probed for E1A and hexon proteins as described in the legend to Fig. 1. E1A (red) andhexon (green) were detected in a small number of suprabasal cells 72 h postinfection but not at earlier time points. (Bottom) In situ hybridizationof adenovirus DNA was performed with biotinylated probes, synthesized by nick translation, and a signal amplification system based onfluorescein-labeled tyramide as described previously (26). White arrowheads point at cells in which viral DNA was detected. At 24 and 48 hpostinfection, only very low levels of adenovirus DNA were detected, but by 72 h, some cells with very high viral DNA content were observed.White arrows point at the basement membrane. (B) Adenovirus E1 (nucleotides [nt] 1 to 532), E2 early (nt 26986 to 27336), and E4 (nt 35581to 35935) promoters were each blunted and cloned at the NruI site in pLN-lacZ (27), a derivative of vector pLNSX (21). The adenoviral promoterswere each placed upstream of the �-galactosidase gene but downstream of the LTR-driven neomycin resistance gene. (Top) Amphotropicretroviruses were generated as described previously (27) and used to infect primary human keratinocytes to make raft cultures as describedpreviously (41). The raft cultures were fixed, stained with X-Gal (5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside), embedded, and sectioned.The sections were lightly counterstained with hematoxylin. (Bottom) The keratinocytes were first infected with pBabePur-18URR-E7 (26)retrovirus and selected in puromycin-containing medium before infection with the pLN-derived retroviruses. Black arrows point at the basalmembrane.
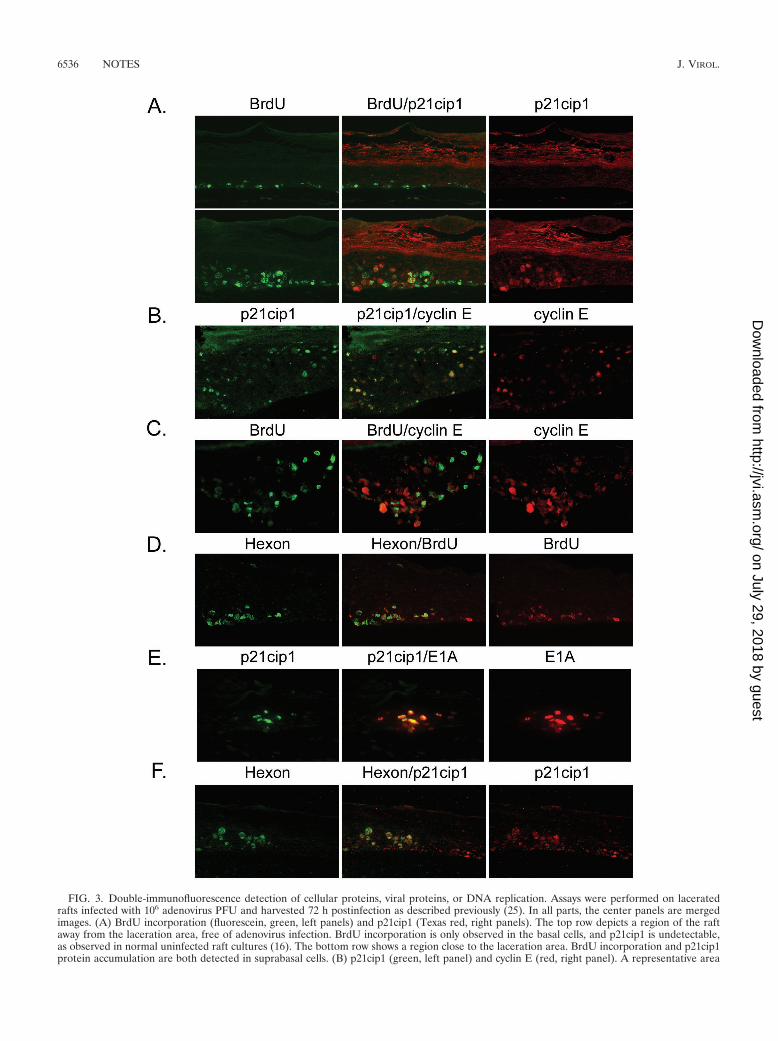
FIG. 3. Double-immunofluorescence detection of cellular proteins, viral proteins, or DNA replication. Assays were performed on laceratedrafts infected with 106 adenovirus PFU and harvested 72 h postinfection as described previously (25). In all parts, the center panels are mergedimages. (A) BrdU incorporation (fluorescein, green, left panels) and p21cip1 (Texas red, right panels). The top row depicts a region of the raftaway from the laceration area, free of adenovirus infection. BrdU incorporation is only observed in the basal cells, and p21cip1 is undetectable,as observed in normal uninfected raft cultures (16). The bottom row shows a region close to the laceration area. BrdU incorporation and p21cip1protein accumulation are both detected in suprabasal cells. (B) p21cip1 (green, left panel) and cyclin E (red, right panel). A representative area
added to the cultures by one of three routes. From below, 106
PFU was mixed with the culture media. From above, 106 PFUsuspended in 40 �l of phosphate-buffered saline (PBS) wasplaced on top of stratified epithelial cultures. Alternatively, theculture was first lacerated with a scalpel before application of106 PFU in 40 �l of PBS from above. The raft cultures wereformalin fixed, paraffin embedded, and cut into 4-�m sections.To reveal the CPE caused by adenovirus, the sections werestained with hematoxylin and eosin. Additional sections wereprobed for the expression of the early protein E1A and the lateprotein hexon by double immunofluorescence. Only culturesthat were lacerated showed signs of infection in regions nearthe laceration (Fig. 1A). Apparently, the virus neither is capa-ble of crossing the collagen matrix underneath the epitheliumnor can penetrate the intact cornified layer that protects thesurface of the tissue. It has been recently demonstrated thatCAR lies below the tight junction seal in adherens junctions(37), in agreement with the notion that a wound in the epithe-lium is necessary to initiate the infection process.
At a high MOI, infection begins in the CAR-positive basaland parabasal layers. In the lacerated raft cultures infectedwith 106 PFU, we observed the expression of E1A 24 h postin-fection in the areas surrounding the cuts (Fig. 1A). By 48 hpostinfection, the expression was stronger and localized to thebasal cells of the raft. Hexon also became detectable at thistime, as well as a fairly mild CPE. By 72 h, E1A expression waspreferentially found in the mid-spinous cells, just above a re-gion strongly positive for hexon. The CPE was more wide-spread, and empty spaces between the epithelium and thedermal equivalent were also observed. This space could havebeen generated when infected basal cells were lysed or de-tached from the collagen. By 96 h, the whole thickness of theepithelium in the area of laceration was affected. E1A was stilldetectable in cell layers above those positive for hexon. Most ofthe basal and parabasal cells were lysed by then (Fig. 1A).
It is interesting that cells weakly positive for hexon were alsoweakly positive for E1A. In contrast, most cells strongly posi-tive for E1A were negative for hexon, and, conversely, cellsstrongly positive for hexon were negative for E1A. We suggestthat these signal patterns reflect the progression of the infec-tion process. At early times, E1A was strong, and there was nohexon (E1A�/hexon�). E1A expression started to declinewhen the late phase initiated (weakly positive for both). Fi-nally, E1A expression was below detection, while hexon be-came highly abundant (E1A�/hexon�). It is likely that, at thislate time, many cells were already dead and only the virusparticles were stable, but other viral proteins were degraded.Paracrystalline arrays of adenovirus particles were observed inthe nuclei of infected cells by electron microscopy of ultrathinsections of fixed tissues (Fig. 1B). From these experiments, it
was quite clear that the primary infection of adenovirus takesplace in the basal layer of the raft and extends upwards withtime.
To investigate the reason for the preferential infection of thebasal and parabasal cells early in the infection, we determinedthe expression pattern of CAR. By using immunohistochemis-try of sections of normal raft cultures, we detected CAR mainlyin the basal and parabasal cell layers (Fig. 1C, top panel), inagreement with a recent report on organotypic cultures oforopharyngeal cells and airway epithelia (15, 37). These resultswould explain, at least in part, the observed preference towardsthese cell layers during the initial infection. In the negativecontrol, CAR was not detected in raft cultures of an HPV-associated cervical carcinoma cell line, SiHa (Fig. 1C). Inter-estingly, submerged cultures of SiHa cells are susceptible toadenovirus infection, but SiHa raft cultures were refractory toinfection even after laceration (N. S. Banerjee, L. T. Chow andT. R. Broker, unpublished observations).
In our time course experiments, we observed that after a fewdays, the infected cells were detected in the spinous and eventhe granular strata. It is not clear how the virus enters thesesuprabasal cells that are low on or devoid of CAR. One pos-sibility is that progeny virus is transmitted to neighboring cellsthrough gap junctions. Using electron microscopy, we exam-ined numerous gap junctions, but we did not capture any viralparticles in close proximity to them. This mode of transmissionhas not been described in monolayer cultures either (39). Thesecond possibility is that the local high concentration of prog-eny virus released from the first round of infection makespossible the infection of surrounding cells in the presence oflow CAR or even in the absence of CAR (2, 8, 11). Alterna-tively, in a fraction of the basal and parabasal cells, the virusmay have remained inactive after gaining entry. Only when theinfected cells or their daughter cells have ascended the epithe-lium and become differentiated does the virus enter into theproductive phase. This possibility is consistent with the CPEfound throughout the entire stratified epithelium when kera-tinocytes were infected with the wild-type virus prior to thedevelopment of raft cultures (3). Had the infection progressedinto the late phase in all basal cells, there would not have beena stratified epithelium. This was not the case. This possibility ofa delayed activation, as reflected by the progression of the CPEin Fig. 1A, is consistent with the observation that PHKs mi-grate up to three cell layers in 48 h under these culture conditions(6). Indeed, a differentiation-dependent virus production wouldbe similar to the case with HPVs, the productive phase of whichis strictly dependent on squamous differentiation (33).
At a low MOI, early signs of adenovirus infection are onlyfound in the suprabasal cells after a lag. To investigate thepossibility that the productive phase of adenovirus infection is
is presented. Most p21cip1-positive cells are also positive for cyclin E. (C) BrdU incorporation (green, left panel) and cyclin E (Texas red, rightpanel). A region close to the laceration area is shown. High levels of cyclin E negatively correlate with high levels of BrdU incorporation, asobserved previously for HPV (17). (D) Hexon (green, left panel) and BrdU (red, right panel). Hexon was detected as described in the legend toFig. 1. BrdU was probed with a mouse monoclonal antibody (1:50 dilution [Zymed Laboratories, Inc., South San Francisco, Calif.]) followed bya 1:75 dilution of biotinylated anti-mouse IgG (Vector Laboratories, Burlingame, Calif.) and Texas red-streptavidin (Vector Laboratories).(E) p21cip1 (green, left panel) and E1A (red, right panel). E1A was detected first as described in the legend to Fig. 1A. The slide was then probedwith anti-p21cip antibody directly conjugated with fluorescein (OP64F; dilution, 1:50 [Oncogene]). (F) Hexon (green, left panel) and p21cip1 (red,panel). The experiment was similar to that described for panel D, but p21cip1 was detected with a monoclonal antibody (OP64, dilution 1:25[Oncogene]).
linked to differentiation, we repeated the infections of raftcultures with a 100-fold-lower MOI. It has been reported thatat a high MOI, the gene expression program of the adenovirusis distorted and early promoters are derepressed (24, 32), thusobscuring a potentially inactive state.
We infected lacerated raft cultures with 104 PFU as de-scribed above. Nine-day-old rafts were harvested at 24, 48, and72 h postinfection. Paraffin-embedded sections were probedwith E1A and hexon by double immunofluorescence. As shownin Fig. 2A, neither viral protein was detectable at 24 and 48 hpostinfection. At 72 h postinfection, both proteins were de-tected in a small number of cells, mostly in the lower andmidspinous strata. In situ hybridization of adenovirus DNArevealed very weak isolated signals in the basal and parabasallayers at 24 h postinfection (Fig. 2A, bottom panel). The iso-lated signals became stronger and clearly suprabasal at 72 hpostinfection, consistent with viral DNA amplification andhexon expression. Thus, at the low MOI, fewer cells wereinfected and infection proceeded much more slowly; however,eventually late genes were expressed.
Taken together, these observations support the interpreta-tion that the virus preferentially infects the basal and parabasalcells but restricts its productive replication until the cellprogresses in its differentiation program and enters the spinouslayers. However, we cannot rule out the possibility that viralpromoter activation and viral DNA amplification initiated inthe basal or parabasal cells, but given the low MOI, the levelsof viral protein and DNA were below the sensitivity of detec-tion. By the time the amounts of protein and DNA weredetectable, the cells had ascended through the stratified epi-thelium.
Activities of E1 and E2a early promoters are differentiationdependent. To distinguish the possible interpretations dis-cussed above, we conducted the following experiments to ex-amine the viral promoter activities. Productive viral DNA am-plification requires the orderly expression of the early proteinsencoded by the E1A, E1B, E2, and E4 open reading frames(reviewed in reference 31). We cloned the �-galactosidasereporter gene under the control of the adenoviral promotersE1, E2 early (at coordinate 75), and E4. These reporter cas-settes were introduced into the raft cultures by using a retro-virus vector that expresses the bacterial neomycin resistancegene. Submerged PHKs were infected with these retroviruses,selected with G418 for 2 days, and the collagen matrix wasseeded with them. Usually, 50 to 70% of the cells survived theselection. We have previously demonstrated that in this setting,the presence of the retroviral LTR does not affect the activitiesof viral or cellular promoters in driving the �-galactosidasegene expression in the appropriate cell strata (27, 40, 41). Inaddition, a myc-tagged human PCNA gene introduced via alentivirus also exhibits an expression pattern similar to that ofthe endogenous gene. Furthermore, both genes were activatedin differentiated cells by the HPV-18 E7 gene driven by thedifferentiation-dependent HPV-18 enhancer promoter locatedin the upstream regulation region (HPV-18 URR-E7) (26).
The results of these experiments are shown in Fig. 2B (toprow). For all three promoters, the reporter activity was ob-served only in the differentiated strata, with E4 being moresporadically distributed than the others. The E1 promoter be-came active in the lower spinous cells, while the E2 early
promoter became active in the mid-spinous layers. Both pro-moters remained active throughout the upper layers of the raft.To verify that the retrovirus backbone did not affect the normalregulation of the promoters, we also transduced the PHKs withHPV-18 URR-E7 via a retrovirus. In monolayer cell lines, E7transactivates the E2 early promoter through deregulation ofthe pRb/E2F pathway (4, 28). Upon E7 gene expression, theE2 early promoter expression became more widespread, be-ginning in the lower spinous layer (Fig. 2B, bottom row), co-inciding with the expression pattern of the HPV-18 URR (27).In contrast, E7 had no effect on the reporter expression patternof the E1 or E4 promoters. We were not able to test theactivation of E2 early promoter by adenovirus E1A protein.We suspect that PHKs transduced with a retrovirus harboringan E1A expression cassette either expressed little or no E1Aprotein or underwent apoptosis, as reported previously in pri-mary mouse fibroblasts (38).
From these observations, we conclude that the differentia-tion-dependent replication exhibited by adenovirus in the raftcultures is due, at least in part, to the differentiation-depen-dent activity of its early promoters. Particularly, in the case ofE1, an AP3-like factor has been identified as being responsiblefor the inhibition of the promoter in undifferentiated cells (13).However, information on the function of this factor in unin-fected cells is still lacking. The constitutive expression of theearly promoters in differentiated cells is an important factor totake into account during the design of oncolytic vectors thatrely on the abrogation of E1A functions to modulate theiractivities. The fact that the E2 early and E4 promoters are stillactive even in the absence of E1A may account in part for theability of altered viruses, such as CB016, to retain the ability toreplicate in normal epithelium (3).
Levels of p21cip1 and cyclin E are increased in adenovirus-infected cells. We have previously shown that HPV inducesS-phase reentry in a fraction of the differentiated cells in thesquamous epithelium in patient specimens. HPV-18 URR-E7is sufficient for this induction in the raft culture setting (5).However, HPV-18 URR-E7 also induces a host defense re-sponse in a separate population of differentiated keratinocytes.This involves the inhibition of cyclin E/cdk2 in a complex withthe cdk2 inhibitor, p27kip1 or p21cip1 (25). In normal squa-mous epithelium and in epithelial raft cultures, p21cip1 mRNAis up-regulated in differentiated cells; however, the p21cip1protein is quickly degraded and is below the level of detectionin situ (16, 30). Its level rises dramatically along with that ofcyclin E in a subset of the differentiated cells of warty patientlesions or of HPV-18 URR-E7-transduced raft cultures. Noaccumulation is observed in the basal strata. Differentiatedcells that exhibit this abnormal accumulation are unable toreenter S phase, as revealed by the lack of bromodeoxyuridine(BrdU) incorporation. This stabilization of cyclin E and p21cipproteins has been attributed to the differentiation-induced sta-ble expression of the p27kip1 protein, which sequesters E7-induced cyclin E into a stable inactive complex with cdk2. Inturn, cyclin E then stabilizes p21cip1 protein in an analogouskinase-inactive complex (25). Since HPV DNA replication alsodepends on an active cyclin E/cdk2 (19, 20), viral DNA isunable to amplify in cells that stably accumulate p21cip1 andcyclin E (17).
We asked whether this cellular response could also be ob-
served during adenovirus DNA infection and whether adeno-virus could overcome it. To answer these questions, we exam-ined raft cultures infected with adenovirus for 3 days at a highMOI. In the last 12 h prior to harvesting, the cultures wereexposed to BrdU. In a series of double immunofluorescencestudies, we probed for p21cip1, cyclin E, or BrdU. As reportedpreviously, neither protein was detected in the mock-infectedraft cultures, and BrdU incorporation was restricted to thebasal layer of the epithelium. Similar observations were madein the areas away from infection sites in the lacerated raftcultures (Fig. 3A, top row). In contrast, high levels of p21cip1were found in and around the regions that show evidence ofCPE (Fig. 3A, bottom row). Cells that showed high levels ofp21cip1 also had high levels of cyclin E (Fig. 3B). BrdU wasalso detected in suprabasal cells in areas of CPE. However,unlike the HPV-host interactions, a significant fraction of cellswere positive for both p21cip1 and BrdU in the lower spinouscells (Fig. 3A, bottom row). Similarly, some of the cyclin E-positive cells were also positive for BrdU (Fig. 3C). Thus, DNAreplication took place in some of the cells with high levels ofcyclin E and p21cip1.
To determine whether the BrdU incorporation represented,at least in part, viral DNA replication, we carried out hexonand BrdU double immunofluorescence, because hexon is ex-pressed late in the infection after the initiation of viral DNAreplication. As shown in Fig. 3D, BrdU incorporation wasobserved in unaffected basal cells (the right side of the panel)as well as in basal cells positive for hexon (the left side of theleft panel). However, suprabasal BrdU incorporation oftencoincided with hexon expression (the left side of the panel).Thus, viral DNA had replicated in these cells.
To investigate the kinetics of p21cip1 induction, we per-formed double immunofluorescence for E1A and p21cip1 andfor hexon and p21cip1. A fraction of cells strongly positive forE1A were also positive for p21cip1, whereas those weaklypositive for E1A were negative for p21cip1 (Fig. 3E). All cellspositive for hexon were also positive for p21cip1, but somep21cip1-positive cells were negative for hexon (Fig. 3F). Takentogether, these data indicate that p21cip1 appears at or shortlyafter E1A is expressed and is maintained throughout the re-mainder of the adenovirus life cycle.
Collectively, these results suggest that as in the case of HPV,the response against unscheduled DNA synthesis in postmi-totic differentiated cells mediated by p27kip1 and p21cip1 isalso triggered by the adenovirus infection. However, this mech-anism seems less effective than in the case of HPV. Althougha number of p21cip1-positive cells were negative for BrdUincorporation and hexon, indicative of a lack of viral amplifi-cation, many p21cip1- and cyclin E-positive cells were positivefor BrdU and hexon. Evidently, adenovirus is better equippedthan HPV to overcome this cellular response. For example,adenovirus carries its own DNA polymerase, terminal protein,and single-stranded DNA binding protein to support viralDNA replication. Currently, there is no information onwhether adenovirus needs an active cyclin E/cdk2 for efficientviral replication. Nevertheless, these results stress the fact thatthe stable expression of p27kip1 protein and the abundance ofp21cip1 mRNA in the differentiated keratinocytes (16, 25)serve the purpose of attempting to prevent nonscheduled DNAreplication in differentiated cells. As such, they act as an anti-
viral mechanism to temper infections. Clearly, this scheme isquite successful during HPV infection, but is less effectiveduring adenovirus infection.
In conclusion, we have shown that epithelial raft cultures aresuitable tools for the study of the different aspects of theadenovirus infectious cycle. First, we have shown that the cor-nified layer effectively prevents adenovirus infection, andwounding of the basal and parabasal strata is required to effectan infection. Second, we have shown that the primary targets ofAd5 are the basal and parabasal cells, where CAR is expressed,in agreement with the results obtained with oropharyngealcells (15). Third, at low MOI, the virus may remain in a rela-tively inactive state until the infected cell or its daughter celldifferentiates and enters the spinous layers. It is in these dif-ferentiated layers where the viral early promoters are activatedand the productive viral DNA amplification takes place. Thiswould explain why this virus encodes gene products that canreestablish the S-phase milieu in cells that are postmitotic or inG0 (35). In contrast, at high MOI, the viral gene expressionprogram is altered. Expression of early genes occurs right afterinfection, bypassing the lag period. Thus, viral amplificationtakes place in the basal and parabasal cells. Fourth, the hostresponds to the infection by accumulating p21cip1 and cyclin Ein a way resembling the response to another small DNA tumorvirus, HPV, although this strategy is much less effective foradenovirus. In summary, this work introduces new tools andconcepts to the study of the basic biology of adenovirus in thebelief that a better understanding of how the virus infects andinteracts with its host cells will greatly improve gene deliverymethods and help in the design of new, clinically safer vectors.
We thank Ge Jin for tissue embedding and sectioning, Brenda Gos-sage for technical help in cloning of the adenoviral promoters, and thenurses in the neonatal nursery of Cooper Green Hospital for collectingforeskins.
This work was supported by Public Health Service grants CA 36200,CA94084, P50 CA83591, CA83821, DAMD 17-00-1-0002, andCA93976 and by support from the Lustgarten Foundation and SusanG. Komen Foundation. The Digital Imaging Microscopy Facility wasestablished with funds provided in large measure by the UAB HealthServices Foundation and by grant DE/CA11910.
REFERENCES
1. Alemany, R., C. Balague, and D. T. Curiel. 2000. Replicative adenovirusesfor cancer therapy. Nat. Biotechnol. 18:723–728.
2. Alemany, R., and D. T. Curiel. 2001. CAR-binding ablation does not changebiodistribution and toxicity of adenoviral vectors. Gene Ther. 8:1347–1353.
3. Balague, C., F. Noya, R. Alemany, L. T. Chow, and D. T. Curiel. 2001.Human papillomavirus E6E7-mediated adenovirus cell killing: selectivity ofmutant adenovirus replication in organotypic cultures of human keratino-cytes. J. Virol. 75:7602–7611.
4. Chellappan, S., V. B. Kraus, B. Kroger, K. Munger, P. M. Howley, W. C.Phelps, and J. R. Nevins. 1992. Adenovirus E1A, simian virus 40 tumorantigen, and human papillomavirus E7 protein share the capacity to disruptthe interaction between transcription factor E2F and the retinoblastomagene product. Proc. Natl. Acad. Sci. USA 89:4549–4553.
5. Cheng, S., D.-C. Schmidt-Grimminger, T. Murant, T. R. Broker, and L. T.Chow. 1995. Differentiation-dependent up-regulation of the human papillo-mavirus E7 gene reactivates cellular DNA replication in suprabasal differ-entiated keratinocytes. Genes Dev. 9:2335–2349.
6. Chien, W.-M., F. Noya, H. M. Benedict-Hamilton, T. R. Broker, and L. T.Chow. 2002. Alternative fates of keratinocytes transduced by human papil-lomavirus type 18 E7 during squamous differentiation. J. Virol. 76:2964–2972.
7. Chow, L. T., and T. R. Broker. 1997. In vitro experimental systems for HPV:epithelial raft cultures for investigations of viral reproduction and pathogen-esis and for genetic analyses of viral proteins and regulatory sequences. Clin.Dermatol. 15:217–227.
8. Dechecchi, M. C., P. Melotti, A. Bonizzato, M. Santacatterina, M. Chilosi,
and G. Cabrini. 2001. Heparan sulfate glycosaminoglycans are receptorssufficient to mediate the initial binding of adenovirus types 2 and 5. J. Virol.75:8772–8780.
9. DeWeese, T. L., H. van der Poel, S. Li, B. Mikhak, R. Drew, M. Goemann, U.Hamper, R. DeJong, N. Detorie, R. Rodriguez, T. Haulk, A. M. DeMarzo, S.Piantadosi, D. C. Yu, Y. Chen, D. R. Henderson, M. A. Carducci, W. G.Nelson, and J. W. Simons. 2001. A phase I trial of CV706, a replication-competent, PSA selective oncolytic adenovirus, for the treatment of locallyrecurrent prostate cancer following radiation therapy. Cancer Res. 61:7464–7472.
10. Dollard, S. C., J. L. Wilson, L. M. Demeter, W. Bonnez, R. C. Reichman,T. R. Broker, and L. T. Chow. 1992. Production of human papillomavirus andmodulation of the infectious program in epithelial raft cultures. Genes Dev.6:1131–1142.
11. Einfeld, D. A., R. Schroeder, P. W. Roelvink, A. Lizonova, C. R. King, I.Kovesdi, and T. J. Wickham. 2001. Reducing the native tropism of adeno-virus vectors requires removal of both CAR and integrin interactions. J. Vi-rol. 75:11284–11291.
12. Graham, F. L., and L. Prevec. 1991. Manipulation of adenovirus vectors.Methods Mol. Biol. 7:109–128.
13. Herbst, R. S., M. Pelletier, E. M. Boczko, and L. E. Babiss. 1990. The stateof cellular differentiation determines the activity of the adenovirus E1Aenhancer element: evidence for negative regulation of enhancer function.J. Virol. 64:161–172.
14. Huber, M., A. Limat, E. Wagner, and D. Hohl. 2000. Efficient in vitrotransfection of human keratinocytes with an adenovirus-enhanced receptor-mediated system. J. Investig. Dermatol. 114:661–666.
15. Hutchin, M. E., R. J. Pickles, and W. G. Yarbrough. 2000. Efficiency ofadenovirus-mediated gene transfer to oropharyngeal epithelial cells corre-lates with cellular differentiation and human coxsackie and adenovirus re-ceptor expression. Hum. Gene Ther. 11:2365–2375.
16. Jian, Y., D.-C. Schmidt-Grimminger, W.-M. Chien, X. Wu, T. R. Broker, andL. T. Chow. 1998. Post-transcriptional induction of p21cip1 protein by hu-man papillomavirus E7 inhibits unscheduled DNA synthesis reactivated indifferentiated keratinocytes. Oncogene 17:2027–2038.
17. Jian, Y., B. A. Van Tine, W.-M. Chien, G. M. Shaw, T. R. Broker, and L. T.Chow. 1999. Concordant induction of cyclin E and p21cip1 in differentiatedkeratinocytes by the human papillomavirus E7 protein inhibits cellular andviral DNA synthesis. Cell Growth Differ. 10:101–111.
18. Laporta, R. F., and L. B. Taichman. 1981. Adenovirus type-2 infection ofhuman keratinocytes: viral expression dependent upon the state of cellularmaturation. Virology 110:137–146.
19. Lin, B.-Y., T. Ma, J.-S. Liu, S.-R. Kuo, L. Jin, T. R. Broker, J. W. Harper, andL. T. Chow. 2000. HeLa cells are phenotypically limiting in cyclin E/CDK2for efficient human papillomavirus DNA replication. J. Biol. Chem. 275:6167–6174.
20. Ma, T., N. Zou, B.-Y. Lin, L. T. Chow, and J. W. Harper. 1999. Interactionbetween cyclin-dependent kinases and human papillomavirus replication-initiation protein E1 is required for efficient viral replication. Proc. Natl.Acad. Sci. USA 96:382–387.
21. Miller, A. D., and G. J. Rosman. 1989. Improved retroviral vectors for genetransfer and expression. BioTechniques 7:980–982.
22. Nelson, C. C., A. W. Braithwaite, M. Silvestro, and A. J. Bellett. 1990.E1a-dependent expression of adenovirus genes in OTF963 embryonal car-cinoma cells: role of E1a-induced differentiation. Proc. Natl. Acad. Sci. USA87:8041–8045.
23. Nemunaitis, J., I. Ganly, F. Khuri, J. Arseneau, J. Kuhn, T. McCarty, S.Landers, P. Maples, L. Romel, B. Randlev, T. Reid, S. Kaye, and D. Kirn.2000. Selective replication and oncolysis in p53 mutant tumors with ONYX-
015, an E1B-55kD gene-deleted adenovirus, in patients with advanced headand neck cancer: a phase II trial. Cancer Res. 60:6359–6366.
24. Nevins, J. R. 1981. Mechanism of activation of early viral transcription by theadenovirus E1A gene product. Cell 26:213–220.
25. Noya, F., W.-M. Chien, T. R. Broker, and L. T. Chow. 2001. p21cip1 degra-dation in differentiated keratinocytes is abrogated by costabilization withcyclin E induced by human papillomavirus E7. J. Virol. 75:6121–6134.
26. Noya, F., W.-M. Chien, X. Wu, N. S. Banerjee, J. C. Kappes, T. R. Broker,and L. T. Chow. 2002. The promoter of the human proliferating cell nuclearantigen gene is not sufficient for cell cycle-dependent regulation in organo-typic cultures of keratinocytes. J. Biol. Chem. 277:17271–17280.
27. Parker, J. N., W. Zhao, K. J. Askins, T. R. Broker, and L. T. Chow. 1997.Mutational analyses of differentiation-dependent human papillomavirus type18 enhancer elements in epithelial raft cultures of neonatal foreskin kera-tinocytes. Cell Growth Differ. 8:751–762.
28. Phelps, W. C., C. L. Yee, K. Munger, and P. M. Howley. 1988. The humanpapillomavirus type 16 E7 gene encodes transactivation and transformationfunctions similar to those of adenovirus E1A. Cell 53:539–547.
29. Scheffner, M., B. A. Werness, J. M. Huibregtse, A. J. Levine, and P. M.Howley. 1990. The E6 oncoprotein encoded by human papillomavirus types16 and 18 promotes the degradation of p53. Cell 63:1129–1136.
30. Schmidt-Grimminger, D.-C., X. Wu, Y. Jian, T. R. Broker, and L. T. Chow.1998. Post-transcriptional induction of p21cip1 protein in condylomata anddysplasias is inversely related to human papillomavirus activities. Am. J.Pathol. 152:1015–1024.
31. Shenk, T. E. 2001. Adenoviridae: the viruses and their replication, p. 1053–1088. In D. M. Knipe and P. M. Howley (ed.), Fundamental virology, 4th ed.Lippincott, Williams & Wilkins, Philadelphia, Pa.
32. Shenk, T. E., N. Jones, W. Colby, and D. Fowlkes. 1980. Functional analysisof adenovirus-5 host-range deletion mutants defective for transformation ofrat embryo cells. Cold Spring Harbor Symp. Quant. Biol. 44:367–375.
33. Stoler, M. H., and T. R. Broker. 1986. In situ hybridization detection ofhuman papillomavirus DNAs and messenger RNAs in genital condylomasand a cervical carcinoma. Hum. Pathol. 17:1250–1258.
34. Taichman, L. B., S. S. Reilly, and R. F. LaPorta. 1983. The role of keratin-ocyte differentiation in the expression of epitheliotropic viruses. J. Investig.Dermatol. 81:137s–140s.
35. Tiainen, M., D. Spitkovsky, P. Jansen-Durr, A. Sacchi, and M. Crescenzi.1996. Expression of E1A in terminally differentiated muscle cells reactivatesthe cell cycle and suppresses tissue-specific genes by separable mechanisms.Mol. Cell. Biol. 16:5302–5312.
36. van den Heuvel, S. J., T. van Laar, W. M. Kast, C. J. Melief, A. Zantema, andA. J. van der Eb. 1990. Association between the cellular p53 and the ade-novirus 5 E1B-55kd proteins reduces the oncogenicity of Ad-transformedcells. EMBO J. 9:2621–2629.
37. Walters, R. W., P. Freimuth, T. O. Moninger, I. Ganske, J. Zabner, and M. J.Welsh. 2002. Adenovirus fiber disrupts CAR-mediated intercellular adhe-sion allowing virus escape. Cell 110:789–799.
38. White, E., R. Cipriani, P. Sabbatini, and A. Denton. 1991. Adenovirus E1B19-kilodalton protein overcomes the cytotoxicity of E1A proteins. J. Virol.65:2968–2978.
39. Wigand, R., and G. Kumel. 1977. The kinetics of adenovirus infection andspread in cell cultures infected with low multiplicity. Arch. Virol. 54:177–187.
40. Zhao, W., L. T. Chow, and T. R. Broker. 1997. Transcription activities ofhuman papillomavirus type 11 E6 promoter-proximal elements in raft andsubmerged cultures of foreskin keratinocytes. J. Virol. 71:8832–8840.
41. Zhao, W., F. Noya, W. Y. Chen, T. M. Townes, L. T. Chow, and T. R. Broker.1999. Trichostatin A up-regulates human papillomavirus type 11 upstreamregulatory region-E6 promoter activity in undifferentiated primary humankeratinocytes. J. Virol. 73:5026–5033.