Page 1
R-State AMP Complex Reveals Initial Steps of the Quaternary Transition
of Fructose-1,6-bisphosphatase‡
Cristina V. Iancu, Susmith Mukund, Herbert J. Fromm,
and Richard B. Honzatko*
Department of Biochemistry, Biophysics, and Molecular Biology,
Iowa State University, Ames, IA 50011
Running title: Fructose-1,6-bisphosphatase
This work was supported in part by National Institutes of Health Research Grant NS 10546.
‡Coordinates and structure factors (accession labels 1YXI, 1YYZ and 1YZO) for the structures
described in this paper have been deposited in the Protein Data Bank, Research Collaboratory for
Structural Bioinformatics (RCSB).
*Corresponding author. Telephone: (515) 294-6116. Fax: (515) 294-0453. E-mail:
[email protected] .
1
JBC Papers in Press. Published on March 14, 2005 as Manuscript M501011200
Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 2
Abstract
AMP transforms fructose-1,6-bisphosphatase from its active R-state to its inactive T-state;
however, the mechanism of that transformation is poorly understood. The mutation of Ala54 to
leucine destabilizes the T-state of fructose-1,6-bisphosphatase. The mutant enzyme retains wild-
type levels of activity, but the concentration of AMP that causes 50% inhibition increases 50-
fold. In the absence of AMP, the Leu54 enzyme adopts an R-state conformation nearly identical
to that of the wild-type enzyme. The mutant enzyme, however, grows in two crystal forms in the
presence of saturating AMP. In one, the AMP-bound tetramer is in a T-like conformation,
whereas in the other, a R-like conformation. The latter reveals conformational changes in two
helices due to the binding of AMP. Helix H1 moves toward the center of the tetramer and
displaces Ile10 from a hydrophobic pocket. The displacement of Ile10 exposes a hydrophobic
surface critical to interactions that stabilize the T-state. Helix H2 moves away from the center of
the tetramer, breaking hydrogen bonds with a buried loop (residues 187–195) in an adjacent
subunit. The same hydrogen bonds reform, but only after the quaternary transition to the T-state.
Proposed here is a model that accounts for the quaternary transition and cooperativity in the
inhibition of catalysis by AMP.
2
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 3
Fructose-1,6-bisphosphatase (D-fructose-1,6-bisphosphate 1-phosphohydrolase, EC 3.1.3.11;
FBPase1) catalyzes a tightly regulated step of gluconeogenesis, the hydrolysis of fructose 1,6-
bisphosphate (F16P2) to fructose 6-phosphate (F6P) and inorganic phosphate (Pi) (1,2). AMP
and fructose 2,6-bisphosphate (F26P2), binding to allosteric and active sites, respectively, inhibit
FBPase while simultaneously activating its counterpart in glycolysis, fructose-6-phosphate 1-
kinase (3,4). Biosynthesis and degradation of F26P2 is subject to hormonal control principally
by glucagon and insulin (4,5). F26P2 enhances the binding of AMP to FBPase by up to an order
of magnitude (6). Hence, although intracellular concentrations of AMP remain relatively
constant, AMP should become a more potent inhibitor of FBPase as concentrations of F26P2
increase. AMP binds 28 Å away from the nearest active site, and perhaps not surprisingly
inhibits catalysis noncompetitively with respect to F16P2. Yet, AMP is a competitive inhibitor of
catalysis with respect to essential divalent cations (Mg2+, Mn2+, or Zn2+), all of which are in
proximity to (and probably coordinate with) the 1-phosphoryl group of F16P2 (7−10).
FBPase is a homotetramer [subunit Mr of 37,000 (11)] and exists in at least two distinct
quaternary conformations called R and T (12−13). AMP induces the transition from the active
R-state to the inactive (or less active) T-state. Substrates or products in combination with metal
cations stabilize the R-state conformation. A proposed mechanism for allosteric regulation of
catalysis involves three conformational states of loop 52−72 called engaged, disengaged, and
disordered (14). AMP alone or with F26P2, stabilizes a disengaged loop (15,16), whereas metals
with products stabilize an engaged loop (10,16−18). In active forms of the enzyme, loop 52−72
probably cycles between its engaged and disordered conformations (14,17). Fluorescence from a
tryptophan reporter group at position 57 is consistent with the conformational states for loop
52−72, observed in crystal structures (19,20). Thus far, the engaged conformation of loop 52−72
3
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 4
has appeared only in R-state crystal structures, and the disengaged conformer only in T-state
structures; however, disordered conformations of the dynamic loop have appeared in both the R-
and T-states (16,17,21,22).
The precise sequence of events that attend the R- to T-state transition in FBPase has been
elusive. Crystal structures of the R- and T-states are the endpoints of the allosteric transition,
and leave much to speculation regarding intermediate conformational states of FBPase. The
immediate consequences of AMP binding to the R-state are unknown. Does the dynamic loop
become disengaged in response to the binding of AMP or in response to the allosteric transition
to the T-state? How does the binding of AMP destabilize the R-state and stabilize the T-state?
A recent study revealed the first intermediate state of porcine FBPase (23), a T-like conformation
due to the binding of an allosteric effector to the center of the tetramer. The results of that study
indicated the potential for trapping intermediate conformational states of FBPase by
crystallization.
The mutation of Ala54 to leucine disrupts key packing interactions of the disengaged loop
conformation. The resulting Leu54 enzyme has wild-type catalytic properties and retains
cooperativity in AMP inhibition, but exhibits a 50-fold increase in the IC50 for AMP. In the
absence of AMP, Leu54 FBPase is in its canonical R-state. Two crystal forms, however, grow in
the presence of saturating AMP. In one crystal form, the enzyme is in its T-state, but with a
disordered dynamic loop. In the other crystal form, the tetramer is in an R-like quaternary state
with an engaged dynamic loop. The latter crystal form reveals the immediate consequences of
AMP association in the absence of an allosteric transition. The observed conformational changes
suggest the mechanism by which AMP leverages the allosteric transition in FBPase.
4
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 5
Experimental
Materials F16P2, F26P2, NADP+ and AMP were purchased from Sigma. DNA-modifying and
restriction enzymes, T4 polynucleotide kinase and ligase were from Promega. Glucose-6-
phophate dehydrogenase and phosphoglucose isomerase came from Roche. Other chemicals
were of reagent grade or equivalent. Escherichia coli strains BMH 71-18 mutS and XL1-Blue
came from Clontech and Stratagene, respectively. The FBPase-deficient E. coli strain DF 657
came from the Genetic Stock Center at Yale University.
Mutagenesis of wild-type FBPase— The mutation of Ala54 to leucine was accomplished by
specific base changes in a double-stranded plasmid containing the gene coding for FBPase using
the Transformer site-Directed Mutagenesis kit (Clontech). The mutagenic primer for
Ala54→Leu was 5'GGCGGGCATCCTGCACCTC3'. (The codon with the point mutation is
underlined in bold typeface). The selection primer for mutagenesis,
5’CAGCCTCGCCTCGAGAACGCCA3’ (digestion site underlined in bold typeface), changed
an original NruI site on the plasmid into a XhoI site. The mutation and the integrity of the
construct were confirmed by sequencing the promoter region and the entire open reading frame.
The Iowa State University sequencing facility provided DNA sequences, using the fluorescent
dye-dideoxy terminator method.
Expression and Purification of wild-type and Leu54 FBPases Cell-free extracts of wild-type
and Leu54 FBPases were subjected to heat treatment (63o C for 7 minutes), followed by
centrifugation. The supernatant solution was loaded onto a Cibracon Blue sepharose column,
previously equilibrated with 20 mM Tris-HCl, pH 7.5. The column was washed first with 20
mM Tris-HCl, pH 7.5. Enzyme was eluted with a solution of 500 mM NaCl and 20 mM Tris-
HCl of the same pH. After pressure concentration (Amicon PM-30 membrane) and dialysis
5
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 6
against 10 mM Tris-HCl, pH 8.0, the protein sample was loaded onto a DEAE sepharose column
equilibrated with 10 mM Tris-HCl, pH 8.0. Purified enzyme was eluted with a NaCl gradient
(0–0.5 M) in 10 mM Tris-HCl, pH 8.0, and then dialyzed extensively against 50 mM Hepes, pH
7.4, for kinetic investigations and for crystallization experiments. Purity and protein
concentrations of FBPase preparations were confirmed by SDS-polyacrylamide gel
electrophoresis (25) and the Bradford assay (26), respectively.
Kinetic experiments Assays for the determination of kcat, and specific activity ratios at pH
7.5/9.5 employed the coupling enzymes, phosphoglucose isomerase and glucose-6-phosphate
dehydrogenase (1). The reduction of NADP+ to NADPH was monitored by absorbance at 340
nm. All other assays used the same coupling enzymes, but monitored the formation of NADPH
by its fluorescence emission at 470 nm, using an excitation wavelength of 340 nm. Assays were
performed at 22 ºC in 50 mM Hepes, pH 7.5, or in 50 mM Caps, pH 9.5. Assay solutions
contained ethylenediamine tetracetic acid (EDTA) and KCl at concentrations of 10 µM and 150
mM, respectively. Initial rates were analyzed with programs written either in MINITAB, using
an α value of 2.0 (26), or by ENZFITTER (27). The kinetic data for AMP inhibition with
respect to Mg2+ and F26P2 inhibition with respect to F16P2 were fit to several models, but only
parameters associated with the best fitting mechanism of inhibition are reported in the results
section.
Crystallization of the product complex Crystals of Leu54 FBPase were grown by the
method of hanging drops. Equal parts of a protein solution and a precipitant solution were
combined in a droplet of 4 µL total volume. Wells contained 500 µL of the precipitant solution.
R-state crystals grew from a protein solution [Leu54 FBPase (10 mg/ml), Hepes (25 mM, pH
7.4), MgCl2 (5 mM), and F16P2 (5 mM)] combined with a precipitant solution [Hepes (100 mM,
6
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 7
pH 7.4), polyethylene glycol 3350 (8% (w/v)), glycerol (27% (v/v)), and t-butanol (5% (v/v))].
Crystals of the R-like AMP complex grew from a protein solution [Leu54 FBPase (10 mg/ml),
Hepes (25 mM, pH 7.4), MgCl2 (5 mM), F16P2 (5 mM), and AMP (5mM)] combined with a
precipitant solution [Hepes (100 mM, pH 7.4), polyethylene glycol 3350 (12% (w/v)), glycerol
(23% (v/v)), and t-butanol (5% (v/v))]. Crystals of the T-state AMP complex grew from a
protein solution [Leu54 FBPase (10 mg/ml), Hepes (25 mM, pH 7.4), MgCl2 (5 mM), F16P2 (5
mM), and AMP (5 mM)] combined with a precipitant solution [Hepes (100 mM, pH 7.4),
polyethylene glycol 3350 (14% (w/v)), 2-methyl-2,4-pentanediol (21% (v/v)), and t-butanol (5%
(v/v))]. Crystals were of equal dimensions (0.2–0.4 mm), growing in approximately three days
at 20 °C. Conditions of crystallization include cryoprotectants; crystals can be transferred
directly from droplet to liquid nitrogen.
Data collection Data were collected at Iowa State University from single crystals on a
Rigaku R-AXIS IV++ rotating anode/image plate system, using CuKα radiation from an Osmic
confocal optics system, and a temperature of 110 K. Data were reduced with the program
package CrystalClear, provided with the instrument.
Structure determination, model building and refinement Crystals of Leu54 FBPase are
isomorphous to either the AMP/Zn2+/product complex (16) or the Zn2+/product complex (10).
Phase angles, used in the generation of initial electron density maps, were based on model 1EYJ
or 1CNQ of the PDB, from which water molecules, metal cations, small-molecule ligands, and
residues 52−72 had been omitted. Residues 52−72 were built into the electron density of omit
maps, using the program XTALVIEW (28). Ligands were added to account for omit electron
density at the active site and/or the AMP site. The resulting models underwent refinement, using
CNS (29) with force constants and parameters of stereochemistry from Engh & Huber (30). A
7
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 8
cycle of refinement consisted of slow cooling from 1000–300 K in steps of 25 K, followed by
120 cycles of conjugate gradient minimization, and concluded by the refinement of individual
thermal parameters. Thermal parameter refinement employed restraints of 1.5 Å2 on nearest
neighbor and next-to-nearest neighbor main chain atoms, 2.0 Å2 on nearest neighbor side chain
atoms, and 2.5 Å2 on next-to-nearest neighbor side chain atoms.
In subsequent cycles of refinement, water molecules were fit to difference electron
density of 2.5σ or better and were added until no significant decrease was evident in the Rfree
value. Water molecules in the final models make suitable donor-acceptor distances to each other
and the protein and have thermal parameters under 60 Å2. Stereochemistry of the models was
examined by the use of PROCHECK (31).
Results
Rationale for the Leu54→Ala mutation— The Cβ atom of Ala54 is at the center of a cluster of
hydrophobic side chains, which forms only when the dynamic loop is in its T-state disengaged
conformation. A mutation at position 54 to a large side chain should disrupt packing interactions
and thereby destabilize the disengaged conformation of the dynamic loop. In contrast, ample
room is available for large side chains at position 54 in the R-state engaged conformation of the
loop. The Ala54→Leu mutation then should shift the equilibrium population of AMP complexes
of FBPase toward the R-state.
Expression and Purification of Wild-Type FBPase Expression and isolation procedures
described above provide wild-type and Leu54 FBPases in at least 95% purity, as judged by SDS-
polyacryamide gel electrophoresis (data not shown). Gels indicated no proteolysis of the
purified enzymes.
Kinetics Experiments Kinetics parameters for Leu54 and wild-type FBPases are in Table I.
8
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 9
The determination of kcat and Km for F16P2 (listed as KmF16P2 in Table I) at pH 7.5 employed a
saturating concentration of Mg2+ (2 mM for Leu54 FBPase and 5 mM for wild-type FBPase) and
concentrations of substrate ranging from 0.5–20 µM. A fit of the Michaelis-Menten equation to
the data provided values for kcat and KmF16P2 (Michaelis constant for F26P2). Ratios of specific
activities at pH 7.5 to 9.5 for wild-type and Leu54 FBPases (each above 3) are indicative of
tetrameric enzymes with intact (non-proteolyzed) polypeptide chains.
The Hill coefficient for Mg2+ was determined at a saturating concentration of F16P2 (20
µM) and concentrations of free Mg2+ ranging from 0.1–5.0 mM. Data were fit to Eq 1:
v/Vm = 1/[1 + (A0.5/A)n] Eq. 1
where v is the velocity, Vm is the maximum velocity at saturating concentrations of F16P2 and
Mg2+, A is the concentration of Mg2+, n is the Hill coefficient for Mg2+, and A0.5 is the
concentration of Mg2+ that gives v/Vm of one-half.
The Hill coefficient for AMP was determined at saturating F16P2 (20 µM), Mg2+
concentrations of 0.8 and 0.15 mM for wild-type and Leu54 FBPases, and AMP concentrations
ranging from 0–500 µM. Data were fit to Eq. 2:
v/V0 = 1/[1 + (I/I0.5)n] Eq. 2
where v is the velocity, V0 is the velocity at an AMP concentration of zero, I is the concentration
of AMP, n is the Hill coefficient for AMP, and I0.5 is the concentration of AMP that gives v/V0 of
one-half.
The kinetic mechanism of AMP inhibition with respect to Mg2+ was determined from
assays that employed saturating (20 µM) F16P2, five different Mg2+ concentrations ranging from
0.8–3.0 mM for wild-type enzyme or 0.2–0.6 mM for Leu54 FBPase, and five different AMP
concentrations ranging from 0–150 µM. A model for linear competitive inhibition (Eq. 3)
9
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 10
provided the best result (goodness-of-fit of less than 4%):
Vm/v = 1 + Ka/A2+ (Ka/KiAMP) (I/A)2 Eq. 3
where v is the velocity, Vm is the velocity at an inhibitor concentration of zero, saturating
concentrations of F16P2 and Mg2+, A is the concentration of Mg2+, I the concentration of AMP,
Ka the Michaelis constant for Mg2+, and KiAMP the dissociation constant for AMP from the
enzyme-inhibitor complex. Eq. 3 constrains the Hill coefficients for Mg2+ and AMP to 2,
consistent with independent determinations of these quantities.
The kinetic mechanism of F26P2 inhibition with respect to F16P2 was determined from
assays that employed saturating Mg2+ (5 mM for wild-type enzyme and 2 mM for Leu54
FBPase), five different concentrations (1–6 µM) of F16P2, and five different concentrations (0–
1.0 µM ) of F26P2. A model for linear competitive inhibition provided the best fit to the data
(goodness-of-fit of less than 3%):
Vm/v = 1 + Kb/B+ (Kb/KiF26P2) (I/B) Eq. 4
where Vm is the velocity at an inhibitor concentration of zero and saturating concentrations of
F16P2 and Mg2+, B is the concentration of F16P2, I the concentration of F26P2, Kb the Michaelis
constant for F16P2, and KiF26P2 the dissociation constant for F26P2 from the inhibitor enzyme-
complex
Product complex of Leu54 FBPase (PDB identifier 1YXI) Crystals belong to the space
group I222 (a=52.8, b=82.8 and c=165.5 Å), and are isomorphous to those of wild-type FBPase
in its R-state, containing one subunit of the tetramer in the asymmetric unit of the crystal (10,16–
18). Electron density for residues 1−6 is weak or absent; the model begins at residue 7 and
continues to the last residue of the sequence. Thermal parameters vary from 10 to 70 Å2. The
model has stereochemistry (as determined by PROCHECK (31)) comparable to that of structures
10
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 11
of equivalent resolution. Statistics for data collection and refinement are in Table II.
The product complex of Leu54 FBPase is identical to that of the wild-type enzyme save
clear electron density showing the leucyl side chain at position 54. One molecule each of F6P
and Pi bind to the active site with three atoms of Mg2+. The dynamic loop (residues 52−72) is in
its engaged conformation. Superposition of the Leu54 subunit onto the wild-type subunit reveals
no deviation in the relative positions of Cα atoms in excess of 0.43 Å. Superposition of the
Leu54 tetramer onto canonical wild-type R- and T-states clearly indicates an R-state complex.
We refer the reader to other descriptions of R-state product complexes (10,16–18) for more
detailed descriptions of comparable structures.
AMP/product R-like complex of Leu54 FBPase (PDB identifier 1YYZ) Crystals belong to the
space group I222 (a=53.8, b=82.6 and c=166.6 Å). They contain one subunit in the asymmetric
unit and are isomorphous to those of wild-type FBPase in its R-state (10,16–18). Electron
density for residues 1−9 is weak or absent; the model begins at residue 10 and continues to the
last residue of the sequence. Thermal parameters vary from 10 to 75 Å2. The model has
stereochemistry comparable to that of structures of equivalent resolution (31).
The subunit of the AMP/product complex of Leu54 FBPase has one molecule each of F6P
and Pi with three atoms of Mg2+ at the active site. In addition, strong electron density is present
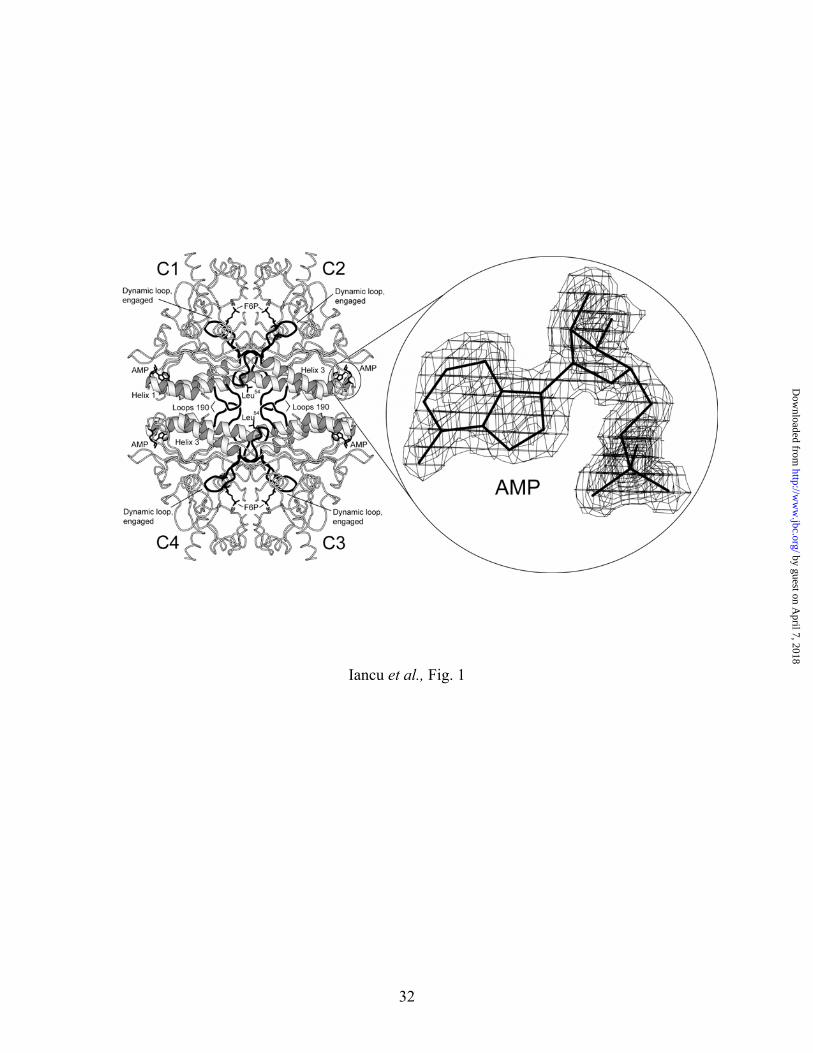
in the allosteric inhibitor pocket, which represents a bound molecule of AMP (Fig. 1). The
dynamic loop (residues 52−72) adopts the engaged conformation. Superposition of the Leu54
tetramer onto canonical wild-type R- and T-states reveals a change in quaternary state (Table
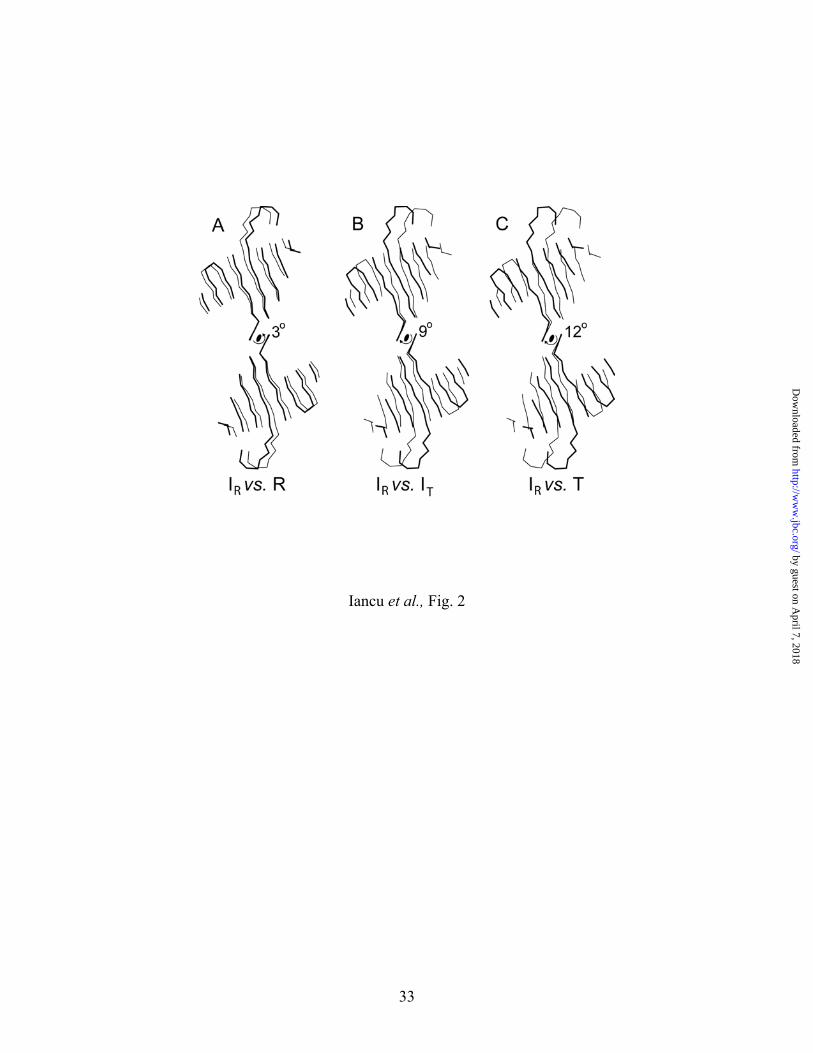
III). Subunit pair C1–C2 has rotated 3º relative to C3–C4 (Fig. 2). The subunit rotation lies
between that of the canonical R-state (0º rotation) and T-state (15º rotation), and differs from the
intermediate quaternary state (9º rotation) stabilized by the allosteric inhibitor OC252 (Fig. 2).
11
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 12
Hereafter, we will use the label IR to represent the R-like state of the AMP/product complex of
Leu54 FBPase, and IT to represent the T-like state of the OC252 complex.
The IR structure reveals the effect of AMP binding in the absence of a complete
quaternary transition. The superposition of the IR subunit onto the subunits of the R-state
tetramer removes coordinate displacements due to the partial (3°) rotation of the subunit pairs in
the IR-state, revealing conformational changes at the tertiary level. In such a comparison,
conformational changes are evident only in the N-terminal element and helices H1 and H2. The
6-amino group of AMP draws backbone carbonyl 17 (helix H1) toward itself, while pushing
away the side chain of Val17 in avoiding unacceptable nonbonded contacts (Fig. 3a). The
interactions between AMP and Val17 translate helix H1 by 0.5 Å toward the center of the
tetramer and move C-terminal end of helix H1 approximately 1.0 Å toward the bound AMP
molecule. The movement compresses the N-terminal end of helix H1 into residues 193–195 of
subunit C1, and displaces the side chain of Ile10 from a hydrophobic cluster of residues (Figs.
3b&c). Helix H2 moves along its axis 0.5 Å away from the center of the tetramer. Helix
movements sever hydrogen bonds between Thr14 and Asn35 and between Thr39 and Glu192, the
latter a linkage between subunits C1 and C4 (Table IV). Moreover, the hydrogen bond between
Thr46 and backbone carbonyl 189 (another C1–C4 contact) may be weakened. Lys42 remains in
its inter-subunit salt link with Glu192, with little change to its other hydrogen bonding
interactions. The loss or weakening of hydrogen bonds involving Thr39, Thr46, and Glu192
observed in the IR structure is not evident in a direct comparison of the canonical R- and T-states
(Table IV).
The superposition of the IR-state subunit onto the subunits of the T-state tetramer of wild-
type FBPase reveals tertiary conformational change due to the 12º subunit-pair rotation. This
12
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 13
includes the movement of the dynamic loop from its engaged to disengaged conformer, a
displacement of more than 30 Å for some Cα carbons of that loop. For the most part, other
conformational changes involve modest displacements in atoms not exceeding 0.5 Å. These
changes involve almost every atom in FBPase in a correlated set of collective movements.
Immediately evident is the additional translation of helix H2, outward from the center of the
tetramer and along its axis, and the occurrence of unacceptable contacts between loops 190 (Fig.
4). The unacceptable contacts are relaxed in the T-state subunit by movements in loops 190 in
subunits C1 and C4 away from a molecular axis of twofold symmetry.
AMP/product T-state complex of Leu54 FBPase (PDB identifier 1YZO) Crystals belong to the
space group P21212 (a=59.7, b=166.1 and c=78.9), and are isomorphous to those of AMP
complexes of FBPase (16,17). The subunit pair C1–C2 is in the asymmetric unit of this crystal
form. Regions of weak or absent electron density include residues 1−8 and 55−72. The model
begins at residue 9 and continues to the last residue of the sequence, but segment 55−72 is
unreliable, as evidenced by thermal parameters as high as 113 Å2. The model has
stereochemistry generally comparable to that of structures derived from data of comparable
resolution (31). Statistics for data collection and refinement are in Table II.
The enzyme in this crystal form is in the T-state (quaternary transition angle of 15º);
however, unlike loop-disengaged AMP complexes of the wild-type enzyme, the dynamic loop in
T-state Leu54 FBPase is disordered. Moreover, hydrogen bonds normally well established in the
T-state of the wild-type enzyme seem to be marginally weaker in T-state Leu54 FBPase. The
active site retains Pi, F6P, and Mg2+ bound to sites 1 and 2.
13
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 14
Discussion
Conformational changes between the R- and IR-states of Leu54 FBPase are consistent with
two models for quaternary change: a concerted model in which AMP drives the quaternary
transition by acting on a set of interconnected levers and a sequential model in which AMP raises
the energy level of the R-state while simultaneously lowering that of the nascent T-state (Fig. 5).
The point of departure for each model is an AMP-induced translation of helices H1 and H2 in
opposite directions, helix H1 toward and helix H2 away from the center of the tetramer. The two
models differ in regard to the consequences of helix H2 movement. For the concerted model,
helix H2 retains its interactions with loops 190, whereas in the sequential model, interactions
between helix H2 and loops 190 are broken.
In the concerted model, two sets of coupling interactions distribute forces throughout the
entire tetramer due to the binding of one AMP molecule. One set of interactions involves Thr39,
Lys42 and Thr46 of helices H2 with Glu192 and backbone carbonyls 189 and 190 of loops 190
(Table IV). These interactions link subunits C1 to C4 and C2 to C3. A second set of interactions
defines the C1–C2 and symmetry-related C3–C4 subunit interfaces (13). The binding of one
molecule of AMP to say subunit C1, results in the aforementioned movements of helices H1 and
H2. The movement in helix H2 of subunit C1 exerts an outward force on loops 190 of subunits
C1 and C4. The C1–C2 and C3–C4 subunit interactions, however, constrain loops 190 to a fixed
distance from the center of the tetramer. Loops 190 can only follow the outward movement of
helix H2 by the rigid body rotations of subunit pairs C1–C2 and C3–C4 (Fig. 5). The two sets of
coupling interactions insure that all loops 190 and their associated subunits undergo rigid-body
motions, and that all helices H2 undergo an outward movement in response to the binding of one
or more molecules of AMP.
14
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 15
The concerted model suffers from two significant shortcomings. Firstly, the coupling
interactions between helix H2 and loops 190 have weakened in the IR state. Only the interactions
involving Lys42 appear unaffected by movements in helices H2, and we suggest below that even
this critical salt link may rupture during the quaternary transition. The weakened linkages
between helices H2 and loops 190, however, may be the consequence of having four AMP
molecules bound to an R-state tetramer. Two bound molecules of AMP convert R-state hybrid
tetramers of FBPase into their T-states (32). Hence, the IR-state of the Leu54 FBPase may
represent an “over-torqued” tetramer, the existence of which is possible only because the
mutation at position 54 eliminates the T-state as a low-energy alternative. The second
shortcoming of the concerted model is not so easy to dismiss. A concerted mechanism for
FBPase requires cooperativity in the binding of AMP molecules to any pair of sites. A hybrid
tetramer of FBPase that constrains AMP-binding to subunits C1 and C2, however, exhibits non-
cooperative inhibition even though it undergoes a quaternary transition (32).
In considering the sequential model for the quaternary transition, we note first that the
subunit rotation observed in the R- to T-state transition cannot happen as a rigid-body motion. In
the R-state, loop 190 from subunit C1 is in contact with the symmetry-related loop from subunit
C4. Progress toward the T-state results in unacceptable contacts between loops 190 from
subunits C1 and C4. Loop 190 must undergo conformational change, but multiple hydrogen
bonds fix its conformation in both the R- and T-states (Table IV). The movement of helix H2
releases a conformational restraint on loop 190 in a neighboring subunit by the disruption or
weakening of hydrogen bonds involving Thr39 and Thr46. In this environment of fewer restraints,
loop 190 is more likely to relax unfavorable contacts that occur during the quaternary transition.
Moreover, the movement in helix H2 favorably positions Thr39 and Thr46 for the formation of
15
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 16
strong hydrogen bonds in the T-state. In fact, the hydrogen bond involving Glu192 and Thr39,
ruptured in the R- to IR-state transition, reforms in the T-state. The mechanism is fully
reversible: loss of AMP from the T-state presumably causes helix H2 to move back toward the
center of the enzyme, breaking or weakening hydrogen bonds between subunits C1 and C4, and
repositioning Thr39 and Thr46 in favor of R-state interactions.
The sequential model can accommodate both cooperative and non-cooperative
mechanisms of AMP inhibition. A mixture of hybrid mutants of FBPase that force AMP-
binding to opposite halves of the tetramer exhibits cooperative inhibition (32). Hence, subunit
coupling must exist between top and bottom halves of the tetramer. Coupling interactions
necessarily involve subunits C1 and C4, as subunits C1 and C3 have no direct interactions in the
R-state. The binding of AMP to subunit C1 may not only disrupt hydrogen bonds between helix
H2 of subunit C1 and loop 190 of subunit C4, but it may also weaken symmetry-related
hydrogen bonds between helix H2 of subunit C4 and loop 190 of subunit C1. Hence, a second
molecule of AMP would divert less of its binding energy to the movement of helix H2 in subunit
C4 and, as a consequence, bind with higher affinity. All reported mutations of Lys42, Arg49,
Glu192, Ile190, and Gly191, and some mutations of Lys50, abolish AMP cooperativity (22,33–35).
These residues are part of helix H2 or loop 190, and are near to or part of the coupling
interactions between subunits C1 and C4.
A second coupling pathway between AMP binding sites may involve Arg22. The
mutation of Arg22 to methionine eliminates cooperativity in AMP inhibition (36). Arg22 is near
the AMP pocket, has high thermal parameters in the R-state, and does not participate in inter-
subunit hydrogen bonds in either the R- or T-state structures. Replacing subunit C1 of R-state
Leu54 FBPase with an IR subunit generates a model that approximates FBPase with one bound
16
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 17
molecule of AMP (Fig. 6). In that model, Arg22 of subunit C1, due to conformational changes
induced by AMP, makes a strong hydrogen bond with backbone carbonyl 108 of subunit C4.
The formation of the symmetry related hydrogen bond involving Arg22 of subunit C4 should
induce conformational changes in subunit C4 that favor the binding of AMP.
The concerted and sequential models both assume an energy barrier between R- and T-
states. In the sequential model, hydrogen bonds involving Thr39 and Thr46 contribute
significantly to the barrier, but not so in the concerted model as these interactions are retained.
At least one other interaction may contribute significantly to the energy barrier between R- and
T-states. Unacceptable contacts between the side chains of Glu192 (subunit C4) and Lys42
(subunit C1) are likely during the quaternary transition (Fig. 4b). A conformational change in
the side chain of Glu192 eliminates bad contacts with Lys42, and re-establishes its hydrogen bond
with Thr39. The conformational change in Glu192 may require a transitory loss or weakening of
its salt-link with Lys42. The presumed loss of this salt-link could favor the dissociation of the
tetramer into subunit pairs (C1–C2 from C3–C4), and thereby represent the first step in FBPase
subunit exchange kinetics (32,37).
The models above have yet to consider conformational change in the dynamic loop
(residues 52–72). In all reported structures of FBPase, the dynamic loop is either engaged or
disordered in R-like states, and either disordered or disengaged in T-like states. The AMP-
induced movement of helix H1, which probably occurs in concert with that of helix H2,
displaces Ile10 from a hydrophobic surface. That surface interacts with side chains of the
dynamic loop in its disengaged T-state conformation (Fig. 4c). In the R-state then, Ile10
effectively blocks the disengaged conformer of the dynamic loop. The formation of the
disengaged conformer appears as a significant thermodynamic driving force in the quaternary
17
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 18
transition to the T-state. A modest change in the conditions of crystallization (substitution of
glycerol for 2-methy-2,4-pentanediol) transforms the AMP/product complex of Leu54 FBPase
from a loop-disordered T-state to a loop-engaged R-state. Direct interactions between
cryoprotectant and enzyme are unlikely as no bound cryoprotectant molecules appear in either
crystal structure. The AMP/product complex of Leu54 FBPase then probably has substantial
populations of IR- and T-states in solution, allowing the growth of different crystal forms under
nearly identical conditions.
The potential significance of C1–C4 interactions in FBPase has been suggested by others
(13,22), but crystal structures of the R- and T-states did not reveal the transitory loss of hydrogen
bonds across the C1–C4 subunit interface. As a consequence, the basis for a sequential
mechanism of quaternary change remained hidden. The sequential model presented here
reconciles properties of AMP inhibition in wild-type, mutant and hybrid FBPases with known
conformational changes in the tetramer. The present study also suggests that AMP-ligation of
the R-state does not displace the dynamic loop from its engaged conformation. Instead, the
dynamic loop leaves the engaged conformation in the T-state, for reasons now poorly
understood.
18
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 19
References
1. Benkovic, S. T., and de Maine, M. M. (1982) Adv. Enzymol. Relat. Areas Mol. Biol. 53,
45-82
2. Tejwani, G. A. (1983) Adv. Enzymol. Relat. Areas Mol. Biol. 54, 121-194
3. Van Schaftingen, E. (1987) Adv. Enzymol. Relat. Areas Mol Biol. 59, 45-82
4. Pilkis, S. J., El-Maghrabi, M. R., and Claus, T. H. (1988) Ann. Rev. Biochem. 57, 755-
783
5. Okar, D. A., and Lange, A. J. (1999) Biofactors 10, 1-14
6. Pilkis, S. J., El-Maghrabi, R. M, McGrane, M. M., Pilkis, J., and Claus, T. H. (1981) J.
Biol. Chem. 256, 3619-3622
7. Liu, F., and Fromm, H. J. (1991) J. Biol. Chem. 266, 11774-11778
8. Liu, F., and Fromm, H. J. (1988) J. Biol. Chem. 263, 9122-9128
9. Scheffler, J. E., and Fromm, H. J. (1986) Biochemistry 25, 6659-6665
10. Choe J-Y., Poland B. W., Fromm H. J., and Honzatko R. B. (1998) Biochemistry 33,
11441-11450
11. Stone, S. R., and Fromm, H. J. (1980) Biochemistry 19, 620-625
12. Ke, H., Zhang, Y. Liang, J-Y., and Lipscomb, W. N. (1991) Proc. Natl. Acad. Sci. U.S.A.
88, 2989-2993
13. Zhang, Y., Liang, J-Y., Huang, S., and Lipscomb, W. M. (1994) J. Mol. Biol. 244, 609-
624
14. Nelson, S. W., Kurbanov, F., Honzatko, R. B. and Fromm, H, J. (2001) J. Biol. Chem.
276, 6119-6124
15. Xue, Y., Huang, S., Liang, J-Y. Zhang, Y., and Lipscomb, W. N. (1994) Proc. Natl.
19
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 20
Acad. Sci. USA 91, 12482-12486
16. Choe, J-Y., Fromm, H. J., and Honzatko, R. B. (2000) Biochemistry 39, 8565-8574
17. Choe, J-Y., Nelson, S. W., Fromm, H. J., and Honzatko, R. B. (2003) J. Biol. Chem. 278,
16008-16014
18. Choe, J-Y., Iancu, C. V., Fromm, H. J., and Honzatko, R. B. (2003) J. Biol. Chem. 278,
16015-16020
19. Nelson, S. W., Choe, J-Y., Iancu, C. V., Honzatko, R. B., and Fromm, H. J. (2000)
Biochemistry. 39, 11100-11106
20. Wen, J., Nelson, S. W., Honzatko, R. B., Fromm, H. J., and Petrich, J. W. (2001)
Photochem. Photobiol. 74, 679-685
21. Ke, H. M., Thorpe, C. M., Seaton, B., Lipscomb, W. N., and Marcus F. (1990) J. Mol.
Biol. 212, 513-539
22. Lu, G., Stec, B., Giroux, E. L., Kantrowitz, E. R. (1996) Protein Science 5, 2333-2342
23. Choe, J-Y., Nelson, S. W, Arienti, K. L., Axe, F. U., Collins, T. L., Jones, T. K.,
Kimmich, R. D. A., Newman, M. J., Norvell, K., Ripka, W. C., Romano, S. J., Short, K.
M., Slee, D. H., Fromm, H. J., and Honzatko, R. B. (2003) J. Biol. Chem. 278, 51175-
51183
24. Laemmli, U. K. (1970) Nature 227, 680-685
25. Bradford, M. M. (1976) Anal. Biochem. 72, 248-252
26. Liu, F., and Fromm, H.J. (1990) J. Biol. Chem. 265, 7401-7406.
27. Leatherbarrow, R.J. (1987) ENZFITTER: A Non-Linear Regression Data Analysis
Program for the IBM PC, pp.13-75, Elsevier Science Publishers B.V. Amsterdam.
28. McRee, D. E. (1992) J. Mol. Graph. 10, 44-46
20
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 21
29. Brünger, A. T., Adams, P. D., Clore, G. M., DeLano, W. L., Gros, P., Grosse-Kunstleve,
R. W., Jiang, J.-S., Kuszewski, J., Nilges, M., Pannu, M. S., Read, R. J., Rice, L. M.,
Simonson, T., Warren, G. L. (1998) Acta Crystallogr. D 54, 905-921
30. Engh, R. A., and Huber, R. (1991) Acta Crystallog. A 47, 392-400
31. Laskowski, R. A., Mac Arthur, M. W., Moss, D. S., and Thornton, J. M. (1993) J. Appl.
Crystallogr. 26, 283-291
32. Nelson, S. W, Honzatko, R. B., and Fromm, H. J. (2002) J. Biol. Chem. 277, 15539-
15545
33. Lu, G., Giroux, E. L., and Kantrowitz, E. R. (1997) J. Biol. Chem. 272, 5076-5081
34. Cárcamo, J. G., Yañez, A. J., Ludwig, H. C., León, O., Pinto, R. O., Reyes, A. M., and
Slebe, J. C. (2000) Eur. J. Biochem. 267, 2242-2251
35. Shyur, L.-F., Aleshin, A. E., Honzatko, R. B., and Fromm, H. J. (1996) J. Biol. Chem.
271, 33301-33307
36. Shyur, L.-F., Aleshin, A. E., Honzatko, R. B., and Fromm, H. J. (1996) J. Biol. Chem.
271, 3005-3010
37. Nelson, S. W., Honzatko, R. B., and Fromm, H. J. (2001) FEBS Lett. 492, 254-258
38. Kraulis, J. (1991) J. Appl. Crystallogr. 24, 946-950
21
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 22
Footnotes
1The abbreviations used are: FBPase, fructose-1,6-bisphosphatase; F16P2, fructose 1,6-
bisphosphate; F6P, fructose 6-phosphate; F26P2, fructose 2,6-bisphosphate; Pi, orthophosphate.
22
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 23
Table I. Kinetic parameters for wild-type and Leu54 FBPases. Parameters are defined in the
results section.
Wild-type Leu54→Ala
Activity ratio, pH 7.5:9.5 3.5±0.5 3.9±0.4
kcat (sec-1) 20±1 11.3±0.6
KmF16P2 (µM) 1.2±0.05 0.94±0.04
A0.5 (mM) 0.84±0.04 0.14±0.03
Hill coefficient Mg2+ 1.7±0.1 2.1±0.2
I0.5 (µM) 1.23±0.04 62±1
Hill coefficient AMP 2.2±0.1 2.5±0.1
Ka (mM2) 0.78±0.2 0.087±0.005
KiAMP (µM2) 0.6±0.1 3000±200
Kb (µM) 2.4±0.4 1.2±0.1
KiF26P2 (µM) 0.23±0.04 0.38±0.04
23
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 24
Table II. Statistics of data collection and refinement for Leu54 FBPase.
Crystalline complexa R-state IR-state T-state
Resolution limit (Å) 2.0 1.85 2.15
Number of measurements 155,545 124,926 193,057
Number of unique reflections 24,972 32,122 41,418
Completeness of data (%):
Overall 99.6 99.1 99.2
Last shell/resolution-range (Å) 96.5/2.07-2.0 91.1/1.96-1.85 98.6/2.25-2.15
Rsymb
Overall 0.066 0.035 0.049
Last shell/resolution-range (Å) 0.228/2.07-2.0 0.211/1.96-1.85
0.197/2.25-2.15
Number of reflections in refinement 22,691 29,277 37,089
Number of atoms 2,796 2,785 5,627
Number of solvent sites 176 296 510
Rfactorc 0.199 0.223 0.212
24
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 25
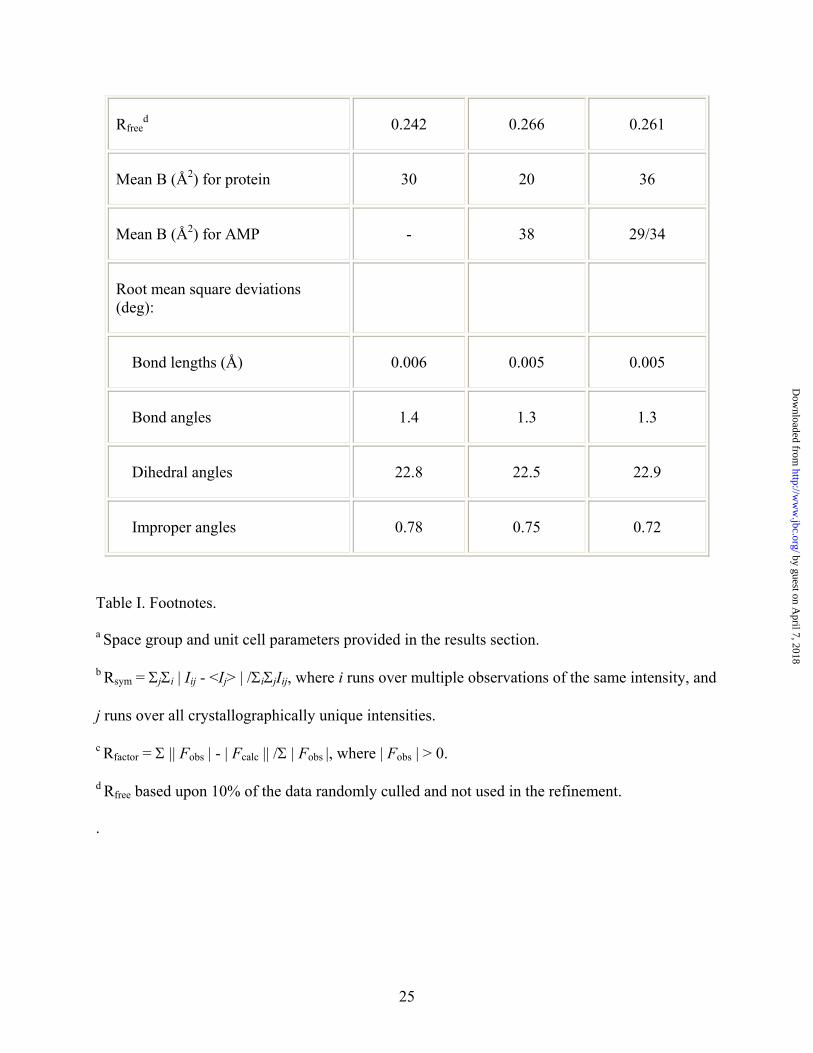
Rfreed 0.242 0.266 0.261
Mean B (Å2) for protein 30 20 36
Mean B (Å2) for AMP - 38 29/34
Root mean square deviations (deg):
Bond lengths (Å) 0.006 0.005 0.005
Bond angles 1.4 1.3 1.3
Dihedral angles 22.8 22.5 22.9
Improper angles 0.78 0.75 0.72
Table I. Footnotes.
a Space group and unit cell parameters provided in the results section.
b Rsym = ΣjΣi | Iij - <Ij> | /ΣiΣjIij, where i runs over multiple observations of the same intensity, and
j runs over all crystallographically unique intensities.
c Rfactor = Σ || Fobs | - | Fcalc || /Σ | Fobs |, where | Fobs | > 0.
d Rfree based upon 10% of the data randomly culled and not used in the refinement.
.
25
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 26
Table III. Root-mean-squared deviations between conformational states of FBPase. RCSB
identifiers 1EYK and 1CNQ represent the canonical T- and R-states, respectively. IT is the T-
like state of the OC252 complex of FBPase (PDB identifier 1Q9D). IR is the R-like state of the
AMP/product complex of Leu54 FBPase reported here. The determined angle of rotation is
sensitive to the subset of Cα atoms used in the calculation of the rotation matrix. The use of all
Cα atoms, including those of the dynamic loop, results in an angle of 18º for the R- to T-state
transition. In contrast, the use of Cα atoms that deviate by less than 1 Å in the initial
superposition gives an angle of 13º. Superpositions and root-mean squared deviations here are
based on Cα atoms from the following residues: 33-49, 75-265, and 272-330. Values in bold
typeface come from superpositions of tetramers. Other values come from superpositions of C1–
C2 dimers. The latter provide an estimate of coordinate uncertainty due to random and
systematic errors.
IR IT T
R 0.30/0.74 0.48/2.16 0.58/2.7
IR 0.43/1.54 0.48/2.09
IT 0.33/0.64
26
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 27
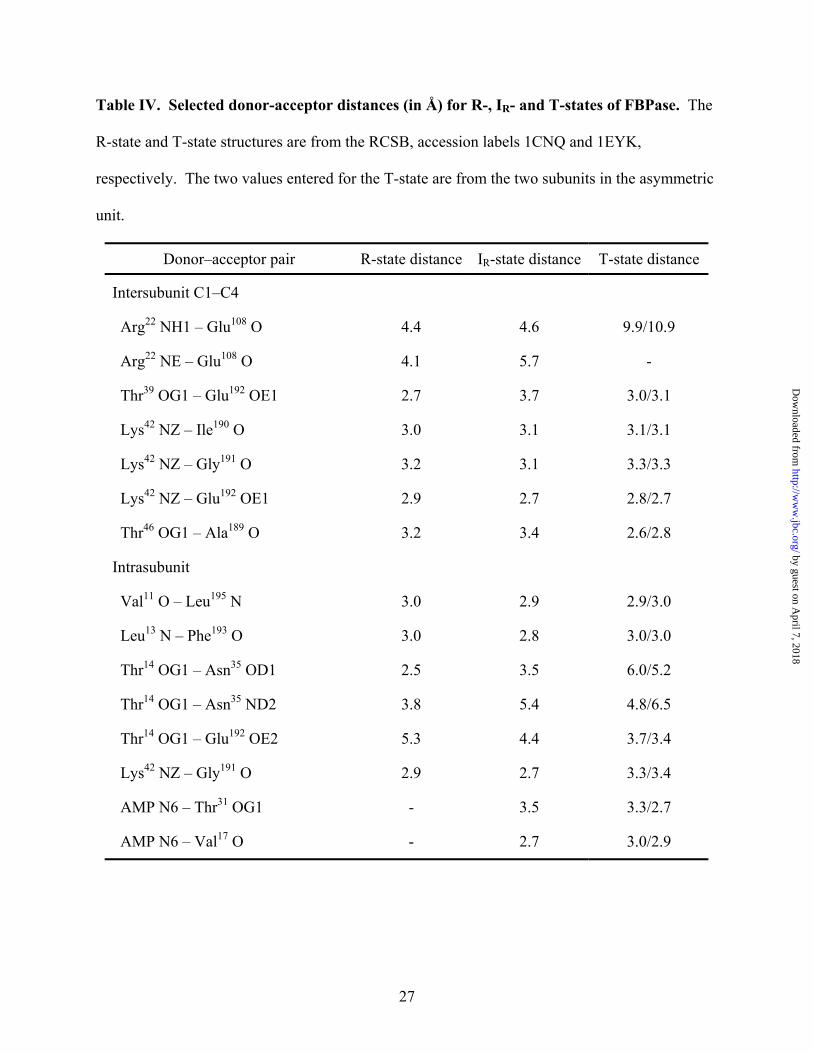
Table IV. Selected donor-acceptor distances (in Å) for R-, IR- and T-states of FBPase. The
R-state and T-state structures are from the RCSB, accession labels 1CNQ and 1EYK,
respectively. The two values entered for the T-state are from the two subunits in the asymmetric
unit.
Donor–acceptor pair R-state distance IR-state distance T-state distance
Intersubunit C1–C4
Arg22 NH1 – Glu108 O 4.4 4.6 9.9/10.9
Arg22 NE – Glu108 O 4.1 5.7 -
Thr39 OG1 – Glu192 OE1 2.7 3.7 3.0/3.1
Lys42 NZ – Ile190 O 3.0 3.1 3.1/3.1
Lys42 NZ – Gly191 O 3.2 3.1 3.3/3.3
Lys42 NZ – Glu192 OE1 2.9 2.7 2.8/2.7
Thr46 OG1 – Ala189 O 3.2 3.4 2.6/2.8
Intrasubunit
Val11 O – Leu195 N 3.0 2.9 2.9/3.0
Leu13 N – Phe193 O 3.0 2.8 3.0/3.0
Thr14 OG1 – Asn35 OD1 2.5 3.5 6.0/5.2
Thr14 OG1 – Asn35 ND2 3.8 5.4 4.8/6.5
Thr14 OG1 – Glu192 OE2 5.3 4.4 3.7/3.4
Lys42 NZ – Gly191 O 2.9 2.7 3.3/3.4
AMP N6 – Thr31 OG1 - 3.5 3.3/2.7
AMP N6 – Val17 O - 2.7 3.0/2.9
27
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 28
Figure Legends
Fig. 1. Overview of the R-like AMP/product complex of Leu54 FBPase. The four subunits of
the tetramer, labeled C1 through C4, are depicted with one molecule each of bound AMP
(allosteric pocket) and F6P (active site). Active-site bound Pi and Mg2+ and side chains
of Leu54 from subunits C1 and C3 are omitted for clarity. Residues 52–72 (dynamic
loop) and 187–195 (loop 190) are in black. The panel to the right shows electron density
covering the AMP molecule from an omit map contoured at a level of 1-σ with a cutoff
radius of 1 Å. This drawing was prepared with MOLSCIRPT (38).
Fig. 2. Quaternary states of FBPase. Various quaternary states of FBPase differ by rotations
of subunit pair C3–C4 relative to subunit pair C1–C2. Superpositions of FBPase
tetramers, using selected Cα atoms from subunit pair C1–C2, reveals significant
displacements in Cα atoms of C3–C4 subunit pairs in instances or differing quaternary
states. Depicted here are displacements in Cα carbons of β-strands of the AMP domain
of the subunit pairs C3–C4 in the IR complex (bold lines) relative to those of FBPase in
the R-state (A) and IT-state (B), and T-state (C). The magnitude and direction of subunit
rotations that transform the IR-state into the three other quaternary states of FBPase is
indicated. The Cα atoms listed in Table III are the basis for superposition of subunit pair
C1–C2. This drawing was prepared with MOLSCIRPT (38).
Fig. 3. Tertiary conformational changes between R- and IR-states. Dotted lines represent
selected donor-acceptor interactions of 3.2 Å or less. Solid green lines represent potential
non-bonded contacts of 2.5 Å or less. Panels A–C: Superposition of the IR-state subunit
(red) onto each subunit of the R-state tetramer (black) reveals tertiary conformational
changes induced by the binding of AMP and the resulting 3º subunit-pair rotation.
28
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 29
Hydrogen bonds (dotted red lines) involving the 6-amino group of AMP and a non-
bonded contact (green line) with the side chain of Val17 induce helix movement and
shears hydrogen bonds between Thr14 and Asn35 (panel A), as well as Glu192 and Thr39
(panels A–C). In addition, the movement of helix H1 displaces Ile10 from its R-state
hydrophobic contacts (panels B and C). This drawing was prepared with MOLSCIRPT
(38). Inset: Shown are regions of the tetramer (purple) and viewing directions (bold
arrows) corresponding to panels A–C. The viewing direction for panel C is 45º inclined
to the plane of the tetramer.
Figure 4. Tertiary conformational changes between T- and IR-states. Superposition of the
IR-state subunit (red) onto each subunit of the T-state tetramer (black) reveals tertiary
conformational changes induced by the 12º subunit-pair rotation. An additional
translation of helix H2 along its axis is evident (panels A–C). The disengaged loop
(residues 52–72) covers the hydrophobic surface exposed by the displacement of Ile10
(panel C). Close contacts between loops 190 of subunits C1 and C4 and Glu192 and Lys42
(solid green lines) are relaxed by conformational changes. The conformational change in
Glu192 re-establishes its hydrogen bond with Thr39 (panels A–C). Regions of FBPase
depicted and viewing directions are as indicated in Fig. 3. This drawing was prepared
with MOLSCIRPT (38).
Fig. 5. Models of concerted and sequential conformational change. The subunits of the
FBPase tetramer are simplified to helix H2 (rectangle) and loop 190 (oval). The viewing
direction is down a molecular axis of twofold symmetry, with subunit C1 and C2 above
the plane (bold lines) and subunit C3 and C4 below the plane. In the concerted model
(left) AMP molecules bind successively in any order to the subunits of tetramer, until the
29
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 30
combined torque (represented by open arrows) on each subunit pair exceeds the energy
barrier that separates the R- and T-states. The binding of at least two AMP molecules is
necessary for the concerted conformational change. In the sequential model (right), the
first molecule of AMP can bind to any subunit with equal affinity, causing the outward
movement of helix H2 of only that subunit (filled arrow). If binding occurs at subunit
C1, interactions between subunit C1 and C4 are weakened. The second molecule of
AMP binds to subunit C4, because less binding energy is spent in the movement of helix
H2 in that subunit. The R- to T-state transition can occur in response to the binding of at
least two molecules of AMP. For the sequential mechanism, transition to the T-state
restores hydrogen bonds lost by the AMP-induced movements of helices H2 in the R-
state.
Fig.6. Stereoview of the proposed role of Arg22 in cooperativity of AMP inhibition. The
view is down a molecular twofold axis toward the center of the tetramer. The
superposition of the IR-state subunit (bold lines) onto subunit C1 of the R-state (fine
lines) represents possible relaxation events due to the binding of AMP to subunit C1
(top). AMP-induced conformational change in helix H1 would allow the formation of a
hydrogen bond between Arg22 (subunit C1) and backbone carbonyl 108 (subunit C4), and
stack the side chains of Arg22 and Phe89 (subunit C4). A second superposition of an IR-
state subunit (bold lines) onto subunit C4 of the R-state (fine lines) represents the
relaxation of subunit C4 to the altered conformation of AMP-bound subunit C1 (bottom).
The proposed interaction involving Arg22 (subunit C1) carries over to the symmetry-
related Arg22 (subunit C4). As a consequence, the AMP pocket of subunit C4 may adopt
30
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 31
a conformation that approximates the AMP-bound conformation of subunit C1, even in
the absence of AMP.
31
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 32
Iancu et al., Fig. 1
32
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 33
Iancu et al., Fig. 2
33
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 34
Iancu et al., Fig. 3
34
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 35
Iancu et al., Fig. 4
35
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 36
Iancu et al., Fig. 5
36
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 37
Iancu et al., Fig. 6
37
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 38
Cristina V. Iancu, Susmith Mukund, Herbert J. Fromm and Richard B. Honzatkofructose-1,6-bisphosphatase
R-state AMP complex reveals initial steps of the quaternary transition of
published online March 14, 2005J. Biol. Chem.
10.1074/jbc.M501011200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from