Page 1
EPIDEMIOLOGI
Analisi Jurnal “Comparative ecology of 11 sympatric species of Macaranga in Borneo: tree distribution in relation to horizontal and vertical resource heterogeneity”
DISUSUN OLEH:
Aulia Herika Putri
Feni Tiara Diah
Povi Olivia
Willa Elisa Br Sembiring
PROGRAM STUDI ILMU KEPERAWATAN
FAKULTAS KEDOKTERAN
UNIVERSITAS SRIWIJAYA
2015
Page 2
DAFTAR ISI
DAFTAR ISI.............................................................................................................................................1
BAB I......................................................................................................................................................2
PENDAHULUAN.....................................................................................................................................2
I.1 Latar Belakang.......................................................................................................................2
I.2 Hipotesis................................................................................................................................3
I.3 Tujuan....................................................................................................................................3
BAB II.....................................................................................................................................................4
METODE................................................................................................................................................4
II.1 Design Studi...........................................................................................................................4
II.2 Populasi dan Sample..............................................................................................................4
II.3 Analisis Data..........................................................................................................................5
BAB III....................................................................................................................................................9
PEMBAHASAN.......................................................................................................................................9
III.1 Hasil Jurnal.............................................................................................................................9
III.2 Pembahasan........................................................................................................................17
III.3 Keterbatasan Penelitian.......................................................................................................22
1
Page 3
BAB I
PENDAHULUAN
I.1 Latar BelakangAdapaun yang melatar belakangi penulis membuat jurnal yang berjudul
“Comparative ecology of 11 sympatric species of Macaranga in Borneo: tree
distribution in relation to horizontal and vertical resource heterogeneity" ialah karena
ada beragam jenis pohon pada hutan tropis, apalagi dalam kapasitas mereka untuk
memanfaatkan sumber daya dan distribusi mereka yang sangat dipengaruhi oleh
heterogenitas spasial dan temporal ketersediaan sumber daya. Heterogenitas spasial
sumber daya tanah ini mempengaruh pola skala luas dari distribusi jenis-jenis pohon
tersbut. Dalam hutan, heterogenitas ketersediaan cahaya di kontinum celah bawah
dapat menentukan distribusi local. Baik skala-variasi kondisi tanah, misalnya dalam
kesenjangan pohon-jatuh, serta juga dapat mempengaruhi pola distribusi spesies.
Selanjutnya, gradien ketersediaan cahaya turun melalui kanopi hutan yang dipengaruh
oleh distribusi vertical. Beberapa penelitian, menunjukkan walaupun , telah
dianggapnya sebuah array dari kedua komponen horisontal dan vertikal dari
ketersediaan sumber daya, dan bagaimana mereka mungkin berinteraksi untuk
mempengaruhi distribusi dan koeksistensi pohon-pohon tropis simpatrik.
Banyak penelitian tentang heterogenitas horizontal ketersediaan sumber daya
telah berfokus pada pentingnya tingkat cahaya, dan variasi konsekuen dalam
kehidupan pohon histories dalam kaitannya dengan gap-tumbuhan bawah kontinum.
Pioneer dan pohon non-perintis spesies telah terbukti berbeda dalam distribusi dan
kinerja pada berbagai tingkat cahaya, Tapi beberapa studi telah meneliti ekologis
kelompok ini 'mirip' spesies seperti non-pionir, dan dianggap jarang memiliki sumber
daya lain selain cahaya. Kohyama (1993) mengemukakan bahwa menggabungkan
struktur horisontal dan vertikal ke dalam model dinamika hutan meningkatkan kondisi
di mana stabil koeksistensi spesies terjadi, bahkan ketika spesies semua non-pionir.
Kami menyelidiki distribusi 11 spesies pelopor Macaranga (Euphorbiaceae) di
Kalimantan, dalam kaitannya dengan berbagai komponen horisontal dan vertikal dari
heterogenitas sumber daya. Kami berpendapat bahwa pemahaman tentang variasi
kehidupan-sejarah dan perannya dalam pemeliharaan keanekaragaman tinggi di hutan
hujan tropis, harus melibatkan analisis rinci tanggapan spesies untuk spektrum penuh
2
Page 4
heterogenitas sumber daya di seluruh siklus hidup mereka (lihat juga Clark & Clark
1992 ).
I.2 HipotesisHorizontal dan vertikal heterogenitas ketersediaan sumber daya, ditambah
dengan penggunaan khusus sumber daya dengan jenis pohon, menghasilkan pola yang
kompleks dari distribusi jenis pohon di hutan hujan tropis. Distribusi horizontal setiap
pohon dinilai sehubungan dengan mahkota tingkat cahaya, microsites pendirian, dan
variasi skala yang lebih luas di tanah properti tekstur. Distribusi horizontal setiap
pohon dinilai sehubungan dengan mahkota tingkat cahaya, microsites pendirian, dan
variasi skala yang lebih luas di tanah properti tekstur.
I.3 TujuanTujuan penelitian ini yaitu untuk membuktikan apakah distribusi pohon
horisontal, seperti yang diharapkan, erat berkorelasi dengan bukaan kanopi dan
lingkungan cahaya tinggi, dan apakah distribusi terhadap microsites fisik tertentu bias
untuk perkecambahan dan pembentukan, misalnya gundukan tip-up atau situs yang
kaya nutrisi.
3
Page 5
BAB II
METODE
II.1 Design StudiJurnal ini menggunakan metode penelitian studi kolerasional dengan
menggunakan pendekan allometric. Peneliti mempelajari studi hubungan antara
beberapa variable yakni menguji distribusi horizontal dan vertikal dari 4.014 individu
di 11 spesies awal, yang mana distribusi horizontal setiap pohon dinilai hubungan
dengan mahkota tingkat cahaya, microsites pendirian, dan variasi skala yang lebih
luas di tanah properti tekstur. Distribusi vertikal dinilai menggunakan pendekatan
alometrik untuk memperkirakan ketinggian maksimum pohon (Hmax) dan
kemiringan hubungan pancang tinggi-diameter.
II.2 Populasi dan SamplePenelitian ini terjadi di Taman Nasional Bukit Lambir (Lambir NP) di
Sarawak, Malaysia timur (4 ° 20'N, 113 ° 50'E), termasuk 6.800 ha terutama dataran
rendah, hutan campuran dipterocarpaceae tropis (MDF), mulai dari dekat permukaan
laut ke ketinggian 465 m, pada berpasir ke tanah liat-(Watson 1985). Kanopi hutan
heterogen adalah 40-60 m dan batang tingkat turnover yang tinggi (Phillips et al.
1994). Floristically, Lambir NP ini sangat beragam (Hall 1991; Davies & Becker
1996), dengan 1.175 jenis pohon D1 diameter cm setinggi dada (dbh) baru-baru ini
dicatat dalam survei 52-ha (LaFrankie et al 1995.).
Lambir NP menerima sekitar 3000 mm curah hujan per tahun, dengan semua
bulan rata-rata> 100 mm (Watson 1985). Suhu yang khas untuk daerah (berarti
maxima harian c. 32 ° C dan berarti minima harian c. 24 ° C) dan tidak menunjukkan
fluktuasi musiman yang besar.
Pada tahun 1991, sebuah proyek penelitian jangka panjang dimulai untuk
memantau spesies kayu di 52 ha MDF dataran rendah. Metodologi ini mirip dengan
yang digunakan pada Barro Colorado Island, Panama (Hubbell & Foster 1983), dan di
Pasoh Forest Reserve, barat Malaysia (Manokaran et al. 1990). Dengan demikian,
semua individu d1 cm d.b.h. yang ditandai, dipetakan untuk ± 10 cm, diidentifikasi
spesies dan diameter diukur untuk ± 1 mm (Condit 1995).
4
Page 6
Selama sensus awal (November 1991-November 1992) semua individu
Macaranga D1 cm dbh (n = 2882) diidentifikasi di lapangan untuk morfospesies.
Pohon di 8 ha yang kemudian diperiksa untuk memastikan korespondensi antara
morfospesies dan spesies taksonomi. Semua individu yang masih ada dari spesies
studi di 52-ha petak diukur kembali dan diberi identifikasi spesies yang benar sekitar
32 bulan setelah sensus awal. Namun, 192 orang mati batang tidak dapat percaya diri
diidentifikasi untuk spesies baik karena tidak ada bahan tanaman yang tersisa atau
nama field spesies 'telah tidak konsisten digunakan (Davies et al. 1995), dan mereka
dikeluarkan dari analisis.
Tujuh plot tambahan didirikan di daerah lain hutan primer dan sekunder di
Lambir NP. Individu dari semua kelas ukuran (termasuk batang ≤1 cm dbh) dari
spesies studi (n = 1132) di plot tambahan dipetakan dan diukur antara November 1991
dan Januari 1992. Sensus kedua dari semua plot dilakukan selama bulan Juli dan
Agustus 1994.
II.3 Analisis DataSelama sensus awal d.b.h. yang dari semua pohon di semua plot diukur.
Ketinggian pohon dari 8 ha plot 52-ha, dan semua plot tambahan, juga diukur. D.b.h.
yang dari semua pohon yang masih ada itu diukur kembali dalam sensus kedua.
Ketinggian semua pohon yang masih ada awalnya diukur untuk tinggi juga diukur
kembali, dan kumpulan data alometri itu dilengkapi dengan pengukuran tinggi dalam
lebih kurang 30 ha dari 52 ha petak-.
Untuk pohon D1 cm dbh, diameter diukur dengan terdekat 1 mm dengan pita
diameter rimbawan logam itu. Untuk bibit dan anakan ≤1 cm dbh, diameter
diperkirakan sebagai mean dari dua pengukuran calliper cepat sekitar 10% dari tinggi
batang. Diameter pohon diukur kembali di 4 ha dari 52 ha petak lama setelah sensus
awal dan akurasi ditemukan menjadi sangat tinggi. Ketinggian pohon ≤6 m tinggi
diukur langsung ke terdekat 1 cm, sedangkan ketinggian pohon tinggi diperkirakan
dengan clinometer (SUUNTO, Finlandia).
Karakteristik Lingkungan
Semua pohon di tujuh plot tambahan dan Subsamples pohon di 52-ha petak dinilai.
Lingkungan cahaya kanopi masing-masing individu yang dicirikan oleh indeks
5
Page 7
pencahayaan mahkota (indeks CI) pada skala 1-5, seperti dalam Clark & Clark
(1992).
Foto ikan-mata setengah bola diambil di berbagai tingkat kanopi-keterbukaan
dengan lensa Spiratone ikan-mata dipasang di tripod. Hitam dan putih negatif yang
dipindai dari gambar video dan dianalisis menggunakan 6.03c SOLARCALC
(Chazdon & Field 1985), dan digunakan untuk mengkalibrasi indeks CI (Tabel 1).
Tabel indeks 1. Crown iluminasi (CI) dikalibrasi terhadap persentase kanopi
keterbukaan dan tertimbang persentase kanopi keterbukaan diperkirakan dari (n)
foto mata ikan hemispherical. Huruf yang berbeda berikut nilai rata-rata
menunjukkan perbedaan yang signifikan pada P = 0,001 untuk anova pada data
log-transformasi.
indeks
CI
Crown deskripsi
lingkungan cahaya
% Kanopi
opennessmean
(SE)
Rata-rata tertimbang
% canopyopenness
(SE)
n
5 Crown ± benar-benar
terkena vertikal dan
lateral (misalnya besar
pembukaan hutan)
35.5 (3.9) a 45.3 (5.2) a 11
4 Mahkota terkena vertikal
dan beberapa cahaya
lateral (misalnya gap
hutan menengah)
15.8 (1.4) b 19.2 (1.9) b 13
3 Mahkota terkena
beberapa vertikal dan
beberapa cahaya lateral
(misalnya gap hutan
kecil)
7.4 (0.3) c 9.0 (0.5) c 34
2 Tidak ada cahaya vertikal
dan cahaya lateral yang
sedang (misalnya tepi
celah kecil)
5.1 (0.4) d 6.7 (0.5) d 24
1 Tidak ada cahaya lateral
yang vertikal dan
2.9 (0.3) e
6
Page 8
minimal (misalnya bawah
hutan)
4.1 (0.4) e 19
Pembentukan microsite dari setiap pohon Macaranga dicetak sebagai:
'terganggu' = di tanah terbuka tanpa gangguan jelas; = 'log' atau memiliki setidaknya
beberapa akar pohon utama langsung bersentuhan dengan kayu dan / atau cabang
utama berasal dari pohon atau cabang jatuh; 'tip-up' = pada atau terkait dengan
gundukan ujung-up atau akar-pit; 'Bencana longsor' = pada longsor sebuah; atau
'aluvial' = di situs aluvial. Metode ini meremehkan proporsi batang pada microsites
terganggu karena bukti gangguan menghilang dari waktu ke waktu. Karena kami tidak
punya perkiraan daerah relatif hutan tertutup oleh masing-masing jenis usaha,
perbandingan statistik hanya dilakukan antara spesies untuk jenis microsite.
Sebuah peta tanah dibangun untuk seluruh 52-ha petak sampling di tengah
masing-masing 1.300 20 m × 20 m kuadrat dalam plot. Tanah 5-15 cm dalam
ditugaskan ke salah satu dari empat kelas tekstur berdasarkan isi pasir jelas, lengket
dan licin tanah (Kimmins 1987). Sampel tanah (n = 145) yang mencakup semua
empat kelas tekstur dianalisis (Tabel 2) menggunakan Satuan LaMotte Tekstur tanah
(LaMotte Co, Chestertown, MD).
Tabel variasi tekstur 2. Tanah di plot 52-ha di Taman Nasional Bukit Lambir,
Sarawak, Malaysia. Berarti (± 1 SE) persentase pasir, lumpur dan tanah liat isinya
ditentukan untuk kelas tekstur tanah empat bidang. Ukuran sampel yang
diberikan dalam tanda kurung. Huruf yang berbeda berikut baris menunjukkan
perbedaan yang signifikan antara kelas tekstur tanah (G-tes; P <0,05)
tekstur kelas
tanah
% pasir % lumpur % tanah liat
1 39 ± 0.8 (56) 25 ± 0.8 (51) 36 ± 0.7 (51) a
2 45 ± 1.0 (23) 37 ± 1.5 (22) 19 ± 1.3 (22) b
3 51 ± 0.8 (35) 22 ± 0.7 (33) 27 ± 0.8 (33) ba
4 64 ± 1.4 (31) 23 ± 1.1 (29) 13 ± 1.0 (29) c
POHON ALLOMETRY
7
Page 9
Allometric hubungan antara diameter tinggi dan batang pohon dinilai
menggunakan serangkaian pendekatan Statistik.Untuk menguji untuk linearitas dari
ketinggian-diameter hubungan, polinomial urutan kedua cocok menggunakan regresi
kuadrat (LS) untransformed dan log-mengubah variabel (Niklas 1995). Hubungan
dianggap non-linear ketika masa urutan kedua adalah signifikan, dan ketinggian
maksimum asimtotik (Hmax) kemudian diperkirakan sebagai H = Hmax * [1 − exp
(−aDb)] mana H adalah pohon tinggi dalam m, D adalah d.b.h. cm, dan dan b
allometric konstanta yang mendekati nilai-nilai konstanta allometric standar untuk
nilai-nilai kecil h (Thomas 1995). Model ini tidak dapat diselesaikan untuk tiga
spesies; Ada juga beberapa besar individu untuk M. hypoleuca, dan tingginya M.
beccariana dan M. winkleri tidak muncul untuk mencapai asymptote sehubungan
dengan diameter. Spesies ini, maksimum pohon tinggi diperkirakan sebagai tinggi
berarti pohon-pohon terbesar 10 sampel (n = 6 untuk M. hypoleuca). Selama delapan
spesies lain metode ini memberikan nilai yang sama dari Hmax yang berasal dari
allometric model. Model allometric asimtotik dengan kesalahan standar diperkirakan
oleh regresi non-linear kuadrat menggunakan modul Nonlin di systat (Wilkinson
1990). Karena ada perubahan ukuran tergantung di lereng hubungan tinggi-diameter,
lereng bagian linier awal (5-40 mm d.b.h.) diperkirakan menggunakan allometric
model: log10 H = log10 B + A * log10 D, dimana H dan D batang tinggi dan diameter
m, dan B dan A konstanta allometric standar. Model ini diperkirakan menggunakan
keduanya berkurang utama axis (RMA) dan regresi kuadrat standar; hanya hasil RMA
yang ditampilkan sebagai pola pada dasarnya sama. Karena kesulitan Statistik
membandingkan parameter regresi non-linear (Ross 1981), perbandingan parameter
non-linear yang dibuat oleh memeriksa untuk tumpang tindih dari 95% confidence
interval.Linear lereng allometric dibandingkan dengan keseragaman lereng tes (Sokal
& Rohlf 1981).
8
Page 10
BAB III
PEMBAHASAN
III.1 Hasil Jurnal
HORISONTAL DISTRIBUSI SPESIES MACARANGA DALAM
HUBUNGANNYA DENGAN MIKRO
Lingkungan cahaya mahkota
Indeks CI sangat berkorelasi dengan kanopi keterbukaan seperti yang
diperkirakan dengan foto-foto setengah bola ikan-mata (Penombak dinaikkan pangkat
korelasi, rs = 0.82). Lima CI kelas rata-rata mewakili tingkat yang berbeda secara
signifikan kanopi keterbukaan (Tabel 1).
Keduanya berarti CI indeks (kisaran 4.2-2.0) dan proporsi pohon di setiap
kelas CI bervariasi antara spesies (Fig. 1), meskipun masing-masing spesies terjadi di
setidaknya empat kelas.Spesies menuntut lebih cahaya tinggi memiliki indeks CI
berarti dekat 4.0 (M. gigantea, M winkleri, dan M. hosei) dan spesies lebih toleran
naungan memiliki indeks CI berarti dekat 2.0 (M. lamellata dan M. kingii), dan
berada di kedua ujung sebuah kontinum dari distribusi sehubungan dengan kanopi
lingkungan cahaya (Fig. 1).
9
Page 11
Gambar 1. Persentase individu-individu yang terjadi di masing-masing kelas
iluminasi lima mahkota (Tabel 1) 11 spesies Macaranga. Spesies yang disusun dalam
rangka penurunan indeks berarti CI, dengan ukuran sampel untuk perkiraan indeks CI
dalam tanda kurung. Spesies ditunjukkan oleh satu huruf: E, M. gigantea; W, M.
winkleri; O, M. hosei; Y, M. hypoleuca; L, M. triloba; B, M. beccariana;
Trachyphylla M., V, M. havilandii; U, M. hullettii; M, M. lamellata; dan K, M. kingii.
es Macaranga juga berbeda secara signifikan dalam lingkungan cahaya mahkota pada
pohon berbeda ukuran (Tabel 3). Semua spesies, dengan pengecualian kecil-statured
M. havilandii, telah secara signifikan lebih besar eksposur light mahkota untuk
pohon-pohon besar. Antara kelas ukuran, namun, perbedaan antara spesies dalam
rata-rata indeks CI tetap dipertahankan (setidaknya di bawah 10 cm d.b.h.; Tabel
3).Urutan peringkat spesies CI indeks tidak berubah secara substansial dengan
meningkatkan ukuran pohon.Spesies lebih toleran naungan memiliki lebih rendah CI
indeks di seluruh semua ukuran kelas (Tabel 3).
10
Page 12
Spesi
11
Tabel 3.Berarti mahkota iluminasi (CI) indeks untuk spesies Macaranga 11 dalam tujuh kelas
ukuran diameter.Spesies diatur seperti Fig. 1 dengan ukuran sampel (n). Nilai-nilai
probabilitas (P *) merujuk pada Kruskal-Wallis non-parametrik anovas; nilai-nilai di ujung
baris dalam spesies perbandingan antara ukuran kelas, dan nilai-nilai di dasar kolom adalah
perbandingan antara spesies di setiap kelas ukuran
Tree diameter size classes0–2 cm 2–4 cm 4–6 c
m6–8 cm 8–10 cm 10–
15 cmd > 15 cm
CI n CI
n CI
n CI n CI
n CI
n CI n P*
gigantea
3.0
8 4.0 8 4.5
13 4.5
4 5.0 1 5.0
1 4.7
7 0.004**
winkleri
3.6 39
3.8
26 4.2
9 4.4 8 4.6
10 4.6
11
– – 0.001**
hosei 3.4 59
4.2
31 4.8
5 4.5 2 – – 5.0
2 5.0 16
<0.001**
hypoleuca
2.6 14
3.8
11 4.0
4 4.0 1 – – 5.0
1 5.0 4 0.001**
triloba 2.7 103
3.4
40 4.0
23
4.4 8 4.4
21 4.5
20
– – <0.001**
beccariana
3.0 107
3.4
58 4.0
28
4.1 17
4.0
7 4.2
4 – – <0.001**
trachyphylla
2.9 96
3.2
57 3.4
23
3.4 20
4.0
5 4.9
19
4.3 6 <0.001**
havilandii
2.8 98
2.7
4 – – – – – – – – – – 0.96
hullettii
1.9 37
2.2
78 3.2
58
3.3 32
3.2
19 3.6
5 – – <0.001**
lamellata
1.5 61
2.1
116 2.5
54
2.8 14
2.9
8 – – – – <0.001**
kingii 1.7 39
2.3
23 2.7
6 2.7 3 – – – – – – 0.004**
P* <0.001**
<0.001**
<0.001**
<0.001**
<0.001**
Page 13
Pendirian microsites
Ada perbedaan yang signifikan antara Macaranga spesies dalam proporsi
pohon di kelas lima pendirian (Tabel 4). Pendirian situs terganggu bervariasi antara
12% dan 43% dari pohon-pohon di antara spesies 11. Empat spesies paling toleran
naungan, M. havilandii, M. hullettii, M. lamellata, dan M. kingii (Fig. 1), memiliki
proporsi yang lebih besar (80-88%) dari orang-orang yang tumbuh di situs tampaknya
terganggu daripada spesies lebih menuntut cahaya (57 – 76%). Antara spesies,
proporsi batang dalam cahaya rendah (CI = 1) sangat positif berkorelasi dengan
proporsi batang di tanah tidak terganggu (Pearson, r = 0.82, P = 0,002). Ini mungkin
diharapkan karena gangguan kanopi yang mengakibatkan peningkatan ketersediaan
ringan sering mengakibatkan gangguan di permukaan tanah.Namun, ada dua spesies
(M. winkleri dan M. beccariana) mana pohon situs terganggu memiliki indeks CI
yang secara signifikan lebih rendah daripada di microsites terganggu (Kruskal-Wallis
tes, P < 0.01).
Tabel 4. Persentase (%) dan jumlah (n) individu dari 11 jenis Macaranga di lima microsites pendirian. Probabilitas (P *) mengacu pada tes Chi-kuadrat dari perbedaan antara spesies dalam proporsi pohon di microsites 'tidak terganggu'. Spesies yang diatur seperti gambar 1
Establishment micrositeUndisturbed Logs Tip-up Landslip Alluvial
Species % n % n % n % n % ngigantea 71.7 33 13.0 6 8.7 4 6.5 3 0.0 0winkleri 71.8 79 15.5 17 10.9 12 1.8 2 0.0 0hosei 74.2 112 21.2 32 4.0 6 0.7 1 0.0 0hypoleuca 56.6 30 17.0 9 13.2 7 11.3 6 1.9 1triloba 59.4 224 8.0 30 0.5 2 2.4 9 29.7 112beccariana 65.6 320 17.4 85 8.4 41 8.6 42 0.0 0trachyphylla 76.1 392 15.3 79 4.5 23 4.1 21 0.0 0havilandii 79.7 94 11.9 14 8.5 10 0.0 0 0.0 0hullettii 82.6 214 11.2 29 2.3 6 3.9 10 0.0 0lamellata 88.4 244 10.9 30 0.4 1 0.4 1 0.0 0kingii 87.5 98 1.8 2 0.0 0 0.0 0 10.7 12P* <0.001**
Spesies diferensial didistribusikan dalam jenis gangguan.Macaranga triloba
telah hampir 30% dari batang yang di aluvial atau secara berkala tergenang situs,
12
Page 14
landslips tua telah relatif lebih batang M. hypoleuca dan M. beccariana dari spesies
lainnya, dan gundukan tip-up yang relatif kaya M. hypoleuca dan M. winkleri (Tabel
4).
Pohon berbagai ukuran mungkin terjadi dalam pendirian berbeda microsites
karena mempertahankan baik diferensial menurut pendirian situs, atau fosil fluktuasi
dalam ketersediaan situs pendirian.Analisis kami untuk spesies dengan ukuran sampel
besar (data tidak ditampilkan) menyarankan bahwa M. beccariana telah secara
signifikan lebih besar individu di landslips (P < 0.001), mungkin karena fosil variasi
karena situs tersebut jarang Lambir NP.Sebaliknya, M. hosei, M. triloba dan M.
trachyphylla situs terganggu (pada log, tip-up dan landslips) telah secara signifikan
lebih besar proporsi individu dalam ukuran kecil atau menengah kelas dari kelas
ukuran lebih besar (P < 0.01), menyarankan mempertahankan diferensial.Namun,
pohon yang lebih besar bisa juga telah menetapkan setelah gangguan bukti-bukti yang
tidak tetap.
Tekstur tanah
Kelas tekstur tanah Field-ditentukan sangat berhubungan dengan perbedaan
dalam kandungan tanah pasir, meningkat dari kelas 1 sampai 4 (Tabel 2). Sekitar 65%
dari petak 1300 dalam 52-ha plot diklasifikasikan di pasir-kaya kelas 4 dan 11% di
tanah liat kaya kelas 1 (Tabel 5). Distribusi sembilan dari 11 jenis Macaranga secara
signifikan bias terhadap tanah tekstur kelas (Tabel 5). Tujuh spesies (M. beccariana,
M. hosei, M. hypoleuca, M. kingii, M. trachyphylla, M. triloba dan M. winkleri)
memiliki individu-individu secara signifikan lebih dari yang diharapkan dalam petak
tanah liat kaya lebih, sedangkan M. lamellata dan M. havilandii punya signifikan
lebih individu di petak pasir yang kaya.
Tabel 5.Distribusi spasial dari 11 jenis Macaranga terhadap tanah tekstur variasi (Lihat
tabel 2) di Taman Nasional Lambir Hills, Sarawak, Malaysia. Proporsi 1300 52-ha petak
20 m × 20 m plot di kelas empat tanah yang tercantum pada baris pertama. Untuk masing-
masing spesies kerapatan individu dalam setiap kelas tanah adalah terdaftar (pohon ha−1).
N adalah jumlah total petak di plot, dan jumlah individu untuk setiap spesies. Probabilitas
(P *) merujuk pada G-tes Apakah spesies distribusi yang berbeda dari yang diharapkan
berdasarkan frekuensi jenis tanah di dalam plot. Spesies yang diatur seperti gambar 1
13
Page 15
Soil texture classes1 2 3 4 N P*
Clay-rich Sand-rich52-ha plot 11.1 14.2 9.4 65.4 1300 –Speciesgigantea 1.2 0.5 0.8 0.4 29 0.16winkleri 8.0 1.9 1.6 1.1 105 <0.001hosei 4.3 3.0 6.1 2.1 147 <0.001hypoleuca 3.0 1.9 0.2 0.8 60 <0.001triloba 5.7 2.0 2.5 2.0 127 <0.001beccariana 18.9 11.8 10.7 7.4 499 <0.001trachyphylla 26.2 17.5 18.0 12.0 775 <0.001havilandii 0.9 0.5 2.9 1.6 77 0.007hullettii 8.2 9.1 10.7 6.9 399 0.053lamellata 2.1 7.1 6.6 7.4 347 <0.001kingii 6.3 3.5 2.5 1.5 125 <0.001
Spesies dengan ukuran sampel yang lebih besar kami menguji apakah ada
hubungan antara kelas tekstur tanah dan ketersediaan cahaya (diperkirakan dari indeks
CI). Dalam kasus tidak (n = 7 jenis) ini berarti CI indeks secara signifikan berbeda
untuk individu pada kelas tekstur tanah yang berbeda (P > 0.1, tes Kruskal – Wallis).
VERTIKAL DISTRIBUSI SPESIES MACARANGA
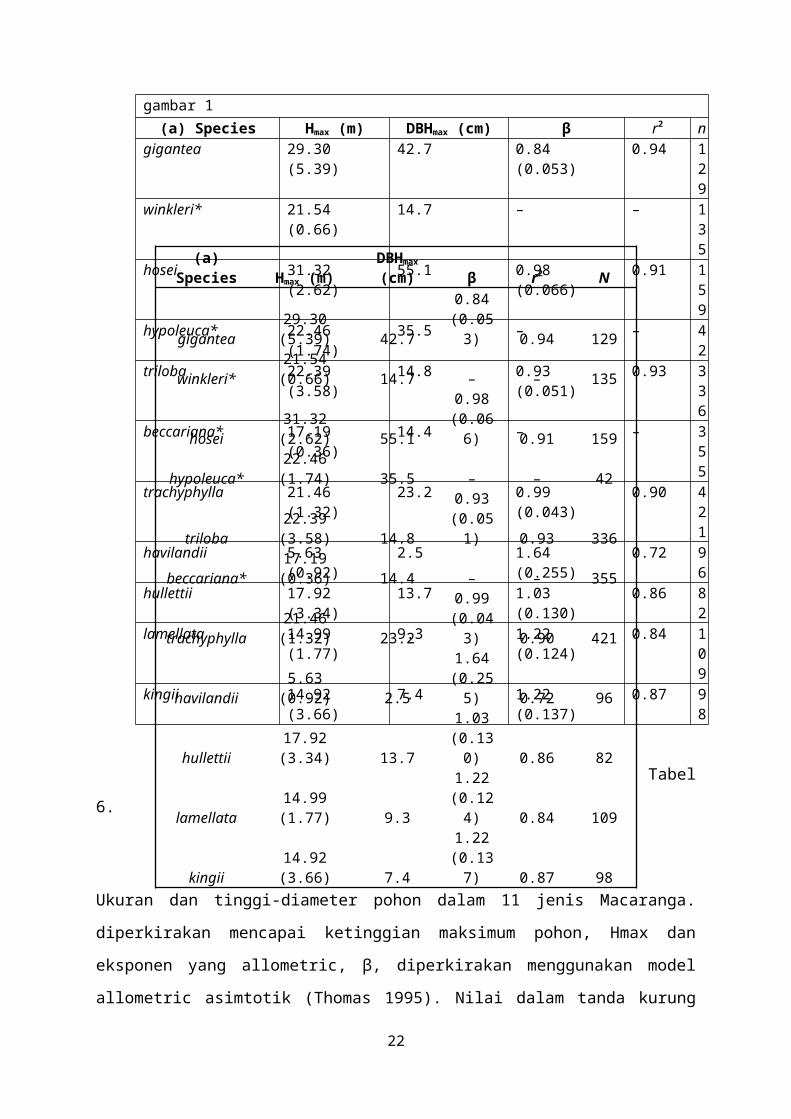
Ukuran maksimum pohon dan allometry dari pohon tinggi dan diameter
bervariasi antara spesies Macaranga (Tabel 6). Semua 11 spesies menunjukkan
perubahan ukuran tergantung yang signifikan di lereng hubungan tinggi-diameter
pohon, seperti yang ditunjukkan oleh regresi kuadrat (P < 0,05). Hal ini karena
kecenderungan untuk pohon tinggi untuk mencapai asymptote dan diameter menjadi
tak tentu (King 1990a).Namun, di M. winkleri dan M. beccariana asymptote sudah
jauh melampaui data diamati dan karena itu dianggap perkiraan ketinggian maksimum
tidak masuk akal.
Tabel 6.Pohon allometry ukuran dan tinggi-diameter dalam 11 jenis Macaranga.() diperkirakan mencapai ketinggian maksimum pohon, Hmax dan eksponen yang allometric, β, diperkirakan menggunakan model allometric asimtotik (Thomas 1995).Nilai dalam tanda kurung adalah kesalahan standar. DBHmax maksimum diamati d.b.h. Untuk spesies yang bertanda asterisk, Hmax dihitung dengan menggunakan nilai rata-rata dari pohon yang lebih besar seperti yang dijelaskan dalam teks. (b) allometry sedang berkembang tinggi-diameter 11 spesies Macaranga dalam ukuran kisaran 5-40 mm d.b.h. Awal lereng dan y-intercept tinggi × diameter hubungan dinilai dengan RMA regresi data log10 berubah. Nilai dalam tanda kurung adalah 95%
14
Page 16
confidence interval untuk koefisien regresi.Ukuran sampel (n) dan r2 terdaftar untuk analisis kedua. Spesies yang diatur seperti gambar 1
(a) Species Hmax (m) DBHmax (cm) β r2 ngigantea 29.30 (5.39) 42.7 0.84 (0.053) 0.94 1
29
winkleri* 21.54 (0.66) 14.7 – – 135
hosei 31.32 (2.62) 55.1 0.98 (0.066) 0.91 159
hypoleuca* 22.46 (1.74) 35.5 – – 42
triloba 22.39 (3.58) 14.8 0.93 (0.051) 0.93 336
beccariana* 17.19 (0.36) 14.4 – – 355
trachyphylla 21.46 (1.32) 23.2 0.99 (0.043) 0.90 421
havilandii 5.63 (0.92) 2.5 1.64 (0.255) 0.72 96
hullettii 17.92 (3.34) 13.7 1.03 (0.130) 0.86 82
lamellata 14.99 (1.77) 9.3 1.22 (0.124) 0.84 109
kingii 14.92 (3.66) 7.4 1.22 (0.137) 0.87 98
Tabel 6. Ukuran dan tinggi-diameter pohon dalam 11 jenis Macaranga. diperkirakan
mencapai ketinggian maksimum pohon, Hmax dan eksponen yang allometric, β, diperkirakan
menggunakan model allometric asimtotik (Thomas 1995). Nilai dalam tanda kurung adalah
kesalahan standar. DBHmax maksimum diamati d.b.h. Untuk spesies yang bertanda asterisk,
Hmax dihitung dengan menggunakan nilai rata-rata dari pohon yang lebih besar seperti yang
dijelaskan dalam teks. (b) allometry sedang berkembang tinggi-diameter 11 spesies
Macaranga dalam ukuran kisaran 5-40 mm d.b.h. Awal lereng dan y-intercept tinggi ×
diameter hubungan dinilai dengan RMA regresi data log10 berubah. Nilai dalam tanda
kurung adalah 95% confidence interval untuk koefisien regresi. Ukuran sampel (n) dan r2
terdaftar untuk analisis kedua. Spesies yang diatur seperti gambar 1.
15
(a) Species Hmax (m) DBHmax (cm) β r2 N
gigantea 29.30 (5.39) 42.70.84
(0.053) 0.94 129winkleri* 21.54 (0.66) 14.7 – – 135
hosei 31.32 (2.62) 55.10.98
(0.066) 0.91 159
hypoleuca* 22.46 (1.74) 35.5 – – 42
triloba 22.39 (3.58) 14.80.93
(0.051) 0.93 336
beccariana* 17.19 (0.36) 14.4 – – 355
trachyphylla 21.46 (1.32) 23.20.99
(0.043) 0.90 421
havilandii 5.63 (0.92) 2.51.64
(0.255) 0.72 96
hullettii 17.92 (3.34) 13.71.03
(0.130) 0.86 82
lamellata 14.99 (1.77) 9.31.22
(0.124) 0.84 109
kingii 14.92 (3.66) 7.41.22
(0.137) 0.87 98
Page 17
Perkiraan ketinggian maksimum pohon (Hmax) berkisar antara 5,5-31.3 m dan
diameter batang maksimum berkisar 2,5-55.1 cm d.b.h. di antara spesies Macaranga
11 (tabel 6a). Macaranga gigantea, dan M. hosei adalah pada salah satu ujung sebuah
kontinum dari pohon ukuran, dengan pohon-pohon dewasa sampai kira-kira 30 m
tingginya dan maksimum diameter batang > 40 cm. Macaranga hypoleuca mirip
dengan spesies ini (S. J. Davies, pengamatan pribadi), meskipun beberapa orang besar
yang terjadi dalam sampel kami. Macaranga havilandii adalah di sisi ekstrem lain,
dengan perkiraan Hmax 5.6 m dan d.b.h. maksimum dari hanya 2,5 cm. Macaranga
lamellata dan M. kingii pohon-pohon yang sedikit lebih besar dengan Hmax 14-15 m
dan diameter batang maksimum 7-10 cm d.b.h. Lima spesies lainnya (M. winkleri, M.
triloba, M. beccariana, M. trachyphylla dan M. hullettii) adalah menengah dalam
ukuran, dengan Hmax mulai dari 17 hingga 23 m dan maksimum d.b.h. 13-23 cm
(tabel 6a). Antara spesies, Hmax sangat positif berkorelasi (r = 0.85, P = 0.001)
dengan proporsi pohon di lingkungan cahaya tinggi (CI = 5).
Keseragaman lereng tes menunjukkan bahwa ada perbedaan yang signifikan
antara spesies Macaranga di lereng dan sumbu y sedang berkembang tinggi-diameter
allometry model (tabel 6b; tes tidak ditampilkan). Selain itu, lereng awal yang
diperkirakan dari model asimtotik berbeda secara signifikan antara spesies (tabel 6a).
Perkiraan ketinggian maksimum pohon negatif berhubungan dengan kedua-sumbu y
(r = −0.79, P = 0.003), dan lereng awal (r = −0.68, P = 0.018) tinggi-diameter
hubungan, menunjukkan bahwa anakan spesies kecil-statured (dan dalam kasus ini
lebih toleran naungan spesies) yang lebih ramping daripada bibit dari spesies besar-
statured (Tabel 6).
Sebagai anakan spesies lebih toleran naungan itu rata-rata yang hidup di
tingkat cahaya yang lebih rendah daripada spesies menuntut lebih tinggi-cahaya (Fig.
16
(b) Species Slope y-intercept r2 Ngigantea 1.00 (0.12) 2.17 (0.24) 0.88 36winkleri 1.10 (0.13) 2.45 (0.24) 0.86 41hosei 0.76 (0.08) 1.86 (0.15) 0.78 80
hypoleuca 0.96 (0.29) 2.29 (0.51) 0.65 19triloba 1.09 (0.10) 2.41 (0.18) 0.80 90
beccariana 1.01 (0.06) 2.30 (0.11) 0.80 235
Page 18
1), mungkin bahwa allometries berbeda sedang berkembang adalah karena lingkungan
cahaya daripada spesies. Meskipun ukuran sampel terbatas, keseragaman lereng tes
menunjukkan tidak ada perbedaan yang signifikan dalam tinggi-diameter hubungan
antara tingkat cahaya untuk lima dari tujuh spesies lebih berlimpah (data tidak
ditampilkan). Hanya M. hosei telah secara signifikan lebih besar allometric lereng di
tingkat cahaya rendah. Analisis ini menunjukkan bahwa spesies yang berbeda, tetapi
ukuran sampel yang lebih besar diperlukan untuk menilai pentingnya lingkungan
cahaya untuk Macaranga sedang berkembang allometries.
III.2 Pembahasan
DISTRIBUSI HORISONTAL
Korelasi kuat ditemukan antara ikan-mata fotografi perkiraan tingkat cahaya
kanopi dan sewenang-wenang mahkota iluminasi indeks. Berdasarkan indeks ini, ada
perbedaan yang signifikan antara Macaranga spesies dalam distribusi individu
sehubungan dengan tingkat cahaya, menyarankan berbagai warna toleransi dalam
kelompok ini pohon hutan hujan tropis. Spesies studi kami berkisar dari sangat cahaya
tinggi menuntut perintis khas kesenjangan besar hutan dan hutan sekunder habitat
(misalnya Macaranga gigantea), melalui apa mungkin dianggap spesies kecil-celah
(misalnya M. trachyphylla), sangat toleran naungan pohon yang bertahan dalam
understorey (misalnya M. kingii). Namun, semua spesies memiliki setidaknya
beberapa individu dalam empat dari lima kelas CI, menunjukkan tumpang-tindih yang
besar, terutama di antara cahaya tinggi lebih menuntut spesies. Dalam sebuah analisis
sebelumnya, menggunakan bibit sembilan dari 11 spesies tumbuh di lingkungan yang
seragam, indeks naungan toleransi ditemukan sangat berkorelasi dengan sifat
fotosintetik dan lain-lain ecophysiological (Davies 1998).
Rata-rata tingkat cahaya dicegat oleh individu-individu yang meningkat
melalui ontogeny di 10 dari 11 spesies Macaranga. Hasil yang sama telah ditemukan
dalam studi non-pelopor spesies (Clark & Clark 1992). Di sebagian besar spesies
Macaranga peningkatan ontogenetik ini adalah karena kedua angka kematian lebih
tinggi dalam lingkungan cahaya rendah (Davies 1996), dan untuk meningkatkan
tingkat cahaya dicegat sebagai pohon tumbuh kearah tajuk (Aoki et al. 1978; Chazdon
1986). Hasilnya adalah bahwa indeks CI berbeda antara spesies di bawah 10 cm d.b.h.
17
Page 19
tetapi tidak di atas ukuran ini (dengan jenis kecil dikecualikan). Ada tidak ada bukti
untuk penurunan indeks CI pada ukuran pohon yang lebih besar, untuk spesies yang
mungkin mengindikasikan over topping karena penutupan kanopi.
Perkiraan kualitatif mahkota lingkungan cahaya (seperti indeks CI) mungkin
tidak sebanding antara studi, terutama jika Indeks tidak dikalibrasi terhadap beberapa
standar seperti ikan-mata foto (Clark et al. 1993). Namun, kita perhatikan bahwa
Clark & Clark (1992), menggunakan analisis lingkungan cahaya yang serupa tetapi
lebih halus untuk spesies mulai dari sangat toleran naungan cukup cahaya-menuntut,
menunjukkan perbedaan yang signifikan antara spesies di microsite hunian
sehubungan dengan mahkota iluminasi. Selain itu, Clark et al. (1993) menemukan
perbedaan yang signifikan antara perintis Cecropia obtusifolia dan C. insignis
distribusi sedang berkembang dengan menghormati untuk lingkungan cahaya.
Brokaw (1987) juga menggambarkan perbedaan dalam persyaratan ukuran celah
antara tiga jenis pionir di Panama, dan perbedaan-perbedaan dalam distribusi jenis
pionir dalam kesenjangan (Popma et al. 1988) menyarankan kebutuhan cahaya yang
berbeda. Sejumlah penelitian lain (misalnya Riddoch et al. 1991; Kitajima 1994;
Reich et al. 1994) telah menemukan perbedaan dalam ecophysiological tanggapan
terhadap cahaya antara tropis spesies perintis, tetapi beberapa telah memeriksa
mereka konsekuensi bagi hutan distribusi spasial (Davies 1998).
Spesies Macaranga berbeda secara signifikan dalam proporsi batang pada
microsites pendirian berbeda. Spesies lebih toleran naungan memiliki batang yang
lebih sedikit terganggu situs daripada spesies menuntut lebih tinggi-cahaya. Ini bukan
hanya karena korelasi antara situs tidak terganggu dan tingkat cahaya rendah karena
pohon dari hanya dua jenis telah secara signifikan lebih rendah berarti CI indeks pada
terganggu dari situs terganggu. Spesies menuntut cahaya tinggi tampaknya lebih
memilih untuk membangun situs terganggu terlepas dari tingkat cahaya. Apakah ini
karena lebih tinggi germinability benih kecil (Davies 1996), diferensial benih hujan
(Levey 1988) atau persyaratan gangguan fisik untuk pengecambahan tidak diketahui.
Dalam kesenjangan, Ellison et al. (1993) kepadatan bibit tertinggi ditemukan untuk
kecil-unggulan Melastomataceae di tanah mineral terkena akar lubang dan gundukan,
meskipun tingkat cahaya lebih rendah dan ketersediaan unsur hara mungkin telah
lebih rendah ini microsites (Vitousek & Denslow 1986). Putz (1983) ditemukan lebih
tinggi jenis pionir kolonisasi terganggu tanah daripada tanah tidak terganggu celah-
celah di Panama. Antara cahaya tinggi lebih menuntut Macaranga spesies, ada
18
Page 20
perbedaan besar dalam proporsi pohon di microsites 'terganggu' berbeda (log, tip-up
dan landslips). Núñez-Farfan & Dirzo (1988) menemukan perbedaan dalam distribusi
dan kinerja berikutnya jenis pionir Cecropia obtusifolia dan Heliocarpus
appendiculatus antara crown dan akar zona hutan kesenjangan, meskipun keumuman
pola ini memerlukan studi lebih lanjut seperti zona unreplicated. Ada perbedaan yang
signifikan antara spesies, tetapi pendekatan skala yang lebih halus diperlukan untuk
menilai relatif pentingnya kondisi fisik untuk pembentukan dan tingkat cahaya dalam
menentukan variasi dalam distribusi horisontal dan kinerja untuk menuntut cahaya
Macaranga spesies yang lebih tinggi.
Biasanya, tanah variasi menyediakan skala kasar heterogenitas sumber daya di
hutan tropis dari pendirian kondisi (Newbery & Proctor 1984; Baillie et al. 1987;
Swaine 1996). Interbedding batu pasir dan serpih batu di situs studi, bagaimanapun,
berarti bahwa tanah di sini bervariasi skala spasial cukup kecil (Watson 1985).
Mencolok, distribusi sembilan dari 11 spesies Macaranga yang signifikan bias
terhadap tekstur tanah, dan hanya salah satu spesies yang ada adalah ukuran sampel
besar, M. hullettii, adalah tidak berkorelasi (P = 0.053). Tekstur tanah berkorelasi
dengan kedua akar mat kedalaman dan daun sampah ketebalan di situs Studi
(Palmiotto 1995). Sambil tanah kadar hara telah ditunjukkan untuk menjadi lebih
tinggi pada bertekstur halus (lebih clay-kaya) tanah di Lambir NP (Hall 1991; Ashton
& Hall 1992), tanah juga memiliki akar-mat tipis dan lapisan humus, dan mungkin
juga berbeda di tanah moisture ketersediaan (Newbery et al. 1996). Percobaan karena
itu diperlukan untuk menyelidiki dasar tanah preferensi di antara spesies Macaranga
ini. Di Malesia barat, Macaranga jauh lebih beragam dan berlimpah dalam suksesi
sekunder lebih kaya gizi situs (serpih dan basal-berasal tanah) daripada situs gizi-
miskin (berasal dari batu pasir tanah), dan hampir tidak ada dari Adinandra-belukar,
masyarakat successional sangat depauperate sekunder Malaya dan Singapura (Wyatt-
Smith 1963; SIM et al., 1992).
PENYEBARAN VERTIKAL
Ada perbedaan yang signifikan antara spesies pohon maksimum ukuran
(Hmax dan Dmax), dan lereng dan y-intercept sedang berkembang tinggi-diameter
hubungan (Tabel 6). Antara spesies, Hmax negatif berhubungan dengan toleransi
naungan; tiga spesies statured kecil itu juga paling toleran naungan (Fig. 2). Thomas
(1993) menemukan hubungan yang positif antara Hmax dan cahaya-jenuh tingkat
19
Page 21
fotosintesis untuk 25 spesies empat genera pohon hutan hujan Malaysia, meskipun
hubungan antara harga fotosintetik dan naungan-toleransi dan distribusi bibit dan bibit
terhadap lingkungan cahaya tidak dipelajari. Meskipun hubungan antara warna-
toleransi dan pohon maksimum ukuran untuk pohon-pohon tropis ini tidak berarti
umum, dengan sejumlah kecil statured cahaya tinggi demanders (Swaine & Whitmore
1988; Clark & Clark 1992), pola yang terlihat dalam genera empat dipelajari oleh
Thomas (1993) diulang dalam hasil kami dengan Macaranga. Apakah pola ini terjadi
di lain genera (atau clades) memerlukan studi lebih lanjut.
Gambar 2. Perwakilan dari tiga dimensi distribusi spesies Macaranga 11
perkiraan maksimum pohon tinggi, proporsi batang di tingkat cahaya tinggi (CI kelas
5) dan proporsi pohon di tanah pasir-kaya (tanah tekstur kelas 4). Spesies ditunjukkan
oleh satu huruf dalam gambar 1.
Meskipun Hmax negatif berkorelasi dengan naungan toleransi, ada variasi
dalam Hmax di antara spesies Macaranga lebih toleran naungan. Macaranga gigantea
dan M. hosei tumbuh setinggi 25-30 m dan > d.b.h. 40 cm, sedangkan M. winkleri
dan M. beccariana jarang mencapai 20 m tinggi dan d.b.h. 15 cm, namun semua
empat spesies sering sympatric dalam lingkungan cahaya sangat tinggi di awal suksesi
sekunder. Empat spesies menetapkan awal setelah pembentukan celah, dan
berkembang sangat pesat. Macaranga beccariana dan M. winkleri memulai reproduksi
pada pohon ukuran sekitar 5-6 cm d.b.h. dan mulai senesce di sekitar 10 cm d.b.h.,
sementara M. gigantea dan M. hosei tidak mulai untuk mereproduksi sampai 10 cm
d.b.h. dan bertahan dalam hutan kesenjangan selepas kematian tetangga pohon M.
winkleri dan M. beccariana (Davies 1996). Ini terkait erat spesies jelas berbagi
sumber daya serupa, tetapi peran disarankan temporal atau vertikal stratifikasi sumber
daya di spesies koeksistensi harus diuji oleh analisis lebih rinci penggunaan sumber
daya diferensial mereka seluruh ontogeny. Pertanyaan yang lebih luas dari sejarah
evolusi ini perbedaan dalam ciri-ciri sejarah hidup mungkin juga dibahas dengan
menggunakan pendekatan filogenetik (Davies 1996).
Perkiraan maksimum pohon tinggi negatif berhubungan dengan lereng sedang
berkembang tinggi-diameter hubungan antara spesies Macaranga 11. Meskipun
ketersediaan cahaya Apakah pengaruh allometries sedang berkembang pada beberapa
spesies, anakan spesies statured kecil selalu cenderung memiliki batang yang lebih
ramping daripada anakan spesies statured yang lebih besar. Pola ini juga ditemukan
untuk 37 spesies pohon hutan hujan bebas-pelopor Malaysia (Thomas 1995). Namun,
20
Page 22
pola berlawanan ditemukan untuk hutan hujan sedang berkembang allometries di
Panama (King 1990b), dimana dua spesies understorey dilaporkan memiliki batang
tebal pada ketinggian tertentu sedang berkembang daripada empat kanopi spesies. Hal
ini berpendapat bahwa ini adalah karena kebutuhan spesies understorey untuk
mendukung mahkota biomassa dan dedaunan area yang lebih besar untuk
memaksimalkan cahaya intersepsi. Thomas (1995), di sisi lain, menunjukkan bahwa
batang lebih ramping spesies understorey statured kecil dalam studi dapat
dipertanggungjawabkan oleh kepadatan kayu mereka umumnya lebih besar dan
karenanya kekuatan, dan manfaat yang tidak proporsional, dalam hal pencegatan
cahaya, yang merupakan cangkokan dari spesies understorey mungkin mendapatkan
dengan memaksimalkan tinggi kenaikan vs diameter kenaikan. Dalam kasus tiga kecil
toleran naungan Macaranga spesies dalam studi ini, semua memiliki arsitektur yang
agak sederhana dengan sejumlah kecil menengah dan daun unlobed yang sangat besar
pada umumnya (S.J. Davies, pengamatan pribadi). Analisis lebih lanjut antar-
hubungan antara tampilan daun, kanopi arsitektur dan allometry sedang berkembang,
dan bagaimana mereka merespons lingkungan cahaya sedang berkembang, diperlukan
untuk menilai relatif pentingnya memaksimalkan diameter batang sehubungan dengan
tinggi (untuk mengaktifkan tampilan biomassa besar daun) vs memaksimalkan
pertumbuhan tinggi (untuk meningkatkan pencegatan cahaya).
Secara keseluruhan, 11 spesies Macaranga yang sympatric pada skala hutan
menunjukkan berbagai macam interspesifik perbedaan dalam kedua pola distribusi
horisontal dan vertikal. Gambar 2 memberikan ringkasan diagram dari pola-pola ini
berfokus pada tiga sumbu utama variasi dalam pola distribusi, mahkota tingkat cahaya
(berdasarkan proporsi pohon di kelas indeks CI tertinggi), jenis tanah distribusi
(berdasarkan proporsi pohon di tanah pasir kaya) dan diperkirakan ketinggian
maksimum pohon. Ada sebuah kontinum dari spesies hunian microsites, dari kecil
statured dan lebih toleran naungan spesies (M. kingii, M. lamellata, M. hullettii, dan
M. havilandii) yang berbeda sangat pada preferensi tekstur tanah, untuk spesies tujuh
lainnya yang memiliki tinggi tuntutan untuk sumber cahaya dan tanah, tapi berbeda
secara signifikan dalam ukuran maksimum pohon, dan oleh karena itu waktu yang
spesies individu mendominasi suksesi. Heterogenitas ketersediaan sumber daya di
hutan dan tingginya jelas spesialisasi spesies ini menunjukkan bahwa semua tiga
sumbu distribusi penting dalam mempengaruhi koeksistensi kelompok beragam
pohon-pohon successional yang awal. Bagaimana variasi ini dalam sumber daya
21
Page 23
heterogenitas mempengaruhi kinerja baik individu dan peraturan dinamika populasi
spesies ini akan memiliki pengaruh yang kuat pada pola pewarisan sekunder di hutan
Kalimantan.
III.3 Keterbatasan PenelitianAdapun beberapa hal yang membatasi keleluasaan penelitian ini yaitu,
penelitian ini hanya menggunakan objek pada hutan tropis dengan beberapa spesies.
Penelitian ini terbatas dengan waktu karena menguji tumbuh kembangnya beberapa
spesies tumbuhan. Penelitian ini terbatas pada tempat dimana sasarannya adalah hutan
tropis yang berada di Macaranga Kalimantan.
22