Research ArticleAnalysis of Argonaute Complex Bound mRNAs in DU145Prostate Carcinoma Cells Reveals New miRNA Target Genes
Jaroslaw Szczyrba12 Volker Jung3 Michaela Beitzinger4 Elke Nolte5
Sven Wach5 Martin Hart1 Sandra Sapich1 Marc Wiesehoumlfer2 Helge Taubert5
Gunther Wennemuth2 Norbert Eichner4 Thomas Stempfl6
Bernd Wullich5 Gunter Meister4 and Friedrich A Graumlsser1
1 Institute of Virology Saarland University Medical School Kirrbergerstrasse Haus 47 66421 HomburgSaar Germany2Institute of Anatomy University Hospital Essen Hufelandstrasse 55 45147 Essen Germany3University Clinic of Urology Saarland University Medical School Kirrbergerstrasse 66421 HomburgSaar Germany4Biochemistry Center Regensburg (BZR) Laboratory for RNA Biology 93053 Regensburg Germany5University Clinic of Urology Friedrich-Alexander-University Erlangen-Nurnberg Krankenhausstrasse 12 91054 Erlangen Germany6Center of Excellence for Fluorescent Bioanalytics (KFB) University of Regensburg 93053 Regensburg Germany
Correspondence should be addressed to Jaroslaw Szczyrba jaroslawszczyrbauk-essendeand Friedrich A Grasser graesserukseu
Received 27 May 2016 Accepted 30 October 2016 Published 5 January 2017
Academic Editor Zhenyu Jia
Copyright copy 2017 Jaroslaw Szczyrba et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Posttranscriptional gene regulation by microRNAs (miRNAs) contributes to the induction and maintenance of prostate carcinoma(PCa) To identifymRNAs enriched or removed fromAgo2-containingRISC complexes these complexeswere immunoprecipitatedfromnormal prostate fibroblasts (PNFs) and the PCa lineDU145 and the boundmRNAswere quantified bymicroarrayThe analysisof Ago complexes derived from PNFs or DU145 confirmed the enrichment or depletion of a variety of mRNAs already known fromthe literature to be deregulated Novel potential targets were analyzed by luciferase assays with miRNAs known to be deregulatedin PCa We demonstrate that the mRNAs of the death effector domain-containing protein (DEDD) the tumor necrosis factorreceptor superfamily member 10b protein (TNFRSF10B) the tumor protein p53 inducible nuclear protein 1 (TP53INP1) and thesecreted protein acidic cysteine-rich (SPARC osteonectin) are regulated by miRNAs miR-148a miR-20a miR-24 and miR-29abrespectivelyTherefore these miRNAs represent potential targets for therapy Surprisingly overexpression of miR-24 induced focusformation and proliferation of DU145 cells while miR-29b reduced proliferation The study confirms genes deregulated in PCa byvirtue of their presenceabsence in the Ago2-complex In conjunction with the already published miRNA profiles of PCa the datacan be used to identify miRNA-regulated mRNAs
1 Introduction
Prostate carcinoma (PCa) is a leading cause of cancermorbid-ity andmortality inmenworldwide [1]The etiology of PCa iscomplex and numerous genetic [2] and epigenetic alterations[3] have previously been reported
The elucidation of possible functional consequences ofgenetic or epigenetic alterations is still a great challengeMicroRNAs (miRNAs) are short noncodingRNAs of 18ndash23 ntlength that posttranscriptionally regulate gene expression
by binding to mRNA targets to inhibit protein synthesis(reviewed in [4]) In the context of tumorigenesis miRNAscan function both as tumor suppressors and as oncogenes [5]They are bound to their target mRNAs in the RISC complexwhich contains anArgonaute (Ago) protein that is tethered tothemRNAby themiRNA Binding of themiRNA to the targetinitially inhibits protein synthesis by reducing translation buteventually leads to degradation of the mRNA target [6]
The expression of miRNAs in PCa has been studiedextensively (reviewed in [7]) We had previously described
Hindawi Publishing CorporationProstate CancerVolume 2017 Article ID 4893921 12 pageshttpdxdoiorg10115520174893921
2 Prostate Cancer
the miRNA profile of PCa at different stages of malignancyand could identify novel targets of miRNAs deregulated inPCa [8ndash12] Although there has been substantial progressin understanding the way how miRNAs are processed andloaded into Ago protein complexes (reviewed in [6 13]) it isstill not fully understood if or how pre-miRNAs are selectedfor loading into Ago complexes and if a deregulated miRNAexpression ultimately leads to an altered abundance of thismiRNA in the active miRISC complexes After constructionof the active miRISC the miRNA component guides theactive miRISC complex to its cognate mRNA target Thereare numerous additional factors that may influence the finalinteraction of miRISC with the target mRNA (reviewed in[14]) miRNA-regulated mRNAs can be isolated by immuno-precipitation of the Ago protein(s) [15ndash18]Therefore the aimof our study was to investigate whether the altered miRNAexpression in PCa leads to an altered spectrum of miRISC-associated mRNAs in cancer cells
2 Materials and Methods
21 Cell Lines The human PCa cell lines DU145 and LNCaPas well as HEK293T cells were purchased from the Ger-man collection of microorganisms and cell cultures (DSMZBraunschweig Germany) Primary human normal prostatefibroblasts (PNF-08) were kindly provided by Prof GerhardUnteregger (Dept of Urology University of SaarlandMedicalSchool) All cells were cultured as previously described [19]For precipitation of the Ago2 containing RISC complexextract of approx 2 times 108 PNF-08 and DU145 cells were used(see below)
22 RNA Extraction Extracts from cell lines were gener-ated using TRIzol (Life Technologies Darmstadt Germany)Extraction of total RNA and protein was carried out accord-ing to the TRIzol manual
23 Analysis of Ago2-Bound mRNAs The Ago2-containingRISC complexes were isolated from the DU145 and PNFcells as previously described [20] Shortly the DU145 andPNF-08 cells were lysed in a buffer consisting of 05NP40 150mMKCl 25mMTris pH 75 2mMEDTA and05mMDTT After centrifugation at 15000timesg at 4∘C for20min the supernatant was incubated for 4 h at 4∘C withprotein G Sepharose (GE Healthcare Solingen Germany)that was previously saturated with Ago2-specific antibody11A9 [16] The beads were washed repeatedly with a buffercontaining 300mMNaCl 50mMTris pH 75 5mMMgCl2and 005 NP40 The beads were then incubated for 15minat 65∘C with Proteinase K digestion buffer (300mMNaCl200mMTris-HCl pH 75 25mMEDTA 2 SDS 20mgmLProteinase K) The RNA was then extracted with TRIzolaccording to the TRIzol manual
24 Microarray Analysis Sample preparation for microarrayhybridization was carried out as described in the AmbionWT Expression Kit Protocol (Thermo Scientific) and theAffymetrix WT Terminal Labeling and Hybridization User
Manual (Affymetrix Inc Santa Clara CA USA) In brief200ndash300 ng of extracted RNAwere reverse transcribed usingan rRNA-depleted random primer mix as provided in theAmbion WT Expression Kit followed by an in vitro tran-scription reaction 8ndash12 120583g of in vitro transcribed antisenseRNA were purified and reverse transcribed into dUTP-containing sense-strand- (ss-) cDNA Purified ss-cDNA wasfragmented using a combination of uracil DNA glycosylase(UDG) and apurinicapyrimidinic endonuclease 1 (APE 1)followed by terminal labeling with biotin This processyielded between 1 and 3 120583g fragmented and labeled ss-cDNAwhich were hybridized to Affymetrix Human Gene 11 STPEG plate arrays Hybridization washing staining and arrayscanning were performed with a GeneTitan instrument(Affymetrix)
Summarized probe set signalswere calculated by applyingthe RMA [21] algorithm as implemented in the AffymetrixGeneChip Expression Console Software The result file wasexported in txt format and fold changesignificance calcu-lations were done in Microsoft Excel
25 Data Analysis First a literature research has been per-formed for mRNAs which were enriched or depleted atleast threefold in the Ago2 complexes of DU145 in com-parison to PNF-08 to identify genes involved in prostatetumorigenesis or in cancer development and progression ingeneralThe genes foundwere checked for their expression inprostatic normal and tumor tissue using an online databasefor mRNA expression from micro arrays oncomineorgThe next step was to analyze the 31015840 untranslated regions(31015840UTRs) of selected genes for putative miRNA target sites(Supplementary Table S2 in Supplementary Material avail-able online at httpsdoiorg10115520174893921) Predic-tion of miRNA target sites in the 31015840UTRs was performedby online algorithms targetscan (httpwwwtargetscanorg)and microRNAorg using binding of the ldquoseed sequencerdquobinding energy and conservation of binding sites as criteriaFor further analysis putative binding miRNAs were selectedwhich have been previously published as deregulated in PCa
26 Quantitative Real-Time PCR (qRT-PCR) Analysis ofmRNA Expression cDNA synthesis was performed withthe DyNAmo cDNA Synthesis Kit (Finnzymes Oy Van-taa Finland) using 200 ng of total RNA and random hex-amer primers Real-time PCRs were performed in triplicateusing TaqMan gene expression assays and TaqMan reagents(Applied Biosystems Foster City CA USA) according tothe manufacturerrsquos protocols with the StepOnePlus Real-Time PCR System (Life Technologies Darmstadt Ger-many) Sequence-specific primers and fluorescence-labeledprobes complementary to EI24 YWHAE TFRC CORO1CS100A16 NLK RAB1B DEDD CUL5 and PRDX3 and theendogenous reference GAPDH were used All PCRs weremeasured in triplicate in a final volume of 10120583L contain-ing 1x TaqMan Universal PCR Master Mix (No AmperaseUNG) and 50 ng cDNA Thermal cycling conditions werechosen according to the manufacturerrsquos recommendationsAs a reference sample cDNA prepared from DU145 cells
Prostate Cancer 3
was included in every reaction plate Calculation of relativeRNA expression levels by applying the ΔΔCt method [22]was performed using the StepOne software v20 (AppliedBiosystems Foster City CA USA)
27 Dual Luciferase Assays Dual luciferase assays employing31015840UTR reporters in pMIR-RNLTK (a dual firefly and renillaluciferase vector) were carried out in HEK293T cells asdescribed [11] A total of 105 HEK293T cells were seededin each cavity of a 24-well tissue culture dish and trans-fected using Nanofectin transfection reagent (GEHealthcareSolingen Germany) with 02 120583gwell reporter vector and atotal of 08 120583g pSG5-miRNA effector plasmid(s) Luciferaseassays were performed 48 hours after transfection using theDual Luciferase Reporter Assay System according to themanufacturerrsquos instructions (Promega Madison WI USA)Assays were conducted in duplicate and carried out at leastfour times for each effectorreporter pair
28 Plasmids The pSG5 plasmids for the expression ofmiRNAs miR-24 and -29b [11] miR-148a [10] miR-200c and-375 [12] have been described previously For the presentstudy miRNAs miR-15b 20a -21 and -29a were PCRamplified from human genomic DNA and inserted intothe vector pSG5 (Agilent Technologies) using the primersshown in Supplementary Table S3 The 31015840UTRs of the genesof the death effector domain-containing protein (DEDD)the tumor necrosis factor receptor superfamily member10b protein (TNFRSF10B) the tumor protein p53 induciblenuclear protein 1 (TP53INP1) the secreted protein acidiccysteine-rich (SPARC osteonectin) the Abelson interactor2 protein (ABI2) and the periredoxin 3 (PRDX3) proteinwere PCR amplified and inserted into the pMIR-RNLTKvector using the primers shown in Supplementary Table S3When necessary the potential binding sites for miRNAs weremutated by PCR the primers used for the mutations are alsoshown in Supplementary Table S3
29 Western Blotting For western blotting 2 times 105DU145cells grown in 10 cm dishes were transfected with 10120583gof plasmid DNA using jetPRIME (Polyplus transfectionSelestat France) After 48 h cells were lysed with 2-fold concentrated lysis buffer (130mM TrisHCl 6 SDS10 3-mercapto-12-propandiol 10 glycerol) and 30 120583g ofextracted proteins were separated on 9 tricine gels andtransferred to nitrocellulose membranes (Invitrogen Carls-bad CA USA) Membranes were blocked with TBST5BSA for 30min and incubated with primary antibodies overnight at 4∘C After washing three times with TBST themembranes were incubated with HRP-conjugated secondaryantibodies for 1 h at room temperature Protein bands weredetected with ECL reagent (GE Healthcare) using FujiFilmLAS-3000 imaging system (FujiFilm Tokyo Japan) Primaryrabbit anti-SPARC antibody was purchased from Cell Sig-naling Technology (D10F10 Danvers MA USA) rat anti-TP53INP1 antibodywas a generous gift fromDr Alice Carrier(Centre de Recherche en Cancerologie de Marseille France)and secondaryHRP-conjugated goat anti-rabbit IgG (31460)
and goat anti-rat IgG (31470) antibodies were purchasedfrom Thermo Scientific Pierce (Fisher Scientific SchwerteGermany)
210 Colony Formation Assay 2 times 106 DU145 cells wereseeded in 10 cm dishes and transfected with 10 120583g plas-mid DNA using jetPRIME (Polyplus transfection SelestatFrance) 24 hours after transfection the cells were detachedby trypsin resuspended in 10mL medium seeded in 6-well plates (2000 cellswell) and cultured for 8 days Aftermedium replacement cultures were stained with 04 cristalviolet fixed with 4 paraformaldehyde for 30 minutesand washed 3 times with PBS Wells were photographedand densitometrically analysed with ImageJ 148v (NationalInstitute of Health USA)
211 Cell Proliferation Assay 15 times 105 DU145 cells wereseeded in 6-well plates transfected with 2 120583g plasmid DNAusing jetPRIME (Polyplus transfection Selestat France) andcultivated for 24ndash72 h For measuring cell numbers on days0 to 3 after transfection cells were detached with trypsin andresuspended in 1mLmediumCell numberswere determinedwith CASY 1 cell counter (Scharfe System Reutlingen Ger-many) in cellsmL
212 Statistics Statistical evaluation of the luciferase assayswas performed with SigmaPlot 10 (Systat) using Studentrsquos 119905-test analysis All statistical tests were performed as two-sidedand 119901 values of lt005 were considered as significant
3 Results
To identify mRNAs differentially enriched or excluded fromthe RISC complex Ago-2 was immunoprecipitated fromextracts derived from normal prostate fibroblasts (PNF-08)and from the PCa cell line DU145 using the previouslydescribed Ago-2 antibody 11A9 and an appropriate isotypecontrol [15 16] The mRNAs present in the Ago-precipitatewere quantitated by microarray analysis The relative abun-dance of RISC-associated mRNAs in the two cell lines arepresented in Supplementary Table S1 Twenty-four mRNAsthat exhibited a pronounced enrichment or depletion in theAgo complexes of DU145 cell are presented in Table 1 Supple-mentary Table S4 lists the functions (if known) of the genes inTable 1 and if they are potential or known targets of miRNAsderegulated in PCa For instance the highest abundance inthe Ago complexes of DU145 cells was observed for lamininalpha 3 (LAMA3) Expression of this gene is among othermembers in its family significantly reduced in prostate carci-noma [23] in line with being a potential target for miRNAsmiR-20a and miR-106a (httpwwwtargetscanorg) bothof which are up-regulated in prostate carcinoma [8 19] Inaddition to the LAMA3 mRNA the highly RISC-enrichedadenylate cyclase 3 (ADCY3) mRNA is a potential target forthe upregulatedmiRNAsmiR-25 andmiR-27ab Converselythe Thy-1 cell surface antigen (THY1) has been shown to beupregulated in PCa [24] Accordingly the THY1 mRNA levelwas reduced in the Ago complex of the DU145 cells
4 Prostate Cancer
Table 1 Abundance of selected mRNAs in AGO complexes of DU145 PCa cells in comparison to prostate normal fibroblasts (PNF08)
Gene title Gene symbol EnrichmentThy-1 cell surface antigen THY1 lt001Matrix Gla protein MGP lt001Periostin osteoblast specific factor POSTN lt001Secreted protein acidic cysteine-rich (osteonectin) SPARC lt001Chemokine (C-C motif) ligand 2 CCL2 lt001Cathepsin K CTSK 0059Tyrosine 3-monooxygenasetryptophan 5-monooxygenase activation protein epsilon polypeptide YWHAE 0093Peroxiredoxin 3 PRDX3 0158Coronin actin binding protein 1C CORO1C 0187Tumor protein p53 inducible nuclear protein 1 TP53INP1 0214Transferrin receptor (p90 CD71) TFRC 0278Laminin alpha 3 LAMA3 6891Etoposide induced 24 mRNA EI24 6813Adenylate cyclase 3 ADCY3 6001Succinate dehydrogenase complex subunit A SDHA 5973Adiponectin receptor 2 ADIPOR2 5187Nemo-like kinase NLK 4582ATM interactor ATMIN 4430RAB1B member RAS oncogene family RAB1B 4293Death effector domain containing DEDD 4019Tumor necrosis factor receptor superfamily member 10b TNFRSF10B 3635abl-interactor 2 ABI2 3267cullin 5 CUL5 3144S100 calcium binding protein A16 S100A16 3069
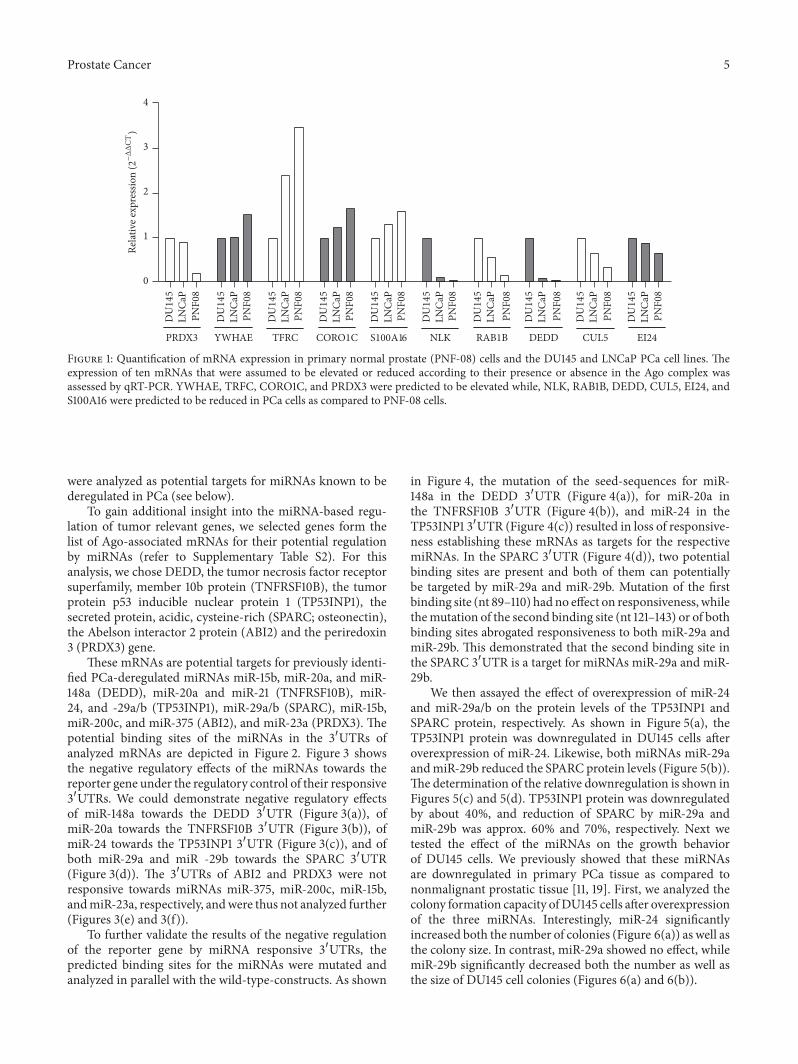
We then selected 10 mRNAs that based on our resultssuggest a possible deregulation in PCa We analyzed byqRT-PCR the relative expression of these genes in PNF-08 cells in comparison with DU145 and LNCaP cells SixmRNAs were selected that exhibited an enrichment in theAgo complexes from PCa cells and were therefore potentiallydownregulated in cell lines and tumors Etoposide induced24 mRNA (EI24 alias p53-induced gene 8 protein) S100calcium binding protein A16 (S100A16) Nemo-like kinase(NLK) RAS oncogene family member 1B (RAB1B) deatheffector domain-containing protein (DEDD) and cullin 5(CUL5) On the other end we selected mRNAs that showeddepletion from Ago complexes of PCa cells and therefore arepotentially overexpressed in PCa cells and tumorsTheseweretyrosine 3-Monooxygenase (YWHAE aka 14-3-3 proteinepsilon) transferrin receptor (TRFC) coronin actin bindingprotein 1C (CORO1C) and periredoxin 3 (PRDX3) Theresults of the analysis are shown in Figure 1
We initially expected that mRNAs enriched in Agocomplexes undergo degradation by deadenylation or by 51015840-decapping which would result in a reduced overall mRNAcontent in whole-cell RNA preparations Interestingly the re-sults of quantitative PCR analyses demonstrated that mRNAsenriched in the Ago complexes of PCa cells were presentat higher amounts in whole-cell RNA and mRNAs depletedfromAgo complexes of PCa cells exhibited a reduced amount
Only twoout of the selected 10mRNAs (S100A16 andPRDX3)followed our initial expectation These results demonstratethat an enrichment of mRNAs in Ago complexes does notnecessarily lead to an enhanced degradation of the respectivemRNA
EI24 as an Etoposide-inducible gene was chosen becauseEtoposide has recently been suggested as one componentof a combination chemotherapy treatment of advanced PCa[25] YWHAE is among the 14-3-3 proteins that regulate theETV1 transcription factor in PCa [26] TRFC was shownoriginally to be significantly induced in DU145 cells [27]CORO1Ccoronin was found to be induced in androgen-insensitive PCa [28] and S100A16 appears to be upregulatedonly in metastatic tumor cells [29]
NLK acts as a 120573-catenin pathway inhibitor throughphosphorylation and degradation of TCFLEF transcriptionfactors [30] NLK and DEDD are suggested to induce apop-tosis [31 32] and loss of RAB1B was shown to enhance theaggressiveness of breast carcinoma [33] A reduction of CUL5expression was found for instance in breast cancer [34] butso far not in prostate carcinoma Finally PRDX3 was chosenas it was previously found to be upregulated in primaryPCa [35] For this gene our initial assumption holds truewhere a reduced abundance of the PRDX3 mRNA in Agocomplexes of PCa cells results in an elevated mRNA contentin whole cells In addition the 31015840UTRs of DEDD and PRDX3
Prostate Cancer 5
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
EI24CUL5DEDDRAB1BNLKS100A16CORO1CTFRCYWHAEPRDX3
0
1
2
3
4
Rela
tive e
xpre
ssio
n (2minusΔΔCT
)
Figure 1 Quantification of mRNA expression in primary normal prostate (PNF-08) cells and the DU145 and LNCaP PCa cell lines Theexpression of ten mRNAs that were assumed to be elevated or reduced according to their presence or absence in the Ago complex wasassessed by qRT-PCR YWHAE TRFC CORO1C and PRDX3 were predicted to be elevated while NLK RAB1B DEDD CUL5 EI24 andS100A16 were predicted to be reduced in PCa cells as compared to PNF-08 cells
were analyzed as potential targets for miRNAs known to bederegulated in PCa (see below)
To gain additional insight into the miRNA-based regu-lation of tumor relevant genes we selected genes form thelist of Ago-associated mRNAs for their potential regulationby miRNAs (refer to Supplementary Table S2) For thisanalysis we chose DEDD the tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10B) the tumorprotein p53 inducible nuclear protein 1 (TP53INP1) thesecreted protein acidic cysteine-rich (SPARC osteonectin)the Abelson interactor 2 protein (ABI2) and the periredoxin3 (PRDX3) gene
These mRNAs are potential targets for previously identi-fied PCa-deregulated miRNAs miR-15b miR-20a and miR-148a (DEDD) miR-20a and miR-21 (TNFRSF10B) miR-24 and -29ab (TP53INP1) miR-29ab (SPARC) miR-15bmiR-200c and miR-375 (ABI2) and miR-23a (PRDX3) Thepotential binding sites of the miRNAs in the 31015840UTRs ofanalyzed mRNAs are depicted in Figure 2 Figure 3 showsthe negative regulatory effects of the miRNAs towards thereporter gene under the regulatory control of their responsive31015840UTRs We could demonstrate negative regulatory effectsof miR-148a towards the DEDD 31015840UTR (Figure 3(a)) ofmiR-20a towards the TNFRSF10B 31015840UTR (Figure 3(b)) ofmiR-24 towards the TP53INP1 31015840UTR (Figure 3(c)) and ofboth miR-29a and miR -29b towards the SPARC 31015840UTR(Figure 3(d)) The 31015840UTRs of ABI2 and PRDX3 were notresponsive towards miRNAs miR-375 miR-200c miR-15bandmiR-23a respectively andwere thus not analyzed further(Figures 3(e) and 3(f))
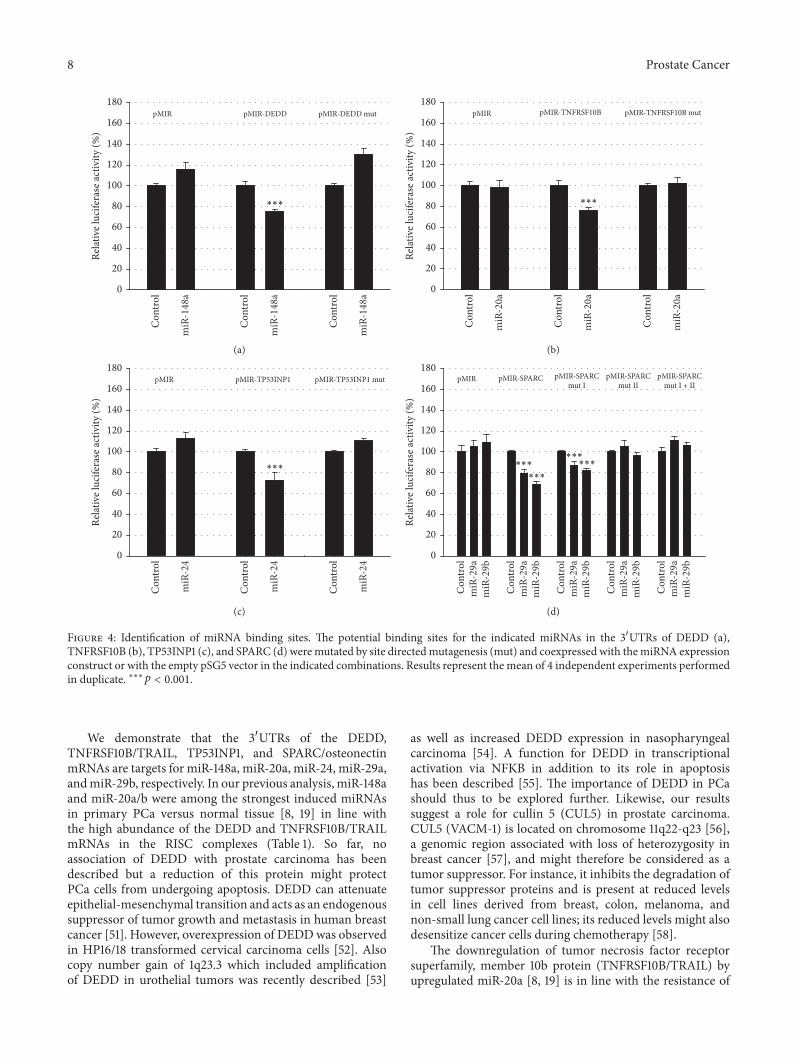
To further validate the results of the negative regulationof the reporter gene by miRNA responsive 31015840UTRs thepredicted binding sites for the miRNAs were mutated andanalyzed in parallel with the wild-type-constructs As shown
in Figure 4 the mutation of the seed-sequences for miR-148a in the DEDD 31015840UTR (Figure 4(a)) for miR-20a inthe TNFRSF10B 31015840UTR (Figure 4(b)) and miR-24 in theTP53INP1 31015840UTR (Figure 4(c)) resulted in loss of responsive-ness establishing these mRNAs as targets for the respectivemiRNAs In the SPARC 31015840UTR (Figure 4(d)) two potentialbinding sites are present and both of them can potentiallybe targeted by miR-29a and miR-29b Mutation of the firstbinding site (nt 89ndash110) had no effect on responsiveness whilethemutation of the second binding site (nt 121ndash143) or of bothbinding sites abrogated responsiveness to both miR-29a andmiR-29b This demonstrated that the second binding site inthe SPARC 31015840UTR is a target for miRNAs miR-29a and miR-29b
We then assayed the effect of overexpression of miR-24and miR-29ab on the protein levels of the TP53INP1 andSPARC protein respectively As shown in Figure 5(a) theTP53INP1 protein was downregulated in DU145 cells afteroverexpression of miR-24 Likewise both miRNAs miR-29aandmiR-29b reduced the SPARC protein levels (Figure 5(b))The determination of the relative downregulation is shown inFigures 5(c) and 5(d) TP53INP1 protein was downregulatedby about 40 and reduction of SPARC by miR-29a andmiR-29b was approx 60 and 70 respectively Next wetested the effect of the miRNAs on the growth behaviorof DU145 cells We previously showed that these miRNAsare downregulated in primary PCa tissue as compared tononmalignant prostatic tissue [11 19] First we analyzed thecolony formation capacity ofDU145 cells after overexpressionof the three miRNAs Interestingly miR-24 significantlyincreased both the number of colonies (Figure 6(a)) as well asthe colony size In contrast miR-29a showed no effect whilemiR-29b significantly decreased both the number as well asthe size of DU145 cell colonies (Figures 6(a) and 6(b))
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
2 Prostate Cancer
the miRNA profile of PCa at different stages of malignancyand could identify novel targets of miRNAs deregulated inPCa [8ndash12] Although there has been substantial progressin understanding the way how miRNAs are processed andloaded into Ago protein complexes (reviewed in [6 13]) it isstill not fully understood if or how pre-miRNAs are selectedfor loading into Ago complexes and if a deregulated miRNAexpression ultimately leads to an altered abundance of thismiRNA in the active miRISC complexes After constructionof the active miRISC the miRNA component guides theactive miRISC complex to its cognate mRNA target Thereare numerous additional factors that may influence the finalinteraction of miRISC with the target mRNA (reviewed in[14]) miRNA-regulated mRNAs can be isolated by immuno-precipitation of the Ago protein(s) [15ndash18]Therefore the aimof our study was to investigate whether the altered miRNAexpression in PCa leads to an altered spectrum of miRISC-associated mRNAs in cancer cells
2 Materials and Methods
21 Cell Lines The human PCa cell lines DU145 and LNCaPas well as HEK293T cells were purchased from the Ger-man collection of microorganisms and cell cultures (DSMZBraunschweig Germany) Primary human normal prostatefibroblasts (PNF-08) were kindly provided by Prof GerhardUnteregger (Dept of Urology University of SaarlandMedicalSchool) All cells were cultured as previously described [19]For precipitation of the Ago2 containing RISC complexextract of approx 2 times 108 PNF-08 and DU145 cells were used(see below)
22 RNA Extraction Extracts from cell lines were gener-ated using TRIzol (Life Technologies Darmstadt Germany)Extraction of total RNA and protein was carried out accord-ing to the TRIzol manual
23 Analysis of Ago2-Bound mRNAs The Ago2-containingRISC complexes were isolated from the DU145 and PNFcells as previously described [20] Shortly the DU145 andPNF-08 cells were lysed in a buffer consisting of 05NP40 150mMKCl 25mMTris pH 75 2mMEDTA and05mMDTT After centrifugation at 15000timesg at 4∘C for20min the supernatant was incubated for 4 h at 4∘C withprotein G Sepharose (GE Healthcare Solingen Germany)that was previously saturated with Ago2-specific antibody11A9 [16] The beads were washed repeatedly with a buffercontaining 300mMNaCl 50mMTris pH 75 5mMMgCl2and 005 NP40 The beads were then incubated for 15minat 65∘C with Proteinase K digestion buffer (300mMNaCl200mMTris-HCl pH 75 25mMEDTA 2 SDS 20mgmLProteinase K) The RNA was then extracted with TRIzolaccording to the TRIzol manual
24 Microarray Analysis Sample preparation for microarrayhybridization was carried out as described in the AmbionWT Expression Kit Protocol (Thermo Scientific) and theAffymetrix WT Terminal Labeling and Hybridization User
Manual (Affymetrix Inc Santa Clara CA USA) In brief200ndash300 ng of extracted RNAwere reverse transcribed usingan rRNA-depleted random primer mix as provided in theAmbion WT Expression Kit followed by an in vitro tran-scription reaction 8ndash12 120583g of in vitro transcribed antisenseRNA were purified and reverse transcribed into dUTP-containing sense-strand- (ss-) cDNA Purified ss-cDNA wasfragmented using a combination of uracil DNA glycosylase(UDG) and apurinicapyrimidinic endonuclease 1 (APE 1)followed by terminal labeling with biotin This processyielded between 1 and 3 120583g fragmented and labeled ss-cDNAwhich were hybridized to Affymetrix Human Gene 11 STPEG plate arrays Hybridization washing staining and arrayscanning were performed with a GeneTitan instrument(Affymetrix)
Summarized probe set signalswere calculated by applyingthe RMA [21] algorithm as implemented in the AffymetrixGeneChip Expression Console Software The result file wasexported in txt format and fold changesignificance calcu-lations were done in Microsoft Excel
25 Data Analysis First a literature research has been per-formed for mRNAs which were enriched or depleted atleast threefold in the Ago2 complexes of DU145 in com-parison to PNF-08 to identify genes involved in prostatetumorigenesis or in cancer development and progression ingeneralThe genes foundwere checked for their expression inprostatic normal and tumor tissue using an online databasefor mRNA expression from micro arrays oncomineorgThe next step was to analyze the 31015840 untranslated regions(31015840UTRs) of selected genes for putative miRNA target sites(Supplementary Table S2 in Supplementary Material avail-able online at httpsdoiorg10115520174893921) Predic-tion of miRNA target sites in the 31015840UTRs was performedby online algorithms targetscan (httpwwwtargetscanorg)and microRNAorg using binding of the ldquoseed sequencerdquobinding energy and conservation of binding sites as criteriaFor further analysis putative binding miRNAs were selectedwhich have been previously published as deregulated in PCa
26 Quantitative Real-Time PCR (qRT-PCR) Analysis ofmRNA Expression cDNA synthesis was performed withthe DyNAmo cDNA Synthesis Kit (Finnzymes Oy Van-taa Finland) using 200 ng of total RNA and random hex-amer primers Real-time PCRs were performed in triplicateusing TaqMan gene expression assays and TaqMan reagents(Applied Biosystems Foster City CA USA) according tothe manufacturerrsquos protocols with the StepOnePlus Real-Time PCR System (Life Technologies Darmstadt Ger-many) Sequence-specific primers and fluorescence-labeledprobes complementary to EI24 YWHAE TFRC CORO1CS100A16 NLK RAB1B DEDD CUL5 and PRDX3 and theendogenous reference GAPDH were used All PCRs weremeasured in triplicate in a final volume of 10120583L contain-ing 1x TaqMan Universal PCR Master Mix (No AmperaseUNG) and 50 ng cDNA Thermal cycling conditions werechosen according to the manufacturerrsquos recommendationsAs a reference sample cDNA prepared from DU145 cells
Prostate Cancer 3
was included in every reaction plate Calculation of relativeRNA expression levels by applying the ΔΔCt method [22]was performed using the StepOne software v20 (AppliedBiosystems Foster City CA USA)
27 Dual Luciferase Assays Dual luciferase assays employing31015840UTR reporters in pMIR-RNLTK (a dual firefly and renillaluciferase vector) were carried out in HEK293T cells asdescribed [11] A total of 105 HEK293T cells were seededin each cavity of a 24-well tissue culture dish and trans-fected using Nanofectin transfection reagent (GEHealthcareSolingen Germany) with 02 120583gwell reporter vector and atotal of 08 120583g pSG5-miRNA effector plasmid(s) Luciferaseassays were performed 48 hours after transfection using theDual Luciferase Reporter Assay System according to themanufacturerrsquos instructions (Promega Madison WI USA)Assays were conducted in duplicate and carried out at leastfour times for each effectorreporter pair
28 Plasmids The pSG5 plasmids for the expression ofmiRNAs miR-24 and -29b [11] miR-148a [10] miR-200c and-375 [12] have been described previously For the presentstudy miRNAs miR-15b 20a -21 and -29a were PCRamplified from human genomic DNA and inserted intothe vector pSG5 (Agilent Technologies) using the primersshown in Supplementary Table S3 The 31015840UTRs of the genesof the death effector domain-containing protein (DEDD)the tumor necrosis factor receptor superfamily member10b protein (TNFRSF10B) the tumor protein p53 induciblenuclear protein 1 (TP53INP1) the secreted protein acidiccysteine-rich (SPARC osteonectin) the Abelson interactor2 protein (ABI2) and the periredoxin 3 (PRDX3) proteinwere PCR amplified and inserted into the pMIR-RNLTKvector using the primers shown in Supplementary Table S3When necessary the potential binding sites for miRNAs weremutated by PCR the primers used for the mutations are alsoshown in Supplementary Table S3
29 Western Blotting For western blotting 2 times 105DU145cells grown in 10 cm dishes were transfected with 10120583gof plasmid DNA using jetPRIME (Polyplus transfectionSelestat France) After 48 h cells were lysed with 2-fold concentrated lysis buffer (130mM TrisHCl 6 SDS10 3-mercapto-12-propandiol 10 glycerol) and 30 120583g ofextracted proteins were separated on 9 tricine gels andtransferred to nitrocellulose membranes (Invitrogen Carls-bad CA USA) Membranes were blocked with TBST5BSA for 30min and incubated with primary antibodies overnight at 4∘C After washing three times with TBST themembranes were incubated with HRP-conjugated secondaryantibodies for 1 h at room temperature Protein bands weredetected with ECL reagent (GE Healthcare) using FujiFilmLAS-3000 imaging system (FujiFilm Tokyo Japan) Primaryrabbit anti-SPARC antibody was purchased from Cell Sig-naling Technology (D10F10 Danvers MA USA) rat anti-TP53INP1 antibodywas a generous gift fromDr Alice Carrier(Centre de Recherche en Cancerologie de Marseille France)and secondaryHRP-conjugated goat anti-rabbit IgG (31460)
and goat anti-rat IgG (31470) antibodies were purchasedfrom Thermo Scientific Pierce (Fisher Scientific SchwerteGermany)
210 Colony Formation Assay 2 times 106 DU145 cells wereseeded in 10 cm dishes and transfected with 10 120583g plas-mid DNA using jetPRIME (Polyplus transfection SelestatFrance) 24 hours after transfection the cells were detachedby trypsin resuspended in 10mL medium seeded in 6-well plates (2000 cellswell) and cultured for 8 days Aftermedium replacement cultures were stained with 04 cristalviolet fixed with 4 paraformaldehyde for 30 minutesand washed 3 times with PBS Wells were photographedand densitometrically analysed with ImageJ 148v (NationalInstitute of Health USA)
211 Cell Proliferation Assay 15 times 105 DU145 cells wereseeded in 6-well plates transfected with 2 120583g plasmid DNAusing jetPRIME (Polyplus transfection Selestat France) andcultivated for 24ndash72 h For measuring cell numbers on days0 to 3 after transfection cells were detached with trypsin andresuspended in 1mLmediumCell numberswere determinedwith CASY 1 cell counter (Scharfe System Reutlingen Ger-many) in cellsmL
212 Statistics Statistical evaluation of the luciferase assayswas performed with SigmaPlot 10 (Systat) using Studentrsquos 119905-test analysis All statistical tests were performed as two-sidedand 119901 values of lt005 were considered as significant
3 Results
To identify mRNAs differentially enriched or excluded fromthe RISC complex Ago-2 was immunoprecipitated fromextracts derived from normal prostate fibroblasts (PNF-08)and from the PCa cell line DU145 using the previouslydescribed Ago-2 antibody 11A9 and an appropriate isotypecontrol [15 16] The mRNAs present in the Ago-precipitatewere quantitated by microarray analysis The relative abun-dance of RISC-associated mRNAs in the two cell lines arepresented in Supplementary Table S1 Twenty-four mRNAsthat exhibited a pronounced enrichment or depletion in theAgo complexes of DU145 cell are presented in Table 1 Supple-mentary Table S4 lists the functions (if known) of the genes inTable 1 and if they are potential or known targets of miRNAsderegulated in PCa For instance the highest abundance inthe Ago complexes of DU145 cells was observed for lamininalpha 3 (LAMA3) Expression of this gene is among othermembers in its family significantly reduced in prostate carci-noma [23] in line with being a potential target for miRNAsmiR-20a and miR-106a (httpwwwtargetscanorg) bothof which are up-regulated in prostate carcinoma [8 19] Inaddition to the LAMA3 mRNA the highly RISC-enrichedadenylate cyclase 3 (ADCY3) mRNA is a potential target forthe upregulatedmiRNAsmiR-25 andmiR-27ab Converselythe Thy-1 cell surface antigen (THY1) has been shown to beupregulated in PCa [24] Accordingly the THY1 mRNA levelwas reduced in the Ago complex of the DU145 cells
4 Prostate Cancer
Table 1 Abundance of selected mRNAs in AGO complexes of DU145 PCa cells in comparison to prostate normal fibroblasts (PNF08)
Gene title Gene symbol EnrichmentThy-1 cell surface antigen THY1 lt001Matrix Gla protein MGP lt001Periostin osteoblast specific factor POSTN lt001Secreted protein acidic cysteine-rich (osteonectin) SPARC lt001Chemokine (C-C motif) ligand 2 CCL2 lt001Cathepsin K CTSK 0059Tyrosine 3-monooxygenasetryptophan 5-monooxygenase activation protein epsilon polypeptide YWHAE 0093Peroxiredoxin 3 PRDX3 0158Coronin actin binding protein 1C CORO1C 0187Tumor protein p53 inducible nuclear protein 1 TP53INP1 0214Transferrin receptor (p90 CD71) TFRC 0278Laminin alpha 3 LAMA3 6891Etoposide induced 24 mRNA EI24 6813Adenylate cyclase 3 ADCY3 6001Succinate dehydrogenase complex subunit A SDHA 5973Adiponectin receptor 2 ADIPOR2 5187Nemo-like kinase NLK 4582ATM interactor ATMIN 4430RAB1B member RAS oncogene family RAB1B 4293Death effector domain containing DEDD 4019Tumor necrosis factor receptor superfamily member 10b TNFRSF10B 3635abl-interactor 2 ABI2 3267cullin 5 CUL5 3144S100 calcium binding protein A16 S100A16 3069
We then selected 10 mRNAs that based on our resultssuggest a possible deregulation in PCa We analyzed byqRT-PCR the relative expression of these genes in PNF-08 cells in comparison with DU145 and LNCaP cells SixmRNAs were selected that exhibited an enrichment in theAgo complexes from PCa cells and were therefore potentiallydownregulated in cell lines and tumors Etoposide induced24 mRNA (EI24 alias p53-induced gene 8 protein) S100calcium binding protein A16 (S100A16) Nemo-like kinase(NLK) RAS oncogene family member 1B (RAB1B) deatheffector domain-containing protein (DEDD) and cullin 5(CUL5) On the other end we selected mRNAs that showeddepletion from Ago complexes of PCa cells and therefore arepotentially overexpressed in PCa cells and tumorsTheseweretyrosine 3-Monooxygenase (YWHAE aka 14-3-3 proteinepsilon) transferrin receptor (TRFC) coronin actin bindingprotein 1C (CORO1C) and periredoxin 3 (PRDX3) Theresults of the analysis are shown in Figure 1
We initially expected that mRNAs enriched in Agocomplexes undergo degradation by deadenylation or by 51015840-decapping which would result in a reduced overall mRNAcontent in whole-cell RNA preparations Interestingly the re-sults of quantitative PCR analyses demonstrated that mRNAsenriched in the Ago complexes of PCa cells were presentat higher amounts in whole-cell RNA and mRNAs depletedfromAgo complexes of PCa cells exhibited a reduced amount
Only twoout of the selected 10mRNAs (S100A16 andPRDX3)followed our initial expectation These results demonstratethat an enrichment of mRNAs in Ago complexes does notnecessarily lead to an enhanced degradation of the respectivemRNA
EI24 as an Etoposide-inducible gene was chosen becauseEtoposide has recently been suggested as one componentof a combination chemotherapy treatment of advanced PCa[25] YWHAE is among the 14-3-3 proteins that regulate theETV1 transcription factor in PCa [26] TRFC was shownoriginally to be significantly induced in DU145 cells [27]CORO1Ccoronin was found to be induced in androgen-insensitive PCa [28] and S100A16 appears to be upregulatedonly in metastatic tumor cells [29]
NLK acts as a 120573-catenin pathway inhibitor throughphosphorylation and degradation of TCFLEF transcriptionfactors [30] NLK and DEDD are suggested to induce apop-tosis [31 32] and loss of RAB1B was shown to enhance theaggressiveness of breast carcinoma [33] A reduction of CUL5expression was found for instance in breast cancer [34] butso far not in prostate carcinoma Finally PRDX3 was chosenas it was previously found to be upregulated in primaryPCa [35] For this gene our initial assumption holds truewhere a reduced abundance of the PRDX3 mRNA in Agocomplexes of PCa cells results in an elevated mRNA contentin whole cells In addition the 31015840UTRs of DEDD and PRDX3
Prostate Cancer 5
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
EI24CUL5DEDDRAB1BNLKS100A16CORO1CTFRCYWHAEPRDX3
0
1
2
3
4
Rela
tive e
xpre
ssio
n (2minusΔΔCT
)
Figure 1 Quantification of mRNA expression in primary normal prostate (PNF-08) cells and the DU145 and LNCaP PCa cell lines Theexpression of ten mRNAs that were assumed to be elevated or reduced according to their presence or absence in the Ago complex wasassessed by qRT-PCR YWHAE TRFC CORO1C and PRDX3 were predicted to be elevated while NLK RAB1B DEDD CUL5 EI24 andS100A16 were predicted to be reduced in PCa cells as compared to PNF-08 cells
were analyzed as potential targets for miRNAs known to bederegulated in PCa (see below)
To gain additional insight into the miRNA-based regu-lation of tumor relevant genes we selected genes form thelist of Ago-associated mRNAs for their potential regulationby miRNAs (refer to Supplementary Table S2) For thisanalysis we chose DEDD the tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10B) the tumorprotein p53 inducible nuclear protein 1 (TP53INP1) thesecreted protein acidic cysteine-rich (SPARC osteonectin)the Abelson interactor 2 protein (ABI2) and the periredoxin3 (PRDX3) gene
These mRNAs are potential targets for previously identi-fied PCa-deregulated miRNAs miR-15b miR-20a and miR-148a (DEDD) miR-20a and miR-21 (TNFRSF10B) miR-24 and -29ab (TP53INP1) miR-29ab (SPARC) miR-15bmiR-200c and miR-375 (ABI2) and miR-23a (PRDX3) Thepotential binding sites of the miRNAs in the 31015840UTRs ofanalyzed mRNAs are depicted in Figure 2 Figure 3 showsthe negative regulatory effects of the miRNAs towards thereporter gene under the regulatory control of their responsive31015840UTRs We could demonstrate negative regulatory effectsof miR-148a towards the DEDD 31015840UTR (Figure 3(a)) ofmiR-20a towards the TNFRSF10B 31015840UTR (Figure 3(b)) ofmiR-24 towards the TP53INP1 31015840UTR (Figure 3(c)) and ofboth miR-29a and miR -29b towards the SPARC 31015840UTR(Figure 3(d)) The 31015840UTRs of ABI2 and PRDX3 were notresponsive towards miRNAs miR-375 miR-200c miR-15bandmiR-23a respectively andwere thus not analyzed further(Figures 3(e) and 3(f))
To further validate the results of the negative regulationof the reporter gene by miRNA responsive 31015840UTRs thepredicted binding sites for the miRNAs were mutated andanalyzed in parallel with the wild-type-constructs As shown
in Figure 4 the mutation of the seed-sequences for miR-148a in the DEDD 31015840UTR (Figure 4(a)) for miR-20a inthe TNFRSF10B 31015840UTR (Figure 4(b)) and miR-24 in theTP53INP1 31015840UTR (Figure 4(c)) resulted in loss of responsive-ness establishing these mRNAs as targets for the respectivemiRNAs In the SPARC 31015840UTR (Figure 4(d)) two potentialbinding sites are present and both of them can potentiallybe targeted by miR-29a and miR-29b Mutation of the firstbinding site (nt 89ndash110) had no effect on responsiveness whilethemutation of the second binding site (nt 121ndash143) or of bothbinding sites abrogated responsiveness to both miR-29a andmiR-29b This demonstrated that the second binding site inthe SPARC 31015840UTR is a target for miRNAs miR-29a and miR-29b
We then assayed the effect of overexpression of miR-24and miR-29ab on the protein levels of the TP53INP1 andSPARC protein respectively As shown in Figure 5(a) theTP53INP1 protein was downregulated in DU145 cells afteroverexpression of miR-24 Likewise both miRNAs miR-29aandmiR-29b reduced the SPARC protein levels (Figure 5(b))The determination of the relative downregulation is shown inFigures 5(c) and 5(d) TP53INP1 protein was downregulatedby about 40 and reduction of SPARC by miR-29a andmiR-29b was approx 60 and 70 respectively Next wetested the effect of the miRNAs on the growth behaviorof DU145 cells We previously showed that these miRNAsare downregulated in primary PCa tissue as compared tononmalignant prostatic tissue [11 19] First we analyzed thecolony formation capacity ofDU145 cells after overexpressionof the three miRNAs Interestingly miR-24 significantlyincreased both the number of colonies (Figure 6(a)) as well asthe colony size In contrast miR-29a showed no effect whilemiR-29b significantly decreased both the number as well asthe size of DU145 cell colonies (Figures 6(a) and 6(b))
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Prostate Cancer 3
was included in every reaction plate Calculation of relativeRNA expression levels by applying the ΔΔCt method [22]was performed using the StepOne software v20 (AppliedBiosystems Foster City CA USA)
27 Dual Luciferase Assays Dual luciferase assays employing31015840UTR reporters in pMIR-RNLTK (a dual firefly and renillaluciferase vector) were carried out in HEK293T cells asdescribed [11] A total of 105 HEK293T cells were seededin each cavity of a 24-well tissue culture dish and trans-fected using Nanofectin transfection reagent (GEHealthcareSolingen Germany) with 02 120583gwell reporter vector and atotal of 08 120583g pSG5-miRNA effector plasmid(s) Luciferaseassays were performed 48 hours after transfection using theDual Luciferase Reporter Assay System according to themanufacturerrsquos instructions (Promega Madison WI USA)Assays were conducted in duplicate and carried out at leastfour times for each effectorreporter pair
28 Plasmids The pSG5 plasmids for the expression ofmiRNAs miR-24 and -29b [11] miR-148a [10] miR-200c and-375 [12] have been described previously For the presentstudy miRNAs miR-15b 20a -21 and -29a were PCRamplified from human genomic DNA and inserted intothe vector pSG5 (Agilent Technologies) using the primersshown in Supplementary Table S3 The 31015840UTRs of the genesof the death effector domain-containing protein (DEDD)the tumor necrosis factor receptor superfamily member10b protein (TNFRSF10B) the tumor protein p53 induciblenuclear protein 1 (TP53INP1) the secreted protein acidiccysteine-rich (SPARC osteonectin) the Abelson interactor2 protein (ABI2) and the periredoxin 3 (PRDX3) proteinwere PCR amplified and inserted into the pMIR-RNLTKvector using the primers shown in Supplementary Table S3When necessary the potential binding sites for miRNAs weremutated by PCR the primers used for the mutations are alsoshown in Supplementary Table S3
29 Western Blotting For western blotting 2 times 105DU145cells grown in 10 cm dishes were transfected with 10120583gof plasmid DNA using jetPRIME (Polyplus transfectionSelestat France) After 48 h cells were lysed with 2-fold concentrated lysis buffer (130mM TrisHCl 6 SDS10 3-mercapto-12-propandiol 10 glycerol) and 30 120583g ofextracted proteins were separated on 9 tricine gels andtransferred to nitrocellulose membranes (Invitrogen Carls-bad CA USA) Membranes were blocked with TBST5BSA for 30min and incubated with primary antibodies overnight at 4∘C After washing three times with TBST themembranes were incubated with HRP-conjugated secondaryantibodies for 1 h at room temperature Protein bands weredetected with ECL reagent (GE Healthcare) using FujiFilmLAS-3000 imaging system (FujiFilm Tokyo Japan) Primaryrabbit anti-SPARC antibody was purchased from Cell Sig-naling Technology (D10F10 Danvers MA USA) rat anti-TP53INP1 antibodywas a generous gift fromDr Alice Carrier(Centre de Recherche en Cancerologie de Marseille France)and secondaryHRP-conjugated goat anti-rabbit IgG (31460)
and goat anti-rat IgG (31470) antibodies were purchasedfrom Thermo Scientific Pierce (Fisher Scientific SchwerteGermany)
210 Colony Formation Assay 2 times 106 DU145 cells wereseeded in 10 cm dishes and transfected with 10 120583g plas-mid DNA using jetPRIME (Polyplus transfection SelestatFrance) 24 hours after transfection the cells were detachedby trypsin resuspended in 10mL medium seeded in 6-well plates (2000 cellswell) and cultured for 8 days Aftermedium replacement cultures were stained with 04 cristalviolet fixed with 4 paraformaldehyde for 30 minutesand washed 3 times with PBS Wells were photographedand densitometrically analysed with ImageJ 148v (NationalInstitute of Health USA)
211 Cell Proliferation Assay 15 times 105 DU145 cells wereseeded in 6-well plates transfected with 2 120583g plasmid DNAusing jetPRIME (Polyplus transfection Selestat France) andcultivated for 24ndash72 h For measuring cell numbers on days0 to 3 after transfection cells were detached with trypsin andresuspended in 1mLmediumCell numberswere determinedwith CASY 1 cell counter (Scharfe System Reutlingen Ger-many) in cellsmL
212 Statistics Statistical evaluation of the luciferase assayswas performed with SigmaPlot 10 (Systat) using Studentrsquos 119905-test analysis All statistical tests were performed as two-sidedand 119901 values of lt005 were considered as significant
3 Results
To identify mRNAs differentially enriched or excluded fromthe RISC complex Ago-2 was immunoprecipitated fromextracts derived from normal prostate fibroblasts (PNF-08)and from the PCa cell line DU145 using the previouslydescribed Ago-2 antibody 11A9 and an appropriate isotypecontrol [15 16] The mRNAs present in the Ago-precipitatewere quantitated by microarray analysis The relative abun-dance of RISC-associated mRNAs in the two cell lines arepresented in Supplementary Table S1 Twenty-four mRNAsthat exhibited a pronounced enrichment or depletion in theAgo complexes of DU145 cell are presented in Table 1 Supple-mentary Table S4 lists the functions (if known) of the genes inTable 1 and if they are potential or known targets of miRNAsderegulated in PCa For instance the highest abundance inthe Ago complexes of DU145 cells was observed for lamininalpha 3 (LAMA3) Expression of this gene is among othermembers in its family significantly reduced in prostate carci-noma [23] in line with being a potential target for miRNAsmiR-20a and miR-106a (httpwwwtargetscanorg) bothof which are up-regulated in prostate carcinoma [8 19] Inaddition to the LAMA3 mRNA the highly RISC-enrichedadenylate cyclase 3 (ADCY3) mRNA is a potential target forthe upregulatedmiRNAsmiR-25 andmiR-27ab Converselythe Thy-1 cell surface antigen (THY1) has been shown to beupregulated in PCa [24] Accordingly the THY1 mRNA levelwas reduced in the Ago complex of the DU145 cells
4 Prostate Cancer
Table 1 Abundance of selected mRNAs in AGO complexes of DU145 PCa cells in comparison to prostate normal fibroblasts (PNF08)
Gene title Gene symbol EnrichmentThy-1 cell surface antigen THY1 lt001Matrix Gla protein MGP lt001Periostin osteoblast specific factor POSTN lt001Secreted protein acidic cysteine-rich (osteonectin) SPARC lt001Chemokine (C-C motif) ligand 2 CCL2 lt001Cathepsin K CTSK 0059Tyrosine 3-monooxygenasetryptophan 5-monooxygenase activation protein epsilon polypeptide YWHAE 0093Peroxiredoxin 3 PRDX3 0158Coronin actin binding protein 1C CORO1C 0187Tumor protein p53 inducible nuclear protein 1 TP53INP1 0214Transferrin receptor (p90 CD71) TFRC 0278Laminin alpha 3 LAMA3 6891Etoposide induced 24 mRNA EI24 6813Adenylate cyclase 3 ADCY3 6001Succinate dehydrogenase complex subunit A SDHA 5973Adiponectin receptor 2 ADIPOR2 5187Nemo-like kinase NLK 4582ATM interactor ATMIN 4430RAB1B member RAS oncogene family RAB1B 4293Death effector domain containing DEDD 4019Tumor necrosis factor receptor superfamily member 10b TNFRSF10B 3635abl-interactor 2 ABI2 3267cullin 5 CUL5 3144S100 calcium binding protein A16 S100A16 3069
We then selected 10 mRNAs that based on our resultssuggest a possible deregulation in PCa We analyzed byqRT-PCR the relative expression of these genes in PNF-08 cells in comparison with DU145 and LNCaP cells SixmRNAs were selected that exhibited an enrichment in theAgo complexes from PCa cells and were therefore potentiallydownregulated in cell lines and tumors Etoposide induced24 mRNA (EI24 alias p53-induced gene 8 protein) S100calcium binding protein A16 (S100A16) Nemo-like kinase(NLK) RAS oncogene family member 1B (RAB1B) deatheffector domain-containing protein (DEDD) and cullin 5(CUL5) On the other end we selected mRNAs that showeddepletion from Ago complexes of PCa cells and therefore arepotentially overexpressed in PCa cells and tumorsTheseweretyrosine 3-Monooxygenase (YWHAE aka 14-3-3 proteinepsilon) transferrin receptor (TRFC) coronin actin bindingprotein 1C (CORO1C) and periredoxin 3 (PRDX3) Theresults of the analysis are shown in Figure 1
We initially expected that mRNAs enriched in Agocomplexes undergo degradation by deadenylation or by 51015840-decapping which would result in a reduced overall mRNAcontent in whole-cell RNA preparations Interestingly the re-sults of quantitative PCR analyses demonstrated that mRNAsenriched in the Ago complexes of PCa cells were presentat higher amounts in whole-cell RNA and mRNAs depletedfromAgo complexes of PCa cells exhibited a reduced amount
Only twoout of the selected 10mRNAs (S100A16 andPRDX3)followed our initial expectation These results demonstratethat an enrichment of mRNAs in Ago complexes does notnecessarily lead to an enhanced degradation of the respectivemRNA
EI24 as an Etoposide-inducible gene was chosen becauseEtoposide has recently been suggested as one componentof a combination chemotherapy treatment of advanced PCa[25] YWHAE is among the 14-3-3 proteins that regulate theETV1 transcription factor in PCa [26] TRFC was shownoriginally to be significantly induced in DU145 cells [27]CORO1Ccoronin was found to be induced in androgen-insensitive PCa [28] and S100A16 appears to be upregulatedonly in metastatic tumor cells [29]
NLK acts as a 120573-catenin pathway inhibitor throughphosphorylation and degradation of TCFLEF transcriptionfactors [30] NLK and DEDD are suggested to induce apop-tosis [31 32] and loss of RAB1B was shown to enhance theaggressiveness of breast carcinoma [33] A reduction of CUL5expression was found for instance in breast cancer [34] butso far not in prostate carcinoma Finally PRDX3 was chosenas it was previously found to be upregulated in primaryPCa [35] For this gene our initial assumption holds truewhere a reduced abundance of the PRDX3 mRNA in Agocomplexes of PCa cells results in an elevated mRNA contentin whole cells In addition the 31015840UTRs of DEDD and PRDX3
Prostate Cancer 5
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
EI24CUL5DEDDRAB1BNLKS100A16CORO1CTFRCYWHAEPRDX3
0
1
2
3
4
Rela
tive e
xpre
ssio
n (2minusΔΔCT
)
Figure 1 Quantification of mRNA expression in primary normal prostate (PNF-08) cells and the DU145 and LNCaP PCa cell lines Theexpression of ten mRNAs that were assumed to be elevated or reduced according to their presence or absence in the Ago complex wasassessed by qRT-PCR YWHAE TRFC CORO1C and PRDX3 were predicted to be elevated while NLK RAB1B DEDD CUL5 EI24 andS100A16 were predicted to be reduced in PCa cells as compared to PNF-08 cells
were analyzed as potential targets for miRNAs known to bederegulated in PCa (see below)
To gain additional insight into the miRNA-based regu-lation of tumor relevant genes we selected genes form thelist of Ago-associated mRNAs for their potential regulationby miRNAs (refer to Supplementary Table S2) For thisanalysis we chose DEDD the tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10B) the tumorprotein p53 inducible nuclear protein 1 (TP53INP1) thesecreted protein acidic cysteine-rich (SPARC osteonectin)the Abelson interactor 2 protein (ABI2) and the periredoxin3 (PRDX3) gene
These mRNAs are potential targets for previously identi-fied PCa-deregulated miRNAs miR-15b miR-20a and miR-148a (DEDD) miR-20a and miR-21 (TNFRSF10B) miR-24 and -29ab (TP53INP1) miR-29ab (SPARC) miR-15bmiR-200c and miR-375 (ABI2) and miR-23a (PRDX3) Thepotential binding sites of the miRNAs in the 31015840UTRs ofanalyzed mRNAs are depicted in Figure 2 Figure 3 showsthe negative regulatory effects of the miRNAs towards thereporter gene under the regulatory control of their responsive31015840UTRs We could demonstrate negative regulatory effectsof miR-148a towards the DEDD 31015840UTR (Figure 3(a)) ofmiR-20a towards the TNFRSF10B 31015840UTR (Figure 3(b)) ofmiR-24 towards the TP53INP1 31015840UTR (Figure 3(c)) and ofboth miR-29a and miR -29b towards the SPARC 31015840UTR(Figure 3(d)) The 31015840UTRs of ABI2 and PRDX3 were notresponsive towards miRNAs miR-375 miR-200c miR-15bandmiR-23a respectively andwere thus not analyzed further(Figures 3(e) and 3(f))
To further validate the results of the negative regulationof the reporter gene by miRNA responsive 31015840UTRs thepredicted binding sites for the miRNAs were mutated andanalyzed in parallel with the wild-type-constructs As shown
in Figure 4 the mutation of the seed-sequences for miR-148a in the DEDD 31015840UTR (Figure 4(a)) for miR-20a inthe TNFRSF10B 31015840UTR (Figure 4(b)) and miR-24 in theTP53INP1 31015840UTR (Figure 4(c)) resulted in loss of responsive-ness establishing these mRNAs as targets for the respectivemiRNAs In the SPARC 31015840UTR (Figure 4(d)) two potentialbinding sites are present and both of them can potentiallybe targeted by miR-29a and miR-29b Mutation of the firstbinding site (nt 89ndash110) had no effect on responsiveness whilethemutation of the second binding site (nt 121ndash143) or of bothbinding sites abrogated responsiveness to both miR-29a andmiR-29b This demonstrated that the second binding site inthe SPARC 31015840UTR is a target for miRNAs miR-29a and miR-29b
We then assayed the effect of overexpression of miR-24and miR-29ab on the protein levels of the TP53INP1 andSPARC protein respectively As shown in Figure 5(a) theTP53INP1 protein was downregulated in DU145 cells afteroverexpression of miR-24 Likewise both miRNAs miR-29aandmiR-29b reduced the SPARC protein levels (Figure 5(b))The determination of the relative downregulation is shown inFigures 5(c) and 5(d) TP53INP1 protein was downregulatedby about 40 and reduction of SPARC by miR-29a andmiR-29b was approx 60 and 70 respectively Next wetested the effect of the miRNAs on the growth behaviorof DU145 cells We previously showed that these miRNAsare downregulated in primary PCa tissue as compared tononmalignant prostatic tissue [11 19] First we analyzed thecolony formation capacity ofDU145 cells after overexpressionof the three miRNAs Interestingly miR-24 significantlyincreased both the number of colonies (Figure 6(a)) as well asthe colony size In contrast miR-29a showed no effect whilemiR-29b significantly decreased both the number as well asthe size of DU145 cell colonies (Figures 6(a) and 6(b))
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
4 Prostate Cancer
Table 1 Abundance of selected mRNAs in AGO complexes of DU145 PCa cells in comparison to prostate normal fibroblasts (PNF08)
Gene title Gene symbol EnrichmentThy-1 cell surface antigen THY1 lt001Matrix Gla protein MGP lt001Periostin osteoblast specific factor POSTN lt001Secreted protein acidic cysteine-rich (osteonectin) SPARC lt001Chemokine (C-C motif) ligand 2 CCL2 lt001Cathepsin K CTSK 0059Tyrosine 3-monooxygenasetryptophan 5-monooxygenase activation protein epsilon polypeptide YWHAE 0093Peroxiredoxin 3 PRDX3 0158Coronin actin binding protein 1C CORO1C 0187Tumor protein p53 inducible nuclear protein 1 TP53INP1 0214Transferrin receptor (p90 CD71) TFRC 0278Laminin alpha 3 LAMA3 6891Etoposide induced 24 mRNA EI24 6813Adenylate cyclase 3 ADCY3 6001Succinate dehydrogenase complex subunit A SDHA 5973Adiponectin receptor 2 ADIPOR2 5187Nemo-like kinase NLK 4582ATM interactor ATMIN 4430RAB1B member RAS oncogene family RAB1B 4293Death effector domain containing DEDD 4019Tumor necrosis factor receptor superfamily member 10b TNFRSF10B 3635abl-interactor 2 ABI2 3267cullin 5 CUL5 3144S100 calcium binding protein A16 S100A16 3069
We then selected 10 mRNAs that based on our resultssuggest a possible deregulation in PCa We analyzed byqRT-PCR the relative expression of these genes in PNF-08 cells in comparison with DU145 and LNCaP cells SixmRNAs were selected that exhibited an enrichment in theAgo complexes from PCa cells and were therefore potentiallydownregulated in cell lines and tumors Etoposide induced24 mRNA (EI24 alias p53-induced gene 8 protein) S100calcium binding protein A16 (S100A16) Nemo-like kinase(NLK) RAS oncogene family member 1B (RAB1B) deatheffector domain-containing protein (DEDD) and cullin 5(CUL5) On the other end we selected mRNAs that showeddepletion from Ago complexes of PCa cells and therefore arepotentially overexpressed in PCa cells and tumorsTheseweretyrosine 3-Monooxygenase (YWHAE aka 14-3-3 proteinepsilon) transferrin receptor (TRFC) coronin actin bindingprotein 1C (CORO1C) and periredoxin 3 (PRDX3) Theresults of the analysis are shown in Figure 1
We initially expected that mRNAs enriched in Agocomplexes undergo degradation by deadenylation or by 51015840-decapping which would result in a reduced overall mRNAcontent in whole-cell RNA preparations Interestingly the re-sults of quantitative PCR analyses demonstrated that mRNAsenriched in the Ago complexes of PCa cells were presentat higher amounts in whole-cell RNA and mRNAs depletedfromAgo complexes of PCa cells exhibited a reduced amount
Only twoout of the selected 10mRNAs (S100A16 andPRDX3)followed our initial expectation These results demonstratethat an enrichment of mRNAs in Ago complexes does notnecessarily lead to an enhanced degradation of the respectivemRNA
EI24 as an Etoposide-inducible gene was chosen becauseEtoposide has recently been suggested as one componentof a combination chemotherapy treatment of advanced PCa[25] YWHAE is among the 14-3-3 proteins that regulate theETV1 transcription factor in PCa [26] TRFC was shownoriginally to be significantly induced in DU145 cells [27]CORO1Ccoronin was found to be induced in androgen-insensitive PCa [28] and S100A16 appears to be upregulatedonly in metastatic tumor cells [29]
NLK acts as a 120573-catenin pathway inhibitor throughphosphorylation and degradation of TCFLEF transcriptionfactors [30] NLK and DEDD are suggested to induce apop-tosis [31 32] and loss of RAB1B was shown to enhance theaggressiveness of breast carcinoma [33] A reduction of CUL5expression was found for instance in breast cancer [34] butso far not in prostate carcinoma Finally PRDX3 was chosenas it was previously found to be upregulated in primaryPCa [35] For this gene our initial assumption holds truewhere a reduced abundance of the PRDX3 mRNA in Agocomplexes of PCa cells results in an elevated mRNA contentin whole cells In addition the 31015840UTRs of DEDD and PRDX3
Prostate Cancer 5
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
EI24CUL5DEDDRAB1BNLKS100A16CORO1CTFRCYWHAEPRDX3
0
1
2
3
4
Rela
tive e
xpre
ssio
n (2minusΔΔCT
)
Figure 1 Quantification of mRNA expression in primary normal prostate (PNF-08) cells and the DU145 and LNCaP PCa cell lines Theexpression of ten mRNAs that were assumed to be elevated or reduced according to their presence or absence in the Ago complex wasassessed by qRT-PCR YWHAE TRFC CORO1C and PRDX3 were predicted to be elevated while NLK RAB1B DEDD CUL5 EI24 andS100A16 were predicted to be reduced in PCa cells as compared to PNF-08 cells
were analyzed as potential targets for miRNAs known to bederegulated in PCa (see below)
To gain additional insight into the miRNA-based regu-lation of tumor relevant genes we selected genes form thelist of Ago-associated mRNAs for their potential regulationby miRNAs (refer to Supplementary Table S2) For thisanalysis we chose DEDD the tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10B) the tumorprotein p53 inducible nuclear protein 1 (TP53INP1) thesecreted protein acidic cysteine-rich (SPARC osteonectin)the Abelson interactor 2 protein (ABI2) and the periredoxin3 (PRDX3) gene
These mRNAs are potential targets for previously identi-fied PCa-deregulated miRNAs miR-15b miR-20a and miR-148a (DEDD) miR-20a and miR-21 (TNFRSF10B) miR-24 and -29ab (TP53INP1) miR-29ab (SPARC) miR-15bmiR-200c and miR-375 (ABI2) and miR-23a (PRDX3) Thepotential binding sites of the miRNAs in the 31015840UTRs ofanalyzed mRNAs are depicted in Figure 2 Figure 3 showsthe negative regulatory effects of the miRNAs towards thereporter gene under the regulatory control of their responsive31015840UTRs We could demonstrate negative regulatory effectsof miR-148a towards the DEDD 31015840UTR (Figure 3(a)) ofmiR-20a towards the TNFRSF10B 31015840UTR (Figure 3(b)) ofmiR-24 towards the TP53INP1 31015840UTR (Figure 3(c)) and ofboth miR-29a and miR -29b towards the SPARC 31015840UTR(Figure 3(d)) The 31015840UTRs of ABI2 and PRDX3 were notresponsive towards miRNAs miR-375 miR-200c miR-15bandmiR-23a respectively andwere thus not analyzed further(Figures 3(e) and 3(f))
To further validate the results of the negative regulationof the reporter gene by miRNA responsive 31015840UTRs thepredicted binding sites for the miRNAs were mutated andanalyzed in parallel with the wild-type-constructs As shown
in Figure 4 the mutation of the seed-sequences for miR-148a in the DEDD 31015840UTR (Figure 4(a)) for miR-20a inthe TNFRSF10B 31015840UTR (Figure 4(b)) and miR-24 in theTP53INP1 31015840UTR (Figure 4(c)) resulted in loss of responsive-ness establishing these mRNAs as targets for the respectivemiRNAs In the SPARC 31015840UTR (Figure 4(d)) two potentialbinding sites are present and both of them can potentiallybe targeted by miR-29a and miR-29b Mutation of the firstbinding site (nt 89ndash110) had no effect on responsiveness whilethemutation of the second binding site (nt 121ndash143) or of bothbinding sites abrogated responsiveness to both miR-29a andmiR-29b This demonstrated that the second binding site inthe SPARC 31015840UTR is a target for miRNAs miR-29a and miR-29b
We then assayed the effect of overexpression of miR-24and miR-29ab on the protein levels of the TP53INP1 andSPARC protein respectively As shown in Figure 5(a) theTP53INP1 protein was downregulated in DU145 cells afteroverexpression of miR-24 Likewise both miRNAs miR-29aandmiR-29b reduced the SPARC protein levels (Figure 5(b))The determination of the relative downregulation is shown inFigures 5(c) and 5(d) TP53INP1 protein was downregulatedby about 40 and reduction of SPARC by miR-29a andmiR-29b was approx 60 and 70 respectively Next wetested the effect of the miRNAs on the growth behaviorof DU145 cells We previously showed that these miRNAsare downregulated in primary PCa tissue as compared tononmalignant prostatic tissue [11 19] First we analyzed thecolony formation capacity ofDU145 cells after overexpressionof the three miRNAs Interestingly miR-24 significantlyincreased both the number of colonies (Figure 6(a)) as well asthe colony size In contrast miR-29a showed no effect whilemiR-29b significantly decreased both the number as well asthe size of DU145 cell colonies (Figures 6(a) and 6(b))
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Prostate Cancer 5
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
DU145
LNCa
PPN
F08
EI24CUL5DEDDRAB1BNLKS100A16CORO1CTFRCYWHAEPRDX3
0
1
2
3
4
Rela
tive e
xpre
ssio
n (2minusΔΔCT
)
Figure 1 Quantification of mRNA expression in primary normal prostate (PNF-08) cells and the DU145 and LNCaP PCa cell lines Theexpression of ten mRNAs that were assumed to be elevated or reduced according to their presence or absence in the Ago complex wasassessed by qRT-PCR YWHAE TRFC CORO1C and PRDX3 were predicted to be elevated while NLK RAB1B DEDD CUL5 EI24 andS100A16 were predicted to be reduced in PCa cells as compared to PNF-08 cells
were analyzed as potential targets for miRNAs known to bederegulated in PCa (see below)
To gain additional insight into the miRNA-based regu-lation of tumor relevant genes we selected genes form thelist of Ago-associated mRNAs for their potential regulationby miRNAs (refer to Supplementary Table S2) For thisanalysis we chose DEDD the tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10B) the tumorprotein p53 inducible nuclear protein 1 (TP53INP1) thesecreted protein acidic cysteine-rich (SPARC osteonectin)the Abelson interactor 2 protein (ABI2) and the periredoxin3 (PRDX3) gene
These mRNAs are potential targets for previously identi-fied PCa-deregulated miRNAs miR-15b miR-20a and miR-148a (DEDD) miR-20a and miR-21 (TNFRSF10B) miR-24 and -29ab (TP53INP1) miR-29ab (SPARC) miR-15bmiR-200c and miR-375 (ABI2) and miR-23a (PRDX3) Thepotential binding sites of the miRNAs in the 31015840UTRs ofanalyzed mRNAs are depicted in Figure 2 Figure 3 showsthe negative regulatory effects of the miRNAs towards thereporter gene under the regulatory control of their responsive31015840UTRs We could demonstrate negative regulatory effectsof miR-148a towards the DEDD 31015840UTR (Figure 3(a)) ofmiR-20a towards the TNFRSF10B 31015840UTR (Figure 3(b)) ofmiR-24 towards the TP53INP1 31015840UTR (Figure 3(c)) and ofboth miR-29a and miR -29b towards the SPARC 31015840UTR(Figure 3(d)) The 31015840UTRs of ABI2 and PRDX3 were notresponsive towards miRNAs miR-375 miR-200c miR-15bandmiR-23a respectively andwere thus not analyzed further(Figures 3(e) and 3(f))
To further validate the results of the negative regulationof the reporter gene by miRNA responsive 31015840UTRs thepredicted binding sites for the miRNAs were mutated andanalyzed in parallel with the wild-type-constructs As shown
in Figure 4 the mutation of the seed-sequences for miR-148a in the DEDD 31015840UTR (Figure 4(a)) for miR-20a inthe TNFRSF10B 31015840UTR (Figure 4(b)) and miR-24 in theTP53INP1 31015840UTR (Figure 4(c)) resulted in loss of responsive-ness establishing these mRNAs as targets for the respectivemiRNAs In the SPARC 31015840UTR (Figure 4(d)) two potentialbinding sites are present and both of them can potentiallybe targeted by miR-29a and miR-29b Mutation of the firstbinding site (nt 89ndash110) had no effect on responsiveness whilethemutation of the second binding site (nt 121ndash143) or of bothbinding sites abrogated responsiveness to both miR-29a andmiR-29b This demonstrated that the second binding site inthe SPARC 31015840UTR is a target for miRNAs miR-29a and miR-29b
We then assayed the effect of overexpression of miR-24and miR-29ab on the protein levels of the TP53INP1 andSPARC protein respectively As shown in Figure 5(a) theTP53INP1 protein was downregulated in DU145 cells afteroverexpression of miR-24 Likewise both miRNAs miR-29aandmiR-29b reduced the SPARC protein levels (Figure 5(b))The determination of the relative downregulation is shown inFigures 5(c) and 5(d) TP53INP1 protein was downregulatedby about 40 and reduction of SPARC by miR-29a andmiR-29b was approx 60 and 70 respectively Next wetested the effect of the miRNAs on the growth behaviorof DU145 cells We previously showed that these miRNAsare downregulated in primary PCa tissue as compared tononmalignant prostatic tissue [11 19] First we analyzed thecolony formation capacity ofDU145 cells after overexpressionof the three miRNAs Interestingly miR-24 significantlyincreased both the number of colonies (Figure 6(a)) as well asthe colony size In contrast miR-29a showed no effect whilemiR-29b significantly decreased both the number as well asthe size of DU145 cell colonies (Figures 6(a) and 6(b))
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Figure 2 Predicted miRNA target sites in 31015840UTRs The 31015840UTR regions of DEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e)and PRDX3 (f) are depicted A schematic representation of the predicted miRNAs and their interaction site(s) is shown
4 Discussion
We could demonstrate that a large amount of the mRNAsthat we identified as differentially represented in the Agocomplexes of DU145 cells have already been described asdifferentially expressed in primary PCa Although miRNAsact as the guiding element for the RISC factor it is stillnot fully understood how the fate of a targeted mRNAis decided The binding of miRISC complexes can equallyaffect only protein translation without measurably alteringthe mRNA content of the cell Another source of variabilityexists in the fact that the level of a given miRNA does notnecessarily reflect its presence in the RISC complexes [36]Furthermore La Rocca and coworkers have demonstratedthat miRNAs in resting tissues are predominantly found inlow-molecular weight complexes not associated with mRNAwhile in growing tissues the majority of miRNAs are foundin high molecular weight complexes bound to mRNAs [37]Here we used growing cells where this effect might bedisregarded
In addition to the alreadymentioned LAMA3mRNA thehighly RISC-enriched adenylate cyclase 3 (ADCY3) mRNAis a potential target for the upregulated miRNAs miR-25 andmiR-27ab [8 19] In contrast overexpression of MICB the
likewise highly enriched mRNA in the Ago complex is con-sidered to have a tumor-promoting rather than suppressingfunction [38] The also enriched succinate dehydrogenasecomplex subunit A flavoprotein (SDHA) a potential tumorsuppressor [39] was not found to be differentially expressedin normal versus PCa tissue [40] However its presence inthe Ago complex might indicate that SDHA is reduced atthe protein level in PCa In line with this assumption it hasbeen demonstrated that loss of SDH expression leads to apseudohypoxia signaling [41] As further examples the Ago-IP enriched mRNAs ATM interactor (ATMIN) consideredto be a tumor suppressor [42] and the adiponectin receptor 2(ADIPOR2) a known inhibitor of proliferation [43] are bothpotential targets for the upregulated miR-200c Howevera recent study described a strong correlation of increasedADIPOR2 expression with poor prognosis of PCa [44] Ofthe genes found to be present at reduced levels in the Agocomplex (Table 1) the Thy-1 cell surface antigen (THY1)[24] chemokine (C-C motif ligand 2 (CCL2 MCP1)) [45]matrix Gla protein (MGP) [46] periostin (POSTN) [47 48]secreted protein acidic cysteine-rich (SPARC osteonectin)[49] or cathepsin K (CSTK) [50] have been found to beoverexpressed in PCa
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Prostate Cancer 7
pMIR pMIR-DEDD 3998400UTR
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
Con
trol
miR
-15b
miR
-20
a
miR
-148
a
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(a)
pMIR pMIR-TNFRSF10B 3998400UTR
Con
trol
miR
-20
a0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
lowastlowastlowast
miR
-21
Con
trol
miR
-20
a
miR
-21
(b)
Con
trol
miR
-29
a
miR
-29b
miR
-24
Con
trol
miR
-29
a
miR
-29b
miR
-24
pMIR pMIR-TP53INP1 3998400UTR
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29
a
miR
-29b
Con
trol
miR
-29
a
miR
-29b
pMIR-SPARC 3998400UTR
lowastlowastlowast
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)pMIR
(d)
Con
trol
miR
-200
c
miR
-375
miR
-15b
Con
trol
miR
-200
c
miR
-375
miR
-15b
pMIR 3998400UTR
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-ABI2
(e)
Con
trol
miR
-23a
Con
trol
miR
-23a
pMIR 3998400UTRpMIR PRDX3
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(f)
Figure 3 Response of 31015840UTRs towardsmiRNAsTheDEDD (a) TNFRSF10B (b) TP53INP1 (c) SPARC (d) ABI2 (e) and PRDX3 (f) 31015840UTRswere cloned behind the luciferase reporter gene of the pMIR vectorThe reporter gene constructs were expressed with the miRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
8 Prostate Cancer
Con
trol
miR
-148
a
Con
trol
miR
-148
a
Con
trol
miR
-148
a
pMIR pMIR-DEDD pMIR-DEDD mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180Re
lativ
e luc
ifera
se ac
tivity
()
(a)
Con
trol
miR
-20a
Con
trol
miR
-20a
Con
trol
miR
-20a
pMIR pMIR-TNFRSF10B pMIR-TNFRSF10B mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(b)
Con
trol
miR
-24
Con
trol
miR
-24
Con
trol
miR
-24
pMIR pMIR-TP53INP1 pMIR-TP53INP1 mut
lowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
(c)
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
Con
trol
miR
-29a
miR
-29b
pMIR pMIR-SPARC
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
0
20
40
60
80
100
120
140
160
180
Relat
ive l
ucife
rase
activ
ity (
)
pMIR-SPARCmut I
pMIR-SPARCmut II
pMIR-SPARCmut I + II
(d)
Figure 4 Identification of miRNA binding sites The potential binding sites for the indicated miRNAs in the 31015840UTRs of DEDD (a)TNFRSF10B (b) TP53INP1 (c) and SPARC (d) weremutated by site directedmutagenesis (mut) and coexpressed with themiRNA expressionconstruct or with the empty pSG5 vector in the indicated combinations Results represent the mean of 4 independent experiments performedin duplicate lowastlowastlowast119901 lt 0001
We demonstrate that the 31015840UTRs of the DEDDTNFRSF10BTRAIL TP53INP1 and SPARCosteonectinmRNAs are targets for miR-148a miR-20a miR-24 miR-29aandmiR-29b respectively In our previous analysis miR-148aand miR-20ab were among the strongest induced miRNAsin primary PCa versus normal tissue [8 19] in line withthe high abundance of the DEDD and TNFRSF10BTRAILmRNAs in the RISC complexes (Table 1) So far noassociation of DEDD with prostate carcinoma has beendescribed but a reduction of this protein might protectPCa cells from undergoing apoptosis DEDD can attenuateepithelial-mesenchymal transition and acts as an endogenoussuppressor of tumor growth and metastasis in human breastcancer [51] However overexpression of DEDDwas observedin HP1618 transformed cervical carcinoma cells [52] Alsocopy number gain of 1q233 which included amplificationof DEDD in urothelial tumors was recently described [53]
as well as increased DEDD expression in nasopharyngealcarcinoma [54] A function for DEDD in transcriptionalactivation via NFKB in addition to its role in apoptosishas been described [55] The importance of DEDD in PCashould thus to be explored further Likewise our resultssuggest a role for cullin 5 (CUL5) in prostate carcinomaCUL5 (VACM-1) is located on chromosome 11q22-q23 [56]a genomic region associated with loss of heterozygosity inbreast cancer [57] and might therefore be considered as atumor suppressor For instance it inhibits the degradation oftumor suppressor proteins and is present at reduced levelsin cell lines derived from breast colon melanoma andnon-small lung cancer cell lines its reduced levels might alsodesensitize cancer cells during chemotherapy [58]
The downregulation of tumor necrosis factor receptorsuperfamily member 10b protein (TNFRSF10BTRAIL) byupregulated miR-20a [8 19] is in line with the resistance of
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Prostate Cancer 9
Control miR-24
DU145
(a)
Control miR-29a miR-29b
DU145
(b)
0
20
40
60
80
100
120
Relat
ive T
P53I
NP1
sign
al in
tens
ity (
)
(c)
0
20
40
60
80
100
120
Relat
ive S
PARC
sign
al in
tens
ity (
)
(d)
TP53
INP1
SPA
RC
miR-24Control miR-29bmiR-29aControl
120573-A
ctin
120573-A
ctin
Figure 5 Regulation of TP53INP1 and SPARC protein expression by miR-24 and 29ab DU145 cells were transfected either with controlvectors or miRNA expression vectors 48 hours after transfection the protein expression of (a) TP53INP1 and (b) SPARC was determined bywestern blot using 120573-actin as loading control The quantification of the western blots in (c) and (d) represents the relative downregulation asdetermined in three independent experiments using 120573-actin as loading control
the PCa cell lines LNCaP DU145 or PC3 towards TRAILTNFRSF10B-induced apoptosis [59] and its downregulationin PCa [60] We further show that SPARCosteonectin andTP53INP1 are targets for miRNAs miR-29ab and miR-24respectively All three miRNAs are downregulated in PCaIn line with these observations we found reduced proteinlevels of TP53INP1 and SPARCosteonectin after overex-pression of the respective targeting miRNAs in DU145 cells(Figure 5) SPARCosteonectin is upregulated and promotesthe malignant behavior of PCa [49] These findings arecorroborated by the known function ofmiRNAsmiR-29abcin inhibiting the growth and invasiveness of PCa [61] andare high-lighted by the inhibition of cell growth by miR-29bOverexpression of TP53INP1 has been observed in PCa andis predictive of biological cancer relapse [62] Interestinglyoverexpression of miR-24 exerted a positive effect on thegrowth behavior of DU145 cells as determined by colonyformation assays (Figure 6) It is possible that the downreg-ulation of miR-24 and its possible tumor-promoting effectworks only in cooperationwith other factors such asmiRNAsIn mammary carcinoma overexpression of miR-24 enhances
proliferation and metastatic potential [63] Likewise miR-24 overexpression increases cell proliferation in non-smallcell lung cancer [64] Furthermore miR-24 targets PTENin human tongue squamous cell carcinoma which couldalso explain the increase in growth rate [65] ConverselymiR-24 functions as a tumor suppressor in nasopharyngealcarcinoma [66]
The data by Flores et al [36] and La Rocca et al [37]show that mRNAs are differentially associated with theRISC complex and that the majority of Ago complexes arenot associated with mRNAs in resting tissues The presentapproach using growing cell lines was chosen to avoid the lat-ter problem In conjunction with our previously establishedmiRNA expression data we were able to mechanisticallyconfirm the regulation of four out of six selected genes bymiRNAs deregulated in PCa Our data might be useful forthe detection of novel deregulated mRNAs as we provide alist of mRNAs highly enriched in the Ago complexes or thosethat are only presented at low levels Our experimental datahowever does not fully support our initial hypothesis that the
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
10 Prostate Cancer
0
02
04
06
08
10
12
14
16
18
Relat
ive c
olou
r uni
ts
Con
trol
miR
-24
miR
-29a
miR
-29b
(a)
0
2
4
6
8
10
12
14
Cel
l num
ber
ControlmiR-24
miR-29amiR-29b
48 72240Time (h)
times105
(b)
Figure 6 Effect of miR-24 and miR-29ab on cell growth DU145 cells were transfected with miRNA expression vectors or control vectorsand seeded in a limited cell number Eight days after seeding colonies were stained with crystal violet (a) Colony formation was quantified bydensitometry analyses Data show the mean and plusmn SEM of the densitometry analysis of CFA (119901 lt 0001) of three independent experimentsCell growth was determined by automated cell counting of parallel experiments at 24 48 and 72 h after transfection (b)
presence or absence of amRNA in these complexes affects theexpression of the corresponding gene product
Competing Interests
The authors declare no conflict of interests
Acknowledgments
This work was supported by Wilhelm-Sander-Stiftung by agrant to Bernd Wullich and Friedrich A Grasser (Grant no200702501) Gunter Meister was supported by the DeutscheForschungsgemeinschaft (DFG Forschergruppe FOR2127)Friedrich A Grasser was also supported by an intramuralgrant (HOMFOR)
References
[1] J Ferlay H-R Shin F Bray D Forman C Mathers and DM Parkin ldquoEstimates of worldwide burden of cancer in 2008GLOBOCAN2008rdquo International Journal of Cancer vol 127 no12 pp 2893ndash2917 2010
[2] A D Choudhury R Eeles S J Freedland et al ldquoThe roleof genetic markers in the management of prostate cancerrdquoEuropean Urology vol 62 no 4 pp 577ndash587 2012
[3] C Jeronimo P J Bastian A Bjartell et al ldquoEpigenetics inprostate cancer biologic and clinical relevancerdquo EuropeanUrology vol 60 no 4 pp 753ndash766 2011
[4] S Jonas and E Izaurralde ldquoTowards amolecular understandingof microRNA-mediated gene silencingrdquo Nature Reviews Genet-ics vol 16 no 7 pp 421ndash433 2015
[5] R Spizzo M S Nicoloso C M Croce and G A CalinldquoSnapShot MicroRNAs in cancerrdquo Cell vol 137 no 3 p 586e12009
[6] G Meister ldquoArgonaute proteins functional insights and emerg-ing rolesrdquo Nature Reviews Genetics vol 14 pp 447ndash459 2013
[7] M Maugeri-Sacca V Coppola D Bonci and R De MarialdquoMicroRNAs and prostate cancer from preclinical research totranslational oncologyrdquo Cancer Journal vol 18 no 3 pp 253ndash261 2012
[8] M Hart E Nolte S Wach et al ldquoComparative micrornaprofiling of prostate carcinomas with increasing tumor stage bydeep sequencingrdquoMolecular Cancer Research vol 12 no 2 pp250ndash263 2014
[9] S Wach E Nolte J Szczyrba et al ldquoMicroRNA profilesof prostate carcinoma detected by multiplatform microRNAscreeningrdquo International Journal of Cancer vol 130 no 3 pp611ndash621 2012
[10] M Hart S Wach E Nolte et al ldquoThe proto-oncogene ERG is atarget of microRNA miR-145 in prostate cancerrdquo FEBS Journalvol 280 no 9 pp 2105ndash2116 2013
[11] J Szczyrba E Nolte M Hart et al ldquoIdentification of ZNF217hnRNP-K VEGF-A and IPO7 as targets formicroRNAs that aredownregulated in prostate carcinomardquo International Journal ofCancer vol 132 no 4 pp 775ndash784 2013
[12] J Szczyrba E Nolte S Wach et al ldquoDownregulation of Sec23Aprotein by miRNA-375 in prostate carcinomardquoMolecular Can-cer Research vol 9 no 6 pp 791ndash800 2011
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
Prostate Cancer 11
[13] A Dueck and G Meister ldquoAssembly and function of smallRNAmdashargonaute protein complexesrdquo Biological Chemistry vol395 no 6 pp 611ndash629 2014
[14] M R Fabian and N Sonenberg ldquoThe mechanics of miRNA-mediated gene silencing a look under the hood of miRISCrdquoNature Structural and Molecular Biology vol 19 no 6 pp 586ndash593 2012
[15] S Rudel A Flatley L Weinmann E Kremmer and G MeisterldquoA multifunctional human Argonaute2-specific monoclonalantibodyrdquo RNA vol 14 no 6 pp 1244ndash1253 2008
[16] M Beitzinger L Peters J Y Zhu E Kremmer and G MeisterldquoIdentification of human microRNA targets from isolated arg-onaute protein complexesrdquoRNABiology vol 4 no 2 pp 76ndash842007
[17] F V Karginov C Conaco Z Xuan et al ldquoA biochemicalapproach to identifying microRNA targetsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 104 no 49 pp 19291ndash19296 2007
[18] G Easow A A Teleman and S M Cohen ldquoIsolation ofmicroRNA targets by miRNP immunopurificationrdquo RNA vol13 no 8 pp 1198ndash1204 2007
[19] J Szczyrba E Loprich S Wach et al ldquoThe microRNA profileof prostate carcinoma obtained by deep sequencingrdquoMolecularCancer Research vol 8 no 4 pp 529ndash538 2010
[20] M Beitzinger and G Meister ldquoExperimental identificationof microRNA targets by immunoprecipitation of argonauteprotein complexesrdquoMetHods In Molecular Biology vol 732 pp153ndash167 2011
[21] RA Irizarry BHobbs F Collin et al ldquoExploration normaliza-tion and summaries of high density oligonucleotide array probelevel datardquo Biostatistics vol 4 no 2 pp 249ndash264 2003
[22] T D Schmittgen and K J Livak ldquoAnalyzing real-time PCR databy the comparative C
119879methodrdquo Nature Protocols vol 3 no 6
pp 1101ndash1108 2008[23] U G Sathyanarayana A Padar M Suzuki et al ldquoAberrant
promoter methylation of laminin-5-encoding genes in prostatecancers and its relationship to clinicopathological featuresrdquoClinical Cancer Research vol 9 no 17 pp 6395ndash6400 2003
[24] LD TrueH ZhangM Ye et al ldquoCD90THY1 is overexpressedin prostate cancer-associated fibroblasts and could serve as acancer biomarkerrdquoModern Pathology vol 23 no 10 pp 1346ndash1356 2010
[25] S A Rosenthal K Bae K J Pienta et al ldquoPhase III Multi-institutional trial of adjuvant chemotherapy with paclitaxelestramustine and oral etoposide combined with long-termandrogen suppression therapy and radiotherapy versus long-term androgen suppression plus radiotherapy alone for high-risk prostate cancer preliminary toxicity analysis of RTOG 99-02rdquo International Journal of Radiation Oncology Biology Physicsvol 73 no 3 pp 672ndash678 2009
[26] S Oh S Shin S A Lightfoot andR Janknecht ldquo14-3-3 proteinsmodulate the ets transcription factor ETV1 in prostate cancerrdquoCancer Research vol 73 no 16 pp 5110ndash5119 2013
[27] H N Keer J M Kozlowski Y C Tsai C Lee R N McEwanand J T Grayhack ldquoElevated transferrin receptor content inhuman prostate cancer cell lines assessed in vitro and in vivordquoThe Journal of Urology vol 143 no 2 pp 381ndash385 1990
[28] H Zhao Y Kim PWang et al ldquoGenome-wide characterizationof gene expression variations and DNA copy number changesin prostate cancer cell linesrdquo Prostate vol 63 no 2 pp 187ndash1972005
[29] D A Smirnov D R Zweitzig B W Foulk et al ldquoGlobal geneexpression profiling of circulating tumor cellsrdquoCancer Researchvol 65 no 12 pp 4993ndash4997 2005
[30] T Ishitani S Kishida J Hyodo-Miura et al ldquoThe TAK1-NLKmitogen-activated protein kinase cascade functions in theWnt-5aCa2+ pathway to antagonize Wnt120573-catenin signalingrdquoMolecular and Cellular Biology vol 23 no 1 pp 131ndash139 2003
[31] K H Emami L G Brown T E M Pitts X Sun R L Vessellaand E Corey ldquoNemo-like kinase induces apoptosis and inhibitsandrogen receptor signaling in prostate cancer cellsrdquo Prostatevol 69 no 14 pp 1481ndash1492 2009
[32] A H Stegh O Schickling A Ehret et al ldquoDEDD a novel deatheffector domain-containing protein targeted to the nucleolusrdquoThe EMBO Journal vol 17 no 20 pp 5974ndash5986 1998
[33] H-L Jiang H-F Sun S-P Gao et al ldquoLoss of RAB1B promotestriple-negative breast cancer metastasis by activating TGF-120573SMAD signalingrdquo Oncotarget vol 6 no 18 pp 16352ndash163652015
[34] M J Fay K A Longo G A Karathanasis et al ldquoAnalysis ofCUL-5 expression in breast epithelial cells breast cancer celllines normal tissues and tumor tissuesrdquoMolecular Cancer vol2 article 40 2003
[35] A Basu H Banerjee H Rojas et al ldquoDifferential expressionof peroxiredoxins in prostate cancer consistent upregulation ofPRDX3 and PRDX4rdquo Prostate vol 71 no 7 pp 755ndash765 2011
[36] O Flores E M Kennedy R L Skalsky and B R Cullen ldquoDif-ferential RISC association of endogenous human microRNAspredicts their inhibitory potentialrdquo Nucleic Acids Research vol42 no 7 pp 4629ndash4639 2014
[37] G La Rocca S H Olejniczak A J Gonzalez et al ldquoInvivo Argonaute-bound microRNAs exist predominantly in areservoir of low molecular weight complexes not associatedwith mRNArdquo Proceedings of the National Academy of Sciencesof the United States of America vol 112 no 3 pp 767ndash772 2015
[38] G Liu S Lu XWang et al ldquoPerturbation of NK cell peripheralhomeostasis accelerates prostate carcinomametastasisrdquo Journalof Clinical Investigation vol 123 no 10 pp 4410ndash4422 2013
[39] N Burnichon J-J Briere R Libe et al ldquoSDHA is a tumorsuppressor gene causing paragangliomardquo Human MolecularGenetics vol 19 no 15 pp 3011ndash3020 2010
[40] F Ohl M Jung C Xu et al ldquoGene expression studies inprostate cancer tissue which reference gene should be selectedfor normalizationrdquo Journal of Molecular Medicine vol 83 no12 pp 1014ndash1024 2005
[41] M Miettinen and J Lasota ldquoSuccinate dehydrogenase deficientgastrointestinal stromal tumors (GISTs)mdasha reviewrdquo Interna-tional Journal of Biochemistry and Cell Biology vol 53 pp 514ndash519 2014
[42] X Liu and S Zha ldquoATMIN a new tumor suppressor indeveloping B cellsrdquo Cancer Cell vol 19 no 5 pp 569ndash570 2011
[43] Q Gao and J Zheng ldquoAdiponectin-induced antitumor activityon prostatic cancers through inhibiting proliferationrdquo CellBiochemistry and Biophysics vol 70 no 1 pp 461ndash465 2014
[44] J R Rider M Fiorentino R Kelly et al ldquoTumor expression ofadiponectin receptor 2 and lethal prostate cancerrdquo Carcinogen-esis vol 36 no 6 pp 639ndash647 2015
[45] I Tsaur A Noack J Makarevic et al ldquoCCL2 chemokine as apotential biomarker for prostate cancer a pilot studyrdquo CancerResearch and Treatment vol 47 no 2 pp 306ndash312 2015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015
12 Prostate Cancer
[46] E N Levedakou T G Strohmeyer P J Effert and E T LiuldquoExpression of the matrix Gla protein in urogenital malignan-ciesrdquo International Journal of Cancer vol 52 no 4 pp 534ndash5371992
[47] T Tsunoda B Furusato Y Takashima et al ldquoThe increasedexpression of periostin during early stages of prostate cancerand advanced stages of cancer stromardquo Prostate vol 69 no 13pp 1398ndash1403 2009
[48] V Tischler F R Fritzsche P J Wild et al ldquoPeriostin is up-regulated in high grade and high stage prostate cancerrdquo BMCCancer vol 10 article no 273 2010
[49] K Jacob M Webber D Benayahu and H K KleinmanldquoOsteonectin promotes prostate cancer cell migration andinvasion a possible mechanism for metastasis to bonerdquo CancerResearch vol 59 no 17 pp 4453ndash4457 1999
[50] K D Brubaker R L Vessella L D True R Thomas and ECorey ldquoCathepsin K mRNA and protein expression in prostatecancer progressionrdquo Journal of Bone and Mineral Research vol18 no 2 pp 222ndash230 2003
[51] Q Lv W Wang J Xue et al ldquoDEDD interacts with PI3KC3to activate autophagy and attenuate epithelial-mesenchymaltransition in human breast cancerrdquo Cancer Research vol 72 no13 pp 3238ndash3250 2012
[52] G Vazquez-Ortiz J A Garcıa C J Ciudad et al ldquoDifferentiallyexpressed genes between high-risk human papillomavirus typesin human cervical cancer cellsrdquo International Journal of Gyne-cological Cancer vol 17 no 2 pp 484ndash491 2007
[53] M Riester L Werner J Bellmunt et al ldquoIntegrative analysis of1q233 copy-number gain in metastatic urothelial carcinomardquoClinical Cancer Research vol 20 no 7 pp 1873ndash1883 2014
[54] Y-B Zhou Z-X Huang C-P Ren B Zhu and K-T Yaoldquo[Screening and preliminary analysis of the apoptosis-andproliferation-related genes in nasopharyngeal carcinoma]rdquoNanfang yi ke da xue xue bao vol 29 no 4 pp 645ndash647 2009
[55] Y Zhan R Hegde S M Srinivasula T Fernandes-Alnemriand E S Alnemri ldquoDeath effector domain-containing proteinsDEDD and FLAME-3 form nuclear complexes with the TFI-IIC102 subunit of human transcription factor IIICrdquo Cell Deathand Differentiation vol 9 no 4 pp 439ndash447 2002
[56] P J Byrd T Stankovic C M McConville A D Smith P RCooper and A M R Taylor ldquoIdentification and analysis ofexpression of human VACM-1 a cullin gene family memberlocated on chromosome 11q22-23rdquo Genome Research vol 7 no1 pp 71ndash75 1997
[57] S L Carter M Negrini R Baffa et al ldquoLoss of heterozygosityat 11q22-q23 in breast cancerrdquo Cancer Research vol 54 no 23pp 6270ndash6274 1994
[58] R S Samant P A Clarke and PWorkman ldquoE3 ubiquitin ligaseCullin-5 modulates multiple molecular and cellular responsesto heat shock protein 90 inhibition in human cancer cellsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 111 no 18 pp 6834ndash6839 2014
[59] A Van Ophoven C P Ng B Patel B Bonavida and ABelldegrun ldquoTumor necrosis factor-related apoptosis-inducingligand (TRAIL) for treatment of prostate cancer first results andreview of the literaturerdquo Prostate Cancer and Prostatic Diseasesvol 2 no 5-6 pp 227ndash233 1999
[60] V Modur R Nagarajan B M Evers and J Milbrandt ldquoFOXOproteins regulate tumor necrosis factor-related apoptosis induc-ing ligand expression Implications for PTEN mutation inprostate cancerrdquo The Journal of Biological Chemistry vol 277no 49 pp 47928ndash47937 2002
[61] R Nishikawa Y Goto S Kojima et al ldquoTumor-suppressivemicroRNA-29s inhibit cancer cell migration and invasion viatargeting LAMC1 in prostate cancerrdquo International Journal ofOncology vol 45 no 1 pp 401ndash410 2014
[62] S Giusiano S Garcia C Andrieu et al ldquoTP53INP1 overexpres-sion in prostate cancer correlates with poor prognostic factorsand is predictive of biological cancer relapserdquo Prostate vol 72no 2 pp 117ndash128 2012
[63] W W Du L Fang M Li et al ldquoMicrorna mir-24 enhancestumor invasion and metastasis by targeting PTPN9 and PTPRFto promote EGF signalingrdquo Journal of Cell Science vol 126 no6 pp 1440ndash1453 2013
[64] G Zhao L Liu T Zhao et al ldquoUpregulation of miR-24promotes cell proliferation by targeting NAIF1 in non-small celllung cancerrdquo Tumor Biology vol 36 no 5 pp 3693ndash3701 2015
[65] X Zheng J Li C Peng et al ldquoMicroRNA-24 induces cisplatinresistance by targeting PTEN in human tongue squamous cellcarcinomardquo Oral Oncology vol 51 no 11 pp 998ndash1003 2015
[66] Y-Q Li J-H Lu X-M Bao X-F Wang J-H Wu andW-Q Hong ldquoMiR-24 functions as a tumor suppressor innasopharyngeal carcinoma through targeting FSCN1rdquo Journalof Experimental andClinical Cancer Research vol 34 article 1302015