Page 1
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata i
ÍNDICE
Lista de abreviaturas iii
Lista de figuras Lista de tabelas Resumo Abstract
iv v vi vii
1- Introdução
1
1.1 - Proteinases cisteínicas
1
1.2 - Inibidores de proteinas em plantas
2
1.3 - Cistatinas: Inibidores de proteinases cisteínicas 1.4 – Vigna vexillata
3 7
2 – Objetivos 3 - Materiais e Métodos
10 10
3.1 - Material Biológico
10
3.1.2- Bactéria Escherichia coli cepa M15 (pREP4) recombinante 3.1.3 - Bactéria Escherichia coli cepa M15 (pREP4) recombinante
10 10
3.2 - Expressão de uma CCPI recombinante em E. coli M15 (pREp4)
10
3.3 - Purificação de uma CCPI recombinante por cromatografia em Ni-NTA agarose
11
3.4- Extração de proteínas extracelulares em sementes de Vigna vexillata
12
3.5-Dosagem de proteínas
12
3.6- Obtenção dos fragmentos de membranas plasmáticas e paredes celulares de cotilédones de V.vexillata
12
3.7 - Visualização eletroforética de proteínas
15
3.8 - Western Blotting
15
3.9 - Ensaio de ELISA para investigação das interações entre cistatina de V. 16
Page 2
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata ii
unguiculata fragmentos de parede e membranas celulares de cotilédones de V. vexillata e insulina bovina 3.10- Purificação de uma cistatina a partir de extrato bruto de sementes de V. vexillata por meio de cromatografia de afinidade
17
3.11 - Ensaio de inibição de crescimento do fungo Fusarium solani utilizando a cistatina purificada de V. vexillata 3.12 - Localização tecidual de cistatinas de V. vexillata por imunohistoquímica
18 19
4 - Resultados
19
5 - Discussão 36 6 - Conclusões
41
7 - Referências Bibliográficas
42
Page 3
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata iii
LISTA DE ABREVIATURAS
• Amp – Ampicilina • BSA – Albumina Sérica Bovina
• CCPI – “Cowpea Cysteine Proteinase Inhibitor”
• DAB - Diaminobenzidina
• DO – Densidade ótica
• ELISA – Ensaio imunológico de ligação a enzima
• GA - Glutaraldeído
• IgG – Imunoglobulina G
• IPTG – Isopropil β D-galactosídeo
• LB – Meio Luria-Bertani
• Ni-NTA – Níquel ácido nitrilotriacético
• PA - Paraformaldeído
• PAGE – Eletroforese em gel de poliacrilamida
• PBS – Tampão fosfato salino
• PMSF – Fenilmetil-sulfonil fluoreto
• mRNA – RNA mensageiro
• SDS – Sódio dodecil sulfato
• TCA – Ácido tricloroacético
• TEMED – N,N,N’,N’- Tetrametiletilenodiamino
• Tris – Tris (hidroximetil) aminometano
• Tween20 – Polioxietileno (20) sorbitano monolaurato
Page 4
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata iv
LISTA DE FIGURAS
Figura 1 - Fotografia de exemplar típico da espécie Vigna vexillata (L.) A. Rich, com
destaque para suas folhas, flor e vagem.
9
Figura 2 – Flor da espécie Vigna vexillata (L.) A. Rich.
9
Figura 3- Preparação dos fragmentos de parede celular e membranas celulares. Metodologia de Okorokov e Iehle (1998).
14
Figura 4 – SDS-PAGE (A) e Western-blotting (B) da fração protéica do extrato
bacteriano, retida em cromatografia de afinidade em coluna de Níquel NTA-Agarose.
M - Marcadores de massa molecular; FPR - Fração protéica retida.
22
Figura 5 – SDS PAGE do perfil protéico das frações obtidas no experimento de fluido apoplástico.
25
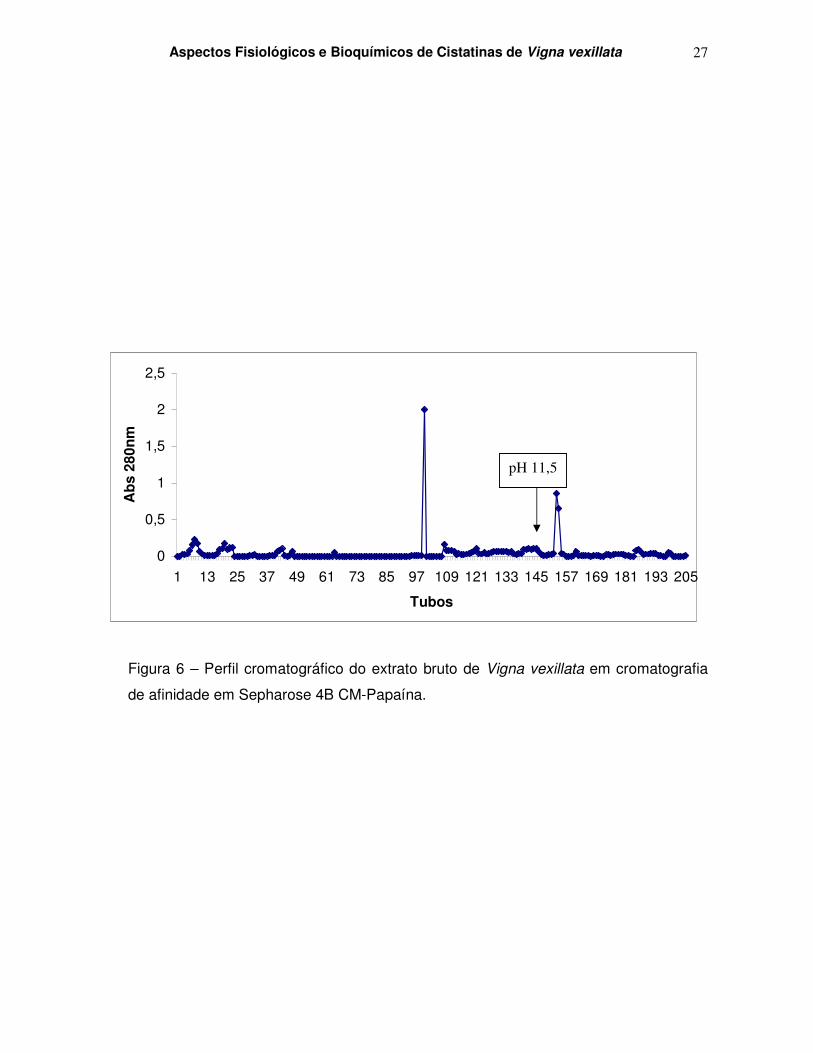
Figura 6 – Perfil cromatográfico do extrato bruto de Vigna vexillata em Sepharose 4B CM-Papaína.
27
Figura 7 - SDS-PAGE (A) e Western-blotting (B) da fração retida na coluna de afinidade Sepharose 4B CM-Papaína.
29
Figura 8 – Ensaio da atividade antifúngica da cistatina purificada de Vigna vexillata
(200 µg de proteína) contra o fungo F. solani .
31
Figura 8 – Ensaio da atividade antifúngica da cistatina purificada de Vigna vexillata
(200 µg de proteína) contra o fungo F. solani .
31
Figura 9 – Fotomicrografia da análise em microscópio ótico do fungo Fusarium solani
crescido em meio de cultura controle (A) e acrescido de cistatina purificada de Vigna
vexillata (200 µg de proteína) (B)
Figuras 10 e 11 – Análise imunohistoquímica de cistatinas de sementes de V.vexillata
32
Page 5
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata v
LISTA DE TABELAS
Tabela 1 – Classificação da espécie Vigna vexillata (L.) A. Rich.
8
Tabela 2- Interações entre a cistatina recombinante, fragmentos de membranas e
paredes celulares de cotilédones de V. unguiculata e insulina bovina por ELISA.
23
Tabela 3- Interações entre a cistatina recombinante, fragmentos de membranas e
paredes celulares de cotilédones de V. vexillata e insulina bovina por ELISA.
24
Tabela 4- Ensaio de atividade inibitória contra papaína nas amostras provenientes da
comatografia de afinidade em Sepharose 4B CM Papaína.
28
Page 6
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata vi
Resumo
Fitocistatinas têm sido relacionadas a diversos papéis fisiológicos, como
controle de proteólise endógena e defesa contra proteases exógenas de predadores de
plantas. A sugestão de uma característica singular para elas tem sido proposta
envolvendo sua participação em processos de transdução celular de sinais e interação
intertecidual. Nós isolamos uma cistatina de Vigna vexiilata usando uma coluna de
afinidade Seprharose 4B CM-Papaína. A extração ocorreu usando Tris-HCl 0.1M, NaCl
0.5M pH 7.2 (300g de farinha/L), 16h. A suspensão foi centrifugada (5,000xg, 30 minutos
/4ºC) e o sobrenadante (10 mL) foi passado pela coluna Sepharose 4B CM-Papaína. O
tampão de equilíbrio foi fosfato de sódio 0,1M, NaCl 0,5M pH 7,0 e as frações retidas
foram eluídas com o mesmo tampão em pH 11.5. A proteína isolada apresentou um peso
molecular de 13 kDa e reagiu com o anticorpo anti-cistatina através de Western blotting.
Ensaios em microplaca de atividade antifúngica demonstraram que a proteína isolada
completamente inibiu o crescimento de F. solani em uma concentração de 200µg.
Page 7
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata vii
Abstract
Phytocystatins have been correlated to several physiological roles, as control of
endogenous proteolysis and defense against exogenous proteases from plant predators. A
suggestion of a singular attribute for them has been proposed involving its participation in
cellular signal transduction and intertissue interaction processes. We isolated a V. vexillata
cystatin by using an affinity Sepharose 4B CM-Papain column. The extraction was
performed using Tris-HCl 0.1M, NaCl 0.5M pH 7.2 (300g of flour/L), overnight. The
suspension was centrifuged (5,000xg, 30 minutes /4ºC) and supernatant (10 mL) was
percolated through the Sepharose 4B CM-Papain column. The equilibration buffer was
sodium phosphate 0,1M, NaCl 0,5M pH 7,0 and retained fractions were eluted by the
same buffer at pH 11.5. The isolated protein had a Mr of 13 kDa and cross-reacted with an
anti-cowpea cystatin antibody by Western blotting. Antifungal activity microplate assays
showed that the isolated protein completely inhibited the growth of F. solani at a
concentration of 200µg. As the V. vexillata cystatin has previously been shown by us to be
located at extracellular space of seed cotyledons, a defense role against predators is
suggested. However, binding of this cystatin to cell wall and cell membrane components
are being tested by ELISA, to investigate its potential participation in the above mentioned
role of involvement in signal transduction and intertissue interaction processes.
Page 8
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 1
1- INTRODUÇÃO
1.1 - Proteinases cisteínicas
O termo protease é usado para se referir tanto a endopeptidases como a
exopeptidades. O termo proteinase é usualmente empregado para se referir a
endopeptidases. Estas se classificam de acordo com o aminoácido envolvido em seu sítio
catalítico e com seu mecanismo de ação em: serínicas, cisteínicas, aspárticas ou
metaloproteinases (Barrett, 1986).
As proteinases cisteínicas são amplamente distribuídas em animais, plantas e
microorganismos, onde participam de variados processos intra e extracelulares de
importância fisiológica. Estas podem ser induzidas em resposta a diferentes fatores
ambientais como calor e frio (Schaffer e Fisher, 1990), seca (Koizumi et al., 1993).
As proteinases cisteínicas não são secretadas como enzimas digestivas em
animais superiores, mas são encontradas nos intestinos médios de algumas espécies das
famílias Hemipterae (Houseman, 1978; Houseman et al., 1980) e Coleopterae (Murdock
et al., 1987). Alguns membros da família Coleopterae são importantes pestes de
sementes e folhas (Murdock et al., 1987); estes utilizam principalmente proteinases
cisteínicas para a sua digestão protéica, não necessitando de proteinases serínicas para
tal propósito. Desta maneira a larva de Callosobruchus maculatus (inseto predador de
sementes de feijão-de-corda), por exemplo, consegue ingerir tecidos com níveis
relativamente altos de inibidores de proteinases serínicas sem sofrer maiores efeitos
(Xavier Filho e Campos, 1984).
Dentre as proteinases de órgãos de reserva em plantas que estão envolvidas
em processos bioquímicos durante a germinação, as principais são as proteinases
cisteínicas (Ryan e Walker-Simmons, 1981). Estas enzimas foram primeiramente
estudadas em sementes germinantes de leguminosas (Tully e Beevers, 1978).
As proteinases cisteínicas estão envolvidas na degradação de proteínas de
reserva durante a germinação de sementes como mostram estudos pioneiros sobre a
degradação protéica de vicilinas por uma peptideohidrolase cisteínica em sementes
germinantes de Vigna radiata (Baumgartner e Chrispeels,1976).
A ação das proteinases cisteínicas na hidrólise de proteínas de reserva
também é relatada em trabalhos sobre a germinação de sementes de feijão comum
Page 9
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 2
(Phaseolus vulgaris) (Csoma e Polgar, 1984) e sobre a ação de duas endopeptidases
cisteínicas na degradação protéica em cotilédones de sementes de Vigna mungo
(Mitsuhashi et al., 1986).
Durante o desenvolvimento de sementes, as proteinases cisteínicas catalizam
os processos pós-transcricionais da maioria dos precursores protéicos, como é mostrado
no trabalho de Abe et al. (1993), onde é estudada uma asparaginil endopeptidase de
sementes de Canavalia ensiformis. Estudos que revelam uma expressão coordenada de
proteinases cisteínicas e inibidores de papaína durante o desenvolvimento de sementes
de feijão-de-corda foram realizados por Fernandes et al. (1991).
As proteinases cisteínicas também estão envolvidas na degradação protéica
durante os processos de senescência, como mostra o trabalho no qual é caracterizada e
purificada uma tiol protease induzida durante a senescência em ovários não polinizados
de Pisum sativum (Cercos e Carbonell,1993).
1.2 - Inibidores de proteinases em plantas
Existem, em plantas, algumas famílias de inibidores de proteinases já
descritas, dentre as quais temos a família de inibidores de tripsina de soja do tipo Kunitz,
família de inibidores de batata I, família de inibidores de batata II, família de inibidores de
proteinases aspárticas e a família de inibidores de proteinases cisteínicas (cistatinas), por
exemplo (De Leo, 2002).
Devido ao fato de que inibidores de proteinases inibem enzimas digestivas de
animais, estes foram e são objetos de inúmeros estudos.
A importância de alguns inibidores de proteinases no controle de pragas fez
com que muitos esforços fossem dirigidos a estudar a expressão, em plantas, de
diferentes genes de inibidores de proteinases. Assim, plantas transgênicas de tabaco
foram utilizadas para avaliar os efeitos inseticidas de vários genes (Hilder et al., 1987)
dentre eles: os de inibidores de proteinases I e II de plantas de tabaco (Johnson et al.,
1989); o gene que codifica para inibidor tríptico de feijão-de-corda (CpTI) (Boulter et al.,
1989); o gene de um inibidor de proteinase cisteínica de sementes de arroz (Masoud et
al., 1993); e o gene que codifica o inibidor bifuncional de proteinases serínicas e de α-
amilases de insetos de milho (Masoud et al., 1993), dentre outros. Outras plantas
transgências, tais como batata, algodão, alfafa, soja, Arabidopsis, batata-doce, morango,
repolho, arroz, apresentaram melhores “performances” em bioensaios utilizando uma
Page 10
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 3
diversa gama de insetos predadores, que plantas controle (não transformadas)
(Mochizuki et al., 1999; Hilder e Boulter, 1999; Lara et al., 2000; Vila et al., 2005; Bu et al.,
2006; Álvarez-Alfageme et al., 2007).
1.3 - Cistatinas: inibidores de proteinases cisteínicas
O termo cistatina foi atribuído pela primeira vez por Barrett et al (1986) a uma
proteína de clara de ovo de galinha, que tinha propriedade de inibir proteinases
cisteínicas.
As cistatinas são, portanto, inibidores competitivos e reversíveis de
proteinases cisteínicas, compondo uma superfamília de proteínas evolutivamente
relacionadas. Esta superfamília, denominada superfamília das cistatinas, está dividida em
três famílias (Barret et al., 1987): a das estefinas, a família das cistatinas e a família dos
cininogênios.
As cistatinas de plantas estão classificadas em grupo à parte, tido com o
grupo das fitocistatinas. Estas proteínas não possuem pontes de enxofre e contêm uma
seqüência conservada L-A-R-[FY]-A-[VI]-X3-N na região N-terminal (Margis et al., 1998).
As evidências que sugerem que as cistatinas supracitadas constituem uma
nova família diferente das estefinas, cistatinas e cininogênios baseiam-se no fato de que
muitas destas possuem características de duas famílias diferentes como estefinas e
cistatinas ,simultaneamente, como citado por Flores (1997) .
Outra característica marcante, diferente, encontrada em algumas cistatinas de
origem vegetal, como a orizacistatina II (Kondo et al., 1991) e a cistatina de milho (Abe et
al., 1992), é a organização genômica evidentemente diferente das cistatinas animais
(Margis et al., 1998 ; Kondo et al., 1991; Abe et al., 1992).
As fitocistatinas têm sido relacionadas com a regulação de proteinases
cisteínicas endógenas envolvidas na mobilização de proteínas de reserva durante a
germinação de sementes ou em outros processos fisiológicos de tecidos vegetais (Abe et
al., 1991; Watanabe et al., 1991; Fernandes et al., 1991; Lim et al., 1996; Ojima et al.,
1997).
Estudos sobre a presença de cistatinas em plantas iniciaram-se apenas na
década de 80. Exemplos de tais registros foram as descobertas das cistatinas I e II de
arroz, conhecidas como orizacistatinas (Abe et al., 1987;1989; Kondo et al., 1991), das
Page 11
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 4
cistatinas I e II de milho (Abe e Whitaker, 1988; Abe e Arai, 1991; Abe et al., , 1996; Irie et
al., 1996), cistatina de Wisteria floribunda (Hirashiki et al., 1990), cistatina de tubérculos
de batata (Rowan et al., 1990; Waldron et al., 1993; Orr et al., 1994; Walsh e Strickland,
1993), cistatina de sementes de feijão–de-corda (Fernandes et al., 1993), cistatinas de
frutos de abacate (Kimura et al., 1995), cistatinas de botões florais de Brassica campetris
(Lim et al., 1996), cistatina de sementes de soja (Hines et al., 1991; Misaka et al., 1996),
cistatinas de sementes de mamão (Song et al., 1995), cistatinas de sementes de
Phaseolus vulgaris (Santino et al., 1998; Brzin et al., 1998), cistatinas de Chelidonium
majus ( Rogelj et al., 1998), cistatinas de sementes de girassóis (Kouzuma et al., 1996,
2000 e 2001) cistatinas de sementes de trigo (Corre-Menguy et al., 2002), cistatinas de
sementes de sésamo (Shyu et al., 2004a), cistatinas de frutos de abacaxi (Shyu et al.,
2004b), cistatina de sementes de cevada (Martinez et al., 2005a), cistatina do tubérculo
Colocasia esculenta (Yang et al., 2005), cistatina de morango (Martinez et al., 2005b),
cistatinas de cana-de-açúcar (Gianotti et al., 2006), cistatina de Celosia cristata
(Gholizadeh et al., 2005), cistatinas de sementes de Phaseolus mungo (Sharma et al.,
2006).
O papel de defesa vegetal destas proteínas contra diversos predadores tem
sido demonstrado por testes de atividades biológicas de cistatinas contra insetos, como
os trabalhos de Hines et al. (1991), Chen et al. (1992), Michaud et al. (1994), Irie et al.
(1996), Kuroda et al. (1996) e Lim et al. (1996), contra nematódios (Urwin et al., 1995,
1997), moluscos (Walker et al., 1999) e fungos (Pernas et al., 1999; Siqueira-Júnior et al.,
2002., Martinez et al, 2005., Christoya et al., 2006).
Paralelamente, também foi demonstrado que em sementes de feijão-de-corda
não há relação entre os níveis de cistatinas encontrados e a resistência destas ao
bruquídeo peste, Callosobruchus maculatus (Xavier-Filho et al.,1989; Fernandes et al.,
1993) e um papel destas fitocistatinas na regulação de proteinases cisteínicas endógenas
durante a maturação das sementes foi ainda sugerido (Fernandes et al., 1991).
No caso específico de feijão-de-corda foram observados padrões complexos e
coordenados de atividades de proteinases cisteínicas e de cistatinas durante o
desenvolvimento de sementes (Fernandes et al., 1991), bem como variada localização em
células e tecidos (Flores et al., 2001). Neste último trabalho observa-se uma distribuição
uniforme de cistatinas em células de cotilédones e eixos embrionários de feijão-de-corda,
exceto por uma maior concentração em células epidérmicas da zona de abscisão entre os
cotilédones. De forma semelhante, Misaka et al. (1996) demonstraram que a cistatina de
Page 12
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 5
soja concentrava-se em uma camada periférica de células cotiledonárias. Entretanto, os
autores não atribuem nenhuma importância especial a tal localização, ao contrário dos
primeiros que sugerem que tal concentração elevada em uma camada celular periférica
seria compatível com um papel de defesa do tecido vegetal. A seqüência de aminoácidos
deduzida de uma cistatina de feijão-de-corda clonada por Fernandes et al. (1993) foi vista
como deficiente em um peptídeo sinal N-terminal, significando que esta isoforma não
deve ser transportada através de um sistema secretório.
Outros resultados demonstraram que, além de uma localização citoplasmática
principal, as cistatinas de sementes de Vigna vexillata também eram encontradas em
depósitos extracelulares nos cotilédones destas sementes e que, portanto, um processo
de translocação, neste caso, deveria ser admitido (Ávila et al., 1999).
O estudo de uma cistatina também extracelular, insolúvel, caracterizada por
Ojima et al. (1997), de tecidos cultivados in vitro de sementes de cenoura, sugeriu seu
envolvimento em processos de transdução de sinais celulares e de interação tecidual
entre células. Esta cistatina foi vista como capaz de interagir com glicoproteínas solúveis
de 57 kDa, que por sua vez, também possuíam a capacidade de ligarem-se a peptídeos
homólogos à insulina (Satoh ,1998).
Após a descoberta da insulina em animais, trabalhos realizados por Collip
(1923), apresentaram resultados que sugeriam a presença de substâncias similares à
insulina em extratos vegetais (folhas de cebola, cevada, alface, trigo e feijão verde; raízes
de cebola e cevada e caules de feijão verde). Nestes trabalhos, observou-se que todos os
extratos promoviam a redução dos níveis de glicose do sangue de coelhos normais e
cães pancreatectomizados. Os níveis de glicose no sangue de cães pancreatectomizados
também retornavam aos níveis normais após o tratamento com extratos de beterraba
(Best, 1924).
No trabalho realizado por Khanna et al. (1976) com sementes de Momordica
charantia, foi isolada uma proteína com massa molecular de aproximadamente 6 kDa que
reagia com anticorpos contra insulina humana. Trabalho realizado por Collier et al. (1987)
relatou o isolamento de proteínas de folhas de espinafre e plantas de Lemna gibba G3,
com massas moleculares semelhantes à insulina, que reagiam com anticorpos anti-
insulina suína e ligavam-se ao receptor de insulina humana. Mas nenhum desses
trabalhos determinou a estrutura primária dessas proteínas. Foi isolada, do tegumento de
sementes de C. ensiformis, uma proteína de massa molecular de 6 kDa que apresentou
100% de homologia com a insulina bovina. Essa mesma fração mostrou um fragmento
Page 13
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 6
polipeptídico de 11 aminoácidos, homólogo a um receptor do tipo proteína quinase
humana (Oliveira et al.,1999). Trabalhos realizados posteriormente pelo mesmo grupo
mostraram a presença de proteínas homólogas à insulina em vagens verdes do feijão de
corda Vigna unguiculata (Venâncio et al., 2003), e em folhas de Bauhinia variegata
(Azevedo et al., 2006). Outros resultados mostraram que antígenos do tipo insulina estão
presentes na torta de filtro, subproduto residual da clarificação de açúcar extraído da
cana-de-açúcar (Lopes, 2001), em folhas de Phaseolus vulgaris, nas gimnospermas
Cupressus sempervirens, Pinus ponderosa, Cycas revoluta, Eycadaceae zamia e
Selaginell sp, na parte aérea das pteridófitas Psilotaceae sp e Equisetoceae sp, na
levedura Saccharomyces cerevisiae, na alga Rodofta sp, no caldo de cana de Saccharum
officinarum, (Silva et al., 2002; Xavier-Filho et al., 2003).
Posteriormente, trabalhos realizados por Oliveira et al. (2004) mostraram que
proteínas imunorrelacionadas à insulina, ao receptor de insulina e a proteínas do tipo
fosfoserina estão localizados na camada interna do tegumento de sementes de C.
ensiformis.
Em 1989, Kagawa e Hirano mostraram a presença de proteínas do tipo
globulinas 7S, em soja, que eram capazes de se ligarem à insulina suína, sugerindo para
essas proteínas uma possível função de receptor para o hormônio insulina. Essa proteína
foi denominada Bg e consiste de duas cadeias polipeptídicas de 16 e 26 kDa, unidas
entre si por pontes dissulfetos. Trabalhos posteriores demonstraram que essas proteínas
apresentavam atividade de proteína quinase semelhante à atividade tirosina quinase,
típica do receptor de insulina animal.
Proteínas do tipo globulinas 7S básicas, imunorrelacionadas à Bg (globulina
7S básica de soja), já foram detectadas em uma grande variedade de sementes, dentre
elas, Vigna radiata, Vigna angularis e Lupinus alobus (Komatsu e Hirano, 1991).
Satoh, em 1998, encontrou em sementes de cenoura uma proteína de 57 kDa,
a qual denominou de EDGP. Tal proteína apresentou homologia de seqüência de
aminoácidos com a Bg e também era capaz de se ligar à insulina e também foi capaz de
ligar-se a cistatina extracelular de sementes de cenoura. Posteriormente, foi isolada uma
proteína de radículas de soja que era capaz de estimular a fosforilação da Bg in vitro, a
qual foi denominada leginsulina (Ilgoutz et al., 1997).
Existe uma complexidade de papéis fisiológicos pensados hoje para as
fitocistatinas, dentre esses, há indícios de que as fitocistatinas estariam participando de
processos de sinalização celular e transdução de sinais. Esta função é particularmente de
Page 14
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 7
interesse em nosso grupo de estudo, já que recentemente isolou-se e caracterizou-se
pela primeira vez, na literatura, uma insulina de origem vegetal, em tegumentos de
sementes de Canavalia ensiformis, em nosso laboratório (Oliveira et al., 1999). Além
disso, como relatado anteriormente, também foi detectada a presença de cistatina
extracelular em Vigna vexillata (Ávila et al., 1999), sendo este o segundo registro de tal
localização para fitocistatinas, de nosso conhecimento.
Além disso, já existem evidências da ligação de uma cistatina recombinante
de Vigna unguiculata com porções de paredes e membranas celulares de células
cotiledonárias de Vigna vexillata e também existem indícios da presença de uma cistatina
endógena em frações de parede celular de Vigna vexillata (Da Cunha, 2004), o que é de
grande interesse, uma vez que não se conhece na literatura nenhuma cistatina localizada
em paredes celulares de sementes.
1.4 - Vigna vexillata
Vigna vexillata é uma das mais importantes espécies pertencentes ao
subgênero Plectotropis. Este subgênero é um dos sete reconhecidos por Maréchal no
gênero Vigna (Maréchal et al., 1978). O subgênero Plectotropis ocupa uma posição
intermediária entre os subgêneros Vigna e Ceratrotopis, os quais englobam espécies
endêmicas da África e Ásia, respectivamente (Maréchal et al., 1978). A classificação da
espécie Vigna vexillata está descrita na Tabela 1.
Espécies não cultivadas do gênero Vigna despertam interesse de
pesquisadores envolvidos com pesquisas visando diferentes objetivos, tais como a
utilização destas espécies como fontes de genes para a manipulação genética de feijões
cultivados, como, por exemplo, o feijão-de-corda (Vigna unguiculata (L.) Walp.) e o feijão
mungo (Vigna radiata (L.) Wilczeck) (Rao e Kammannanauar, 1990; James e Lawan,
1991; Oghiakhe et al., 1993; Thottappilly et al., 1994). Tais projetos de manipulação
teriam como principais metas a melhoria da capacidade defensiva contra pestes e pragas,
e a elevação do valor nutritivo de suas sementes e folhagens (Siddhuraju et al., 1994;
Vanderborght, 1989). Além disso, muitas destas espécies apresentam propriedades
medicinais (Kokwaro, 1976) e são também utilizadas como forragens em cultivos
(Vanderborght, 1989).
A espécie Vigna vexillata é caracterizada por enormes variações morfológicas
principalmente no que diz respeito à forma dos folíolos. Esta variabilidade resultou em
Page 15
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 8
múltiplas nomenclaturas no passado, as quais foram drasticamente reduzidas por
Verdcourt (1971) e depois por Marechal et al. (1978), que identificaram várias variedades
botânicas como sendo Vigna vexillata.
A espécie Vigna vexillata não é cultivada para propósito de consumo e não
passou por nenhum processo de cultivo agrícola (Marechal et al., 1978).
As Figuras 1 e 2 mostram as características físicas da espécie Vigna vexillata
(L.) A. Rich.
Tabela 1 – Classificação da espécie Vigna vexillata (L.) A. Rich.
Classificação: Vigna vexillata (L.) A. Rich.
Reino Plantae – Plantas Sub-reino Tracheobionta – Plantas Vasculares
Super-divisão Spermatophyta – Plantas com sementes Divisão Magnoliophyta – Plantas com flores
Classe Magnoliopsida – Dicotiledôneas Subclasse Rosidae
Ordem Fabales Família Fabaceae
Subfamília Faboideae Gênero Vigna
Espécie Nome vulgar
Vigna vexillata (L.) A. Rich. Feijão-miúdo, violeta do campo (“zombi pea”)
Adaptado de Albuquerque, V.M. (1987).
Page 16
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 9
Figura 1 - Fotografia de exemplar típico da espécie Vigna vexillata (L.) A.
Rich, com destaque para suas folhas, flor e vagem.
Fonte: http://www.nfc.co.kr/nat/flora/flora&04/ros/legu/legu&23/
Figura 2 – Flor da espécie Vigna vexillata (L.) A. Rich.
Fonte: http://www.nfc.co.kr/nat/flora/flora&04/ros/legu/legu&23/
2 – OBJETIVOS
Page 17
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 10
2.1- Purificar e caracterizar uma cistatina extracelular de V.vexillata.
2.2– Determinar o potencial fungicida da cistatina purificada contra fungos
fitopatogênicos de relevância agronômica.
2.3 - Avaliar as interações entre a cistatina recombinante de V.unguiculata com
preparações de paredes e membranas celulares de células cotiledonárias das
mesmas sementes.
2.4 - Imunolocalização de cistatina de Vigna vexillata.
3- MATERIAIS E MÉTODOS
3.1- Materiais Biológicos
3.1.2- Material Botânico
A espécie Vigna vexillata não é cultivada para propósito de consumo e não passou
por nenhum processo de cultivo agrícola (Marechal et al., 1978).
Para o presente trabalho os frutos já maduros de Vigna vexillata são constantemente
coletados nas dependências do campus da UENF, onde tal espécie cresce, como erva
daninha, em determinadas épocas do ano.
3.1.3 - Bactéria Escherichia coli cepa M15 (pREP4) recombinante
Os clones bacterianos de E.coli (cepa M15[pREP4]) transformados com o plasmídeo
pQ30 contendo um cDNA codificante de uma cistatina de feijão-de-corda (CCPI)
(Fernandes et al., 1993) foram cedidos pelo Dr. Peter Urwin, da Universidade de Leeds –
UK.
3.2 - Expressão de uma CCPI recombinante em E. coli M15 (pREp4)
O gene codificador de uma cistatina de sementes de V. unguiculata foi clonado
(Fernandes et al., 1993) e expresso em células de E. coli (Urwin et al., 1997).
A proteína recombinante foi obtida pela transferência, segundo metodologia descrita
por Flores (1997), de uma alíquota da bactéria crescida em meio sólido onde foram
mantidas colônias bacterianas transformadas com o gene codificador de cistatina, para 3
mL de meio LB (10g de bactotriptona; 10g de NaCl; 5g de extrato de levedura; pH 7,0). A
este foram previamente adicionados 7µL de ampicilina (retirados de uma solução estoque
Page 18
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 11
de concentração de 50 mg de ampicilina por mL de água) e 15µL de kanamicina (retirados
de uma solução estoque de 100 mg de kanamicina por mL de água) . A suspensão foi
mantida em estufa incubadora a 37oC, por 16 h e o crescimento foi avaliado por leitura a
600nm em espectrofotômetro (OD = 0,6).
O próximo passo foi preparar as placas de Petri para efetivar o crescimento das
colônias em meio sólido. Foram utilizadas duas placas autoclavadas, 30mL de meio LB,
7µL de ampicilina (50mg/mL), 12,87µL de kanamicina (100mg/mL) e 0,45g de bacto-agar.
15mL dessa preparação foram distribuídos em cada uma das placas que foram então
vedadas e deixadas em estufa a 37oC, por 16 h.
As bactérias foram transferidas para o meio sólido e deixadas em estufa a 37ºC, por
16h. Após o crescimento das colônias nas placas contendo o meio sólido, foi realizada
mais uma transferência para o meio líquido, desta vez para multiplicação das células
bacterianas.
As bactérias nessa cultura foram então induzidas a expressar a proteína
recombinante, pela adição de 0,5 mM de IPTG. Novamente houve uma incubação sob
agitação por 3h, até o alcance de uma OD600 = 0,8. A suspensão foi centrifugada a 3000 x
g, por 15 min, tendo-se descartado o sobrenadante e ressuspendido o precipitado em 20
mL de Tris-HCl 10 mM, fosfato de sódio dibásico 0,1M, uréia 8M, pH 8,0.
A cultura foi então sonicada em intensidade máxima (30 burst) por três vezes, por 1
min cada, dando um espaço de 1 min de intervalo entre cada sonicação. As células
lisadas foram sedimentadas por centrifugação a 12000 x g por 5 min e o extrato contendo
os produtos induzidos foi congelado até ser utilizado.
A metodologia descrita baseou-se na utilização por Flores et al. (2002).
3.3 - Purificação da CCPI recombinante por cromatografia em Ni-NTA agarose
A cromatografia foi realizada em tubos de microcentrufugação em sistema de
“batching” utilizando 500 µL de matriz em cada tubo. A matriz foi equilibrada com 1 mL de
tampão imidazol 10mM sob agitação leve, em temperatura ambiente, durante 5 minutos.
Após este tempo a solução foi centrifugada a 7000 x g por 30 s. Estas etapas de equilíbrio
e centrifugação foram repetidas por mais duas vezes.
Em seguida adicionou-se 780 µL do extrato bacteriano em cada tubo, os quais
foram mantidos, sob leve agitação, durante 45 min. Após este tempo as suspensões
Page 19
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 12
foram centrifugadas a 7000 x g por 30 s. O sobrenadante foi reservado e denominado não
retido. Após estas etapas foram realizadas 4 lavagens com 1 mL de tampão imidazol 20
mM, sob agitação, por 10 min, e as suspensões foram centrifugadas a 7000 x g por 30 s.
As lavagens foram reunidas e denominadas de fração de lavagem.
Outras 5 eluições foram realizadas com 200 µL de tampão imidazol 250 mM sob
agitação por 5 min. Depois de centrifugação por 30 s, a 7000 x g, os sobrenadantes foram
reunidos e denominados de fração retida.
3.4- Obtenção do fluido apoplástico em sementes de Vigna vexillata
As proteínas exudadas e/ou extracelulares de sementes de Vigna vexillata foram
extraídas segundo a metodologia descritas por Wälti et al (1995). As sementes foram
embebidas em tampão de extração de fluido apoplástico (0,14 M de carbonato de sódio,
NaCl 0,5 M, 0,33% Tween 20 e 0,2% β-mercaptoetanol, pH 5,5) por 1h e após este tempo
foram submetidas a um tratamento de baixa pressão, através de uma bomba à vácuo.
Elas, então, foram transferidas cuidadosamente para seringas e estas para tubos de
microcentrífuga e, então, foram submetidas à centrifugação (700 x g, 5 min, 4ºC). O fluido
apoplástico foi então coletado no fundo do tubo de microcentrífuga.
3.5-Dosagem de proteínas
As determinações das quantidades totais de proteínas foram feitas pelo
método descrito por Bradford (1976), usando-se albumina sérica bovina como proteína de
referência.
3.6- Obtenção dos fragmentos de membranas plasmáticas e paredes celulares de
sementes de Vigna unguiculata e vexillata.
De acordo com a metodologia de Okorokov e Iehle (1998) as sementes foram
embebidas em água para a preparação do homogenato que sofreu, então, um processo
de centrifugação e conseqüentemente de filtração para a obtenção do primeiro sedimento
que foi continha porções de paredes celulares. O sobrenadante desta primeira
centrifugação foi novamente centrifugado e o sedimento proveniente foi considerado
como porções de membranas celulares maiores. Realizou-se então mais um processo de
centrifugação e este último sedimento foi considerado com porções de membranas
menores. O sobrenadante foi então descartado.
Page 20
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 13
Os procedimentos empregados para a obtenção de fragmentos de
membranas plasmáticas e paredes celulares estão mostrados esquematicamente na
figura 3.
Page 21
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 14
Figura 3- Preparação dos fragmentos de parede celular e membranas celulares.
Metodologia de Okorokov e Iehle (1998).
Sobrenadante
Sedimento (Porções de membranas celulares menores)
Sedimento (Porções de membranas celulares maiores)
3g de sementes sem os tegumentos
Preparação, em água, do homogenato
1/3 (β−β−β−β−mercaptoetanol 25mM e PMSF 2mM) 15min. 4ºC.
F i l tr aç ã o do h omo g e na t o e m 9 c a ma d as de g a ze e m u m f u n i l p lá s t i c o .
Centrifugação a 2470 x g / 15 min.
S e di me nt o ( Po rç õe s d e par e de ce l u l ar )
Sobrenadante – Centrifugação a 11000 x g/ 45min.
S o b r e na d a n te – C e n t r i f ug a çã o a
2 5 00 xg / 30 mi n
Page 22
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 15
3.7- Visualização eletroforética de proteínas
Amostras da fração protéica do extrato bacteriano retida em cromatografia de
afinidade, das frações obtidas do experimento do fluido apoplástico e da fração retida na
coluna de afinidade Sepharose 4B CM-Papaína; todas em uma concentração de 20 µg de
proteína por poço, foram visualizadas por eletroforese em gel de poliacrilamida (12%) na
presença de SDS, segundo método descrito por Laemmli (1970). A eletroforese foi
realizada em um sistema vertical Mini Protean II da BIORAD. A separação protéica
ocorreu sob uma corrente constante de 15 mA nos primeiros 30 minutos e 30 mA no
tempo restante.
O gel foi corado em solução corante de Azul Brilhante de Coomassie G a 2%
em água, metanol e ácido acético (6:3:1, v/v/v), por 6 horas e descorado em solução
descorante composta de água destilada, metanol e ácido acético na mesma relação de
proporcionalidade usada na preparação da solução corante.
3.8- Western blotting
As proteínas separadas por eletroforese em gel de poliacrilmida-SDS foram
transferidas para uma membrana de nitrocelulose, de acordo com Towbin et al. (1979). O
sistema utilizado foi o de transferência semi-seca Trans-blot Sigma.
A membrana e o gel foram equilibrados em tampão de transferência (Tris
25mM, glicina 192mM e metanol 20%) para equlíbrio durante 20 minutos. Em seguida
foram mergulhadas 9 folhas de papel filtro no mesmo tampão.
O sanduíche de transferência foi montado da seguinte maneira: três pedaços
de papel de filtro, seguidos da membrana de nitrocelulose, do gel e outros três pedaços
de papel de filtro umedecidos no tampão de transferência. Após 1h de transferência em
amperagem de 1mA/cm2 do gel, o que correspondeu a uma amperagem constante de
38mA, a membrana foi corada com Ponceau S (Sigma) para verificação da transferência.
Após confirmar-se que as proteínas tinham sido transferidas para a membrana
de nitrocelulose, esta foi submersa em solução bloqueadora (tampão fosfato de sódio 0,1
M, NaCl 0,5 M pH 7,2 [PBS], com 2% de leite em pó desnatado), por 2 horas.
Posteriormente foram feitas quatro lavagens em tampão PBS e a membrana foi colocada
em contato com solução bloqueadora contendo anticorpo contra cistatina de feijão-de-
Page 23
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 16
corda produzido em coelho e cedido pelo Dr. Robert Thornburg, da Iowa State University,
na diluição de 1:2000, e mantida nesta solução durante a noite (16 horas, 4ºC).
No dia seguinte a solução bloqueadora foi descartada e quatro lavagens por 5
min cada com PBS foram efetuadas. Nova solução bloqueadora, desta vez contendo
anticorpo anti-IgG de coelho conjugado com peroxidase na diluição de 1: 2000, foi
despejada sobre a membrana e deixada sob agitação leve, por duas horas.
Para a revelação da reação imunoquímica utilizou-se o método de revelação
por diaminobenzidina (DAB), onde foram utilizados 100 µL do tampão Tris-HCl 2mM pH
7,5, 5 mg do corante DAB, 0,3 mL de imidazol 0,1M, 4,9 mL de água destilada e 5 �L de
água oxigenada 30%. Esta solução (10 mL) foi vertida sobre a membrana e mantida por
aproximadamente 10 minutos. A reação foi parada com substituição da solução
reveladora por água destilada.
3.9- Ensaio de ELISA para investigação das interações entre cistatina recombinante
de V. unguiculata, fragmentos de paredes e membranas celulares de V. vexillata,
unguiculata e insulina bovina
Foram realizados ensaios de ELISA modificados para investigação de
possíveis ligações entre a cistatina de V. unguiculata e insulina bovina comercialmente
adquirida, antes e após a ligação da cistatina a fragmentos de paredes e membranas
celulares de V. vexillata e unguiculata.
Para o ensaio de ELISA propriamente dito, poços de micro placas (NUNC
Maxi-Sorb) foram sensibilizados com 20 µg de proteína /100 µL de tampão carbonato
bicarbonato 0,05M pH 9,6 dos diversos antígenos utilizados (cistatina; cistatina retida em
parede; cistatina retida em membrana e insulina bovina comercial).
A sensibilização procedeu-se por 16 horas a 4ºC. Após a sensibilização os
poços foram lavados com PBS (fosfato 0,1M/ NaCl 0,5M/ pH 7,6) contendo Tween 0,05%
(280 µL) por 60 min. Após este período foram adicionados 300 µL do tampão bloqueador
(gelatina 1% no mesmo tampão PBS/Tween citado acima) As reações foram mantidas por
60 min, a temperatura ambiente, e uma nova lavagem com PBS/Tween (280 µL). por 60
min, foi realizada. Após esta etapa, foram adicionados, quando previsto na estratégia do
ensaio experimental, os ligantes (20 µg de insulina bovina ou cistatina). A reação de
Page 24
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 17
ligação foi aguardada durante 2 h, a temperatura ambiente. Os poços foram então
lavados com PBS-Tween 0,05% (280 µL) por 2 h.
Em casos específicos, quando tratando-se de cistatina como antígeno, as
preparações de paredes e membranas celulares foram adicionadas após o período de
bloqueio com PBS-Tween-gelatina, quando previsto na estratégia experimental mostrada
na Tabela 3 (Poços 9 e 10). Nestes ensaios, especificamente, pretendeu-se investigar se
a ligação de insulina bovina dava-se ao nível de cistatinas isoladas ou associadas a tais
preparações de paredes e membranas.
Em seguida, adicionou-se os anticorpos anti-cistatina ou anti-insulina humana
diluídos (1:2000) em tampão bloqueador (50 µL) durante 2 h, a 37ºC. Depois de expostas
aos anticorpos, as placas foram lavadas 6 x com PBS/Tween (280 µl) a cada 5 min, e
incubadas com uma solução contendo o complexo anti-IgG de coelho + peroxidase
(quando o 1º anticorpo for anti-cistatina) ou anti-IgG de guinea pig + peroxidase (quando o
1º anticorpo for anti-insulina), diluído em tampão bloqueador (1:2000), a 37ºC, por 2 h.
Lavagens posteriores (6 x) com PBS/Tween (280 µL) foram novamente
realizadas a cada 5 min. A revelação foi feita usando-se 50 µL de uma solução de OPD
(orto fenil diamina) (5 mg de OPD/ 12,5 mL de tampão ácido [3,25 mL de ácido cítrico 0,1
M; 3,5 mL de fosfato de sódio 0,2 M; 5 µL de H2O2 30%; 5,75 mL de H2O pH 5,0]).
A revelação durou aproximadamente 10 minutos, em uma reação mantida em
ausência de luz. Esta reação foi parada com 50 µL de H2SO4 3 N e os produtos foram
avaliados por sua absorbância a 492 nm, lidos em um leitor de microplaca.
3.10- Purificação de uma cistatina a partir do extrato bruto de sementes de Vigna
vexillata por meio de cromatografia de afinidade.
Para a obtenção do material a ser aplicado na coluna, retirou-se o tegumento
das sementes de Vigna vexillata para a preparação da farinha. Foi realizada uma primeira
extração com metanol 80%, 1:5 (m/v), para a remoção da clorofila. Essa extração foi
repetida três vezes. Em seguida foi realizada uma segunda extração, na proporção de
300g de farinha para 1L de tampão de extração (Tris-HCl 0,1M, NaCl 0,5M pH 7,2), sob
agitação, em geladeira, por 16 horas. Após essa extração, esse material foi centrifugado
a 5.000xg por 30 minutos a 4ºC. O precipitado foi descartado e o sobrenadante fervido
durante 5 minutos. Fez-se uma nova centrifugação nas mesmas condições citadas acima.
Page 25
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 18
O precipitado foi descartado e o sobrenadante foi então liofilizado. Esse material foi
submetido à dosagem de proteínas e ensaio de atividade inibitória contra papaína.
A coluna de afinidade Sepharose 4B – CM-Papaína, previamente preparada e
disponível em nosso laboratório, foi equilibrada com tampão fosfato de sódio 0,1M, NaCl
0,5M pH 7,0. Foram aplicados na coluna 10 mL do material obtido como descrito acima. A
coluna foi então novamente lavada com tampão pH 7,0, para assim haver a liberação do
material que não ficou retido na coluna. Quando esse material parou de ser liberado, foi
aplicado o tampão de eluição da coluna (tampão fosfato de sódio 50 mM, NaCl 0,5 M pH
11,5), que liberou as proteínas retidas na matriz, mais especificamente cistatinas presas à
papaína carboximetilada imobilizada na Sepharose. A coleta foi realizada em sistema de
coletor automático com um volume coletado de 1 mL. Todo o material liberado da coluna
foi analisado em espectrofotômetro a 280 nm para assim medir as devidas absorbâncias,
bem como em sua atividade inibitória sobre papaína por ensaios in vitro.
3.11- Ensaio de inibição de crescimento do fungo F. solani utilizando a cistatina
purificada de Vigna vexillata.
3.11.1 - Obtenção de esporos filamentosos
O fungo F. solani foi crescido em uma placa de Petri contendo ágar
sabouraud por 10 dias a 30 ºC. Após esse período 10 mL de salina 0,15 M foram vertidos
sobre essa placa e os esporos foram liberados com o auxílio de uma alça de Drigalski.
Essa suspensão com os esporos foi devidamente filtrada em gases para a purificação de
restos miceliais que pudessem estar vindo junto com os esporos. Esses esporos foram
contados em câmara de Newbauer.
3.11.2 - Ensaio de inibição
Em placas de cultura de células (96 poços), contendo 200 µL de meio de
cultura específico para o crescimento do fungo utilizado, foi adicionada uma amostra
contendo a cistatina purificada de Vigna vexillata em uma concentração de 200 µg de
proteína/mL e 1 x 104 esporos/mL de F. solani. Para a observação da inibição do
crescimento dos fungos, foi determinada a densidade ótica calculadas a partir de leituras
em “um leitor de ELISA” a 670 nm a cada 6 horas, por um período de 60 horas. Todo o
procedimento, desde a obtenção dos esporos até a montagem do experimento na placa
foi feito sobre condições assépticas em capela de fluxo laminar.Controles foram feitos
sem a adição da cistatina.
Page 26
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 19
As amostras obtidas foram também gotejadas sobre lâminas microscópicas
para visualização em microscópio ótico e análise morfológica dos prováveis efeitos da
cistatina sobre o crescimento do fungo.
3.12 – Localização tecidual de cistatinas de V. vexillata por imunohistoquímica
As sementes de V. vexillata foram analisadas no que se refere à localização
tecidual de cistatinas. Para tal, as sementes foram embebidas em água destilada por 8h e
posteriormente fixadas por 3h em tampão cacodilato 0,01M pH 7,2 ; glutaraldeído 0,01%
e paraformaldeído 4%. Após a fixação, as sementes foram submetidas a três lavagens
(30 min cada) em tampão cacodilato 0,05M para posteriormente serem desidratadas
seqüencialmente com metanol a 50% (30min), 70%(60min), 90%(60min) e tratadas com
50% de metanol + 50% de resina LR Gold (18h), 30% de metanol + 70% de resina LR
Gold (16h), 100% de resina LR Gold (2 a 3 dias) e 100% resina + catalizador (10h). Todos
os passos foram feitos a -20°C. Cortes semifinos são preparados e incubados com PBS-
BSA (tampão fosfato-salino com albumina sérica bovina) por 40min, e então incubados
com anticorpo anti-cistatina (1:200) por 2h. Os cortes são então incubados em cloreto de
amônio por 30 min, incubados com PBS-BSA por 20 min, incubados com anticorpo
primário anticistatina por 2h (1:100), lavados 10 vezes com PBS-BSA por 10 min cada,
incubados com anticorpo secundário conjugado a ouro 5nm (1:300), repete-se a lavagem
com PBS-BSA por 10 min cada, lava-se com PBS (tampão fosfato salino) 10min cada
lavagem e lava-se com água milli Q por 10 min cada. As incubações que envolvem
anticorpos, são realizadas em câmara úmida (um recipiente contendo algodão
embebecido em água). Ao término das etapas de preparação, acrescenta-se um a gota
de 0,2 M de N-propilgalato e cobrem-se as lâminas com lâminulas. O material deve ser
guardado em geladeira (4ºC) até a visualização, após coloração por prata. Os resultados
foram observados por microscopia óptica.
4 – RESULTADOS 4.1 - Análise da expressão da cistatina recombinante de Vigna unguiculata em E.
coli, por SDS-PAGE
A Figura 4A mostra a eletroforese desnaturante em gel de poliacrilamida
demonstrando a purificação da cistatina recombinante de Vigna unguiculata por
Page 27
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 20
cromatografia de afinidade, em resina de Ni-NTA agarose. O poço 2, referente a amostra
proveniente do pico retido na resina apresenta a banda protéica de 13 kDa,
aproximadamente (seta).
A natureza da banda protéica apontada pela seta, como uma cistatina, foi
confirmada por experimento de Western blotting, após transferência de proteína do gel
para uma membrana de nitrocelulose. O reconhecimento imunológico positivo por um
anticorpo específico anti-cistatina (1:2000) é observado na Figura 4B. A banda referente a
cistatina recombinante, reconhecida pelo anticorpo, é apontada na região de massa
molecular próxima a 13 kDa (seta).
4.2 –Verificação das ligações estabelecidas entre cistatinae de Vigna unguiculata,
fragmentos de paredes e membranas celulares de V. vexillata, unguiculata e
insulina bovina
As Tabelas 2 e 3 apresentam os valores obtidos com o ensaio de ELISA.
Os ensaios mostram a existência de uma ligação da cistatina a preparações
de membranas e paredes de células cotiledonárias de V. vexillata e unguiculata indicados
a partir dos resultados dos ensaios anteriores. Nota-se também que não há ligação direta
da cistatina com insulina bovina, nem vice-versa (Poços 6 e 14, respectivamente). Porém
observa-se a ligação da insulina diretamente às preparações de membranas e paredes
(Poços 12 e 13). Tal interação é confirmada quando analisamos a ligação da insulina com
os complexos cistatina-parede e cistatina-membrana (Poços 7 e 8) e percebemos que ela
é bem menor do que quando a insulina liga-se diretamente, sem a presença de cistatina,
às preparações de paredes e membranas (Poços 12 e 13). Isto indicaria que a cistatina
estaria competindo com a insulina por sítios de ligação nestas frações subcelulares, o que
também reforça, outra vez, a ligação da cistatina a paredes e membranas. Um
interessante dado é o apresentado no experimento do Poço 3, cujo valor elevado de
detecção pelo anticorpo anti-cistatina sugere a presença de uma cistatina endógena
ligada à preparação de parede em V. vexillata, uma vez que comparando-se os valores de
absorbância da ligação da cistatina recombinante com o anticorpo anti-cistatina (Poço 1:
0,960) e os valores da absorbância da ligação do complexo cistatina-parede com o
mesmo anticorpo (Poço 3: 1,312), observa-se um valor muito mais alto para a segunda
situação experimental.
Page 28
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 21
Quando incubamos primeiramente a insulina nas preparações de paredes e
membranas celulares e depois incubamos a cistatina (Poços 9 e 10) observamos também
valores de ligação que estariam indicando a formação de um complexo supra-molecular
entre as estruturas celulares estudadas, a cistatina e a insulina bovina.
4.3 - Eletroforese das frações provenientes dos ensaios de fluido apoplástico
realizados em sementes de Vigna vexillata.
A figura 5 mostra a eletroforese em gel de poliacrilamida das frações
provenientes dos ensaios de fluido apoplástico, extraído de sementes de Vigna vexillata.
A visualização de tais frações por SDS-PAGE mostra o perfil protéico destas amostras e
sugere a presença de moléculas do tipo cistatina, uma vez que bandas com massas
moleculares características destas, de aproximadamente 14kDa (seta), aparecem nos
poços referentes às frações dos fluidos. Tal proteína foi detectada nos três diferentes
fluidos, exsudatos quando as sementes foram embebidas em diferentes soluções, como
especificado no item 3.4. O resultado precisa ser confirmado com a realização de Western
blotting, assim que tivermos posse do anticorpo anti-cistatina.
Page 29
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 22
A B M FPR FPR
Figura 4 – SDS-PAGE (A) e Western-blotting (B) da fração protéica do extrato
bacteriano, retida em cromatografia de afinidade em coluna de Níquel NTA-Agarose. M -
Marcadores de massa molecular; FPR - Fração protéica retida.
Obs: Os marcadores de massa molecular encontram-se na seguinte ordem de cima para
baixo: 66kDa: Albumina sérica bovina, 45kDa: Ovoalbumina, 29kDa: Anidrase carbônica,
24kDa: Tripsinogênio, 18kDa: Inibidor de tripsina e 14kDa: Lactoalbumina.
Page 30
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 23
Tabela 2 - Interações entre a cistatina recombinante, fragmentos de
membranas e paredes celulares de cotilédones de V. unguiculata e insulina bovina por
ELISA.
Poços Antígeno Incubação Anticorpo Absorbância *
1
Cistatina
-
Anti-cistatina
0,921
2
Insulina
-
Anti-cistatina
0
3
Cistatina + Parede
-
Anti-cistatina
0,917
4
Cistatina + Membrana
-
Anti-cistatina
0,812
5
Cistatina
-
Anti-insulina
0
6
Cistatina
Insulina
Anti-insulina
0
7
Cistatina + Parede
Insulina Anti-insulina
0,299
8
Cistatina + Membrana
Insulina Anti-insulina
0,323
9
Cistatina
Membrana +Insulina
Anti-insulina
0,378
10
Cistatina
Parede + Insulina
Anti-insulina
0,387
11
Insulina
-
Anti-insulina
0,598
12
Membrana
Insulina
Anti-insulina
0,465
13
Parede
Insulina
Anti-insulina
0,478
14
Insulina
Cistatina
Anti- insulina
0,520
Page 31
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 24
Tabela 3 - Interações entre a cistatina recombinante, fragmentos de
membranas e paredes celulares de cotilédones de V. vexillata e insulina bovina por
ELISA.
Poços Antígeno Incubação Anticorpo Absorbância *
1
Cistatina
-
Anti-cistatina
0,960
2
Insulina
-
Anti-cistatina
0
3
Cistatina + Parede
-
Anti-cistatina
1,312
4
Cistatina + Membrana
-
Anti-cistatina
0,848
5
Cistatina
-
Anti-insulina
0
6
Cistatina
Insulina
Anti-insulina
0
7
Cistatina + Parede
Insulina Anti-insulina
0,340
8
Cistatina + Membrana
Insulina Anti-insulina
0,341
9
Cistatina
Membrana +Insulina
Anti-insulina
0,390
10
Cistatina
Parede + Insulina
Anti-insulina
0,370
11
Insulina
-
Anti-insulina
0,595
12
Membrana
Insulina
Anti-insulina
0,450
13
Parede
Insulina
Anti-insulina
0,421
14
Insulina
Cistatina
Anti- insulina
0,590
Page 32
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 25
M F1 F2 F3
Figura 5 – SDS PAGE do perfil protéico das frações obtidas no experimento de fluido
apoplástico. (M – marcador de peso molecular [66kDa: Albumina sérica bovina, 45kDa:
Ovoalbumina, 29kDa: Anidrase carbônica, 24kDa: Tripsinogênio, 18kDa: Inibidor de
tripsina, 14kDa: Lactoalbumina); F1: fluido apoplástico 1; F2: fluido apoplástico 2; F3:
fluido apoplástico 3.
66
45
29
24 14
Page 33
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 26
4.4 – Purificação da cistatina de Vigna vexillata
4.4.1 – Cromatografia de afinidade em CnBr Sepharose 4B – CM Papaína.
A figura 6 mostra o perfil cromatográfico de extrato bruto de Vigna vexillata (10 mL)
submetido à cromatografia de afinidade em coluna Sepharose 4B CM-Papaína ativada
com CnBr. O gráfico representa a leitura, em absorbância, dos tubos coletados durante o
curso da cromatografia, evidenciando-se, após a seta vertical, os materiais eluídos pela
adição do tampão de eluição da coluna (tampão fosfato de sódio 50mM, NaCl 0,5M, pH
11,5). Observa-se um pico proeminente em torno dos tubos 149, 150, 151 e 152. Esse
material foi recolhido e utilizado em teste enzimático que mede a atividade inibitória contra
papaína (Tabela 4). Os ensaios confirmam a atividade inibitória de papaína, típica de
cistatinas, nos tubos 155, 156 e 157. Em seguida o pico referente a estes tubos contendo
a atividade inibitória foi dialisado e submetido a eletroforese e Western Blotting.
4.4.2 - Eletroforese proveniente da fração retida nos ensaio de cromatografia de afinidade para cistatina em extrato de sementes de Vigna vexillata.
A figura 7A mostra a eletroforese em gel de poliacrilamida das frações obtidas
na coluna de afinidade Sepharose 4B CM-Papaína.
A visualização das frações por SDS-PAGE mostra o perfil protéico destas
amostras e mostra a presença da cistatina, uma vez que uma banda com a massa
molecular característica desta cistatina, entre 14 e 18 kDa, aparece no poço referente à
fração. A natureza da banda protéica apontada pela seta (Figura 7B), como sendo uma
cistatina, foi confirmada por experimento de Western blotting, após transferência de
proteína do gel para uma membrana de nitrocelulose. O reconhecimento imunológico
positivo por um anticorpo específico anti-cistatina (1:2000) é observado na Figura 7B.
Uma segunda banda de massa molecular aproximada de 30 kDa, vista na
fração retida na figura 7A, pode representar a formação de um dímero, o que já foi
previamente demonstrado para a cistatina recombinante de Vigna unguiculata por Flores
et al. (2001) e Aguiar et al. (2006).
Page 34
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 27
0
0,5
1
1,5
2
2,5
1 13 25 37 49 61 73 85 97 109 121 133 145 157 169 181 193 205
Tubos
Ab
s 28
0nm
Figura 6 – Perfil cromatográfico do extrato bruto de Vigna vexillata em cromatografia
de afinidade em Sepharose 4B CM-Papaína.
pH 11,5
Page 35
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 28
Tabela 4 – Ensaio de atividade inibitória contra papaína nas amostras provenientes da
cromatografia de afinidade em Sepharose 4B CM-Papaína. UAI (Unidade de Atividade
Inibitória).
TUBOS UAI 1-154 0 155 224,5 156 321 157 325,8
158-205 0
Page 36
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 29
M ET FR M ET FR
Figura 7 - SDS-PAGE (A) e Western-blotting (B) da fração retida na coluna de
afinidade Sepharose 4B CM-Papaína. M - Marcadores de massa molecular; ET – Extrato
Total; FR - Fração protéica retida.
66
45
29 24 18 14
A B
Page 37
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 30
4.5 – Ensaio de atividade antifúngica da cistatina purificada de Vigna vexillata
contra o fungo Fusarium solani.
A figura 8 mostra a atividade antifúngica da cistatina purificada de Vigna
vexillata contra o fungo Fusarium solani. Pode-se observar, que nas mesmas condições
de tempo e temperatura, a fração da cistatina purificada foi capaz de inibir praticamente
100% o crescimento do fungo quando comparado com o controle.
As figuras 9A e 9B mostram através da análise microscópica o perfil do ensaio
antifúngico, comprovando novamente a ação inibitória da cistatina de Vigna vexillata
sobre o crescimento do fungo Fusarium solani. Na figura 9B, podemos notar nitidamente a
total inibição de formação de hifas do fungo em estudo, quando o meio de crescimento foi
acrescido da fração purificada da cistatina de Vigna vexillata.
Page 38
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 31
0
0,2
0,4
0,6
0,8
1
1,2
0 10 20 30 40 50 60
Tempo, h
Abso
rbân
cia,
670
nm
Controle
Cistatina
Figura 8 – Ensaio da atividade antifúngica da cistatina purificada de Vigna
vexillata (200 µg de proteína) contra o fungo F. solani .
Page 39
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 32
Figura 9 A – Fotomicrografia da análise em microscópio ótico do fungo
Fusarium solani crescido em meio de cultura controle.
Figura 9B - Fotomicrografia da análise em microscópio ótico do fungo
Fusarium solani crescido em meio de cultura controle, acrescido de cistatina
purificada de Vigna vexillata (200 µg de proteína)
Page 40
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 33
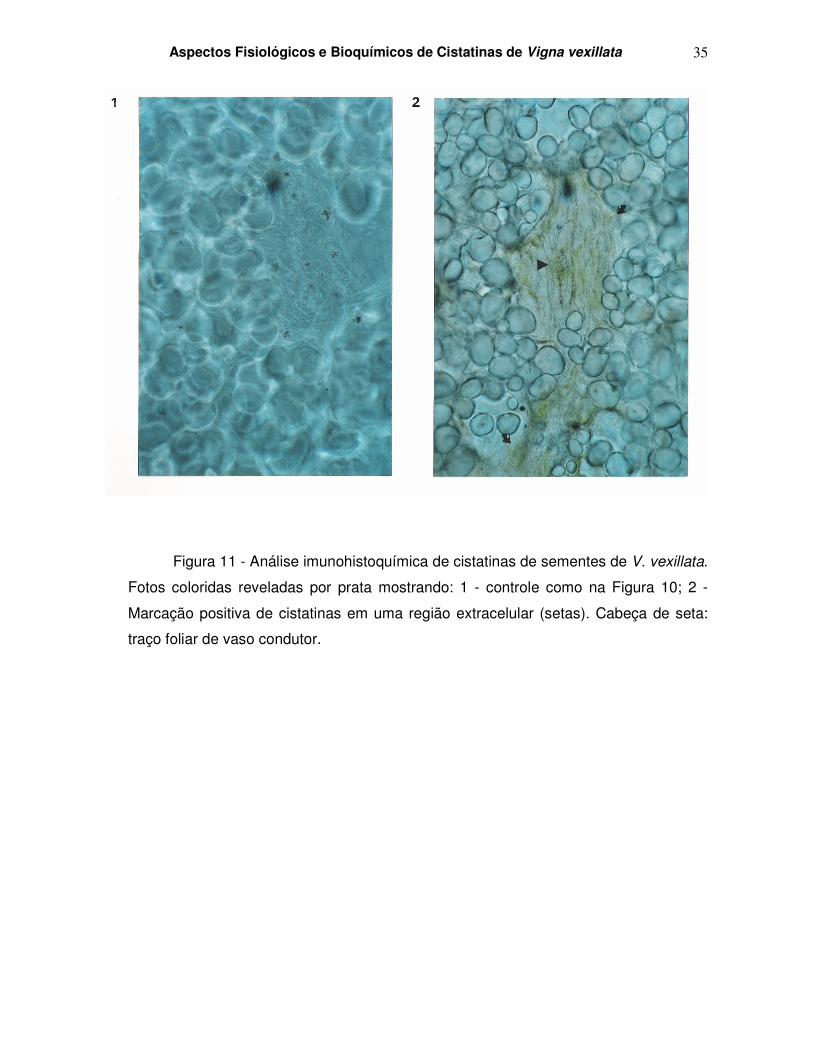
4.6 – Localização tecidual de cistatinas de V. vexillata
Nos estudos ultraestruturais, uma marcação positiva foi vista em citoplasma e
espaço extracelular cotiledonário de sementes de V. vexillata ao utilizar-se um anticorpo
anti-cistatina de V. unguiculata. As Figuras 10 e 11 mostram a análise imunohistoquímica
de cistatinas de sementes de V. vexillata, inicialmente mostradas por fotos em preto e
branco, reveladas por prata (Figura 10) e em seguida por fotos coloridas (Figura 11),
mostrando a marcação por setas e cabeças de setas.
Page 41
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 34
Figura 10 - Análise imunohistoquímica de cistatinas de sementes de V. vexillata.
Fotos em preto e branco, reveladas por prata mostrando: 1 - Controle sem anticorpo
primário; 2 - Marcação positiva no citoplasma e em camada circundante das células
cotiledonárias (setas).
Page 42
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 35
Figura 11 - Análise imunohistoquímica de cistatinas de sementes de V. vexillata.
Fotos coloridas reveladas por prata mostrando: 1 - controle como na Figura 10; 2 -
Marcação positiva de cistatinas em uma região extracelular (setas). Cabeça de seta:
traço foliar de vaso condutor.
Page 43
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 36
5 - DISCUSSÃO
Cistatinas são proteínas com elevados potenciais defensivos contra pestes e
patógenos que vêm sendo empregadas em programas de transformação genética de
plantas visando a melhoria de suas habilidades defensivas contra predadores (Hines et al,
1981). Uma das principais forças motrizes de tal aplicação biotecnológica é o fato de
considerarem-se os genes de cistatinas como os, talvez, melhores candidatos aos
programas de transformação de plantas visando o incremento de suas habilidades
defensivas (Urwin et al., 1997).
Existe uma grande diversidade de papéis fisiológicos propostos para tal classe
de proteína. Um desses papéis foi proposto em trabalho de Satoh (1998). Este autor
verificou uma capacidade de ligação entre uma cistatina de culturas de células de
sementes de cenoura com glicoproteínas solúveis extracelulares, que por sua vez, ligava-
se a proteínas homólogas à insulina. É de nosso grande interesse confirmar esta
capacidade de ligação uma vez que nosso estudo demonstrou que, além de uma
localização citoplasmática principal, as cistatinas de sementes de Vigna vexillata também
são encontradas em depósitos extra-celulares nos cotilédones destas sementes e que,
portanto, um processo de translocação deve ser admitido (Figuras 10 e 11). Nosso grupo
também isolou e caracterizou pela primeira vez, na literatura, uma insulina de origem
vegetal, em tegumentos de sementes de Canavalia ensiformis (Oliveira et al., 1999). Tais
resultados serão bastante úteis para a avaliação de mais uma possível função para a
classe das cistatinas, que é a de estas proteínas serem participantes de processos de
interação tecidual e transdução de sinais.
Em virtude das sementes de V. vexillata apresentarem dimensões diminutas e
as cistatinas serem encontradas em concentrações muito baixas em sementes
(Fernandes et al., 1991), para este trabalho foi preciso, inicialmente, expressarmos uma
cistatina recombinante de V. unguiculata através da utilização de células bacterianas de
E. coli, que já haviam sido previamente transformadas, para expressarem a proteína de
interesse (gentilmente cedidas pelo Dr. Peter Urwin - UK). A expressão de proteínas em
bactérias é estratégia freqüentemente adotada diante da necessidade de obtenção de
quantidades suficientes de uma dada proteína. Neste sentido podemos mencionar as
expressões em bactérias da cistatina de milho (Abe et al., 1994), cistatina de folha de
mamão (Song et al., 1995), orizacistatina I e II de arroz (Michaud et al., 1994), cistatinas
Page 44
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 37
de folhas de repolho (Lim et al.,1996), de sementes de soja (Misaka et al., 1996) e de
sementes de feijão-de-corda (Urwin et al., 1995).
O isolamento da cistatina recombinante expressa por tais células foi alcançado
pela percolação do extrato bacteriano, cujas células haviam sido lisadas por sonicação,
através de uma coluna de níquel NTA-agarose. Tal isolamento pode ser verificado por
SDS-PAGE e por Western-blotting (Figura 4) onde verificou-se a presença de uma banda
de aproximadamente 13 kDa, responsiva ao anticorpo anti-cistatina de Vigana vexillata.
É importante ressaltar que a cistatina que está sendo empregada neste projeto
é uma cistatina recombinante de Vigna unguiculata que já havia sido clonada por
Fernandes et al. (1993), que demonstraram ser, esta, uma proteína que não possuía um
peptídeo sinal, portanto não podendo ser extracelular. Tal fato é de extrema relevância
uma vez que estamos detectando ligação desta cistatina a elementos constituintes de
paredes e membranas em uma tentativa de investigar uma proposição da função para
cistatinas descrita por trabalho de Satoh (1998). A diferença fundamental é que a cistatina
estudada por Satoh (1998) é uma cistatina extracelular obtida a partir de tais culturas de
células de cenoura.
Nossos ensaios de ligação sugeriram potencialidades de ligação da cistatina
recombinante de Vigna unguiculata tanto às paredes como às membranas celulares de
ambas as sementes, V. unguiculata e V. vexillata (Tabelas 2 e 3). Os dados também
apontam claramente para a ligação tanto da cistatina recombinante, quanto da insulina
bovina, às preparações de paredes e membranas celulares de cotilédones de V. vexillata
e unguiculata. Deve-se assumir, portanto, a presença de ligantes para tais proteínas nas
diferentes frações subcelulares. Nota-se, ainda, que o valor de absorbância-controle da
reação da cistatina recombinante com o anticorpo anti-cistatina é mais baixo que aquela
referente à reação da cistatina incubada às preparações de paredes e o mesmo anticorpo
na espécie V. vexillata, sugerindo que o anticorpo estaria reconhecendo uma outra
cistatina, que provavelmente é a cistatina endógena, sugerida como presente em
preparações de paredes celulares de cotilédones de V. vexillata. Também é possível que
esta cistatina seja externalizada, como suspeitado pela presença de bandas de massa
molecular em torno de 13 kDa, visualizada no fluido apoplástico de sementes de V.
vexillata (Figura 5)
Tal consideração é de grande interesse para estudos futuros, uma vez que
não se conhece na literatura nenhuma cistatina localizada em paredes celulares de
sementes.
Page 45
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 38
Em 1989, Kagawa & Hirano mostraram a presença de proteínas do tipo
globulinas 7S, em soja, que eram capazes de se ligarem à insulina suína, sugerindo para
essas proteínas uma possível função de receptor para o hormônio insulina. Essa proteína
foi denominada Bg e consiste de duas cadeias polipeptídicas de 16 e 26 kDa, unidas
entre si por pontes dissulfetos. Trabalhos posteriores demonstraram que essas proteínas
apresentavam atividade de proteína quinase semelhante à atividade tirosina quinase,
típica do receptor de insulina animal.
Proteínas do tipo globulinas 7S básicas, imunorrelacionadas a Bg (globulina
7S básica de soja), já foram detectadas em uma grande variedade de sementes, dentre
elas, Vigna radiata, Vigna angularis e Lupinus alobus (Komatsu & Hirano, 1991).
Satoh, em 1998, encontrou em sementes de cenoura uma proteína de 57 kDa,
a qual denominou de EDGP. Tal proteína apresentou homologia de seqüência de
aminoácidos com a Bg e também era capaz de se ligar à insulina. Posteriormente, foi
isolada uma proteína de radículas de soja que era capaz de estimular a fosforilação da Bg
in vitro, a qual foi denominada leginsulina (Ilgoutz et al., 1997).
Acreditamos, com isso, que também existam, em nossas preparações de
paredes e membranas, proteínas similares que estejam sendo ponte de ligação para a
insulina e também exista a presença de um ligante, nestas frações subcelulares, para a
cistatina.
A formação de um complexo supra-molecular observada entre as frações
subcelulares analisadas, a cistatina e a insulina bovina, evidenciada principalmente pelos
nossos ensaios de ELISA, vem reforçar a proposta de Satoh (1998) que sugere que
cistatinas extracelulares de células de cenoura estariam envolvidas em processos de
transdução de sinais celulares e de interação tecidual entre células.
A utilização da cromatografia de afinidade em Sepaharose CM-papaína foi bem
sucedida no que se refere à purificação da cistatina de Vigna vexillata. A proteína
purificada apresentou massa molecular em torno de 15kDa, e a formação de um dímero
de cerca de 30 kDa foi visualizada por experimentos de eletroforese sob condições
desnaturantes, a exemplo do já observado para a cistatina de Vigna unguiculata por
Flores et al. (2001) e Aguiar et al. (2006). A proteína eluída desta cromatografia de
afinidade foi imunoreativa com o anticorpo anti-cistatina de Vigna unguiculata, quando
submetida a experimento de Western blotting.
A cistatina de V. vexillata foi vista como capaz de inibir em 100% o crescimento
do fungo fitopatogênico F. solani, causando também um drástico efeito sobre a formação
Page 46
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 39
de hifas, em concentração de 200 µg/mL. Os ensaios de atividade inibitória de
crescimento deste fungo reforçam uma das funções já descritas para as fitocistatinas, que
é o papel de defesa vegetal contra diversos predadores, como os fungos e ácaros
(Pernas et al., 1999; Siqueira-Junior et al., 2002; Soares-Costa et al., 2002; Martinez et al.,
2003; Yang & Yeh, 2005; Christova et al., 2006; Abraham et al., 2006). Martinez et al.
(2005) mostraram que a cistatina recombinante de morango, FaCPI-1, inibiu quase
completamente o crescimento dos fungos Botritis cinérea, a uma concentração de 3 µM, e
Fusarium oxysporum, em concentrações mais elevadas, de 6 µM. A exemplo do
observado neste trabalho, com a cistatina de V. vexillata, o efeito inibitório observado deu-
se sobre ambos os processos de germinação de esporos e desenvolvimento micelial. É
de extrema importância comparar o potencial inibitório da cistatina de V. vexillata com
outras cistatinas já estudadas, tais como a cistatina de tomate, como descrito por
Siqueira-Júnior et al. (2002), uma vez que com 200 µg de cistatina de V. vexillata o
crescimento do fungo foi quase 100% inhibido enquanto que a cistatina de tomate, nas
mesmas concentrações não inibe nem 50% do crescimento dos fungos Trichoderma
viride (non-phytopathogenic) and Fusarium solani (phytopathogenic). A cistatina de tomate
(TC), como visto por tais autores, causa uma redução de cerca de 18% no crescimento de
T. viride e de 45% no crescimento de F. solani quando testada em concentrações de
150 µg ml–1. Abraham et al. (2006) demonstraram que seis de sete cistatinas de cevada
foram capazes de inibir o crescimento dos fungos B. cinerea e F. oxysporum, sendo as
cistatinas HvCPI-2 e HvCPI-6 as duas mais eficazes quando calculadas as Concentrações
Efetivas para inibição de 50% do crescimento (EC50), as quais foram <1.5 µM para ambos
os fungos. Os experimentos realizados por Yang & Yeh (2005), com cistatinas de
tubérculos de inhame (Colocasia esculenta) mostraram que em concentrações de 150
µg/ml ou 200 µg/ ml, o crescimento micelial de Sclerotium rolfsii Sacc. era fortemente
inibido. A morfologia das hifas, observada por microscopia óptica microscope, revelou
filamentos mais curtos e delgados.
Embora o mecanismo de atividade antifúngica destas fitocistatinas ainda não
esteja completamente estabelecido, Martinez et al. (2003) demonstraram por mutagênese
sítio-dirigida que a inibição de Botrytis cinerea pela cistatina de cevada não está
associada com a propriedade inibitória de proteinases cisteínicas desta proteína.
Especula-se que alterações na permeabilidade de membranes fúngicas possam ser a
origem dos problemas causados pelas fitocistatinas nestes organismos. O inibidor de
Page 47
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 40
tripsina conhecido como SAP16 de Helianthus annuus modifica a permeabilidade da
membrana celular de Sclerotinia scleroticum e reprime a germinação de ascósporos
(Giudici et al., 2000). Alterações de permeabilidade de membrana foram também
detectadas quando a orizacistatina OC-I foi expressa no compartimento citossólico de
células de folhas de tabaco (van de Vyver et al., 2003). No entanto, Yang and Yeh (2005)
sugeriram recentemente que a inibição de crescimento do fungo fitopatogênico Sclerotium
rolfsii causada por uma cistatina de tubérculos de Colocasia esculenta relacionava-se
diretamente a inibição de uma proteinase cisteínica do fungo. Conclui-se, portanto, que
trabalhos adicionais serão necessários para o estabelecimento dos mecanismos de ação
antifúngica de cada fitocistatina sobre diferentes patógenos.
Também faz-se importante ressaltar que, a localização da cistatina de V.
vexillata nos espaços extracelulares (Figuras 10 e 11), por experimentos preliminares,
condiz muito bem com uma função de defesa contra tais organismos patogênicos.
Trabalhos de imunolocalização de cistatinas em plantas, em geral, apontam para
localizações citoplasmáticas, tais como o de Boulter and Harvey (1985), acerca de uma
multicistatina de batata e o de Madureira et al. (2006), acerca de uma cistatina de folhas
de tomate de 87 kDa. Tais localizações citoplasmáticas também indicam correlações com
potenciais papéis defensivos, ao contrário das sugestões de papéis fisiológicos propostos
para cistatinas de sementes de soja (Misaka et al., 1996) e de Vigna unguiculata (Flores
et al., 2001), as quais sugerem-se estar envolvidas na regulação de proteinases
cisteínicas endógenas, importantes para os processos de desenvolvimento e germinação
de sementes. A localização em espaços extra-celulares e cotiledonários das cistatinas de
V. vexillata, aqui apresentadas, bem como as de cistatinas de tomate (Madureira et al.,
2006) e batata (Boulter and Harvey, 1985), diferem significativamente de trabalhos
anteriores investigativos da compartimentalização de outros inibidores de proteases.
Diversos destes inibidores, tais como os inibidores de proteases serínicas induzidos por
lesão ou metil-jasmonato de folhas de tomate, foram encontrados em vacúolos centrais,
inclusive associados a corpos protéicos (Walker-Simmons and Ryan, 1977; Narváez-
Vásquez et al., 1993; Villanueva et al., 1998) Um inibidor de proteinases cisteínicas de
batata, mas que no entanto não apresenta homologia com cistatinas de plantas, foi
também principalemente localizado em vacúolos de plantas tratadas com metil jasmonato
(Gruden et al., 1997).
Page 48
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 41
6 – Conclusões
1. A ligação de uma cistatina recombinante de V. unguiculata com porções de
paredes e membranas celulares de células cotiledonárias de V. vexillata é
fortemente sugerida pelas diferentes estratégias metodológicas apresentadas no
trabalho.
2. Diferentes componentes de paredes e membranas celulares devem estar
envolvidos em tais interações.
3. Não observou-se ligação direta da insulina com a cistatina recombinante de V.
unguiculata, ou vice-versa, por ensaio de ELISA modificado.
4. Ligações entre insulina bovina e preparações de paredes e membranas celulares
de células cotiledonárias de Vigna vexillata foram mostradas por ensaio de ELISA
modificado.
5. Há indícios de uma cistatina endógena ligada às preparações de paredes celulares
de sementes de Vigna vexillata.
6. Foi verificado através de ensaios imunohistoquímicos que a cistatina se localiza
principalmente no citoplasma das sementes de Vigna vexillata.
Page 49
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 42
6 -REFERÊNCIAS BIBLIOGRÁFICAS
ABE, M., Kuroda, M., Arai, S. (1992) Corn kernel cysteine proteinase inhibitor as a novel
cystatin superfamily member of plant origin. Molecular cloning and expression studies.
Eur. J. Biochem. 209:933-937.
ABE, M., Abe, K., Iwabuchi, K., Domoto, C., Arai, S. (1994) Corn cystatin I expressed in
Eschechia coli: Investigation of its inhibitory profile and occurrence in corn kernels. J.
Biochem. 116: 488-492.
ABE, M. and ARAI, S. (1991) Some properties of a cysteine proteinase inhibitor from corn
endosperm. Agric. Biol. Chem. 55: 2417-2418.
ABE, M., Abe, K.,. Domoto, C., Arai, S. (1995) Two distinct species of corn cystatin in corn
kernels. Biosci. Biotech. Biochem. 59: 756-758.
ABE, M., Emori, Y., Kondo, H., Suzuki, K., Arai, S. (1987) Molecular cloning of a cystein
proteinase inhibitor of rice (oryzacystatin). Homology with animal cystatins and transient
expression in the ripening process of rice seeds. J. Biol. Chem. 262: 16793-16797.
ABE, Y., Shirane, K., Yokosawa, H., Matsushita, H., Mitta, M., Kato, I., Ishii, S. (1993)
Asparaginyl endopeptidase of Jack bean seeds. J. Biol. Chem. 268: 3525-3529.
ABE, M. and WHITAKER, J. R. (1988) Purification and characterization of a cysteine
proteinase inhibitor from the endosperm of corn. Agric. Biol. Chem. 52: 1583-1584.
ABE, M., Domoto, C., Watanabe, H., Abe, K., Arai, S. (1996) Structural organization of the
gene encoding corn cystatin. Biosci. Biotech. Biochem. 60: 1173-1175.
Page 50
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 43
ABRAHAM, Z., Martinez, M., Carbonero, P., Diaz, I. (2006) Structural and functional
diversity within the cystatin gene family of Hordeum vulgare. J. Exp. Bot. 57:4245-4255
AGUIAR, J.M., Franco, O.L., Rigden, D.J., Bloch Jr. C., Monteiro, A.C.S., Flores, V.M.Q.,
Jacinto, T., Xavier-Filho, J., Oliveira, AE.A., Grossi-de-Sá, M.F., Fernandes, K.V.S. (2006)
Molecular modeling and inhibitory activity of cowpea cystatin against bean bruchid pests.
Proteins: Struct. Funct. Bioinform. 63: 662-670.
ÁLVAREZ-ALFAGEME, F., Martinez, M., Pascual-Ruiz, S., Castanera, P., Diaz, I., Ortego,
F. (2007) Effects of potato plants expressing a barley cystatin on the predatory bug
Podisus maculiventris via herbivorous prey feeding on the plant. Transg. Res. 16: 1-13.
ÁVILA, D.S., Cunha, M., Jacinto T., Xavier-Filho J., Fernandes, K. V.S., Expression of
phytocystatins in cultivated and wild legume seeds. Caxambu, 1999. Resumos de Anais -
XXVIII Reunião Anual da SBBq, E-64, 35.
AZEVEDO, C.R.; Maciel, F. M.; Silva, L. B.; Ferreira, A. T. S.; Da Cunha, M;Machado, Ol
L. T.; Fernandes, K. V. S; Oliveira, A. E. A.; Xavier-Filho, J. (2006). Isolation anda
intracellular localization of insulin-like proteins from leaves of Bauhinia variegata. Braz.
Jour. Med. Biol. Res. 39:1435-1444.
BARRETT, A. J. (1986) The classes of proteolytic enzymes. In: Plant Proteolytic Enzymes.
Ed. M. J. Dalling. 1: 1-16 Boca Raton, F1: CRC Press .
BARRETT, A. J. and Salvesen, G (1986) In: Proteinase Inhibitors, New York: Academic
Press.
BARRETT, A. J., Rawlings, N. D., Davies, M. E., Machleidt, W., Salvesen, G., Turk, V
(1987). In: Cysteine proteinase inhibitors of the cystatin superfamily 515-569.
BEST, C.H. (1924). Recent work on insulin. Endocrinology. 1: 617-629.
BOULTER, D.,Gatehouse, A. M. R., Hilder, V. (1989) Use of cowpea trypsin inhibitor
(CpTI) to protect plants against insect predation. Biotechnol. Adv. 7: 489-497.
Page 51
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 44
BOULTER, D. and Harvey, P.J. (1985) Accumulation, structure and utilisation of tuber
storage proteins with particular reference to Dioscorea rotundata, Physiol. Veg. 23: 61–74.
BRADFORD, M. M. (1976) A rapid and sensitive method for quantitation of microgram
quantities of protein utilizing of dye binding. Anal. Biochem., 72: 248-254.
BRZIN, J., Popovic, T., Ritonja, A., Puizdar, V. and Kidric, M. (1998) Related cystatin
inhibitors from leaf and seed of Phaseolus vulgaris L. Plant Science 138: 17-26.
BU, Q-Y., Wu, L., Yang, S-H., Wan, J-M. (2006) Cloning of a potato proteinase inhibitor
gene PINII-2x from diploid potato (Solanum phurejia L.) and transgenic investigation of its
potential to confer insect resistance in rice; J. Integrat. Plant Biol. 48: 732-739.;
CERCOS, M. and Carbonell, J. (1993) Purification and characterization of a thiol-protease
induced during senescence of unpollinated ovaries of Pisum sativum. Plant Physiol. 88:
267-274.
CHEN, M.S., Johnson, B., Wen, I., Muthukrishnan, S., Kramer, K.J., Morgan, T.D., Reeck,
G.R., (1992) Rice cystatin - bacterial expression, purification, cysteine proteinase inhibitory
activity, and insect growth suppressing activity of a truncated form of the protein. Prot.
Expr. Purif. 3:41-49.
CHRISTOVA, P.K, Christov, N.K., Imai, R. (2006) A cold inducible multidomain cystatin
from winter wheat inhibits growth of the snow mold fungus, Microdochium nivale. Planta
223: 1207–1218.
COLLIER, E.; Watkinson, A.; Cleland, C.F.; Roth, J. (1987). Partial purification and
characterization of an insulin-like material from spinach and Lemna gibba G3. J. Biol.
Chem. 262: 6238-6247.
COLLIP, J.B. (1923) Glucokinin. A new hormone present in plant tissue. Preliminary
paper. J. Biol. Chem. 56: 513-543.
Page 52
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 45
CORRE-MENGUY, F., Cejudo, F. J, Mazuber,t C., Vidal, J., Lelandais-Briere, C., Torres,
G., Rode, A., Hartmann, C. (2002) Characterization of the expression of a wheat cystatin
gene during caryopsis development. Plant Mol. Biol. 50: 687-698.
CSOMA, C. and Polgar, L. (1984) Proteinase from germinating bean cotyledons. Biochem.
J. 122: 769-776.
CUATRECASAS, P. (1970) Protein purification by affinity chromatography. J. Biol. Chem.
245: 3059-3065.
DA CUNHA, P. (2005) Interações de cistatinas com componentes de membranas e
paredes celulares de sementes do gênero Vigna vexillata. Monografia apresentada ao
Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense
para obtenção do Grau de Bacharelado em Ciências Biológicas.
DE LEO, F. (2002) Plant protease inhibitor family. Nucleic Acids Res. 30: 347-348.
FERNANDES, K. V. S., Campos F. A. P., do Val, R. R., Xavier-Filho, J. (1991) The
expression of papain inhibitors during development of cowpea seeds. Plant Sci., 74: 179-
184.
FERNANDES, K.V.S., Sabelli P. A., Barratt, D.H.P., Richardson, M., Xavier-Filho, J.,
Shewry, P. (1993) The resistance of cowpea seeds to bruchid beetle is not related to
levels of cysteine proteinase inhibitors. Plant Mol. Biol. 23:215-219.
FLORES, V. M. Q. (1997) Cistatinas de Vigna unguiculata: expressão temporal e tecidual
em sementes e heteróloga em Escherichia coli. Fortaleza, 1997. Tese de Mestrado.
Departamento de Bioquímica e Biologia Molecular do Centro de Ciências – Universidade
Federal do Ceará.
FLORES, V. M. Q., Louro, R. P., Xavier-Filho, J., Barratt, D. H. P., Shewry, P. R.,
Fernandes, K. V. S. (2001) Temporal and tissue localization of a cowpea (Vigna
unguiculata) cystatin. Physiol. Plantarum 112: 195-199.
Page 53
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 46
FLORES, V. M. Q., Fernandes, R. C. S. C., Macedo, Z. S., Medina-Acosta, E. (2002)
Expression and purification of the recombinant enteropathogenic Escherichia coli vaccine
candidates BfpA and EspB. Prot. Expr. Purif. 25: 16–22.
GIANOTTI, A., Rios, W.M., Soares-Costa, A., Nogaroto, V., Carmona, A.K., Oliva, M.L.V.,
Andrade, S.S., Henrique-Silva, F. (2006) Recombinant expression, purification, and
functional analysis of two novel cystatins from sugarcane (Saccharum officinarum). Prot.
Expr. Purif.
47: 483-489.
GIUDICI, A.M., Regente, M.C., Canal, L. (2000). A potent antifungal protein from
Helianthus annuus flowers is a trypsin inhibitor. Plant Physiol. Biochem. 38: 881–888.
GHOLIZADEH, A., Santha, I.M., Kohnehrouz, B.B., Lodha, M.L., Kapoor, H.C. (2005)
Cystatins may confer viral resistance in plants by inhibition of a virus-induced cell death
phenomenon in which cysteine poteinases are active: cloning and molecular
characterization of a cDNA encoding cysteine proteinase inhibitor (celostatin) from Celosia
cristata (crested cock´s comb). Biotechnol. Appl. Biochem. 42: 197-204.
GRUDEN, K., Strukelj, B., Ravnikar, M., Poljsak-Prijatelj, M., Mavric, I., Brzin, J.,
Pungercar, J. Kregar, I. (1997) Potato cysteine proteinase inhibitor gene family: molecular
cloning, characterisation and immunocytochemical localisation studies. Plant Mol. Biol. 34:
317–323.
HILDER, V.A., Boulter, D. (1999) Genetic engeneering of crop plants for insect resistance
- a critical review. Crop Protect. 18: 177-191.
HILDER, V. A., Gatehouse, A. M. R., Sherman, S. E., Barker, R. F., Boulter, D. (1987) A
novel mechanism of insect resistance engeneered into tobacco. Nature 330: 160-163.
HINES, M.E., Osuala, C. J., Nielsen, S.S. (1991) Isolation and partial characterization of a
soybean cystatin cysteine proteinase-inhibitor of coleopteran digestive proteolytic activity.
J. Agric. Food. Chem. 39:1515-1520.
Page 54
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 47
HIRASHIKI, I., Ogata, F., Yoshida, N., Makisumi, S. and Ito, A. (1990) Purification and
complex formation analysis of a cysteine proteinase inhibitor (Cystatin) from seeds of
Wisteria floribunda. J. Biochem. 108: 604-608.
HONG, J.K., Hwang, J.E., Lim, C.J., Yang, K.A., Jin, Z-L., Kim, C.Y., Koo, J.C., Chung,
W.S., Lee, K.O., Lee, S.Y., Cho, M.J., Lim, C.O. (2007) Over-expression of Chinese
cabbage phytocystatin 1 retards seed germination in Arabidopsis. Plant Sci. 172: 556-563.
HOUSEMAN, J. G. (1978) A thiol – activated digestive proteinase from adults of Rhodnius
prolixus Satl (Hemiptera: Reduviidae). Can. J. Zool. 56: 1140-43.
HOUSEMAN, J. G. and Downe, A. E. R. (1980) Cathepsin D-like activity in the posterior
midgut of hemiptera insects. Comp. Biochem. Physiol. B. 75: 509-12.
ILGOUTZ, S.C.; Knittel, N.; Lin, J.M.; Sterle, S.; Gayler, K.R.; (1997). Transcription of
genes for conglutinin ץ and a leginsulin-like protein in narrow-leafed lupin. Plant. Mol. Biol.
34: 613-627.
IRIE, K., Hosoyama, H., Tackeuchi,T. ,Jwabuchi, K., Watanabe, H., Abe, M., Abe, K. and
Arai, S. (1996) Transgenic rice established to express corn cystatin exhibits strong
inhibitory activity against gut proteinases. Plant Mol. Biol. 30:149-157.
JAMES, A. T. and R. J. Lawn. (1991) Inheritance of select traits in accessions of Vigna
vexillata (L.) A. Rich. of Australian and African origin. Austr. J.Bot. 39:415-429.
JOHNSON, R., Narvaez, J. Na, J., Ryan, C. A. (1989) Expression of proteinase inhibitors I
and II in transgenic tobacco plants: Effects on natural defense against Manduca sexta
larvae. Proc. Natl. Acad. Sci. USA 86:9871-9875.
KAGAWA, H. and HIRANO, H. (1989). Sequence of cDNA encoding soybean basic 7S
globulin. Nucl. Acids Res. 17: 8868-8872.
KHANNA, P.; Nag, T.N.; Chandrajaia, S.; Mohan, S. (1976). Process for isolation of insulin
from plant source. United States Patent. 3: 945-988.
Page 55
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 48
KIMURA, M., Ikeda, T., Fukumoto, D., Yamasaki, N. and Yokemura, M. (1995) Primary
structure of cysteine proteinase inhibitor from the fruit of avocado (Persea Americana Mill).
Biosci. Biotech. Biochem. 59: 2328-2329
KOIZUMI, M., Yamaguchi-Shinozaki, K., Tsuji, H.. Shinozaki, K. (1993) Structure and
expression of the two genes that encode distinct drought-inducible cysteine proteinases in
Arabidopsis thaliana. Gene 129: 175-182.
KOKWARO, J. O. (1976) Medicinal plants of East Africa. East African Literature Bureau,
General Printers, Kampala, Nairobi, Dar Es Salam.
KONDO, H., Abe, K., Emori, Y.. Arai, S. (1991) Gene organization of oryzacystatin-II, a
new cystatin superfamily member of plant origin, is closely related to that of oryzacystatin-I
but different from those of animal cystatin. FEBS Lett. 278: 87-90.
KOMATSU, S. and HIRANO, H.; (1991). Plant basic 7S globulin-like proteins have insulin
and insulin-like growth factor binding activity. Science 294: 210-212.
KOUZUMA, Y., Kawano, K., Kimura, M., Yamasaki, N., Kadowaki T., Yamamoto K. (1996)
Purification, characterization and sequencing of two cysteine proteinase inhibitors, Sea
and Scb, from sunflower (Heliantus annuus) seeds. J. Biochem. 119: 1106-1113.
KOUZUMA, Y., Inanaga, H., Doi-Kawano, K., Yamasaki, N., Kimura, M. (2000) Molecular
cloning and functional expression of cDNA enconding the cysteine proteinase inhibitor with
three cystatin domains from sunflower seeds. J. Biochem. 128: 161-166.
KOUZUMA, Y., Tsukigata, K., Inanaga, H., Doi-Kawano, K., Yamasaki, N.and Kimura M.
(2001) Molecular clonig and functional expression of cDNA enconding the cysteine
proteinase inhibitor Sca from sunflower seeds. Biosc. Biotecn. Biochem. 65: 969-972.
KURODA, M., Ishimoto, M., Suzuki, K., Kondo, H., Abe, K., Kitamura, K. and Arai, S.
(1996) Oryzacystatins exhibit growth-inhibitory and lethal effects on different species of
Page 56
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 49
bean insect pests Callosobruchs chinenis (Coleoptera) and Riptortus clavatus
(Hemiptera). Biosci. Biotech. Biochem. 60:209-212.
LAEMMLI, U.K. (1970) Cleavage of structural protein during the assembly of the head of
bacteriophage T4. Nature, 680-685.
LARA, P., Ortego, F., Gonzalez-Hidalgo, E., Castañera, P., Carbonero, P., Diaz, I. (2000)
Adaptation of Spodoptera exigua (Lepidoptera : Noctuidae) to barley trypsin inhibitor BTI-
CMe expressed in transgenic tobacco. Transgen. Res. 9: 169-178.
LIM, C. H., Lee, S. J., Chung, W. S., Park, S. H., Hwang, J., Cho, M. J. (1996)
Characterization of a cDNA enconding cysteine proteinases inhibitor from Chinese
cabbage (Brassica campestris L. ssp. Pekinensis) flower buds. Plant Mol. Biol. 30:373-
379.
LOPES, M.A.C. Avaliação preliminar do conteúdo protéico da torta de filtro, um
subproduto da indústria açucareira. Campos dos Goytacazes, 2001.
MADUREIRA, H.C., Da Cunha, M., Jacinto, T. (2006) Immunolocalization of a defense-
related 87 kDa cystatin in leaf blade of tomato plants. Environm. Exp. Bot. 55: 201-208.
MARECHAL, R., Mascherpa, J. M. and Stainer, F. (1978) Etude taxonoique d’un grupe
complexe d’espécies des genres Phaseolus et Vigna (Papilionaceae) sur la base de
donnès morphologiques et polliniques, traitées par l’analyse informatique. Boissiera 28: 1-
273.
MARGIS, R., Reis, E. M., Villeret, V. (1998) Structural and phylogenetic relationships
among plant and animal cystatins. Arch. Bioch. Bioph. 359: 24-30.
MARTINEZ, M., Lopez-Solanilla, E., Rodriguez-Palenzuela, P., Carbonero, P., Diaz, I.
(2003a) Inhibition of plant-pathogenic fungi by the barley cystatin HvCPI (gene Icy) is not
associated with its cysteine proteinase inhibitory properties. Mol. Plant–Microbe Interac.
16: 876–883.
Page 57
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 50
MARTINEZ, M., Abraham, Z., Gambardella, M., Echaide, M., Carbonero, P. and Diaz, I.
(2005b) The strawberry gene Cyf1 encodes a phytocystatin with antifungal properties, J.
Exp. Bot. 56 1821–1829.
MARTINEZ, M., Rubio-Somoza, I., Fuentes, R., Lara, P., Carbonero, P. and Diaz, I.
(2005a) The barley cystatin gene (Icy) is regulated by DOF transcription factors in
aleurone cells upon germination, J. Exp. Bot. 56 547–556.
MASOUD, S. A., Johnson, I. B., White, F. F. and Reeck, G. R. (1993) Expression of a
cysteine proteinase inhibitor (Oryzacystatin I) in transgenic tobacco plants. Plant Mol. Biol.
21, 655-663.
MICHAUD, D., Nguyen-Guoc, B., Bernier-Vadnais, N., Faye, I. and Yelle, S. (1994)
Cysteine proteinase forms in sprouting potato tuber. Physiol. Plant. 90, 497-503.
MISAKA, T., Kuroda, M., Iwabuchi, K., Abe, K., Arai, S. (1996) Soyacystatin, a novel
cysteine proteinase inhibitor in soybean, is distinct in protein structure and gene
organization from other cystatins of animal and plant origin. Eur. J. Biochem. 240: 614–
690.
MITSUHASHI, W., Koshiba, T., Minamikawa, T. (1986) Separation and characterization of
endopeptidases from cotyledons of germinating Vigna mungo seeds. Plant Physiol. 86:
628-634.
MOCHIZUKI, A., Nishizawa, Y., Onodera, H., Tabei, Y., Toki, S., Habu, Y., Ugaki, M.,
Ohashi, Y. (1999) Transgenic rice plants expressing a trypsin inhibitor are resistant against
rice stem borers, Chilo suppressalis. Entomol. Exp. Appl. 93: 173–178.
MURDOCK, L. L., Brookhart, G., Dunn, P. E., Foard, D. E. and Kelley, S. (1987) Cysteine
digestive proteinases in coleoptera. Comp. Biochem. Physiol. B 87: 783-87.
NARVÁEZ-VÁSQUEZ, J., Franceschi, V.R., Ryan, C.A. (1993) Proteinase-inhibitor
synthesis in tomato plants: evidence for extracellular deposition in roots through the
secretory pathway. Planta 189: 257–266.
Page 58
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 51
OGHIAKHE, S., Jackai, L.E.N., Hodgson, C.J. and Q.N. Ng. (1993) Anatomical and
biochemical parameters of resistance of the wild cowpea, Vigna vexillata Benth. (acc.
TVNu 72) to Maruca testualis Geyer (Lepidoptera: pyralidae). Insect Sci. Appl. 14: 315-
323.
OJIMA, A., Shiota, H., Higashi, K. and Kamada, H. (1997) An extracellular insoluble
inhibitor of cysteine proteinases in cell cultures and seeds of carrot. Plant Mol. Biol. 34:99-
109.
OKOROKOV, A. I. and Iehle, I. (1998) Ca2+-ATPases of Saccharomyces cerevisiae:
diversity and possible role in protein sorting. FEMS Microbiol. Lett. 162: 83-91.
OLIVEIRA, A.E.A., Machado O.L.T, Gomes, V.M., Xavier- Neto J., Pereira, A.C., Vieira,
J.G.H., Fernandes, K.V.S. and Xavier-Filho, J. (1999) Jack bean seed coat contains a
protein with complete sequence homology to bovine insulin. Prot. Pept. Lett. 6:15 – 21.
OLIVEIRA, A.E.A.; Ribeiro, E.S.; Da Cunha, M.; Gomes, V.M.; Fernandes, K.V.S.; Xavier-
Filho, J. (2004). Insulin accelerates Canavalia ensiformis seeds germination and
development. Plant Growth Regul. 43: 57-62.
ORR, G. L., Srickland, J. A. and Walsh, T. A. (1994) Inhibitor of Diabrotica larval growth by
a multicystatin from potato tubers. J. Insect Physiol. 40: 893-900.
PERNAS, M., Lópes-Solanilla, E., Sánchez-Minge, R., Salcedo, G., Rodriguez-
Palenzuela, P. (1999) Antifungal activity of a plant cystatin. Mol. Plant-Microbe Inter. 12:
624-627.
RAO, M. R. G. and Kammannanauar, P. Y. (1990) Vigna vexillata: a source of desirable
characters for breeding mung bean. Current Res. 19: 169-171.
RICHARDSON, M. (1980) Protein inhibitors of enzymes. J. Food Chem. 6: 235-253.
Page 59
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 52
ROGELJ, B., Popovic, T., Ritonja, A., Strukelj, B. and Brzin J. (1998) Chelidocystatin, a
novel phytocystatin from Chelidonium majus. Phytochemistry.49: 1645-1649.
ROWAN, A. D., Brzin, J., Buttle, D. J. and Barret, A. J. (1990) Inhibitor of cysteine
proteinases by a protein inhibitor from potato. FEBS Let. 269: 328-330.
RYAN, C. A. (1973) Proteolytic Enzymes and their inhibitors in plants. Annu. Rev Plant
Physiol. 24: 173-196.
RYAN, C. A. and Walker-Simmons, M. (1981) Plant Proteinases. In: The biochemistry of
plants. P. K. Stumpf and E. I. Conn Eds. 6: 321-350.
RYAN, C. A. Proteinase inhibitor gene families: strategies for transformation to improve
plant defenses against herbivores. Bio Essays Vol. 1, Nº1, 1989.
SANTINO, A., Gallo, A., Leone, A., Poltronieri, P. and Zacheo G. (1998) Purification and
characterization of a novel papain inhibitor from commom bean (Phaseolus vulgaris) seed.
Physiol. Plant. 104: 293-298.
SATOH, S. (1998) Functions of the cell wall in the interactions of plant cells: analysis using
carrot cultured cells. Plant Cell Physiol. 39:361-368.
SCHAFFER, M. A. and Fisher, R. L. (1990) Transcriptional activation by heat and cold of a
thiol protease gene in tomato. Plant Physiol. 93: 1486-1491.
SHARMA, S., Rashid, F., Bano, B. (2006) Studies on low molecular mass phytocystatins
purified from Phaseolus mungo (Urd). Biochemistry 71: 406-413.
SIDDHURAJU, P., Vijayakumari K. and Janardhanan, K. (1994) Chemical analysis and
nutritional assessment of the less know pulses, Vigna aconitifolia (Jacq.) Maréchal and
Vigna vexillata (L) A. Rich. Plant Foods Hum. Nutr. 45: 103-111.
SILVA, L.B.; Santos, Azevedo, C.R.; Lopes, M.A.C.; Venâncio, T.M.; Cavalcante, C.P.;
Uchoa, A.F.; Astolfi-Filho, S.; Oliveira, A.E.A.; Fernandes, K.V.S.; Xavier-Filho, J. (2002).
Page 60
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 53
The leaves of green plants as well as cianobacterium, a red alga, and fungi contain insulin-
like antigens. Braz. J. Med. Biol. Res. 35: 297-303.
SIQUEIRA-JÚNIOR, C.L., Fernandes, K.V.S. Machado, O.L.T., Da Cunha, M., Gomes,
V.M., Jacinto, T. (2002) 87 kDa tomato cystatin exhibits properties of a defense protein
and forms protein crystals in prosystemin overexpressing transgenic plants. Plant Physiol.
Biochem., 40: 247-254.
SOARES-COSTA, A., Beltramini, L.M., Thiemann, O.H., Enrique-Silva, F. (2002) A
sugarcane cystatin: recombinant expression, purification and antifungal activity. Biochem.
Biophys. Res. Comm. 296: 1194–1199.
SONG, I., Taylor, M., Baker, K. and Bateman Jr., R. C. (1995) Inhibition of cysteine
proteinases by Carica papaya cystatin produced in Escherichia coli. Gene 162: 221-224.
SHYU, D. J. H., Chou, W. M., Viu, T. J., Lin, C. P. C., Tzen, J. T. C. (2004a) Cloning,
functional expression, and characterization of cystatin in sesame seed. J. Agric. Food
Chem. 52: 1350-1356.
SHYU, D. J. H., Chyan, C. L., Tzen, J. T. C., Chou, W. M. (2004b) Molecular cloning,
expression, and functional characterization of a cystatin from pineapple stem. Biosci.
Biotecnol. Biochem. 68 (8): 1681-1698
TOWBIN, H., Staehelin, T. and Gordon, J. (1979) Electroforetic transfer of proteins from
polyacrilamide gels to nitocellulose sheets: procedure and some applications. Proc. Natl.
Acad. Sci., USA, 76: 4350-4354
THOTTAPPILY, G., Ng, N.Q., Rossel, H.W. (1994) Screening germplasm of Vigna
vexillata for resistance to cowpea mottle virus. Int. J. Trop. Plant Diseas. 12: 75-80.
TULLY, R. E. and Beevers, H. (1978) Proteases and peptidases of castor bean
endosperm. Enzyme characterization and changes during germination. Plant Physiol. 62:
746-750.
Page 61
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 54
URWIN, P.E., Atkinson, H.J., Waller, D.A., McPherson, M.J. (1995) Engineered
oryzacystatin-I expressed in transgenic hairy roots confers resistence to Globodera pallida.
The Plant J., 8: 121-131.
URWIN, P. E., Lilley, C. J., Mcpherson, M. J. and Atkinson, H. J. (1997) Resistance to
both cystatin and root-knot nematodes conferred by transgenic Arabidopsis expressing a
modified plant cystatin. The Plant J. 12:455-461.
VANDERBORGHT, T. (1989) Some observations on seedlings of Vigna vexillata (L.) A.
Rich. (Fabaceae). Bull. Jard. Bot. Nat. Belg. 59: 179-187.
VAN DER VYVER, C, Schneidereit, J., Driscoll, S., Kurner, J., Kunert, K., Foyer, C.H.
(2003) Oryzacystatin I expression in transformed tobacco produces conditional growth
phenotype and enhances chilling tolerance. Plant Biotechn. J. 1: 101–112.
VENÂNCIO, T.M.; Oliveira, A.E.A.; Silva, L.B.; Machado, O.L.T.; Fernandes, K.V.S.;
Xavier-Filho, J. (2003). A protein with sequence homology to bovine insulin is found in the
legume Vigna unguiculata (cowpea). Braz. J. Med. Biol. Res. 36: 1167-73.
VILA, L., Quilis, J., Breitler, J.C., Marfà, V., Murillo, I., Vassal, J.M., Messeguer, J.,
Guiderdoni, E., San Segundo, B. (2005) Expression of the maize proteinase inhibitor (mpi)
gene in rice plants enhances resistance against the striped stem borer (Chilo
suppressalis): effects on larval growth and insect gut proteinases. Plant Biotechnol. J. 3:
2005. 187–202.
VILLANUEVA, J., Canals, F., Prat, S., Ludevid, D., Querol, E., Avilés, F.X. (1998)
Characterization of the wound-induced metallocarboxypeptidase inhibitor from potato.
FEBS Lett. 440: 175–182.
WÄLT, M., Roulin, S., Feller, U. (2002) Effects of pH, light and temperature on (1->3,1-
>4)-b-glucanase stability in wheat leaves, Plant Physiol. Biochem. 40: 363-371
Page 62
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 55
WALKER, A.J., Urwin, P.E., Atkinson, H.J., Brain, P.M., Glen, D.M., Shewry, P.R. (1999)
Transgenic Arabidopsis leaf tissue expressing a modified oryzacystatin shows resistance
to the field slug Deroceras reticulatum (Müller). Transg. Res., 8: 95-103.
WALKER-SIMMONS, M. and Ryan, C.A. (1977) Wound-induced accumulation of trypsin
inhibitor activities in plant leaves. Plant Physiol. 59: 437–439.
WALDRON, C., Wegrich, L. M,. Merlo, P. A. O. and Walsh, T. A. (1993) Characterization
of a genomic sequence coding for potato multicystatin, an eight-domain cysteine
proteinase inhibitor. Plant Mol. Biol. 23: 801-812.
WALSH, T. A. and Strickland, J. A. (1993) Proteolysis of the 85-kilodalton crystalline
cysteine proteinase inhibitor from potato releases functional cystatin domains. Plant
Physiol. 103: 1227-1234.
WATANABE, H., Abe, K., Emori, Y., Hosoyama, H. and Arai, S. (1991) Molecular cloning
and gibberellin-induced expression of multiple cysteine proteinases of rice seeds
(oryzains). J. Biol. Chem. 266:16897-16902.
XAVIER-FILHO, J. and CAMPOS, F. A. P. (1984) Inibidores de enzimas proteolíticas em
plantas. Arq. Biol. Tecnol 27: 407-17.
XAVIER-FILHO, J., Campos, F.A.P, Ary, M.B, Silva, C.P, Carvalho, M.M.M, Macedo,
M.L.R., Lemos, F.J.A. and Grant, G. (1989) Poor correlation between the levels of
proteinase inhibitors found in seeds of different cultivars of cowpea (Vigna unguiculata)
and the resistance/susceptibility to predation by Callosobruchus maculatus. J. Agric. Food
Chem. 37:1139-1143.
XAVIER-FILHO, J.; Oliveira, A.E.A.; Silva, L.B.; Azevedo, C.R.; Venâncio, T.M.; Machado,
O.L.T.; Oliva, M.L.; Fernandes, K. V.S.; Xavier-Neto, J. (2003). Plant insulin or glucokinin:
a conflicting issue. Braz. J. Plant. Physiol. Minireview. 15: 67-78.
Page 63
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 56
YANG, A.H., Yeh, K.W. (2005) Molecular cloning, recombinant gene expression, and
antifungal activity of cystatin from taro (Colocasia esculenta cv. Kaosiung no. 1), Planta
221: 493–501.
Page 64
Aspectos Fisiológicos e Bioquímicos de Cistatinas de Vigna vexillata 1