aDepartment of Nutrition and Physiology, Faculty of Home Economics, Kyoritsu Women's University, Tokyo 101-8433, JapanbDepartment of Medical Physiology, Graduate School of Medicine, Chiba University, Chiba 260-8670, JapancFaculty of Health Care, Kiryu University, Midori 379-2392, JapandNational Institute of Health and Nutrition, Tokyo 162-8636, Japan
A R T I C L E I N F O
⁎ Corresponding author at: Faculty of Health C48 9122.
Article history:Accepted 7 September 2011Available online 14 September 2011
Liver has a high regenerative capacity and restores its mass and function shortly afterpartial hepatectomy through increased proliferation and metabolic modification ofhepatocytes. The proliferation of hepatocytes can be triggered by its mass reduction afterhepatectomy or by the neural factors including lesioning of the ventromedialhypothalamus (VMH). In the present study, we examined the effect of VMH lesioning onliver regeneration in hepatectomized rats by evaluating liver function and morphology.We found that functional deficits caused by partial hepatectomy [prolonged prothrombintime (PT), increased indocyanine green (ICG) retention, and decrease in PAS (periodicAcid-Schiff staining)-positive hepatocytes] were restored by VMH lesioning at 1 week afterthe surgery, whereas these alterations disappeared at 4 weeks. Morphologically, lipidmicrodroplets, which are considered to be important for maintaining contiguous liver func-tion via supplying fuel for cell proliferation, were found to accumulate in hepatocytes of thehepatectomized rats at early period (1 day) after partial hepatectomy. Interestingly, suchlipid microdroplets were also detected in the VMH lesioned rats and the more abundantlyin the VMH lesioned, hepatectomized rats up to 1 week after the surgery. In conclusion,our results suggest that VMH lesioning in rats promotes recovery of liver anatomicallyand functionally after partial hepatectomy by promoting cell proliferation process.
83B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
1. Introduction
The liver is the largest organ in the body and plays an essentialrole in regulation of energy homeostasis, synthesis of mostsecreted proteins in the blood, excretion of bile acids requiredfor lipid absorption, and elimination of extrinsic or intrinsictoxic substances (Kuntz and Kuntz, 2008). Impairment of liverfunction leads to liver failure, which is a fatal condition if nottreated properly. A decline in liver function may be caused bydysfunction of existing hepatocytes or by a decrease in thenumber of functioning hepatocytes.
A common cause of loss of hepatocytes in human disease issurgical resection of part of the liver harboring malignanttumors. Although the loss of liver mass would cause attenua-tion of liver function, liver possesses a high regenerative capac-ity. In rodents, liver restores its mass within a week, even if 2/3of the liver is resected (Higgins and Anderson, 1931). Themech-anism of liver regeneration has been studied extensively. Liverregeneration is a sequential process involving various genesthat promote liver regeneration. These include hepatocytegrowth factor (HGF), ligands of the epidermal growth factorreceptor (EGFR) such as EGF, TGFα, and HB-EGF; tumor necrosisfactor (TNF); and interleukin-6 (IL-6) (Michalopoulos, 2007).
Despite the highly regenerative properties of hepatocytes,surgery is unsuccessful in some cases because of excessive liverloss due to massive hepatectomy (Michalopoulos, 2010). There-fore, amethod to promote liver regeneration is desirable. In addi-tion to the internal mechanism in the liver, proliferation ofhepatocytes is also regulated by organs outside the liver.We pre-viously reported that VMH lesioning increased DNA synthesismainly through cholinergic nerves (Kiba et al., 1992) and thatthe expression levels of genes involved in neural developmentand immune systemwere increased in the liver of VMH lesionedrats (Kiba et al., 2009). We also reported that regeneration of liverafter partial hepatectomywas facilitated by VMH lesioning alongwith increased DNA synthesis, as assessed by thymidine uptakein the residual liver (Kiba et al., 1994). However, the effects ofVMH lesioning on the function andmorphology of thehepatecto-mized liver have not been reported. In this study, we examinedfunctional parameters [serum albumin level, prothrombin time,and indocyanine green (ICG) retention rate], the morphology ofthe liver [stainingwithmethylene blue using epoxy resin embed-ded sectionsorwithperiodic acid-Schiff reaction (PAS) usingordi-nary paraffin sections], and proliferating cell nuclear antigen(PCNA), amarker of cell proliferation in rats subjected to simulta-neous VMH lesioning and/or partial hepatectomy. In order toobserve the accumulation of lipid microdroplets in the livers ofVMH lesioned rats, the rats were maintained under pair-feedingconditions to exclude the effect of hyperphagia caused by VMHlesioning. The morphological alterations were examined byboth light- and electron-microscopy.
2. Results
2.1. Changes in body weight after the surgery
The body weights in the 4 experimental groups before andafter the surgery (1 or 4 weeks) are shown in Fig. 1. The body
weights of sham VMH lesioned and sham hepatectomized(SV-SH, designated as controls) and sham VMH lesioned andhepatectomized (SV-H) rats returned to basal levels within aweek after the surgery, while those of VMH lesioned andsham hepatectomized (V-SH) (342.2±12.1 g, n=6), and VMHlesioned and hepatectomized (V-H) (330.6±8.9 g, n=6) rats at1 week after the surgery were significantly higher comparedwith SV-SH (279.4±1.5 g, n=6) and SV-H (274.1±8.1 g, n=6)rats, respectively (Fig. 1A). Four weeks after the surgery, thebody weights of the VMH lesioned rats (V-SH and V-H rats)were further increased compared with their counterparts(SV-SH and SV-H rats, respectively) (Fig. 1B). We alsomeasured food intake at 1 week after the surgery, and foundthat V-SH and V-H rats ate more than SV-SH and SV-H rats(Fig. 1C). These results are consistent with the previous re-ports that VMH lesioning caused obesity associated with hy-perphagia in animals (Inoue et al., 1977a; King, 2006).
2.2. Effects of VMH lesioning and partial hepatectomy onweights of liver and other tissues
Liver weights of the free feeding rats weremeasured at 1 weekafter the surgery. Although approximately 70% of the liver wasremoved by the hepatectomy, the liver weight of the SV-H ratshad recovered to 86% of that of the SV-SH rats within a weekafter the surgery (Fig. 2A). In V-SH rats, VMH lesioning in-creased the liver weight by 50% compared with that of SV-SHrats (Fig. 2A). Similarly, VMH lesioning in hepatectomizedrats had the same effect on liver weight, with a 64% increase(V-H vs. SV-H).
In addition to the effect on liver weight, VMH lesioningcaused a significant increase in the weight of the pancreasand stomach (Fig. 2B). However, we found only a marginalincrease in the weight of the heart or kidney and none for thelung. These results correspond to previous findings showingincreased weights of the pancreas and stomach as well asliver in VMH lesioned rats (Kiba et al., 1993, 1996) and mice(Suzuki et al., 2011).
2.3. Effect of VMH lesioning on liver function after partialhepatectomy
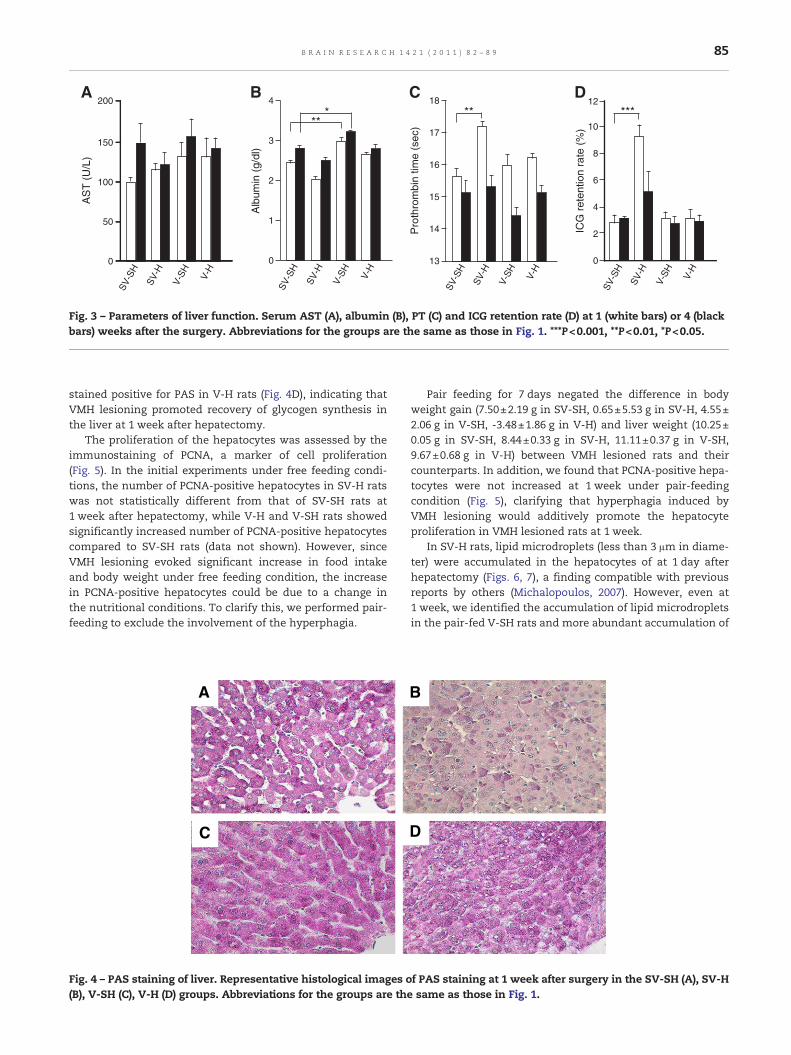
We next evaluated liver function at 1 and 4 weeks after thesurgery. There were no differences in serum aspartate amino-transferase (AST) among the 4 groups of rats (Fig. 3A). Thealbumin levels at 1 week tended to be lowered (by 16%) by hep-atectomy (2.44±0.07 g/dl in SV-SH; 2.04±0.09 g/dl in SV-H, n=6),but thedifferencewasnot significant. However, the serumalbu-min levels in V-SH rats (3.0±0.11 g/dl) at 1 week were signifi-cantly higher (P<0.01) than those in SV-SH rats, suggestingthat VMH lesioning increased serum albumin at this timepoint (Fig. 3B). At 4 weeks after the surgery, the same trendsfor serum albumin were present in the 4 groups (Fig. 3B).
We also examined prothrombin time (PT) to assess liverfunction. Hepatectomy exacerbated PT at 1 week after thesurgery (17.2±0.17 sec in SV-H rats vs. 15.6±0.27 sec in SV-SHrats, n=5, P<0.01) (Fig. 3C). In contrast, VMH lesioning alone didnot affect PT (16.0±0.35 sec in V-SH rats vs. 15.6±0.27 sec in SV-SH rats, not significant). However, VMH lesioning reversed theelongated PT caused by hepatectomy (16.2±0.18 sec in V-H rats;
A
100
200
300
400
Bod
y w
eigh
t (g)
**
SV-S
H
SV-H
V-SH V-
H0
**
SV-S
H
SV-H
V-SH V-
H
Bod
y w
eigh
t (g)
0
100
200
300
400
500
B
**
SV-S
HSV
-HV-
SH V-H
Foo
d in
take
(g/
day)
0
10
20
30
40
50C
Fig. 1 – Changes of body weight and food intake after surgery. (A, B) Body weights at baseline (before surgery) and 1 (A) and 4(B) weeks after surgery. White bars, before surgery; black bars, 1 (A) and 4 (B) weeks after surgery. * P <0.001. (C) Food intake at1 week after surgery. SV-SH, shamVMH lesioned and sham hepatectomized rats; SV-H, shamVMH lesioned and hepatectomizedrats; V-SH, VMH lesioned and sham hepatectomized rats; V-H, VMH lesioned and hepatectomized rats. *P<0.001.
84 B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
not significant vs. SV-SH rats), suggesting that VMH lesioningspecifically promotes recovery of coagulability. Four weeks afterthe surgery, PT returned to a normal level (that of SV-SH rats),even in SV-H rats. Therefore, PT did not differ significantlyamong the 4 groups at 4 weeks (Fig. 3C, black bars). The ICG reten-tion ratemarkedly increased in SV-H rats compared to SV-SH rats(controls) (P<0.001) at 1 week (Fig. 3D, white bars). Furthermore,VMH lesioning facilitated the restoration of ICG retention ratecaused by hepatectomy at 1 week (3.15±0.70% in V-H rats vs.9.31±0.93% in SV-H rats, n=5, P<0.001). The ICG retention rate4 weeks after surgery did not differ significantly among the 4groupsof rats, similar to the results of PT (Fig. 3D). Taken together,
SV-S
H
SV-H
V-SH V-
H
*****
0
5
10
15
20
Live
r w
eigh
t (g)
A
0
0.5
Panc
reas
**
Tis
sue
wei
ght (
g)
1
1.5
2
B
Fig. 2 – Weights of liver and other tissues. The weights of the liverAbbreviations for the groups are the same as those in Fig. 1. ThewhSV-H, V-SH and V-H groups, respectively. *** P<0.001, **P<0.01, *P
the data for PT and ICG retention rate show that VMH lesioninghas a stimulatory effect on the recovery of liver function afterpartial hepatectomy.
2.4. Histological examination of the liver
The ability of the liver to synthesize glycogen was evaluatedat 1 week after the surgery by PAS staining. In SV-SH rats,most hepatocytes stained positive for PAS (Fig. 4A), whilePAS-positive hepatocytes were rarely detected in SV-H rats(Fig. 4B). VMH lesioning alone did not affect the PAS stainingpattern as shown in V-SH rats (Fig. 4C), but most hepatocytes
Hea
rt
Kidn
ey
Stom
ach
***
Lung
******
(A) and other tissues (B) are shown at 1 week after surgery.ite, grey, black and hatched bars denoteweights in the SV-SH,
<0.05.
B
***
SV-S
H
SV-H
V-SH V-
H
0
1
2
3
4
Alb
umin
(g/
dl)
C***
SV-S
H
SV-H
V-SH V-
H
0
2
4
6
8
10
12
ICG
ret
entio
n ra
te (
%)
A
0
50
100
150
200
AS
T (
U/L
)
SV-S
H
SV-H
V-SH V-
H
SV-S
H
SV-H
V-SH V-
H
**
13
14
15
16
17
18
Pro
thro
mbi
n tim
e (s
ec)
D
Fig. 3 – Parameters of liver function. Serum AST (A), albumin (B), PT (C) and ICG retention rate (D) at 1 (white bars) or 4 (blackbars) weeks after the surgery. Abbreviations for the groups are the same as those in Fig. 1. ***P<0.001, **P<0.01, *P<0.05.
85B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
stained positive for PAS in V-H rats (Fig. 4D), indicating thatVMH lesioning promoted recovery of glycogen synthesis inthe liver at 1 week after hepatectomy.
The proliferation of the hepatocytes was assessed by theimmunostaining of PCNA, a marker of cell proliferation(Fig. 5). In the initial experiments under free feeding condi-tions, the number of PCNA-positive hepatocytes in SV-H ratswas not statistically different from that of SV-SH rats at1 week after hepatectomy, while V-H and V-SH rats showedsignificantly increased number of PCNA-positive hepatocytescompared to SV-SH rats (data not shown). However, sinceVMH lesioning evoked significant increase in food intakeand body weight under free feeding condition, the increasein PCNA-positive hepatocytes could be due to a change inthe nutritional conditions. To clarify this, we performed pair-feeding to exclude the involvement of the hyperphagia.
A
C
Fig. 4 – PAS staining of liver. Representative histological images o(B), V-SH (C), V-H (D) groups. Abbreviations for the groups are the
Pair feeding for 7 days negated the difference in bodyweight gain (7.50±2.19 g in SV-SH, 0.65±5.53 g in SV-H, 4.55±2.06 g in V-SH, -3.48±1.86 g in V-H) and liver weight (10.25±0.05 g in SV-SH, 8.44±0.33 g in SV-H, 11.11±0.37 g in V-SH,9.67±0.68 g in V-H) between VMH lesioned rats and theircounterparts. In addition, we found that PCNA-positive hepa-tocytes were not increased at 1 week under pair-feedingcondition (Fig. 5), clarifying that hyperphagia induced byVMH lesioning would additively promote the hepatocyteproliferation in VMH lesioned rats at 1 week.
In SV-H rats, lipid microdroplets (less than 3 μm in diame-ter) were accumulated in the hepatocytes of at 1 day afterhepatectomy (Figs. 6, 7), a finding compatible with previousreports by others (Michalopoulos, 2007). However, even at1 week, we identified the accumulation of lipid microdropletsin the pair-fed V-SH rats and more abundant accumulation of
B
D
f PAS staining at 1 week after surgery in the SV-SH (A), SV-Hsame as those in Fig. 1.
PC
NA
pos
itive
hep
atoc
yte
(%)
0
1
2
3
4
5
SV-SH SV-H V-HV-SH
EA B
C D
Fig. 5 – PCNA staining of liver. (A–D) Representative histological images of PCNA staining at 1 week after the surgery in the SV-SH(A), SV-H (B), V-SH (C), V-H (D) groups under pair-fed conditions. PCNA-positive hepatocytes are indicated by arrows. (E) Thenumber of PCNA-positive hepatocytes/1000 cells. Abbreviations for the groups are the same as those in Fig. 1.
86 B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
microdroplets and medium-sized droplets (approximately10 μm in diameter) in the pair-fed V-H rats, indicating thatVMH lesioning promoted the accumulation of lipid microdro-plets in regenerating liver.
3. Discussion
The VMH of the hypothalamus is known to be a metabolicsensor that regulates body fat mass and food intake. Whenthe VMH is ablated in experimental animals, the animalsdevelop marked obesity associated with hyperphagia (Inoueet al., 1977a; King, 2006). In accordance with the previousreports (Higgins and Anderson, 1931), we found that the liver
A
E F G
B C
Fig. 6 – Methylene blue staining of liver. Representative histologicthe surgery in the SV-SH (A, E), SV-H (B, F), V-SH (C, G) and V-H (
weight of SV-H rats recovered within a week after partial hep-atectomy (Fig. 2), suggesting a potent regenerative ability ofthe liver itself. VMH lesioning increased the liver weight inV-SH and V-H rats under free feeding conditions. In the pre-sent study, VMH lesioning exerted an additive effect on liverweight recovery after hepatectomy, and also increased theweights of the pancreas and stomach, but only slightly affectedthe heart, kidney or lung, suggesting that VMH lesioning withaccompanying hyperphagia increased cell proliferation inpancreas and stomach, as well as in liver (Kiba et al., 1992,1993, 1996; Suzuki et al., 2011).
After partial hepatectomy, increased proliferation of hepato-cytes occurs for restoring the liver mass. In this study, the liverweight in SV-H rats was comparable with that for SV-SH rats
H
D
al images of HE staining at 1 day (A–D) and 1 week (E–H) afterD, H) groups.
A B C D
Fig. 7 – Electron microscopic examination of liver. Representative ultrastructural images of electron microscopy at 1 day aftersurgery in the SV-SH (A), SV-H (B), V-SH (C) and V-H (D) groups. Arrows indicate lipid microdroplets. Bar, 2 μm.
87B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
at 1 week after partial hepatectomy (Fig. 2A). The increase inproliferation of hepatocytes after hepatectomy (assessed bythymidine incorporation) peaks at 36–48 hr (Chijiiwa et al.,1994). Although we previously observed increased PCNA stain-ing at 1 week after VMH lesioning in the previous (Kintakaet al., 2009; Suzuki et al., 2011) studies, our present study withpair-feeding suggested that hyperphagia caused byVMH lesion-ing exerts a synergistic effect with VMH lesioning for theincrease in PCNA staining at 1 week.
In the present study, we evaluated liver function in 4experimental groups at 1 and 4 weeks after the surgery, andfound an inconsistency between recovery of liver weight andthat of function, as assessed by serum levels of albumin,AST, PT, and ICG retention rates (Fig. 3). This indicates thatliver regeneration after hepatectomy seems to show a littledifferent time courses for recovery of function andmorphology.The serum albumin level was higher in V-H rats than in SV-Hrats, suggesting that the VMH lesioning may protect againsthypoalbuminemia due to loss of liver mass. However, thisassumption is unlikely because the serum albumin level wassimilarly higher in V-SH rats than in SV-SH rats. Alternatively,VMH lesioning and accompanying hyperphagia may increasethe liver weight and serum albumin level with or withoutconcurrent hepatectomy.
PT and activated thromboplastin time (aPTT) are goodmarkers for protein synthesis capacity after hepatectomy (Kimet al., 2010). Unlike the changes in serum albumin and AST, wefound that elongation of PT caused by hepatectomywas reversedby concurrent VMH lesioning (Fig. 3C), suggesting that VMHlesioning specifically promotes recovery of blood coagulability.We also found that VMH lesioning ameliorated increased ICGretention induced by hepatectomy (Fig. 3D). Since the ICG clear-ance test is widely performed before hepatectomy for estimatinghepatic functional reserve capacity and this parameter is a riskfactor for survival (Suda et al., 2008), these results show thatVMH lesioning has a stimulatory effect on recovery of liverfunction after partial hepatectomy.
We examined more specified morphological changes ofthe regenerating liver in rats with or without VMH lesioning,and found that glycogen synthesis (assessed by PAS staining)was not completely recovered at 1 week after hepatectomyalone. Since glycogen is a major source of glucose underfasted conditions, this result suggests that hepatocytes inSV-H rats had impaired capacity for carbohydrate metabolismat 1 week after hepatectomy. However, hepatocytes in V-Hrats stained positive for PAS at 1 week (Fig. 4), indicating thatVMH lesioning promoted recovery of glycogen synthesis in
the liver after hepatectomy. More interestingly, we identifiedmarked accumulation of lipid microdroplets in the hepato-cytes of V-H rats at 1 week after the surgery (Fig. 6H). Hepato-cytes have been reported to accumulate fat during liverregeneration after hepatectomy and this deposit of lipidmicrodroplets is now considered to be important for fulfillingboth hepatocyte proliferation essential for liver regenerationand maintenance of liver functions (Michalopoulos, 2007).The importance of this lipid deposition has been demonstrat-ed by several studies on liver regeneration. It has beenreported that the liver regeneration after partial hepatectomyis impaired by exogenous leptin administration (Shteyer et al.,2004), by liver specific glucocorticoid receptor disruption(Shteyer et al., 2004), or by genetic disruption of caveolin-1,all of which through the lack of lipid microdroplets accumula-tion (Fernández et al., 2006). Taken together, the markedincrease in lipid microdroplets in the hepatocytes of V-H ratsappears to be a desirable adaptive response for recovery ofliver morphology and function after hepatectomy. Thesemorphological features (the increase in PAS-positivehepatocytes and lipid microdroplets) in V-H rats are in accor-dancewith reversal of the functional properties of a prolongedPT and increased retention of ICG.
The functional relationship between the hypothalamus andthe liver has been examined in several studies. Obici et al.(2002) found that insulin signaling in the hypothalamus inhib-ited glucose production in the liver. The involvement of auto-nomic nervous system in the regulation was directly shown byPocai et al. (2005). They showed that activation of ATP-sensitiveK+ channels in the hypothalamus inhibits the gluconeogenesisin the liver through efferent vagal input to the liver. Inoue et al.(2006) showed that centrally-administrated insulin suppresseshepatic glucose production through IL-6/STAT3 signaling in theliver. All these results support the notion that the hypothalamusis the key regulator of energymetabolism in the liver. In additionto the role of the hypothalamus in maintaining energy homeo-stasis, the present study revealed that VMH lesioning promotesrecovery of synthesis of secretory proteins in the blood stream(such as prothrombin) and excretion of toxic substance (suchas ICG) and recovery of reduced glycogen storage in the liverafter partial hepatectomy in rats. Therefore, the VMH issuggested to play a critical role in regulating a variety of liverfunctions through the autonomic nervous system.
In theprevious studies,we found enhanced cell proliferationin visceral organs (liver, stomach, small intestine and pancreas)in VMH lesioned rats using [3H]-thymidine uptake (Kiba et al.,1992, 1993, 1996). We also found that the cell proliferation of
88 B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
these visceral organs in VMH lesioned rats was inhibitedcompletely by sub-diaphragmatic vagotomy and almostcompletely by administration of atropine, a cholinergic blocker,but not by elimination of hyperphagia via pair feeding or byelimination of hyperinsulinemia via administration of neutral-izing antibody against insulin, suggesting that cell proliferationof these visceral organs by VMH lesioning is derived from vagalhyperactivity (Kiba et al., 1992, 1993, 1996). Consequently, weinfer that a similar mechanism is involved in the liver regener-ation in VMH lesioned, hepatectomized rats.
In conclusion, VMH lesioning facilitates simultaneous res-toration of the liver morphologically and functionally afterpartial hepatectomy possibly through hyperactivity of para-sympathetic nerves induced by VMH lesioning. VMH lesioningmay provide a new tool for identifying factors involved inhepatocyte regeneration.
4. Experimental procedures
4.1. Animals
Female Sprague–Dawley (SD) rats aged 12–13 weeks andweighing about 250 g (Clea Japan Inc., Tokyo, Japan) werehoused in an air-conditioned (23±2 °C) room under an auto-matic lighting cycle (8:00 a.m. –8:00 p.m.). The animals weregiven free access to normal pellets (CE-2; Clea Japan Inc.)and drinking water. The rats were subjected to the studieswhen their weight reached 270–300 g. Female rats were cho-sen, because it is previously known that VMH lesioninginduces hyperphagic more markedly in female rats than inmales. For pair-feeding experiment, the food consumption ofVMH lesioned (V-SH and V-H) rats was adjusted so as tomatch with that of control (SV-SH).
4.2. Partial hepatectomy in rats
A70%hepatectomywas performedunder inhalation anesthesiawith isoflurane (Forane; Dinabot, Osaka, Japan). Aftermaking anupper midline incision, 70% of the liver was resected followingthe procedure established by Higgins and Anderson (1931). Theleft lateral lobe and median lobe of the liver were excised. TheVMH was lesioned immediately after hepatectomy.
4.3. VMH lesioning in rats
VMH lesioned rats were prepared following the method ofInoue et al. (1977b). The stereotaxic coordinates were as fol-lows: at the bregma anteriorly, 0.75 mm lateral to the mid-sagittal line, and 1.0 mm up (in the Z-axis) from the cranialbase. Sham VMH lesioned rats were similarly prepared withoutpassing a current. After the surgery, the rats were returned totheir cages and allowed free access to food and water. After allexperiments were completed, the area and extent of the hypo-thalamic lesions were examined histologically in all animals.
4.4. Experimental protocols
VMH lesioning or sham-VMH lesioning was performed in ratswith 70% hepatectomy or sham hepatectomy and a total of 4
groupswereprepared: shamVMH lesionedand shamhepatecto-mized (SV-SH) (control) group; shamVMH lesioned and hepatec-tomized (SV-H) group; VMH lesioned and sham hepatectomized(V-SH) group (each group, n=6) and VMH lesioned and hepatec-tomized (V-H) group (n=6). One or 4 weeks after the surgery,blood was drawn from femoral vein under anesthesia withisoflurane for measuring liver function parameters and an ICGtest was performed. At the end of the study, the rats wereeuthanized and the brain and liverwere sampled for histologicalexaminations. Tissueweights (pancreas, stomach, kidney, heart,and lung) were also measured. Pair-feeding experiments wereconducted in order to exclude the effects of hyperphagia usuallyseen in VMH lesioned animals (Inoue et al., 1977b; King, 2006;Suzuki et al., 2011) and examined the effect of VMH lesioningitself on morphological alterations in the liver.
4.5. Measurements
The serum level of AST, albumin, PT, and the ICG retention ratewere measured to assess liver function. Under anesthesia withisoflurane, 2 ml of blood was collected for measuring AST,albumin, and PT. This was followed by intravenous injectionof ICG, and blood was collected through femoral vein at15 min after ICG injection for calculating the ICG retentionrate. AST and albumin were measured using a Hitachi 7020autoanalyzer (Hitachi Co., Tokyo, Japan). PT was measured foroverall evaluation of blood coagulability. For measuring PT,100 μl of plasma was mixed with 200 μl of 11.6 mM CaCl2 withrabbit brain thromboplastin (Sigma-Aldrich, St. Louis, MO,USA) and clot formation was detected with a BBL fibrometer(Becton Dickinson, Cockeyville, MD, USA). ICG (0.25mg/100 g;Daiichi Sankyo Co., Tokyo, Japan) was diluted with physiologi-cal saline and injected into the femoral vein. The 15-minretention rate was adopted in this ICG test. The plasma ICGconcentration (the ICG retention rate 15min after femoralinjection, ICG R15) was determined using a spectrophotometerat 805 nm.
4.6. Histological examination
Tissue was fixed and subjected to histological examination forassessment of proliferation, function, and morphologicalstatus of the regenerated liver. PAS staining was performedfor evaluation of glycogen synthesis in hepatocytes. Morpho-logical examination by light microcopy was performedwith PCNA, a marker of cell proliferation, and methyleneblue staining using epoxy resin embedded sections, inwhich tissue samples were dissected and fixed in 2%paraformaldehyde/2.5% glutaraldehyde and post-fixed in 1%osmium tetroxide (that binds to unsaturated fatty acid andshows metachromasia of greenish yellow for lipid microdro-plets) and embedded in epoxy resin for preparation of 1.5 μmsections. Electronmicroscopy was performed under standard-ized methods.
4.7. Statistical analysis
All data are expressed as the mean±SEM. Comparisonbetween groups was made using one-way ANOVA followedby a Dunnett or Tukey test.
89B R A I N R E S E A R C H 1 4 2 1 ( 2 0 1 1 ) 8 2 – 8 9
Acknowledgments
This workwas supported in part by a Grant-in-Aid for ScientificResearch on Innovative Areas (3217) from the Ministry ofEducation, Science, Sports, Culture, and Technology.
Appendix A. Supplementary data
Supplementary data to this article can be found online atdoi:10.1016/j.brainres.2011.09.012.
R E F E R E N C E S
Chijiiwa, K., Nakano, K., Kameoka, N., Nagai, E., Tanaka, M., 1994.Proliferating cell nuclear antigen, plasma fibronectin, and liverregeneration rate after seventy percent hepatectomy innormal and cirrhotic rats. Surgery 116, 544–549.
Fernández, M.A., Albor, C., Ingelmo-Torres, M., Nixon, S.J.,Ferguson, C., Kurzchalia, T., Tebar, F., Enrich, C., Parton, R.G.,Pol, A., 2006. Caveolin-1 is essential for liver regeneration.Science 313, 1628–1632.
Higgins, G.M., Anderson, R.M., 1931. Experimental pathology ofthe liver. I; Restoration of the liver of the white rat followingpartial surgical removal. Arch. Pathol. 12, 186–202.
Inoue, H., Ogawa, W., Asakawa, A., Okamoto, Y., Nishizawa, A.,Matsumoto, M., Teshigawara, K., Matsuki, Y., Watanabe, E.,Hiramatsu, R., Notohara, K., Katayose, K., Okamura, H., Kahn,C.R., Noda, T., Takeda, K., Akira, S., Inui, A., Kasuga, M., 2006.Role of hepatic STAT3 in brain-insulin action on hepaticglucose production. Cell Metab. 3, 267–275.
Inoue, S., Mullen, Y.S., Bray, G.A., 1977a. Effect of transplantationof pancreas on development of hypothalamic obesity. Nature266, 742–744.
Inoue, S., Campfield, L.A., Bray, G.A., 1977b. Comparison ofmetabolic alterations in hypothalamic and high fatdiet-induced obesity. Am. J. Physiol. 233, R162–R168.
Kiba, T., Tanaka, K., Endo, O., Inoue, S., 1992. Role of vagus nervein increased DNA synthesis after hypothalamic ventromediallesions in rat liver. Am. J. Physiol. 262, G483–G487.
Kiba, T., Tanaka, K., Endo, O., Inoue, S., 1993. Ventromedialhypothalamic lesions increase gastrointestinal DNAsynthesis through vagus nerve in rats. Gastroenterology 104,475–484.
Kiba, T., Tanaka, K., Numata, K., Hoshino, M., Inoue, S., 1994.Facilitation of liver regeneration after partial hepatectomy by
ventromedial hypothalamic lesions in rats. Pflugers Arch. 428,6–9.
Kiba, T., Tanaka, K., Numata, K., Hoshino, M., Misugi, K., Inoue, S.,1996. Ventromedial hypothalamic lesion-induced vagalhyperactivity stimulates rat pancreatic cell proliferation.Gastroenterology 110, 885–893.
Kiba, T., Kintaka, Y., Suzuki, Y., Nakata, E., Ishigaki, Y., Inoue, S.,2009. Ventromedial hypothalamic lesions change theexpression of neuron-related genes and immune-relatedgenes in rat liver. Neurosci. Lett. 455, 14–16.
Kim, Y.K., Shin, W.J., Song, J.G., Jun, I.G., Kim, H.Y., Seong, S.H.,Sang, B.H., Hwang, G.S., 2010. Factors associated with changesin coagulation profiles after living donor hepatectomy.Transplant. Proc. 42, 2430–2435.
King, B.M., 2006. The rise, fall, and resurrection of theventromedial hypothalamus in the regulation of feedingbehavior and body weight. Physiol. Behav. 87, 221–244.
Kintaka, Y., Osaka, T., Zuzuki, Y., Hashiguchi, T., Niijima, A.,Kageyama, H., Takenoya, F., Shioda, S., Inoue, S., 2009. Effectsof gastric vagotomy on visceral cell proliferation induced byventromedial hypothalamic lesions: Role of vagalhyperactivity. J. Mol. Neurosci. 38, 243–249.
Kuntz, E., Kuntz, H.-D., 2008. Biochemistry and functions of theliver, In: Kuntz, E., Kuntz, H.-D. (Eds.), Hepatology-Textbookand Atlas, 3rd Edition. Springer Medizine Verlag, Wetzlar,Germany, pp. 35–76.
Michalopoulos, G.K., 2007. Liver regeneration. J. Cell. Physiol. 213,286–300.
Michalopoulos, G.K., 2010. Liver regeneration after partialhepatectomy: critical analysis of mechanistic dilemmas. Am. J.Pathol. 176, 2–13.
Pocai, A., Lam, T.K., Gutierrez-Juarez, R., Obici, S., Schwartz, G.J.,Bryan, J., Aguilar-Bryan, L., Rossetti, L., 2005. Hypothalamic K(ATP) channels control hepatic glucose production. Nature 434,1026–1031.
Shteyer, E., Liao, Y., Muglia, L.J., Hruz, P.W., Rudnick, D.A., 2004.Disruption of hepatic adipogenesis is associated with impairedliver regeneration in mice. Hepatology 40, 1322–1332.
Suda, K., Ohtsuka, M., Ambiru, S., Kimura, F., Shimizu, H.,Yoshidome, H., Miyazaki, M., 2008. Risk factors of liverdysfunction after extended hepatic resection in biliary tractmalignancies. Am. J. Surg. 197, 752–758.
Suzuki, Y., Inoue, S., Shimizu, H., Ishizuka, N., Kasahara, Y.,Takahashi, T., Arai, K., Kobayashi, Y., Kishi, M., Imazeki, N.,Senoo, A., Osaka, T., 2011. Cell proliferation in visceral organsinduced by ventromedial hypothalamic (VMH) lesions:development of electrical VMH lesions in mice and resultingpathophysiological profiles. Endocr. J. 58, 247–256.