Biological Roles for Recombination 1. Generating new gene/allele combinations (crossing over during meiosis) 2. Generating new genes (e.g., Immuno- globulin rearrangement) 3. Integration of a specific DNA element (or 4. virus) 5. DNA repair
Transcript
Biological Roles for Recombination
1. Generating new gene/allele combinations (crossing over during meiosis)
2. Generating new genes (e.g., Immuno- globulin rearrangement)
3. Integration of a specific DNA element (or
4. virus)
5. DNA repair
Why Recombination?
Two Broad Categories Homologous (or general) Site-specific (e.g. phage genomes into bacterial chromosomes)
Mutation happens - without recombination, mutation targetwould increase from gene to entire chromosome
Recombination allows favorable and unfavorable mutationsto be separated
Provides a means of escape, to generate new combinationsof genes, and spreading of favorable alleles
Homologous Recombination
Required for DNA replication, repairs accidents atreplication fork
Repairs double strand DNA (dsDNA) breaks
Occurs at meiosis (cross-overs)
Happens at “four strand” stage of meiosis, involvestwo of four strands
Occurs randomly between homologous sequences
Double strand break repair (DSBR) model
Allelic and non-allelic (ectopic recombination)
• Homologous recombination
• Nonhomologous recombination.
• Homologous recombination is based on sequence complementarity, and can be further divided into:
• generalized recombination
• site- specific recombination.
Generalized recombination refers to DNA exchange between molecules with extended sequence homology.
For example, transformation and conjugation between related bacterial strains.
Site-specific recombination refers to DNA recombination between molecules that shared limited regions of sequence homology.
For example, conjugation between two different species.
Two models have been proposed for generalized recombination:
a. Single stranded break model:
Recombination starts with the nick or break of homologous strands at a single correspondent
point. This breakage allows each free end to pair
with its complement in the other duplex).
B. The double stranded break model DNA exchange is initiated by an endonuclease-induced, double strand break in one moleculecalled recipient duplex.
The breakage is enlarged by the endonuclease to a gap with 3’ end free ends.
Nonhomologous recombination refers to the process by which a DNA molecule is incorporated into an unrelated genome with no sequence homology.
For exp., transposition of viral genome into human chromosome, or transposition.
Mechanisms of generalized recombination: Recombiantion involves breakage and rejoining of homologous DNA molecules. The intermediate stage during homologous recombination where the two homologous molecule covalent linked to each other is called Holiday Structure.
Examples of (mostly) Homologous Recombination Fig. 22.1
The Holliday model The Holliday model
Two homologous duplexes are alignedTwo homologous duplexes are aligned
Strand exchange leads to an intermediate Strand exchange leads to an intermediate with crossed strandswith crossed strands
This branch can move: Branch migrationThis branch can move: Branch migration
The branch is resolved by cleavage and The branch is resolved by cleavage and sealingsealing
R. Holliday (1964)
Holliday Model
- Holliday Junctions form during recombination
- HJs can be resolved 2 ways, only one produces true recombinant molecules
patch
The recBCD Pathway of Homologous Recombination
Part I: Nicking and Exchanging
recBCD Pathway of Homologous Recomb. Part I: Nicking and Exchanging
1. A nick is created in one strand by recBCD at a Chi sequence (GCTGGTGG), found every 5000
bp.
2. Unwinding of DNA containing Chi sequence by recBCD allows binding of SSB and recA.
3. recA promotes strand invasion into homologous DNA, displacing one strand.
4. The displaced strand base-pairs with the single strand left behind on the other chromosome.
5. The displaced and now paired strand is nicked (by recBCD?) to complete strand exchange.
Recombination is initiated by double-strand breaks in DNA
Double-stranded (dsDNA) breaks are not uncommon
Meiosis
Created by topoisomerase-like enzymes
Mitosis
Radiation
Mutagens (e.g. chemicals)
Stalled replication forks
Specialized endonucleases (eg site-specific HO endonuclease in switching of yeast matting type (MAT) genes)
Recombination requires DNA binding proteins
Extensively studied in model organisms, E. coli and yeast
Bacterial recombination enzymes identified by Rec - mutations
At least 25 proteins are involved in homologous recombination in E. coli
Remember four; RecBCD and RecA
3 member protein complex with endonuclease andhelicase activity
essential for 99% of recombination eventsoccurring at double-stranded breaks in bacteria
binds double stranded break
unwinds and degrades DNA
Pauses at chi sequence
Loads RecA on 3’ ssDNA extensions
RecBCD
Initiation of recombination by the RecBCD enzyme
RecA
Involved in SOS response; required for nearly ALL homologous
recombination in bacteria
Single-strand DNA binding protein, DNA dependent ATPase
Multiple DNA binding sites
Initiates the exchange of DNA between two recombining DNA double helixes
RecA enables single stranded DNAto invade DNA helix
Eukaryotes have multiple homologs of bacterial RecA
(Rad51 is best studied)
Chi site Χ
Recombination hotspot
Modifies RecBCD enzymatic activity
5’ GCTGGTGG 3’
1009 chi (Χ) sites in E. coli genome
Χ homologs in other bacteria
Targeted gene disruption by homologous recombination
Lodish et al. Molecular Cell Biology
Gene Conversion
A special type of homologous recombination
Non-reciprocal transfer of genetic material from a ‘donor’sequence to a highly homologous ‘acceptor’ sequence
Initiated by double strand DNA (dsDNA) breaks
5’ > 3’ exonucleases
3’ ssDNA tail strand invasion (RAD51 and others)
Outcome: portion of ‘donor’ sequence copied to ‘acceptor’and original ‘donor’ copy unchanged
donor acceptor
geneconversion
recBCD Pathway of Homologous Recom. Part II: Branch Migration and Resolution
1. Nicks are sealed Holliday Junction
2. Branch migration (ruvA + ruvB)
3. Resolution of Holliday Junction (ruvC)
RecBCD : A Complex Enzyme
• RecBCD has:
1. Endonuclease subunits (recBC) that cut one DNA strand close to Chi sequence.
2. DNA helicase activity (recD subunit) and
a DNA-dependent ATPase activity– unwinds DNA to generate the 3’ SS tails
RecA• 38 kDa protein that polymerizes onto SS DNA 5’-3’• Catalyzes strand exchange, also an ATPase• Also binds DS DNA, but not as strongly as SS
RecA binds preferentially to SS DNA and will catalyze invasion of a DS DNA molecule by a SS homologue.
Important for many types of homologous recombination, such as during meoisis (in yeast).
RecA Function Dissected
• 3 steps of strand exchange:1. Pre-synapsis: recA coats single-stranded
DNA (accelerated by SSB, so get more relaxed structure).
2. Synapsis: alignment of complementary sequences in SS and DS DNA (paranemic or side-by-side structure).
3. Post-synapsis or strand-exchange: SS DNA replaces the same strand in the duplex to form a new DS DNA (requires ATP hydrolysis).
Meiotic Recomb. in Yeast
- is initiated by a double-strand break (DSB)
Repair of double-strand breaks (DSBs)in non-dividing or mitotic cells
DSBs probably most severe form of DNA damage, can cause loss of genes or even cell death (apoptosis)
DSBs caused by:- ionizing radiation- certain chemicals- some enzymes (topoisomerases, endonucleases)- torsional stress
2 general ways to repair DSBs:
1. Homologous recombination (HR) - repair of broken DNA using the intact homologue, very similar to meiotic recombination. Very accurate.
2. Non-homologous end joining (NHEJ) - ligating non-homologous ends. Prone to errors, ends can be damaged before religation (genetic material lost) or get translocations. (Mechanism in Fig 20.38)
Usage: NHEJ >> HR in plants and animals
DNA Recompilation applications
What the is Recombinant DNA?
Recombinant DNA is what you get when you combine DNA from two different sources.
For example:Mouse + Human DNA
Human + Bacterial DNAViral + Bacterial DNA
Human + (other) Human DNA
Why Make Recombinant DNA?
Recombinant DNA Technology May Allow Us To:
• Cure or treat disease• Genetically modify our foods to increase flavor, yield, nutritional value or shelf-life• Better understand human genetics• Clone cells or organs
Molecular Biology’s Best Friends: Bacteria
Why use bacteria?
• They’re relatively simple organisms.• They reproduce very quickly and asexually (this means that the “daughter” cells will contain the exact same DNA as the “parent” cell).• It’s pretty easy to get DNA back into the bacteria after you’ve changed it.• We can mess around with their DNA and kill a lot of them during our experiments and nobody gets mad.
Insulin for Diabetics: The New Way
Step 1:
Isolate (find) the human gene responsible for producing insulinand decide where you want to put it.
In this case, we decide to put our human DNA into the plasmid of E. coli, a verycommon bacterium.
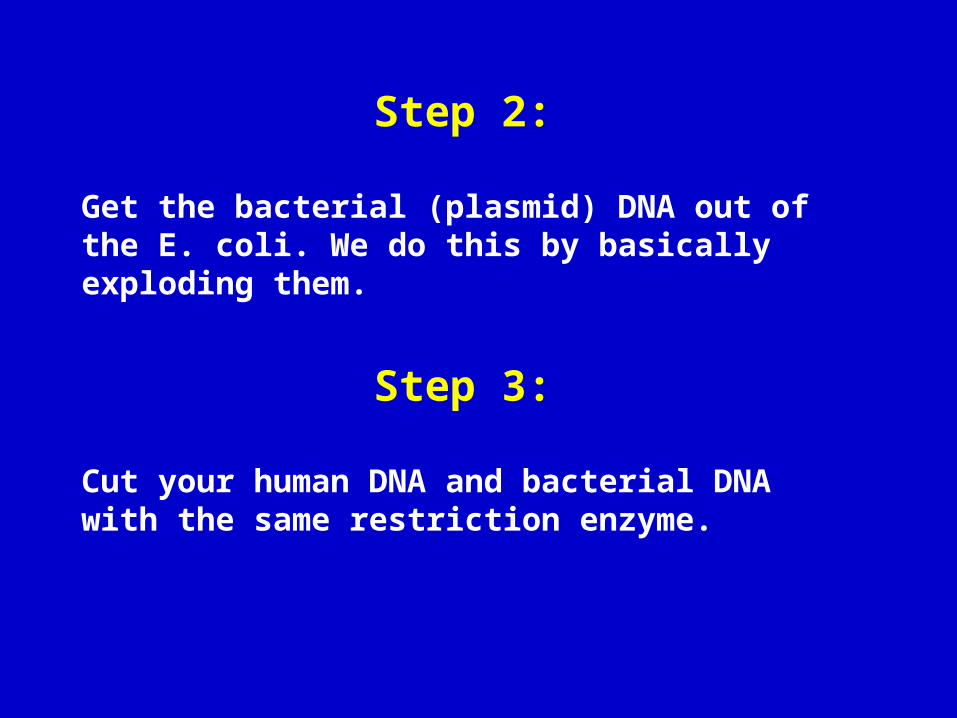
Step 2:
Get the bacterial (plasmid) DNA out of the E. coli. We do this by basically exploding them.
Step 3:
Cut your human DNA and bacterial DNA with the same restriction enzyme.
Step 4:
Mix the cut human DNA, which contains the insulin gene, with the cut bacterial DNA.
They’ll stick together because they were cut with the same restriction enzyme.
Step 5:
Get your new recombinant plasmid back into the bacteria.
This is easy because bacteria will take in DNA that’s floating around near them. We call this “transformation”.
Step 6
• Find the bacteria that have taken up your recombinant plasmid amongst the riff-raff in the petri dish.
Now your E. coli will use its new DNA to make human insulin!Because they reproduce so quickly, you’ll soon have thousands, millions, or billions of human insulin making machines.
By filtering out the bacteria after they’ve made insulin, you’ve got clean human insulin that can be packaged and given to diabetic patients.



























































![Thymoglobulin (anti-thymocyte globulin [rabbit]) · 2020. 12. 14. · DESCRIPTION . Thymoglobulin® (Anti-thymocyte globulin [rabbit]) is a purified, pasteurized, gamma immune globulin](https://static.documents.pub/doc/80x56/60c2dece3812e518472963b9/thymoglobulin-anti-thymocyte-globulin-rabbit-2020-12-14-description-thymoglobulin.jpg)



