Original Research Article BMP-2 modulates expression of other growth factors in a rat fracture healing model Jessica A. Cottrell a,1 , Owen Keane b , Sheldon S. Lin c , J. Patrick O'Connor a, * a Department of Biochemistry & Molecular Biology, Rutgers University, New Jersey Medical School and Graduate School of Biomedical Sciences, Newark, NJ, United States b Trinity College, University of Dublin, Dublin, Ireland c Department of Orthopaedics, Rutgers University, New Jersey Medical School and Graduate School of Biomedical Sciences, Newark, NJ, United States Introduction The osteo-inductive properties of bone morphogenetic pro- tein-2 (BMP-2) are well described (Urist, 1965, Rosen, 2009). Use of recombinant human BMP-2 (rhBMP-2) to promote spinal arthrodesis or fracture repair is an established therapeutic strategy (Lo et al., 2012). Other growth factors also have osteo-inductive or osteogenic properties and can be used to enhance or aid bone regeneration, such as BMP-7 (alternative name is OP-1), PTH (parathyroid hormone), FGF-2 (basic fibroblast growth factor), PDGF (platelet derived growth factor), CTGF (connective tissue growth factor), VEGF (vascular endo- thelial growth factor), HGF (hepatocyte growth factor), and TGF- b 3 (transforming growth factor b 3 ) (Kawaguchi et al., 2001; j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x a r t i c l e i n f o Article history: Received 6 June 2013 Received in revised form 14 October 2013 Accepted 17 October 2013 Available online xxx Keywords: Fracture healing Recombinant human bone morphogenetic protein-2 Growth factors Rat RT-qPCR a b s t r a c t Recombinant human bone morphogenetic protein-2 (rhBMP-2) is an osteo-inductive reagent that can be used therapeutically to promote spinal arthrodesis and fracture healing. Despite its clinical use, the effect of rhBMP-2 on expression of endogenous osteogenic growth factors is poorly described. The goal of this study was to determine if local rhBMP-2 treatment modulates expression of pro-osteogenic growth factors during fracture healing. Rat femur fractures were wrapped with control or rhBMP-2 collagen soaked sponge. Radiographic observations demon- strated that rhBMP-2 treatment enhanced fracture callus formation and decreased bridging time. Expression of fracture site growth factor mRNAs was determined by quantitative poly- merase chain reaction. The gene expression data demonstrated that rhBMP-2 treatment significantly increased mRNA levels of BMP-2 at day 4, BMP-4 at days 2, 4, 7, and 10, BMP-7 at day 2, and TNF-a at day 7. Treatment with rhBMP-2 was shown to significantly reduce mRNA levels of BMP-2 on day 2 and 10, TNF-a on day 14, and BMP-6 on days 2, 10, 14, and 21. TGFb-1 was also significantly reduced on days 2 and 21. In conclusion, treatment with rhBMP-2 appears to enhance expression of other osteogenic factors that could contribute to bone formation. # 2014 Faculty of Health and Social Studies, University of South Bohemia in Ceske Budejovice. Published by Elsevier Urban & Partner Sp. z o.o. All rights reserved. * Corresponding author at: Department of Biochemistry & Molecular Biology, Rutgers University-NJMS, MSB E659, 185 South Orange Avenue, Newark, NJ 07103, United States. Tel.: +1 973 972 5011; fax: +1 973 972 5294. E-mail address: [email protected](J.P. O'Connor). 1 Current Address: Department of Biological Sciences, Seton Hall University, School of Arts and Science, South Orange, NJ, United States. JAB-24; No. of Pages 9 Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates expression of other growth factors in a rat fracture healing model. J. Appl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003 Available online at www.sciencedirect.com ScienceDirect journal homepage: http://www.elsevier.com/locate/jab 1214-021X/$ – see front matter # 2014 Faculty of Health and Social Studies, University of South Bohemia in Ceske Budejovice. Published by Elsevier Urban & Partner Sp. z o.o. All rights reserved. http://dx.doi.org/10.1016/j.jab.2014.02.003
Transcript
JAB-24; No. of Pages 9
Original Research Article
BMP-2 modulates expression of other growth factors in a ratfracture healing model
Jessica A. Cottrell a,1, Owen Keane b, Sheldon S. Lin c, J. Patrick O'Connor a,*aDepartment of Biochemistry & Molecular Biology, Rutgers University, New Jersey Medical School and Graduate School of Biomedical Sciences,Newark, NJ, United StatesbTrinity College, University of Dublin, Dublin, IrelandcDepartment of Orthopaedics, Rutgers University, New Jersey Medical School and Graduate School of Biomedical Sciences, Newark, NJ,United States
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x
a r t i c l e i n f o
Article history:
Received 6 June 2013
Received in revised form
14 October 2013
Accepted 17 October 2013
Available online xxx
Keywords:
Fracture healing
Recombinant human bone
morphogenetic protein-2
Growth factors
Rat
RT-qPCR
a b s t r a c t
Recombinant human bone morphogenetic protein-2 (rhBMP-2) is an osteo-inductive reagent
that can be used therapeutically to promote spinal arthrodesis and fracture healing. Despite its
clinical use, the effect of rhBMP-2 on expression of endogenous osteogenic growth factors is
poorly described. The goal of this study was to determine if local rhBMP-2 treatment modulates
expression of pro-osteogenic growth factors during fracture healing. Rat femur fractures were
wrapped with control or rhBMP-2 collagen soaked sponge. Radiographic observations demon-
strated that rhBMP-2 treatment enhanced fracture callus formation and decreased bridging
time. Expression of fracture site growth factor mRNAs was determined by quantitative poly-
merase chain reaction. The gene expression data demonstrated that rhBMP-2 treatment
significantly increased mRNA levels of BMP-2 at day 4, BMP-4 at days 2, 4, 7, and 10, BMP-7 at
day 2, and TNF-a at day 7. Treatment with rhBMP-2 was shown to significantly reduce mRNA
levelsof BMP-2 onday2 and10,TNF-a on day 14,and BMP-6 ondays 2, 10, 14,and 21.TGFb-1 was
also significantly reduced on days 2 and 21. In conclusion, treatment with rhBMP-2 appears to
enhance expression of other osteogenic factors that could contribute to bone formation.
# 2014 Faculty of Health and Social Studies, University of South Bohemia in Ceske
Budejovice. Published by Elsevier Urban & Partner Sp. z o.o. All rights reserved.
The osteo-inductive properties of bone morphogenetic pro-tein-2 (BMP-2) are well described (Urist, 1965, Rosen, 2009). Useof recombinant human BMP-2 (rhBMP-2) to promote spinalarthrodesis or fracture repair is an established therapeuticstrategy (Lo et al., 2012). Other growth factors also have
* Corresponding author at: Department of Biochemistry & MolecularAvenue, Newark, NJ 07103, United States. Tel.: +1 973 972 5011; fax: +
E-mail address: [email protected] (J.P. O'Connor).1 Current Address: Department of Biological Sciences, Seton Hall Univ
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates eAppl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003
1214-021X/$ – see front matter # 2014 Faculty of Health and Social Stuby Elsevier Urban & Partner Sp. z o.o. All rights reserved.http://dx.doi.org/10.1016/j.jab.2014.02.003
osteo-inductive or osteogenic properties and can be used toenhance or aid bone regeneration, such as BMP-7 (alternativename is OP-1), PTH (parathyroid hormone), FGF-2 (basicfibroblast growth factor), PDGF (platelet derived growth factor),CTGF (connective tissue growth factor), VEGF (vascular endo-thelial growth factor), HGF (hepatocyte growth factor), and TGF-b3 (transforming growth factor b3) (Kawaguchi et al., 2001;
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x2
JAB-24; No. of Pages 9
Gandhi et al., 2004; Alkhiary et al., 2005; Li et al., 2007; Manabeet al., 2007; Barnes et al., 2008; Hollinger et al., 2008a,b; Anituaet al., 2012). This has lead to testing mixtures of growth factors asspinal fusion or fracture healing treatments. For instance, theefficacy of platelet-rich plasma to treat fractures is thought tooccur by release of multiple growth factors following plateletdegranulation (Wrotniak et al., 2007). In addition, the osteo-inductive effects of demineralized bone matrix (DBM) productsare thought to occur by release of multiple growth factors,including BMPs, from the bone matrix (Bae et al., 2006). Thus, it isreasonable to hypothesize that increasing endogenous expres-sion of multiple osteogenic growth factors would betterstimulate bone formation and healing.
The effect of rhBMP-2 on expression of endogenousosteogenic growth factors is poorly understood. Gene expres-sion data obtained from a rabbit spinal fusion model showedthat the expression of BMP-6 increased in fusion beds treatedwith rhBMP-2 (Morone et al., 1998). This one study suggests thatapplication of a single growth factor, rhBMP-2, can induceexpression of other growth factors that have been associatedwith bone formation. Many in vitro studies have shown thatBMP-2 can induce expression of genes involved in osteogenesis.For example, Bais et al. (2009) demonstrated that inhibition ofBMP-2 expression with lentiviral BMP-2 shRNA prevented bothmineralized nodule formation in vitro and bone formation in vivoand blocked the expression of Runx2 and Osterix, which aretranscription factors necessary for terminal osteogenic differ-entiation. However, these studies have not focused on theexpression of other osteogenic factors. Use of additional growthfactors in combination with rhBMP-2 may not be necessary toachieve superior clinical efficacy if BMP-2 treatment will induceendogenous expression of these same factors. To test thishypothesis, the expression of osteogenesis related growthfactors during fracture repair was compared between controlfracture calluses and fracture calluses treated with rhBMP-2.The data indicate that rhBMP-2 treatment alters the expressionof other growth factors and promotes BMP-4 expression.
Materials and methods
Animals
All animal procedures were reviewed and approved by theUniversity's Institutional Animal Care and Use Committee andmet or exceeded all federal guidelines for the humane use of
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates
animals in research. Female Sprague–Dawley rats were fed astandard diet and kept caged in pairs in a constant tempera-ture and humidity environment. All rats were between 250 and300 g and approximately 12 weeks old at the beginning of theexperiment. A total of 74 female Sprague–Dawley rats wereused in this study (Table 1). One rat died post-operatively andis not included in the study. Thus specimens from 73 rats wereused in the final gene expression analysis (average sacrificeweight 304 � 27 g).
Surgical procedure/fracture model
A standard, closed femur fracture model was employed toproduce unilateral fractures as modified for female Sprague–Dawley rats (Bonnarens and Einhorn, 1984; Bergenstock et al.,2005). Briefly, rats were anesthetized by intraperitoneal injec-tion of ketamine (50 mg/kg) and xylazine (10 mg/kg). Underaseptic conditions, a medial parapatellar incision (approxi-mately 1 cm) was made in the right hind limb and the patellawas dislocated laterally. The medullary canal was enteredthrough the intercondylar notch and reamed with a 21 gaugeneedle. A 0.71 mm diameter stainless steel pin was theninserted into the canal and was secured in the greatertrochanter by tamping. The distal portion of the pin was cutflush within the intercondylar notch and the patella reduced.The soft tissue and skin were closed in two layers usingresorbable sutures. The diaphysis of the pinned femur wasfractured immediately using a 3-point bending device. Thefracture site was immediately exposed through a lateral incisionand treated with a direct onlay of clinically used and FDAapproved absorbable collagen sponge (ACS, 1 cm � 1 cm �0.35 cm) soaked with 0.1 ml of saline (control group) or with0.1 ml rhBMP-2 solution at a concentration of 0.2 mg/ml (rhBMP-2 treatment group) to provide a nominal 20 mg rhBMP-2 dose.The animals were caged in pairs and allowed to ambulate freelypost-surgery. Animals were euthanized by CO2 inhalation.Femurs were dissected free and processed for quantitative realtime PCR. Previous studies have demonstrated that 6 samplesfrom each treatment group are required at each time point toidentify significant variation in gene expression (Balaburski andO'Connor, 2003). Thus at least 6 rats were used for each timepoint. Time points included 2, 4, 7, 10, 14, and 21 days afterfracture which span the early phase of healing when cellmigration and proliferation predominate and the subsequentregenerative phase when chondrocyte differentiation andendochondral ossification occurs.
expression of other growth factors in a rat fracture healing model. J.
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x 3
JAB-24; No. of Pages 9
Radiography
Immediately following surgery, radiographs were made toverify the position and quality of each fracture. Radiographswere made using a Packard Faxitron (McMinnville, OR) andKodak MinR-2000 mammography film (Eastman Kodak Co.,Rochester, NY) while rats were under anesthesia. Additionalradiographs were taken before sacrifice at each time point todetermine the degree of healing. Radiographs taken at 21 dayswere scored to identify degree of healing as describedpreviously (Bergenstock et al., 2005). In brief, the radiographswere scored 0 (non-union), 1, 2, 3, or 4 (complete union) anddata between treatments were compared using the Mann–Whitney Rank Sum Test (SigmaPlot version 12.5, SystatSoftware, Inc., San Jose, CA).
RNA preparation
Rats were euthanized at each time point, the femur quicklyresected, the proximal and distal ends removed to isolate thefracture callus, and then the callus was flash frozen in liquidnitrogen. Total RNA was purified from each callus aspreviously described (Balaburski and O'Connor, 2003). Briefly,the callus was pulverized in a liquid nitrogen cooled appara-tus, extracted with TriZol reagent, and then further purifiedusing a Qiagen RNeasy kit. An aliquot of total RNA wasseparated by agarose gel electrophoresis and stained withethidium bromide to assess RNA quality (intact rRNA). OnlyRNAs with acceptable characteristics including intact 18S and28S ribosomal RNA were used for further analysis (Table 1).
cDNA for each sample was prepared using 1 mg aliquots oftotal RNA as described previously using an oligo(dT)20 primer,RNase inhibitor (SuperaseIn, Ambion Inc., Austin, TX), and M-MLV reverse transcriptase (New England Biolabs, Beverly, MA)(Balaburski and O'Connor, 2003). Reactions were allowed toproceed for 1 h at 42 8C and were stopped by incubation at 92 8Cfor 10 min. cDNA was stored at �20 8C until use. The cDNAfrom each callus specimen was tested for suitability by PCRamplification of GAPDH cDNA and visualization of the PCRproduct by agarose gel electrophoresis and ethidium bromidestaining. This procedure was repeated for each sample until acDNA preparation suitable for the subsequent qPCR analysiswas obtained.
RT-qPCR procedures
Quantitative polymerase chain reaction (qPCR) was performedusing the Absolute QPCR SYBR Green mix (ABgene, Rochester,
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates eAppl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003
NY) and an Applied Biosystems 7500 Real-Time PCR System(Foster City, CA). The target mRNAs that were quantified in thisstudy are listed in Table 2. Reactions contained an aliquot ofcDNA corresponding to 1.25 ng of total RNA for GAPDH or6.25 ng of total RNA for all other target mRNAs in a 25 mlreaction volume. The increase in SYBR green fluorescence wasmeasured over 40 cycles of PCR. Reactions were performed intriplicate for each mRNA target and for each sample. Thequality of each qPCR reaction was assessed by a post-amplification melting step. Reactions that failed were repeatedat least once. Samples in which one or more of the qPCRreactions repeatedly failed were removed from the study.
Data analysis
For each qPCR reaction, the threshold cycle (Ct) wasautomatically determined using the Applied Biosystemssoftware. All amplification curves were manually reviewedto insure that calculated Ct values appeared appropriate. MeanCt values were calculated using the supplied Applied Biosys-tems software. The corresponding qPCR Ct value for GAPDHwas subtracted from the target mRNA Ct values for thatspecimen to obtain the DCt value. Since there is an inverserelationship between DCt values and expression, the meaninverse DCt values (1/DCt value) were plotted over time forclarity. Gene expression comparisons were made betweentreatment groups across time points by utilizing the DCt valuesand two-way ANOVA with parametric post hoc tests.
Results
Disposition of animals and femur samples
Table 1 summarizes the experimental design. Fracture callusspecimens were successfully harvested from 73 rats. RNApreparations from 68 of the 73 specimens (93%) were ofsatisfactory quality for RT-qPCR analysis.
Radiographic observations
Radio-opaque fracture calluses were evident in the control andrhBMP-2 treatment groups by day 7 (Fig. 1). However, the radio-opaque callus in the rhBMP-2 treatment groups on days 7 (J)and 10 (K) were larger than their matched controls (D and E).Additionally, the fracture line in the rhBMP-2 treatment groupwas resolved by day 21(L) unlike the control treatment group(F). Treatment with rhBMP-2 at the fracture site appeared to
xpression of other growth factors in a rat fracture healing model. J.
Fig. 1 – Radiographic time course of fractures. (A–F) Radiographs from control-treated rats at 2, 4, 7, 10, 14, and 21 days afterfracture. (G–L) Radiographs from rhBMP-2-treated rats at 2, 4, 7, 10, 14, and 21 days after fracture.
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x4
JAB-24; No. of Pages 9
enhance fracture callus development and bridging. Scoring ofthe 21 day radiographs using a 0–4 point scale indicated thatrhBMP-2 treatment significantly enhanced bone healing(P = 0.016). The rhBMP-2 treated specimens had a meanradiographic score of 3.3 (0.7 standard deviation) as comparedto 0.6 (0.8 standard deviation) for the control treated speci-mens.
RT-qPCR analysis of osteogenic growth factor mRNA levels
Growth factor mRNA levels in the fracture calluses of thecontrol and rhBMP-2 treatment groups were compared overthe course of healing by RT-qPCR. mRNA levels werenormalized to GAPDH mRNA levels. GAPDH mRNA RT-qPCRCt values were similar between control and rhBMP-2 treatedsamples (Table 3). Growth factor mRNA DCt values werecompared within the control-treated rats using one-wayANOVA and post hoc Fisher tests to identify changes ingrowth factor mRNA levels with healing time. To identifyeffects of exogenous rhBMP-2 treatment on growth factormRNAs levels, DCt values were compared between treatment
Table 3 – GAPDH expression in control and BMP-2 treatedfractures (Ct values).
groups and time after fracture using two-way ANOVA analysisand post hoc Holm–Sidak tests to isolate specific differences.Mean DCt values and two-way ANOVA P values are reported inTables 4 and 5.
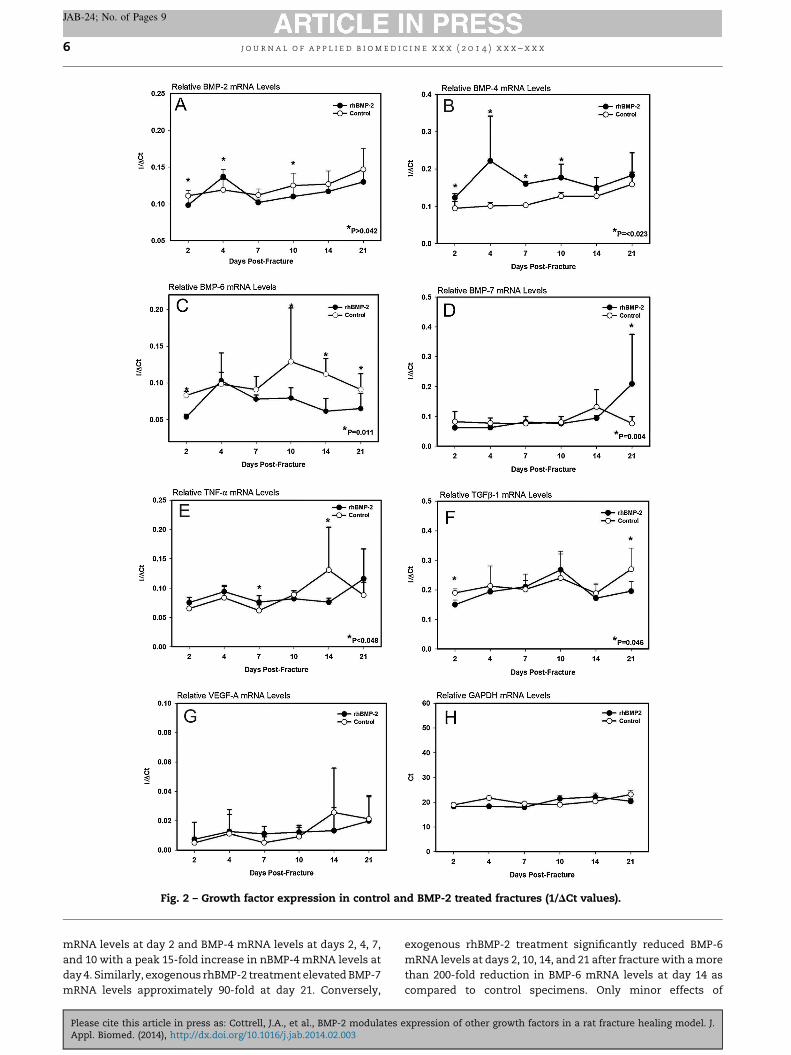
Only mRNA levels for BMP-2, BMP-4, and TNF-a varied withtime after fracture in the fracture calluses of the control-treated rats. BMP-2 and BMP-4 mRNA levels peaked at day 21(Fig. 2). The day 21 BMP-2 mRNA levels were significantlyhigher than the 2, 4, or 7 day time point levels (P = 0.002, 0.017,and 0.002, respectively) and was approximately 4-fold higherthan at day 2. BMP-4 mRNA levels at days 10, 14, and 21 weresignificantly higher than at days 2, 4, or 7 (P < 0.025 for allcomparisons) and were approximately 20-fold greater at day21 than at day 2. TNF-a mRNA levels were significantly lower atday 2 as compared to days 10, 14, and 21 (P = 0.021, 0.005, and0.048, respectively) and at day 7 as compared to days 4, 10, 14,and 21 (P < 0.033 for all comparisons). There was approxi-mately 13-fold more TNF-a mRNA in the fracture callus at day21 as compared to day 2. The increase in BMP-2, BMP-4 andTNF-a mRNA levels with time after fracture corresponds withthe period of maximal bone formation and osteoclast-mediated calcified cartilage resorption and callus bonemodeling. BMP-6 mRNA levels also appeared to be highestat days 10 and 14 after fracture and BMP-7 mRNA levelsappeared highest at day 14, but these increases were notsignificant. Surprisingly, TGF-b1 mRNA levels did not changeeven during the chondrogenic phase (days 7–14) of fracturehealing. We expected VEGF-A mRNA levels to increase in thelater stages of healing since angiogenesis is essential forfracture healing (Hausman et al., 2001). Though VEGF-A mRNAlevels were highest at days 14 and 21 after fracture, theincreased levels were not significant.
Exogenous rhBMP-2 treatment decreased callus BMP-2mRNA levels on days 2 and 7 after fracture (Table 4). Incontrast, exogenous rhBMP-2 treatment elevated BMP-2
expression of other growth factors in a rat fracture healing model. J.
Table 4 – BMP mRNA levels in control and rhBMP-2 treated fractures. Italicized P values indicate mean DCt values forcontrol-treated samples that were significantly higher (reduced mRNA levels) than rhBMP-2 treated samples. Bolded Pvalues indicate mean DCt values for control-treated samples that were significantly lower (increased mRNA levels) thanrhBMP-2 treated samples.
mRNA and days after fracture Control treated DCt rhBMP-2 treated DCt Holm–Sidak
Table 5 – Additional growth factor mRNA levels in control and rhBMP-2 treated fractures. Italicized P values indicate meanDCt values for control-treated samples that were significantly higher (reduced mRNA levels) than rhBMP-2 treated samples.Bolded P values indicate mean DCt values for control-treated samples that were significantly lower (increased mRNA levels)than rhBMP-2 treated samples.
mRNA and days after fracture Control treated DCt rhBMP-2 treated DCt Holm–Sidak
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x 5
JAB-24; No. of Pages 9
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates expression of other growth factors in a rat fracture healing model. J.Appl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003
Fig. 2 – Growth factor expression in control and BMP-2 treated fractures (1/DCt values).
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x6
JAB-24; No. of Pages 9
mRNA levels at day 2 and BMP-4 mRNA levels at days 2, 4, 7,and 10 with a peak 15-fold increase in nBMP-4 mRNA levels atday 4. Similarly, exogenous rhBMP-2 treatment elevated BMP-7mRNA levels approximately 90-fold at day 21. Conversely,
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates
exogenous rhBMP-2 treatment significantly reduced BMP-6mRNA levels at days 2, 10, 14, and 21 after fracture with a morethan 200-fold reduction in BMP-6 mRNA levels at day 14 ascompared to control specimens. Only minor effects of
expression of other growth factors in a rat fracture healing model. J.
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x 7
JAB-24; No. of Pages 9
exogenous rhBMP-2 treatment on TGF-b1 and TNF-a mRNAlevels were detected (Table 5). There were approximately 2-fold decreases in TGF-b1 mRNA levels at days 2 and 21 ascompared to control specimens. TNF-a mRNA levels were 8-fold higher at day 7 in the rhBMP-2 treated specimens but were11-fold lower on day 14. No effect of rhBMP-2 treatment onVEGF-A mRNA levels was noted.
Discussion
Exogenous rhBMP-2 treatment induces bone formation asshown in numerous pre-clinical and clinical studies (Axelradand Einhorn, 2009). The osteo-inductive effects of rhBMP-2may include regulating expression of other growth factorsimportant for osteogenesis, chondrogenesis, and angiogenesisin order to promote osteogenesis over other differentiation orhomeostatic pathways. The present study supports thisconcept in that exogenous rhBMP-2 treatment of rat femurfractures significantly affected the expression of differentgrowth factors throughout the first 21 days of fracture healing,which encompasses the regenerative phase of healing.Specifically, exogenous rhBMP-2 treatment significantly in-creased mRNA levels for BMP-2 on day 4, BMP-4 on days 2through 10, and BMP-7 on day 21 after fracture. In contrast,exogenous rhBMP-2 treatment reduced BMP-6 mRNA levels ondays 2, 10, 14, and 21 and TGF-b1mRNA levels on days 2 and 21after fracture. No effect of exogenous rhBMP-2 treatment onVEGF-A mRNA levels was found though TNF-a mRNA levelsvaried from control values at 7 and 14 days after fracture. Thusexogenous rhBMP-2 treatment enhanced the expression of 3osteogenic growth factors with apparently lesser effects onmRNA levels for chondrogenic TGF-b1, angiogenic VEGF-A,and inflammatory TNF-a.
The effects of rhBMP-2 treatment appear to be complex inthat rhBMP-2 treatment caused temporal elevations andreductions in osteogenic growth factor mRNA levels. Compli-cating any interpretation of these data is that rhBMP-2released from the collagen sponge to the fracture site isexpected to occur by an initial burst release followed by alogarithmic release of rhBMP-2 over 10 days with less than 5%remaining after 10 days based upon in vivo testing in rats(Friess et al., 1999). A similar burst release profile for rhBMP-2from collagen sponge has been observed in vitro (Yang et al.,2012). Thus, temporal effects on growth factor expressionlikely reflect variation in rhBMP-2 dose present as well as thephase of fracture healing when RNA was isolated for analysis.
Our data correlate with in vitro studies in which rhBMP-2treatment of murine marrow stromal cells increased endoge-nous BMP-2 expression (Edgar et al., 2007). Increased BMP-2expression caused by rhBMP-2 treatment should promoteosteoblast differentiation and osteogenesis (Rosen, 2009).However, rhBMP-2 treatment decreased BMP-4 expression inmarrow stromal cell cultures but increased endogenous BMP-4expression during fracture healing (Fig. 2) (Edgar et al., 2007).This suggests that the exogenous rhBMP-2 is acting on adifferent cell type to induce BMP-4 expression during fracturehealing or that the rhBMP-2 is indirectly promoting BMP-4expression perhaps by directly stimulating expression of othergrowth factors. BMP-4 can promote chemotactic migration of
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates eAppl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003
mesenchymal progenitor cells (Fiedler et al., 2002), suggestingthat elevated BMP-4 levels may promote additional cellrecruitment to the tissues stimulated with rhBMP-2.
BMP-6 is important for periosteal bone formation in mousegrowth plates, can induce ectopic bone formation, andpromote osteoclastogenesis (Jane et al., 2002; Wutzl et al.,2006; Perry et al., 2008). BMP-6 expression was highest in thecontrol group between days 10 and 21. This delayed peak inBMP-6 expression during fracture healing correlates withprevious results in a rabbit spinal fusion model in which BMP-6mRNA levels peaked 5 weeks after surgery (Morone et al.,1998). In this study, BMP-6 levels were similar between bothgroups at days 4 and 7. However, treatment with rhBMP-2reduced BMP-6 expression by more than 15-fold on days 2, 10,14, and 21. The basis for this rhBMP-2 induced reduction inBMP-6 expression is not understood. The over 100-foldreduction in BMP-6 mRNA levels at day 2 in the rhBMP-2treated samples is more likely to be caused by a direct effect ofrhBMP-2, as rhBMP-2 levels should still be present at thefracture site. However, the reductions in BMP-6 mRNA levels atthe later times may reflect alterations in the healing responseand differential cell populations within the callus. Promotersfor the human BMP-2, BMP-4, and BMP-6 genes have similarstructures suggesting that any differential regulation of BMP-6mRNA levels may be occurring via a post-transcriptionalmechanism (Tamada et al., 1998).
The significant increase in BMP-7 mRNA levels at day 21 isunlikely caused by a direct effect of rhBMP-2. In mouse tibiafracture studies, BMP-7 mRNA levels remained consistentlylow during healing until days 14 and 21 after fracture whenlevels dramatically increased (Cho et al., 2002). BMP-7 wasrecently shown to suppress chondrocyte hypertrophy inATDC5 cell cultures (Caron et al., 2013). Thus, expression ofBMP-7 after peak cartilaginous callus formation may benecessary to avoid negative effects on endochondral ossifica-tion. Since rat fracture healing occurs over a slightly longertime span, we suspect that the almost 100-fold increase inBMP-7 mRNA levels in the day 21 rhBMP-2 treated samplesreflects an advanced stage of healing caused by the priorrhBMP-2 treatment as compared to the control-treated speci-mens.
In mouse tibia fractures, TGF-b1mRNA levels peak at day 10when callus cartilage volume is also maximal (Cho et al., 2002,Manigrasso and O'Connor, 2008). In this study, TGF-b1 mRNAlevels peaked at day 21 in the control-treated specimens whichwould correlate with the period when cartilage volume ishighest during rat femur fracture healing. The reduced levelsof TGF-b1 mRNA in the rhBMP-2 treated specimens at day 21likely reflects an increased proportion of bone in the callus ascompared to cartilage.
TNF-a expression levels in the rhBMP-2 groups trendedhigher than control groups at days 2 and 4 and wassignificantly higher at day 7. However, when TNF-a expressionpeaked in control rats on day 14 the rhBMP-2 treated group wassignificantly lower.
TNF-a is necessary for fracture healing since loss of TNF-asignaling impairs healing (Gerstenfeld et al., 2003). The preciserole of TNF-a in fracture healing is not known. TNF-a signalingmay be necessary to induce COX-2 expression, promotechondrocyte apoptosis, or aid in recruiting muscle-derived
xpression of other growth factors in a rat fracture healing model. J.
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x8
JAB-24; No. of Pages 9
stromal cells to the fracture site (Geng et al., 1995; Simon et al.,2002; Gerstenfeld et al., 2003; Glass et al., 2011). Despite thenecessity of TNF-a, exogenous rhBMP-2 treatment had littleeffect on TNF-a mRNA levels suggesting that rhBMP-2treatment has no effect on TNF-a expression. Similarly, VEGFis necessary for endochondral ossification and angiogenesis isnecessary for fracture healing (Gerber et al., 1999; Hausmanet al., 2001). Despite this, rhBMP-2 treatment had no effect onVEGF mRNA levels.
In conclusion, treatment with rhBMP-2 increased expres-sion of endogenous BMP-2 and BMP-4 during fracture healingin this rat model. These data suggest that rhBMP-2 may act topromote bone healing by inducing expression of endogenouspro-osteogeneic growth factors in addition to its establisheddirect effects on cell differentiation.
r e f e r e n c e s
Alkhiary, Y.M., Gerstenfeld, L.C., Krall, E., Westmore, M., Sato,M., Mitlak, B.H., Einhorn, T.A., 2005. Enhancement ofexperimental fracture-healing by systemic administration ofrecombinant human parathyroid hormone (PTH 1-34). J.Bone Joint Surg. Am. 87, 731–741.
Anitua, E., Alkhraisat, M.H., Orive, G., 2012. Perspectives andchallenges in regenerative medicine using plasma rich ingrowth factors. J. Control. Release 157, 29–38.
Bae, H.W., Zhao, L., Kanim, L.E., Wong, P., Delamarter, R.B.,Dawson, E.G., 2006. Intervariability and intravariability ofbone morphogenetic proteins in commercially availabledemineralized bone matrix products. Spine 31, 1299–1306discussion 1307–1308.
Bais, M.V., Wigner, N., Young, M., Toholka, R., Graves, D.T.,Morgan, E.F., Gerstenfeld, L.C., Einhorn, T.A., 2009. BMP2 isessential for post natal osteogenesis but not for recruitmentof osteogenic stem cells. Bone 45, 254–266.
Balaburski, G., O'Connor, J.P., 2003. Determination of geneexpression variation during fracture healing. Acta Orthop.Scand. 74, 22–30.
Bergenstock, M., Min, W., Simon, A.M., Sabatino, C., O'Connor, J.P., 2005. A comparison between the effects ofacetaminophen and celecoxib on bone fracture healing inrats. J. Orthop. Trauma 19, 717–723.
Bonnarens, F., Einhorn, T.A., 1984. Production of a standardclosed fracture in laboratory animal bone. J. Orthop. Res. 2,97–101.
Caron, M.M., Emans, P.J., Cremers, A., Surtel, D.A., Coolsen, M.M.,van Rhijn, L.W., Welting, T.J., 2013. Hypertrophicdifferentiation during chondrogenic differentiation ofprogenitor cells is stimulated by BMP-2 but suppressed byBMP-7. Osteoarthritis Cartilage 21, 604–613.
Cho, T.-J., Gerstenfeld, L.C., Einhorn, T.A., 2002. Differentialtemporal expression of members of the transforming growthfactor b superfamily during murine fracture healing. J. BoneMiner. Res. 17, 513–520.
Edgar, C.M., Chakravarthy, V., Barnes, G., Kakar, S., Gerstenfeld,L.C., Einhorn, T.A., 2007. Autogenous regulation of a networkof bone morphogenetic proteins (BMPs) mediates the
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates
osteogenic differentiation in murine marrow stromal cells.Bone 40, 1389–1398.
Fiedler, J., Roderer, G., Gunther, K.P., Brenner, R.R., 2002. BMP-2,BMP-4, and PDGF-bb stimulate chemotactic migration ofprimary human mesenchymal progenitor cells. J. Cell.Biochem. 87, 305–312.
Friess, W., Uludag, H., Foskett, S., Biron, R., Sargeant, C., 1999.Characterization of absorbable collagen sponges asrecombinant human bone morphogenetic protein-2 carriers.Int. J. Pharm. 185, 51–60.
Gandhi, A., O'Connor, J.P., Parsons, J.R., Lin, S.S., 2004. Localizedinsulin delivery normalizes growth factor expression in theearly phase of diabetic fracture healing. Trans. Orthop. Res.Soc. 29, 0452.
Geng, Y., Blanco, F.J., Cornelisson, M., Lotz, M., 1995. Regulationof cyclooxygenase-2 expression in normal human articularchondrocytes. J. Immunol. 155, 796–801.
j o u r n a l o f a p p l i e d b i o m e d i c i n e x x x ( 2 0 1 4 ) x x x – x x x 9
JAB-24; No. of Pages 9
Morone, M.A., Boden, S.D., Hair, G., Martin Jr., G.J., Racine, M.,Titus, L., Hutton, W.C., Hutton, W.C., 1998. The Marshall R.Urist Young Investigator Award. Gene expression duringautograft lumbar spine fusion and the effect of bonemorphogenetic protein 2. Clin. Orthop. Relat. Res. 351, 252–265.
Perry, M.J., McDougall, K.E., Hou, S.C., Tobias, J.H., 2008. Impairedgrowth plate function in bmp-6 null mice. Bone 42, 216–225.
Rosen, V., 2009. BMP2 signaling in bone development and repair.Cytokine Growth Factor Rev. 20, 475–480.
Simon, A.M., Manigrasso, M.B., O'Connor, J.P., 2002. Cyclo-oxygenase 2 function is essential for bone fracture healing. J.Bone Miner. Res. 17, 963–976.
Tamada, H., Kitazawa, R., Gohji, K., Kamidono, S., Maeda, S.,Kitazawa, S., 1998. Molecular cloning and analysis of the
Please cite this article in press as: Cottrell, J.A., et al., BMP-2 modulates eAppl. Biomed. (2014), http://dx.doi.org/10.1016/j.jab.2014.02.003
50-flanking region of the human bone morphogeneticprotein-6 (BMP-6). Biochim. Biophys. Acta 1395, 247–251.
Urist, M.R., 1965. Bone: formation by autoinduction. Science 150,893–899.
Wrotniak, M., Bielecki, T., Gazdzik, T.S., 2007. Current opinionabout using the platelet-rich gel in orthopaedics and traumasurgery. Ortop. Traumatol. Rehabil. 9, 227–238.
Wutzl, A., Brozek, W., Lernbass, I., Rauner, M., Hofbauer, G.,Schopper, C., Watzinger, F., Peterlik, M., Pietschmann, P.,2006. Bone morphogenetic proteins 5 and 6 stimulateosteoclast generation. J. Biomed. Mater. Res. A 77, 75–83.
Yang, H., La, W., Cho, Y., Shin, W., Yeo, G., Kim, B., 2012.Comparison between heparin-conjugated fibrin and collagensponge as bone morphogenetic protein-2 carriers for boneregeneration. Exp. Mol. Med. 44, 350–355.
xpression of other growth factors in a rat fracture healing model. J.