VOL 27 NO 1 MARCH 2006 ISSN 0965-0989 Brucellosis – new paradigms for a classical pathogen Sally J Cutler, PhD Bacterial Zoonoses Research Team Leader, Statutory & Exotic Bacterial Diseases, Veterinary Laboratories Agency, Addlestone, Surrey, UK Introduction Globally, brucellosis remains one of, if not the major bacterial zoonosis, still devastating productivity of livestock and affecting mankind, either directly through infection, or indirectly through reduction of productivity among food- producing livestock. It is estimated by the WHO that more than 500,000 new cases of human brucellosis occur each year. Many nations have deployed eradication campaigns, with some countries such as the UK and several other Northern European countries successfully gaining brucellosis-free status. Even in these countries, comprehensive surveillance is an essential prerequisite to maintain this status, exemplified by recent introductions of Brucella-infected cattle to the UK, despite pre- and post-export screening. The aetiological agent of brucellosis is a small, pleomorphic Gram-negative rod, which was first isolated from human clinical cases of undulant fever by David Bruce (1887). Later, in 1904, the Mediterranean Fever Commission under the leadership of David Bruce, identified the reservoir of infection in goats and subsequent transmission to man through consumption of unpasteurised milk. In subsequent years, similar microbes were identified in cattle (B. abortus) and swine (B. suis). This microbe belongs within the α-2 proteobacteria clustering with Bartonella, Ochrobactrum, Agrobacteria and Rhizobia as phylogenetic neighbours. The genus was originally divided into species based on various biochemical capabilities, dye and phage susceptibilities and host preferences (see Table). These species in some cases were further divided into biovars (see Table). Following whole genomic hybridization studies, the high degree of homogeneity among the brucellae was noted, prompting the re-classification into a single species, B. melitensis 1 . Although justifiable according to phylogenetic criteria, this classification has proved unpopular, largely through distinct host susceptibilities and differences in host-pathogen interactions. Brucellae generally possess two chromosomes with a large replicon of 2.1 Mbp (chromosome 1) and a smaller replicon of 1.2 Mbp (chromosome 2); however, B. suis biovar 3 possesses a single replicon of 3.1 Mbp. Plasmids have not been reported within the brucellae. Diagnostic Challenges Clinical Diagnosis: Human clinical disease is characterized by undulant fever, but in many instances other presentations will be predominant including spondylitis, neuropsychiatric complaints, arthralgia, arthritis, respiratory signs and in chronic cases, focal signs often with abscess formation. However, in many cases, clinical signs are protean and often non-specific. In livestock, the disease typically manifests as reproductive failure, often through abortion or weak, infected offspring, but orchitis, epididymitis and lameness can also occur. During acute infection with B. melitensis, blood cultures will yield brucellae in 70-80% of individuals. However, this is significantly reduced with infection with other Brucella species. Importantly, recovery of brucellae from clinical material is often lengthy, necessitating extended incubation periods where brucellosis is suspected. Serology: Serological diagnostics have evolved over time from tests for agglutinins through to numerous ELSIA formats and fluorescent antibody polarization assays 2 . Associated with this evolution is increased specificity; however, even this new generation of assays are prone to detecting false-positive serologically reactive samples. These have been recently reviewed elsewhere 3 . The antigen offering superior IN THIS ISSUE Brucellosis - new paradigms for a classical pathogen: Sally J Cutler Clostridium difficile – new challenges for hyper-virulent strains: Ian Poxton Table. Species, biovars and reservoirs of the brucellae. Brucella species Biovars Reservoir host Brucella melitensis 3 Sheep, goats, cattle Brucella abortus 7 Cattle Brucella suis 5 Swine Brucella canis 1 Dogs Brucella ovis 1 Sheep Brucella neotomae 1 Rodents Brucella pinnipediae* Not determined Otter, seal Brucella cetaceae* Not determined Dolphin, porpoise * Species names not yet approved.
Transcript
V O L 2 7 N O 1 M A R C H 2 0 0 6
I S S N 0 9 6 5 - 0 9 8 9
Brucellosis – new paradigms for a classical pathogenSally J Cutler, PhDBacterial Zoonoses Research Team Leader, Statutory & Exotic Bacterial Diseases, Veterinary Laboratories Agency,Addlestone, Surrey, UK
IntroductionGlobally, brucellosis remains one of, if not the
major bacterial zoonosis, still devastatingproductivity of livestock and affecting mankind,either directly through infection, or indirectlythrough reduction of productivity among food-producing livestock. It is estimated by the WHOthat more than 500,000 new cases of humanbrucellosis occur each year. Many nations havedeployed eradication campaigns, with somecountries such as the UK and several otherNorthern European countries successfully gainingbrucellosis-free status. Even in these countries,comprehensive surveillance is an essentialprerequisite to maintain this status, exemplifiedby recent introductions of Brucella-infected cattleto the UK, despite pre- and post-export screening.
The aetiological agent of brucellosis is a small,pleomorphic Gram-negative rod, which was firstisolated from human clinical cases of undulantfever by David Bruce (1887). Later, in 1904, theMediterranean Fever Commission under theleadership of David Bruce, identified the reservoirof infection in goats and subsequent transmissionto man through consumption of unpasteurisedmilk. In subsequent years, similar microbes wereidentified in cattle (B. abortus) and swine (B. suis).
This microbe belongs within the α-2proteobacteria clustering with Bartonella,Ochrobactrum, Agrobacteria and Rhizobia asphylogenetic neighbours. The genus wasoriginally divided into species based on variousbiochemical capabilities, dye and phagesusceptibilities and host preferences (see Table).These species in some cases were further divided
into biovars (see Table). Following whole genomichybridization studies, the high degree ofhomogeneity among the brucellae was noted,prompting the re-classification into a singlespecies, B. melitensis1. Although justifiableaccording to phylogenetic criteria, thisclassification has proved unpopular, largelythrough distinct host susceptibilities anddifferences in host-pathogen interactions.Brucellae generally possess two chromosomeswith a large replicon of 2.1 Mbp (chromosome 1)and a smaller replicon of 1.2 Mbp (chromosome 2);however, B. suis biovar 3 possesses a singlereplicon of 3.1 Mbp. Plasmids have not beenreported within the brucellae.
Diagnostic ChallengesClinical Diagnosis:
Human clinical disease is characterized byundulant fever, but in many instances otherpresentations will be predominant including
spondylitis, neuropsychiatric complaints,arthralgia, arthritis, respiratory signs and inchronic cases, focal signs often with abscessformation. However, in many cases, clinical signsare protean and often non-specific. In livestock,the disease typically manifests as reproductivefailure, often through abortion or weak, infectedoffspring, but orchitis, epididymitis and lamenesscan also occur. During acute infection with B.melitensis, blood cultures will yield brucellae in70-80% of individuals. However, this issignificantly reduced with infection with otherBrucella species. Importantly, recovery ofbrucellae from clinical material is often lengthy,necessitating extended incubation periods wherebrucellosis is suspected.
Serology:Serological diagnostics have evolved over
time from tests for agglutinins through tonumerous ELSIA formats and fluorescentantibody polarization assays2. Associated with thisevolution is increased specificity; however, eventhis new generation of assays are prone todetecting false-positive serologically reactivesamples. These have been recently reviewedelsewhere3. The antigen offering superior
IN THIS ISSUEBrucellosis - new paradigms for a classical pathogen: Sally J Cutler
Clostridium difficile – new challenges for hyper-virulent strains: Ian Poxton
Table. Species, biovars and reservoirs of the brucellae.
Brucella species Biovars Reservoir host
Brucella melitensis 3 Sheep, goats, cattle
Brucella abortus 7 Cattle
Brucella suis 5 Swine
Brucella canis 1 Dogs
Brucella ovis 1 Sheep
Brucella neotomae 1 Rodents
Brucella pinnipediae* Not determined Otter, seal
Brucella cetaceae* Not determined Dolphin, porpoise
* Species names not yet approved.
sensitivity is lipopolysaccharide with its o-polysaccharide side chain of homopolymers ofN-formyl-perosamine [N-formylated 4-amino, 4,6-dideoxyglucose] found either in α 1-2linkages or α (1-2) linkages together with α (1-3)usually in a 4:1 ratio. These variations result in theA and M serotype specificity of brucellae. The o-polysaccharide side chain shows distinctstructural similarity with the N-formyl-perosaminehomopolymer of N-formyl-perosamine of Yersiniaenterocolitica O:9. This microbe is frequentlyfound in association with swine, but also in otherlivestock species. It is likely that exposure to thismicrobe, or similar ones, may account for theserological “false alarms” encountered withinbrucellosis-free countries. The search forimproved diagnostic antigens has becomesomething of a search for the “Holy Grail”.Several different antigens have been assessed;however, the overall consensus of opinion is thatthey fail to offer diagnostic advantage over currentserological assays4.
Serodiagnosis is further complicated whenused in countries where the disease is endemic.Here significant proportions of the population willnaturally have elevated titres against brucellae.Furthermore, brucellosis eradication effortsutilizing live attenuated vaccine strains (exceptRB51), will result in positive titres in recentlyvaccinated livestock.
Immunological approaches:In order to overcome the problems outlined
above, investigations have been undertaken toevaluate use of other immunodiagnostic methods.As Brucella stimulate a strong Th1 response, thepotential of using elevated interferon-gamma(IFN-gamma) levels following specific stimulationhas been explored. Early results look promising,with this approach clearly differentiating betweenbrucellosis and Yersinia infections, however, noimprovements in test sensitivity were offered.Further limitations arise from the undulating levelsobserved with IFN-gamma responses and theirrequirements for stimulation of blood cells withina tight time window.
2
Vol 27 No 1
Polymerase chain reaction assays: Many PCR assays have been published for the
detection of brucellae using various targets,including the intragenic spacer, IS711, outermembrane protein BCSP315. Typically these willdetect levels of 10fg, which equates to threegenomes. The limitation of this approach is thelikelihood of sufficient bacteria present in blood orserum to enable detection. Brucellae are rapidlyinternalized by host cells, whereby they areinstrumental in development of their particularreplicative niche. Foci of infection are often foundin lymph nodes, bone marrow and varioustissues, especially reproductive tissues inlivestock. Application of PCR to these sampleswill be much more likely to detect Brucella DNA.
Typing methods:Classical biotyping has been the “gold
standard” approach for distinguishing differentisolates into their respective species and biovars.However, this needs highly skilled staff, largeamounts of viable organism requiringcontainment level three facilities and lengthyincubation periods. Given the reputation ofBrucella as the most frequently encounteredlaboratory-acquired infection6, this approach isnot to be undertaken lightly and is probably bestreserved for reference facilities. Various microbialtyping methods, including outer membraneprotein typing; ribotyping; AFLP; PFGE-RFLP;insertion sequence typing; to name but a few,have been applied to the Brucella with mixedsuccess7-9. These approaches were able tosuccessfully speciate isolates, but proved unableto offer differentiation down to biovar levels.
Recent application of PCR-based variablenumber tandem repeat typing (VNTR) orhypervariable octameric oligonucleotide fingerprints (Hoof Prints)10 has proved to be a valuablemethod for traceback studies when outbreaksoccur. However, on a more cautionary note, thosetargets most suited for epidemiological profilingwill not necessarily identify isolates, but are bestsuited for microbial forensic typing. Selection ofmore slowly evolving repeats, or even use of
single nucleotide polymorphisms, can howeverprovide a molecular equivalent to biotyping.
Brucellosis and the “omics” revolutionGenomes of the three principal pathogenic
species have now been published11-13, while thatof B. ovis is currently in progress. Comparativegenomics of these three species has confirmedthe remarkable homogeneity among thesespecies, with closest similarity between B. abortus and B. melitensis12. Surprisingly, only101 unique genes were present/absent in one ofthese three genomes. Transcripts were detectedfrom 54 of these using reverse transcriptase PCR,with one unique transcript to each of B. abortusand B. melitensis and a further 20 in B. suis.Thus, it is likely that the intriguing differences inhost specificity may results from either subtlevariations within this conserved DNA, or throughdifferential expression of conserved genes ratherthan possession of unique genomic DNA.
Studies of the proteome (secretome andcellular proteome), have revealed differencesbetween wild type and attenuated vaccine strainsof B. melitensis. Notably, expression differenceswere demonstrated in iron regulatory proteins,sugar binding, lipid degradation and proteinbiosynthesis between wild type and attenuatedstrains. Comparative proteomic analysis of B. abortus and B. melitensis produced 312differentially expressed proteins. Whether theseare representative of the species and theirsignificance in host specificity of these microbesremains to be determined.
Analysis of the “omic” approaches washeralded as the likely way to dissect the basis forhost specificity and virulence for these microbes;however, the biological basis for these traitsremains elusive. Scrutiny of the Brucella genomesfor conventional microbial virulence genes hasmet with limited success, although genes forcapsules, toxins and other conventional virulencedeterminants have failed to produce insights intothe pathogenesis of these microbes. Surprisingfindings, such as the presence of genes necessaryfor expression of flagellum in this non-motileorganism, were detected. Upon furtherinvestigation, it appears that these are transientlyexpressed and may indeed have a role in thepathogenesis of these microbes14. Certainly thesurface lipopolysaccharide plays a crucial role invirulence of the brucellae together with expressionof type IV secretion system15. Microarray analysisof different Brucella species hybridized to B. melitensis 16M, again show limited diversityamong species. This work identified differencesbetween species suggestive of horizontal geneticacquisition, that appeared to be clustered intogenomic islands; however, these differences alone
Scanning and transmission electron microscopy ofBrucella species.
may not be sufficient to account for the hostspecificity observed within the brucellae16.
Control strategiesEradication campaigns have been introduced
in many countries with varying degrees ofsuccess. Typically these are based on vaccinationof susceptible hosts, surveillance to detectinfected livestock and their subsequent slaughter,and movement restrictions where localizedinfection has been detected. The combination ofthese methods can lead to eventual freedom fromdisease, although the likelihood of success islargely dependent on compliance of manydifferent groups (farmers, import/export,veterinary workers, public health workers,scientists and government). Without thisconcerted, joined-up approach, chances ofsuccess are limited.
Current challengesOur new challenges are the growing incidence
of B. melitensis infection in cattle, particularly inareas such as the Middle East. Vaccine efficacy ischallenged by this new threat as cattle areroutinely immunized with attenuated B. abortusstrains (either S19, or RB51). The ability of thesevaccines to protect against B. melitensis challengein bovines remains largely untested.
A further challenge is posed by the threat ofinfection from wild-life reservoirs. This is a majorchallenge for the control of brucellosis worldwide.Particular problems have been documented withinfected bison re-introducing brucellosis to cattlein USA and with B. suis infected wild boar orEuropean hares posing significant infection risksto outdoor piggeries.
New potential threats are emerging with thesignificant infection levels found among marinemammals. Infection levels of around 26% havebeen reported among samples submitted to our
laboratory. Although experimental studies havedemonstrated infection among terrestriallivestock17, whether this poses a significant riskunder natural circumstances remains to beestablished. Interestingly, human infection withthese strains is possible and consequently, agenuine zoonotic threat exists18.
Possibly the largest challenge is how to tacklethe huge disease burden in developing countries.Here the consequences of brucellosis impactdramatically on both human health and on thelivestock on which man depends. Diagnosticcapability is sporadic and disease epidemiologylargely unknown. Introduction of pen-sidediagnostic assays and education focused towardsthe spread of infection and methods to controldisease in livestock and reduce transmission toman are urgently required.
Recommended Further Reading:Brucella Molecular and Cellular Biology (2004)
Edited by I. López-Goñi and I. Moriyón, HorizonBioscience, Norfolk, England. ISBN: 1-904933-04-1
References1. Verger J., Grimont F., PAD G. and Grayon M.
(1985) Brucella, a monospecific genus as shown by
deoxyribonucleic acid hybridization. International
Journal of Systematic Bacteriology. 35. 292–295.
2. Nielsen K. and Gall D. (2001) Fluorescence
polarization assay for the diagnosis of brucellosis: a
review. J Immunoassay Immunochem. 22. 183–201.
3. Nielsen K. (2002) Diagnosis of brucellosis by
serology. Veterinary Microbiology. 90. 447–459.
4. Letesson J.J. et al. (1997) Humoral immune
responses of Brucella-infected cattle, sheep, and goats
to eight purified recombinant Brucella proteins in an
indirect enzyme-linked immunosorbent assay.
Clinical Diagnostic and Laboratory Immunology. 4.
556–564.
5. Al Dahouk S., Tomaso H., Nockler K. and
Neubauer H. (2004) The detection of Brucella spp.
using PCR-ELISA and real-time PCR assays.
Clinical Laboratory 50. 387–394.
6. Ergonul O., Celikbas A., Tezeren D., Guvener E.
and Dokuzoguz B. (2004) Analysis of risk factors for
Growth of Brucella species on serum dextrose agarshowing characteristic morphology.
Clostridium difficile – new challenges for hyper-virulent strainsIan R Poxton BSc PhD DScProfessor of Microbial Infection and ImmunityUniversity of Edinburgh College of Medicine and Veterinary MedicineCentre for Infectious Diseases, Chancellor’s Building, 49 Little France Crescent, Edinburgh, UK
IntroductionClostridium difficile, a strictly anaerobic,
Gram-positive spore-forming bacterium hasbeen recognised as the cause of antibiotic-associated diarrhoea and colitis for about 30years. The spectrum of disease – ranging frommild, self-limiting diarrhoea, to seriousdiarrhoea, to life-threatening pseudo-membranous colitis – is generally referred to asC. difficile-associate disease (CDAD). Initiallyuncommon, but with a few noteworthyoutbreaks, it has been steadily increasing inprevalence since the early 1990s, reaching levelsin many of our hospitals that are generallyhigher than the better-known methicillin-resistant Staphylococcus aureus (MRSA). It isthe most common cause of health-care-associated (nosocomial) diarrhoea and figuresfor Scotland are summarised in Figure 1. Theshape of the curve is similar for most othercountries in the developed world. CDAD causesmuch morbidity in elderly hospital patients andsome mortality has been associated with thedisease. The Health Protection Agency issued a
press release in December 20051 highlightingthe current status of the organism and disease.(In this web reference there is a link to theHealthcare Commission’s full report on:"Management, prevention and surveillance ofClostridium difficile – Interim findings from anational survey of NHS acute trusts in England")
The recent recognition of a hyper-virulentstrain firstly in Canada and the USA2,3 and morerecently in England, the Netherlands andBelgium has brought the organism to theattention of the general public as the latest“superbug”. For the most recent findings go tothe “News and Activities” section of theEuropean Study Group for Clostridium difficileweb site4.
Historical aspects“Bacillus difficilis” was first recognised in
1935 as a normal component of the stools ofyoung babies, and during the following 40 yearsit received scant attention in the medicalliterature. As recently as 1976 it was considerednon-pathogenic. However, just a year or so later
it was shown to be the cause ofpseudomembranous colitis, a rare but life-threatening condition5. This discovery was aresult of the efforts of researchers investigatinga significant increase in a previously very rarecondition: antibiotic-associated pseudo-membranous colitis, a disease which at that timewas being linked to the use of specificantibiotics, especially clindamycin and therelated lincomycin. C. difficile was subsequentlyrecognized as the major cause of hospital-acquired, antibiotic-associated diarrhoea, alesser form of the more rare and seriouspseudomembranous colitis.
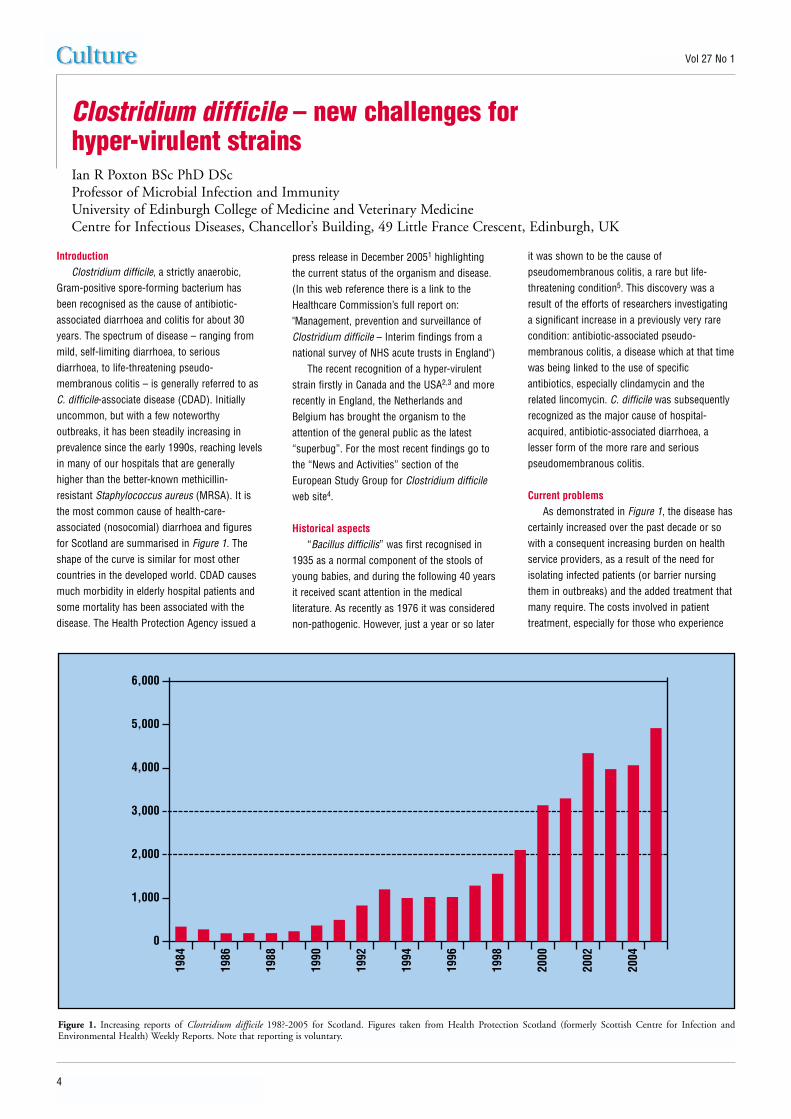
Current problemsAs demonstrated in Figure 1, the disease has
certainly increased over the past decade or sowith a consequent increasing burden on healthservice providers, as a result of the need forisolating infected patients (or barrier nursingthem in outbreaks) and the added treatment thatmany require. The costs involved in patienttreatment, especially for those who experience
4
Vol 27 No 1
Figure 1. Increasing reports of Clostridium difficile 198?-2005 for Scotland. Figures taken from Health Protection Scotland (formerly Scottish Centre for Infection andEnvironmental Health) Weekly Reports. Note that reporting is voluntary.
6,000
5,000
4,000
3,000
2,000
1,000
0
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
recurring disease, are considerable and in 1996were reported to be in the region of £4000 perpatient6. Recent unpublished estimates from theUSA are in the region of US$6000. If this wasn’tbad enough, a new dimension to the problemhas become apparent in the last year or so. In2004 a major outbreak began to unfold in theMontreal region of Quebec, and the CanadianBroadcasting Corporation news serviceannounced “7000 sufferers and 600 deaths inQuebec”. These were four times and six timesthe expected levels, respectively. Because of theunusual severity of the symptoms, clinicians
familiar with the disease suggested early in theoutbreak that a strain of increased virulence wasprobably responsible for the problem. This strainis now known in North America as BI/NAP1. Anapparently identical strain known in the UK asribotype 027 has been recognised as causing anongoing problem at Stoke Mandeville Hospital inBuckinghamshire where at least 12 deaths werereported from 150 cases in the period February– June 2004 (Personal communication, JonBrazier). Since then, other centres haveexperienced problems with this 027 strain –notably in Exeter, Devon and Romford, Essex.
The Secretary of State for Health was questionedin the House of Commons and in the writtenanswer 12 other hospitals throughout Englandwere reported to have the strain – from Truro inthe South West to Newcastle and Sunderland inthe North East (Reported in Hansard 27th June20057). Currently at least nine hospitals in TheNetherlands and six in Belgium are experiencingproblems with the same strain. Although the 027ribotype has been known for several years, therecently-recognised hypervirulent epidemic strainis distinct from earlier isolates in that only recentlyhas it acquired resistance to fluoroquinoloneantibiotics. At the time of writing this article (Jan2006) none has been identified in Scotland.
It appears that the new hyper-virulent strain(BI/NAP1 or ribotype 027) has the followingcharacteristics:
● It produces higher than normal levels oftoxins in vitro and almost certainly in vivo.
● There is a deletion in the gene (tcdC) whichnormally negatively regulates toxinproduction – resulting in constant maximumlevel of transcription of toxins
● It belongs to ribotype 027 and toxinotype III● It is resistant to fluoroquinolone antibiotics
such as ciprofloxacin and moxifloxicin● It produces the binary toxin● The North American and European isolates
are probably identical● The disease that it causes is more severe
with more colectomies required and moredeaths attributable to it.
It is becoming apparent that the deletion inthe toxin-regulating gene is not restricted to the027 ribotype, so a family of super strains maysoon become apparent.
The disease: pathogenesis and epidemiologyThere are several recent reviews of the
crucial processes in the cause of the disease(pathogenesis)8 and briefly they conclude that:
● The normal protective properties of the gut –the colonisation resistance promoted by thenormal resident bacteria of the healthy gut –are compromised by antibiotics or, lessfrequently, other therapeutic agents
● The colon becomes colonised with C. difficile● C. difficile evades the immune response and
multiplies, producing toxins A and B● If a patient is susceptible – i.e. is unable to
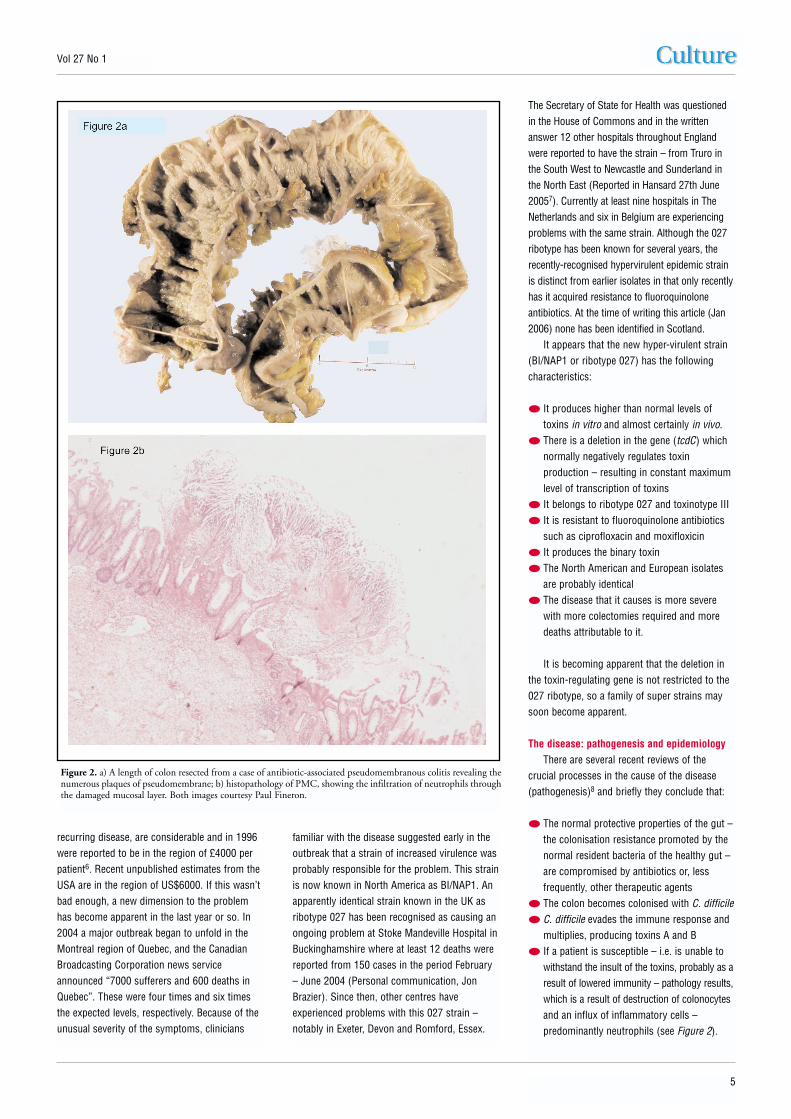
withstand the insult of the toxins, probably as aresult of lowered immunity – pathology results,which is a result of destruction of colonocytesand an influx of inflammatory cells –predominantly neutrophils (see Figure 2).
5
Vol 27 No 1
Figure 2. a) A length of colon resected from a case of antibiotic-associated pseudomembranous colitis revealing thenumerous plaques of pseudomembrane; b) histopathology of PMC, showing the infiltration of neutrophils throughthe damaged mucosal layer. Both images courtesy Paul Fineron.
Our early understanding of the diseaseassumed that the bacterium was of endogenousorigin: small numbers of C. difficile that werepresent in the colon were allowed to flourishafter the normal microbiota was severelyaffected by antibiotic exposure. However, it soonbecame apparent that this was unlikely as it didnot explain the occurrence of outbreaks. Whencarefully looked for in the healthy adult theorganism is rarely encountered. One of ourearliest publications on C. difficile in 1984demonstrated that a single strain wasresponsible for an outbreak9. It is now acceptedthat C. difficile is an infectious agent and istransmitted by faecal-oral transmission of sporesfrom patient to patient, or contaminatedenvironment to patient.
Once established in the gut, the majorvirulence factors of the organism – toxins A andB – are elaborated. The genes encoding andcontrolling these toxins are carried on apathogenicity island referred to as thepathogenicity locus or “PaLoc”10. As well as thetwo genes encoding the toxins (tcdA and tcdB),there is both a positive regulator tcdR and anegative regulator tcdC. A final gene tcdEencodes a holing-like protein that is probably
involved with export from the cell. Normallytoxins A and B are produced on entry intostationary phase, and the transcription of theirgenes is repressed by tcdC. However, in theabove-mentioned hyper-virulent strain, earlytranscription of the toxin genes is a result of adeletion in the tcdC gene.
Laboratory detectionInitially, laboratory diagnosis was based on
traditional bacterial culture on a selectivemedium such as the cycloserine, cefoxitin,fructose agar (CCFA) developed by George et alin 197911. Confirmation that the isolatesproduced toxin was by demonstration of anantitoxin-neutralisable cytopathic effect in tissueculture monolayers. These techniques are highlyspecific and sensitive but labour-intensive andtherefore expensive. However, following thewidespread introduction of rapid toxin detectionby immunoassay, laboratory detection movedaway from culture/cytotoxin detection to thesemore rapid methods. Culture methods have nowlargely been abandoned in the UK, most othercountries in Europe and the USA and diagnosisis relying entirely on detection of toxin(s) instool filtrates by an immuno method. Initially,
only toxin A was detected, but most kits alsonow measure A and B. This followed therecognition of A-negative, B-positive strains ascausing disease. Certainly, detection of toxin(s)is the cheapest option and the results areavailable rapidly. The improved sensitivity ofmany of the commercially available kits meansthat in most cases an extremely reliablediagnosis can be achieved. However, suchmethods do have a serious drawback. As noorganism is isolated it cannot be “typed” toindicate whether it belongs to a hyper-virulentphenotype – one that should be dealt with moreseriously than normal. It also does not permitsurveillance of antibiotic resistance pattern orany other epidemiological or pathogenic marker.
Recommendations for the diagnosticlaboratory● Investigate all stools by a rapid
immunoassay detecting toxins A and B or bytissue culture cytotoxin test from a) hospitalpatients with diarrhoea, b) any other patientswhen requested by a clinician, c) patientswith liquid stools from the community, whenthere is a recent history of antibiotic use.
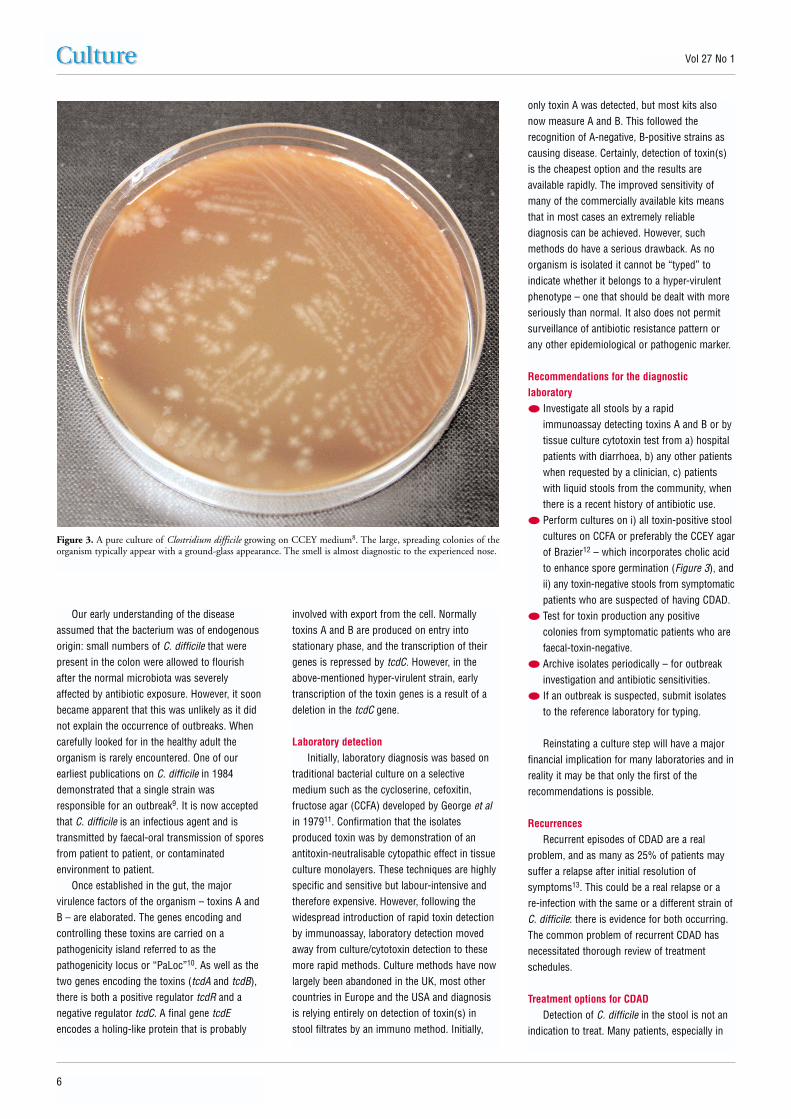
● Perform cultures on i) all toxin-positive stoolcultures on CCFA or preferably the CCEY agarof Brazier12 – which incorporates cholic acidto enhance spore germination (Figure 3), andii) any toxin-negative stools from symptomaticpatients who are suspected of having CDAD.
● Test for toxin production any positivecolonies from symptomatic patients who arefaecal-toxin-negative.
● Archive isolates periodically – for outbreakinvestigation and antibiotic sensitivities.
● If an outbreak is suspected, submit isolatesto the reference laboratory for typing.
Reinstating a culture step will have a majorfinancial implication for many laboratories and inreality it may be that only the first of therecommendations is possible.
Recurrences Recurrent episodes of CDAD are a real
problem, and as many as 25% of patients maysuffer a relapse after initial resolution ofsymptoms13. This could be a real relapse or a re-infection with the same or a different strain ofC. difficile: there is evidence for both occurring.The common problem of recurrent CDAD hasnecessitated thorough review of treatmentschedules.
Treatment options for CDADDetection of C. difficile in the stool is not an
indication to treat. Many patients, especially in
6
Vol 27 No 1
Figure 3. A pure culture of Clostridium difficile growing on CCEY medium8. The large, spreading colonies of theorganism typically appear with a ground-glass appearance. The smell is almost diagnostic to the experienced nose.
wards for the elderly, carry the organismasymptomatically. They may pose a risk to otherpatients, but patients without symptoms do notrequire antibiotic therapy. However, in the futurewhen other (safer) therapies may exist,treatment may be an option.
If the disease is mild, no treatment may benecessary as the symptoms may resolvenaturally. For more severe disease, stopping theadministration of the precipitating antibiotic isthe first option. If this is not possible, orineffective, the first line of treatment isadministration of metronidazole or vancomycin:the former is used more typically these days.Various comparative studies have been donewith these agents and most conclude that thereis not much between them, but some sayvancomycin works a little faster, and should beconsidered for more serious disease.
The real problem of patients sufferingrecurrences after apparent initial resolution withantibiotics – together with the anathema oftreating a disease caused by antibiotics withanother antibiotic – has promoted muchresearch into the development of alternativetypes of treatments. These fall into four mainareas: i) prebiotics and probiotics, ii) absorbentsfor toxin – to eliminate them from the gut, iii)“faecal transplants/enemas” – where stoolsdonated from a healthy donor are placed in thebowel in an attempt to restore the normalmicrobiota, and iv) immunotherapy – eitheractive or (probably more usefully) passiveimmunisation. Lynn McFarlane has recentlyreviewed “alternative treatments” for CDAD14.Currently none of these alternatives has becomeroutine, but efforts are progressing in earnest.Anti-peristaltic drugs are strongly contra-indicated as they may precipitate toxicmegacolon and perforation.
Infection control and the future?Despite our increasing knowledge of the
organism and the disease, it is unlikely that theproblem will disappear in the near future. Manyof our hospitals and institutions caring for theelderly are now generally highly contaminatedwith spores of C. difficile and increasingnumbers of susceptible, antibiotic-treatedpatients are propagating the organism. Infectioncontrol methods are difficult for this disease buttheir importance cannot be stressed too much.They include regular surveillance, isolation orbarrier nursing, personal hygiene, and intensivecleaning of affected wards to remove thebacterial spore load. Note that the spores of C. difficile are resistant to alcohol-basedantiseptics (alcohol hand-washing gels areineffective), and chlorine-based disinfectants are
only partially effective. The use of hydrogenperoxide vapour is being proposed for roomsterilisation – but only after patients and beddinghave been removed! Adherence to strictantibiotic policies: restricting the use of thoseantibiotics such as oral cephalosporins andclindamycin, which are known to precipitate thedisease, is also crucially important. However,more recently the use of fluoroquinolones hasbeen seen to select for the new 027 strains.
Even in areas without the epidemic 027strain, anecdotal reports suggest the incidenceand severity of CDAD has been increasing inrecent years. The situation is likely to worsenbefore it gets better. Major targets for researchmust include: the development of more effective,patient-friendly disinfectants with which to cleanhospitals; new therapies that are not based onantibiotics – such as the use of prebiotics,immunotherapy or toxin inactivators; and moresensitive (molecular) methods such as real-timePCR for accurate diagnosis. Crucially, muchmore funding is required for this organism anddisease, which to date has been minimal in theUK. Together with further research there shouldalso be better surveillance and mandatoryreporting of the organism.
AcknowledgementsI am grateful to the many colleagues, PhD
students and fellow committee members of theEuropean Study Group for Clostridium difficile(http://www.escmid.org/sites/index_f.aspx?par=2.5) who have contributed to my knowledge ofthe organism. Thanks to Paul Fineron for theimages in Figure 2 and thanks also to the ChiefScientist Office of the Scottish Executive HealthDepartment for past and current funding.
About CultureCulture has been produced by Oxoid for over 25 years. Past issues are now a rarity, but recent issues are now available to view, download and print out via
the Oxoid website (www.oxoid.com) Alternatively, if you would like to receive Culture each time it is produced, please speak to your local Oxoidrepresentative or send your name, address, telephone and E-mail details to Martin Cunningham at the address shown below.
Culture is published by Euromed Communications, Haslemere, UK.Edited by Joe Ridge and peer-reviewed by Dr Eric Y Bridson, Professor Grahame W Gould and Mr David E Post.
Folio No. LT 1086A
New Oxoid Dry-Bags simplify media preparationNew Oxoid Dry-Bags greatly reduce the timeand labour involved in preparing large volumesof primary enrichment media and diluents forfood microbiology testing. Pre-weighed, steriledehydrated media are supplied in lightweight,transparent plastic bags – all that is required isthe addition of water and the medium ordiluent is ready for use.
Oxoid Dry-Bags completely eliminate the needfor glassware, which reduces hazards in thelaboratory and removes the need for washingand sterilisation. Water is simply addedthrough a special filter and the bag is ready touse in just minutes. Each bag makes 20 litresof medium or diluent, which can be dispensedeasily through the outlet tube. Once empty,the bag can be disposed of with normallaboratory waste.
Oxoid Dry-Bags are supplied in compact boxesof 5 or 10 bags, which are lightweight and easyto transport, providing ultimate convenience inthe laboratory but taking up much less spacethan fully-prepared media products or mediapreparation facilities.
To find out about the range of media anddiluents available as Dry-Bags please telephone +44 (0) 1256 841144, email [email protected] or visit the Oxoid website at www.oxoid.com