Converted to digital format by Aaron M. Feuer (NOAA/RSMAS) in 2005. Copy available at the NOAA Miami Regional Library. Minor editorial changes may have been made. Changes in Cayo Enrique, La Parguera, Puerto Rico, From 1936 to 1980 Using Aerial Photoanalysis Roy A. Armstrong University of Puerto Rico, Mayaguez Campus
Transcript
Converted to digital format by Aaron M. Feuer (NOAA/RSMAS) in 2005. Copy available at the NOAA Miami Regional Library. Minor editorial changes may have been made.
Changes in Cayo Enrique, La Parguera, Puerto Rico, From 1936 to 1980 Using Aerial Photoanalysis
Roy A. Armstrong University of Puerto Rico, Mayaguez Campus
CHANGES IN CAYO ENRIQUE, LA PARGUERA, PUERTO RICO,
FROM 1936 TO 1980 USING AERIAL PHOTOANALYSIS
By
Roy A. Armstrong
A thesis submitted in partial fulfillment of the requirements for the degree of
MASTER OF SCIENCE
in the Department of Marine Sciences
UNIVERSITY OF PUERTO RICO MAYAGUEZ CAMPUS
December 1981
SUMARIO
Fotografias aereas de Cayo Enrique, La Parguera, Puerto Rico, se obtuvieron de
los anos 1936, 1951, 1963, 1971, 1978, 1979 y 1980. De estas se hicieron ampliaciones a
una escala aproximada de 1:4,000 para diferenciar las distintas zonas del arrecife; sin
embargo, en esta escala solo las comunidades mayores se pudieron medir con presicion.
Los rasgo vistos en las fotos aereas fueron corroborados por reconocimiento en el campo.
Se notaron -altas variaciones en las areas de Rhizophora mangle y Thalassia testudinum
a to largo de un perlodo de 44 anos. La tasa de aumento en estas dos comunidades
sugiere que Cayo Enrique se aproxima a una zonaci6n climax dominada primordialmente
por especies de plantas.
Cambios detectables en Cayo Enrique, usando una serie cronologica de fotos aereas,
incluye los efectos de los huracanes. Estos efectos consisten de acumulaci6n de coral muerto
("boulder ramparts") en el nano arrecifal y una disminuci6n en el area de mangle vivo. Distur-
bios periodicos aumentan la diversidad de especies del fronton arrecifal y promueven el
establecimiento de comunidades de coral en la laguna y el nano arrecifal.
Se discute el uso de pelicula en blanco y negro, diapositivas a color y pelicula
infrarroja de falso color. Informaci'on obtenida previamente mediante el uso de fotografias
aereas de arrecifes de coral provee los estudiosfundamentales necesarios para evaluar los
efectos actuales de interferencia humana y las catastrofes naturales en ecosistemas de
arrecifes coralinos.
ABSTRACT
Aerial photographs of Cayo Enrique, La Parguera, Puerto Rico, were obtained for
the years 1936, 1951, 1963, 1971, 1978, 1979 and 1980. Enlargements were made to the
approximate scale of 1:4000 to differentiate the various reef zones; however, only reef
macro-assemblages could be accurately measured at this scale. Features seen in the
photographs were substantiated by surface reconnaissance. Rhizophora mangle and
Thalassia testudinum areas, measured with a planimeter, showed the maximum variation
of all reef zones for a 43 year period. The rate of increase in these two communities
suggests that Cayo Enrique is approaching a climax zonation dominated primarily by plant
species.
Detectable changes in Cayo Enrique, using a chronological series of aerial
photographs, include the effects of hurricane disturbances. Hurricane effects in Cayo
Enrique consist of boulder rampart deposition on the reef flat and an overall reduction of
living mangrove area. Periodic disturbances increase the species diversity of shallow fore-
reef areas and promote the establishment of coral communities on reef-flat and lagoonal
zones.
The use of black and white, color reversal and false-color infra-red films in aerial
photoanalysis of coral reefs is discussed. Past aerial coverage of coral reefs provides the
baseline data needed to assess present day effects of human interference and natural
catastrophes on coral reef communities.
ii
ACKNOWLEDGEMENTS
I wish to thank all members of my committee for their time and assistance; M. J.
Cerame-Vivas for his suggestions and providing necessary equipment and materials, C. E.
Cutress for his constant advice on photographic methods and critically reviewing the thesis,
L. R. Almodovar for his helpful suggestions and advice. For piloting the airplanes used in this
work I thank M. Ewing and V. Fingerhut. Special gratitude to L. Riggs for her field assistance
and support. I also wish to thank M. Hernandez-Avila, director of the Department of Marine
Sciences for providing funds to purchase the old aerial photographs used in this thesis.
iii
TABLE OF CONTENTS
Page
LIST OF FIGURES ..................................................................................................................v LIST OF TABLES ................................................................................................................... vi INTRODUCTION .....................................................................................................................1 REVIEW OF LITERATURE ………………………………………………………………………..3 MATERIALS AND METHODS ................................................................................................6
Aerial photoanalysis Field survey The study site
RESULTS……………………………………………………………………………………………17
Photointerpretation of reef zones and features
I. Black and white film II. Color film III. False-color infrared film
Practicality and limitations of aerial photoanalysis in coral reef studies Changes in a 44 year period
I. Thalassia testudinum areas II. Rhizophora mangle areas III. Hurricane disturbances IV. Human interference
Theoretical aspects and future trends
CONCLUSIONS ....................................................................................................................46 LITERATURE CITED .............................................................................................................48 APPENDIX .............................................................................................................................53
iv
LIST OF FIGURES
Figure Page
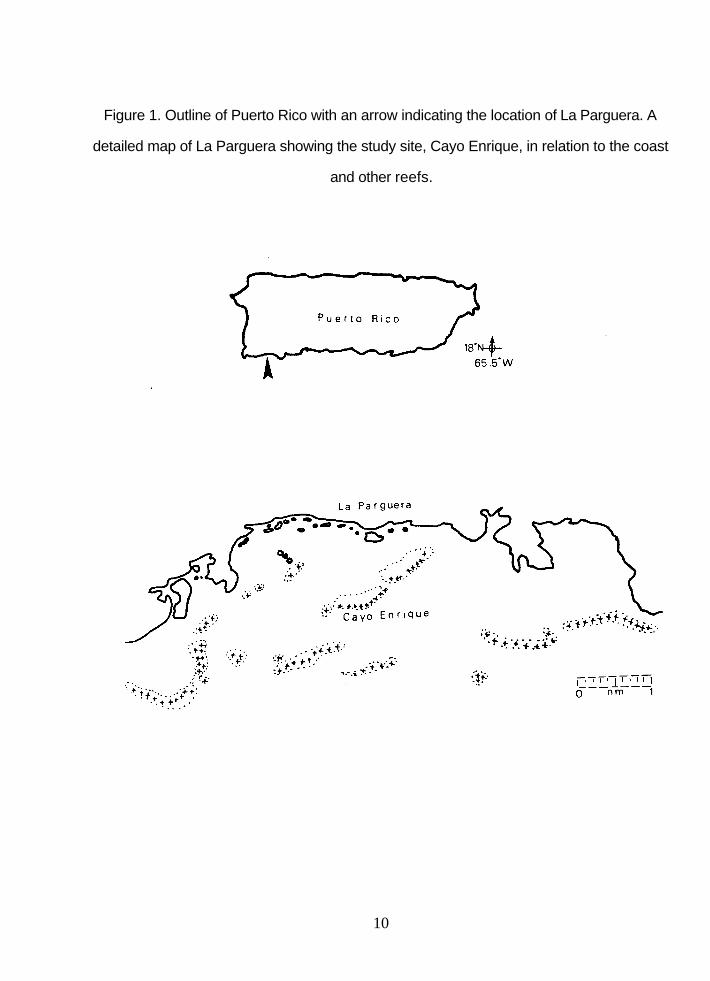
1. Outline of Puerto Rico with an arrow indicating the location of La Parguera. A detailed map of La Parguera showing the study site, Cayo Enrique, in relation to the coast and other reefs ................................................................10
2. 1978 aerial photograph and diagram of Cayo Enrique showing the various zones and features ................................................................11
3. Low-altitude oblique black and white aerial photograph of Cayo Enrique taken in 1980. The different reef-flat zones are discussed in the text ................................................................14
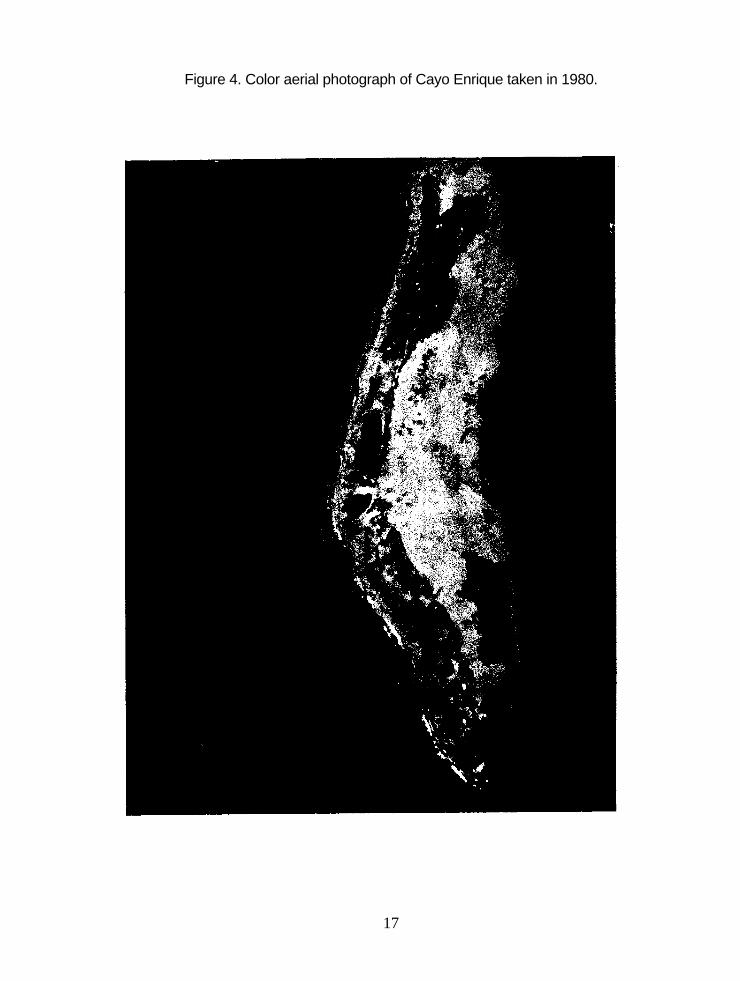
4. Color aerial photograph of Cayo Enrique taken in 1980 . . . 17
5. Infrared color aerial photograph of Cayo Enrique (a portion of a NASA high-altitude infrared color photograph taken in 1974) ................................................................................................20
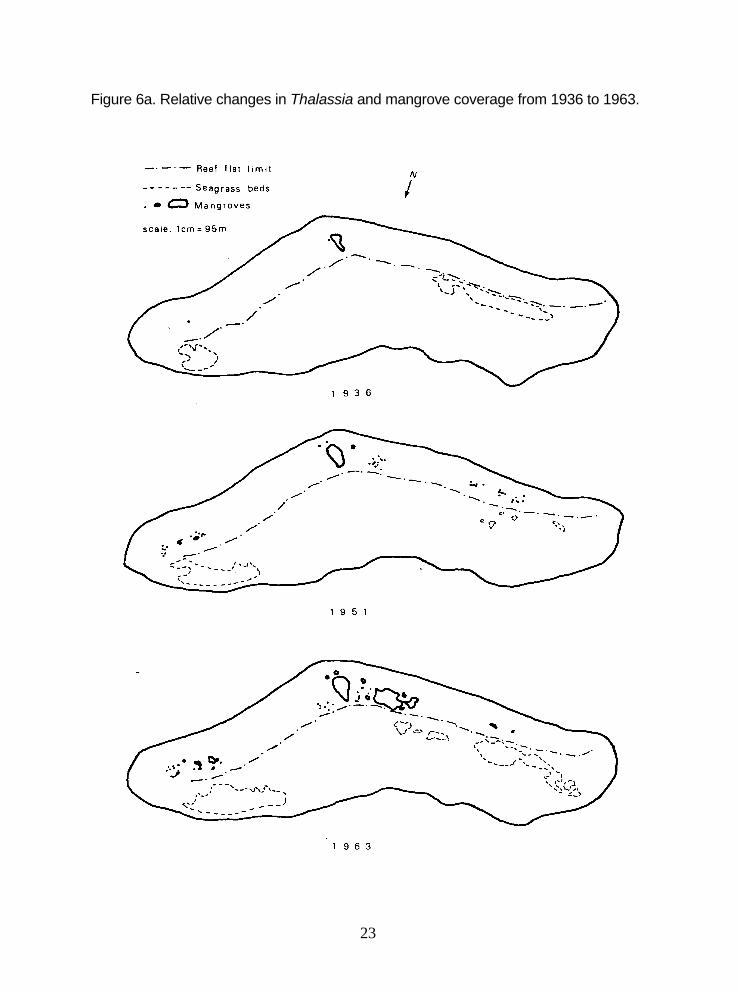
6a. Relative changes in Thalassia and mangrove coverage from 1936 to 1963 ................................................................................................23
6b. Relative changes in Thalassia and mangrove coverage from 1971 to 1979................................................................................................24
7. Area measurements of Thalassia beds in Cayo Enrique . . . . 25
8. Area measurements of mangroves in Cayo Enrique .............................................................27
9. Oblique low-altitude aerial photographs of Cayo Enrique showing boulder rampart formations on the reef flat ..........................................................................................................................31
10. Accumulation of Thalassia and Syringodium blades on Cayo Enrique after Hurricane David ................................................................33
11. Histogram of constituents of the boulder ramparts at Cayo Enrique ................................................................................................34
v
LIST OF TABLES Table Page
1. Criteria used to define reef features and zones of Cayo Enrique using black and white photographs ...................................................................................................... 8
2. Criteria used to define reef features and zones
of Cayo Enrique using color photography. *Color classification after Maertz and Paul (1950) .............................................................................................................. 18
1
INTRODUCTION
Coral reefs are among the most biologically productive and taxonomically diverse of
ecological communities. They are extremely efficient ecological units which support a large
biomass in the relatively barren marine environment (Klim, 1969; Odum & Odum, 1955).
Coral reefs form barriers to waves and currents, protecting coastal areas against
erosion. Reefs also modify adjacent sedimentary and depositional patterns creating sandy
cays and islands. Coral reef ecosystems, which may comprise over 3,000 species of plants
and animals, are increasingly used for recreational purposes and for scientific research.
Coral reefs are absent from the north coast of Puerto Rico. They are common on the
eastern and southern sides of the island, reaching their maximum development on the
southwestern coast off La Parguera. The study site, Cayo Enrique, is a typical coral reef of La
Parguera frequently visited by local fishermen, pleasure boats, and marine scientists.
This study attempts to measure changes in the reef morphology and features of Cayo
Enrique over a 44 year period using aerial photo-analysis. This technique has been used
before in coral reef studies only to map and differentiate reef zones and for illustrative
purposes. A series of vertical panchromatic aerial photographs, dating from 1936 to present,
were used to measure various reef macroassemblages such as Thalassia beds and
mangrove areas.
2
Detectable changes in the reef morphology and features of Cayo Enrique are
correlated to the meteorological record and/or possible human interference. Catastrophic
effects of hurricanes are of particular interest in the recent history of Cayo Enrique.
Hurricanes David in 1979 and Allen in 1980 caused major coral destruction. Boulder ram-
part islets were formed on the reef-flat, composed mostly of dead pieces of Acropora
palmata. Although mangroves and other reef organisms were adversely affected by the
hurricane, Thalassia beds appeared to be undisturbed.
Human activities in Cayo Enrique appear to have had no detectable effects on the
reef environment. Hurricane disturbances of moderate intensity are an important factor in
maintaining high species diversity in coral reefs.
The practicality and limitations of aerial photoanalysis in reef studies is discussed.
When available, a chronological series of aerial photographs is a useful tool in detecting
and measuring trends in coral reefs and provides baseline data by which present and future
comparisons can be made.
3
REVIEW OF LITERATURE
Verrill (1869), one of the earliest workers on West Indian coral reefs, compared the
coral fauna of the Atlantic and Pacific coasts off the Isthmus of Darien. Rathbun (1879)
described Brazilian corals and coral reefs. Vaughan (1901) was the first investigator to
sample and describe Puerto Rican corals.
In Jamaica, Goreau and co-workers did extensive field work on the coral reefs of the
north coast of the island. Their exploration of the deeper reef zones revealed active reef
framebuilding to a depth of 100 m. Several new coral species were discovered, bringing to
62 the species of scleractinian corals known from Jamaica (Goreau and Wells, 1967).
In Puerto Rico, Almy and Carrion-Torres (1963) studied the coral reefs around the
island and reported 38 coral species. Pressick (1970) found 18 scleractinian species in a
reef near Icacos Island, on the northeast coast of Puerto Rico. Glynn (1973a, 1973b)
described the Porites reef-flat biotope of Cayo Laurel in La Parguera. The effects of
sedimentation on Puerto Rican reef corals has been discussed by Loya (1976) and Rogers
(1977).
Cayo Enrique, besides being one of the largest and best developed reefs of La
Parguera, has been studied very little. Wallace et al. (1973) described the zonation of Cayo
Enrique's fore reef. The effect of burrowing organisms on the reef apron sands of Enrique
was described
4
by Morelock and Mathews (1973) and Mathews (1974). Morelock et al. (1977)
described the Enrique reef environment and divided it into three different depositional and
morphological areas: the reef apron, the reef flat and shoals, and fore reef.
Aerial photography and other remote sensing techniques have been used very little
in coral reef studies. As early as 1925, vertical aerial photographs were taken of the Great
Barrier Reef in Australia (Hopley, 1977). After the Second World War, many coral reef
papers used aerial photography to differentiate reef zones and for illustrative purposes (e.g.
Steers, 1945; Teichert and Fairbridge, 1948, 1950).
Kumpf and Randall (1961) used a series of aerial photographs in charting the marine
biota around St. John, U.S. Virgin Islands. Kelly and Conrod (1969) made a qualitative
analysis of the bottom communities on the Bahama Banks using aerial photography.
Guidelines for the use of aerial photography in reef studies are provided by Hopley (1977)
and Hopley and Steveninck (1977).
Hurricane effects on coral reefs and shallow-water benthic communities have been
documented by many investigators. In a series of papers, Stoddart (1962, 1963, 1974)
studied the effects of a severe hurricane that affected reefs in the British Honduras region
in 1961. In 1972 a re-survey of these reefs showed that little or no recovery had taken
place on reefs subjected to massive damage, whereas complete recovery had occurred on
moderately or slightly damaged reefs (Stoddart, 1974).
Other studies on hurricane effects on Caribbean coral reefs and
5
benthic communities include Thomas et al. (1961), Tabb and Jones (1962),
Oppenheimer (1963), Vermer (1963), Ball et al. (1967), Perkin and Enos (1968), and, in
the Pacific Ocean, Flood and Jell (1976), Randall and Eldredge (1977), and Ogg and
Koslow (1978).
In Puerto Rico, the effects of Hurricane Edith on La Parguera reefs were documented
by Glynn et al. (1964). They report minor damage to Cayo Enrique and no coral-shingle islet
formation. Glynn (1973, p. 306) reported that extensive areas on the windward and back
side of Cayo Laurel were affected after the passage of Hurricane Beulah on September 10,
1967. Twelve years later, Hurricane David (August 25-September 8, 1979) passed 70 to 80
miles south of Puerto Rico on a west-northwesterly course. The effects of this and other
hurricanes on Cayo Enrique are documented here by the author (see "Hurricane distur-
bances, p. 40).
6
MATERIALS AND METHODS
Aerial Photoanalysis
Vertical panchromatic aerial photographs were analyzed from the years 1936, 1951,
1963, 1971, 1978, 1979 and 1980. The 1936-1971 aerial photographs were obtained from
the Photogrammetry Division, Department of Public Works. The 1978 aerial photograph was
obtained from Mark Hurd Aerial Surveys Incorporated, San Juan, Puerto Rico. The author
photographed the 1979-1980 aerial coverage from a small high-winged airplane. This
included vertical and low-altitude oblique panchromatic and color reversal photographs.
Kodak Plus-X Pan Professional Film 4147 in 4 X 5 format, and Kodak Kodachrome color
reversal and Plus-X Pan Film in 35 mm format were used. A polarizing filter was used with
color reversal film to minimize the effects of sun glare.
All photographs were enlarged to the approximate scale of 1:4000. A more exact
scale was computed for each photograph using man-made structures, which were
measured in the field and compared to their equivalent photographic image measurement
using the formula:
S = D/Pm
where D = the ground measurement and Pm = the photographic image measurement.
The 1979-1980 photographic scale was computed by using:
S = F/H
where F = the focal length of the camera and H = the flying altitude.
A NASA high-altitude infrared color photograph was used to show this film 's ability
to differentiate shoreline and reef mangrove
7
vegetation. This photograph was obtained from EROS Data Center, U.S.
Geological Survey. Seagrass and mangrove areas were traced from each
photograph and measured with a planimeter. A 10 x illuminated hand magnifier
was used for detailed observation of the photographs. Different reef features and
zones were defined in all aerial photographs according to their tone and their
location on the reef (Table 1). The meteorological data was obtained from the
Ensenada and Magueyes Island stations of the U.S. National Weather Service.
Field Survey
Cayo Enrique was surveyed by walking on the reef-flat, snorkeling on the
back-reef, and SCUBA diving on the fore-reef. Transects were made on the fore-reef
using a 10 m long plastic chain placed parallel to depth contours at 2 m intervals.
Percentage coral coverage and species diversity were recorded for each transect.
Species diversity was measured using the Shannon-Weaver (1949) function:
H' = pi In pi
where pi = ni/N. Evenness was calculated as
H'/H' max
where H' max is the diversity that would exist if all the species pre-sent were
distributed equally (Pielou, 1966). Depth was measured using an oil-filled depth
gauge. The width of the various reef-flat zones was measured using a 30 m
measuring tape and reel. Boulder rampart composition was determined by randomly
picking 100 fragments from the eastern and middle boulder ramparts.
8
Table 1. Criteria used to define reef features and zones of Cayo Enrique using
panchromatic photographs.
Feature Location Tone
Acropora palmata zone
windward side of reef to 3 m depth
medium to dark gray, contrast- ing with light substrate
Millepora-Palythoa zone
reef crest light gray, usually defined by white line of breaking waves
Living and dead Porites and other corals
leeward and adja-
cent to reef crest
light gray mixed with sandy areas
Thalassia-Zoanthus
zone
most of reef flat areas
medium to dark gray
mangroves limited to reef flat areas
even medium gray tone, mottled
appearance
Thalassia seagrass beds
east and west ends of back reef lagoon
light to medium gray tones
sandy areas back reef lagoon and slope
white to very light gray tones that vary with depth
small patch reefs back reef lagoon dark to very dark spotted pat- tern over light gray lagoon sands
boulder ramparts reef flat areas behind reef crest
brilliant white tones with dark edges
9
The 1979 and 1980 aerial photographs were taken to the field to correlate the
features seen in the photographs to the field observations. Color slide film was used to
document the effects of Hurricanes David and Allen on the reef.
The Study Site
Cayo Enrique is located 1.5 km south of the village of La Parguera. It is
approximately 1.4 km long by 0.4 km at its widest point, and is aligned almost parallel to
shore (Fig. 1).
Cayo Enrique was selected for this study because of its relatively large size and the
presence of various biotic macroassemblages, such as seagrass beds and mangrove areas.
Due to its proximity to land, it also appears in most aerial photographs of the area.
The temperature-salinity characteristics of La Parguera are indicative of a mild
hydrographic climate (Glynn, 1973). A continuous surface current flows over the reefs; this
and a maximum daily tidal range of only 40 cm prevent marked temperature and salinity
differences.
A stable reef-flat and back-reef environment is thus provided. Although heavy mortality
of reef-flat organisms related to midday extreme low tidal exposure do occur (Glynn, 1968),
their effect on the reef's macro-flora and fauna is negligible when compared to the
catastrophic effects of hurricanes.
Cayo Enrique can be classified as an apron reef with a shallow (0.5-3 m deep)
area of sand deposition leeward of the reef flat
(Fig. 2). These sediments are medium and coarse-grained sands composed mainly
of Acropora, Porites and Halimeda fragments (Morelock et al.,
10
Figure 1. Outline of Puerto Rico with an arrow indicating the location of La Parguera. A
detailed map of La Parguera showing the study site, Cayo Enrique, in relation to the coast
and other reefs.
11
Figure 2. 1978 aerial photograph and diagram of Cayo Enrique showing the various zones and features.
12
1977). Both seagrass beds and small patch reefs occur in this zone. Thalassia
testudinum areas are found on both ends of the sandy lagoon and on the reef flat. The
seagrass, Syringodium filiforme, and the algae, Dictyota divaricata and Spyridia
filamentosa, are usually found associated with Thalassia. The sea urchins, Diadema
antillarum, Tripneustes ventricosus and Lytechinus variegatus, are common in the sandy
lagoon.
The reef flat. is composed mainly of living Thalassia, Zoanthus, Porites, and,
occasionally, Halimeda clumps. Rhizophora mangle occurs in the middle and both ends of
the reef flat. A more detailed description of the reef-flat zone, based on aerial
photointerpretation, is found on page 17.
The reef crest is dominated by the hydrocoral, Millepora complanata. The
zoanthidian, Palythoa caribbea, is found near the reef crest encrusting dead corals.
At the eastern end, the fore-reef platform of Cayo Enrique is relatively broad and
gradually slopes to a depth of 20 m while be-coming narrower and steeper to the west.
This reef lacks spur-andgroove development of the fore-reef but has a well-defined coral
zonation. Acropora palmata occurs seaward of the reef crest to a depth of 3 in, followed
by Acropora cervicornis to a depth of 5 m. A zone of massive corals occurs from 5 to 15
m and is composed mainly of Montastrea, Diplopia and Agaricia.
The lagoon coral patch-reefs are dominated by the coral, Montastrea annularis, in
addition to numerous sponges and gorgonians. Other corals present include Acropora
cervicornis, Siderastrea siderea, and Diploria labyrinthiformis. Dead coral heads
covered with algae are common in this area.
13
RESULTS
Photointerpretation of reef zones and features
I. Black and white film
Black and white (panchromatic) film was the principal film used in this research. This
is the easiest to handle, most economical, and most widely used film in aerial surveys. Reef
features and zones are recorded on this film as different tones of gray. Tone and location on
the reef was used to differentiate between the various reef features and zones (Table 1).
Tones vary from a brilliant white for boulder ramparts to medium and dark gray for
mangroves and living coral, respectively. Hopley (1977) stated that dead coral, having a
greater light reflectance, appears grayer than living corals in this film.
A vertical panchromatic aerial photograph and diagram of Cayo Enrique was
used to show major reef macroassemblages (Fig. 2). This photograph, taken in 1978,
can be used in conjunction with the criteria listed in Table 1 to differentiate most reef
features.
A low-altitude oblique black and white aerial photograph shows the different reef-flat
zones of Cayo Enrique more clearly (Fig. 3). A transect was made across the reef flat on the
western part of the reef to interpret the zones seen in the oblique aerial photograph. The reef
crest is approximately 8 m wide and is dominated by the hydro-coral, Millepora complanata.
The zoanthidian, Palythoa caribbea, is found here encrusting dead corals. Small colonies of
Porites
14
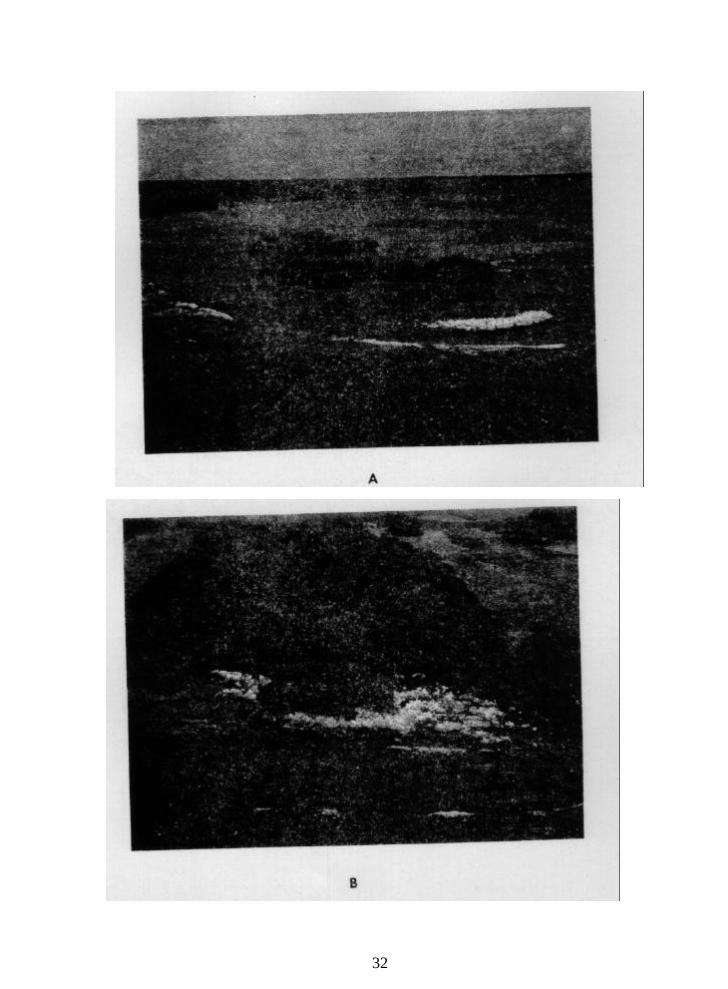
Figure 3. Low-altitude oblique black and white aerial photograph of Cayo Enrique
taken in 1980. The different reef-flat zones are discussed in the text.
(A) Boulder rampart at the eastern end of the reef, and
(B) boulder rampart at the south windward side of the reef.
32
33
Figure 10. Accumulation of Thalassia and Syringodium blades on Cayo
Enrique after Hurricane David.
34
Figure 11. Histogram of constituents of the boulder ramparts of Cayo Enrique.
35
pica) and fragments of Montastrea.
An approximate decrease of 1,560 m2 in living Rhizophora mangle area (see
Fig. 8) occurred due to the catastrophic effects of Hurricane David. While some
mangroves were completely uprooted, most of the damage was caused by
hurricane winds and sea-spray scalding of the leaves with subsequent defoliation
(Almodovar, personal communication). Mechanical damage of Rhizophora prop-
roots resulted from abrasion and piling-up of coral boulders against them. Mangrove
defoliation also occurred in exposed localities after the passage of Hurricane Edith
(Glynn et al., 1964). Wadsworth and Englerth (1959) reported breakage and
uprooting of Laguncularia racemosa trees on the southeastern coast of Puerto Rico
after Hurricane Betsy. Craighead and Gilbert (1962) reported widespread
destruction of mangroves in southern Florida after Hurricane Donna. In addition to
defoliation, they claimed that mechanical damage, root damage, and oxygen
deficiency resulted in mangrove mortalities of 25-75% over large areas, reaching up
to 90% locally. Stoddart (1971) observed that mangroves that were completely
defoliated in the British Honduras cays after Hurricane Hattie suffered permanent
damage. He claimed that dead mangroves, in the absence of fires, can remain in
place for years or even decades.
No reduction in Thalassia coverage resulted from the passage of
Hurricane David. This agrees with the Thomas et al. (1961) report of light damage
to the Thalassia beds of Biscayne Bay after Hurricane Donna. Oppenheimer
(1963) stated that, after Hurricane Carla, not only did the grass flats remain intact
36
but they appeared more healthy than at any time during the previous three years.
He suggested that wave motion apparently removed old and unattached grasses
and algae, leaving clean grass flats. While this is true for effects of minor to
moderate disturbances on protected seagrass beds, the effects of major distur-
bances on exposed seagrass beds could be catastrophic.
Several weeks after Hurricane David, there was a bloom of the alga,
Trichosolen duchassaingii. on the reef-flat of Cayo Enrique and on other reefs of
La Parguera. This alga is an opportunistic species that covers newly available
substrates (Matta, personal communication).
A re-survey of Cayo Enrique was done on August 4, 1980, almost one year
after Hurricane David. On the fore-reef zones, massive and hemispherical corals
were alive and looked healthy, while the mixed coral zone was mostly reduced to
Acropora cervicornis rubble. Several dead colonies in growing position were
covered with algae. The A. palmata zone was predominantly covered by
fragmented in situ colonies of the same species. Regeneration was evident, as
some A. palmata colonies showed a low-relief healthy growth. This probably
resulted from fragments of A. palmata that survived the hurricane and remained in
situ. Highsmith et al. (1980) reported 39% survivorship of coral fragments and
detached colonies after Hurricane Greta in Belize. High survivorship, plus fast
calcification rates in Acropora, promotes rapid reef recovery.
Numerous mangrove seedlings were starting to grow near the man-
groves and on the boulder ramparts. It was evident that just one year after
Hurricane David corals and mangroves were starting to colonize the reef again.
On August 5, 1980, Hurricane Allen, one of the severest storms of the
37
century, passed approximately 200 miles south of La Parguera with maximum
winds of 148 knots near its center. Although the total rainfall recorded on Magueyes
Island Station was only 0.04 m on the day of
the storm, strong wave action affected the reefs. The following day a survey of
Cayo Enrique showed the following effects: (1) the boulder rampart on the middle
part of the reef was moved leeward and piled-up against the mangroves. A new
boulder rampart was deposited to windward near the reef-crest. This was
composed mostly of old dead coral fragments (resulting from Hurricane David)
which were transported by wave action from the fore-reef; (2) mangrove seedlings
growing on the boulder ramparts were destroyed by abrasion and burial, while
seedlings that were established near the mangroves to leeward survived the hurri-
cane; and (3) on the eastern reef-flat area there were uprooted, small, mangrove
trees, dead sea fans and gorgonians, and other dead invertebrates.
Unfortunately, the 1980 aerial photograph of Cayo Enrique (Fig. 4) could not
be used to measure the seagrass and mangrove areas after Hurricane Allen
because the photograph is slightly oblique. As mentioned before, only vertical aerial
photographs can be used to measure reef areas accurately.
In the recent history of Cayo Enrique, two hurricanes (David, 1979
and Allen, 1980) were responsible for considerable damage to various reef
features and zones. These hurricane-generated changes were detectable in
the aerial photographs of those years. Other hurricanes have also affected
the reefs of La Parguera since 1936 (e.g. Hurricane Edith in 1963), but none
have caused as much damage to Cayo Enrique as Hurricanes David and
38
Allen. Hurricane David also caused extensive damage to Acropora
cervicornis colonies at a depth of 20 m at the shelf edge off La Parguera
(Boulon, 1981). Hurricane David was undoubtedly the largest storm to affect
La Parguera reefs during the last four decades.
Hurricanes Betsy (1956), Edith (1963), Beulah (1967), David (1979), and
Allen (1980) have been reported in the literature or in this study as hurricanes
affecting La Parguera reefs. Glynn (1973, p. 110) reports damage to many areas
of Laurel reef due to Hurricane Beulah (September 10, 1967). The tracts of these
hurricanes and their minimum distances from the south coast of Puerto Rico (see
Appendix,
p. 69) have been used to establish the distance of 60 nautical miles as the
minimum distance from shore for hurricanes to have any effects on the reefs of La
Parguera. This arbitrary distance of 60 nautical miles is an approximation, since not
only is the distance from shore important, but also the magnitude of the storm and
the speed of translocation. Hurricane Allen (1980) was of such great magnitude
that, even at a distance of 180 nautical miles it did considerable damage to the
reefs of La Parguera. A total of 17 hurricanes have potentially affected Cayo
Enrique and the other reefs of La Parguera over the last 100 years. This represents
an average of 1.7 hurricanes of minor to moderate intensity per decade. Using this
rationale, it can be expected that the reefs of La Parguera will suffer the effects of
cyclonic disturbances on the average of one to two per decade.
Major hurricanes affecting Puerto Rico are very rare. Hurricanes San
Ciriaco of 1899 and San Felipe of 1928 are known to have caused widespread
destruction to the island. San Felipe (1928) is claimed to have been the worst
39
hurricane affecting the southwestern coast of Puerto Rico (Cote, personal
communication). After these two hurricanes David (1979) is perhaps the worst
hurricane affecting the La Parguera area in the last 100 years. This suggests that
the frequency of major hurricanes affecting the reefs of La Parguera is between
two and three per century.
IV. Human interference
In the central reef-flat area of Cayo Enrique there is a patch of dry land
within the mangroves where a single tree of Avicennia germinans is living along
with nearly a dozen damaged Laguncularia racemosa trees. Both species lack
many branches and leaves and there is evidence that they were cut for firewood or
other purposes. Be-cause there are few leaves and branches remaining, and
because they are nearly covered by dense Rhizophora foliage, Avicennia and
Laguncularia do not appear in the aerial color infrared photograph of Cayo
Enrique. The colonization and establishment of these mangroves in Cayo Enrique
appears to be limited by man's activities and not by natural factors.
Cayo Enrique is frequently visited by pleasure boats and yachts that stay
up to 3-4 days at anchor in the calm sandy lagoon. No detrimental effects on the
reef have been observed that can be related to boating activities. Boats tend to
avoid Thalassia beds in favor of sandy areas that provide a better holding ground
for anchoring. Destruction of Thalassia caused by boat propellers in shallow bays
has been reported by Phillips (1960). This was not observed in Cayo Enrique,
probably due to the deeper distribution of Thalassia there.
Local fishermen frequently walk on the reef-flat in search of octopus and
40
other edible mollusks. Although these activities could potentially destroy fragile
corals such as Porites furcata and other reef organisms, its real effect is yet to be
determined.
Oil pollution, in the form of tar balls, was observed on the boulder ramparts
during the last two years. This could represent, if it continues and increases over
the years, a major source of pollution and an unnecessary stress upon the reef
environment.
In just a decade, human and industrial wastes caused the death of fringing
and patch reefs off the coast of Venezuela (Weiss and Goddard, 1977). The main
causes of pollution there were particulate and soluble organic wastes that entered
the water by dumping, drainage, and sewage. The Parguera area is beginning to
suffer problems similar to the ones
reported from Venezuela. Numerous "casetas" (wooden houses on stilts) are built
along the coast among the mangroves. These are the main source of nutrients that
enter the water by the dumping of sewage and household detergents. As a result
of this, the coastal Thalassia beds are mostly covered by floccules of the blue-
green algae Microcoleus lyngbyaceus, an indicator of organic pollution. The
concentration of the coliform bacteria in these waters has been found to be up to
200 bacteria/ml (Imam, personal communication). According to the EPA Water
Quality Criteria (1974), this value is 100 times the maximum acceptable limit for
water contact sports. Destruction of Thalassia by boat propellers is common along
the shallow, coastal seagrass beds.
Sediment resuspension by boat traffic creates high levels of turbidity that
decreases the amount of light required by Thalassia for photosynthesis. Turbidity is
41
highest during weekends when boat traffic reaches its maximum. These effects are
restricted to the nearshore coastal area of La Parguera due to the prevailing winds
and currents.
Cayo Enrique and the outer reefs, located upcurrent and windward of the
pollution source, appear not to be affected by these factors. However, some
recreational activities, if unregulated, can potentially damage the reef community.
These include overfishing, shell and coral collecting, cutting of mangroves, and
dumping of sewage and garbage by visiting pleasure boats. As this reef, and the
other reefs of La Parguera, become more accessible to a larger number of persons,
the effects of human interference will influence, to a greater extent, the future
health of the reef.
Theoretical aspects and future trends
The main ecological concept considered here is whether Cayo Enrique can
be considered a high-diversity community and how this diversity is maintained. The
concept of reefs as highly diverse communities evolved mostly from studies of the
rich coral assemblages of fore-reef areas. Theories explaining the existence of high
local diversity in coral reefs and tropical rain forests are discussed by Connell
(1978).
Before Hurricane David, the diversity, as measured by Shannon and
Weaver (1949) on the fore-reef of Cayo Enrique, was .18 for the Acropora palmata
zone and .51 for the mixed zone, with an evenness of .12 and .32 respectively
(Goenaga and Cintron, 1979). These low diversity and evenness values indicate
high dominance by A. palmata and, in the mixed zone, by A. cervicornis. A slightly
42
higher diversity and evenness value in the mixed zone indicates the presence of
other corals not found in the previous zone. Goenaga and Cintron (1979) also
report Agaricia agaricites, Montastrea annularis, and Diploria sp. in the mixed coral
zone.
A year and a half after Hurricane David, along with the cumulative
effects of Hurricane Allen, a re-survey was done on the same fore-reef zones of
Cayo Enrique. Species diversity in the A. palmata zone increased to 1.33 since
the stress, while the value for the mixed coral zone was .79. Evenness values
were .96 and .46 respectively. The A. palmata zone increased its diversity
substantially from .18 to 1.33. The evenness value for this zone (.96) indicates
high evenness, or least dominance by any species. While the diversity for the
mixed zone increased slightly to .79, the evenness remained low (.46), indi-
cating moderate to high dominance. In this case, the massive coral Montastrea
annularis, representing 79% of the living coral cover, was the dominant species.
The results discussed here were obtained using 10 m transects
made parallel to depth contours at two and four meters depth on the middle
fore-reef zone of Cayo Enrique. These results are thus preliminary and
suggest only that hurricane disturbances are capable of increasing the
diversity of shallow fore-reef areas.
Qualitative observations on the mixed zone of Cayo Turrumote made two
years after Hurricane David leads to similar conclusions. This zone is deeper (10
m) and was exposed to heavier wave action than Cayo Enrique during the
hurricanes. A total of 13 species of juvenile Scleractinian corals were found in an
area that was dominated by A. cervicornis before Hurricane David. The larvae of
43
these corals settled on the newly available substrate composed of dead A.
cervicornis branches increasing the species richness and, undoubtedly, its diver-
sity.
Moderate hurricane disturbances can increase the species diversity of a
reef by interrupting the competitive process that leads to dominance by one or few
species and by providing new substrata (a major limiting factor) for the settlement
of new coral recruits. Major disturbances can drastically prolong the recovery
process. Recovery periods for coral reefs can range from a few years to several
decades, depending on the magnitude of the storm and the distance from its cen-
ter. Ten years after Hurricane Hattie, Stoddart (1974) found little or no recovery on
reefs subjected to massive damage, while complete recovery had taken place in
little or moderately damaged reefs. He suggests that "there is a threshold of
damage beyond which storm effects are likely to be prolonged." Recovery in
severely damaged reefs will largely depend upon recruitment by larvae from a
distant source, while, on moderately damaged reefs, a high survivorship of coral
fragments may explain the rapid recovery of reefs (Highsmith et al., 1980). They
propose a model in which periodically disturbed reefs will have the highest long-
term calcification and growth rates.
Periodic hurricane disturbances of moderate intensity are important
factors in maintaining high species diversity in fore-reef areas. In the reef-flat
and back-reef areas, moderate disturbances have fewer effects in the
community structure and species diversity than in fore-reef areas.
The rate by which seagrasses are covering lagoon areas, and the extent of
mangrove colonization on the reef flat, may indicate that Cayo Enrique is
44
approaching a climax zonation dominated primarily by plant species. The reef-flat
and lagoon areas are becoming dominated by two species, Rhizophora mangle and
Thalassia testudinum. On a large scale, this represents a low diversity equilibrium
condition for the reef in general. However, when viewed on a small scale, both man-
grove and seagrass communities increase the species diversity of the reef by
stabilizing the seabed and increasing the surface area available for marine
organisms to live. This biotic substrate extension supports diverse epiphytic algal,
fish, and invertebrate assemblages by providing either food, substrate, shelter, or a
combination of these. Dominance by Rhizophora and Thalassia, and whether this
dominance increases or decreases the total diversity of the reef, is largely a problem
of scale.
During Hurricane David, coral boulders were transported to the reef-flat and
lagoon areas, where they provided new substrate for othercorals to settle. An actual
example of this occurred after Hurricane Beulah when the reef flat in Cayo Laurel
was extended laterally 0.5 m (Glynn, 1973, p. 306). He proposes that leeward
expansion of the reef flat could be a result of deposits becoming stabilized and
providing substrate for new coral growth. Davies (1977) goes one step further,
proposing that under the present hydraulic regime, reefs in the Great Barrier Reef
are growing backwards (i.e. on their lee sides). He claims that windward reef
extension is unlikely and that spur and groove formations, originally described as of
constructional origin (Goreau, 1959; Shinn, 1963), could represent the original front
of an eroding reef.
The future of Cayo Enrique, based on observed trends in the past 44 years,
and if unaffected by man, will largely depend on the occurrence and frequency of
45
physical disturbances. Hurricanes have been the main agents of change for the
reef zones and features of Cayo Enrique. Although major hurricanes can be
catastrophic and prolong the recovery process, they are rare--on the order of 2-3
per century. Moderate hurricane disturbances are more common and actually
benefit the reef in many ways. They increase the species diversity of the fore-reef,
and, to a lesser extent, of the reef-flat and back-reef lagoon areas. The rate at
which mangroves and seagrasses are colonizing available substrate may lead to a
climax zonation until a major disturbance interrupts this process. Long term
transport of coral colonies to leeward shores by hurricane waves provides the
necessary substrate for-the establishment of "patch reefs" or coral communities.
This, and theslow but definite expansion of the reef flat, indicates that Cayo Enrique
could also be growing backwards.
46
CONCLUSIONS
Two major communities of Cayo Enrique, the reef-flat Rhizophora mangle and the
lagoonal Thalassia testudinum areas, showed the maximum variations of all reef zones over
the last 44 years as was determined by a chronological series of aerial photographs. The rate
of increase of these biotic macroassemblages suggests that Cayo Enrique is approaching a
climax zonation dominated primarily by plant species. Hurricane disturbances of moderate
intensity are an important factor in maintaining high species diversity in fore-reef areas, and,
to a lesser extent, in reef-flat and lagoonal areas. Moderate disturbances also affect the reef
by depositing dead corals on the reef-flat, forming boulder ramparts that can become
colonized by mangroves. Hurricanes also adversely affect mangrove areas, but appear not to
affect lagoonal seagrass beds significantly. A new transport of corals to leeward by hurricane
waves enhances the expansion of the reef flat and the establishment of coral communities on
the back-reef lagoon.
Human activities in Cayo Enrique appear to have no significant effects on the reef
environment. Whether this can also be said in fu-ture years will depend on whatever
measures are taken today to protect this reef, as well as the other reefs of La Parguera,
from future ad-verse direct and indirect effects of man.
The use of aerial photoanalysis in coral reef studies provides a mechanism by
means of which reefs can be studied over a long time period. Black and white film is the
easiest and most widely used film in aerial surveys. Color film offers no significant
advantages over black and white film except for greater water penetration. False-color
infrared film is best for differentiating shoreline and reef-flat man-grove vegetation. Its
major disadvantage is that water readily absorbs near-infrared radiation, preventing
submerged vegetation from being recorded on this film. High-altitude aerial and satellite
photography of coral reefs have the advantage of covering large areas in a single
47
photographic frame at the expense of not effectively resolving individual reef features and
zones. I propose a combination of high-altitude and low-altitude photography of coral reefs
to maximize the amount of information obtained from this remote sensing technique.
When available, a chronological series of aerial photographs is
a useful tool in detecting and measuring trends in coral reefs and provides baseline data by
which present and future comparisons can be made.
48
LITERATURE CITED Almy, C. C., and C. Carrion-Torres. 1963. Shallow-water stony corals of Puerto Rico.
Caribbean Journal of Science 3: 133-162. Ball, M. M., E. A. Shinn, and K. W. Stockman. 1967. The geologic effects of Hurricane
Donna in South Florida. Journal of Geology 75: 583-597. Boulon, R. H., Jr. 1980. Patterns of coral community structure and species diversity of a
submerged shelf edge reef off south-western Puerto Rico. M.S. thesis, University of Puerto Rico. 61 pp.
Camp, D. K., S. P. Cobb, and J. F. van Breedveld. 1973. Overgrazing of seagrasses by a
regular urchin, Lytechinus variegatus. Bioscience 23: 37-38. Connell, J. H. 1978. Diversity in tropical rain forests and coral reefs. Science 199:
1302-1310. Craighead, F. C., and V. C. Gilbert. 1962. The effects of Hurricane Donna on the
vegetation of southern Florida. Quarterly Journal, Florida Academy of Science 25: 1-28.
Davies, P. R. 1977. Modern reef growth--Great Barrier Reef. Proceedings, Third
International Coral Reef Symposium 2: 325-330. Flood, P. R., and J. S. Jell. 1977. The effect of Cyclone "David" (January, 1976) on the
sediment distribution patterns on Heron Reef, Great Barrier Reef, Australia. Proceedings, Third Inter-national Coral Reef Symposium 2: 119-125.
Glynn, P. W. 1968. Mass mortalities of echinoids and other reef flat organisms coincident
with midday, low water exposures in Puerto Rico. Marine Biology 1: 226-243. _______. 1973a. Ecology of a Caribbean coral reef. The Porites reef-flat biotope: Part I.
Meteorology and hydrography. Marine Biology 20: 297-318. _______.1973b. Ecology of a Caribbean coral reef. The Porites reef-flat biotope: Part II.
Plankton community with evidence for depletion. Marine Biology 22: 1-21.
49
Glynn, P. W., L. R. Almodovar, and J. G. Gonzalez. 1964. Effects of Hurricane Edith on marine life in La Parguera, Puerto Rico. Caribbean Journal of Science 4: 335-345.
Goenaga, C., and G. Cintron. 1979. Inventory of the Puerto Rican coral reefs. Report
submitted to the coastal zone management of the Department of Natural Resources, Commonwealth of Puerto Rico. 190 pp.
Goreau, T. F. 1959. The ecology of Jamaican coral reefs: I. Species composition and
zonation. Ecology 40: 67-90. Goreau, T. F., and J. W. Wells. 1967. The shallow-water Scleractinia from Jamaica: Revised
list of species and their vertical distribution range. Bulletin of Marine Science 17(2): 442-453.
Highsmith, R. C., A. C. Riggs, and C. M. D'Antonio. 1980. Survival of hurricane-generated
coral fragments and a disturbance model of reef calcification/growth rates. Oecologia 46: 322-329.
Hopley, D. 1977. Aerial photography and other remote sensing techniques. Pages 23-44
in D. R. Stoddart and R. E. Johannes, eds. Handbook of Coral Reef Research Methods. UNESCO Monographs on Oceanographic Methodology.
Hopley, D., and A. V. Steveninck. 1977. Infrared aerial photography of coral reefs.
Proceedings, Third International Coral Reef Symposium 2: 305-311. Kelly, M. G. 1970. Patterns of distribution of coastal biota, remote sensing, and
conservation of resources. Pages 105-113 in W. G. Weist and P. E. Greeson, eds. Hydrobiology. Miami Beach, Fla.
Klim, D. G. 1969. Interactions between seawater and coral reefs in Kaneohe Bay, Oahu,
Hawaii. University of Hawaii, Hawaii Institute of Geophysics, report No. 69-19. Kumpf, H. E., and H. A. Randall. 1961. Charting the marine environments of St. John,
U.S. Virgin Islands. Bulletin of Marine Science 11: 543-551. Loya, Y. 1976. Effects of water turbidity and sedimentation on the community structure of
Puerto Rican corals. Bulletin of Marine Science 26(4): 450-466. Maertz, A., and M. Paul. 1950. A Dictionary of Color. McGraw-Hill, New York, 2nd ed.
50
Matthews, S. A. 1974. Sediment reworking rates and effects of three burrowing organisms of La Parguera, Puerto Rico. M.S. thesis, University of Puerto Rico, Mayaguez.
Morelock, J., and S. Matthews. 1973. Bioturbation of carbonate reef sands. National
Geographic Society Research Reports, vol. 14. Morelock, J., N.-Schneidermann, and W. R. Bryant. 1977. Shelf reefs, southwestern
Puerto Rico. American Association of Petroleum Geologists, Studies in Geology, No. 4: 17-25.
Odum, H. T., and E. P. Odum. 1955. Trophic structure and productivity of a windward coral
reef community on Eniwetok Atoll. Ecological Monographs 25: 291-320. Ogg, J. G., and J. A. Koslow. 1978. The impact of Typhoon Pamela (1976) on Guam's coral
reefs and beaches. Pacific Science 32(2): 105-118. Oppenheimer, C. H. 1963. Effects of Hurricane Carla on the ecology and distribution of the
Florida seagrasses. Professional Papers Service, Florida Board of Conservation 2: 1-72.
Perkins, R. D., and P. Enos. 1968. Hurricane Betsy in the Florida-Bahama area--
geologic effects and comparison with Hurricane Donna. Journal of Geology 76: 710-717.
Phillips, R. C. 1960. Observations on the ecology of the Florida seagrasses. Professional
Papers Service, Florida Board of Conservation 2: 1-72. Pielou, E. C. 1966. The measurement of diversity in different types of biological
collections. Journal of Theoretical Biology 13: 145-163. Pressick, M. L. 1970. Zonation of stony corals of a fringing reef southeast of Icacos
Island, Puerto Rico. Caribbean Journal of Science 10: 137-140. Randall, R. H., and L. G. Eldredge. 1977. Effects of Typhoon Pamela on the coral reefs of
Guam. Proceedings, Third International Coral Reef Symposium 2: 525-531. Rathbun, R. 1879. Brazilian corals and coral reefs. American Naturalist 13: 535-551. Rogers, C. S. 1977. A response of a coral reef to sedimentation. Ph.D. dissertation,
University of Florida.
51
Shannon, C. E., and W. Weaver. 1949. The Mathematical Theory of Communication. University of Illinois Press, Urbana, Ill.
Shinn, E. A. 1963. Spur and groove formation of the Florida reef tract. Journal of Sedimentary
Petrology 33: 291-303. Steers, J. A. 1945. Coral reefs and air photography. Geographical Journal 106: 232-235. Stoddart, D. R. 1962. Catastrophic storm effects on the British Honduras reefs and cays.
Nature 196(4854): 512-515. _______. 1963. Effects of Hurricane Hattie on the British Honduras reefs and cays.
October 30-31, 1961. Atoll Research Bulletin, Washington, D.C. 95: 1-142. _______. 1971. Coral reefs and islands and catastrophic storms. Pages 155-197 in J. A.
Steers, ed. Applied Coastal Geomorphology. McMillan, London. _______. 1974. Post-hurricane changes on the British Honduras reefs: re-survey of
1972. Proceedings, Second International Coral Reef Symposium 2: 473-483. Tabb, D. C., and A. C. Jones. 1962. Effect of Hurricane Donna on the aquatic fauna of
North Florida Bay. Transactions, American Fisheries Society 91(4): 375-378. Teichert, C., and R. W. Fairbridge. 1948. The Low Isles of the Great Barrier Reef: a new
745. Thomas, L. P., D. R. Moore, and R. C. Work. 1961. Effect of Hurricane Donna on the turtle
grass beds of Biscayne Bay, Florida. Bulletin of Marine Science Gulf Caribbean 11(2): 191-197.
U.S. Department of Commerce Coast and Geodetic Survey. 1929. Coast Pilot--West
Indies. U.S. Government Printing Office, 3rd ed., serial no. 446, 213 pp. Vaughan, T. H. 1901. Stony corals of the Porto Rican waters. U.S. Fish Commission, Bulletin
for 1900. Part 2, pp. 289-320. Vermer, D. E. 1963. Effects of Hurricane Hattie, 1961, on the cays of British Honduras.
Zeitschrift fur Geomorphologie 7(4): 332-354. Verrill, A. E. 1869. Comparison of coral faunae of the Atlantic and Pacific coasts of the
Isthmus of Darien. American Naturalist 3: 499-500.
52
Wadsworth, F. H., and G. H. Englerth. 1959. Effects of the 1956 hurricane on forests in Puerto Rico. Caribbean Forester January-June, pp. 38-51.
Wallace, R., T. Morgan, M. Silverstone, J. Morelock, and N. Schneidermann. 1973. Cayo
Enrique sediments and samples. Proceedings of the Association of the Island Marine Laboratories of the Caribbean 10: 30; abst.
Weiss, M. P., and D. A. Goddard. 1977. Man's impact on coastal reefs--an example from
Venezuela. Pages 111-124 in S. H. Frost, M. P. Weiss, and J. B. Saunders, eds. Reefs and related carbonates--ecology and sedimentology. The American Association of Petroleum Geologists, Tulsa, Oklahoma.
53
Appendix. A list of hurricanes, with their tracks, that have been reported in the literature
or in this study as hurricanes affecting La Parguera reefs.