CHROMOSOME NUMBERS, KARYOTYPE AND CROSSABILITY OF SOME MANIHOT SPECIES A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN HORTICULTURE MAY 1972 By Effiong Etim Umanah Thesis Committee: Richard W. Hartmann, Chairman Yoneo Sagawa James C. Gilbert Douglas J. C. Friend
Transcript
C HRO M O S O M E NUMBERS, K A R Y O T Y P E AND C R O S S A B I L I T Y
OF SOME M A N I H O T SPECIES
A THESIS S U B MITTED TO THE G R A D U A T E D I V I S I O N OF THE U N I V E R S I T Y OF HAWAII IN PARTIAL F U L F I L L M E N T
OF THE R E Q U I R E M E N T S FOR THE DEGREE OF
M A S T E R OF SCIENCE
IN H O R T I C U L T U R E
M A Y 1972
By
Effiong Etim Umanah
Thesis Committee:
R i c h a r d W. Hartmann, C h a irman Yoneo Sagawa J ames C. Gilb e r t Douglas J. C. Friend
We certify that we have read this thesis and that in
our opin i o n it is s a t i s f a c t o r y in scope and quality as a
thesis for the degree of M a s t e r of Science in H o r t i c u l t u r e
THE S I S COM M I T T E E
Cha irman
PAGE
L I S T OF T A B L E S ................................................... iv
L I S T OF I L L U S T R A T I O N S ........................................ v
I N T R O D U C T I O N ...................................................... 1
R E V I E W OF L I T E R A T U R E ........................................... 3
M A T E R I A L S AND M E T H O D S ........................................ 8
R E S U L T S ........................................................... 13
C h r o m o s o m e numbers and karyo t y p e s ................... 13K a r y o t y p e of M a n i h o t esculenta ..................... 13K a r y o t y p e of M a n i h o t g l a z i o v i i ..................... 14P o l l i n a t i o n experi m e n t s ................................ 16O b s e r v a t i o n s .............................................. 17Po l l e n p h y s i o l o g y ........................................ 18
D I S C U S S I O N ........................................................ 33
P o l y p l o i d nature of M a n i h o t s p e c i e s ................ 34F e r t i l i t y of selfs and crosses ..................... 38Pollen p h y s i o l o g y of M a n i h o t s p e c i e s ............. 40
S U M M A R Y ........................................................... 44
A P P E N D I X ........................................................... 45
L I T E R A T U R E CITED ................................................ 46
T ABLE OF CONTEN TS
TABLE D E S C R I P T I O N PAGE
1 M a n i h o t species and va r i e t i e s used inc ytological studies ................................ 9
2 C h r o m o s o m e numbers in M a n i h o t species . . . . 19
3 M e a s u r e m e n t s of somatic c hromosomes inM a n i h o t esculenta ................................... 20
4 M e a s u r e m e n t s of somatic chromo s o m e s inMa n i h o t g l a z i o v i i ................................... 21
5 C o m p a r i s o n of chromosome data of M a n i h o ts p e c i e s .................................. ! ! ! . . . 22
6 Fruit set, seed set and g e r m i n a t i o n ofseeds from reci p r o c a l i n t e r s p e c i f i c and i n t r a s p e c i f i c crosses and selfs ................ 23
7 S t a i n a b i l i t y , g e r m i n a b i l i t y and size ofM a n i h o t species pollen ........................... 24
L I S T OF TABLES
LI S T OF I L L U S T R A T I O N S
PLATE FIGURE D E S C R I P T I O N PAGE
Somatic chromosomes of M a n i h o t species p r e t r e a t e d with8 - o x y q u i n o 1 ine (1650x) 26M a n i h o t g l a z i o v i i (ceara rubber)
2 M. esculenta v a r . Itu3 M. esculenta f. v a r i e g a t e d4 M. es culenta var. A m a r i l l o5 M. esculenta var. ? ex El Salvadore6 M. esculenta var. Ceiba7 M. esculenta v a r . Ma m e y a8 M. esculenta v a r . N ina9 M. esculenta var. Pata de Palona
Somatic chromo s o m e s of M a n i h o tspecies with p r e t r e a t m e n t andw i t h o u t p r e t r e a t m e n t with8 - o x y q u i n o l i n e and some meio t i cc o n f i g u r a t i o n s in m i c r o s p o r o g e n e s i sof M a n i h o t spp ( 1 6 5 0 x ) ................... 28
10 M. esculenta var. Seda11 M. esculenta var. Puna cassava12 M. esculenta var. ?13 M. g l a z i o v i i14 M. esculenta17 Early A n a p h a s e I in M. esculenta18 M e t a p h a s e II (polar view) in
M. e s c ulenta (n = 18)19 M e t a p h a s e I in M. g l a z i o v i i 18II20 M e t a p h a s e II in M. gla z i o v i i (polar
view) n = 18
K a r y o t y p e s of M a n i h o t species ( 5 5 0 0 x ) . 3015 M. esculenta16 M. g l a z i o v i i
V e g e t a t i v e m o r p h o l o g y of M a n i h o t s p e c i e s .................... "T ! T . . . 32
21 M a t u r e plant of M. g l a z i o v i i withflowers and fruits (O.lx)
22 Young potted plant of M. g l a z i o v i i (O.lx)2 3 M. esculenta f . v a r i e gated (O.lx)24 M. esculenta var. Itu with
flowers and fruits (O.lx)
I N T R O D U C T I O N
The genus M a n i h o t is important beca u s e of the species
M a n i h o t esculenta Crantz. This species is b e l i e v e d to have
o r i g i n a t e d from Brazil and Central A m e r i c a (Pierre Martyr,
1494; Barnes, 1954; Rogers, 1964; and Schwerin, 1970).
P u r s e g l o v e (1968) indicates that it was g rown as a crop in
P eru some 4000 years ago and in M e x i c o 2000 years ago and that
it is not known in a w ild state. It is a w i d e l y cult i v a t e d
tropical and subtropical root crop known by such names as
cassava, ma n i o c or tapioca in different parts of the world.
It is used as a v e g e t a b l e in the areas of its c u l t i v a t i o n and
as a source of high quality starch for industrial processes
of vari o u s types.
A l t h o u g h some b r e e d i n g w ork for improved yield and quality
of the tubers has b een done, no study has b een made of the
c y t o g e n e t i c s of the species in relation to other species of
the genus and hence i n t e rspecific r e l a t i o n s h i p s of the genus
for p u r poses of inter s p e c i f i c h y b r i d i s a t i o n and i n c o r p o r a t i o n
of desirable genes for specific economic characters and disease
resistance. W h e n c h romosome m o r p h o l o g y is compared in the
different taxa, e s pecially at the species and lower levels, an
estimate of the r e l a t i o n s h i p of these taxa may be obtained.
Thus an evaluation of the c omparative k a r y o m o r p h o l o g y among
the various taxa is desirable. This n e c e s s i t a t e s the precise
id e n t i f i c a t i o n of each of the pairs of chromosomes.
This type of study will indicate to the plant bree d e r the
r e l a t i o n s h i p s to M. esculenta of other species of the genus
and the p r o b a b i l i t y of o b taining some d e s i r a b 1 e r e s u 1 ts from
inter s p e c i f i c crosses w h i c h are a p r e r e q u i s i t e in incorporating
d e sirable genes in the commercial varieties. F u r t h ermore,
this type of study may open the way to locating m a r k e r genes
on the different linkage groups of this species and also the
g e n etics of i nheritance of economic characters in a d d i t i o n to
ass i s t i n g in a p h y l o g e n e t i c c l a s s i f i c a t i o n of the t a x a .
2
T a x o n o m y of M a n i h o t
The genus M a n i h o t of the family E u p h o r b i a c e a e was
d e s c r i b e d by Pax (1910) to contain 128 species, Rogers and
A p p a n (1970) made e x tensive exp l o r a t i o n s and collec t i o n s of
the various species in their native habitat w hich extends
from southern A r i z o n a in the United States of Amer i c a through
M e x i c o and South A m e r i c a to n o r t h e r n Argen t i n a . They based
their c l a s s i f i c a t i o n on the m o d e r n species concept of a
closed gene pool, ecol o g i c a l and g e o g r a p h i c a l distribution.
C o n s e q u e n t l y they r e c o g n i z e 75 species and a number of sub
specific taxa in the genus. They point out that several of
Pax's species were synonyms of one or another of the species,
since o u t c r o s s i n g is the b r e e d i n g system of the genus and
the plants are, therefore, very h e t e r o z y g o u s and h e t e r o g e n e o u s
They found at least 10 different names given at one time or
another to M a n i h o t esculenta.
C h r o m o s o m e numbers of M a n i h o t
REVIEW OF L I T E R A T U R E
Perry (1943) d e t e r m i n e d the c h romosome numbers of 90
species and v a r i e t i e s of 19 genera of the family E u p h o r b i a c e a e
using a c o m b i n a t i o n ofvarious methods and materials. He
showed that the c hromosome number in the family ranged from
2n = 12 in Eup h o r b i a dulcis to 2n = ca. 224 in A c a l y p h a
w i l k s i a n a and suggested that the primary basic number for the
family is 8 with secondary basic numbers of 6, 7, 9, 10, and
11. For the genus M a n iho t he reported a basic number of 9
with 2n = 36 in all the five species and three v a rieties
examined. Gra n e r (1935, 1941) and B o i t e a u (1938, 1941) made
c hro m o s o m e counts of one species and three species r e s p e c t i v e
ly of M a n iho t and arrived at different numbers. Wher e a s
Gr a n e r o b t a i n e d a somatic number of 2n = 36 for all the
v a r i e t i e s of one species examined, B o i t e a u o b t a i n e d somatic
numbers of 2n = 4, 8, 16, 24, and 32 for the three species
examined. P erry thus seems to confirm G r a n e r ' s results while
Bo i teau's results seem erroneous. H o w e v e r chrom o s o m e karyo-
logy of the genus Ma n iho t has not been r e p o r t e d in the
1 itera t u r e .
I n t e r s p e c i f i c h y b r i d i s a t i o n in the genus M a n i h o t
A t t e m p t s at i n c o rporating genes for m o s a i c disease
r e s i s t a n c e in M a n i h o t esculenta and higher prot e i n contents
of the tubers n e c e s s i t a t e d i n t e rspecific h y b r i d i s a t i o n in the
genus M a n i h o t . The fact that Brandes and Sartoris (1936)
s u c ceeded in transferring genes for m o s a i c virus r e s i s t a n c e
from S a ccharum s p o n t a n e u m to S. o f f i c i n a r u m by i n t e rspecific
h y b r i d i s a t i o n of S . offic ina r urn x S . spon t a n e u m s t i m u l a t e d the
interest of cassava b r e e d e r s in a t tempting to carry out a
similar h y b r i d i s a t i o n program. Both crops are v e g e t a t i v e l y
propagated.
K och (1934) a t t e m p t e d interspecific h y b r i d i s a t i o n of
M a n i h o t esculenta and M. g l a s i o v i i . His M. g l a z i o v i i x
4
M. esculenta cross set no seeds but the r e c i p r o c a l cross set
3.2 percent seeds. The Fi plants were much like M. g l a z i o v i i .
N i c h o l s (1947) carried out both intra- and inter- specific
crosses. In M. esculenta x M. esculenta v a r i e t a l crosses,
seed set ranged from 0 - 5 6 percent. In M. esculenta x
M. g l a z i o v i i he r e p o r t e d 1.8 percent seed set and only 0.4
perc e n t in the r eciprocal cross. In M. dichotoma x M.
glaz i o v i i , 10.7 percent seed set was o b t a i n e d w h e r e a s in the
r e c i p r o c a l cross only 1.8 percent was obtained. Koch was
h o w e v e r not successful in obtaining any seed set in M.
dicho toma x M . esculenta cross. Nich o l s (1947) also carried
out M. saxicola x M. esculenta cross and o b t a i n e d very low
seed set. He was a t t e m p t i n g to breed for hi g h e r protein
content in the tubers.
B olh u i s (1953) surveyed attempts to breed cassava
cultivars wit h a higher prot e i n content in the roots. He
found tha t M. saxicola freely crossed with M. esculenta.
P r o t e i n content was higher in Fl seedling r o o t s , 0.67 - 1.32
percent dry matter, but this was not m a i n t a i n e d in the clones
w h e n p r o p a g a t e d v e g e t a t i v e l y . H o w e v e r high HCN content was
m a i n t a i n e d in the clones. J e n n i n g s (1957) carried crosses of
M . glaz iovii x M. esculenta to advanced generations. The
b a c k c r o s s to M. esculenta gave better yields in the 3rd
b a c k c r o s s than the Fl plants but r e s i s t a n c e to cassava mo s a i c
virus was low. At b e.s t field r esistance was obtained.
5
M. dichotoma x M, e s c ulenta gave very low seed set and even
after 3 b a c k c r o s s e s the roots were still fibrous and woody.
He o b t a i n e d best results w h e n the F4 were crossed with the
3rd b a c k c r o s s of M. g l a z i o v i i x M. esculenta in both yield
and r e s i s t a n c e to virus mosaic.
M. catingea x M. e s c ulenta gave tree-like FI plants
similar to the M. g l a z i o v i i x M. esculenta Fl h y b r i d s with
w o o d y n o n - t u b e r o u s roots and m i l d transient m o s a i c symptoms.
The Fl hybrids were crossed to r e sistant clones of M. glazio-
vii X M. m e l a n o b a s i s so that plants with improved tuber
y i e l d s were obtained. In general no truly r e s i s t a n t s elec
tions were o b t a i n e d from these series of i n t e r s p e c i f i c
hybr idisat ions .
A b r a h a m (1957) carried out a series of i n t e r s p e c i f i c
crosses b e t w e e n M. esculenta and M. g l a z i o v i i . His
M. g l a z i o v i i x M . esculenta cross set no seeds but seed set
of about 1.0 percent was o b t a i n e d in the r e c i p r o c a l crossi
By the 4th b a c k c r o s s to M. esculenta i n d i v iduals with improved
tuber yields were obtained. J e n n i n g s (1959) showed that
M. m e l a n o b a s i s and M. esculenta h y b r i d i s e d easily and the
hybr i d s were fertile. In vie w of the r e a d i n e s s with w hich the
two species h y b r i dised, he doubted w h e t h e r their s eparation
as distinct species was justified. Bolhuis (1953) reac h e d
the same c o n c l u s i o n for M. saxicola and M. esc u l e n t a w here
the inter c r o s s i n g was also easy.
6
H o w e v e r in most of the above crosses the p e r c e n t a g e seed
set was low. A similar s i t uation was o b s e r v e d by Stout and
Clark (1924) in potato. They a t t r i b u t e d the re d u c t i o n in
fertility to the complete r e p l a c e m e n t of seed p r o p a g a t i o n by
v e g e t a t i v e m eans and hence no selection p r e ssure for fertility
in the course of evolution. They concluded, "since potato is
c l o nally propagated, selection and evo l u t i o n have tended to
be a g a i n s t seed set as a result of internal r e g u l a t i o n of
d e v e l o p m e n t and the influence of v e g e t a t i v e vigor may result
in a syst e m a t i c or p l e t h o r i c sterility w h i c h in the case of
p otato seems to affect m a l e n e s s more than femaleness and with
time this has become hereditary".
7
M A T E R I A L S AND M E T H O D S
Cuttings of the cassava v a r i e t i e s (Manihot esculenta
Crantz) and ceara rubber (M a n i h o t g l a z i o v i i Muell, Arg.)
used in these studies were c o llected from various parts of
H a w a i i b e t w e e n D e c e m b e r 1970 and M a r c h 1971, (Table 1).
Some of these cuttings were rooted in verm i c u l i t e ,
the rest were rooted in a 2 : 1 : 1 m i x t u r e of v e r m i culite,
sand and peat moss. W here necessary, slow release fertilizer
(Osmocote) was added to promote growth. All rooting was done
in plastic p o t s .
For somatic c h r o m o s o m e studies, large root tips with
slight y e l l o w tinge indicating vigorous growth were selected
and cut about 2 mm in length. Some were p r e t r e a t e d with
0.002 m olar 8 - o x y q u i n o l i n e solution a c c o r d i n g to the m e t h o d
of Tjio and L e v a n (1950) for 3 hours at 18° C. A f t e r this
t r e atment these root tips and others w h i c h were not p r e t r e a t e d
were fixed in Carnoy solution (three parts absolute alcohol,
one part c h l o r o f o r m and one part glacial acetic acid) for 1
hour at 18° C. They were then wa s h e d in d i s t i l l e d w ater and
h y d r o l y s e d with 1 N h y d r o c h l o r i c acid at 60° C for 10 minutes.
Each root tip was m a c e r a t e d on a m i c r o s c o p e slide after
r e m oving the root cap, stained with 1 percent a c e t o - o r c e i n in
45 percent acetic acid and a cover slip applied. Excess
stain was absorbed using b i b ulous paper. A p p l i c a t i o n of
Table 1, M a n i h o t species and v a r i e t i e s used in c ytological studies
S p e c i e s / V a r i e t i e s Source
M. g l a z i o v i i (ceara rubb er)
M. esculenta
1A m a r illoC e iba (C eb ia)M a m e y aNinaPata de Palona SedaPuna cassava
1f. v a r i e g a t e d
W a i m a n a l o Expe r i m e n t Station
W a i m a n a l o Exper i m e n t Station" ex El Salvadore
K a u a i E x p e r i m e n t Station
H a w a i i (Big Island)H o n o l u l uH o n o l u l u
p r e ssure using the w o o d e n end of the needle spread the cells.
The slide was passed through alcohol lamp flame for a few
seconds to destain the cyt o p l a s m and further spread the cells.
The slide was ringed with sticky-wax around the cover slip
to prevent drying out during examination.
For o b s e r v a t i o n of m e i o t i c b e h a v i o r of pollen mother
cells (PMC) of the species, flower buds were c o l lected at
noon and fixed in Carnoy s o l ution (three parts absolute
alcohol, one part glacial acetic acid and one part chloroform)
for 24 hours at room temperature. Each anther from the fixed
buds was placed in a drop of 1 percent a c e t o - o r c e i n in
45 p e r c e n t acetic acid on a m i c r o s c o p e slide, cut into two
h alves and the pollen m o t h e r cells squeezed out of the
anthers using needles. A n o t h e r debris was remo v e d before
coverslip was applied and excess stain a b s o r b e d using
b i b u l o u s paper. The slide was then rapi d l y passed through
alcohol lamp flame to destain the c y t o p l a s m and spread the
cells. The coverslip w a s s e a l e d with Kerr s t i c k y wax.
R oot tips and flower buds which were not used immediately
after fixation were stored in 70 percent alcohol at 4° C in
the r e f r i g e r a t o r and used to prepare slides as required. All
e x a m i n a t i o n s and anafysis of the chromo s o m e s were carried out
on temporary preparations. W i t h the aid of optical c o m b i n a
tions of Zeiss ocular lOx, plan obj e c t i v e lOOx and 1.30 oil
immersion the chromosomes were counted and studied. Using
10
Zeiss p h o t o m i c r o g r a p h i c camera attached to the above
m i c r o s c o p e , p h o t o g r a p h s were taken of cells wit h well spread
ch r o m o s o m e s at 550x on K o d a k High C o n t r a s t Copy film.
To make k a r y o t y p e s the p h o t o m i c r o g r a p h i c neg a t i v e s were
e n l arged to a m a g n i f i c a t i o n of 5500x using a L e i t z V a l o y II
enlarger and line tracing of w h o l e image lengths of the
c hromo s o m e s with sharp poin t e d pencil, on standard w hite paper
was made and the p o s i t i o n of the c e ntromere was indicated
(Stewart, 1 9 4 7 ) . Each c hromosome from the tracing was then
m e a s u r e d with great care using a me t r i c ruler and flexible
wires. The standard dev i a t i o n was c a l c u l a t e d on the basis
of the mea n chrom o s o m e length of each c h romosome type in each
cell. The c h r o m o s o m e s were then a r r anged in order of d e c r e a s
ing m e a n length, the longest being the first and the shortest,
the last. The Form P e r c e n t (F%) was used to r e p r e s e n t the
p e r c e n t a g e of the short arm length to the entire c hromosome
length (Huziwara, 1 9 5 6 ) . In order to compare the relative
length of the various c h r o m o s o m e s in the c o m p l e m e n t the
ratio in perc e n t a g e of the length of individual chrom o s o m e
to that of the longest c h r o m o s o m e in the c o m p l e m e n t was used
(Huziwara, 1956). In this met h o d the length of the longest
chro m o s o m e was taken as 100 and the r e l ative lengths of all
the r e m aining chromosomes were calc u l a t e d in r e l a t i o n to this
value. All k a ryotype analysis was done on u n p r e t r e a t e d
c hromosomes only.
11
Bud p o l l i n a t i o n was carried out both in the field in the
A r b o r e t u m at the E x p e r i m e n t Station, W a i m a n a l o (about 22 miles
from M a n o a campus) and at M a n o a campus on potted plants. Only
three of the plants (M. g l a z i o v i i and esculenta var i e t i e s
Itu and ? ex El Salvadore) under study were p o l l i n a t e d in
the field at W aimanalo. These three together with the other
9 v a r ieties of M. e s c u lenta were grown in pots and poll i n a t i o n s
were carried out at M a n o a campus. The plants b l o o m e d b e t w e e n
M a y and September.
Seed set o b t a i n e d from the crosses and selfings was
de t e r m i n e d about 3 months after p o l l i n a t i o n s w h e n the capsules
were col l e c t e d before seeds dehisced. D e h i s c e n c e of the
capsules o c c u r r e d in the labor a t o r y and seeds were counted.
P e r c e n t a g e success in capsule and seed set was d e t e r m i n e d by
c o mparing the number of fruits and seeds a c t u a l l y c o llected
with the number of p o l l i n a t i o n s made. Each p o l l i n a t i o n was
p o t e n t i a l l y capable of y i e lding three seeds from the trilocu-
lar ovaries. Seed v i a b i l i t y was tested by g e r m i n a b i l i t y .
P o l l e n fertility was a s s e s s e d in vitro by m e a s u r i n g
s t a i n a b i l i t y using te t r a z o l i u m chloride, b e n z i d i n e d i h y d r o
chloride, and cotton blue in lactophenol. A t t e m p t s were
also made at in vitro g e r m i n a t i o n of the pollen, using
Br e w b a k e r and K w a c k (1964) C a - s u p p 1emented m e d i u m (see Appendix).
12
RESU L T S
C h r o m o s o m e numbers and k aryotypes
The somatic c h romosome c omplement of all the species and
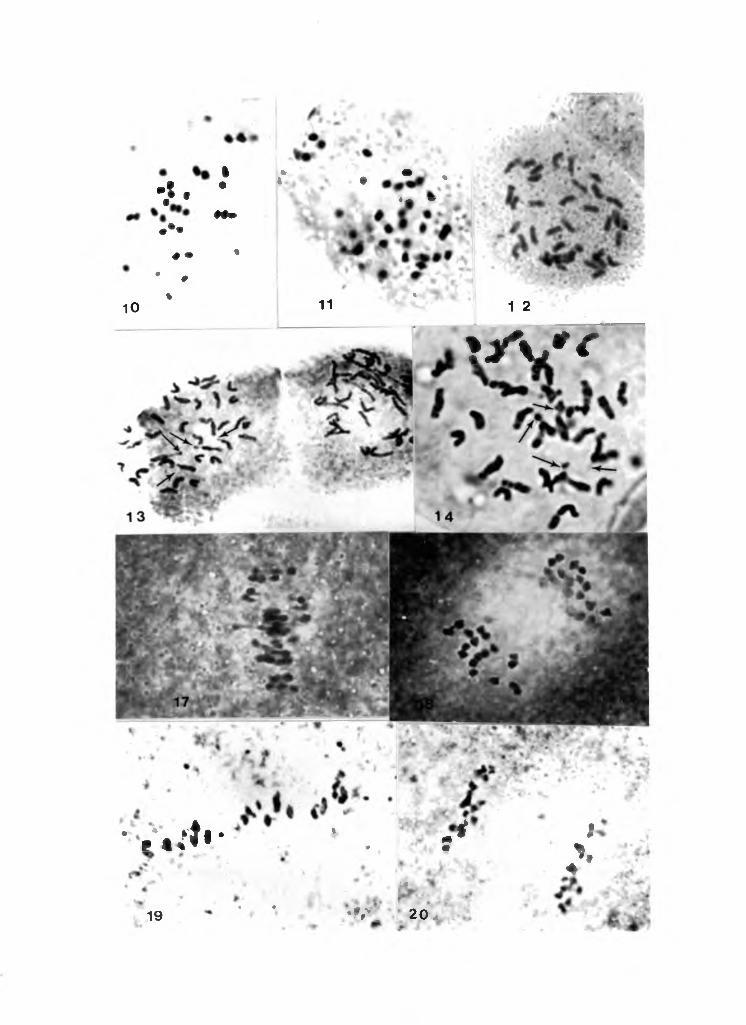
v a r i e t i e s studied is 2n = 36 (Fig. 1 - 12). The chromo s o m e s
on the average are small, ranging in M a n i h o t esculenta from
1.6 to 4.0 m i c r o n s (Table 3) and in M. g l a z i o v i i from 1.4 to
3.8 m i c r o n s (Table 4). B a s e d on the length of the chromosomes,
c e n t r o m e r e position, p r e sence of secondary c o n s t r i c t i o n and
satellites, the chromo s o m e s of each species were divided into
types. The c e n t r o m e r e p o s i t i o n was used to d e termine the Form
P e r c e n t (F7») ; 46 - 50 percent being c l a s s i f i e d as met a c e n t r i c ,
26 - 45 perc e n t as s u b m e t a c e n t r i c , 11 - 25 percent as sub
terminal or a c r o centric, and less than 10 percent as telo
centric, (Huziwara, 1956). Fol l o w i n g M c C l i n t o c k (1929) the
c h r o m o s o m e s were n u m b e r e d starting from the longest to the
shortest.
1. K a r y o t y p e of M. esculenta
The somatic c hromosome c o mplement of this species b ased
on average m e a s u r e m e n t s of chromo s o m e s of one cell each of
11 v a r i e t i e s may be divided into 13 types as follows:
Type I: A pair of longest chromo s o m e s (1,2) 4.0 micronswith s u b m e t a c e n t r i c centromere.
Type II: A pair of chromo s o m e s (3,4) of average length3.3 m i c r o n s wit h a c r o c e n t r i c (s u b t e r m i n a 1) c e n t r o m e r e .
Type III: A pair of c hromosomes (5,6) of length 3.1 micr o n swit h a c r o c e n t r i c centromere.
14
Type IV:
Type V:
Type VI:
Type VII:
Type VIII^
Type IX:
Type X:
Type XI:
Type XII:
Type XIII:
A pair of s atellited c hromosomes (7,8) with s e condary c o n s t r i c t i o n and s u b m e t a c e n t r i c c e n t r o m e r e .
T hree pairs of c h r o m osomes (9,10; 11,12; 13,14)with s u b m e t a c e n t r i c centromeres.
T hree pairs of ch r o m o s o m e s (15,16; 17,18; 19,20)w ith s u b m e t a c e n t r i c centromeres,
A pair of chromosomes (21,22) with a c r o c e n t r i c c e n t r o m e r e ,
A pair of s a t e l l i t e d c h r o m o s o m e s (23,24) with s u b m e t a c e n t r i c c e ntromere and s e c ondary c o n s t r i c tion.
Two pairs of ch r o m o s o m e s (25,26; 29,30) withs u b m e t a c e n t r i c centromeres,
A pair of m e d i u m - s i z e d ch r o m o s o m e s (27,28) w ith m e t a c e n t r i c centromeres,
A pair of small c hromosomes (31,32) with s u b m e t a c e n t r i c centromere,
A pair of short c hromosomes (33,34) with s u b m e t a c e n t r i c centromere,
A pair of shortest chromo s o m e s (35,36) 1,6 micronsin length with m e t a c e n t r i c centromere.
No c h r o m o s o m e with a te l o c e n t r i c c e ntromere was observed,
2. K a r y o t y p e of M, g l a z i o v i i
The somatic chrom o s o m e c o mplement of this species based
on average m e a s u r e m e n t s of ch r o m o s o m e s of two well spread
cells is divided into 13 types as follows:
Type I: A pair of longest c hromosomes (1,2) wit h secondaryc o n s t r i c t i o n and satellites. The c e ntromere p o s i t i o n is s u b m e t a c e n t r i c ,
Type II: A pair of chromo s o m e s (3,4) w ith a c r o c e n t r i ccentromere.
Type III: A pair of c hromosomes (5,6) wit h m e t a c e n t r i cc e n t r o m e r e .
Type IV: A pair of s a tellited chromosomes (7,8) withs u b m e t a c e n t r i c c entromere and secondary c o n s t r i c tion.
Type V: A pair of c hromosomes (9,10) with a c r o c e n t r i cc e n t r o m e r e .
Type VI: Two pairs of chromosomes (11,12; 13,14) witha c r o c e n t r i c centromere.
Type VII: Two pairs of c h r o m osomes (15,16; 17,18) withs u b m e t a c e n t r i c centromere.
Type VIII: Two pairs of c hromosomes (19,20; 21,22) withm e t a c e n t r i c centromeres.
Type IX: Two pairs of chromo s o m e s (23,24; 25,26) withsu b m e t a c e n t r i c centromeres.
Type X: Two pairs of chromosomes (27,28; 29,30) withsu b m e t a c e n t r i c centromeres.
Type XI: A pair of m e d i u m - s i z e d ch r o m o s o m e s (31,32) withs u b m e t a c e n t r i c centromeres.
Type XII: A pair of short c hromosomes (33,34) withs u b m e t a c e n t r i c centromeres.
15
Type XIII: A pair of shortest chromosomes (35,36) withs u b m e t a c e n t r i c centromeres.
No chromo s o m e s with teloce n t r i c centromeres were observed.
The c h r o m osomes of the two species M. esculenta and
M. g l a z i o v i i are similar in gross morphology. Of the 36
chromo s o m e s of M. esculenta 4 have m e t a c e n t r i c centromeres,
26 have s u b m e t a c e n t r i c centromeres, and 6 have ac r o c e n t r i c
centromeres. In M. g l a z i o v i i 6 chromosomes have m e t a c e n t r i c
centromeres, 22 have su b m e t a c e n t r i c centro m e r e s and 8 have
a c r o c e n t r i c c entromeres (Table 5). The m e a n length of the
M. es c u l e n t a c h romosome compl e m e n t is 2.5 m i c r o n s with a rangp
of 4.0 to 1.6 wher e a s in M. gla z i o v i i the mea n length is 2.6
m i c r o n s with a range of 3.8 to 1.4 microns. Two pairs of
ch r o m o s o m e s in each species possess satellites with secondary
c o n strictions. (Fig. 13-14).
M i c r o s p o r o g e n e s i s in the two species was normal
(Fig. 17-20). E i g h t e e n biv a l e n t s were reg u l a r l y observed
at MI and regular d i s j u n c t i o n at AI. M e t a p h a s e II and the
rest of the m e i o t i c phases were normal ending up in the for
m a t i o n of tetrads. Cell wall formation of the m i c r o s p o r e
tetrads was simultaneous. On the basis of these c o m p a rative
data of the two species, they must differ in gene co m p o s i t i o n
which should then account for their m o r p h o l o g i c a l plant
c h aracter d i f f e r e n c e s (Fig. 21-24).
P o l l i n a t i o n experiments
Resu l t s of p o l l i n a t i o n s which were carried out in both
the field at W a i m a n a l o and in potted plants at M a n o a campus
are shown in Table 6, together with g e r m i n a t i o n of the seeds.
The h i g h e s t p e r c e n t a g e capsule set was 42.8 in M. esculenta
i nt e r v a r i e t a 1 cross of Seda x Itu and v a r i e g a t e d x M. g l a z i o v i i
and zero percent where M. g l a z i o v i i was used as a female
parent in a cross with var i e t i e s of M. e s c u l e n t a . R e ciprocal
cross of M. g l a z i o v i i as male parent with v a r i e t i e s of
M. esc u l e n t a as female parents gave co m p a r a t i v e perce n t a g e
capsule set of 14.2, 24.1, and 42.8.
16
V a r i e t i e s of M. esculenta freely i n t e r c r o s s e d among
themselves reciprocally. These results seem to suggest that
r e s t r i c t i o n of r e ciprocal cross betw e e n M. g l a z i o v i i and
v a r i e t i e s of M. esculenta was p r o bably due to m a t ernal
p r o p e r t i e s of M . glaz iovii w h e n M. esculenta was used as
po l l e n parent. This u n ilateral i n c o m p a t i b i l i t y p r o bably
r esults from intera c t i o n of pollen with m a t e r n a l tissues. In
all cases where this i n c o m p a t i b i l i t y was exhibited, it was
o b s e r v e d that the female flowers dropped w i t h i n 5 days after
pollination. In this study no attempt was made to investigate
the causes of the u n i l a t e r a l i n c o m p a t i b i l i t y except that pollen
v i a b i l i t y and size for the species were c o m pared (Table 7).
O b s e r v a t i o n
In general it was o b s erved that there a p p eared to be
p e r i o d i c i t y of flowering in M a n i h o t species. F l owering
started in M a y and c o ntinued through August. By late S e p t e m
ber, all young flower buds dropped p r e m a t u r e l y w i t h o u t r e a c h
ing anthesis. The dropping of the flowers may be closely
c o r r e l a t e d with the drop in temperatures and daylength, during
the Fall through W i n t e r months. M a x i m u m temperatures during
M a y to A u g u s t ranged from 81.7 to 84.6° C in W a i m a n a l o and
83.7 to 88.0° F in M a n o a campus whereas day l e n g t h for the
same period was 12 to 13.5 hours as compared to highest Wi n t e r
temperatures of 76.5° to 79.4° F in W a i m a n a l o and 81.7 to
83.0° F in M a n o a and d a ylength of 10.5 to 12 hours.
17
P o l l e n p h ysiology
R esults of pollen stainability and g e r m i n a b i 1 ity are
r e c o r d e d in Table 7. A l t h o u g h the p e r c e n t a g e of pollen
stained was high in all the stains used, showing different
acti v i t i e s of the pollen, their g e r m i n a b i l i t y was zero
percent. In fact, most pollen grains burst in the g e r m i n a t i o n
m e d i u m w i t h o u t germination.
18
Table 2, Chrom o s o m e numbers in M a n i h o t spp.
19
S p e c i e s / V a r i e t i e s Source 2n n
M a n i h o t esculenta Crantz
Itu W a i m a n a l o Expt. Station 36 18? Wa i m a n a l o Expt, Station
ex El Salvador 36 18
A ma r il1o Ka u a i Expt. Station 36 18
Ceiba (Cebia) II 36 18
M a m e y a 11 36 18
N ina It 36 18
Pata de Palona 11 36 18
Seda It 36 18
Puna cassava H a w a i i (Big Island) 36 187 H o n o l u l u 36 18
f. v a r i e g a t e d Ho no 1 ulu 36 18
M. g l a z i o v i i Muell-Arg. (ceara rubber) W a i m a n a l o Expt. Station 36 18
20
Table 3. M e a s u r e m e n t s of somatic c hromosomes in M. esculenta
Centro- R e l a t i v e 1 eng thC h r o m osomes M e a n length in microns mere_______F%________ percent_____
Table 4. M e a s u r e m e n t s of somatic c h r o m o s o m e s in M. gla z i o v i i
Centro- R e l a t i v eC h r o m o s o m e s M e a n length in microns mere F% 1 ength percent
Shor t Lon g arm arm Total
1,2 sat. 1.5 2.3 3.8 + 0.15 sm 39.5 100.0
3,4 0.6 2.7 3.3 + 0.13 st 18.2 86.8
5,6 1.5 1.6 3 . 1 + 0 . 1 0 m 48,4 81.6
7,8 sat, 1.3 1.8 3,1 + 0,10 sm 41.9 81,6
9,10 0.7 2.4 3.1 + 0,10 st 22.6 81.6
1 1 - 14 0.5 2.2 2 . 7 + 0 . 0 8 st 18.5 71.1
15 - 18 0.9 1.8 2.7 + 0.08 sm 33.3 71.1
19 - 22 1.2 1.3 2.5 + 0.09 m 48.0 68.4
2 3 - 2 6 0.9 1.6 2 . 5 + 0 . 0 5 sm 36.0 65.8
27 - 30 1.1 1,3 2.4 + 0.06 sm 45.8 63.2
31,32 0.6 1.6 2 . 2 + 0 . 0 4 sm 27.3 57.9
33,34 0.7 1.1 1.8 + 0.02 sm 38.9 47.4
35,36 0.6 0.8 1.4 + 0.03 sm 42.9 36.8
22
Table 5. C o m p a r i s o n of c hromosome data of M a n i h o t spp
Species 2nNu m b e r of chromosomes d i fferent centromere
withpositions
Av e r a g e length in microns
subme ta- m e t a c e n t r i c centric
s ub- t e r m i n a 1
M. esculenta 36 4 26 6 2 . 5
M . g 1 a z iov i i 36 6 22 8 2 . 6
Table 6. Fruit set, seed set and g e r m i n a t i o n of seedsfrom r e c i p r o c a l i n t e r s p e c i f i c and intras p e c i f i c crosses and selfs.
S p e c i e s / V a r i e t y
N u m b e r of flower s p o l l i n a t e d
N u m b e r ofcapsulesset
P e r c e n t a g ecapsulesset
N u m b e r of seeds set
M e a n No. of seeds per capsule
N u m b e r of seedsg ermina ted
M. g l a z i o v i i 8 55 9 16.3 21 2.3 19Itu a 42 5 11. 9 13 2.6 13Itu X M. g l a z i o v i i 29 7 24.1 15 2 . 1 15M. gl a z i o v i i x Itu 12 0 0 0 0 0Seda Q 31 9 29.3 23 2.6 20M. g l a z i o v i i x Seda 15 0 0 0 0 0Seda X M. g l a z i o v i i 21 3 14. 2 8 2 . 7 8V a r i e g a t e d 0 5 2 40.0 5 2.5 5M. g l a z i o v i i x V a r i e g a t e d 8 0 0 0 0 0V a r i e g a t e d x M. g l a z i o v i i 7 3 42. 8 7 2.3 7? ex El Salvadore 0 60 15 25.0 37 2 . 4 33Itu X Seda 11 4 36.3 10 2.5 8Seda X Itu 14 6 42. 8 15 2.5 13
N3OJ
Table 7. S t a i n a b i 1 i t y , g e r m i n a b i l i t y and size of M a n i h o t spp, pollen
S p e c i e s / V a r i e t yColor reaction/
S t a i n a b i l i t y perceht G e r m i n a b i l i t y
M e a n po l l e n size in m icrons
tetraz o l i u mchloride
b e n z i d i n e d ihydro- chlor ide
cotton blue in 1 a c to- p h e n o 1 t P= 0 . 0 5
M. g l a z i o v i i p i n k / 98 b l u e / 96 b l u e / 98 0 153 . 9
M. esculenta:0.74 2.37
Itu p i n k / 9 7 blue / 9 5 b l u e / 9 8 0 147.5 /Seda p i n k / 9 8 blue/96 blue/97 0 1 4 6 . 8 /V a r i e g a t e d p i n k / 9 7 b l u e / 98 b l u e / 96 0 1 4 6 . 4 ^
S .E . = + 10.12
to4̂-
25
Plate 1. Somatic c hromosomes of M a n i h o t species p r e t r e a t e d with 8 - o x y q u i n o 1 ine (1650X)
Figure:1. M. g l a z i o v i i (ceara rubber)
2. M. esculenta var. Itu
3. M. esculenta f. v a r i e g a t e d
4. M. e s culenta var. A m a r i l l o
5. M. esculenta var. 1 ex El Salvadore
6. M. esculenta var. Ceiba
7. M. esculenta var. Ma m e y a
8. M. esculenta var. Nina
9. M. esculenta var. Pata de Palona
0
m • •v''^.
« *
•» A
• %• •
#%•#
%
<•
7
I ; - . / - ^
5 ' - . . > , i 4 i ;
- # # •l, 'J •
€
y ♦
27
Plate 2. Somatic c hromosomes of M a n i h o t species with p r e t r e a t m e n t and w i t h o u t p r e t r e a t m e n t with 8 - ox y q u i n o l i n e and some meiotic co n f i g u r a t i o n s in m i c r o s p o r o g e n e s i s of M a n i h o t spp. (1650x)
Figure:
10. M. esculenta v a r . Seda (pretreated with 8 - oxyquinoline)
11. M. esculenta var. Puna cassava (pretreated with 8 - oxyquinoline)
12. M. esculenta var. ? (pretreated with 8 - oxyquinoline)
13. M. g l a z i o v i i (without p r e t r e a t m e n t with 8 - oxyquinoline)
14. M. e s c ulenta (without p r e t r e a t m e n t with 8 - oxyquinoline)
17. Early A n a p h a s e I in M. esculenta
18. M e t a p h a s e II (polar view) in M. esculenta (n = 18)
19. M e t a p h a s e I in M. g l a z i o v i i 18 II
20. M e t a p h a s e II in M. g l a z i o v i i (polar view) n = 18
A r r o w s indicate satellites.
10
1 * •
' . * •• * • •
A A W— m
• • •# • %
• « « •
11 1 2
\ r \ j (
• . ' . • • I ' - ■ . 5 *
19• •
u
*V_ 20..
I
; •
29
Plate 3. K a r y o t y p e s of M a n i h o t species (5500X)
Figure;
15. M. esculenta
16. M . g 1a z iov i i
16
31
Plate 4. V e g e t a t i v e m o r p h o l o g y of M a n i h o t spp
F ig ur e ;
2 1 .
2 2 .
23 .
24.
M a t u r e plant of M. g l a z i o v i i with flowers and fruits (O.lx).
Young potted plant of M. g l a z i o v i i (O.lx).
M. esc u l e n t a f. v a r i e g a t e d (O.lx).
M. e s culenta vari e t y Itu with flowers and fruits ( O . l x ) . -
D I S C U S S I O N
The number of somatic chromo s o m e s in all the var i e t i e s
and form of M a n i h o t e s culenta and M a n i h o t g l a z i o v i i d e t e r
mined is 2n = 36. The game t i c number is n = 18. These
findings agree with those of P erry (1943) and Cr a n e r (1935,
1941). H o w e v e r B o i t e a u (1938, 1941) in M a d a g a s c a r found
somatic numbers of 2n = 4, 8, 16, 24, and 32 for the three
species of M a n i h o t studied. These findings seem to be
e r roneous for if such wide v a r i a t i o n o c c u r r e d in the genus,
it should be found in the original habitat of the genus, in
this case, Brazil and Central America, rather than in
M a d a g a s c a r w here some species of the genus w ere only
intro d u c e d in c o m p a r a t i v e l y recent times.
C h r o m o s o m e s of the species of the genus d e t e r m i n e d in
this study are rather small. For the va r i e t i e s and form of
M a n i h o t e s culenta the mean chromosome length is 2.5 micr o n s
with a range of longest to shortest of 4.0 to 1.6 microns.
For M a n i h o t g l a z i o v i i the mea n length is 1.6 m i c r o n s and a
range of 3.8 to 1.4 microns. Thus the ratio of longest to
shortest c h romosome in M. esculenta is 2.5 : 1 and in
M. g l a z i o v i i it is 2.7 : 1. Except for 2 and 3 pairs o’f
ch r o m o s o m e s r e s p e c t i v e l y in each of the species w hich have
m e t a c e n t r i c centromeres, all the rest have either submeta-
centric or ac r o c e n t r i c centromeres. In M. esculenta the
Form Percent (F7») ranges from 15.3 to 50.0 wher e a s in
M. g l a z i o v i i the range is from 13.3 to 50.0. No chromosomes
with t e l o c e n t r i c centro m e r e s were observed.
T hese facts w o u l d seem to place these k a r y o t y p e s in
a s y m m e t r y class 2b a c cording to Stebbins (1958) c l a s s i f i c a t i o n
of karyotypes. In an e x tensive review of ev o l u t i o n of
k a t y o t y p e s in d i f f e r e n t taxa of the plant kingdom, Stebbins
(1971) suggests that symmetrical k a ryotypes are more p r i m i
tive than a s y m m e t r i c a l ones such that t h e e v o l u t i o n a r y trend
is from symmetry to greater asymmetry, a l t h o u g h r e versals
of this trend may occur periodically. K a r y o t y p e s of M a n i h o t
species may therefore be consi d e r e d as advanced.
P o l y p l o i d nature of M a n i h o t species
K a r y o t y p e s of M a n i h o t species studied show that each
species p o ssesses 2 pairs of satellited chromosomes. The
two pairs are different from each other in gross m orphology.
Since the dis c o v e r y of satellites by N a v a s h i n (1912) in
G a l t o n i a candicans this c hromosomal structure has been used
as one of the features to d e t ermine the level of ploidy of
plants. N u m e r o u s cases of the presence of s atellited c h r o m o
somes have been r e p orted in plants. A few cases have also
been r e p o r t e d in which no satellites have been observed.
H eitz (1931) stated that all plants must have satellites,
haplo i d s with one, diploids with two, tripolids with three,
and tetrap l o i d s with four, etc. A c c o r d i n g to N a n d i (1936)
34
the absence of satellite chromosomes as has been r e p o r t e d in
a few species is not therefore due to their real absence but
due to the d i f f i c u l t y of o b taining good m a t e r i a l s for their
detection. An d it may be added, the technique in p r eparing
the plant m a t e r i a l s for k a r y o l o g i c a l studies, as is evident
in the present study (Figs. 1 - 16). Root tips p r e t r e a t e d
w ith 8 - o x y q u i n o l i n e before fixation s h ortened the c h r o m o
somes to the extent that satellites were not visi b l e
(Figs. 1 - 12), whereas direct fixation w i t h o u t p r e t r e a t m e n t
p e r m i t t e d most m o r p h o l o g i c a l structures of the c hromosomes
to be seen at m e t a p h a s e (Figs. 13 - 14). The present study
w o u l d therefore seem to agree with the statement of Heitz
(1931) that all plants should have satellites and that their
number probably coincides with the number of genomes present
in the species.
Berger (1941) pointed out that in Spinacia oleracia root
tips of which 2n, 4n, and 8n cells occur, the satellites in
resting nuclei could be used as a method of d e t e r mining the
ploidy of the cells. W i l k i n s o n (1941) found that in Salix
alba and in S. fragilis both of which are a 11 o t e t r a p 1 o id s
w ith 2n = 76, two pairs of satellited ch r o m o s o m e s were
present. L e v a n (1931) working with various species of
A l l i u m showed that triploid nutans forms each had 3 - s a t e l l i t e d
chromosomes, in A. v a l i d u m a tetraploid, 2 - s a t e l l i t e d
ch r o m o s o m e s were present in pollen. H o w e v e r some h e x a p l o i d
35
A. nutans types had only 4 satellites w h i l e some diploid
A. narciss if lorum had no satellites..
S a tellites may be de s c r i b e d as minute p o r tions of
c h r o m o s o m e s a t t ached to the c hromosomes by a tenuous thread.
It is usually a t t a c h e d to the distal end of the chromosome.
H o w ever, satellites may sometimes appear at places other than
the distal end of the chromosome. Thus D a r l i n g t o n (1937)
showed interc a l a r y satellite (trabant) in Solanum l y c o p e r s i c o n
and in Rhoeo species.
S a tellites have been shown to play a vital role in the
o r g a n i z a t i o n of the cell. H e i t z (1931) d i s c o v e r e d that each
nuc l e o l u s arose on a s a t ellite chromosome in telophase at
the point w h e r e the satellite thread emerged. This point
was later called the n u c l e o l a r organ i z i n g body. By using a
c o m b i n a t i o n of genetic, c ytological and b i o c h e m i c a l i n v e s t i
gations, B r o w n and G u r d o n (1964) showed that in a m u t a n t of
clawed frog Xenopus laevis w h i c h lacks a n u cleolus, its
l e t h a l i t y in the h o m o z y g o u s c o n d i t i o n w a s r e l a t e d to a
defic i e n c y in the r i bosomes w hich control protein synthesis
through m e s s e n g e r R N A . R i t o s s a et al (1966) showed by
m a p p i n g techniques that b o b b e d locus in D r o s o p h i l a is at or
near the nucleolus organ i z i n g region and that bobbed flies have
less r i b osomal RNA than normal flies. These flies were also
shown to be d e ficient in DNA which codes for ribosomal RNA.
This evidence w ould seem to indicate that bo b b e d m u t a t i o n s
are defi c i e n c i e s for part of the n u c leolar o r ganizer region
36
and that the DNA of this region codes for r i b osomal R N A .
Perry (1962) showed that ribosomal RNA exists in high
c o n c e n t r a t i o n s in the n u c l e o l i and is r e l e a s e d into the
cytoplasm, from w h i c h evidence it may be c o ncluded that
n u c l e o l i are sites for synthesis of r i b o n u c l e a r proteins and
ribosomes. L a n c e (1957) d e m o n s t r a t e d that in higher plants
n u c l e i of the apical m e r i s t e m during trans i t i o n from the
v e g e t a t i v e to the r e p r o d u c t i v e state contain larger n u c leoli
at a time w h e n large amounts of prot e i n are being synthesized.
These pieces of evidence seem to support H eitz (1931)
h y p o t h e s i s that all plants, at least eucaryotes, must contain
satellites.
M e i o s i s in the species and var i e t i e s of M a n i h o t studied
was regular in so far as there were 18 b i v a l e n t s o b s erved
at m e t a p h a s e I and d i s j u n c t i o n at anaphase I. There were no
qua d r ivalents t h roughout the meio t i c process.
Perry (1943) s u ggested a basic number of 9 for the genus
Ma n iho t and basic numbers of 6, 7, 8, 10, and 11 for other
genera of the family E u p h o r b i a c e a e . C o n s i d e r i n g the 2 pairs
of d i ssimilar satel l i t e d chromosomes and regular formation
of b i v alents and disjunction, it is suggested that M a n i h o t
species under i n v e s t i g a t i o n are a 11o t e t r a p 1o i d , wit h a basic
chromosome number of x = 9. Whet h e r these a 1 1 o t e t r a p 1oids
are true or segmental (Stebbins 1950) will have to await
further investigations. Further evidence for the p o lyploid
37
nature of M a n i h o t species studied include small chromosome
size, v e g e t a t i v e vigor, and perennial growth habit, which
a c cording to Stebbins (1971) favour p o l yploid formation in
m any groups of plants.
F e r t i l i t y of selfs and crosses
F e r t i l i t y in the genus M a n i h o t is rather low. W h e n the
different v a r i e t i e s and species were selfed and/or crossed,
a g e n e r a l l y low perce n t a g e seed set was o b t ained (Table 6).
M a n i h o t esculenta crossed only u n i l a t e r a l l y as female parent
with M. g l a z i o v i i producing low p ercentage seed set. Such
unilateral c o m p a t i b i l i t y has been o b s erved in numer o u s other
species of veg e t a b l e and other crops. Ricks (1960) showed
that L y c o p e r s i c o n e s c u l e n t u m crossed u n i l a t e r a l l y as female
parent with Solanum p e n n e l l i i . The Fl hybrids crossed as
female parents to S. pen n e l l i i but only as male parents to
L. esculenturn. Both parents and the Fl hybrids each had 12
pairs of chromosomes. It was observed that failure to effect
a cross was due either to failure of pollen grains to
ger m i n a t e or very slow growth of the pollen tubes. W h e n the
m e i o t i c chromosome cytology of the two parents and the Fl
hybrids were compared (Kush and Rick, 1963) it was observed
that p a chytene c hromosomes were very similar in gross
m orphology. Several of the S . p e n nellii c hromosomes were
found to have somewhat larger chromatic regions with discrete
chromomeres, however. The chromomere patt e r n was different
38
in most cases. M e i o s i s in the hybrids was strictly regular
but only size inequalities occurred in certain bivalents. On
the basis of chromosome pairing, h y b r i d i s a t i o n compatibility,
hybrid fertility and plant m o r p h ology, they suggested a
r e v i s i o n of the taxonomy so as to place both S. p e n nellii
and L. esculenturn in the same genus. L. esculent urn crosses
u n i l a t e r a l l y whe n used as female parent with L. p e r u v i a n u m
and embryo culture is nec e s s a r y to produce Fl hybrids
(Smith, 1944). L i n d q v i s t (1960) showed that L a c t u c a sativa
and L . serriola separ a t e l y crossed with L . sa 1 igna only when
the latter species was used as female parent. He observed
that only slight differ e n c e s exist b e t w e e n L. sativa, L.
s a l i g n a , and L. serriola in chromosome m o r p h o l o g y each species
of w h i c h has a somatic number of 2n = 18. H o w e v e r L. sativa
crossed r e c i p r o c a l l y with L . s e r r i o l a . In the genus Solanum
section Tub er ar i u m , L a m m (1945) showed that Solanum r h y b i n i i
(diploid) crossed easily with related species r e c i p r o c a l l y but
S ■ aj a n h u i r 1 was o n l y s u c c e s s f u l w h e n u s e d as a f e m a l e p a r e n t .
The unilateral c o m p a t i b i l i t y b e t w e e n M. esculenta and
M. g l a z i o v i i is rather similar to the above cases siice their
c h romosome numbers and m o r p h o l o g y are e s s e n t i a l l y the same.
In a review of the p h e n o m e n o n of hybrid i nviability and w e a k
ness, Stebbins (1958) divided the causes into three categories
of: lack of hormony b e t w e e n genes or chromo s o m e s of the
parent species; c ytoplasmic or plastid di f f e r e n c e s between
39
the parent species; and d i s h a r m o n y b e t w e e n the hybrid embryo
and the surrou n d i n g tissues. The first category is recog n i z e d
by the fact that the crosses are equally u n s u ccessful in both
directions wher e a s there is a difference b e t w e e n r e ciprocal
crosses if the cause of inviability is of the kinds included
in the last two categories. The cross b e t w e e n M. esculenta
and M. g l a z i o v i i comes under the latfer categories. Causes
of the unilateral i n c o m p a t i b i l i t y are not known and will
have to await future investigations. H o w e v e r the low seed
set observed in crosses and selfs may be due to red u c t i o n in
fertility in the species as a result of r e p l a c e m e n t of seed
p r o p a g a t i o n by v e g e t a t i v e p r o p a g a t i o n and hence s e l ection
against fertility in the course of evolution. Stout and
C lark (1924) suggested this reason as a p r o b a b l e e x p l a n a t i o n
of low fertility in Solanum t u berosum and its wil d relatives.
P o l l e n p h y s i o l o g y of M a n i h o t species
Pollen grains of M a n i h o t species studied are rather
large, of the order of 147 microns for M. esculenta and 154
microns for M. g l a z i o v i i , (Table 7). There was no signif i c a n t
difference in size b e t w e e n pollen of the two species. They
are spherical and smooth. Sta i n a b i l i t y of newly sampled
pollen at the time of anthesis was high but g e r m i n a b i 1 ity
in vitro was nil. The cytology of M a n i h o t pollen has been
reported by Schnarf (1939) who showed that the pollen grains
of this genus are t r.inuc 1 e a t e . In a c o n s i d e r a t i o n of pollen
40
c ytology and v i a b i l i t y in vitro, B r e wbaker and M a j u m b e r (1961)
concluded that most pollen grains w hich are exceed i n g l y
dif f i c u l t to g e r minate in vitro are trinucleate. B r e wbaker
(1967) summarized major c h a r a c t e r i s t i c s of t rinucleate pollen
as absence of v i a b i l i t y in vitro, absence of storage longevity,
site of self i n c o m p a t i b i l i t y i nhibition the stigma, and type
of self i n c o m p a t i b i l i t y control sporophytic. He concluded
that trinucleate pollen grains demand an unusual e n v i r onment
for g e r m i n a t i o n - one w hich has not been d u p l i c a t e d e f f e c t i v e l yI
in vitro - and these more hi g h l y evolved t rinucleate plants
may have a more s p e c i alized b r e eding habit in this respect.
M a n i h o t species under inves t i g a t i o n certainly have some of the
above char a c t e r i s t i c s which may account for the failure of
their pollen to ge r m i n a t e in vitro and perhaps also partly
for the low seed set observed in c ontrolled pollinations.
It has been shown above that the c h r o m osomes of the
species studied are equal in number and similar in karyotype.
From cytological c o n s i d e rations, therefore, it seems clear
that these species are closely related and for breeding
purposes m ight be expected to cross reciprocally. H o w e v e r
results of i n t e rspecific crosses reported show that r e c i p r o
cal crossing betw e e n them is not practicable. The species
only cross u n i l a t e r a l l y whe n M. g l a z i o v i i was used as pollen
parent and M. esculenta as seed parent. A l t h o u g h no
i n v e s t i g a t i o n was ca.rried out to determine the cause of
41
failure of reciprocal crosses, pollen analysis of the species
showed no significant differences in size b e t w e e n them.
These results would seem to indicate that s i milarity of
k a r y o t y p e s b e t w e e n species cannot n e c e s s a r i l y give indication
of their reciprocal crossability.
Since the subject of i n t e rspecific h y b r i d i z a t i o n is
rather important in veg a t a b l e crops breeding, particularly,
for purposes of transferring economic characters and disease
r e s i s t a n c e from the wild to cultivated crops species, several
techniques have been devised to enhance it. These techniques
include: altering chromosome numbers of parental species,
as in Solanum tuberosum breeding (Livermore and J ohnstone,
1940; Hougas and Peloquin, 1960; in L y c o p e r s i c o n e s c u l e n t u m ,
Bohn, 1948; and Soost, 1958);
use of grafts or chimeras, (Gunther, 1964, in L y c o p e r s i c o n
esculent urn; Lamm, 1941, in Solanum tuberosum);
use of gametic diversity, (Wall and York, 1960, in Cucurbita
spp; H o n m a and H e e c k t , 1959, in P h a s e o l u s spp);
ab atement of c o m p a t i b i l i t y barriers by radiation, (Pandey,
1960 , in Solanum simp 1 i c i f o 1 ium X S. chacoense cross);
use of bridging species, (Rhodes, 1959, in Cuc u r b i t a spp;
Dionne, 1963, in Solanum tuberosum and its wild relatives);
embryo culture, (Smith, 1944, in L y c o p e r s i c o n escul e n t u m
crosses with its wild relatives; Yamane, 1953, and Wall, 1954,
in Cucurb i ta) .
42
43
Some of these techniques might be useful in i
h y b r i d i z a t i o n in M a n i h o t species where i n c o m p a t i b i l i t y is
demons t r a t e d .
SUMMARY
The chromosome numbers, k a ryotype and c r o s s a b i l i t y of
some M a n i h o t species were investigated.
Of the two species and 11 var i e t i e s studied, chromosome
number of 2n = 36 and n = 18 were recorded. Two pairs of
s atel l i t e d chromo s o m e s are reported for the first time for
each of the two species and karyo t y p e s for both species were
similar. M e i o s i s was normal in pollen mother cells showing
18 biv a l e n t s at MI and regular d i s j u n c t i o n at A I . On the
basis of these findings, it is suggested that the M a n i h o t
species under inves t i g a t i o n are a l l o p o l y p l o i d s with a basic
chrom o s o m e number of x = 9.
In spite of the simil a r i t y in k a r y o t y p e s of the two
species, they cross only unilaterally. M. g l a z i o v i i crossed
as pollen parent wit h v a r i e t i e s of M. e s culenta as seed
parent. M . esculenta var i e t i e s freely i n t e r crossed
reciprocally. The u n ilateral i n t e rspecific cros s a b i l i t y of
these species is d i scussed in relation to other such
u n ilateral c o m p a t i b i l i t y in the literature.
P ollen of neither species g e r m i n a t e d in vitro in
c a 1c i u m - s u p p 1 emented medium.
A P P E N D I X
Ca 1 c i u m - s u p p 1 emented m e d i u m for g e r m i n a t i o n of pollen in vitro
Sucrose 10 percent
B oric acid 100 ppm
C a l c i u m nitrate (Ca (NO^) 2 • 2 0) 300 ppm
M a g n e s i u m sulfate 200 ppm
Po t a s s i u m nitrate 100 ppm
D i s t i l l e d wa ter
pH commonly 5 - 7 . But some species are best at pH 8-9.
Best growth o b t ained in standing aqueous drop in petri dish
lined with m oist filter paper at room temperature.
L I T E R A T U R E CITED
A b r a h a m , A., 1957: B r e eding of Tuber Crops in India.In d i a n J o u r n a l of Genet, and Plant B r e e d i n g 17, 212-217.
Afify, A., 1933: The C y t o l o g y of the H y b r i d Betw e e nL. e s c u l e n t u m and L. r a c e m i g e n u m in R e l a t i o n to Its Parents. G e n e t i c a 15, 225-240.
Barnes, R. H . , 1954: The D i f f u s i o n of M a n i o c Plants fromSouth A m e r i c a to Africa: An Essay in E t h n o b o t a n i c a 1 CultureH istory. D i s s e r t a t i o n A b s t r a c t s 14, (10), 1496.
Berger, C. A., 1941: Some Criteria for J u d g i n g the Degreeof P o l y p l o i d y of Cells in the Rest i n g Stage. Amer. Nat.75 , 93- 96.
Bohn, G. W., 1948: S e s q u i d i p o i d Fl hybrid of L. e s c u l e n t u mand L. P e r u v i a n u m . J. Agr. Res. 77, 33-53.
B o i teau, P., 1938: C y t o l o g i e de Manioc. Chron i c a Bot. 4,386-387.
Boiteau, P., 1941: N o v e l l e s O b s e r v a t i o n s C y t o l o g i q u e s Surle M a n i o c Cultive. C h r onica Bot. 6, 388.
Bolhuis, G. G., 1953: A Survey of Some A t t e m p t s to BreedCassava Var i e t i e s with a Hig h Content of Prot e i n in the Root: E u p h y t i c a 2, 107-112.
Brandes, E. W., and Sartoris, 1936: Sugarcane: Its Originand Improvement. In U.S.D.A. Year B o o k of Agric. 1936,561-611 .
B r e w b a k e r , J. L., and S. K. Maj umb er, 1961: I n c o m p a t i b i l i t yand the Pollen Grain, pp. 1503-1508. I^ Rec e n t A d v a n c e s in Botany. Univ. T o r o n t o Press, Toronto, Canada.
B r e w b a k e r , J. L ., 1967: The D i s t r i b u t i o n and P h y l o g e n e t i cS i g n i f i c a n c e of B i n u c l e a t e and T r i n u c l e a t e Po l l e n Grains in the A n g i o s p e r m s . Amer. J. Bot. 54 (a) 1069-1083.
B rew b a k e r , J. L. and B. H. Kwack, 1964: The Calc i u m Ion andSubst a n c e s In f l u e n c i n g P o l l e n Growth. In Pollen P h y s i o l o g y and F e r t i l i z a t i o n , pp. 143-151. N or t h-lTo 11 and P u b l i s h i n g Co Ams ter d a m . 1964.
Brown, D. D., and Gurdon, J. C., 1964: A b s e n c e of Rib o s o m a lRNA Syn t h e s i s in the A n u c l e o l a t e M u t a n t of X e n o p u s laevis. Proc. Natn. Acad. Sci. U. S. A., 15, 139-14*6”!
Brown, S. W., 1949: The S t r ucture and M e i o t i c Beh a v i o u r ofthe D i f f e r e n t i a t e d C h r o m o s o m e s of Tomato. Genetics, 34, 437-461.
Burnham, C. R. and Hagbert, A., 1956: C y t o g e n e t i c N otes inC h r o m o s o m a l I n t e r c h a n g e s in Barley. B e r e d i t a s 42, 467-482.
D a r l i n g t o n , D. C. 1937: Mitosis: The C o n s t a n c y of theC hromosomes. In Re c e n t A d v a n c e s in Cytology, pp. 36-37. B l a c k i s t o n ' s Son & Co., Inc. Philadelphia.
Dionne, L. A., 1963: Studies on the Use of Sola n u m acauleas a Bridge B e t w e e n S. tu b e r o s u m and Species Tn the Series B u l b o c a s t a n a , C a r d i o p h y 11 a , and P i n n a t i s e c t a . E u p h y t i c a 12, 263- 269.
E m s w e l l e r , S. L. and H. A. Jones, 1935: A n I n t e r s p e c i f i cH y b r i d in A l l i u m Hilgardia. 9, 265-273.
E msweller, S. L. and H. A. Jones, 1935: M e i o s i s in A l l i u mfistulos urn, A. cepa and their Hybrid. H ilgardia. 9”!! 2 7 7- 2 94
Gilbert, J. C. and D. C. M c G u i r e , 1956: I n h e r i t a n c e ofR e s i s t a n c e to Severe R o o t k n o t from M e l o i d o g y n e incognita in C o m m e r c i a n Type Tomatoes. Proc. A m e r . So c . Hor t . Sci.,68, 437-442.
G o t t s c h a l k , W. 1951: U n t e r s u c h u n g e n am P a c h y t a n N o r m a l e rund R o n t g e n b e s t r a h l t e r Pollen Mu t t e r Zellen Von Solanum l y c o p e r s i c o n . Chromosama. 4, 298-341.
Graner, E. A., 1935: C o n t r i b u i c a o Para 0 Estudo C y t o l o g i c ode Mandioca. Pub. Escola Superior de A g r i c u l t u r a "Luiz de Qu e i r o z . " P i r aciaba S. Paulo, Brazil.
Graner, E. A., 1941: P o l y p l o i d cassava: Indu c e d by C o l c h i cine Treatment. J. Heredity. 32, 281-288.
Gunther, E., 1964: R e z i p r o k e B a s tarde Z w i s c h e n L y c o p e r s i c o nesculentum. Mill, und L y c o p e r s i c o n p e r u v i a n u m (l7) Mill. N a turwiss. 51, 443-444"*!
Hall, S. H. and G. J. Galletta, 1971: C o m p a r a t i v e Chro m o s o m eM o r p h o l o g y of Dipl o i d V a c c i n i u m spp. J. Amer. S o c . Hort. Sci. 96, 289-292.
47
Heitz, E., 1931: N u k l e o l e n und C h r o m o s o m e n in der GattungVicia. Planta. 15, 495-505.
Honma, S. and 0. Hee c k t 1959: I n t e r s p e c i f i c H y b r i d B e t w e e nPha s e o l u s coccineus and P. lunatus. J. Heredity. 50, 233-237.
Hougas, R. W., and S. J. Peloquin, I960: C r o s s a b i l i t y ofSolanum t u berosum H a p l o i d s with Diploid Solanum Species.E u r . Potato J . 3, 325-330.
Hu z i w a r a , Y., 1956: K a r y o t y p e Analy s i s in Some Genera ofCompositae. 1. K a r y o t y p e of J a p a n e s e Eupatorium. Cytologia.21, 114-123.
Hyde, B. B., 1953: D i f f e r e n t i a t e d C h r o m o s o m e s in Plantagoova t a . Amer. J. Bot. 40 , 809-315.
J en n i n g s , D. L., 1957: Further Studies in B r e e d i n g Cassava for Virus Resistance. E. Afri. Agric. and F o r e s t r y J.,22, 213-219.
J e n n i n g s , D. L., 1959: M. mel a n o b a s i s , a Useful Pa r e n t forC assava Breeding. EuphyticT] 8^ 157-162.
Jones, H. A. and A. E. Clarke, 1942: A N a t u r a l A m p h i d i p l o i dfrom an O nion spp. Hybrid. A. cepa x A. fistulosum. J. H e r e d 33, 25-32.
Khush, G. S. and C. M. Rick, 1963: M e i o s i s in H y b r i d s B e t w e e nL . e s c u l e n t u m and Solanum p e n n e l l i i . Genetica. 33, 167-183.
Koch, L., 1934: Cassave Selectie. V e e n m a n and Zonen,W a g e n i n g e n 1934.
Lamm, R . , 1941: Varying Cy t o l o g i c a l Beh a v i o u r in R e c i p r o c a lSolanum Cross. H ereditas, 27, 202-208.
Lamm, R., 1945: C y t o g e n e t i c Studies in Solanum SectionTuberarium. Hereditas. 31, 1-128.
Lance, A., 1957: R e c h e r c h e s C y t o l o g i q u e s Sur L ' e v o l u t i o n deQ u e l q u e s M e r i s t e m e s A p i c a u x et sur ses V a r i a t i o n s P rovoquees par des T r a i t e m e n t s P h o t o p e r i o d i q u e s . Anna l e s des Sciences N a t u r e l l e s B o t a n i q u e Ser. 11, 18, 91-422.
Lesley, M. M. and J. W. L e s l e y . , 1943: H y b r i d s of the ChileanTomato. J. Hered. 34, 199-205.
Levan, A., 1931: C y t o l o g i c a l Studies in Allium: A P r e l i m i n a r yNote. Hereditas. 15, 347-356.
48
L i m a - d e-Faria , A., 1952; C h r o m o m e r e A n a l y s i s of the Chrom o s o m eCo m p l e m e n t of Rye. C h r o m o s o m a 5, 1-68.
L i n d q u i s t , K., 1960; C y t o g e n e t i c Studies in the Serriolagroup of Lactuca. H e r e d i t a s 46, 75-151.
L i v e r m o r e , J. R. and F. E. J ohnstone, 1940; The Effect of C h r o m o s o m e D o u b l i n g on the C r o s s a b i l i t y of Sola n u m chacoense,S. J a m e s i i and S. b u l b o c a s t a n u m with S. tub ero s u m . Arne r .P otato J. 17, 170-173.
M a c Arthur, J. W. and L. P. Chaiasson, 1947: C y t o g e n e t i c sNotes on Tomato Species and Hybrids. G e n e t i c s 32, 165-177.
M c C l i n t o c k , G., 1929: C h r o m o s o m e M o r p h o l o g y in Zea maysScience, n.s. 69, 629.
Nandi, H. K., 1936: The Chrom o s o m e M o r p h o l o g y , S e c ondaryA s s o c i a t i o n and Origin of C u l t i v a t e d Rice. J. Genet. 33, 315-336.
Nav a s h i n , S., 1912: Sur le D i m o r p h i s m e N u c l e a i r e des CellulesSo matiques de G a l t o n i a Candicans. Bull. Acad. Sci. St.Pete r s h 6, 373-375.
Nichols, R. F. W., 1947: B r e e d i n g Cassava for Virus Resistance.E. Afri. J. 12, 184-194.
Pandey, K. K., 1960; Wide I n t e r s p e c i f i c H y b r i d i z a t i o n .Ann. Rept. J o h n Innes Inst. 51, 31.
Perry, A. B., 1943: C h r o m o s o m e Number and P h y l o g e n e t i cR e l a t i o n s h i p s in the E u p h o r b i a c e a e . Amer. J. Bot. 30,527-543.
Perry, R. P. , 1962: The Cellular Sites of Synthesis of R i b o s o m a l and 4s R N A . Proc. Natn. Acad. Sci. U. S. A. 48,2179-2185.
Price, S . , 1956: A L e a f Squash T e c h n i q u e for Studies ofSomatic C h r o m o s o m e s in Saccharum. I n t e r n a t i o n a l Soc. Sugar Cane Technol. Proc. 9th. Cong., India.
Purseglove, J. W., 1968: Manihot. In Tropical Crops. I.Di c o t y l e d o n s , pp. 171-180. J o h n W iley & Sons Inc. New York.
Rhodes, A. M., 1959: Species H y b r i d i z a t i o n and I n t e r s p e c i f i cGene Transfer in the Genus Cucurbita. Proc. Amer. Soc. Hort. Sci. 74, 546-551.
49
Rick, C. M., 1967: Fruit and Pedical C h a r a c t e r s Deri v e dfrom G a l a p a g o s Tomato. Econ. Bot. 21, 171-184.
Rick, C. M., 1967: E x p l o i t i n g Species H y b r i d s for V e g e t a b l eImprovement. Proc. XVII Internet. Hort. Cong. Vol. Ill,pp. 217-229.
Rick, C. M., 1960: H y b r i d i z a t i o n Between L y c o p e r s i c o ne s c u l e n t u m and Solanum pennellii. P h y l o g e n e t i c and C y t o g e n e t i c Significance. Proc. Natn. Acad. Sci. U. S. A., 46, 78-82.
R itossa, F. M., K. S. A t w o o d and S. Spiegelman, 1966: AM o l e c u l a r E x p l a n a t i o n of the Bobbed M u t a n t s of D r o s o p h i l a as Partial D e f i c i e n c i e s of R i b o s o m a l DNA. Genetic"s~! 5 4, 8T9- 834.
Rogers, D. J. 1963: Studies of M a n i h o t esculenta Crantz and R e l a t e d Species. Bull. To r r e y Bot. Club 90, 43-54.
Rogers, J. and S. G. Appan, 1970: U n t a p p e d G e n e t i c R e s o u r c e sfor Cassava Improvement. Proc. Sec. Inter. S y m p . on T r o p .Root and Tuber Crops. 1, 72-75.
Schnarf, K., 1939: V a r i a t i o n im Bau des P o l l e n k o r n e s derA n g i o s p e r m e n . Tabul. Biol. 17, 72-89.
Schwerin, K. H., 1970: A p u n t e s Sobre la Yuca Y Sus Origenes.Trop. Roots and Tuber Crops N e w s l e t t e r 3, 4-12.
Smith, P. G . , 1944: Embryo Culture of a Tomato SpeciesHybrid. Proc. Amer. Soc. Hort. Sci. 44, 413-416.
Soost, R. K., 1958: P r ogenies from Sesquid ip 1oid Fl H y b r i dof L . e s c u l e n t u m and L. p e r u v i a n u m . J. Hered. 49, 208-213.
Stebbins, G. L., 1950: V a r i a t i o n and E v o l u t i o n in P l a n t s , pp. 251-297. C o l umbia U n i v e r s i t y Press, N e w York.
, 1958: The Inviability, W e a k n e s s and S t erility
50
of I n t e r s p e c i f i c Hybrids. A d v a n c e s in G e n e t i c s 9, 147-215.
___________ , 1959: The Role of H y b r i d i z a t i o n in Evolution.Proc. Amer. Phil. Soc. 103, 231-257.
, 1958: L o n g e v i t y , H a b i t a t and R e l e a s e of Gene t i cV a r i a b i l i t y in the Hig h e r Plants. Cold Spring Harb . Symp. Quan. Biol. 23, 365-378.
______________, 1971: The M o r p h o l o g i c a l , P h y s i o l o g i c a l andC y t o g e n e t i c S i g n i f i c a n c e of Polyploidy. In C h r o m o s o m a l E v o l u t i o n in Higher Plants, pp. 124-154. A d d i s o n - W e s 1ey P u b l i s h i n g Co. Reading, M a s s a c h u s e t t s .
, 1971; The E v o l u t i o n of Karyotypes. In Chromo s o m a l
51
E v o l u t i o n in Higher Plants, pp. 85-123. A d d i s o n - W e s 1 ey P ubl i s h i n g Co., Reading, M a s s a c h u s e t t s .
Stewart, R. N., 1947; The M o r p h o l o g y of Somatic Chromo s o m e s in L i l i u m . Amer. J. Bot. 34 , 9- 26.
Stout, A. B. and C. F. Clark, 1924: Sterility of W ild andC u l t i v a t e d Potatoes with R e f e r e n c e to Breeding from Seed.Bull. U. S. Dept, of Agric. No. 1195, (1924) 1-32.
Tjio, J. H., and A. Levan, 1950: The Use of O x y q u i n o l i n e inC h r o m o s o m e Analysis. A n a l e s de la E s t a c i o n E x p e r i m e n t a l de Aula Dei. 2, (1) 21-64.
Wall, J. R., 1954: I n t e r s p e c i f i c H y b r i d s of C u c urbitaO b t a i n e d by Embryo Culture. Proc. Amer. Soc. Ho r t . ^ i .63 , 427- 430.
Wall, J. R., and T. L. York, I960: C a m e t i c D i v e r s i t y as anAid to I n t e r s p e c i f i c H y b r i d i s a t i o n in Pha s e o l u s and Cucurb i t a . Proc. Amer. Soc. Hort. Sci. 75, 419-428.
Wh i t a k e r , T. W., and C. W. Bohn, 1950: The Taxonomy,Genetics, P r o d u c t i o n and Uses of the C u l t i v a t e d Species of Cucurb i t a . Econ. Bot. 4, 52-81.
, 1930: C h r o m o s o m e Numb e r s in C u l t i v a t e d Cucurbits.Amer. J. Bot. 17, 1033-1040.
, 1931: Sex Ratio and Sex E x p r e s s i o n in theC u l t i v a t e d Cucurbits. Amer. J. Bot. 18, 359-366.
, and I. C. dagger, 1937: B r e e d i n g and Im p r o v e m e n tof Cucurbits. U. S. Dept. Agric. Year B ook 1937, 207-232.
W ilk i n s o n , J., 1941: The C y t ology of the Cricket Bat W i l l o w(Salix alba var. caerulea). Ann. Bot. 5, 149-165.
Yamane, Y., 1952; Studies in Species H y b r i d s of Cucurbita.1. C. maxima x C. m o s c h a t a . Seiken Ziho, 5, 94-99. ^
V e n k a t a s w a r a l u , J. and V. R. Reddi, 1956: M o r p h o l o g y of thePa c h y t e n e C h r o m o s o m e and M e i o s i s in Sorghum s u b g l a b r a s c e n s , a E u - S o r g h u m . J. Indian Bot. Soc., 3 5 , 3 44- 356.