164
Copyright 2010 Amrita Das

Copyright 2010 Amrita Das

ESTROGEN SIGNALING DURING DECIDUALIZATION
BY
AMRITA DAS
DISSERTATION
Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in VMS-Veterinary Biosciences
in the Graduate College of the University of Illinois at Urbana-Champaign, 2010
Urbana, Illinois
Doctoral Committee:
Professor Indrani C. Bagchi, Chair Professor Jodi A. Flaws, Co-Chair Professor Milan K. Bagchi Associate Professor Humphrey H. Yao

ii
Abstract
The process of implantation begins with the attachment of the embryo to the uterine lumen.
This event is followed by the development of the implanted blastocyst in the differentiated
endometrium. Decidualization, a signatory event during this process is characterized by the
deciduous transformation of fibroblast endometrial stromal cells to morphologically and
functionally unique decidual cells along with the formation of extensive neo-angiogenic
networks that play pivotal role in nourishing the developing embryo. Steroid hormones
estrogen (E) and progesterone (P) acting through their cognate nuclear receptors critically
modulate this maternal-fetal interaction that entails the initial success of procreation. While E
is primarily involved in cell proliferation and acquisition of epithelial receptivity, P critically
governs stromal differentiation in addition to contributing to E action in preparing the uterus
prior to the attachment reaction. Thus, a thriving implantation is the manifestation of
complementary steroid hormone action both during the proliferation and differentiation
phases. The role of E in decidualization however remained elusive. Our studies indicated that
the decidual uterus harbors steroid biosynthetic machinery driving the local production of
intra-uterine E in both mouse and human endometrium. This local steroid acting through the
estrogen receptor alpha (ER) plays a major role in sustaining decidualization and uterine
vascularization, absence of which resulted in embryo resorption. To identify the downstream
signaling pathway(s) and molecular candidate(s) that mediate the action of local E, we
performed gene expression profiling, using decidual tissues obtained from mice treated with
or without letrozole, a specific inhibitor to the key biosynthetic enzyme, cytochrome P450
aromatase. Of the number of molecules identified from the microarray analysis likely to be
involved in cell differentiation, the c-Fos family member Fos related antigen-1 (FRA-1), was
significantly regulated by local E. Our studies further revealed that FRA-1 is a direct target of
the E/ER signaling in the decidual uterus and plays a major role in sustaining the stromal
differentiation process. FRA-1, which exhibits strong nuclear expression in the uterine
decidual cells, also governed the extent of embryo invasion by modulating the stromal cell
migration and remodeling. Additionally gene profiling experiments indicated that uterine E
regulated the development of vascular network via the expression of cytokines and paracrine
factors from the differentiated stroma. To understand the direct implication of this study in
reproductive health of adult individuals, we next monitored the presence of endogenous E

iii
signaling in the human endometrium using primary stromal cells isolated from endometrial
biopsies of healthy females. In this in vitro model system, decidualization can be induced by
administering a cocktail of steroid hormones and cAMP. We were able to demonstrate
significant induction of aromatase in the differentiating cells which resulted in local
biosynthesis of E. Further, siRNA mediated gene silencing of aromatase or ER expression
led to a significant down regulation of human endometrial decidual bio-markers, indicating a
major contribution of this intra-uterine E signaling in mediating stromal cell differentiation.
Together, this study has helped us to gain novel insights of steroid hormone regulation of
endometrial stromal cell differentiation and angiogenesis in mouse and human implantation.

iv
Acknowledgments
This dissertation would have never been possible without the support of many wonderful
people I came across in the academic field. While it will be impossible to mention all, several
deserve special attention for their contribution.
I am thankful to Dr. Indrani Bagchi for her enormous support, encouragement and for
helping me structure myself through out graduate school. She had a big contribution in my
scientific and personal development. Looking back, it seemed she always knew when I
needed the advice and when I was better off figuring things out for myself. I am equally
grateful to Dr. Milan Bagchi for mentoring me and sharing critical insights that have
significantly helped me develop this project. I am also thankful to the members of my PhD.
committee- Drs. Jodi Flaws and Humphrey Yao for their valuable suggestions and for helping
me prepare the postdoctoral interviews. I would also like to thank Dr. Aslam Hassan for
being the Director of Graduate Studies Program in the department and for his critical
suggestions.
I am extremely fortunate to have Dr. Quanxi Li as my colleague and friend. In addition to
his constructive criticism that has always encouraged me to do unbiased science, he has
guided me enormously in developing my scientific writing. I would have never made it into
science without the encouragement and confidence of Dr. Alok Ghosh Chaudhuri and Prof.
Chanchal K. Dasgupta from University of Calcutta, India. I am extremely thankful for their
guidance in my formative years as a student in science. While the list grows, I would like to
mention my friends and colleagues including Dr. Athi Kannan, Mary Laws, Santhi Sridharan,
Chia Feng Liu and Rituparna Moitra for their technical and personal support through out my
graduate studies. Further, I am also thankful to Billie Field Memorial Foundation for
providing me financial support.
This acknowledgment remains incomplete without mentioning my family. My father Arun
Kumar Das and my mother Jayasree Das always had their strongest belief and confidence in
me. I cannot thank them enough for whatever I have achieved today. I am also extremely
thankful to my sister Arpita, for always being my trusted friend and Soham, for being the
most adorable nephew.
Finally, my long term friend and now husband Akash has been the source of my strength
through all the years since college. Without him I cannot imagine myself completing graduate
school. I am indebted to his support and for always standing by me.

v
To Baba and Maa

vi
Table of Contents
Introduction ...................................................................................................................................1
CHAPTER 1: Literature Review: Molecular regulators of stromal differentiation and angiogenesis ....................................................................................................................................4 1.1 Abstract ......................................................................................................................................5 1.2 Extensive uterine remodeling characterizes implantation .........................................................6 1.3 Steroid hormone action regulate implantation...........................................................................7 1.3.1 P signaling during implantation ........................................................................................7
1.3.1.a Regulation of PR expression..................................................................................7 1.3.1.b Downstream mediators of P action ........................................................................8
1.3.2 E signaling during implantation......................................................................................11 1.3.2.a Intrauterine E is critical during decidualization-emerging concepts....................13 1.3.2.b Intrauterine E regulates angiogenesis during decidualization .............................13 1.3.2.c Downstream mediators of E action ......................................................................14
1.4 Additional regulators critical for stromal cell differentiation ..................................................16 1.5 Additional regulators critical for angiogenesis ........................................................................17 1.6 Decidualization in human endometrial stromal cells...............................................................17 1.6.1 Biomarkers of human stromal cell decidualization ........................................................18 1.6.2 Hormonal regulation of the human menstrual cycle.......................................................19 1.6.3 Molecules governing human stromal cell decidualization..............................................20 1.6.4 Angiogenesis and development of spiral arteries during human menstrual cycle..........23 1.7 Figures......................................................................................................................................26
CHAPTER 2: De Novo synthesis of estrogen in pregnant uterus is critical for stromal cell decidualization.......................................................................................................................32 2.1 Abstract ....................................................................................................................................33 2.2 Introduction..............................................................................................................................34 2.3 Experimental Procedures .........................................................................................................35 2.4 Results......................................................................................................................................37 2.4.1Ovarian E is not essential for decidualization..................................................................37 2.4.2 Evidence for biosynthesis of E in the decidual uterus ....................................................38 2.4.3 Uterine aromatase activity is critical for successful implantation ..................................40 2.4.4 Inhibition of uterine aromatase activity blocks stromal differentiation..........................40 2.5 Discussion ................................................................................................................................41 2.6 Figures......................................................................................................................................45
CHAPTER 3: Fos-related antigen 1 (FRA-1) is an estrogen regulated transcription factor playing a critical role in mediating uterine decidualization .........................................53 3.1 Abstract ....................................................................................................................................54 3.2 Introduction..............................................................................................................................55 3.3 Experimental Procedures .........................................................................................................56 3.4 Results......................................................................................................................................60

vii
3.4.1 Intrauterine E regulates FRA-1 expression during decidualization................................60 3.4.2 Induction of FRA-1 during decidual phase of pregnancy...............................................61 3.4.3 E/ER signaling regulates FRA-1 expression in differentiating stromal cells...............62 3.4.4 ERK-MAPK pathway dependent activation of FRA-1 in differentiating stromal cells .63 3.4.5 FRA-1 is a critical mediator of stromal cell decidualization ..........................................64 3.4.6 FRA-1 regulates stromal cell remodeling through extracellular matrix proteins ...........65 3.5 Discussion ................................................................................................................................66 3.6 Figures......................................................................................................................................70
CHAPTER 4: Intra-uterine estrogen signaling critically regulates decidual angiogenesis ..85 4.1 Abstract ....................................................................................................................................86 4.2 Introduction..............................................................................................................................87 4.3 Experimental Procedures .........................................................................................................88 4.4 Results......................................................................................................................................91 4.4.1 Endothelial cell population in decidual uterus................................................................91 4.4.2 Isolation of endothelial cells from decidual uterus .........................................................91 4.4.3 Intrauterine E biosynthesis plays a critical role in decidual vascular growth.................92 4.4.4 Plausible mechanisms of E action in modulating uterine vascular development ...........93 4.4.5 Expression of angiogenic markers is concentrated to the mesometrium........................94 4.5 Discussion ................................................................................................................................94 4.6 Figures......................................................................................................................................98
CHAPTER 5: Estrogen signaling plays a critical role in mediating human endometrial stromal cell decidualization.......................................................................................................109 5.1 Abstract ..................................................................................................................................110 5.2 Introduction............................................................................................................................111 5.3 Experimental Procedures .......................................................................................................112 5.4 Results....................................................................................................................................115 5.4.1 Cytochrome P450 aromatase is induced in differentiating endometrial stromal cells.115 5.4.2 Functional E biosynthetic machinery in differentiating endometrial stromal cells ......116 5.4.3 Endogenous E plays a major role in mediating stromal differentiation........................116 5.4.4 Optimal concentration of local E critically regulates stromal differentiation...............117 5.4.5 ER plays a major role in stromal cell differentiation .................................................118 5.5 Discussion ..............................................................................................................................119 5.6 Figures....................................................................................................................................123
Summary ....................................................................................................................................131
Bibliography ...............................................................................................................................134
Appendix.....................................................................................................................................145

viii
List of figures Figure 1.1: Morphological structure of uterine cross-sections during implantation......................26 Figure 1.2: Major hallmarks of implantation.................................................................................27 Figure 1.3: Mouse in vitro stromal cell culture..............................................................................28
Figure 1.3A: Morphological change ......................................................................................28 Figure 1.3B: Expression of biochemical markers..................................................................28
Figure 1.4: Steroid hormone nuclear receptor expression during implantation.............................29 Figure 1.4A: Expression of ER ...........................................................................................29 Figure 1.4B: Expression of PR ..............................................................................................30
Figure 1.5: Steroid hormone profile during implantation..............................................................31 Figure 2.1: Administration of exogenous P sustains decidualization in ovariectomized pregnant mice.................................................................................................................................45
Figure 2.1A: Experimental scheme........................................................................................45 Figure 2.1B: Gross uterine morphology ................................................................................45 Figure 2.1C: Expression of decidual bio-markers .................................................................45
Figure 2.2: Evidence for local E biosynthesis in decidual uterus ..................................................46 Figure 2.2A: Schematic representation of E biosynthetic machinery....................................46 Figure 2.2B: Steroid biosynthetic enzyme expression...........................................................46 Figure 2.2C: Aromatase expression by Northern Blot...........................................................46 Figure 2.2D: Laser Capture Micro-dissection RT PCR.........................................................47 Figure 2.2E: Expression of P450 c17 in decidual uterus .......................................................48 Figure 2.2F: Expression of P450 aromatase in decidual uterus, stromal cells ......................48
Figure 2.3: Blockade of aromatase function leads to loss of pregnancy .......................................49 Figure 2.4: Inhibition of aromatase activity impairs uterine decidualization ................................50
Figure 2.4A: Experimental design .........................................................................................50 Figure 2.4B: Decidual response in stimulated horn...............................................................50 Figure 2.4C: Quantitative analysis of uterine weight gain ....................................................50 Figure 2.4D: Expression of decidual bio-markers .................................................................51 Figure 2.4E: Immunolocalization of Cx-43 in control and inhibitor treated uteri.................51
Figure 3.1: Intrauterine E regulates FRA-1 expression during decidualization ............................70 Figure 3.2: FRA-1 expression is induced in the decidual uterus ...................................................72
Figure 3.2A: During pregnancy .............................................................................................72 Figure 3.2B: During artificially induced decidualization ......................................................72
Figure 3.3: FRA-1 is down stream of E signaling in mouse uterine stromal cells ........................73 Figure 3.4: FRA-1 is directly regulated by E/ER signaling during decidualization ...................74
Figure 3.4A: FRA-1 expression is induced in mouse stromal cells as decidualization progresses in vitro ..................................................................................................................74 Figure 3.4B: ER expression is induced in mouse stromal cells as decidualization progresses in vitro ..................................................................................................................75 Figure 3.4C: FRA1 is regulated by ER in differentiating mouse uterine stromal cells ......76
Figure 3.5: Recruitment of ER to Fra-1 promoter.......................................................................77 Figure 3.6: ERK-MAPK pathway dependent regulation of FRA-1 activation in differentiating stromal cells ...................................................................................................................................78
Figure 3.6A: Expression profile of FRA-1 and ERK1/2 .......................................................78 Figure 3.6B: Gel shit assay, indicating DNA binding activity of FRA-1..............................79
Figure 3.7: Fra-1 is critical for mouse uterine stromal cell differentiation....................................80

ix
Figure 3.8: FRA-1 modulates uterine stromal cell mobility through extracellular matrix proteins ...............................................................................................................................82
Figure 3.8A: Wound healing assay after Fra-1 silencing ......................................................82 Figure 3.8B: Quantitative representation of wound healing assay ........................................82 Figure 3.8C: Ki-67 staining after wound healing assay.........................................................83 Figure 3.8D: Expression of extracellular modulators after Fra-1 silencing...........................83
Figure 3.9: Expression of FRA-1 in the mesometrium of Day 8 uterus ........................................84 Figure 4.1: Extensive endothelial cell population in decidual uterus ............................................98
Figure 4.1A: Day 8 whole mount deciduoma........................................................................98 Figure 4.1B: Co-culture of stromal-endothelial cells in vitro................................................98
Figure 4.2: Isolation of endothelial cells from decidual uterus .....................................................99 Figure 4.3: Classification of the angiogenic molecules downregulated after AI administration.101 Figure 4.4: Stromal cell differentiation induces the release of angiogenic modulators, regulated by endogenous E action ...............................................................................................104 Figure 4.5: Stromal cell derived factors are modulated by E action............................................105 Figure 4.6: Intrauterine E regulates the expression of endothelial cell specific angiogenic factors...........................................................................................................................................106 Figure 4.7: Expression of angiogenic markers in specific uterine zones in the presence and absence of aromatase inhibitor.....................................................................................................107 Figure 5.1: Cytochrome P450 aromatase is induced in differentiating human endometrial stromal cells .................................................................................................................................123
Figure 5.1A: Human endometrial stromal cells differentiates in vitro in the presence of steroids and cAMP...............................................................................................................123 Figure 5.1B: Induction of the human stromal cell differentiation markers .........................123 Figure 5.1C: Cytochrome P450 aromatase is induced during in vitro differentiation.........123 Figure 5.1D: Immunolocalization of P450 aromatase in differentiating stromal cells........124
Figure 5.1E: Expression of P450 aromatase in the human endometrium............................124 Figure 5.2: Presence of a functional steroid biosynthetic machinery in the human endometrial stromal cells .................................................................................................................................125 Figure 5.3: Endogenous E plays a major role in mediating stromal differentiation ....................126 Figure 5.4: Optimal concentrations of E sustain stromal differentiation.....................................127 Figure 5.5: Expression of ER during stromal cell differentiation .............................................128 Figure 5.6: ER regulates human endometrial stromal cell differentiation ................................129
Figure 5.6A: siRNA mediated silencing of ER in stromal cells .......................................129 Figure 5.6B: Morphological change after ER silencing....................................................129 Figure 5.6C: Expression of decidualization markers after ER silencing ..........................129 Figure 5.6D: Over-expression of ER induces differentiation............................................129
Figure 6: Summary: Estrogen mediated transcriptional network in the decidual endometrium .133 Figure A-1: Silencing ER expression in mouse stromal cells ...................................................146 Figure A-2: Cellular architecture of cells treated with ER siRNA............................................147 Figure A-3: ER critically modulates mouse stromal cell differentiation ..................................147 Figure A-4: C/EBP regulates aromatase expression in decidual uterus ....................................148 Figure A-5: Map of mouse ERG-1 clone.....................................................................................150

x
List of Tables Table 2.1: Measurement of aromatase activity in uterus during early pregnancy .........................52 Table 4.1: Percentage of endothelial cells in decidual uterus......................................................100 Table 4.2: Angiogenic factors regulated by intrauterine E ..........................................................108 Table A.1: Mouse primer sequences............................................................................................151 Table A.2: Human primer sequences...........................................................................................153

1
Introduction
Infertility and altered reproductive health in adult women are some of the leading concerns of
today’s society. Although modern medicine led to tremendous technological advancements in
developing assisted reproductive techniques, the success rates amongst infertile individuals
remain fairly low. One of the major causes of the failed conception is impaired endometrial
modification to support the development of a healthy and competent blastocyst. The study of
implantation focuses on understanding the maternal physiology that coordinates embryo
sustenance before placentation. Decidualization, the major event during this process is
characterized by differentiation of stromal cells to the decidual cells accompanied by
extensive development of vascular networks. Identification of the signaling pathway(s) and
molecular candidate(s) regulating the uterine preparation for pregnancy would thus
effectively contribute to the development of therapeutics for treating implantation failure and
pathological situations like endometriosis.
Steroid hormones estrogen (E) and progesterone (P) orchestrate the implantation process
by regulating the terminal differentiation of uterine fibroblast stromal cells to epitheloid
decidual cells. While the pivotal role of P in inducing decidualization is widely established,
the role of E is primarily focused on acquisition of uterine epithelial receptivity and embryo
attachment. The contribution of the later in stromal differentiation process however, remains
unclear. Recent studies in our laboratory have revealed that cytochrome P450 aromatase, the
cardinal enzyme regulating E biosynthesis, is expressed and results in synthesis of E in the
uterus during decidual phase of pregnancy. To analyze the functional significance of this
local steroid action, the goal of this overall dissertation project is centered on the question:
“Does E signaling play a role in decidualization?”
A detailed overview of the established molecular mechanisms during implantation,
primarily in the decidualization phase has been reviewed in Chapter 1 of the dissertation
while Chapters 2 to 5 address the role of E signaling in the regulation of decidual program in
mice and human.
Letrozole, a specific enzyme inhibitor to aromatase action, disrupts E biosynthetic
machinery. By implementing this phenomenon we first addressed the role of decidual E

2
during early pregnancy in mice. We observed that letrozole treatment had a severe
consequence in sustaining stromal differentiation and blastocyst development. To identify the
possible downstream targets involved in mediating this effect, we subjected uterine RNA
treated with or without the inhibitor drug to microarray-based gene expression profiling. In
Chapter-2 we detail the signaling mechanisms that mediate endogenous E action during
stromal cell differentiation. The primary objective was to identify critical down stream targets
of uterine E that mediate stromal differentiation process.
Gene profiling experiments identified an early inducible gene that belongs to the c-FOS
family of transcription factors, Fra-1, as a potential downstream target of aromatase-derived
E. FRA-1 signaling is critical for cell differentiation in several systems including osteoblast
differentiation. Our studies showed prominent expression of this molecule in the
decidualizing stromal bed during early pregnancy. Employing an in-vitro decidualization
approach and siRNA mediated blockade of gene expression, we evaluated the functional
contribution of FRA-1 in the endometrium. In Chapter 3 we have discussed our results
showing the critical role of this transcription factor in governing stromal cell decidualization.
Angiogenesis, the development of new vascular network from pre-existing structures is a
critical physiological process characterizing decidualization. Additionally, the decidua-
derived paracrine factors play a major role in regulating this neo-vascularization process.
Inhibition of aromatase expression in the uterus resulted in a severe defect in the development
of endometrial vasculature during early pregnancy. Letrozole treatment downregulated the
expression of several angiogenic factors in the decidual uteri. In Chapter 4, the candidate
molecules that mediate local E action in controlling uterine vascular growth is addressed.
Further, to identify the endothelial cell specific E target molecules, which contribute to
decidual vascularization, differential expression of these regulatory molecules in blood vessel
rich mesometrial region of the uterus in control and aromatase inhibitor treated animals was
analyzed.
Although insights from the murine research-models are important in understanding of
human physiological processes, direct analysis of human tissues undoubtedly gives us a
better understanding of the actual scenario. The final part of this dissertation thus focused on
identifying the contribution of E signaling during human endometrial stromal cell
decidualization. Stromal cells isolated from endometrial biopsies of healthy individuals

3
undergo in-vitro decidualization in response to steroid hormone P and cAMP. The cellular
differentiation is demarcated by transition of the fibroblast morphology to an epitheloid
shape. Interestingly, we observed a significant induction of P450 aromatase expression in
these differentiating stromal cells, indicating that similar to the rodent model, local E
signaling might contribute to human decidualization. In Chapter 5, we focused on the
importance of local E signaling via its predominant receptor ER in mediating human
endometrial stromal cell differentiation.
In summary, this dissertation study has focused on the critical role of stertoid signaling in
the regulation of uterine function during implantation. The experiments performed have
significantly contributed to this goal by testing the hypothesis that intrauterine E signaling
and its downstream target molecules critically regulate endometrial stromal differentiation
and uterine neovascularization during implantation in mouse and humans.

4
Chapter-1
Literature Review- Molecular regulators of stromal differentiation and angiogenesis

5
1.1. ABSTRACT
The close association of the fertilized embryo and the receptive maternal uterus during the
“window of implantation” governs the initial success of procreation. Although, the extent of
trophoblast invasion differs between species, yet this event universally regulates a thriving
conception. The overall implantation process is defined by a series of physiological events
initiated with the attachment of the embryo to the uterus followed by deciduous
transformation of a non-pregnant uterine tissue to a specialized chamber supporting embryo
growth. As stromal cell proliferation and differentiation proceed modifying the uterine
cellular mass, new blood vessels originate to support the growth of the developing embryo.
Unlike rodents, in humans, differentiation of stromal cells is initiated in regular menstrual
cycles through a process known as pre-decidualization. Steroid hormone orchestration
critically regulates the implantation process. In addition to the ovarian source of estrogen (E)
and progesterone (P) that prepares the uterine cells for embryo receptivity, recent studies
have identified an intrauterine origin of E significantly contributing to the decidualization
process. This hormone action regulates uterine cellular differentiation, vascular permeability
and angiogenesis to sustain embryo growth and thereby the successful initiation of
pregnancy. In this chapter we will focus on the major findings in the field of implantation
biology that pictures the interplay of steroid hormone signaling paradigms during early
pregnancy.

6
1.2. Extensive uterine remodeling characterizes implantation
Implantation is initiated in mouse when the blastocyst enters the uterine cavity 4 days after
fertilization. Embryo attachment to the uterine wall initiates apoptosis of the adjacent luminal
epithelial cells and proliferation of the surrounding stromal population. With the progression
of pregnancy, a subset of these proliferating cells enters the differentiation route to form
polyploid and secretory decidual cells (1, 2). Subsequently, the wave of differentiation
progress to the deeper layers of the implantation chamber forming the secondary decidual
zone (Fig. 1.1). Stromal cell decidualization has been assessed by monitoring the enzymatic
activity of alkaline phosphatase and the expression of prolactin related protein, two most well
characterized markers of decidualization (Fig. 1.2). By day 10 of pregnancy, the terminally
differentiated stromal cells undergo apoptosis to accommodate the growing embryo and
placentation ensues to maintain the conceptus till term. The morphological transition of
isolated stromal cells during differentiation can be well appreciated in vitro primary mouse
stromal cell cultures. Stromal cells enzymatically isolated from day 4 pre-implantation mouse
uterus differentiate when cultured in the presence of steroid hormones (Fig 1.3). As the cell
decidualizes, they form multinucleated giant cells that exit from the cell cycle and enter
endoreduplication resulting in the polyploid nucleus with a DNA content as much as n=64
(3).
In the human endometrium, decidualization is however initiated independent of a
blastocyst in the superficial endometrial stromal layer surrounding the spiral arteries during
the mid-secretory phase of every menstrual cycle. In the event a blastocyst invades through
the uterine epithelium, steroid hormone regulated stromal differentiation involves the entire
endometrium accompanied by growth of uterine spiral arteries and infiltration of the uterine
stroma by macrophages, lymphocytes and natural killer cells (4). In the absence of a
blastocyst, the hormone level declines, the endometrium is sloughed off and the menstrual
cycle repeats.
Formation of an extensive angiogenic network during the decidual transformation of
stromal cells is a key physiological consequence that supports early pregnancy and the initial
embryo development. At this time a large population of cytokines, growth factors and other
angiogenic cues are locally released in the uterus that drives proliferation and maturation of
nascent vascular structures to mature blood vessels. In humans, the cyclical change of the

7
endometrium is accompanied by the proliferation, growth and coiling of the spiral arteries to
develop the angiogenic network as the stromal cells undergo pre-decidualization in the mid-
secretory phase.
1.3. Steroid hormone action regulate implantation
Steroid hormones estrogen (E), progesterone (P) and their downstream signaling players are
pivotal in coordinating the physiological events during implantation. The schematic
representation of the hormonal profiles during implantation are represented in Fig 1.5A
(mouse) and B (human). The biological effects of these steroids are classically known to be
mediated by the respective nuclear receptors ER and PR, which act like ligand inducible
transcription factors (5). E mediates its action through two different receptors, ER alpha
(ER) and ER beta (ER). While it has been widely documented that the ER is the critical
receptor governing uterine proliferation and embryo attachment, studies have addressed the
important role of ER in the growth and differentiation of uterine epithelium (6). Ligand
bound ER undergoes a conformational change and binds to the E response elements of target
gene as a receptor dimer. This mechanism of action is referred to as the classical pathway.
Alternatively in some cases ER can also regulate gene expression through the non-canonical
pathway by interacting with AP-1 or Sp1 protein complexes (7). P mediates its effect
through the two isoforms PRA and PRB; PRA is the major isoform involved in the regulation
of uterine physiology. The mechanism of PR action also involves the classical response
element mediated signaling, where in the presence of ligand, PR dissociates from the heat
shock chaperone proteins, dimerizes and binds to P response elements, recruit co-regulators
and govern the transcriptional fate of target genes. PR can also act through the non-classical
pathway, when it undergoes ligand independent activation and translocates to the nucleus to
regulate gene expression (8). Gene silencing studies have been instrumental in understanding
the importance of these proteins during implantation, as will be discussed shortly. Fig 1.4
depicts the expression profile of the steroid receptors ER (A) and PR (B) during early
pregnancy.
1.3.1. P signaling during implantation

8
P signaling through PR plays an authoritative role throughout implantation governing the
transformation of the stromal cells to the decidual cell mass. The role of PR in regulating
decidualization has been widely addressed by several laboratories including ours. Absence of
PR obliterates the implantation process with severe impairment in uterine receptivity, embryo
attachment and decidualization (9). Administration of the well known PR antagonist RU486
also exhibited a severe defect in implantation, similar to the knock out model. Gene
expression analysis of mouse uterus treated with this inhibitor drug revealed a large repertoire
of genes with pivotal role in maintenance of the implantation process (10). Some of these
molecules have been widely explored and will be discussed briefly in this chapter.
1.3.1.a. Regulation of PR expression
PR remains protected by a cohort of chaperone proteins in the absence of endogenous ligand.
These constitute a 90kd heat shock protein (Hsp 90), a co-chaperone and an immunophilin
with tetratricopeptide repeat (TPR) domain, FKBP52 or FKBP51 also known as FKBP4 and
FKBP5, respectively. The uterine localization of FKBP52 exhibits maximal similarity to the
pattern of PR expression during decidualization and is considered as the major chaperone
protein involved in maintaining the functional form of the nuclear receptor competent for
hormone binding. Knockout studies have indicated that while FKBP51 null animals are
fertile, the FKBP52 null mice develop normally to the adult stage but exhibit a severe defect
in implantation. Further, these null mice are also unable to respond to experimentally induced
decidualization (11). PR activity in these mice is severely impaired along with the inhibition
of target genes amphiregulin, IHH and HOXA10. Further, lack of PR activity also reversed P
dependent antagonism of E actions resulting in enhanced E mediated proliferation of uterine
epithelial cells in the knockout animal along with the increased expression of E responsive
epithelial marker lactotransferrin. Together the study indicated that FKBP52 plays a major
role in regulating PR function in the decidual uterus and thereby significantly contributes to
the maintenance of implantation.
1.3.1.b. Downstream mediators of P action
Indian hedgehog (IHH)
A member of the hedgehog morphogens, IHH is a P regulated gene localized in the uterine
luminal and glandular epithelium during the implantation window (12). Conditional ablation
of Ihh in the PR responsive cells of the uterus resulted in uterine infertility (13). The
blastocyst is unable to attach to the uterine wall in the absence of IHH due to an increased

9
expression of MUC-1. Additionally administration of an artificial decidual stimulus failed to
elicit a response, indicating complete failure of the implantation process. The receptor,
Patched homolog 1 (Ptch1), a known target of hedgehog action is expressed in the uterine
stromal cell compartment along with other downstream targets of IHH signaling like the
Hedgehog interacting protein (Hip) and Chicken Ovalbumin Upstream Promoter-
Transcription Factor II (COUP-TF II). Ablation of IHH expression in the uterus impaired the
expression of Ptch1 and COUP-TF II in the stromal compartment, indicating IHH signaling is
an effective mediator of epithelial-stromal communication. The hedgehog pathway in the
uterus also regulates uterine cell proliferation by governing the EGFR signaling (13). In
addition to a defect in cell proliferation and differentiation, ablation of IHH in the uterus
impaired development of the angiogenic network due to the down regulation of Tie2, a well
characterized receptor mediating angiopoeitin action.
In order to understand the mechanism through which P regulates IHH expression in the
uterus, a recent study addressed the involvement of PR in bringing about this effect (14). PR
is expressed both in the epithelial and stromal compartment in the uterus. Since IHH was
confined to the epithelial cell type, a direct PR regulation from the same compartment was
obvious. The regulation of IHH expression by the specific PR population was addressed by
tissue recombination experiments. Interestingly, reciprocal recombination of the wild type
epithelium and PR knock out (PRKO) stroma indicated that PR expressed in the stromal
compartment alone regulates IHH expression in the epithelial layer. Further, this epithelial
morphogen acts through its stromal receptor Ptch1 to regulate the expression of target
molecules like COUP-TF II in the stroma. Identification of the PR-IHH-COUP-TF II
signaling during implantation unveiled a unique stromal-epithelial inter compartmental
crosstalk and its pivotal contribution to stromal cell differentiation.
Chicken ovalbumin upstream transcription factor –II (COUP- TF II)
COUP-TF II is an orphan nuclear receptor with known function in regulating epithelial
mesenchymal cross talk in many tissue systems. Global knockout models of COUP-TF II are
embryonic lethal due to severe defects in cardiovascular and angiogenic development. The
importance of this molecule in adult uterus has been addressed by conditional knock out
mouse models. COUP-TF II is expressed in the uterine stromal cell compartment during
implantation. Uterine stromal cell specific silencing of COUP-TF II impaired embryo
attachment and decidualization (15). At the molecular level, this defect in implantation has

10
been accounted to decreased PR expression in the stroma. During early pregnancy, stromal
PR attenuates the ER activation in the epithelium, antagonizes E regulated epithelial
proliferation and further governs down regulation of ER regulated glycoproteins like
MUC1, a prerequisite for embryo attachment. In the absence of this orphan nuclear receptor,
low levels of PR releases the epithelial inhibition resulting in high ER expression and
increased ER activity in the epithelium leading to a complete implantation failure. Exogenous
administration of BMP2 can ameliorate this defect, indicating the critical role of the TGF
family morphogen in mediating the effect of COUP-TF II action in decidualizing uterus.
Bone morphogenetic protein 2 (BMP2)
Bone morphogenetic proteins (BMP) are well characterized mediators of differentiation in
the development and organization of various tissue systems. BMP2 and its receptor BMPRII,
belonging to the TGF family, are two well characterized targets of P signaling in mouse
uterus, expressed particularly in the stromal cell compartment during the early phase of
decidualization. With the advancement of cell differentiation BMP2 expression sequentially
progresses to the neighboring secondary decidual zone, indicating a pivotal role of this
morphogen in decidualization. The importance of BMP2 during decidualization has been well
characterized by in vitro mouse primary stromal cell cultures and by silencing BMP2
expression in the uterine stromal cells through the PR driven cre recombinase in vivo.
Embryo attachment remains unaffected after the conditional deletion of BMP2 however the
differentiation of stromal cells to the decidual phenotype is greatly impacted by the
attenuation of this molecule (16). Stromal cells isolated from day 4 morning pre-implantation
mouse uterus, in the presence of exogenous steroids undergo in vitro differentiation. In
addition to the morphological change of the stromal cells, the expression of decidual markers
like alkaline phosphatase (Alkp) and prolactin related protein (Prp) are significantly induced
over time. Studies employing this in vitro primary cell culture model indicated that
administration of recombinant BMP2 accelerated the differentiation of stromal cells with a
significantly enhanced the expression of the bio-markers of decidualization (17).
BMP2 action in cell differentiation has been shown to be coordinated by a member of the
Wingless (Wnt) family of glycoprotein, Wnt4. Wnt proteins are highly conserved secreted
molecules that regulate cell-cell interactions during organ development. They bind to the Fzd
receptors on cell membrane and act either through the canonical beta catenin or the non-

11
canonical signaling through the calcium pathway. Over-expression of BMP2 significantly
enhanced Wnt4 expression in primary stromal cell cultures, while silencing the morphogen
suppressed it. Attenuation of Wnt4 expression by siRNA mediated gene silencing exhibited a
marked impairment of stromal cell differentiation in vitro indicating that it might be the key
regulator of BMP2 mediated decidualization (17).
Homeobox gene: HOXA10
The AbdominalB-like homeobox gene, HOXA10 is expressed in the uterine stromal cells
during decidualization and is a target of P signaling mediated through PR. HOXA10 is
involved in the development of genitourinary and adult female reproductive tracts. Null
mutation of Hoxa10 results in a severe defect in decidualization (18). Mechanistic studies
have revealed that this defect in implantation is caused by the altered cell proliferation in
response to exogenous steroids. Although the expression of certain differentiation related
molecules are unchanged, the expression of the PGE2 receptor subtypes EP3 and EP4 are
aberrantly expressed in the null uterus. It is concluded that HOXA10 signaling acts to
mediate P response and thus the activation of the steroid targets EP3 and EP4. The expression
of COX2 and the co-chaperone FKBP52 is altered in the HOXA10 deficient mouse uterus
which further contributes to the implantation defect (18) (19).
CCAAT/ enhancer-binding protein beta (C/EBP)
The transcription factor C/EBP is a critical mediator of steroid hormone action in the
decidual uterus. It belongs to the family of basic leucine zipper domain proteins that is
important in regulating cellular processes like proliferation, differentiation, inflammation and
apoptosis. The expression of C/EBP is induced in the mouse uterus at the time of
implantation and is under the dual regulation of both E and P, thereby contributing to the
differential regulation of cellular functions in the two uterine cell types. Studies in our
laboratory involving the global knock out of C/EBP have identified that it plays a critical
role in uterine epithelial and stromal cell proliferation and in the differentiation of stromal
cells, absence of which leads to a complete failure of the implantation process and a steroid
unresponsive uterus (20).
1.3.2. E signaling during implantation

12
Epithelial cell proliferation is an important event coordinating uterine preparation for
implantation. E plays the predominant role in this process by controlling the expression of
cyclin and cyclin dependent kinases (CDK) (21). Epithelial cell proliferation in response to E
and its stromal receptor ER is mediated by the nuclear localization of cyclin D and
activation of the corresponding CDK4 and CDK6. In a recent study, Zhu and Pollard
identified the signaling pathway that regulates this inter-compartmental crosstalk in
governing cell proliferation. E/ER mediated activation of insulin like growth factor 1 (IGF-
1) pathway activates epithelial PI3-kinase/AKT signaling (22). In the scheme of events,
activated PI3K regulates phosphorylation of the inhibitory domain of GSK3 rendering it
unstable, which further governs the nuclear expression of cyclin D1 and thus progression of
cell cycle. P antagonizes this pathway by inhibiting the phosphorylation of PI3kinase
resulting in stabilization of GSK3 and thus the subsequent degradation of cyclin D and lack
of cell proliferation (23). Thus, a balance in these two steroid dependent regulatory
mechanisms is critical in preparing the epithelial receptivity prior to blastocyst attachment.
Classical studies using the delayed model of implantation have established that E is also
critical in mediating the attachment of the dormant blastocyst in the P primed ovariectomized
pregnant mouse uterus (24, 25). In addition, studies by Lubahn et. al. identified that absence
of ER in mouse results in a hypoplastic uterus with severe impairment to exogenous
hormone treatment and a uterus refractory to embryo attachment (26).
Following blastocyst attachment, decidualization is the most essential prerequisite
governing the implantation success. It requires a controlled orchestration of the two steroids
to mediate the cell differentiation. P from the corpora lutea becomes the predominant
hormone regulating decidualization while the role of E in this phase remained unclear.
Although administration of ER antagonist ICI obliterated stromal decidual response in mice,
the uteri of the ER knockout animals did respond to an artificial stimulus inducing
decidualization (27, 28). However, studies have later identified the presence of a truncated
but functional ER transcript in these knock out mouse capable of mediating DNA binding
activity in the uterus (29). In order to address the possibility that the decidualization observed
in this global knock out model might be a consequence of residual ER activity, we generated
a conditional ER knock out mouse using the Cre-loxP technology. Ablation of ER in the
stromal cells revealed a severe defect in uterine decidual response and thus established the

13
pivotal role of this hormone receptor in stromal cell differentiation (Laws M.J and Bagchi
I.C, unpublished observation). The obvious importance of ER during decidualization
thereby questioned the probable mechanism of its action when the circulatory level of the
ovarian steroid is almost undetectable.
1.3.2.a Intrauterine E is critical during decidualization - emerging concepts
In a recent study from our laboratory, we identified that during the decidual phase of early
pregnancy, the uterus harbors the entire steroid biosynthetic machinery resulting in the
biosynthesis of local E. Further, as will be discussed further in the following chapters, this
intra-uterine E plays pivotal role in mediating stromal cell differentiation and uterine neo-
angiogenesis to critically sustain implantation.
The intrauterine E from differentiating stromal cells drives ER function during
decidualization (30). In addition to the expression of the major enzymes like side chain
cleavage (P450 SCC), StAR protein, 3 hydroxysteroid dehydrogenase (3HSD) and 17
lyase, the key enzyme regulating the conversion of testosterone to E, P450 aromatase is also
induced in decidual uterus. Blockade of aromatase activity impaired intrauterine E action
resulting in a severe decidualization defect and embryo resorption. The altered stromal
differentiation was identified by significant reduction of the decidual mass and down
regulation of known bio-markers of decidualization like Alkp and Prp.
1.3.2.b. Intrauterine E regulates angiogenesis during decidualization
During early pregnancy while the implantation chamber undergoes morphological
transformation, neo-angiogenesis becomes essential for the sustenance of the embryo.
Initially, the microvascular permeability occurs at specific regions of the uterine horn to
demarcate the implantation sites in rodents. With the progression of decidualization, new
blood vasculature begins to accumulate primarily at the mesometrial end. Steroid hormones
are well characterized regulators of vasculogenesis in pregnant uterus. Ovarian E is a known
enhancer of the vascular permeability at the site of blastocyst attachment. However, hormonal
regulation of blood vessel formation in ovariectomized mice have pointed that E might be a
negative regulator of angiogenesis, while P mediates it positively (31). Recent studies from
our laboratory have identified that the local E is critical in sustaining the decidual blood
vasculature by regulating the expression of a large repetoire of angiogenic modulators (30).

14
1.3.2.c. Downstream mediators of E action
Progesterone receptor
PR, the critical mediator of the implantation process, is regulated by E signaling. Prior to
implantation, ovarian E acting through ER governs the expression of stromal PR and
mediates the downstream signaling events. Following blastocyst attachment the declining
level of ovarian E switches with the uterine source and sustains PR expression to regulate the
decidualization process. Consequently, ablation of aromatase enzymatic activity altered PR
expression and thus inhibited a cohort of differentiation responsive genes downstream of PR
action. These included known markers like BMP2 and Wnt4.
Connexin 43 (CX43)
The gap junction protein CX43 is a well characterized E regulated gene in the uterus (32).
Close association of this membrane protein with the differentiating stromal cells indicated a
predominant role in decidualization. Recent studies in our lab have identified that disruption
of CX43 expression in mouse uterus by conditionally deleting the protein in PR positive
stromal cells results in altered cell differentiation. The expression of Prp is significantly de-
regulated in CX43 conditional knockout mice compared to the wild type littermate. Further,
the expression of two well characterized molecules of decidualization, BMP2 and HOXA10
are also significantly impaired after silencing of CX43 gene. CX43 plays a pivotal role in
decidual cell communication by regulating the passage of small molecules, ions and second
messenger signaling candidates. Disruption of this cellular crosstalk resulted in altered
stromal cell differentiation as observed in the null uteri. This defect in the decidual response
further regulated the release of angiogenic modulators like VEGF, HIFs and angiopoetins to
contribute to the uterine vascular growth. Accordingly, deletion of CX43 in decidual cells
had a severe consequence in the endothelial cell maturation. This defect in vascular
networking in the knockout animals resulted in embryo resorption by day 8 of pregnancy and
the consequent infertility (33). Administration of letrezole impairing aromatase enzymatic
activity had a severe effect in regulating uterine vascular growth. The down regulation of
CX43 in the aromatase inhibitor treated uterus indicated that this molecule is one of the prime
mediators of the local E action contributing to the development of uterine angiogenic network
indirectly via the decidual cell regulation.

15
VEGF signaling
Vascular endothelial growth factor (VEGF) is the most potent mitogen for endothelial cells
and a key regulator of angiogenesis (34). Extensive analyses on the expression of VEGF, its
steroid hormone regulation and the involvement of specific receptors during implantation, has
been extremely informative in understanding the functioning of the VEGF system in pregnant
uterus (35). VEGF, a heparin binding homodimeric glycoprotein, mediates its action through
the tyrosine kinase receptors namely VEGFR1 (Flt-1), VEGFR2 (Flk-1) and the glycoprotein
co-receptor Neuropilin-1 (NRP1). VEGFR-2 is the predominant receptor mediating VEGF
action in the uterus (36). Angiopoeitins are another class of angiogenic regulators that
complement the VEGF signaling for the development of angiogenesis. It mediates its action
through the tyrosine kinase receptor Tie2. Angiopoietin 2 collaborates with VEGF in front of
invading vascular sprouts to promote vascular remodeling during implantation (37). Pregnant
uterus exhibits prominent expression of Ang-1, Ang-2, Ang-3 in differentiating stromal cells
along with the expression of Tie2. Hypoxia inducible factors (HIFs) are major transcriptional
regulators of VEGF. HIFs are primarily involved in governing the physiological balance of
oxygen tension and are closely associated with angiogenesis. HIF2 complements the action
of HIF1 to regulate VEGF expression (38). Our studies indicated that angiopoeitins and
HIF2 are regulated by the local E during decidualization thereby contributing to VEGF
regulation and vasculogenesis.
Adrenomedullin (ADM)
ADM is a 52 aminoacid vasodialator peptide with major role in uterine vascular
development. ADM binds to the G protein coupled receptor calcitonin receptor like receptor
(CL) which is associated with a receptor activity modifying protein (RAMP). During early
pregnancy, ADM exhibits strong expression in the stromal cells at the immediate vicinity of
the embryo, while its receptor proteins are restricted to the endothelial cell layers (39, 40).
Heterozygote mice lacking one functional allele of ADM exhibited impaired implantation
due to uterine receptivity failure. The localization pattern of the ADM signaling components
strongly suggests the indirect mechanism of endothelial cell maturation during
decidualization. ADM and RAMP2 are known regulators of E action in uterus. The
regulation of ADM by the intrauterine E indicated its major contribution in the development
of vascular structures in the decidual uterus.

16
1.4. Additional regulators critical for stromal cell differentiation
Interleukin-11 (IL-11)
IL-11 is a pleiotropic cytokine with predominant role in inflammation and wound repair. IL-
11 action is mediated by its receptor alpha chain IL11R and gp130. Expression of both the
ligand and the receptor is induced following embryo attachment during stromal
differentiation. While the expression of the ligand in the uterus is restricted to the primary
decidual region, IL-11R is confined to the secondary zone. The precise expression pattern
indicated that induction of IL-11 adjacent to the embryo acted on the concentric outer layer
expressing the receptor to regulate progression of stromal differentiation. Deletion of IL-
11R disrupts IL-11 signaling and renders mice infertile with a severe defect in
decidualization (41, 42). Vascular permeability surrounding the implantation sites on day 4
was reduced in these receptor null mice indicating reduced capillary permeability and blood
flow at the site of blastocyst apposition. Although the attachment reaction did occur in the
null uteri, the formation of the secondary decidual bed was significantly impaired. By day 8.5
of pregnancy, the blastocysts in the null uterus were nectrotic and the decidual mass was
infiltrated by inflammatory cells resulting in regression of the decidual mass and blastocyst
resorption.
Cyclooxygenase-2 (COX-2)
Prostaglandins are biosynthesized from arachidonic acid via the cyclooxygenase (COX)
pathway. The COX enzymes govern the conversion of arachidonic acid to prostaglandin H2,
which is acted upon by various prostaglandin synthases to form prostaglandins. The two
isoforms of COX enzymes, COX-1 and COX-2, are expressed in the mouse uterus during
implantation. While ablation of the first isoform does not exhibit a fertility defect, mutation
of the later has severe consequences in blastocyst attachment and decidualization (43). COX-
2 is expressed in the luminal epithelium and subepithelial stroma at the time of blastocyst
attachment and initiation of decidualization. Administration of the prostacyclin analogue
PGI2 restores the decidualization defect in the COX-2 null uteri, indicating that it might be
the major functional prostaglandin during implantation. Although COX-2 expression was
induced after E treatment surrounding the embryo during delayed implantation, in the ovex
model the steroid regulation of COX-2 expression was not apparent (44). Recent studies have
identified that the COX enzymes are post-transcriptionally regulated by two microRNAs

17
(miRNA) mmu-miR-101a and mmu-miR-199a, expressed in the uterus during the
implantation window (45).
1.5. Additional regulators critical for angiogenesis
Uterine dendritic cells
During the implantation a special population of hematopoietic cells known as uterine
dendritic cells (uDc) are involved in the coordination of innate, adaptive immune responses
and induction of immunologic tolerance (46). In recent studies the function of these uDc were
analyzed by conditional depletion of these cells during implantation. In addition to impaired
decidualization, the study reported that this cell population controlled the vessel maturation
during uterine angiogenesis. The parallel expression of an angiogenic agonist (TGF1) and
antagonist (sFlt1) in these cell population indicated that uDcs are involved in controlling the
extent of vascular growth during decidualization (47). Removal of uDCs suppressed
angiogenic networking in the decidual uterus. Modulation of uterine vasculature by uDcs
suggested a parallel regulatory mechanism in addition to the steroid hormones.
COX-2
As decidualization progresses the COX-2 localization switches from the antimesometrial to
the mesometrial pole, suggesting a probable role of the enzyme in angiogenesis. Studies in
the knockout model have identified that COX-2 regulates the vasculogenic development
during early decidualization. This effect is primarily brought about by regulating the VEGF
expression in the mesometrial region and to a lesser extent by governing the expression of
molecules involved in the angiopoeitin system (48).
1.6. Decidualization in human endometrial stromal cells
The molecular cross talk between the human endometrium and the conceptus directs a
comprehensive stromal differentiation program in pregnant women. During every menstrual
cycle, however, the orchestrations of cell differentiation independent of pregnancy are
governed by local molecular cues from the epithelial compartment and the steroid hormones.

18
The increasing exposure of the endometrium to cytokines, local endocrine factors like
prostaglandin E2, relaxin and corticotrophin releasing hormone (CRH) stimulates the
production of a second messenger cyclic adenosine mono-phosphate (cAMP). Activation of
the PKA pathway through cAMP is an obligatory signaling event driving the differentiation
of human endometrial stromal cells. Consequently it has been observed that elevated levels of
cAMP resulting in sustained activation of the PKA pathway and cAMP induced sensitization
of cells to steroid action are an absolute necessity to drive these stromal cells differentiation
(4).
In vitro human stromal cell decidualization
Undifferentiated stromal cells isolated from endometrial biopsies of healthy individuals at the
proliferative stage of the menstrual cycle can be differentiated in vitro in response to a
hormone cocktail containing E (10nM) P (1μM) and a membrane permeable analogue of
cAMP, 8Br-cAMP (0.5mM). The induction of differentiation is observed by a distinct
transition of the cellular morphology from spindle like fibroblast appearance to terminally
differentiated plump epitheloid shape. Additionally the expression of the various markers of
human stromal differentiation like prolactin (PRL), insulin like growth factor binding protein-
1 (IGFBP-1), cytokine interleukin-11 (IL-11), vascular endothelial growth factor (VEGF) are
progressively induced as differentiation progresses in response to steroid hormones and
cAMP.
1.6.1. Bio-markers of human stromal cell decidualization
Insulin like growth factor binding protein-1 (IGFBP-1)
Regulation of insulin like growth factor (IGF) signaling by the IGF binding proteins
(IGFBPs) is a critical aspect of human endometrial differentiation. As stromal cells begin to
differentiate it secretes increasing amount of IGFBP-1 and this protein is undoubtedly
considered as the major regulator of the human endometrial decidualization (49). At the
molecular level, several signaling pathways converge in inducing IGFBP-1 expression during
stromal cell differentiation in humans and other primates. Steroid hormone receptor PR
directly regulates IGFBP-1 expression by binding to the PRE in a ligand dependent manner.
Downstream target of PR, the Forkhead transcription factor, FOXO-1 interacts with HOXA-
10 to regulate IGFBP-1 expression (50). Activation of PKA pathway further accentuates the
expression level of this secretory protein.

19
Prolactin (PRL)
PRL is a peptide hormone and cytokine, involved in regulating growth and differentiation of
a number of epithelio-mesenchymal organs. Secretion of PRL under the control of the tissue
specific decidual PRL (dPRL) promoter indicates the extent of stromal cell differentiation in
human endometrium (51). PRL secretion is induced by P, insulin, IGF-1 and PRL releasing
factor (52). Activation of the PKA pathway through cAMP further enhances the expression of
this decidual marker. PR indirectly regulates PRL expression by activating FOXO-1, which
potently enhances the PKA dependent activation of decidual PRL promoter. FOXO-1
interacts with a CCAAT family transcription factor C/EBP beta and subsequently binds to the
proximal promoter of dPRL to regulate PRL expression during stromal cell differentiation
(53).
Interleukin-11 (IL-11)
IL-11 is a cytokine with pleiotropic actions on various tissues. During the mid-secretory
phase, the differentiating stromal cell expresses increasing concentration of IL-11 in response
to cAMP. It has also been observed that addition of exogenous IL-11 in vitro endometrial
cultures induces the expression of decidual markers, PRL and IGFBP-1(54).
Vascular Endothelial Growth Factor (VEGF)
VEGF is another cytokine and angiogenic factor with prominent expression in the
differentiating stromal cells. Since the expression of VEGF is induced during stromal
differentiation in response to steroid hormones and cAMP, this molecule is also considered as
a major marker of decidualization (55).
1.6.2. Hormonal regulation of the human menstrual cycle
The cyclical change of the endometrium is coordinated by a series of hormone action.
Activation of the anterior pituitary by the gonadotropin releasing hormone (GnRH) from the
hypothalamus elicits the release of follicle stimulating hormone (FSH) and leuteinizing
hormone (LH) that acts on the ovarian follicles to initiate its maturation and the subsequent
release of the ovarian steroids. E released from these mature follicles in turn regulates the LH
surge by a feedback mechanism that organizes the follicular rupture. The circulating levels of
E reach around 300pg/ml in the follicular phase and it actively participate in governing the
growth and proliferation of the endometrium. Following ovulation, the newly formed corpus
luteum releases high level of P and to a lesser extent E. Under the influence of P, stromal

20
cells transform to decidual cells thereby demarcating the secretory phase of the cycle. During
the early to mid secretory phase, the level of E remains around 150pg/ml while the levels
decreases dramatically in the late secretory phase to around 50pg/ml (56). This decline in the
E level at a stage when cells are actively involved in differentiation indicates a possible
replenishment of E level through other mechanisms. Studies by Tseng et al. have identified
increasing aromatase enzymatic activity in differentiating cultured human endometrial
stromal cells in response to forskolin and P.
Steroid hormones in human reproductive cycle mediate their effect both through the
classical and non classical pathway. P which is the major steroid during decidualization
mediates its action through PR. Human endometrium has predominant expression of PR-A
and is considered as the major functional nuclear receptor in vivo and in vitro (57). In
response to cAMP signaling, PR-A directly recruits other activated transcription factors like
STAT 5, CEBP, FOXO1 to mediate stromal decidualization. E mediates its action though
the ER and ER. Although ER is the functional form regulating E action in the
proliferative phase, the secretory phase has prominent expression of both ER and ER.
Differentiating human endometrial stroma expresses the major enzyme regulating E
biosynthesis P450aromatase resulting in picogram/ml levels of endogenous E production.
Silencing of aromatase expression in cultured stromal cell identified its contribution in
mediating cell differentiation and regulation of VEGF expression. Endogenous E drives ER
function during human decidualization. siRNA mediated silencing of ER resulted in a
severe defect in decidualization. In addition to an obvious defect in cell shape indicating
more fibroblast morphology compared to decidual, there was a significant impairment in the
expression of the markers of stromal differentiation like IGFBP-1, PRL, IL-11, VEGF,
FOXO1 and others.
1.6.3. Molecules governing human stromal cell decidualization
Of the several signatory molecules that define human differentiation program, the major ones
are discussed below.
CCAAT/ enhancer-binding protein beta (C/EBP)

21
C/EBP is involved in regulating the expression of both PRL and IGFBP-1. These decidual
markers have prominent C/EBP binding sites in their promoter. C/EBP associates with PR-
A and regulates promoter interaction. It exhibits strong nuclear localization in the stromal
cells of the late secretory phase. Additionally in vitro differentiation of stromal cells in
response to P and cAMP also induces C/EBP expression, indicating that this transcription
factor might play a central role in mediating human endometrial stromal cell decidualization.
Transforming growth factor (TGF) family
The TGF family of proteins including the sub-families of TGF, activin, BMP and GDF have
known contribution in apoptosis, proliferation, differentiation and tissue remodeling. The
expression profiles of these molecules have been widely studied in the endometrium to
address their probable role in human stromal cell differentiation.
Activins constituting the subunits A and B, dimerize to form activin A, B or AB. This
subunit is expressed in the glandular epithelium of human endometrium in the proliferative
phase. Their expression in the stromal cells is particularly induced in the mid-late secretory
phase when the cells enter the differentiation program. In-vitro differentiation of endometrial
stromal cells in response to cAMP also induces activin A indicating a predominant role of the
molecule during decidualization (58). Further, exogenous administration of recombinant
activin to cultured stromal cells undergoing decidualization enhanced the secretion of known
decidual markers PRL and IGFBP1. The effect of activin A in cell differentiation can be
attenuated by administration of follistatin, a naturally occurring activin antagonist (59). Thus
parallel expression of these two antagonistic molecules regulates the extent of stromal
differentiation in human endometrium.
Bone morphogenetic proteins, (BMPs) play a critical role in stromal cell differentiation.
The differentiating cells in the secretory phase exhibit high expression of BMP2 and BMP4,
although the expression of the latter is not induced but constitutively expressed in the stromal
cells (60). BMP2 is a phylogenetically conserved morphogen important in both mouse and
human endometrium. Early induction of BMP2 during in vitro differentiation suggested a
possible steroid regulation of this molecule. Administration of recombinant BMP2 in cultured
stromal cells also enhances the expression of decidual markers PRL and IGFBP1 indicating
this morphogen is a major mediator of endometrial stromal cell differentiation (17).

22
In addition to activin and BMPs, TGF1 and GDF 5, 8 and 11 are expressed in the stromal
cells with prominent induction in decidualizing cells of the secretory phase. The strong
expression of these molecules further confirmed their role in coordinating the endometrial
decidualization process in humans.
Forkhead transcription factor forkhead box O1 (FOXO1)
The winged helix/Forkhead class of transcription factors constituting the FOXO proteins
have important role in cell fate decisions in response to environmental factors and hormonal
signals. In the human endometrium several FOXO proteins are induced during the stromal
cell decidulization in the secretory phase. The major ones include FOXO1, FOXO3a and
FOXO4. FOXO1 is significantly induced in the secretory phase of the menstrual cycle in
vivo. FOXO1 expression in cultured human stromal cells is under the direct regulation of the
PKA pathway. P accentuates this induction however P alone is unable to mediate an
enhanced expression of the transcription factor (61). FOXO1 regulates the expression of the
two major differentiation markers PRL and IGFBP-1 (62). Biochemical analysis of the PRL
promoter has indicated that FOXO1 in cooperation with CEBP enhances the dPRL promoter
activity (53). FOXO1 also stimulates IGFBP-1 expression by a direct interaction with
HOXA10. Additionally in a recent study by (63), it was shown that in association with PR,
FOXO1 regulates cell cycle molecules and differentiation in cultured stromal cell
decidualization. These observations indicate that FOXO1 plays a major role in mediating
human stromal cell decidualization.
Signal transducer and activation of transcription-5 (STAT5)
The STAT family of proteins also plays an important role by regulating genes involved in
growth and differentiation. Receptor associated activation of janus kinases further induces the
phosphorylation of a tyrosine residue in STAT protein and its subsequent activation.
Activated STAT dimerizes and translocates to the nucleus to mediate the regulation of its
target genes. In the human endometrium, STAT5 is induced in the glandular epithelium.
During the secretory phase however, it shows predominant expression in the stromal cells
that express the PRL receptor. In vitro cultured stromal cells indicated a strong induction of
STAT5 expression followed by the activation and nuclear localization of STAT5 in response
to cAMP (64). STAT5 interacts with PR and plays a major role in activating PRL promoter.

23
v-src sarcoma (Schmidt-Ruppin A-2) viral oncogene homolog (SRC)
SRC is a non-receptor tyrosine kinase associated with integrins and G protein coupled surface
receptors. It plays an important role in converting extracellular stimuli to intracellular signals.
Activation of SRC in the presence of ligands involves dephosphorylation of the tyrosine
residues at the carboxy terminal. Recent in vitro studies in human endometrium identified
SRC as an important candidate mediating stromal cell differentiation. SRC expression is
regulated by the cAMP and PKA signaling pathways. Additionally, P also induces
SCR/MAPK activity by direct or indirect interaction of SRC with the ligand bound PR.
Administration of a dominant negative mutant of SRC kinase in cultured stromal cells
completely impaired the transformation of the fibroblast cells to the decidual cell type. The
mechanism of SRC action in the human endometrial cultures involved STAT 5 activation
another important regulator of stromal differentiation. Activation of STAT5 by
phosphorylation of a specific tyrosine residue is governed by SRC and in the presence of a
dominant negative SRC mutant this active form was absent (65).
p53
The transcription factor p53 is induced in decidualizing stromal cells in vitro in response to
cAMP treatment. p53 also shows strong nuclear expression in the stromal cells of the
secretory endometrium, indicating a possible role in stromal differentiation. Classically the
cell cycle repressor is involved in apoptosis however a similar possibility in differentiating
stromal cells is remote considering very minute fraction of cells undergoes cell death. The
major function of p53 in stromal decidualization remains unclear, although studies have
identified a direct physical interaction and transrepression between p53 and CEBP (66). In
mouse stromal cell differentiation, p53 is involved in regulating the transcription of LIF, a
major cytokine governing embryo reception (67). In a recent study, it has been shown that
high incidence of single nucleotide polymorphisms in the p53 pathway is more prevalent
among infertile individuals with unsuccessful in vitro fertilization (68).
1.6.4. Angiogenesis and development of spiral arteries during human menstrual cycle
The blood supply to the uterus is furnished by the arteries from ovarian and uterine origin.
The uterine arteries that enter the myometrium gives rise to the arcuate arteries which further

24
branch to form the radial arteries. The radial artery bifurcates to the basal arteries that supply
blood to the basal portion of the endometrium and governs endometrial regeneration after
menstruation; the other half forms the spiral arteries that supply blood to the superficial
functional endometrium (69). The spiral arteries eventually form subepithelial capillary
plexeus that plays a critical role in regulating blood supply to the endometrium and is the
initial site of stromal cell differentiation during the mid-secretory phase.
A functional interface between the maternal and fetal circulations is extremely critical for
the maintenance of pregnancy. This involves major physiological changes in the maternal
vasculature as the endometrium prepares itself for the blastocyst during each mid-secretory
phase of the menstrual cycle. Angiogenic development in the cyclical uterus sequentially
occurs during the (a) regeneration of blood vessel after menstrual sloughing, (b) the
proliferative phase when endometrium grows predominantly under the influence of E, and
finally during (c) the secretory phase, when the development of the spiral arteries becomes
important for the initiation of stromal cell transformation. Some of the known mediators of
the angiogenic signals in the human endometrium include VEGF, its receptors Flk-1, Flt-1,
bFGF and angiogenin.
VEGF is the major cytoine regulating angiogenic network in the cycling human
endometrium. While the superficial stromal cells express high levels of VEGF, its high-
affinity tyrosine kinase receptors receptors Flk-1 and Flt-1 are expressed in the endothelial
cells, indicating a probable paracrine interaction between the two cell compartments (70).
VEGF expression in the human endometrium is cyclical with strong expression in the late
proliferative phase that coincides with the edema and hypervascular permeability. VEGF
expression is also significantly induced in the mid-to late secretory phase and aids in
regulating the angiogenesis and development of the spiral arteries in this phase (55). The gap
junction protein CX43 also regulates the VEGF expression in human endometrium and
significantly contributes to cell differentiation (33).
Basic FGF is a18kd peptide belonging to the heparin-binding family of growth factors with
known mitogenic properties. Along with VEGF it also plays major role in angiogenesis in
vitro and in vivo. In the human endometrium, bFGF is induced in the proliferative phase, but
the expression is persistent in the secretory phase and contributes towards capillary
angiogenesis (71).

25
Angiogenin (Ang) is another characteristic polypeptide with predominant role in vascular
growth. In the human endometrium, Ang is induced in the secretory phase. In vitro studies
have also identified that the expression of Ang is regulated by P, which accentuates the
polypeptide expression as the stromal cells undergo differentiation. Ang is involved in the
convulation and thickening of arterioles in the secretory phase (72).

26
1.7. FIGURES
Figure 1.1: Morphological structure of uterine cross-sections during implantation
Figure 1.1: Morphological structure of uterine cross-sections during implantation Uterine cross-sections from pregnant CD-1 females were obtained. Sections from day 2, and implantation sites on day 4 midnight, day 5, day6, day7 and day8 were stained with hematoxylin and eosin and the representative images are shown at the same magnification.
D2 D4 midnt D5
D6 D7 D8
Figure 1.1Figure 1.1

27
Figure 1.2: Major hallmarks of implantation
Figure 1.2: Major hallmarks of implantation Frozen uterine cross sections isolated on day7 of pregnancy were subjected to alkaline phosphatase enzymatic activity assay (panel A) and immunostaining for PECAM-1 (panel B) to monitor the extent of cell differentiation and angiogenesis, respectively.
A B
Figure 1.2Figure 1.2

28
Figure 1.3: Mouse in vitro stromal cell culture
Figure 1.3: Mouse in vitro stromal cell culture Stromal cells isolated from pre-implantation pregnant mouse were cultured in complete medium. Decidualization was induced by the addition of P. Cells were collected at 5 hrs and 24hr intervals thereafter to monitor A: the morphological changes. Cultured cells were stained with phallodin (green), counter stained with DAPI (nucleus) and images were procured at the same magnification. B: the expression profile of the differentiation markers alkaline phosphatase (upper) and Prp (lower).
72hr 96hr
48hr
5hr
24hr
Figure 1.3A
0hr 24hr 48hr 72hr
Alkp
PrP
0hr 24hr 48hr 72hr 96hr
Figure 1.3B
Figure 1.3A
*
*
*
p<0.001 n= 8
*

29
Figure1.4: Steroid hormone nuclear receptor during implantation
Figure 1.4: Steroid hormone nuclear receptor during implantation-ER Estrogen receptor alpha expression in the pre-implantation (day 1 and day 2) and implantation uterus (day 4 at 0:00hrs, day 5, day 6, day 7 and day 8 at 9:00hrs). Strong nuclear expression is predominantly observed in the luminal epithelium in day 1. The stromal expression is prominent from day 2 along with strong glandular expression. By day 5, wide spread expression of ER is observed over the entire uterine stroma, while day 6 onwards it becomes more restricted to the mesometrium. The figure shows the low (4X) and high (40X) magnification of a representative field from each pregnant sample.
D1 D1 D5 D5
D2 D2D6 D6
D4mn D4mn D8 D8 meso
Figure 1.4AFigure 1.4A

30
Figure1.4: Steroid hormone nuclear receptor during implantation
Figure 1.4: Steroid hormone nuclear receptor during implantation-PRA Progesterone receptor A expression in the pre-implantation (day 1 and day2) and implantation uterus (day 5, day 6, day 7 and day 8 at 9:00hrs). Strong nuclear expression is predominantly observed in the stromal cells surrounding the embryo on day 5. The expression remains persistent at the antimesometrial end on days 7 and 8 of pregnancy. Strong expression surrounding the blood vessels is prominent on day 8 at the mesometrial end. The figure shows the low (4X) and high (20X and 40X) magnification of a representative field from each pregnant sample.
D1 D1 D6 D6
D2 D2 D7 D7 ameso D7 meso
D5 D5 D8 D8 ameso D8 meso
Figure 1.4BFigure 1.4B

31
Figure 1.5: Steroid hormone profile during implantation
Figure 1.5: Steroid hormone profile during implantation Schematic representation of the hormonal profile during early pregnancy. A: High levels of ovarian E and rising level of P are exhibited during the pre-implantation phase. On day 4, a transient surge of ovarian E is occurs specifically in mice. Following attachment, the P level rises significantly and predominates the differentiation phase, while the level of E declines to very low levels. B: The human menstrual cycle is dominated by high E levels in the proliferative phase. Following ovulation, the P levels become dominant along with low levels of E in the secretory phase.
Day 1Day 8
Attachment
EE
PP
Preimplantation Phase Decidualization
Ovarian steroids E and P
Un-detectable level of ovarian E
Figure 1.5A
Estrogen
Progesterone
Proliferative Phase Secretory PhaseO days 4 days 21 days
14 days28 days
Menstruation
Figure 1.5B
Figure 1.5A
Figure 1.5B

32
Chapter-2
De Novo synthesis of estrogen in pregnant uterus is critical for stromal cell
decidualization
______________________________________ A part of this chapter has been published as “De novo synthesis of estrogen in pregnant uterus is critical for stromal decidualization and angiogenesis”, authored by Amrita Das, Srinivasa Raju Mantena, Athilakshmi Kannana, Dean B. Evans, Milan K. Bagchi and Indrani C. Bagchi in the Proceedings of National Academy of Sciences, July 28, 2009, doi: 10.1073/pnas.0901647106

33
2.1. ABSTRACT
Implantation is initiated when the embryo attaches to the uterine luminal epithelium during
early pregnancy. Following this event, uterine stromal cells undergo steroid hormone-
dependent transformation into morphologically and functionally distinct decidual cells in a
unique process known as decidualization. An angiogenic network is also formed in the
uterine stromal bed, critically supporting the early development of the embryo. The steroid-
induced mechanisms that promote stromal differentiation and endothelial proliferation during
decidualization are not fully understood. Although the role of ovarian progesterone (P) as a
key regulator of decidualization is well established, the requirement of ovarian estrogen (E)
during this process remains unresolved. Here we show that the expression of P450 aromatase,
a key enzyme that converts androgens to E, is markedly induced in mouse uterine stromal
cells undergoing decidualization. The aromatase then acts in conjunction with other steroid
biosynthetic enzymes present in the decidual tissue to support de novo synthesis of E. This
locally produced E is able to support the advancement of the stromal differentiation program
even in the absence ovarian E in an ovariectomized, P-supplemented pregnant mouse model.
Administration of letrozole, a specific aromatase inhibitor, to these mice blocked the stromal
differentiation process. Collectively, these studies identified the decidual uterus as a novel
site of E biosynthesis and uncovered E-regulated maternal signaling pathways that critically
control uterine differentiation and angiogenesis during early pregnancy.

34
2.2. INTRODUCTION
Implantation involves a series of complex interactions between the developing embryo and
the uterus, leading to the establishment of pregnancy (1, 2, 73, 74). Although the details of
the implantation process vary between species, the basic features of blastocyst attachment
and penetration of the uterine surface epithelium are common to many mammals. In mice,
implantation is initiated four days after fertilization when the blastocyst reaches the uterus.
The concerted actions of the steroid hormones estrogen (E) and progesterone (P) via their
cognate receptors orchestrate the changes in the uterine tissue that make it competent to
attach to the blastocyst and initiate the process of implantation.
In the mouse, an experimentally induced delayed implantation model provided the
evidence that E plays an essential role in triggering the attachment of the embryo to the
uterine luminal epithelium (24, 25). In these mice, which have undergone ovariectomy on day
4 of gestation, implantation does not occur in the absence of ovarian E. continued
administration of p allows the blastocysts to remain viable within the uterus but they fail to
attach to the luminal epithelium in the absence of E. administration of E to these
ovariectomized pregnant mice allows attachment of the blastocyst trophectoderm to the
luminal epithelium within 12-24 hours, demonstrating a critical function of e-dependent
signaling in initiating the implantation process (3, 4).
Once the embryo attaches itself to the uterine epithelium, the underlying stromal cells start
to proliferate and then differentiate into unique decidual tissue that form the implantation
chamber. Paracrine factors secreted by the decidual cells are critical regulators of uterine
remodeling, maternal immune response, uterine angiogenesis and early embryonic growth.
Decidualization is thus a prerequisite for successful implantation and establishment of
pregnancy.
Previous studies established that P acting via its receptor plays a central role in regulating
decidualization (5). In contrast, a functional requirement of E beyond the embryo attachment
step remained ambiguous. An earlier study showed that administration of P alone to
ovariectomized mice sustained the decidual response during experimentally induced
decidualization, indicating a non-obligatory role of exogenous E in regulating this process
(27). Conversely, it was reported that administration of ICI 182780, an estrogen receptor

35
(ER) antagonist, to mice severely impairs the formation of the decidual tissue, hinting that E
acting via ER regulates this process (75). These apparently contradictory reports prompted us
to further investigate the role of E in the uterus during decidualization.
In this study, we demonstrate, for the first time, that the decidual uterus is a novel site of de
novo synthesis of E. The expression of P450 aromatase, a key enzyme that converts
testosterone into E, is markedly induced in the pregnant mouse uterus during decidualization
and plays an essential role in this process. Even in the absence of ovarian E, this locally
synthesized E is able to support the advancement of the stromal differentiation program in an
ovariectomized, P-supplemented pregnant mouse model.
2.3. EXPERIMENTAL PROCEDURES
Reagents-
Progesterone and 17--estradiol were purchased from Sigma Chemical Co., St. Louis, MO.
Letrozole was synthesized in laboratories of Novartis Pharma AG, Basel, Switzerland.
Animals and Tissue Collection-
All experiments involving animals were approved by the Animal Care Committee at the
University of Illinois at Urbana-Champaign and the studies were conducted in accordance with
the National Institutes of Health standards for the use and care of animals. Female mice (CD-1
from Charles River, Wilmington, MA), in proestrus, were mated with adult males. The presence
of a vaginal plug after mating was designated as day 1 of pregnancy. The animals were killed at
various stages of gestation and the uteri were collected. The uteri were freed of embryos by
repeated flushing.
In some experiments, mice were ovariectomized on day 5 (morning) of pregnancy and injected
daily with P or P in combination with letrozole (20 mg/kg body weight) from days 5-8. Mice
were killed 6 h after the last injection and uteri were isolated.

36
Artificial Decidualization-
Decidualization was experimentally induced in non-pregnant mice as described previously
(76). Mice were ovariectomized and 14 days later were subcutaneously injected with E (2
g/kg body weight) in 0.1 ml of sesame oil for three consecutive days. This was followed by
daily injections of P (40 mg/kg body weight) for three consecutive days. Decidualization was
then initiated in one horn by injection of 20l oil. The other horn was left unstimulated. The
animals were treated with P or P plus letrozole for additional three days following decidual
stimulation and then killed to collect the uterine tissue.
Laser Capture Micro dissection RTPCR-
Sections of uteri collected on d 4, 5, and 6 of pregnancy were subjected to LCM. Frozen
sections (10 m) containing the implantation sites were placed on polyethylene naphthalate
membrane slides (Molecular Devices) and dehydrated serially in ethanol and xylene. Using
the Veritas microdissection instrument (Molecuar Devices), the mesometrial and anti-
mesometrial regions were excised and the decidual tissues were obtained. RNA was isolated
from these tissues using a Pico Pure RNA isolation kit according to the manufacturer’s
instructions. cDNA was synthesized from the isolated RNA using the Sensiscript reverse
transcriptase kit (Qiagen) and subjected to real-time PCR using gene-specific primers.
Immunohistochemistry-
Polyclonal antibodies against aromatase (Abcam Inc, Cambridge, MA), Ki67 (BD Pharmingen,
San Jose, CA), connexin-43 (Zymed Laboratories INC, Carlsbad, CA), and PECAM-1 (BD
Pharmingen, San Jose, CA) were used for immunohistochemistry. Paraffin-embedded uterine
tissues were sectioned at 4 µm and mounted on slides. Sections were washed in PBS for 20 min
and then incubated in a blocking solution for 40 min before incubation with primary antibody
overnight at 40C. Immunostaining was performed using an Avidin-Biotin kit for rabbit primary
antibody (Vector Laboratories, Burlingame, CA). Sections were counterstained with
hematoxylin, mounted, and examined under bright field. Red deposits indicate the sites of
immunostaining.

37
Microarray Analysis-
Mice were treated with or without letrozole and subjected to experimentally induced
decidualization. Uteri were collected from untreated or treated animals at 72 h following the
application of decidual stimulus (n=5). Total RNA was prepared from these tissues and subjected
to microarray analyses using Affymetrix mouse arrays (GeneChip® Mouse Genome 430 2.0
Array) following the Affymetrix protocol as described previously (58).
Assay of Aromatase Activity-
The aromatase activity in uterine and ovarian homogenates was determined by the tritiated water
release assay as described previously (77). Briefly, 250 l of uterine or ovarian homogenates was
incubated with 300 pmol of [1-3H] androstenedione for 6 h at 37 C. The results were calculated
as fmol of [3H] water released per 24 h per mg of tissue. The aromatase activity was expressed as
meanSEM of data derived from three independent experiments.
Estradiol Measurement in Uterine Homogenates-
The measurements of estradiol in uterine homogenates were performed by the Center for
Research in Reproduction Ligand Assay and Analysis Core of the University of Virginia,
Charlottesville, VA.
2.4. RESULTS
2.4.1. Ovarian E is not essential for decidualization
In the mouse, attachment of the embryo to the uterine epithelium occurs on day 4 (midnight)
of pregnancy. This event initiates the process of decidualization, which proceeds through
progressive phases during days 5-8 of gestation. To assess the role of ovarian E during
decidualization, mice were ovariectomized on the morning (6 AM) of day 5, approximately 6
h following the attachment of the embryo to the uterine epithelium and then treated with
exogenous P for three consecutive days (Fig. 2.1A). We found that administration of P alone
to the ovariectomized pregnant mice maintained the decidualization process, and the growth
and development of the implanted embryos (Fig. 2.1B). In these P-treated ovariectomized

38
uteri, the spatio-temporal expression of known markers of decidualization, such as
progesterone receptor (PR) and prolactin-like protein type B (PLP-B), was found to be
similar to that seen in normal pregnant uterus, indicating that P is sufficient to sustain
decidualization in the absence of exogenous E (Fig. 2.1C). Collectively, these results
indicated that ovarian E is not essential for maintenance of decidual response.
2.4.2. Evidence for biosynthesis of E in the decidual uterus
We next considered the possibility that production of E from an extra-ovarian source, such as
the uterus itself, may contribute to the decidualization process. To investigate whether the
uterus has the capacity to synthesize E de novo, we monitored the expression of various
steroid biosynthetic enzymes in this tissue during early pregnancy. Total RNA was obtained
from pregnant uteri on day 4 (morning) preceding implantation and on different days during
the decidualization phase. The RNA samples were analyzed by reverse-transcription-coupled
polymerase chain reaction (RT-PCR) to monitor the expression of various steroidogenic
factors (Fig. 2.2A). We observed prominent expression of StAR (steroidogenic acute
regulatory protein), P450SCC (P450 side chain cleavage enzyme), P450C17 (17lyase), 3 HSD
(3beta-hydroxysteroid dehydrogenase), and 17-HSD-1 (17beta-hydroxysteroid
dehydrogenase type 1) in the pregnant uterus in both preimplantation (day 4 morning) and
decidualization (days 6 and 7) stages (Fig. 2.2B). Interestingly P450 aromatase, the product
of the CYP19 gene, which converts testosterone to biologically active E exhibited markedly
altered expression in pregnant uterus. The expression of aromatase mRNA was undetectable
in the preimplantation uterus on day 4. However, a robust induction of this mRNA was
observed in the decidual uterus on days 6 and 7 of pregnancy. Further analysis by Northern
blotting confirmed that the expression of aromatase mRNA is initiated on day 5 of pregnancy
and increases further on day 6 as decidualization progresses. It was diminished significantly
on day 10 with the cessation of the decidual phase of gestation (Fig. 2.2C).
To further establish that the decidual cells are the actual sites of aromatase mRNA
expression during pregnancy, we performed laser capture microdissection (LCM) to isolate
these cells from uterine sections. Total RNA was prepared from the excised tissue and the
expression of mRNAs corresponding to aromatase and alkaline phosphatase, a well-
established biomarker of decidual cells (8), was assessed by real-time PCR. A significant rise

39
in the level of aromatase mRNA expression, relative to its level on day 4 of pregnancy, was
observed in the stromal tissue excised from the antimesometrial region on day 5 of gestation
(Fig. 2.2D). A dramatic increase (~12 fold compared to day 4) in the level of both aromatase
and alkaline phosphatase mRNAs was seen in the decidual cells collected from the
antimesometrial area of uterine sections on day 6 of pregnancy. Collectively, these results
confirmed that decidual cells are the actual sites of aromatase mRNA expression in the
pregnant uterus.
We next examined the spatial expression of the P450C17 and aromatase proteins in normal
pregnant mouse uterus using immunohistochemistry (Figs. 2.2E and 2.2F). The P450C17
antibody, as expected, showed specific immunostaining in the Leydig cells of testis (Fig.
2.2E panel a). Probing of uterine sections on day 6 of pregnancy with this antibody revealed
prominent expression of P450C17 in decidual cells surrounding the implanted embryo (panel b
and c). The authenticity of the aromatase antibody was first confirmed by immunostaining of
sections of ovaries obtained from mice treated with pregnant mare serum gonadotropin
(PMSG) (Fig. 2.2F panel a). As expected, intense aromatase expression was observed in the
granulosa cells of the ovarian follicles stimulated with PMSG. Sections of ovaries collected
from mice treated with human chorionic gonadotropin (hCG), which induces follicular
rupture, luteinization, and suppression of aromatase expression, showed very little aromatase-
specific immunostaining (panel d). Immunohistochemical analysis of uterine sections showed
no detectable aromatase immunostaining on day 4 (morning) of pregnancy prior to embryo
attachment (panel b). However, on day 5 (panel c) and day 6 (panels e and f) of pregnancy,
prominent expression of this enzyme was seen in decidualizing stromal cells at both
antimesometrial and mesometrial regions surrounding the implanted embryo. Additionally,
when primary stromal cells were isolated from pregnant uteri (preimplantation stage, day 4)
and subjected to steroid-induced decidualization in vitro, the cytoplasmic staining of
aromatase was clearly evident in the decidual cells (right panel).
The finding that aromatase is expressed in the decidual uterus raised the possibility that
this tissue acquires the ability to produce E locally as it undergoes differentiation. We,
therefore, assessed the enzymatic activity of aromatase in the uterus during decidualization.
Mice were ovariectomized on day 5 (morning) of pregnancy, and treated with P in the
presence or absence of letrozole, a well-known aromatase inhibitor. The uteri were collected

40
on day 6 and tissue extracts were prepared. As shown in Table I, extracts of day 6 pregnant
uteri exhibited significant aromatase activity. Treatment of mice with letrozole resulted in a
drastic reduction of this activity. We also determined the intrauterine levels of E (17-
estradiol) by radioimmunoassay. A significant amount of E (12 pg per ml) was detected in the
extracts of decidual uteri on day 6. When the ovariectomized pregnant mice were treated with
letrozole, the E (17-estradiol) level was undetectable in the uterine extracts. Taken together,
these results confirmed that a functionally active aromatase enzyme is present in the decidual
uterus and it catalyzes the production of E within this tissue.
2.4.3. Uterine aromatase activity is critical for successful implantation
We next investigated whether the intrauterine E produced by aromatase is necessary for the
maintenance of pregnancy. Mice were ovariectomized on day 5 (6 AM) of pregnancy and
given daily injections of P in the presence or absence of letrozole. Uteri were collected on
days 8 and 10 of pregnancy and analyzed for the presence of implanted embryos.
Morphological and histological analyses of letrozole-treated and untreated uteri revealed that
administration of this drug severely impaired embryo implantation. Inhibition of aromatase
activity led to a significant reduction in decidual mass and the majority of the implanted
embryos failed to develop properly and were resorbed by day 10 (Fig. 2.3). This result
indicated that the aromatase activity and local E production is essential for endometrial
functions that support embryonic development during early pregnancy.
2.4.4. Inhibition of uterine aromatase activity blocks stromal differentiation
To determine whether the locally produced E controls endometrial functions independent of
embryonic development, we subjected mice to experimentally induced decidualization in
which a mechanical stimulation of the steroid-primed uteri triggers a decidual response in the
absence of the implanting embryo (10). This artificial stimulus mimics the embryonic signal
during implantation and sets in motion the decidualization program.
Following this uterine stimulation, the mice were treated with P alone or P plus letrozole for
three consecutive days (Fig. 2.4A). Uterine response was assessed at 72 h following the
decidual stimulation. As shown in Fig. 2.4B, the uterine horns of the P-treated animals

41
exhibited a robust decidual response (left panel). In contrast, decidualization was severely
compromised in mice that received P plus letrozole (right panel). Treatment with letrozole
strongly inhibited the uterine wet weight gain, a classical hallmark of uterine decidual
response (Fig. 2.4C).
We further assessed the impact of the loss of aromatase activity on decidual response by
monitoring the uterine expression of alkaline phosphatase and decidual prolactin-related
protein (PRP, 11), well-established biochemical markers of uterine stromal differentiation, in
the presence or absence of letrozole. We also examined the expression of two additional
factors: bone morphogenetic protein 2 (BMP2), a morphogen, and connexin 43 (Cx43), a gap
junction protein, which are induced in stromal cells during decidualization and are known to
play critical regulatory roles during this process (10, 12, 13). We found that the expression of
mRNAs encoding alkaline phosphatase, PRP, BMP2, and Cx43 was markedly reduced in the
letrozole-treated uteri (Fig. 2.4D). Consistent with these results, we observed a drastic
reduction in the intensity of Cx43 immunostaining in the uterine sections of letrozole-treated
mice (Fig. 2.4E). In contrast, the expression of ER mRNA was not significantly altered in
response to the inhibitor. Taken together, these results indicated that aromatase-driven
intrauterine E synthesis plays an important regulatory role in stromal cell differentiation.
2.5. DISCUSSION
In cycling rodents, the circulating E and P are produced and secreted by the concerted actions
of ovarian granulosa and theca cells (78). The adrenal glands in rodents are incapable of
synthesizing significant levels of these steroid hormones (79). In pregnant mice, the corpora
lutea develop fully by day 3 of pregnancy and start to produce P, the level of which rises and
remains elevated until mid-gestation (80). In contrast, the level of serum E, which is high on
day 1 of pregnancy due to the preovulatory hormonal surge, declines and reaches ~15 pg/ml
on days 2 and 3. The serum E level rises transiently to ~22 pg/ml on day 4 of pregnancy and
plays a critical role in embryo attachment. Once the embryo attaches to the uterus, the
ovarian E synthesis declines. During the decidualization phase, which lasts from days 5
through 8 of gestation, the circulating level of E remains low at ~15 pg/ml (80). When the

42
ovaries were removed after embryo attachment, administration of exogenous P was found to
be sufficient to sustain decidualization in pregnant mice (Fig. 2.1). The ovarian E, therefore,
has no evident role in regulating uterine functions following the embryo attachment step. The
present study reveals that the mouse uterus is able to carry out de novo synthesis of E during
decidualization and it is this locally produced hormone, not the ovarian E, which critically
supports the stromal differentiation process.
It was previously documented that during early pregnancy mouse endometrial stromal cells
acquire the ability to express the steroidogenic enzymes required for the synthesis of P
starting from cholesterol (81, 82). These factors are StAR, P450scc, and 3 HSD. The
maximal activity of 3 HSD, which catalyzes the conversion of pregnenolone to progesterone,
was detected in decidual tissue on days 6.5-7.5 of pregnancy (83, 84). Although these earlier
reports provided evidence for the potential de novo production of P in the decidua, the
physiological significance of this locally produced hormone remained unclear in the face of
high serum levels of P originating from the corpora lutea during early phases of pregnancy.
The expression of 17-HSD in the decidual tissue was also described previously (21). Our
present study extends these earlier observations to demonstrate that the decidua expresses
additional steroidogenic enzymes P450C17 and P450 aromatase. Therefore, a full complement
of steroidogenic enzymes is expressed in the decidual tissue, which allows conversion of P to
the androgenic precursors and their eventual aromatization to E.
The P450 aromatase, encoded by the CYP19 gene, is the key enzyme that catalyzes the
conversion of C19 steroids to E (22, 23). Previous studies have shown that E is synthesized in
a number of extragonadal sites such as breast, brain, and bone (85). This extragonadal E acts
locally within the tissue in a paracrine or intracrine fashion. Although only a small amount of
E is synthesized at these extragonadal sites, it is possible to attain high local concentrations of
the hormone, which then exerts important biological effects within the tissue. It is noteworthy
that aromatase expression in the pregnant uterus is initiated in the decidua on day 5 of
gestation, immediately following implantation, and continues to increase as the stromal
differentiation program advances during days 6 and 7. Our study is the first report
documenting the expression of a functionally active aromatase in the maternal decidua. Since
the ovarian E secretion drops to very low levels during decidualization, the local production

43
of this hormone in the decidual tissue at the implantation sites assumes high physiological
significance.
Previous studies using the aromatase-null mice provided important insights into the role of
E in various reproductive tissues. As expected, the homozygous mutant females were infertile
(86). Histology of the reproductive tract of these mice demonstrated evidences of follicular
depletion and the presence of hemorrhagic follicles in the ovaries, and diminution of uterine
weight. Supplementation with E rescued the development of ovarian follicles and allowed the
recovery of uterine weight, but did not ameliorate the reproductive failure in the mutant
females (86, 87). Interestingly, transplantation of wild-type ovaries into aromatase-null
female mice, which produces a circulating hormonal profile similar to that in wild-type mice,
resulted in only a poor rescue of the pregnancy outcome (27). These results hinted at
additional reproductive abnormalities in the mutant females, presumably at the level of the
uterus, and are consistent with our current findings demonstrating a critical functional role of
local E produced by uterine aromatase during implantation.
Our study revealed that the aromatase-driven intrauterine E plays an important role in
decidualization. The expression of alkaline phosphatase and PRP, two well-characterized
biomarkers of decidual response, was compromised when ovariectomized pregnant mice
were treated with letrozole (8, 11). Furthermore, the expression of BMP2 and Cx43, critical
regulators of stromal differentiation, were severely reduced in the presence of letrozole,
indicating that decidualization is impaired when this inhibitor blocks aromatase activity.
Previous studies reported that conditional ablation of BMP2 expression in the mouse uterus
leads to infertility due to lack of decidualization (16). Recently we have shown that
conditional loss of expression of Cx43, a major gap junction component in uterine stromal
cells, impairs decidualization and angiogenesis during early pregnancy (88). The uterine
expression of Cx43 is regulated by E (32, 89). It is, therefore, conceivable that the local E
produced by the aromatase in the decidua controls Cx43 expression, which in turn contributes
to the progression of decidualization by promoting gap junction communication between the
stromal cells.
Another major finding of this study is that de novo production of E plays a novel and
central role in the regulation of uterine neovascularization during early pregnancy. Although

44
a functional link between steroid hormone action and uterine angiogenesis in rodents has long
been speculated, the precise nature of this regulation remained unclear. The detailed
molecular candidates which are involved in modulating this decidual angiogenesis have been
discussed in Chapter 4 of the dissertation.
Our study, therefore, uncovers important pathways regulated by local E signaling to
regulate endometrial stromal differentiation and promote the establishment of new vascular
structures within the decidual tissue, thereby critically sustaining decidualization during early
pregnancy.

45
2.6. FIGURES
Figure 2.1: Ovarian E is not necessary for decidualization
Figure 2.1: Administration of exogenous P sustains decidualization in ovariectomized pregnant mice. A: Experimental scheme. Pregnant mice were ovariectomized on day 5 (D5) morning following the attachment of the embryo to the uterine epithelium on day 4 mid night. Mice were then treated with P (40 mg/kg bodyweight) for three days and uteri were collected on day 8 (D8). B: Gross morphology indicating the implantation sites in normal D8 uterus (left panel) and ovariectomized P-treated pregnant uterus (right panel). C: Sections of ovariectomized P-treated D8 and normal D8 uteri were subjected to immunohistochemical analysis using antibodies specific for progesterone receptor (PR, upper panel) and prolactin like protein type B (PLP-B, lower panel). am, m, and D denote antimesometrial, mesometrial, and decidua, respectively.
Figure 2.1A
Figure 2.1C
Figure 2.1BNormal: D8 Ovex-P: D8Normal: D8 Ovex-P: D8
D5
Ovex
P P P D8D4
Attachment
E+P
D5
Ovex
P P P D8D4
Attachment
D5
Ovex
P P P D8D4
Attachment
E+P
Normal D8 Ovex D5+P
PLP
PR
a b
c d
Normal D8 Ovex D5+P
PLP
PR
a b
c d
Figure 2.1C
Figure 2.1A Figure 2.1B

46
Figure 2.2: P450 aromatase is induced in the decidual uterus
Figure 2.2: Evidence for local E biosynthesis in decidual uterus. A: Schematic representation of the Estradiol biosynthetic machinery. B: Total uterine RNA obtained on day 4 (D4), day 6 (D6) and day 7 (D7) of pregnancy were analyzed by RT-PCR for the expression of StAR, P450SCC, P450C17, 3 HSD, 17-HSD-1, and P450 aromatase. C. RNA from d 2 to d 10 of pregnancy was analyzed by Northern blotting with cDNA probes specific for P450 aromatase (Upper) and 36B4 (Lower).
CHOLESTEROL
Pregnenolone
P450SCC
P450C17
17-OH-Pregnenolone
P450C17
Dehydroepiandrosterone
3-HSD
Androstenedione
17-HSD
Testosterone
Estradiol
P450arom
CHOLESTEROL
Pregnenolone
P450SCC
P450C17
17-OH-Pregnenolone
P450C17
Dehydroepiandrosterone
3-HSD
Androstenedione
17-HSD
Testosterone
Estradiol
P450arom
D4 D6 D7
StAR
P450SCC
P450C17
3-HSD
P450arom
36 B4
17-HSD
D4 D6 D7
StAR
P450SCC
P450C17
3-HSD
P450arom
36 B4
17-HSD
Figure 2.2A Figure 2.2B
Figure 2.2CFigure 2.2C
Figure 2.2A Figure 2.2B

47
Figure 2.2 (cont.)
Figure 2.2: Evidence for local E biosynthesis in decidual uterus (cont). D: Top panel: Sections of uteri collected on days 4, 5, and 6, are shown before and after laser capture micro dissection (LCM). Bottom panel: Quantitation of the results of real-time PCR using gene-specific primers for aromatase and alkaline phosphatase. AM, M, E corresponds to antimesometrium, mesometrium and embryo respectively. (n=3, p<0.01)
Day5 EEam
Day6
EE
am
Laser captured tissue
Day4
Day5 EEam
Day6
EE
am
Laser captured tissue
Day4
Figure 2.2D
**

48
Figure 2.2 (cont.)
Figure 2.2: Evidence for local E biosynthesis in decidual uterus.
E. Immunohistochemical analysis using the P450C17 antibody (Santa Cruz Biotech); sections of testes from male mice and uteri obtained from day 6 pregnant mice (panels a and b, respectively). F. Immunohistochemical analysis using the P450 aromatase antibody (Abcam); sections of ovaries collected 48 h after PMSG treatment and 16 h after hCG treatment (panels c and d, respectively), sections of uteri collected on day 4 (panel e), day 5 (panels f and g), and day 6 (panels h and i) of pregnancy. Panel j shows aromatase immunostaining in decidual cells when primary stromal cells isolated from uteri of day 4 pregnant mice were subjected to in vitro decidualization for 72 h. E, denote embryo.
E
E
Figure 2.2E
a b c
E
E
E
IHC: Aromatase
a b c
d e f
Figure 2.2F
24hr 24hr
48hr 48hr
Right panelLeft panel
Figure 2.2E
Figure 2.2F

49
Figure 2.3: Blockade of aromatase function leads to loss of pregnancy.
Figure 2.3: Blockade of aromatase function leads to loss of pregnancy.
Pregnant mice (day 5 morning) were ovariectomized and treated with exogenous P in combination with or without letrozole, an aromatase inhibitor (AI). The uteri were collected on day 10 of gestation (D10). The sections of D10 uteri treated with P only (Panel Day10-P) or P and AI (Panel D10-P+AI) were analyzed with eosin and hematoxylin stain. E, AM and M denote embryo, antimesometrial and mesometrial, respectively.
Figure 2.3
E+P
+AI
D5
Ovex
P P PD10D4
Attachment
E+P
+AI
D5
Ovex
P P PD10D4
Attachment
E
E
Day10-P
Day10-P+AI
E
E
Day10-PE
E
Day10-P
Day10-P+AI
Figure 2.3

50
Figure 2.4: Inhibition of aromatase activity impairs uterine decidualization.
Figure 2.4: Inhibition of aromatase activity impairs uterine decidualization.
A: The hormonal regimen used in the artificial decidualization protocol is shown. Mice were treated with or without letrozole (20 mg/kg body weight). Uteri were collected 72 h after the application of stimulus. B: The extent of decidual response in ovariectomized mice treated with P (left panel) and P plus letrozole (P+AI, right panel) is shown. “s” and “us” denote stimulated and unstimulated uterine horns, respectively. C: The quantitative analysis of the average weight gain of stimulated relative to unstimulated horns in mice (n=5) subjected to artificial decidualization with or without letrozole treatment. The data are represented as meansSEM.
+AI (letrozol)
OvexP P P PPPE E E
stimulus
+AI (letrozol)
OvexP P P PPPE E E
+AI (letrozol)
OvexP P P PPPE E E
stimulus
S
PUS
P+AIUS
SS
PUS
P+AIUS
S
0
50
100
150
Ute
rin
e W
eigh
t G
ain
(S
/US
)
-ArI+ArI
Figure 2.4A
Figure 2.4B
Figure 2.4CFigure 2.4C
Figure 2.4A
Figure 2.4B
*
p<0.05 n=5

51
Figure 2.4 (cont.)
Figure 2.4: Inhibition of aromatase activity impairs uterine decidualization (cont.) D: Uterine RNA was isolated 72 h after the initiation of decidualization and subjected to quantitative PCR analysis using gene-specific primers for ER, alkaline phosphatase (Alk), BMP2, connexin 43 (Cx43) and prolactin-related protein (PRP). P and P+AI represent uterine RNA from ovariectomized mice treated with P and P plus letrozole, respectively. E: Immunolocalization of Cx-43 protein in control (panels a and b) and letrozole treated (panels c and d) decidual uterus.
Figure 2.4D
ER Alkp BMP2 Cx43 PRP
Figure 2.4E
a b
c d
-AI
+AI
Figure 2.4D
Figure 2.4E
*
p<0.01 n=5
*
*

52
Table 2.1: Measurement of aromatase activity in the uterus during early pregnancy. The tissue homogenates were incubated with [1-3H] androstenedione for 6 h at 37 C to estimate the water release per mg of tissue. The intrauterine levels of estrogen were analyzed by radioimmunoassay.
Tissue fmol [H3] water Estradiol released/mg net wt/24 h (pg/ml
uterine extract)------------------------------------------------------------------------------------------------
Uterus (Day 6) 243.2 12.123.22
Uterus (Day 6+AI) 8.71.5 Undetectable
Ovary 455.4 Not analyzed
Table ITable 2.1

53
Chapter-3
Fos-related antigen 1 (FRA-1) is an estrogen regulated transcription factor
playing a critical role in mediating uterine decidualization

54
3.1. ABSTRACT
During early pregnancy, the concerted actions of steroid hormones estrogen (E) and
progesterone (P) via their cognate receptors orchestrate changes that make the uterine
epithelium competent to blastocyst attachment and initiate implantation. Following this event,
uterine stromal cells undergo steroid hormone-dependent transformation into
morphologically and functionally distinct decidual cells in a unique process known as
decidualization. An angiogenic network is also formed in the uterine stromal bed, critically
supporting early development of the embryo. Our recent studies in the mouse have revealed
that, following embryo attachment, de novo synthesis of E by the uterus drives stromal cell
differentiation and neovascularization during the decidual phase of pregnancy. To address the
mechanism of local E action in mediating this physiological process, we performed gene
expression profiling and identified Fos-related antigen 1 (FRA-1), a member of the Fos
family of transcription factors, as a downstream target of intrauterine E signaling. Further
analysis using chromatin immunoprecipitation (ChIP) indicated direct recruitment of E
receptor alpha (ER) to the Fra-1 promoter. Our study also revealed that signaling by
ERK/MAPK pathway activates FRA-1 protein in differentiating endometrial stromal cells.
To investigate the role of FRA-1 in decidualization, we attenuated Fra-1 expression by
administering small interfering RNA (siRNA) in primary cultures of endometrial stromal
cells. Our study demonstrated that signaling by ER regulates Fra-1 expression in the
decidual uterus which plays a critical role in stromal differentiation and migration during
early pregnancy.

55
3.2. INTRODUCTION
Implantation, a critical event in early pregnancy, is initiated by attachment of the blastocyst to
the wall of the uterine lumen. Subsequently, as the embryo invades through the luminal
epithelium, the underlying stroma undergoes dramatic remodeling to form the specialized
implantation chamber. Fibroblastic stromal cells proliferate and then differentiate to form the
secretory decidual cells that support embryo growth until placentation ensues (2, 73, 74, 76).
In addition, uterine endothelial cells proliferate forming an extensive vascular network in the
decidual tissue to nourish the embryo and sustain its development (90, 91). This
morphological and functional transformation of the pregnant uterus is regulated by concerted
action of the steroid hormones estrogen (E) and progesterone (P) and their cognate nuclear
receptors E receptor (ER) and P receptor (PR), respectively (2, 73, 92).
Ovarian E, along with P, regulates epithelial receptivity and embryo attachment during the
pre-implantation period. Following embryo attachment, the role of progesterone (P) as a
critical regulator of decidualization is well established but the effect of ovarian estrogen (E)
during this process is not clear. Our recent studies, however, have shown that the decidual
uterus expresses P450 aromatase, a key enzyme converting androgens to E, and is a novel site
of E biosynthesis (8). This locally produced intrauterine source of E is able to support
advancement of stromal differentiation even in the absence of ovarian E in an
ovariectomized, P-supplemented pregnant mouse model. Administration of letrozole (a
specific aromatase inhibitor) to these mice blocked stromal differentiation and impaired
development of the angiogenic network during decidualization (8).
To further understand the mechanism of E action and to identify the downstream targets of
intrauterine E during decidualization, we performed gene expression profiling of uterine
tissue in the presence or absence of letrozole (8). Our study revealed Fos-related antigen 1
(Fra-1) as one of the genes whose expression was significantly down-regulated in the
letrozole-treated decidual uteri, indicating Fra-1 as a potential target of intrauterine E
regulation during decidualization. FRA-1 belongs to the FOS family of transcription factors
that includes c-FOS, FOS B, and FRA-2 as its other members. FRA-1 exhibits maximum
homology to c-FOS, a known target of E/ER signaling in various cell types (93, 94).
Activation of FOS proteins is mediated by mitogen-activated protein kinase (MAPK)

56
pathways. Phosphorylation of FRA-1 initiates its activation and heterodimerization by the
highly conserved leucine zipper domains with the JUN proteins to constitute activator
protein-1 (AP-1). Previously regarded as the negative regulator of AP-1 activity, the FRA-1
comprising AP-1 complex was subsequently found to have important transforming activities
in many tissues, governing cell proliferation, differentiation, and remodeling (95-97).
In this study we address the regulation and functional significance of FRA-1 during
endometrial stromal cell differentiation. We provide strong evidence that locally produced
intrauterine E signals via ER to regulate Fra-1expression, which is then activated by the
ERK/MAPK pathway in differentiating uterine stromal cells. To elucidate the function of
FRA-1 in decidualization, we utilized a primary culture system in which undifferentiated
stromal cells isolated from pregnant mouse uterus undergo decidualization. Using this in vitro
system, we provide evidence that FRA-1 plays a critical role in stromal differentiation and
cell migration during early pregnancy. Additionally, we demonstrated that VEGFa an
important regulator of angiogenesis is down regulated by FRA-1 indicating this transcription
factor might contribute to the development of the decidual vascularization.
3.3. EXPERIMENTAL PROCEDURES
Reagents-
Progesterone and 17--estradiol were purchased from Sigma Chemical Co. (St. Louis, MO).
Antibodies against FRA-1, p-ERK1/2, ERK1/2, ERand Calnexin were obtained from Santa
Cruz Biotechnology Inc., (Santa Cruz, CA).
Animals and Tissue Collection-
All experiments involving animals were approved by the Animal Care Committee at the
University of Illinois at Urbana-Champaign and the studies were conducted in accordance
with the National Institutes of Health standards for the use and care of animals. Female mice
(CD-1 from Charles River, Wilmington, MA), in proestrus, were mated with adult males. The
presence of a vaginal plug after mating was designated as day 1 of pregnancy. The animals

57
were killed at various stages of gestation and the uteri were collected after removal of the
embryos by repeated flushing.
Artificial Decidualization-
Decidualization was experimentally induced in hormone primed mice as described previously
(98). Briefly, mice were first ovariectomized and two weeks following ovariectomy, animals
were injected with 100 ng of E in 0.1 ml of sesame oil for three consecutive days. This was
followed by daily injections of 1 mg of P for three consecutive days. Decidualization was
then initiated in one horn by injection of 50 l oil. The other horn was left unstimulated. The
animals were treated with P for additional 3 days post stimulation and then killed to collect
the uterine tissue.
Mouse primary stromal cell culture-
Isolation of mouse primary stromal cells was performed as discussed before (99). Briefly, day
4 pregnant mouse uterus in the pre-implantation phase was subjected to enzymatic digestion
using pancreatin and dispase enzymes for 1 hr at room temperature. The reaction was
neutralized with 10% fetal bovine serum (FBS) and the supernatant containing the epithelial
cell clumps were discarded. The partially digested tissue was then incubated with collagenase
at 37C for 45 mins. The reaction was again neutralized with 10% FBS and vortexed to
dispense stromal cells in the suspension. The resultant solution was passed through a 80m
sieve to obtain a pure stromal cell population. Stromal cells obtained were cultured in
complete medium containing 2% serum, and differentiation was induced by adding P.
Immunohistochemistry-
Polyclonal antibodies against FRA-1 (1:500 dilution) was used for immunohistochemistry as
discussed before (100). Briefly, paraffin-embedded uterine tissues were sectioned at 4 µm
and mounted on slides. Sections were washed in PBS for 20 min and then incubated in a
blocking solution containing 10% normal goat serum for 30 min before incubation in primary
antibody overnight at 40C. Immunostaining was performed using avidin-biotin kit for rabbit
primary antibody (Vector Laboratories, Burlingame, CA). Sections were counterstained with
hematoxylin, mounted, and examined under bright field. Red deposits indicate the sites of
positive immunostaining.

58
Western Blotting-
Whole cell extracts were prepared from mouse primary stromal cell cultures undergoing in
vitro differentiation. Briefly, cells were washed with ice-cold balanced solution and lysed
with the RIPA buffer (0.1% SDS, 0.5% Na-deoxycholate, 1% NP-40 in PBS) containing
protease inhibitor cocktail, phenylmethylsulfonylfluoride (PMSF 0.1 mg/ml), and
phosphatase inhibitor (1:1000; Sigma-Aldrich). This was followed by passing the cells
through a 25 gauze syringe and centrifugation at 12000 rpm for 10 mins to remove the cell
debri. Typically 20-50g of the protein extract was analyzed by SDS-PAGE and transferred
to polyvinylidene fluride membrane (Amersham Biosciences Inc., Piscataway, NJ). The
membrane was blocked with 5% milk in Tris buffered saline with 0.1% Tween 20 for 1 hr at
room temperature, followed by incubation with a primary antibody against FRA-1, p-Erk1/2,
Erk1/2, ER or calnexin. The blot was then incubated with HRP-conjugate secondary
antibody for 45 mins at room temperature. The HRP was detected by chemiluminescence.
Alkaline Phosphatase Activity Assay-
For detecting alkaline phosphatase activity, cultured stromal cells were fixed in 4%
formaldehyde for 10 mins and were incubated in the dark at 37 C for 60 m in 2 mM a-
Napthyl phosphate and 4 mM Fast violet in 0.1 M Tris HCl, pH 8.7. In the presence of
substrate a-Napthyl phosphate, the uterine alkaline phosphatase activity releases
orthophosphate and napthol derivatives from the substrate. The napthol derivatives are
simultaneously coupled with the diazonium salt present in the incubating medium to form a
dark dye marking the site of enzyme action. The slides were rinsed in water to terminate the
enzymatic reaction and visualized under a microscope.
siRNA transfection-
siRNA corresponding to mouse FRA-1 (5'-3' sense GGAAGGAACUGACCGACUUtt, and
antisense AAGUCGGUCAGUUCCUUCCtc), ER(5’-3’ sense
GGGAGAAUGUUGAAGGCACAtt, and antisense UGUGCUUCAACAUUCUCCCtc) and
a scrambled negative control were obtained from Ambion (Ambion Inc., Austin, TX) and
transfected using the siPORT NeoFX transfection reagent according to the manufacturer’s
instruction. Briefly, the annealed oligonucleotide was complexed with the transfecting
reagent in Opti MEM I reduced serum medium. The complex containing 20nM of siRNA in

59
5l of the reagent was dispersed into culture plates while the primary cells were attaching.
The cells were again subjected to a second silencing hit after 36 hrs with the Silent Fect
reagent (Bio-Rad Inc., Hercules, CA) and cultured for an additional 24 to 48 hrs before
analyzing gene expression.
Chip Assays-
Primary stromal cells isolated from day 4 pregnant mouse uterus were cultured for 24 hrs in
the presence of steroid hormones and ChIP was performed using a commercially available kit
from Upstate Biotech according to the manufacturer’s instruction (Millipore Inc, Billerica,
MA). Briefly, 1.5x10^7 to 2x10^7 primary stromal cells were isolated from day 4 pregnant
mouse uterus and cultured in 15cm plates in the presence of P. After 24hrs, the chromatin
was crosslinked by adding 1% formaldehyde to the tissue culture medium for 10 mins at
room temperature followed by adding 1M of glycine for 5 mins. The crosslinked chromatin
was sonicated using a sonic Dismembrator Model 100 (Fisher Scientific) at power 4 and 5
pulses (each for 12 seconds) to shear the DNA and obtain fragments from 200bp-1kb. The
sonicated chromatin was diluted in ChIP dilution buffer and pre-cleared using protein G
agarose. The input sample is 1% of the pre-cleared supernatant. Immunoprecipitation was
performed over night at 4C with 5g of ER antibody (Santa Cruz Biotechnology Inc., Santa
Cruz, CA) and 1g of each RNAPolII and the non specific anti mouse IgG antibodies. The
immune complexes were further precipitated using protein-agarose beads and washed several
times using different buffers, followed by elution of the protein/DNA complexes. Reverse
crosslinking was then performed at 65C for 5 hrs and the DNA was purified using spin
columns. Quantitative and regular PCR was performed on these purified DNA samples using
the specific primers.
Gel shift assay-
Primary stromal cells were treated with DMSO or the ERK inhibitor for 2 hrs and subjected
to EMSA following the manufacturer’s instruction (Thermo Scientific Inc., Rockford, IL).
Briefly, nuclear extract was obtained using the NE-PRE nuclear and cytoplasmic extraction
reagent kit from Pierce (Thermo Scientific Inc., Rockford, IL). EMSA was performed using
equal amount (2-3g) of the nuclear extract in 20l of binding buffer (containing 50%
glycerol, 1% NP40, 1M Kcl, 100mM Mgcl2 and 200mM EDTA) on ice for 10 mins before
adding the biotinylated double stranded AP-1 probe (5’-CGCTTGATGAGTCAGCCGGAA-

60
3’). After incubating for an additional 20 mins at room temperature, the samples were
resolved on a 4% polyacrylamide gel. After the gel separation, the samples were transferred
to a Nylon membrane (Ambion Inc., Austin TX) and crosslinked at 120 mJ/cm2 using a
commercial UV-light cross-linker instrument. The biotin labeled DNA was then detected by
chemiluminescence. To demonstrate the specificity of the DNA:protein complex, the nuclear
extract was incubated for 10 mins with a 200 fold molar excess of unlabeled oligo before
adding the labeled probes. To further demonstrate the presence of a specific protein in the
complex, the nuclear extracts were mixed with 2g of anti-FRA-1 antibody and incubated for
20mins on ice prior to adding the labeled probe.
Wound healing assay –
Primary stromal cells were cultured in the presence of steroid hormones to induce
differentiation. FRA-1 expression was silenced in these cells using siRNA transfection. The
confluent monolayer of mouse stromal cells were uniformly scratched with a 10l sterile
pipette tip, the cell debris washed and the remaining cells were further cultured in fresh
medium for an additional 48hrs. Wound width was monitored by phase contrast microscopy
at regular intervals. The extent of migration was analyzed by calculating the wound closure
and expressed as a percentage of the initial wound.
3.4. RESULTS
3.4.1. Intrauterine E regulates FRA-1 expression during decidualization
Our previous studies have shown that the mouse uterus is able to carry out de novo synthesis
of E during decidualization, and it is this locally produced hormone that critically supports
stromal differentiation (8). To identify potential downstream targets of intrauterine E that
contribute to decidualization, we examined changes in uterine mRNA expression profiles in
response to letrozole, which inhibits aromatase activity and thereby blocks estrogen
synthesis. Uteri were collected from control or letrozole-treated mice 72 h following
administration of the decidual stimulus. Total RNA isolated from these tissues was subjected
to microarray analysis using Affymetrix murine GeneChip arrays. We identified several
mRNAs whose expression was altered significantly in the decidual uterus in response to
letrozole. One of the mRNAs whose level was markedly down-regulated encoded FRA-1, a

61
member of the FOS family of transcription factors that regulates a wide variety of biological
processes such as cell proliferation, differentiation, and migration (101).
We verified the results of the microarray analysis by performing q-PCR analysis of RNA
obtained from uteri of control and letrozole-treated mice. While significant down-regulation
of Fra-1 mRNA was observed in the absence of intrauterine E, expression of ER remained
unaltered in either treatment (Fig. 3.1A upper panel). Regulation by endogenous E was
further confirmed by monitoring the expression of FRA-1 protein in uterine sections of mice
treated with or without the inhibitor (Fig. 3.1A lower panel). A high nuclear accumulation of
FRA-1 protein surrounding the site of decidual stimulation was observed in the control
uterine sections (left panel). However, uterine expression of FRA-1 was significantly down-
regulated after letrozole treatment (right panel). We also confirmed intrauterine estrogenic
regulation of FRA-1 by employing a well-established in vitro decidualization system. We had
previously shown that primary cultures of stromal cells isolated from pregnant uteri (pre-
implantation, day 4) and subjected to P-induced decidualization exhibit strong aromatase
expression; inhibition of this aromatase activity by letrozole severely impairs decidualization
(8). As shown in Fig. 3.1B, administration of letrozole to primary cultures of stromal cells
impaired FRA-1 expression in a dose-dependent manner, indicating that FRA-1 is a novel
target of intrauterine E action during early pregnancy.
3.4.2. Induction of FRA-1 during decidual phase of pregnancy
We next studied the spatial expression of FRA-1 in pregnant mouse uterus using
immunohistochemistry (Fig 3.2A, panel a-f). The nuclear expression of FRA-1 was induced
in the stromal cells surrounding the embryo on day 4 midnight (panel a). As decidualization
progressed, FRA-1 expression became prominent at the antimesometrial end on day 5 of
pregnancy (panel b). The expression intensified in the decidual cells surrounding the embryo
on day 6 (panel c and d) and gradually progressed from the antimesometrial end to the
mesometrial region on day 7 (panel e and f). The nuclear FRA-1 expression was finally
restricted in the mesometrial decidua by day 8 (Fig S1). We further monitored the expression
of FRA-1 during experimentally induced decidualization in mouse uterus (Fig 3.2B, panel a-
c). Consistent with the expression profile during pregnancy, nuclear FRA-1 was initially
observed surrounding the lumen 24hrs after stimulus (panel a) and the expression expanded
to the surrounding layers by 64 and 72 hrs (panel b and c). These observations indicated that

62
the precise pattern of FRA-1 expression in the decidual cells is not induced by the embryo
derived signals but a possible prerequisite for the uterine transformation during
decidualization.
3.4.3. E/ ER signaling regulates FRA-1 expression in differentiating stromal cells
To further investigate steroid hormone regulation of FRA-1 in the uterus, we subjected mice
to ovariectomy and treated them with E (100 ng) or P (2 mg) or vehicle for three days.
Uterine stromal cells isolated from these animals were subjected to q-PCR analysis. As
shown in Fig. 3.3A, the expression of P receptor (PR), a well-characterized E target, was
significantly induced after E treatment, while ER expression remained unchanged in
response to hormone treatments. E administration also led to marked induction of Fra-1,
while P had no effect on Fra-1 expression. Immunohistochemical analysis of FRA-1 in the
uterine sections from mice treated with steroid hormones revealed that E treatment led to
strong nuclear localization of FRA-1 in the stromal compartment as well as in the epithelial
cells (Fig. 3.3B, middle panel). In contrast, vehicle- or P-treated uterine sections exhibited
markedly reduced, as well as cytoplasmic, expression of FRA-1 in stroma and luminal
epithelium (left and right panels, respectively).
Using an in vitro decidualization system, we next investigated whether regulation of Fra-1
by E in endometrial stromal cells is mediated via ER. We first monitored expression of Fra-
1 in stromal cell cultures from 24 h to 72 h, and found marked induction of Fra-1 by 24 h in
culture (Fig. 3.4A, upper panel). Fra-1 expression was also observed at 48 h to 96 h, although
at a reduced level. Immunolocalization studies indicated that nuclear expression of FRA-1
protein was induced as early as 5 h in culture and then intensified by 24 h to 48 h of culture
(Fig. 3.4A, lower panel). Consistent with the early induction of FRA-1, expression of ER
mRNA and protein was induced significantly in endometrial stromal cells by 24 h of culture
and declined by 72 h (Fig. 3.4B).
Using this in vitro primary cell culture model we wanted to address E mediated FRA-1
regulation during decidualization. E action in the uterus is primarily mediated through the
predominant intercellular nuclear receptor ER, thus we first wanted to examine if the E
induced FRA-1 expression is regulated by the stromal ERs. A well characterized ER
antagonist, ICI 182 780 was administered to primary mouse stromal cells undergoing in vitro

63
decidualization. 48 hrs after ICI treatment, the cells were harvested and the isolated RNA was
subjected to real time PCR analysis. We observed a significant down regulation of the Fra-1
transcript after treating cells with the ER antagonist (Fig 3.4C). We next investigated the role
of ER in regulating Fra-1 expression by employing an RNAi-mediated gene knockdown
strategy. Cultured endometrial stromal cells were treated with control scrambled siRNA or
ER-specific siRNA for 48 h. We observed that cells transfected with ER siRNA exhibited
more than 80% reduction in ER mRNA expression compared to cells transfected with
scrambled siRNA (Fig. 3.4D, top panel). Down-regulation of ER also led to more than 60%
reduction in Fra-1 mRNA expression in ER siRNA-treated cells compared to cells treated
with scrambled siRNA. The expression of GAPDH remained unaltered in either treatment
condition (Fig. 3.4D, top panel). Consistent with the mRNA expression profile, the protein
levels of ER and FRA-1 were significantly down-regulated in the ER siRNA-treated cells
(Fig. 3.4D, bottom panel). Taken together, these results indicate that E signaling via ER
regulates expression of FRA-1 in differentiating uterine stromal cells.
To examine the possibility that FRA-1 is a direct target of ER action during mouse
stromal cell decidualization, we first analyzed the region from -2000 to +200 bp in the Fra-1
gene promoter using a bioinformatic tool (Transcription Element Search System
http://www.cbil.upenn.edu/cgi-bin/tess/tess) and observed six potential ER binding sites. Fig.
3.5A shows the location of these sites. We then performed a ChIP experiment to identify the
predominant ER binding site in the Fra-1 promoter. Mouse primary stromal cells were
fixed after 24 h of culture and the sheared chromatin was immunoprecipitated with antibodies
against ER, RNA Pol II, and IgG. The precipitated and reverse cross-linked DNA was
purified and subjected to RT-PCR followed by agarose gel analysis using six different primer
sets, each flanking a potential ER binding site in the Fra-1 promoter. The pre-cleared
supernatant served as the input control for each antibody. RNA Pol II and IgG were used as
positive and negative controls for the ChIP experiment, respectively. As shown in Fig. 3.5B,
amplicon 3, which encompassed an E response element (ERE) at about -1400 bp, exhibited a
robust signal, indicating a potential ER binding site in the Fra-1 promoter (Fig. 3.5B upper
panel). Q-PCR analysis from a number of independent experiments further confirmed this
site for predominant ER occupancy in the Fra-1 promoter (lower panel).
3.4.4. ERK-MAPK pathway dependent activation of FRA-1 in differentiating stromal cells

64
FRA-1 is known to be activated by the ERK1/2-regulated MAPK pathways in many cell
types (102). To investigate whether MAPK pathways are involved in regulating FRA-1
activity during endometrial stromal cell differentiation, we first monitored activation status of
the ERK/MAPK pathway in mouse stromal cells. Protein extracts were prepared at regular
intervals from cultured stromal cells undergoing in vitro differentiation and subjected to
Western blot analysis using antibodies against the phosphorylated and total forms of ERK
protein. As shown in Fig. 3.6A, the level of total ERK remained unaltered for the duration of
culture. However, a marked elevation in the level of phospho-ERK1/2 was observed 12 h
after initiation of culture and was sustained until 96 h. Consistent with the phospho-ERK
status, we observed a similar expression profile of FRA-1, raising the possibility that FRA-1
is activated by the ERK1/2 pathway during in vitro endometrial stromal differentiation.
To investigate whether FRA-1 activation is linked to the ERK pathway, we performed a
gel shift assay to investigate whether the ERK pathway affects DNA binding activity of FRA-
1 (Fig. 3.6B). Nuclear extracts isolated from mouse primary stromal cells treated or untreated
with the ERK inhibitor, UO126, were incubated with the biotinylated oligonucleotide-
containing FRA-1 binding site, the AP-1 probe. The protein-DNA complexes were analyzed
by gel electrophoresis. In the presence of nuclear extract and the probe an intense signal was
detectable, indicating formation of a protein-DNA complex (lanes 2-7), while no signal was
detected in the absence of nuclear extract (lane 1). Complex formation was significantly
reduced in response to a 200-fold molar excess of unlabeled AP-1 probe (compare lanes 2, 3
to 4, 5) or after addition of anti-FRA-1 antibody (compare lane 2 to 6 and lane 3 to 7),
indicating that the protein-DNA complex formation is specific to the AP-1 probe and to
binding of FRA-1 protein to the AP-1 probe, respectively. Addition of ERK inhibitor,
UO126, further reduced the signal intensity, suggesting that binding activity of FRA-1 is
compromised in the presence of ERK inhibitor (compare lane 6 to 7). Together, these
experiments indicated that the ERK1/2 pathway is important in activating FRA-1 in
endometrial stromal cells undergoing decidualization.
3.4.5. Fra-1 is a critical mediator of stromal cell decidualization
We next addressed the role of FRA-1 in stromal differentiation by RNA interference-
mediated knockdown of its gene expression. Primary stromal cells were isolated from uteri of
day 4 pregnant mice as described previously and transfected with siRNA targeted specifically
to the Fra-1 mRNA. Control cells were transfected with a scrambled siRNA (NC). Cells

65
transfected with Fra-1 siRNA exhibited a remarkable decrease in Fra-1 mRNA expression
(Fig. 3.7A, upper panel) and FRA-1 immunostaining (Fig. 3.7A, lower panel), whereas
transfection with NC siRNA did not affect Fra-1 expression. We next investigated the
functional consequences of this blockade of Fra-1 expression during stromal differentiation.
The siRNA-mediated down-regulation of Fra-1 in stromal cells led to a significant decline in
the level of alkaline phosphatase (ALP) (Fig. 3.7B) and PRP (Fig. 3.7C), the well-
characterized markers of stromal decidualization. In contrast, the ER level remained
unaltered in cells treated with either Fra-1 or NC siRNA. These results indicated that Fra-1
expression is critical to successful progression of stromal differentiation during
decidualization. VEGFa, a known cytokine mediating the development of angiogenesis
during decidualization was also significantly repressed after FRA-1 attenuation (Fig 3.7C).
Additionally, impairment of FRA-1 activity by disrupting ERK phosphorylation through the
administration of UO126 also altered stromal cell decidualization in vitro (Fig 3.7D). The
defect was again addressed by examining enzymatic activity of Alkp (upper panel) and Prp
expression (lower panel) after inhibitor treatment in stromal cells.
3.4.6. Fra-1 regulates stromal cell remodeling through extracellular matrix proteins
Invasion of an embryo into the uterine stroma necessitates extensive remodeling of the
differentiated decidual cells to accommodate the growing embryo. Because FRA-1 is known
to regulate cell migration and motility in a variety of cell types, we next investigated the role
of FRA-1 in stromal cell remodeling, using an in vitro wound-healing assay. As shown in
Fig. 3.8A, stromal cells transfected with Fra-1 siRNA displayed a significant decrease in cell
motility compared to cells transfected with NC siRNA. Quantification of wound closure
demonstrated that cells treated with the NC siRNA exhibited more than 80% wound closure
at 24 h and almost complete closure by 48 h, while those treated with Fra-1 siRNA exhibited
less than 30% wound closure even after 48 h of culture (Fig. 3.8A and Fig. 3.8B).
To further clarify this impairment in wound closure in Fra-1-silenced cells, we assessed
proliferation of these cells. Stromal cells transfected with control NC or Fra-1 siRNA were
fixed at 48 h after the wound-healing assay and subjected to immunocytochemical analysis
using the anti-Ki67 antibody. Drawing from five independent fields of NC and Fra-1 siRNA
transfected cells, the cells that displayed positive Ki67 staining were then analyzed by paired
t-tests (Fig. 3.8C). We failed to observe any significant difference (P = 0.132) in cell

66
proliferation between the two siRNA-treated cell populations, indicating that the defect in
wound healing in Fra-1-silenced cells might instead be due to impaired cell migration.
To identify possible downstream mediators of Fra-1 involved in stromal cell migration and
remodeling, cells treated with NC- and Fra-1-specific siRNAs were subjected to a wound-
healing assay, followed by RNA extraction and q-PCR analysis to measure expression of
several matrix metalloproteinases (MMPs). We observed significant down-regulation in the
expression of MMP7, MMP9, and MMP13, while expression of MMP2 remained unaltered
(Fig. 3.8D). CD44, a glycoprotein molecule involved in cell migration and a well-
characterized target of FRA-1 in various cell types, was also significantly down-regulated in
cells transfected with Fra-1 siRNA compared to cells treated with NC siRNA (Fig. 3.8D).
Collectively these results indicated that FRA-1 plays a critical role in stromal cell migration
by influencing expression of extracellular matrix modulators, thereby controlling invasion of
the embryo during decidualization.
3.5. DISCUSSION
During early pregnancy in the mouse, E along with P plays a pivotal role in preparing the
uterus for embryo implantation. While ovarian E is critical during the pre-implantation
period, we recently reported that a local source of intrauterine E driven by the presence of
P450 aromatase in decidual uterus contributes significantly, after embryo attachment, to
sustaining stromal differentiation and uterine neovascularization (30). To identify
downstream targets of this local intrauterine E, we disrupted the functional E biosynthetic
pathway by administering letrozole, a potent aromatase inhibitor, and performed gene
expression profiling. Expression of the transcription factor Fra-1 in the decidual uterus was
significantly repressed in response to letrozole, indicating Fra-1 as a possible mediator of E
action. Our study also showed that during decidualization FRA-1 is localized primarily in the
nuclear compartment of differentiating stromal cells, which also express high levels of ER
(Fig. 3.2). Therefore, it is reasonable to speculate that, with the onset of differentiation, FRA-
1 is induced in response to E/ER signaling in decidual cells. In the classical signaling
pathway, ER regulates gene expression by binding to EREs in the upstream regulatory
regions of target genes. So far, PR is the only factor that has been identified as a direct target

67
of ER in endometrial stromal cells. We are now reporting that ER regulates Fra-1 in
uterine stroma by binding to the ERE located 1400 bp upstream in the Fra-1 promoter (Fig.
3.5B).
FRA-1 undergoes phosphorylation at the serine and threonine residues to achieve its
activation and sustained nuclear expression (103). ERK/MAPK regulates activation of FRA-1
by phosphorylation and also governs the stabilization of this nuclear protein by preventing its
proteasomal degradation (102, 104). In the uterus during implantation, induction of ERK
could result from a variety of stimuli such as growth factors, mitogens, or cytokines. E is
known to regulate EGF and IGF signaling in mouse uterus, which might govern the
activation status of ERK1/2 (22, 105, 106). Our study revealed that ERK signaling is
significantly induced while stromal cells differentiate, and thereby activates FRA-1 and
sustains its nuclear expression during decidualization (Figs 3.6A). Activated FRA-1
dimerizes with members of the JUN family of proteins to constitute the AP-1 transcription
factor complex, and binds to AP-1 consensus sites in the target gene promoter. Gel shift
analysis showed binding activity of FRA-1 and specific complex formation during stromal
cell differentiation. Because ERK regulates activation of FRA-1, disruption of the ERK
pathway inhibited the DNA binding activity of FRA-1 and severely impacted complex
formation (Fig. 3.6B). It is, however, possible that inhibition of the ERK pathway may
additionally alter the activation status of the JUN partner, resulting in decreased DNA
binding and complex formation.
FRA-1 is important in regulating cell proliferation, differentiation, and migration in
various cell types. Previous studies indicated that mice lacking c-FOS develop osteoporosis
due to a defect in osteoclast lineage. This defect in differentiation could be rescued by over-
expression of FRA-1, suggesting overlapping functions of c-FOS and FRA-1 in osteoclast
differentiation (107). Additionally, accelerated differentiation of osteoprogenitors to form
mature osteoblasts occurs in response to ectopic expression of FRA-1 (108). Consistent with
these observations, studies employing a conditional knockout model of Fra-1 clearly
established the critical role of FRA-1 in osteoblast differentiation (109). While expression of
c-FOS and JUN proteins in the uterus had been previously reported, the expression and
function of FRA-1 in stromal differentiation was not investigated until now (110, 111). In this
study we observed robust induction of FRA-1 in differentiating endometrial stromal cells

68
(Fig. 3.7A). Attenuation of FRA-1 expression in these cells severely influenced the
expression of decidualization biomarkers, indicating a critical role of Fra-1 in stromal cell
differentiation (Fig. 3.7B).
Decidualization is accompanied by an extensive uterine neo-vascularization (35, 112).
Abundant VEGF expression in the differentiating stromal cells critically regulates the
angiogenic growth, reduction of which results in a severe impairment of the vascular
structure development (33). FRA-1 is known to be involved in angiogenesis during tumor
progression by regulating VEGF expression (113). In our studies we observed that FRA-1
also governs VEGF expression in endometrial differentiating stromal cells. Additionally,
during decidualization, the gradual shift in FRA-1 expression from the antimesometrium to
the mesometrial decidua further supports the previous observation, indicating a probable role
of the transcription factor in the development of uterine angiogenesis (Fig. 3.9). We speculate
that FRA-1 in the decidual cells at the mesometrial pole regulate the VEGF production which
acts on the surrounding endothelial population to modulate the uterine angiogenic
development. Further, these studies also indicated FRA-1 as one of the mediators of
intrauterine E action in regulating the decidual vasculature.
Our study also revealed the critical role of Fra-1 in stromal cell migration during
decidualization. During decidualization, invasion of the embryo through the maternal
endometrium requires extensive uterine remodeling, particularly near the site of embryo
attachment. This remodeling involves reorganization of the differentiating stromal cells to
accommodate the growing embryo. A recent study in a co-culture of human blastocyst on a
monolayer of endometrial stromal cells used time-lapse microscopic images to capture the
migration of stromal cells away from the site of the embryo (114). It is likely that such
remodeling is also operational during implantation in the mouse; however, the regulators of
stromal cell migration during decidualization remain poorly understood. This study used the
wound-healing assay, a well-established experimental read-out of cell migration and
remodeling, to assess the role of Fra-1 in cellular migration. Quantification of wound closure
in stromal cells transfected with Fra-1 siRNA revealed significant impairment in cell
mobility compared to the control siRNA-treated stromal cells (Fig. 3.8). Furthermore, our
studies showed that the migration defect of FRA-1-deficient stromal cells is due to improper
expression of factors that critically regulate cellular remodeling and motility.

69
Previous studies in various cell types have shown that MMPs and the tissue inhibitors of
metalloproteinases (TIMP) play active roles in cell migration and remodeling (37, 38).
Interestingly, it has been reported that administration of hydroxamic acid, an inhibitor of
metalloproteinases, to pregnant mice severely impacts the progress of decidualization
indicating a critical role of MMPs in maintaining the implantation chamber (115). CD44, a
principal hyaluronic receptor with known functions in cell adhesion, trafficking, and
migration, is another strong candidate for regulating endometrial remodeling, and prominent
expression of CD44 has been reported in decidual cells surrounding the embryo (116).
Previous studies have also reported that FRA-1 governs the expression of MMP9, MMP13,
and CD44 in various cell types and in pathological situations such as mesothelioma (117-
119). In this study, we found that expressions of several MMPs and CD44 are regulated by
FRA-1 in differentiating stromal cells. Thus, FRA-1 governs stromal cell migration during
decidualization by regulating the expression of these molecules.
In summary, our study indicates that intrauterine E, functioning via ER, regulates the
expression of transcription factor FRA-1 in the decidual uterus. FRA-1 is then activated by
ERK/MAPK signaling pathways. The activated FRA-1 plays a pivotal role during early
pregnancy by regulating gene expression to promote stromal cell differentiation and
migration during decidualization.

70
3.6. FIGURES
Figure 3.1: Intrauterine Estrogen signaling regulates FRA-1 expression during stromal cell decidualization.
Figure 3.1: Intrauterine Estrogen signaling regulates FRA-1 expression during stromal cell decidualization. Mice were ovariectomized and subjected to artificially induced decidualization as described in experimental procedures. Cohorts of these animals were treated with P alone (P) or P along with aromatase inhibitor (P+AI) and the uterine horns were separately collected after 72hrs of stimulus (n=5). A: Stimulated uterine samples were subjected to RNA isolation and quantitative PCR (q-PCR) analysis using Fra-1 and ER specific primers (upper panel) and to immunohistochemical analysis using the anti-FRA-1 antibody (lower panel). Lu indicates the lumen of the uterus. (n=3, p<0.05)
Figure 3.1A
qPCR, 72hr ADR
P P+AI
IHC: Fra-1, 72hr ADR
Fra1ER
Lu
Lu
Figure 3.1A
*

71
Figure 3.1(cont.)
Figure 3.1: Intrauterine Estrogen signaling regulates FRA-1 expression during stromal cell decidualization. B: Stromal cells isolated from day 4 of pregnancy were subjected to in vitro decidualization in the presence of increasing concentration of AI for 72hrs. The RNA was analyzed with q-PCR to monitor the expression of Fra-1 transcript. (n=3, p<0.05)
Figure 3.1BFigure 3.1B
*

72
Figure 3.2: FRA-1 expression is induced in mouse stromal cells as decidualization progresses.
Figure 3.2: FRA-1 expression is induced in mouse stromal cells as decidualization progresses. A: Uterine cross sections were obtained from mice on different days of pregnancies (a-f) and immunohistochemical analysis was performed against FRA-1 on day 4 midnight (a), day 5 (b), day 6 (c, d), day 7 (e, f). Higher magnification of days 6 and 7 are shown in d and f respectively. B: Sections from mice subjected to experimentally induced decidualization (a-c) were obtained and FRA-1 localization at 24hr (a), 64 hrs (b) and 72hrs (c) after application of decidual stimulus was monitored.
a c e
b d f
Figure 3.2A
b
Figure 3.2B
ca
Figure 3.2A
Figure 3.2B

73
Figure 3.3: FRA-1 is a downstream target of E in mouse uterine stromal cells
Figure 3.3: FRA-1 is a downstream target of E in mouse uterine stromal cells Non-pregnant animals were ovariectomized and treated with vehicle (oil), E or P (n=5). Stromal cells were isolated 24 hrs after the last treatment. A: RNA extracted from isolated stromal cells was subjected to q-PCR analysis using specific primers for ER, PR and Fra-1. B: Immunohistochemical analysis for FRA-1 in the uterine cross sections of mice treated with vehicle, E and P. (n=8, p<0.05)
Figure 3.3A
Fra-1 PR ER-qPCR
Vehicle E PFigure 3.3B IHC: FRA-1
Figure 3.3A
Figure 3.3B
**

74
Figure 3.4: Fra-1 is directly regulated by E/ERa signaling during decidualization
Figure 3.4: Fra-1 is directly regulated by E/ERa signaling during decidualization Primary stromal cells isolated from day4 pre-implantation mouse uterus were subjected to in vitro differentiation in the presence of P. A and B: Cells were isolated at regular intervals to analyze the mRNA level of Fra-1(Fig 3.4A) and ER (Fig 3.4B) expression by q-PCR (upper panel) and protein by immunocytochemistry (ICC) (lower panel).(n=7, p<0.05)
0
5
10
15
20
25
30
35
0hr 24hr 48hr 72hr 96hr
Rel
ativ
e le
vel
of
exp
ress
ion
0hr 24hr 48hr 72hr 96hr
q-PCR
a
c
b
d
Figure 3.4A
FRA-1 immunocytochemistry
a
c
b
d
Figure 3.4A
FRA-1 immunocytochemistry
Figure 3.4A
*

75
Figure 3.4 (cont.)
0hr 24hr 48hr 72hr
24hr
48hr
72hr
Figure 3.4B
a b
c d
e f
ER alpha immunocytochemistry
Figure 3.4B
* *

76
Figure 3.4 (cont.)
Figure 3.4: Fra-1 is directly regulated by E/ERa signaling during decidualization C: Stromal cells isolated from the pre-implantation uterus were cultured for 48hrs in the presence of vehicle (V) or P plus antiestrogen ICI-182 780 (ICI). RNA was extracted and the transcript level of Fra-1 was analyzed by q-PCR. D and E: The endogenous expression of ER was silenced by transfecting a siRNA specific to its transcript in mouse primary stromal cells. After 48hrs of transfection, RNA was subjected to q-PCR analysis using gene specific primers for ER, Fra-1 and GAPDH. The relative levels of gene expression in the ER siRNA treated samples compared to the negative control (NC) siRNA treated samples are shown. (average ± SD) (D). Whole cell protein extract obtained from ER siRNA and NC siRNA treated cells were subjected to Western blot (WB) analysis against FRA-1, ER and the control protein calnexin (E). (p<0.01, n=3)
0
0.2
0.4
0.6
0.8
1
1.2
ER-a Fra-1 GAPDH
Re
lati
ve
lev
el o
f E
xp
res
sio
n
NC-siRNA
ERa siRNA
Figure 3.4C
0
0.2
0.4
0.6
0.8
1
1.2
EP PI
Re
lati
ve
lev
el o
f E
xp
res
sio
n
ER Fra-1 GAPDH
Figure 3.4D
Western Blot
ER
Fra1
Calnexin
NC siRNA ER siRNA
Western Blot
ER
Fra1
Calnexin
NC siRNA ER siRNA
Figure 3.4E
V ICI
Figure 3.4C
Figure 3.4D
Figure 3.4E
*
*
*

77
Figure 3.5: Recruitment of ER to Fra-1 promoter
Figure 3.5: Recruitment of ER to the Fra-1 promoter A: Schematic representation of the potential E response elements from -2000bp to +200bp of Fra-1 gene promoter (accession no: AF017128). The candidate binding sites comprising of half EREs are located at -1829bp (ERE 1), -1677bp (ERE 2), -1408bp (ERE 3), -1114bp (ERE 4), -994 bp (ERE 5), -158bp (ERE 6) and the corresponding PCR amplicons are amplicon 1 (-1964 to -1729), amplicon 2 (-1694 to -1505), amplicon 3 (-1523 to -1373), amplicon 4 (-1204 to -1050), amplicon 5 (-1071 to -931), amplicon 6 (-275 to -118) flanking each potential binding site. B: Mouse primary stromal cells were cultured in presence of P for 24 hrs, formaldehyde fixed and subjected to ChIP protocol. Immunoprecipitation (IP) was performed using the ER antibody. IP with RNA PolII and IgG antibodies were used as the positive and negative control respectively. RT-PCR using 32 cycles was performed to amplify the purified DNA. The input lane represents 1 % of each soluble chromatin. IP lane represented the amplified PCR product for each amplicon (upper panel). The immunoprecipitated promoter-DNA fragments were reverse crosslinked and amplified by q-PCR using a pair of primers for each amplicon and the fold change was calculated (lower panel). The data shown is a representative of 3 independent experiments.
-2000bp +2bp
+200bpChip Primers
Amplicon: 1 2 3 4 5 6
ER
E1
ER
E 2
ER
E 3
ER
E 4
ER
E 5
ER
E 6
Fig. 3.5A
Input
IP
RNAPolymerase
IgGER ERE 1 -1829 bp
ERE 2 -1677 bpERE 3 -1408 bpERE 4 -1114 bpERE 5 -994 bpERE 1 -158 bp
IgG RPII 1 2 3 4 5 6
IgG Rna Pol 1 2 3 4 5 6
Figure 3.5B
IgG Rna Pol 1 2 3 4 5 6
Figure 3.5B
Figure 3.5A
Figure 3.5B

78
Figure 3.6: ERK-MAPK pathway dependent regulation of FRA-1 activation in differentiating stromal cells.
Figure 3.6: ERK-MAPK pathway dependent regulation of FRA-1 activation in differentiating stromal cells. A: Equal amounts of whole cell protein extract isolated from cultured stromal cells were analyzed by Western blotting (WB) with antibodies directed to phosphorylated form of ERK1/2 (pERK1/2), total ERK1/2 and FRA-1. Calnexin was used as a loading control.
0hr 12hr 24hr 48hr 72hr 96hr
p-ERK1/2
ERK1/2
Fra-1
Calnexin
0hr 12hr 24hr 48hr 72hr 96hr
p-ERK1/2
ERK1/2
Fra-1
Calnexin
Figure 3.6AFigure 3.6A

79
Figure 3.6 (cont.)
Figure 3.6: ERK-MAPK pathway dependent regulation of FRA-1 activation in differentiating stromal cells. B: Equal amount of the nuclear extract treated with vehicle DMSO (lanes 2, 5 and 6) and the inhibitor UO126 (lanes 3, 4, and 7) were subjected to gel shift assay to analyze the protein binding to the biotinylated labeled probe containing AP-1 elements. The representative image illustrates the formation of a protein-DNA complex to the biotinylated free probe. Lane 1 serves as a control showing no complex formation in absence of the nuclear extract. Presence of 200-fold molar excess of unlabeled AP-1 probes competed out the biotinylated probe binding indicating that the complex formation is specific to AP-1 (lanes 4 and 5). Incubation with anti-FRA-1 specific antibody (lanes 6 and 7) reduced the extent of complex formation indicating FRA-1 complexes with the nuclear extract and in presence of the antibody, it reduced the extent of protein-DNA complex formation (compare lane 2 with lane 6). Addition of ERK inhibitor UO126 significantly reduced the complex formation indicating binding activity of FRA-1 is compromised in the presence of ERK inhibitor (compare lane 6 to lane 7).
Fig 3.6B
Protein-DNA complex
Free probe
Gel-shift assay
Figure 3.6B

80
Figure 3.7: FRA-1 is a critical regulator of mouse stromal cell decidualization
Figure 3.7: FRA-1 is a critical regulator of mouse stromal cell decidualization
Primary mouse stromal cells transfected with FRA-1 and NC specific siRNA, were collected 48hrs after transfection. A: Down regulation of Fra-1 mRNA (upper panel) and protein (lower panel) was assessed by qPCR, ICC and WB respectively. B: Alkaline phosphatase (Alkp) activity in NC (a) and FRA-1(b) siRNA treated cultured cells. C: Expression of decidualization markers like Alkp, Prp and VEGF. D: Mouse cultured stromal cells were treated with vehicle or ERK1/2 inhibitor UO126 for 48hrs and assessed for enzymatic activity of Alkp (upper panel) and transcript levels of the decidualization markers Alkp and Prp (lower panel). (n=3, p<0.05)
Neg siRNA-30nM FRA-1 siRNA-30nM
0
0.2
0.4
0.6
0.8
1
1.2
Neg siRNA-30nM Fra-1siRNA 30nM
Rel
ativ
e le
vel
of
exp
ress
ion
RNA profile
IHC-Fra-1
Neg siRNA-30nM FRA-1 siRNA-30nM
NC 20 Fos 10 Fos20
Fra-1
Calnexin
Western Blot
Immunocytochemistry
Figure 3.7A
NC siRNA Fra1 siRNA
Figure 3.7B
Figure 3.7C
Fra-1 Alkp PRP VEGF a ER
Figure 3.7A
Figure 3.7B
Figure 3.7C
** * *
*

81
Figure 3.7 (cont.)
Figure 3.7: FRA-1 is a critical regulator of mouse stromal cell decidualization D: Inhibition of FRA-1 activity further impaired stromal cell differentiation as indicated by the decreased enzymatic activity of Alkp (left panel) and expression of decidual biomarkers (right panel). (n=3, p<0.05)
Fig 3.7D
Vehicle ERK inhibitor UO126
Alkp PRP ERa
Figure 3.7D
*
*

82
Figure 3.8: FRA-1 modulates uterine stromal cell mobility through extracellular matrix proteins
Figure 3.8: FRA-1 modulates uterine stromal cell mobility through extracellular matrix proteins. Primary mouse stromal cells were transfected with FRA-1 specific siRNA and NC siRNA or incubated with a transfection reagent serving as a mock control. siRNA transfected confluent monolayers of primary stromal cells were subjected to wound healing assay to monitor the cell migration. A: Cells were photographically recorded under phase contrast microscopy after 0hr, 24hr and 48hr of the wound. B: Quantitative representation of stromal cell migration after scratching from three independent experiments are shown *p<0.01, **p<0.001.
0hr 24hr 48hr
NC siRNA
Fra-1 siRNA
Time points 0hr 24hr 48hr
NC siRNA
Fra-1 siRNA
Time points
Wound healing assayFigure 3.8A
*
P<0.01
Figure 3.8B
Figure 3.8A
Figure 3.8B

83
Figure 3.8 (cont.)
Figure 3.8: FRA-1 modulates uterine stromal cell mobility through extracellular matrix proteins C: Cells were fixed after 48hrs of the migration assay and subjected to immunocytochemical analysis using anti-Ki67 antibody. Left panel: Red dots indicate cells positive for Ki67 and undergoing active proliferation after transfection with control (panels a and b) and FRA-1 siRNA (panels c and d). Right panel: Photomicrographic representation of the staining quantitatively expressing the Ki67 positive cells in NC and FRA-1 siRNA treated cells. D: RNA isolated was subjected to q-PCR to assess the relative levels of the gene expression of MMP2, MMP7, MMP9, MMP13 and CD44 compared to the mock control (average ± SEM). (n=3, p<0.05).
Figure 3.8D
NC siRNA
FRA-1 siRNA
Ki67 staining after wound healing assay
0
0.05
0.1
0.15
0.2
0.25
0.3
NC siRNA FRA-1 siRNAP
os
itiv
e c
ell
po
pu
lati
on
NC siRNA FRA-1 siRNA
Figure 3.8CKi67 immunocytochemistry
a b
c d
Figure 3.8C
Figure 3.8D
*
* *
*

84
Figure 3.9: Expression of FRA-1 in the mesometrium on Day 8 FRA-1 expression on day 8 uterus is restricted to the mesometrial end surrounding the blood vessels as shown in the right panel (arrow pointing the blood vessels)
S1: Expression of FRA-1 in the mesometrium on Day 8
EM
M
Figure 3.9: Expression of FRA-1 in the mesometrium on Day 8
Figure 3.9

85
Chapter-4
Intra-uterine estrogen signaling critically regulates decidual angiogenesis

86
4.1. ABSTRACT
During implantation, the development of new blood vessel plays an important role
contributing to embryo growth and sustaining early pregnancy. Endothelial cells, the major
cell type in these blood vessels and capillary networks are the primary target of several cues
in the uterine milieu that modulate active vessel formation. Fluorescence assisted cell sorting
indicated around 20% of the cells in an isolated deciduoma is endothelial in nature. Although
the regulation of these cells by the steroid hormones estrogen (E) and progesterone (P) has
been addressed in previous studies, the underlying mechanism directing uterine neo-
angiogenesis is not well characterized. In a recent study we identified the local biosynthesis
of E in uterine decidual cells, and its pivotal role in sustaining implantation. Microarray
analysis after disrupting this E biosynthetic machinery revealed significant population of
molecules involved in regulating angiogenesis, indicating a significant contribution of this
steroid in the formation of decidual vasculature. Here, we have addressed the possible
mechanisms through which local E govern angiogenesis. The regulation primarily involved
the expression of paracrine factors from decidualized stromal cells that act on the endothelial
population governing its proliferation and maturation. In addition, we also monitored a subset
of players that directly regulate endothelial cell growth, down regulated in the absence of E
signaling. Collectively, this study addressed novel E regulated maternal signaling pathways
that are critical in governing uterine vascularization and angiogenesis during early pregnancy.

87
4.2. INTRODUCTION
Differentiation of the fibroblast stromal cells to the specialized cell typically characterizes the
deciduous transformation of the uterine endometrium in early pregnancy. The development of
blood vessels during this process is considered as a major hallmark event, significantly
contributing to the formation of an implantation chamber (73, 74). In mouse, the initial
vasculature is identified early on day 4 evening demarcating the implantation sites in uterine
horns. Following embryo attachment, with progression of decidualization, a dramatic
vascular remodeling prepares the uterine endometrium to support embryo growth. Formation
of this vasculature and capillary network becomes predominant by day 7 of pregnancy at the
mesometrial pole, also demarcating the future site of placentation (2, 76, 120).
Development of extensive blood vessels in a relatively a-vascular structure sustains growth
of a developing tissue. In normal physiological condition such spurious growth of vasculature
or angiogenesis is characteristic in the adult female reproductive cycles during cyclical
remodeling of the growing endometrium and during corpus luteum formation in ovary (3,
121). Vascular network develops by the migration of endothelial cells from pre-existing
blood vessel structures and their subsequent morphogenesis at a distant site. The process
requires coordinated execution of a series of cellular events beginning with degradation of the
endothelial basement membrane and extracellular matrix surrounding the parent vessel.
Endothelial cells then migrate from the origin and proliferate along the chemo-attractant
gradient towards an angiogenic signal, resulting in the formation of nascent capillary sprouts
(17).
Steroid hormone signaling critically orchestrates the overall implantation process. In
addition to its contribution in regulating stromal cell differentiation, estrogen (E) and
progesterone (P) also play major roles in the vascular development and angiogenesis. While
studies have indicated that the hormonal regulation of blood vessel formation in non-pregnant
ovariectomized mice is regulated by P (31), during pregnancy, prior to the embryo
attachment, ovarian E is the known enhancer of the vascular permeability at sites of
blastocyst attachment (35, 122). Additionally, previous observations in our laboratory have
identified the major contribution of a E regulated gap junction molecule, Connexin-43 (Cx-
43), in governing uterine angiogenesis (33). Conditional ablation of Cx-43 protein in the
decidual uterus resulted in a severe impairment of vasculature resulting in embryo resorption

88
and subsequent infertility. During decidualization, the expression of Cx-43 is regulated by the
intrauterine E source, indicating a plausible mediator of steroid regulation during decidual
angiogenesis (30).
In this chapter, we identified the contribution of the local E in regulating uterine vascular
growth during decidualization. We observed that this regulatory event was mediated either
through the differentiated stromal cells, by the release of paracrine mediators that modulate
vascular growth or by a direct governance of the endothelial population by local E. The study
addresses some of the key observations that establish the significant contribution of E
signaling in modulating decidual angiogenesis.
4.3. EXPERIMENTAL PROCEDURES
Reagents-
Progesterone and 17--estradiol were purchased from Sigma Chemical Co. (St. Louis, MO).
Animals and Tissue Collection-
All experiments involving animals were approved by the Animal Care Committee at the
University of Illinois at Urbana-Champaign and the studies were conducted in accordance
with the National Institutes of Health standards for the use and care of animals. Female mice
(CD-1 from Charles River, Wilmington, MA), in proestrus, were mated with adult males. The
presence of a vaginal plug after mating was designated as day 1 of pregnancy. The animals
were killed at various stages of gestation and the uteri were collected after removal of the
embryos by repeated flushing.
Artificial Decidualization-
Decidualization was experimentally induced in hormone primed mice as described previously
(98). Briefly, mice were first ovariectomized and two weeks following ovariectomy, animals
were injected with 100 ng of E in 0.1 ml of sesame oil for three consecutive days. This was
followed by daily injections of 1 mg of P for three consecutive days. Decidualization was
then initiated in one horn by injection of 50 l oil. The other horn was left unstimulated. The

89
animals were treated with P for additional 3 days post stimulation and then killed to collect
the uterine tissue.
Mouse primary stromal cell culture-
Isolation of mouse primary stromal cells was performed as discussed before (99). Briefly, day
4 pregnant mouse uterus in the pre-implantation phase was subjected to enzymatic digestion
using pancreatin and dispase enzymes for 1 hr at room temperature. The reaction was
neutralized with 10% fetal bovine serum (FBS) and the supernatant containing the epithelial
cell clumps were discarded. The partially digested tissue was then incubated with collagenase
at 37C for 45 mins. The reaction was again neutralized with 10% FBS and vortexed to
dispense stromal cells in the suspension. The resultant solution was passed through a 80m
sieve to obtain a pure stromal cell population. Stromal cells obtained were cultured in
complete medium containing 2% serum, and differentiation was induced by adding P.
Immunofluorescence-
Polyclonal antibodies against PECAM-1 (BD Biosciences) was used for
immunocytochemistry as discussed before (100). Briefly, cultured cells were fixed in 4%
paraformaldehyde solution and permeabilized with 0.25% Triton X-100. The culture dishes
were washed in PBS for 20 min and then incubated in a blocking solution containing 10%
normal goat serum for 30 min before incubation in primary antibody overnight at 40C.
Immunostaining was performed using a FITC conjugated secondary antibody against the
primary anti PECAM-1 antibody. Sections were counterstained with 4',6-diamidino-2-
phenylindole (DAPI), mounted, and examined under green fluorescence filter. Bright green
deposits indicate the sites of positive immunofluorescence.
Immunohistochemistry-
Paraffin-embedded uterine tissues were sectioned at 4 µm and mounted on slides for
immunostaining of Ki-67, Ang2 (Santa Cruz Biotech), Claudin 5 (Invitrogen Inc.) and frozen
sections for PECAM-1 (BD Biosciences Inc). Sections were washed in PBS for 20 min and
then incubated in a blocking solution containing 10% normal goat serum for 30 min before
incubation in primary antibody overnight at 40C. Immunostaining was performed using

90
Avidin-Biotin kit for rabbit primary antibody (Vector Laboratories, Burlingame, CA).
Sections were counterstained with hematoxylin, mounted, and examined under bright field.
Red deposits indicate the sites of positive immunostaining.
Endothelial cell isolation-
The isolation of endothelial cells was performed as described before with some modifications
(123). Briefly, the cells were obtained from day 8 uterus of pregnant CD-1 females by
mechanical separation, enzymatic digestion and fluorescence activated sorting. Individual
implantation sites were washed in ice cold hanks solution and the myometrium removed to
reveal the deciduoma. The embedded blastocyst was carefully removed under microscope
and the remaining spongy tissue was minced and subjected to enzymatic digestion in 10ml of
DMEM F-12 medium supplemented with 2% Type IA crude collagenase (Sigma), 1% heat
inactivated fetal bovine serum and 1% penicillin and streptomycin in for 1hr at 37C with
intermittent shaking. The reaction was neutralized with 10% FCS in balanced hanks medium,
and the tissue clumps were sheared by repeated pipeting; the undigested tissue was allowed to
settle by gravity and the supernatant was collected, this wash was repeated thrice to obtain the
heterogenous cell population. The supernatant was further digested in 0.25% trypsin EDTA
solution for 20 mins at 37C to obtain a single cell suspension. After digestion, the solution
was filtered by passing through a 80m sieve, washed thrice and the cells were cultured in
gelatin or matrigel coated plates. The culture medium was specific for endothelial cell growth
containing 2mM glutamine, 1% penicillin streptomycin, 1mM sodium pyruvate, 20mM
HEPES, 1% non-essential aminoacid, 50mM beta mercaptoethanol, 20% fetal calf serum,
150m/ml of Endothelial cell growth factor and 12U/ml heparin.
Flow cytometry-
Dissociated cells in single suspension were incubated in ice with rat anti-mouse PECAM-1
antibody for 90 mins, followed by a second incubation with FITC conjugated anti-rat IgG for
30 mins in ice. Cells were washed thrice with 0.1% sodium azide and 1% FBS in balanced
solution after each incubations and were analyzed and separated using FCS express software
(De Novo Software Inc). Establishment of the gates was based on the staining profiles of the
negative controls and cells labeled with propium iodide to eliminate low forward scatter
signal events, debris and apoptotic cells.

91
LCM RTPCR-
Laser assisted microdissection was performed as discussed before (30). Briefly, sections of
uteri collected on day 8 of pregnancy from mice treated with P or P+AI were subjected to
LCM. Frozen sections (10 m) containing the implantation sites were placed on polyethylene
naphthalate membrane slides (Molecular Devices) and dehydrated in ethanol and xylene.
Using the Veritas microdissection instrument, the mesometrial and anti-mesometrial regions
were excised and the decidual tissues were obtained. RNA was isolated from these tissues
using a Pico Pure RNA isolation kit according to the manufacturer’s instructions. cDNA was
synthesized from the isolated RNA using the Sensiscript reverse transcriptase kit (Qiagen)
and subjected to real-time PCR using gene-specific primers.
4.4. RESULTS
4.4.1. Endothelial cell population in decidual uterus
With progression of pregnancy, the decidual mass recruits several factors that govern uterine
neo-angiogenesis to nourish embryo growth and survival. As shown in the whole mount day
8 deciduoma (Fig 4.1A), the mesometrial region exhibits predominant blood vessel formation
and extensive capillary networks demarcating active site of vascular remodeling. To establish
a model for studying the hormonal regulation of endothelial cells during decidualization, we
isolated the uterine cells from day 8 pregnant decidua and cultured them in complete medium
supplemented by endothelial cell growth factors. 5 days after culture, the endothelial cells
clumped together forming the specific cobble stone morphology (Fig 4.1B). Immunostaining
of these cells with Platelet derived endothelial cell adhesion molecule-1 (PECAM-1), an
endothelial cell specific membrane protein also known as CD-31, exhibited positive cell
clusters at the immediate vicinity of the decidual cells (Fig 4.1C).
4.4.2. Isolation of endothelial cells from decidual uterus
In order to quantify the population of uterine endothelial cells during decidualization, we
separated the deciduoma from the myometrium and the embedded blastocyst. The spongy
tissue was then enzymatically digested as described in experimental procedures to yield a
single cell suspension of heterogenous uterine cells and subjected to flow cytometry analysis.
As shown in Fig 4.2A, the forward vs. side scatter plot indicated three major cell population

92
based on the differential size and granularity. Cells were stained with anti-CD31 antibody,
further tagged with a fluorescein dye, FITC and subjected to fluorescence assisted cell sorting
(FACS). Only the live cell population which was gated after staining with propium iodide
were sorted (Fig. 4.2B).
A histogram plotting the cell count on the y-axis to the intensity of the fluorescence on the
x axis represented the CD-31 positive and negative cell populations (Fig.4.3). The total cell
population (M2) and the cells which exhibit higher than the minimal threshold intensity for
FITC (M1) were defined. Before sorting majority of the population were FITC negative
(upper panel), while sorting yielded around 90% pure endothelial cells (lower panel). Further,
in the control sample stained with fluorophore tagged anti-rat IgG, the M1 population was
less than 1%, while after staining with anti-CD31 antibody, around 15% of the gated cells
were M1 positive, representing the endothelial cell population in the decidual uterus (Table
4.1). We performed 4 independent experiments and observed that endothelial cells comprised
around 15-20% of the total number of cells in a day 8 pregnant uterus.
4.4.3. Intrauterine E biosynthesis plays a critical role in decidual vascular growth
A recent study in our laboratory identified the biosynthesis of local E during early pregnancy
and its pivotal contribution in sustaining implantation. To identify the potential targets of E
action in mediating this process, we performed a microarray analysis with the uterine RNA
isolated from the decidual tissue of mice treated with or without the aromatase inhibitor,
letrozole. We identified more than 6000 mRNA transcripts including ESTs which were
altered at a fold change of 1.5 fold or more when compared to the vehicle treated control
decidual uterus. Of the 450 genes with complete coding sequences down regulated in the
absence of E signaling, a significant population of molecules (about 120 genes) was involved
in the development of angiogenesis. These putative E regulated genes were involved in
various aspects of angiogenesis and were broadly classified into factors that contribute to
endothelial cell proliferation, extracellular matrix remodeling, regulation of VEGF
expression, migration and maturation of endothelial cells. A quantitative analysis of the
number of genes involved in each biological function is represented in Fig 4.3A.
To validate the expression profile of the angiogenic markers in the absence of E signaling,
we subjected mice to artificially induced decidualization program and treated a subset of
them with aromatase inhibitor. Q-PCR analysis of the RNA obtained from uterine extracts of

93
the control and inhibitor treated mice confirmed the significant down regulation of several
angiogenic markers like hypoxia inducible factor (HIF2a), Claudin-5, angiopoeitin-2 and 4
(Ang-2 and Ang-4) and adrenomedulin (Adm) (Fig. 4.3B).
The regulation of E action on decidual vascular growth was again confirmed by monitoring
the expression of PECAM-1. As shown in Fig. 4.3C the control uterine horn exhibited intense
PECAM-1 expression, demarcating vascular network (panel a and c); treatment with
aromatase inhibitor resulted in a significant reduction in PECAM-1 expression and intensity,
indicating that the expansion of the uterine angiogenic network was impaired in the absence
of local E production (panel b and d).
Development of vascular network necessitates active proliferation of the endothelial
population in the mesometrial zone of the pregnant uterus. We next assessed the extent of this
cell proliferation in the presence of aromatase inhibitor by immunostaining for Ki67 (Fig.
4.3D). Uterine sections obtained from P-treated mice exhibited robust Ki67 immunostaining
in endothelial cells, consistent with microvascular proliferation (panel A), while those
obtained from aromatase inhibitor-treated animals showed greatly reduced Ki67 expression
(panel B).
4.4.4. Plausible mechanisms of E action in modulating uterine vascular development
Stromal cells isolated by enzymatic digestion from day 4 morning pre-implantation mouse
uterus, were cultured in vitro in the presence of steroids to induce decidualization. The
expression of the angiogenic markers were compared in the P treated control versus
aromatase inhibitor treated samples (Fig 4.4). A significant inhibition of HIF2a, Adm and
Ang 2 and 4 were observed along with the other differentiation markers like Alkp, Prp,
BMP2 and Cx-43 after disrupting the endogenous E biosynthesis, indicating that the release
of paracrine factors from the decidual cells is regulated by the local source of E. This
observation was further validated by monitoring the protein expression of some of these
molecules during normal pregnancy. Pregnant mice were ovariectomized on day 5 early
morning, 6-8 hrs following the embryo attachment. Pregnancy was sustained with an
exogenous administration of P. To a subset of these animals, aromatase inhibitor was
administered to disrupt the local E biosynthesis (Fig 2.1A). Uterine samples collected on day
8 of these mice were subjected to immunohistochemical analysis using antibodies against
Ang-2 a critical player in the development of angiogenic network. Ang-2 exhibited intense

94
staining in the mesometrial decidual cells and in the ectoplacental cone in the control uteri
(Fig 4.5A). Administration of letrozole resulted in the significant down regulation of Ang-2
expression in the mesometrium, while the expression in the ectoplacental cone was retained,
further confirming a significant contribution of endogenous E in regulating the expression of
these molecules and in coordinating endothelial cell growth and maturation.
From our microarray analysis, we identified certain endothelial cell specific molecules
which were down regulated in the presence of letrozole. One such molecule was Claudin 5,
an endothelial cell specific tight junction molecule with major contribution in cell to cell
communication. The expression of this molecule in the control uterus was restricted to the
endothelial cell membrane junctions at the mesometrial end of day 8 uterus (Fig 4.6, panels a
and b). Administration of the inhibitor drug significantly altered Claudin 5 expression,
indicating a severe impairment in endothelial cell to cell communication (panels c and d).
Collectively, these results indicated that aromatase-mediated local E in the uterus plays a
critical role in angiogenesis by governing the expression of stromal cell derived factors and
endothelial cell specific angiogenic modulators.
4.4.5. Expression of angiogenic markers is concentrated to the mesometrium
As we observed in Fig. 4.1A, the maximum concentration of uterine blood vasculature is
restricted to the mesometrial pole, it is thus plausible that the cytokines and other critical
modulators of uterine angiogenesis are primarily accumulated in that region. To confirm this
speculation we captured specific uterine zones by laser assisted microdissection (LCM) and
analyzed the gene expression. Frozen sections collected from d5 ovariectomized-d8 uterus
were processed in xylene and ethanol and subjected to laser capture. The region of intense
blood vasculature comprising the zone immediately outside the ectoplacental cone upto the
beginning of the deep stroma was compared to the decidual cells of the antimesometrium (Fig
4.7A). The angiogenic markers in the blood vessel concentrated mesometrium exhibited a
significant high expression compared to the antimesometrial decidual cells in the normal P
treated d8 uterus (Fig. 4.7B). As expected, treatment with aromatase inhibitor impaired the
gene expression in the mesometrium, confirming the pivotal role of endogenous E signaling
in regulating uterine vascular network (Fig.4.7C).
4.5. DISCUSSION

95
Decidual vasculature is an essential criterion for sustaining early pregnancy. Endothelial cells
near the mesometrium undergo extensive remodeling to form nascent capillary network
demarcating the future site of placentation. While the uterine angiogenesis has been
established in the field of implantation biology, the molecular mechanisms that underlie this
physiology remain largely uncategorized. Our previous study identified an intrauterine source
of E during decidualization. This endogenous steroid is critical in maintaining the
implantation chamber, absence of which results in embryo resorption (30). It is likely that this
dramatic consequence on pregnancy outcome is due to an overall impairment of the
decidualization process. While in the previous chapter the contribution of E in regulating
stromal cell differentiation was focused, the concentration of the present involves on
identifying the morphological features of isolated uterine endothelial cells and its regulation
by the local steroid. The major finding of this study is that de novo production of E plays a
novel and central role in the regulation of uterine neovascularization during early pregnancy.
Separation of the endothelial population from a heterogenous single cell suspension of the
uterine cell mass was achieved by FACS. It is a powerful tool that enables simultaneous
quantification and multiparametric categorization of different populations of cells by labeling
them with fluorescent conjugated antibodies specific to a particular cell type. Using this
method of cell separation we were able to sort a pure population of CD-31 positive
endothelial cells which comprised about 20% of the entire endometrial cellular mass during
decidualization. FACS sorted endothelial cells when grown in culture did form the long
cylindrical tube like processes, but owing to the low cell number that survived after the
sorting process, we were unable to monitor the characteristic cobble stone morphology in
these isolated cells, later addressed in co-culture experiments. Decidual cell plays a major
role in morphogenesis and formation of uterine vasculature. Accordingly, in the stromal-
endothelial co-culture experiments, 5 days of culture promoted the aggregation of endothelial
cells in small clumps and development of their characteristic pavement stone like
morphology (Fig 4.1B). The experimental method of endothelial cell isolation and separation
during decidualization can be used as a powerful tool to study the steroid regulation
specifically to these cell types in future studies.
Steroid hormone regulation is the polar event that organizes endothelial networking during
pregnancy. Although a functional link between the hormone action and uterine angiogenesis

96
in rodents has long been speculated, the precise nature of this regulation remained unclear.
Establishment and remodeling of blood vessels during angiogenesis requires the coordinated
execution of a series of cellular processes. Endothelial cells migrate from the parent vessel
and proliferate in response to one or more paracrine signal(s), resulting in the formation of
nascent capillary sprouts (124). Treatment of decidual uterus with aromatase inhibitor
significantly altered expression of several molecules involved in endothelial cell maturation
and morphogenesis, broadly categorized into factors that promote endothelial cell
proliferation, molecules involved in endothelial-extracellular matrix remodeling, important
players that regulate the expression of VEGF and the associated angiopoeitin-Tie2 signaling
(125).
During angiogenesis, the tip cell that directs the migration of a large population of endothelial
cells towards an environmental cytokine has often been compared to an axonal growth cones
that discerns the direction of axon outgrowth (124). Some of these molecules that regulate the
migration event include semaphorins, ephexin, ninjurin, netrin, slit and robo proteins and
other molecules involved in the Notch and Ephrin signaling (125-129). Interestingly, most of
these factors were found to be down regulated in the absence of intrauterine E signaling
(Table 4.2). This is a critical observation of steroid regulation on the endothelial population,
indicated for the first time that local E signaling governs the directionality of blood vessel
formation during decidualization.
As stromal cells differentiate, they secrete increasing amount of cytokines, transcriptional
regulators of angiogenic factors and other mitogenic agents that act via paracrine signaling on
endothelial cells to regulate angiogenesis. We identified that the decidual E critically
mediated differentiation of stromal cell population which further governed the release of
these paracrine factors, regarded as the indirect mechanism of E action on vascular
development. The alternative mechanism would be a direct regulation of endothelial cells by
E action. We hypothesized that a balanced interplay of these two pathways are responsible in
governing the angiogenic development of the decidual uterus.
Consistent with this speculation, we have identified a number of stromal factors that are
induced in response to aromatase-derived E and are likely regulators of neovascularization in
the decidua. Our study showed that the intrauterine E controls the stromal expression of

97
HIF2, a transcription factor that regulates VEGF production (38). VEGF is a potent mitogen
for endothelial cells, and is a prime regulator of angiogenesis during implantation and
decidualization (35, 37). In many tissues, VEGF acts in concert with angiopoietins to regulate
angiogenesis (37, 130). While angiopoietin 2 collaborates with VEGF to advance the
invasion by the vascular sprouts and promote vascular remodeling, angiopoetin 4 serves an
important function during endothelial cell migration (131). We present evidence that
aromatase-driven E controls the expression of both angiopoetin 2 and angiopoetin 4 in uterine
stromal cells. The stromal expression of adrenomedullin, a factor involved in angiogenesis
and important regulator of uterine function during implantation (40), is also controlled by the
E generated in the uterus. Another critical regulator of angiogenesis regulated by local E is
the gap junction molecule Cx-43. Previous studies in our laboratory have shown that Cx-43
expression in the pregnant uterus is restricted to the decidual population and it significantly
contributes to neo-vascularization, by modulating the paracrine release of angiogenic factors
(132).
Endothelial cell to cell communication is critical in mediating the vascular network and
coordinate endothelial cell migration and growth (133). This communication is primarily
mediated by a group of cell adhesion molecules forming various adhesion complexes. We
found that treatment with aromatase inhibitor severely impaired expression of several
endothelial cell adhesion molecules, indicating a direct regulation of intrauterine E on this
population. Among the other prominent players we identified Claudin-5, vasculo-endothelial
cadherins and several independent cell adhesion molecules like PECAM, CeaCAM and
ICAM, important in controlling endothelial cell communication (134-136). It is possible that
this direct regulation of intrauterine E is mediated by the expression of ER in mouse uterine
endothelial cells. Studies in our laboratory are currently underway exploring this direct aspect
of hormonal regulation in endothelial cells.

98
4.6. FIGURES
Figure 4.1: Extensive endothelial cell population in decidual uterus
Figure 4.1: Extensive endothelial cell population in decidual uterus The implantation sites were isolated from a day 8 pregnant CD-1 mouse and the myometrium was removed to reveal the deciduoma. A: Whole mount picture of an isolated day 8 deciduoma. The figure shows extensive blood vasculature in the mesometrial end compared to the antimesometrium. B: Co-culture of endothelial and stromal cells in vitro. The deciduoma was enzymatically digested after the removal of the blastocyst and the heterogenous population was cultured in endothelial cell specific medium for 5 days. (i) Phase contrast microscopy indicating the cobble stone morphology of endothelial cells. (ii) Immunostaining with FITC conjugated anti-PECAM-1 antibody further characterized these clusters as endothelial cells. The overlay panel clearly indicates strong membrane specific expression in endothelial cells.
Figure 4.1AFigure 4.1A
i
ii
PECAM DAPI Overlay
Co-culture of stromal-endothelial cells in vitro
a b c
Figure 4.1BFigure 4.1B

99
Figure 4.2: Fluorescence activated cell sorting of uterine endothelial cells
Figure 4.2: Fluorescence activated cell sorting of uterine endothelial cells A: 2D Scatter Plot from the day 8 decidua cell extract. The forward vs. side scatter plot indicates a heterogenous population with 3 major cell clusters grouped based on their varying cell size and internal complexity. B: Cells were stained with propidium iodide and the live population is gated to be sorted in the flow. C: The heterogenous cell population isolated from normal day 8 pregnant uterus was stained with anti-CD31 antibody conjugated to a fluroscein dye FITC. Cells were sorted using the flowcytometer. M1 is defined as the cell population that emits fluorescence higher than the threshold intensity, while M2 is the total number of cells sorted. Upper panel: The distribution of cell population before sorting. Lower panel: Cell population positively sorted for CD-31 cells emitting high fluorescence. FACS resulted in a 90% separation of the endothelial cells from the heterogenous mixture.
Feb 14_PECAM +PI.fcs
PI-A
FSC-A
100 101 102 103 104 105
0
65536
131072
196608
262144
All cells
Figure 4.2BFigure 4.2A
Feb 14_None + pi.fcs
SSC-A
FS
C-A
100
101
102
103
104
105
0
65536
131072
196608
262144
Feb 14_None + pi.fcs
SSC-A
FS
C-A
100
101
102
103
104
105
0
65536
131072
196608
262144
R 2
1 0 0 1 0 1 1 0 2 1 0 3 1 0 4
F IT C
0
3 7
7 5
1 1 2
1 5 0
Counts
Pre-sorting uterine single cell suspension
Post-sorting uterine endothelial cells
Figure 4.2C
M2
M1
M1
Figure 4.2A Figure 4.2B
Figure 4.2C

100
Table 4.1: Percentage of endothelial cells in decidual uterus The table represents the number of cells present and the percentage of the gated cells positive in the M1 and M2 population. In the control sample, stained with the isotype control for PECAM-1, less than 1% of the cells exhibited fluorescence, while in the CD-31 sample, almost 15% of the cells exhibited fluorescence, indicating the percentage of CD-31 positive cells present in the heterogenous population.
Control Sample
Control Sample
CD-31 Sample
CD-31 Sample
Control Sample
Control Sample
CD-31 Sample
CD-31 Sample
Table 4.1Table 4.1

101
Figure 4.3: Classification of the angiogenic molecules down regulated after AI administration
Figure 4.3: Classification of the angiogenic molecules down regulated after AI administration Mice were ovariectomized and subjected to artificially induced decidualization as described in experimental procedures. Cohorts of these animals were treated with P alone (P) or P along with aromatase inhibitor (P+AI) and the uterine horns were separately collected after 72hrs of stimulus (n=5). The isolated mRNA from the stimulated uterine horns was subjected to microarray analysis. The down regulated gene list was analyzed at a threshold cut off of 1.5 folds. A: The genes likely to be involved in angiogenesis were identified and further sub-categorized according to their function, as represented in the pie diagram. B: The factors identified in the microarray analysis were further confirmed by their differential mRNA expression in the control and AI treated uterine extracts. (n=3, p<0.01)
Figure 4.3A
Distribution of genes involved in angiogenesis
Cell proliferation10%
Rho proteins11%
ECM remodeling15%
Endothelial cell-cell communication
13%
Neural guidance10%
Cell migration17%
others in angiogenesis9%
VEGF regulators15%
Figure 4.3B
0
0.2
0.4
0.6
0.8
1
1.2
1.4
HIF-2a Ang-2 Ang-4 Claudin-5 Adrenomedullin
Rel
ativ
e le
vel
od
exp
ress
ion
P
AI
Figure 4.3A
Figure 4.3B
**
*
**

102
Figure 4.3(cont.)
Figure 4.3: Angiogenic network is regulated by intrauterine E C: Uteri from mice treated with P and the inhibitor drug were collected at 72 h following the decidual stimulation and frozen tissue sections were subjected to immunohistochemical analysis using an antibody specific for PECAM-1. Panels a and c indicate the PECAM-1 expression in uteri of mice without letrozole treatment; Panel b and d indicate the PECAM-1 expression in uteri of mice treated with letrozole. M indicates mesometrial end.
Figure 4.3C
-AI +AI
a b
c d
-AI +AI
a b
c d
Figure 4.3C

103
Figure 4.3: (cont.)
Figure 4.3: Endothelial cell proliferation is regulated by local E D: Pregnant mice were ovariectomized on day 5 morning, 12 hrs following the blastocyst attachment. To a subset of these animals, P was administered to sustain pregnancy or treated with P along with aromatase inhibitor (P+AI). The uterine horns were collected after 3days on day 8 (n=5). Uterine cross-sections containing a well defined embryo from the control and AI treated animals were subjected to immunohistochemical analysis using anti Ki-67 antibody. Expression of Ki-67 in the control (panel a) and letrozole treated (panel b) uterus.
Figure 4.3D
a b
-AI +AI
Figure 4.3D

104
Figure 4.4. Stromal cell differentiation induces the release of angiogenic modulators
Fig 4.4: Stromal cell differentiation induces the release of angiogenic modulators Stromal cells isolated from uteri of day 4 pregnant mice were subjected to in vitro decidualization in the presence or absence of letrozole for 72 h. RNA was prepared from these cells and subjected to real-time PCR analysis using gene-specific primers to assess the expression of ER, aromatase (arom), alkaline phosphatase (Alkp), prolactin related protein (PRP), BMP2 and angiogenic regulators: HIF2, angiopoeitin 2 (Ang-2), angiopoeitin 4 (Ang-4), and adrenomedullin (adm).
ER Alkp PRP BMP2 HIF2 Ang2 Ang4 Adm Arom
Figure 4.4Figure 4.4
**
* *
p<0.05

105
Figure 4.5: Stromal cell derived factors are modulated by intrauterine E
Figure 4.5: Stromal cell derived factors are modulated by intrauterine E Angiopoeitin 2 exhibited strong expression in the ecto-placental cone and in the mesometrial blood vessels in uterus (upper panel) and in the decidual cells surrounding the lumen in samples undergoing artificially induced decidualization (lower panel). Control uteri (panels a-c) exhibited strong expression of Ang-2 while those treated with aromatase inhibitor exhibited a significant impairment (panels d-f).
P
P+AI
EM
EMEM
EM
Meso
Meso
Figure 4.5, upper panel
a b
d e
c
f
Lu
Lu
P P+AI
Figure 4.5, lower panel
Figure 4.5, upper panel
Figure 4.5, lower panel

106
Figure 4.6: Intrauterine E regulates the expression of endothelial cell specific angiogenic
molecules
Figure 4.6: Intrauterine E regulates the expression of endothelial cell specific angiogenic molecules Pregnant mice were ovariectomized on day 5 morning, 12 hrs following the blastocyst attachment. To a subset of these animals, P was administered to sustain pregnancy or treated with P along with aromatase inhibitor (P+AI). The uterine horns were collected after 3days on day 8 (n=5). Uterine cross-sections containing a well defined embryo from the control and AI treated animals were subjected to immunohistochemical analysis using anti-Claudin 5 antibody. The protein was specifically localized in the endothelial cell junctions at the mesometrium region in the control (panels a and b) and the aromatase inhibitor treated uteri (panels c and d).
P P+AI
Figure 4.6
a c
b d
Figure 4.6

107
Figure 4.7: Expression of the angiogenic markers in specific uterine zones in the presence and absence of aromatase inhibitor
Figure 4.7: Expression of the angiogenic markers in specific uterine zones in the presence and absence of aromatase inhibitor Uterine cross sections of control and aromatase inhibitor treated samples were subjected to LCM RTPCR. The mesometrial blood vessel region and the antimesometrial stromal cell population were separately captured by the laser dissection. A: The expression of the angiogenic markers were compared in the mesometrial blood vessel compared to the antimesometral stroma. B: Comparison of the expression amongst the control and aromatase inhibitor treated mesometrial blood vessel regions.
P P+AI
EMEM
P P+AI
EMEM
Figure 4.7A
Figure 4.7B
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Ang2 Ang4 Adm Claudin 5 Prp
Rel
ativ
e le
vel
of
exp
ress
ion
Mesometrium bloodvessels
Antimesometrium
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Ang2 Ang4 Adm Claudin5 PrP HIF2a
Rel
ativ
e le
vel of ex
pre
ssio
n
8P-BV
8AI-BV
Figure 4.7C
Figure 4.7A
Figure 4.7B
Figure 4.7C
* *
p<0.01
*
*
**
p<0.05
**

108
Table: 4.2: Molecules involved in axonal guidance also modulates decidual vasculature and is regulated by intrauterine E
Mus musculus semaphorin IV isoform b
Semaphorins are important guidance factors in axon guidance. Semaphorin 4 and semaphorin 3 are also important in vascular development
Mus musculus galanin receptor 2 (Galr2)
galanin is a neuropeptide which is upregulated following a nerve injury, GalR2 expression reduces apoptosis by inducing the Akt pathway, induces neurite outgrowth
Mus musculus ninjurin 1 (Ninj1) axonal guidance and cell migration
Mus musculus neuronal guanine nucleotide exchange factor (Ngef)
Also known as Ephexin A. governs axonal cell growrh and migration. Ephexins are involved in Ephrin signaling pathways
Mus musculus transient receptor potential cationchannel, subfamily V, member 2, mRNA.
calcium-permeable cation channels with polymodal activation properties, governs membrane polarization, present in growth axon cones and directs migration
Mus musculus dynactin 3 (Dctn3)
axonal migration; dynactin proteins are more aggregated in the axonal growth cones and govern the penetration of the microtubules in the peripheral zone of differentiating axonal growth cones
Mus musculus chordin (Chrd)governs neural plate morphogenesis and venous and patterning in cardiovascular development
Mus musculus semaphorin IV isoform b
Semaphorins are important guidance factors in axon guidance. Semaphorin 4 and semaphorin 3 are also important in vascular development
Mus musculus galanin receptor 2 (Galr2)
galanin is a neuropeptide which is upregulated following a nerve injury, GalR2 expression reduces apoptosis by inducing the Akt pathway, induces neurite outgrowth
Mus musculus ninjurin 1 (Ninj1) axonal guidance and cell migration
Mus musculus neuronal guanine nucleotide exchange factor (Ngef)
Also known as Ephexin A. governs axonal cell growrh and migration. Ephexins are involved in Ephrin signaling pathways
Mus musculus transient receptor potential cationchannel, subfamily V, member 2, mRNA.
calcium-permeable cation channels with polymodal activation properties, governs membrane polarization, present in growth axon cones and directs migration
Mus musculus dynactin 3 (Dctn3)
axonal migration; dynactin proteins are more aggregated in the axonal growth cones and govern the penetration of the microtubules in the peripheral zone of differentiating axonal growth cones
Mus musculus chordin (Chrd)governs neural plate morphogenesis and venous and patterning in cardiovascular development
Table 4.2Table 4.2

109
Chapter-5
Estrogen signaling plays a critical role in mediating human endometrial stromal cell
decidualization

110
5.1. ABSTRACT
Implantation, a series of physiological events that entails the initial success of pregnancy is
initiated with the attachment of the embryo to the uterine wall and is critically involved in
sustaining the blastocyst growth before onset of placentation. In the human endometrium
however, pre-decidualization of superficial stromal cells precedes embryo attachment during
the late-secretory phase of every menstrual cycle, while in the event of a successful
conception, the cellular differentiation proceeds through the entire endometrium. The cyclical
modification of the human endometrium is controlled by steroid hormones estrogen (E) and
progesterone (P). Although E gains prominence in the proliferative and P in the secretory
phase of the menstrual cycle, the ultimate outcome is the manifestation of a controlled
interplay of the two steroids. In this study we show that the key enzyme driving the
conversion of androgens to E, P450 aromatase is significantly induced during human
endometrial stromal cell differentiation. The biosynthetic machinery results in local
production of the endogenous steroid in the cultured endometrial stromal cells that reaches
the maximum concentration with the continued presence of P and cAMP. Disruption of
aromatase expression markedly impaired intrauterine E production and stromal cell
differentiation. Further, this local E also drives ER function in coordinating decidualization.
Using small interfering RNA silencing technology we are able to show that the endogenous
E/ER signaling plays a critical role in regulating the expression of several critical factors
mediating stromal differentiation and thereby significantly contributing to endometrial
preparation during the late-secretory phase of the cycle.

111
5.2 INTRODUCTION
Decidualization is the key physiological event that characterizes implantation across different
species. While in rodents it is initiated following embryo attachment to uterine luminal
epithelium, in humans, the endometrium prepares for an incoming blastocyst during every
menstrual cycle through the differentiation of the superficial stromal cells, a process known
as pre-decidualization. Since in humans the stromal cell differentiation occur independent of
pregnancy, the local molecular cues from the epithelial compartment and the steroid
hormones, play the important role in orchestrating the cellular transformation in each late
secretory phase (137, 138).
The cyclical change of the human endometrium is coordinated by a series of hormone
action. Activation of the anterior pituitary by the gonadotropin releasing hormone (GnRH)
from the hypothalamus elicits the release of follicle stimulating hormone (FSH) and
leutenizing hormone (LH) which acts on the ovarian follicles to initiate its maturation and the
subsequent release of steroids. Estrogen (E) released from these mature follicles in turn
regulates the LH surge by a feedback mechanism that regulates follicular rupture. The
cyclical release of steroid hormones from the ovary defines the ovarian follicular cycle that
corresponds to the endometrial growth phases, regarded as the menstrual cycle or endometrial
cycle. During the follicular phase, also known as the endometrial proliferative phase, the
circulating concentration of E reaches high levels and it actively participates in the
regeneration of the endometrium by extensive proliferation of the stromal cells and growth of
the blood vessels (56). Following ovulation, the newly formed corpus luteum releases high
level of P and to a lesser extent, E. Under the influence of these steroid hormones endometrial
stromal cells transforms to the decidual cells which release cytokines, growth factors, and
other signaling molecules, demarcating the secretory phase of the cycle.
The increasing exposure of the endometrium to cytokines, local endocrine factors like
prostaglandin E2, relaxin and corticotrophin releasing hormone (CRH) together stimulates
the production of a second messenger, cyclic adenosine mono-phosphate (cAMP) that
induces the PKA pathway. It has thus been observed that elevated levels of cAMP leading to
sustained activation of the PKA pathway and cAMP induced sensitization of cells to steroid
action are absolutely critical for inducing decidualization in the human endometrium during
the late-secretory phase (139). However, when pregnancy is initiated with an invading

112
blastocyst, the differentiation in the superficial layers extends throughout the entire
endometrium accompanied by myriad physiological modifications that support the embryo
growth until placentation ensues. In the absence of the blastocyst, the endometrium sloughs
off along with the onset of a new menstrual cycle.
The contribution of E signaling in governing stromal differentiation has not been well
explored during implantation. A recent study from our laboratory have shown that murine
decidual uterus harbors the entire steroid biosynthetic machinery driving local E production
that critically sustains decidualization (30). It has been reported that differentiating human
endometrial stromal cells exhibits aromatase enzymatic activity, which is significantly
potentiated by P and relaxin (140-146). However, the expression or the role of aromatase in
human endometrial function has not been investigated. In this study we report for the first
time that like rodents, the human endometrial stromal cells expresses the key enzyme P450
aromatase, resulting in an optimal concentration of local E, which significantly contributes to
endometrial stromal cell transformation. Furthermore, this endogenous E mediates ER
action in bringing about the differentiation of stromal cells to decidual cells. Together, our
findings revealed a pivotal contribution of E regulated factor(s) and signaling pathway(s) that
play important role in sustaining decidualization of fibroblast uterine stromal cells across
species.
5.3. EXPERIMENTAL PROCEDURES
Reagents-
Progesterone, 17--estradiol and 8Br-cAMP were purchased from Sigma Chemical Co. (St.
Louis, MO). Antibodies against P450 aromatase, ER and Calnexin were obtained from
Abcam Inc. (Cambridge, MA) and Santa Cruz Biotechnology Inc. (Santa Cruz, CA),
respectively.
Human endometrial stromal isolation and cell culture-
Human endometrial cell cultures and endometrial biopsies are procured according to the
regulations directed for the protection of human subjects participating in clinical research and
are approved by the Institutional Review Boards (IRBs) of Emory University and University

113
of Illinois. Endometrium samples from the early proliferative stage of the menstrual cycle
were obtained at the Emory University Medical Center from fertile volunteers by Pipelle
biopsy. Stromal cells from the endometrium were isolated by Dr. Taylor’s laboratory (147).
Human endometrial stromal cell differentiation in vitro-
Isolated endometrial stromal cells were cultured in complete medium along with 5% FBS. To
induce differentiation cells were transferred to a 2% medium and incubated with 1M of P
and 0.5mM of a membrane permeable analogue of cyclic adenosine monophosphate (cAMP),
8Br-cAMP. Cells were cultured for an additional 4 days and gene expression was analyzed
using markers of differentiation.
siRNA transfection-
siRNA corresponding to human ER (siERα forward sequence:
UCAUCGCAUUCCUUGCAAAdTdT; and the reverse sequence:
UUUGCAAGGAAUGCGAUGAdTdT.), human P450 aromatase (siAromatase forward
sequence GUAUGUUCCACAUAAUGAAtt and the reverse sequence
UUCAUUAUGUGGAACAUACtt) and a scrambled negative control were obtained from
Dharmacon (Dharmacon Inc., Lafayette, CO) and Ambion (Ambion Inc., Austin, TX),
respectively. The oligos were transfected using the silentFect transfection reagent (Bio-Rad
Inc., Hercules, CA) according to the manufacturer’s instruction. Briefly, the annealed
oligonucleotide was complexed with the transfecting reagent in Opti MEM I reduced serum
medium. The complex containing 20nM of siRNA in 5l of the reagent was dispersed into
culture plates containing stromal cells at 70-80% confluency. 24hrs after transfection, cells
were treated with a hormone cocktail containing 1M P and 0.5mM cAMP and cultured for
an additional 5days before RNA extraction. For aromatase siRNA transfection experiments,
cells were maintained in low serum conditions before gene silencing.
Adenoviral transduction-
Cultured human endometrial stromal cells were transduced with the human ER or the GFP
expressing adenoviruses at a MOI of 50:1. Cells were treated with steroid hormones and
cAMP for an additional 2 days to induce differentiation. The extracted RNA was analyzed
for the expression of various differentiation markers.

114
Estradiol ELISA-
Estradiol levels were monitored by collecting the condition media of differentiating stromal
cells after every 3 days of culture until 9 days. E level in the condition medium was
determined using Estradiol ELISA kit (Calbiotech Inc., Spring Valley, CA) according to the
manufacturer’s instruction. Briefly, equal volume of the concentrated condition media and 6
estradiol reference standard solutions were added to wells coated with goat-anti rabbit IgG in
triplicates and incubated with estradiol-HRP conjugate and rabbit anti-estradiol. Absorbance
measured at 450 nm is a readout of the competitive binding between the HRP-labeled E and
endogenous E in the condition medium. E concentration in the samples was then obtained by
plotting the concentration of the standard versus the absorbance.
Western Blotting-
Whole cell extracts were prepared from mouse primary stromal cell cultures undergoing in
vitro differentiation. Briefly, cells were washed with ice-cold balanced solution and lysed
with the RIPA buffer (0.1% SDS, 0.5% Na-deoxycholate, 1% NP-40 in PBS) containing
protease inhibitor cocktail, phenylmethylsulfonylfluoride (PMSF 0.1 mg/ml), and
phosphatase inhibitor (1:1000; Sigma-Aldrich). This was followed by passing the cells
through a 25G syringe and centrifugation at 12000 rpm for 10 mins to remove the cell debri.
Typically 20-50g of the protein extract was analyzed by SDS-PAGE and transferred to
polyvinylidene fluoride membrane (Amersham Biosciences Inc., Piscataway, NJ). The
membrane was blocked with 5% milk in Tris buffered saline with 0.1% Tween 20 for 1 hr at
room temperature, followed by incubation with a primary antibody against ER or calnexin.
The blot was then incubated with HRP-conjugate secondary antibody for 45 mins at room
temperature. The HRP was detected by chemiluminescence.
Immunohistochemistry and immunocytochemistry-
Polyclonal antibodies against Aromatase was used for immunohistochemistry as discussed
before (100). Paraffin-embedded human endometrial tissues were sectioned at 4 µm and
mounted on slides. Sections were washed in PBS for 20 min and then incubated in a blocking
solution containing 10% normal goat serum for 10 min before incubation in primary antibody
overnight at 40C. Immunostaining was performed using Avidin-Biotin kit for rabbit primary
antibody (Vector Laboratories, Burlingame, CA). Sections were counterstained with

115
hematoxylin, mounted, and examined under bright field. Red deposits indicate the sites of
immunostaining. For immunocytochemistry, cultured cells were fixed in 4%
paraformaldehyde for 10 mins and permeablized using 0.25% Triton X before blocking and
staining with the primary antibody.
5.4. RESULTS
5.4.1. Cytochrome P450 aromatase is induced in differentiating endometrial stromal cells
To explore the mechanism of human endometrial stromal cell decidualization, we obtained
undifferentiated stromal cells from healthy individuals undergoing endometrial biopsies
during the proliferative stage of their menstrual cycle. Cells were cultured in complete
medium and differentiation was induced by adding hormone cocktail containing P (1μM) and
a membrane permeable analogue of cAMP, 8Br-cAMP (0.5mM). As shown in Fig 5.1A,
administration of the hormone cocktail induced a visible transition of the cellular morphology
from spindle like fibroblast appearance (panel a) to plump epitheloid shape (panels b and c),
indicating differentiation of stromal cells to the decidual cell type. We further characterized
decidualization by analyzing the mRNA expression profile of various decidual bio-markers
like prolactin (PRL) (Fig 5.1B, panel A), insulin like growth factor binding protein-1
(IGFBP-1) (panel B), cytokine interleukin-11 (IL-11) (panel C), and vascular endothelial
growth factor (VEGF) (panel D). The expressions of these molecules were significantly
induced as the differentiation progressed 4, 6 and 9 days after the hormone cocktail treatment.
The morphological transformation of the cultured cells along with the increased expression
of factors actively involved in decidualization indicated a faithful in vitro representation of
the cellular mechanisms functional in the late-secretory phase. We next monitored the
expression of P450 aromatase in this cell culture model. Aromatase mRNA was induced
within 2 days of P+cAMP treatment and progressively increased with differentiation,
reaching the maximal levels around 6-9 days of culture (Fig 5.1C). Immunocytochemical
analysis of aromatase protein in cultured stromal cells demonstrated prominent cytoplasmic
expression of the enzyme in differentiating stromal cells. Consistent with the mRNA pattern,
the protein expression was also significantly induced after 6 days and reached the maximum
levels at 9 days of P and cAMP treatment (Fig 5.1D).

116
The expression of aromatase in the endometrium was further confirmed by localizing
aromatase protein in tissue sections obtained from endometrial biopsies of healthy individuals
in the late secretory phase of menstrual cycle (Fig 5.1E). The cytoplasmic aromatase protein
expression was restricted to the stromal cells surrounding the spiral arteries (panels A and B),
while the epithelial cells were devoid of positive staining.
5.4.2. Functional E biosynthetic machinery in differentiating endometrial stromal cells
Induction of aromatase in differentiating stromal cells raised the possibility of a functional E
biosynthetic machinery in the endometrium. We first examined the expression of enzymes
responsible for steroid biosynthesis in cultured endometrial stromal cells. Consistent with the
previous reports, strong induction of StAR (steroidogenic acute regulatory protein), P450scc
(P450 side chain cleavage), P450c17 (17-lyase) and 3-HSD (3b-hydroxysteroid
dehydrogenase) was observed in differentiating stromal cells in response to P and cAMP (Fig
5.2A). More importantly, to address the local steroid hormone biosynthesis in these decidual
cells, the medium conditioned by stromal cells undergoing differentiation was collected every
three days and subjected to E (17 estradiol) measurement using an ELISA kit as detailed in
experimental procedures. While the steroid levels in the medium were very low to negligible
prior to 3 days of hormone cocktail treatment, the estradiol level was significantly elevated
after 6 days and reached the maximum concentration around 9 days of culture (Fig 5.2B). We
were able to detect 30-40 pg/ml of E in differentiated stromal cells. Together these
experiments indicated that induction of the key enzyme P450 aromatase along with other
steroid biosynthetic components governed endogenous E production in differentiating human
endometrial stromal cells.
5.4.3. Endogenous E plays a major role in mediating stromal differentiation
To explore the functional significance of local E biosynthesis in human endometrial stromal
cells, we silenced aromatase expression in cultured cells by administering a siRNA targeted
to its mRNA. Differentiation was induced 24 hrs after siRNA transfection by administering P
and cAMP. Cells were cultured for an additional 6 days before analyzing the extent of
decidualization. Administration of aromatase siRNA to cells led to more than 87% of gene
silencing when compared to the cells transfected with a control scrambled siRNA (NC
siRNA) (Fig 5.3A, left panel). The E levels, determined by subjecting the condition media to

117
ELISA assay, were also significantly down regulated in the aromatase siRNA treated cells,
compared to scrambled siRNA transfection (right panel). To address the effect of aromatase
knockdown on cell differentiation we analyzed the mRNA expression of the well established
decidual bio-markers. IGFBP-1, PRL were significantly inhibited along with other markers
like IL-11, VEGFa, FOXO1 and Wnt4. The expression of ER, however, remained unaltered
after silencing of the aromatase gene. These experiments indicates that aromatase derived E is
important in sustaining human endometrial decidualization.
5.4.4. Optimal concentration of local E critically regulates stromal differentiation
To address the fact that stromal differentiation defect observed after silencing aromatase
expression is a consequence of impaired E signaling, we next performed a rescue experiment.
Aromatase siRNA treated cells were cultured in a medium supplemented with 10nM of
exogenous E along with the differentiating cocktail. As expected, cells transfected with
aromatase siRNA exhibited a significant down regulation of PRL and IGFBP-1 when
cultured in presence of P and cAMP (Fig 5.4A). However, administration of E to these
differentiating cells restored the inhibition of the expression of the decidual bio-markers,
indicating a critical role of endogenous E in mediating stromal decidualization.
Endometriosis, a pathological condition that results in impaired endometrial
differentiation, is characterized by increased aromatase activity and E levels in both eutopic
and ectopic stromal cells (148). To address this apparent contradiction with our observation
in the healthy endometrium, we cultured fibroblast stromal cells with steroid hormones and
cAMP for 5days and monitored expression of the decidual bio-markers. As indicated in Fig
5.4B, in the presence of complete medium with FBS (None), there was no induction of
decidualization. Culturing these cells with P and cAMP along with increasing concentration
of E indicated that 10 and 50nM of E was capable of inducing a complete decidual response
in stromal cells. However, E dosage 100nM and above impaired the induction of IGFBP-1
and PRL mRNA expression, indicating that an optimal concentration of E is necessary to
induce complete decidual response in the human stroma.
5.4.5. ER plays a major role in stromal cell differentiation
Nuclear steroid receptors play pivotal role in mediating the hormone action in reproductive
tissues. E signaling is mediated via two receptor isoforms ER and ER. In order to

118
understand the signaling mechanism of endogenous E in human endometrium, we next
studied the expression of ER, the predominant form of ER in the endometrium, during
human stromal cell decidualization. Primary stromal cell cultures were subjected to
decidualization in response to with P and cAMP. The expression pattern of ER mRNA (Fig
5.5 upper panel) and protein (lower panel) was investigated on different days of hormone
treatment. The expression of ER mRNA and protein was elevated in cultured stromal cells
with progression of differentiation.
To address the role of ER in human decidualization, we employed the RNA interference
strategy and silenced the endogenous transcript by administering ER siRNA. 24hrs after
administration of siRNA to the fibroblast stromal cells, differentiation was induced by adding
P and cAMP and cultured in the presence of hormone cocktail. As shown in Fig 5.6A, the
expression of ER was significantly down regulated both at the transcript (upper panel) and
protein levels (lower panel) after treating with the ER siRNA. While cells transfected with
the NC siRNA underwent the usual cellular transformation after 6 days of culture in the
presence of hormone cocktail, transfection of ER siRNA significantly impaired the
morphological transition, indicating a severe impairment in stromal cell differentiation (Fig
5.6B). We also measured the gene expression of the well characterized bio-markers of human
stromal decidualization after silencing ER in cultured stromal cells. Our studies revealed
that while the expression of MMP2 and GAPDH remained unaltered, the expression of PR,
PRL, IGFBP-1, VEGFa, IL-11, Wnt4 and FOXO1 were significantly down regulated in these
cells indicating a critical role of ER in regulating stromal cell differentiation program (Fig
5.6C).
We further confirmed the above observation by over expressing ER in cultured stromal
cells via viral transduction. Adenovirus encoding the ER (Ad-ER) and the GFP sequence
(Ad-GFP) were transduced in cultured stromal cells at a multiplicity of infection (MOI) of
50:1. 48 hrs after viral administration to the differentiating stromal cells, RNA was isolated
and expression of the major decidual bio markers was analyzed by qPCR. Consistent with our
previous observation, we found that over expression of ER in these cultured stromal cells
accentuated the differentiation process resulting in the increased expression of factors that
promote endometrial decidualization (Fig 5.6D).

119
5.5. DISCUSSION
Although certain aspects of establishment of pregnancy in mouse and human are dissimilar,
yet the orchestrations of steroid hormones in regulation of this event remain comparable in
both the species. While E plays the major role in controlling cell proliferation to prepare the
uterus for pregnancy, P critically regulates cell differentiation to sustain the blastocyst growth
before placentation ensues. However in either case, the physiological response achieved is the
consequence of a controlled interplay of these two steroids. The contribution of E in stromal
cell differentiation remained ambiguous in the field of implantation biology until a recent
study showed the induction of aromatase expression in mouse decidual uterus and the pivotal
role of intrauterine E action in modulating the decidualization process. This raised an
interesting possibility that perhaps endogenous E signaling is important in human stromal cell
decidualization.
In previous studies Tseng et. al. reported aromatase enzymatic activity in cultured
endometrial stromal cells in response to forskolin and MPA (141-143, 145). In a more recent
study Aghajanova et.al, studied the comparative expression profile of the various
steroidogenic enzymes in the endometrium of diseased and healthy individuals (149).
Isolated stromal cells were cultured either in the presence of cAMP alone for 96 hrs or P for
14 days. Although most of the biosynthetic enzymes were induced in either sample, the
extent of induction was undoubtedly higher in the endometriotic cells. Our studies employing
isolated human endometrial stromal cells cultured in the presence of P and cAMP showed a
significant induction of aromatase expression both in the transcript and protein levels (Fig
5.1). Additionally the expression of StAR, SCC, 3bHSD and lyase was also prominent in
these differentiating stromal cells, similar to previous reports. Bulun and co-workers
however, were unable to detect any aromatase transcript in the normal endometrial stromal
cells (150). While our findings remain consistent with other previous observations, the
difference could be accounted to the varying treatment parameters, experimental design and
reduced sensitivity of the semi quantitative PCR compared to the real time PCR analysis
performed in this study. To confirm the expression of aromatase in differentiating human
endometrial stromal cells, we validated the amplified cDNA fragment by PCR-sequencing
and identified a single band that shows 100% homology with human Cyp19A1 gene sequence
(NCBI Acc no: DQ118405). Additionally, immunolocalization of aromatase protein

120
specifically to endometrial stromal cells further confirmed the expression of aromatase in the
normal cycling individuals.
The hormone levels in stromal cells obtained from diseased and healthy individuals
cultured in the presence of PGE2 was recently studied by Attar et.al (151). Stromal cells from
the healthy endometrium exhibited low E levels and 48 hrs of vehicle or PGE2 treatment
resulted in less than 10 pg/ml of E in the culture medium; a concentration significantly lower
when compared to those from endometriotic cells. In our studies, we were unable to detect E
in cultured medium in the absence of P and cAMP. After 3 days of hormone cocktail
administration the E concentration was detected below 10 pg/ml. The prolonged treatment
with P and cAMP for 9 days however resulted in almost 40 pg/ml of E in the condition
medium.
In the human during menstrual cycle, E is primarily synthesized by the developing ovarian
follicle and to a reduced extent by the corpora lutea in the early to mid-secretory phase of the
endometrium. However, this circulatory steroid declines precipitously towards the late-
secretory phase while stromal cells are undergoing active differentiation. Following a
successful implantation, the placenta becomes the predominant site of E biosynthesis
throughout the first trimester of pregnancy. The decidua-parietalis, regarded as the gestational
endometrium, maintains a close association with the fetal membrane throughout pregnancy.
A complete differentiation of this maternal compartment is necessary to sustain pregnancy. It
is thus possible that the presence of functional endogenous E biosynthetic machinery in the
endometrial decidual cells probably acts as a functional bridge, to support the transition of the
steroid source from the maternal tissue to placenta.
Administration of aromatase inhibitor letrozole to pregnant mouse results in embryo
resorption and a significant reduction in the decidual cell mass. Molecular analysis revealed
that the expressions of the major markers of mouse decidualization including PRP, Alkp, PR,
BMP2 and Cx43 were significantly impaired after inhibitor treatment. Additionally,
disruption of the intrauterine E biosynthesis severely impacted the development of mouse
decidual angiogenesis and blood vasculature formation (30). The present study indicated that
similar to its contribution in murine decidualization, aromatase derived intrauterine E also
plays a critical role in human endometrial stromal cell differentiation. RNAi mediated gene

121
silencing of aromatase led to a severe defect in the expression of the various decidualization
markers due to altered E biosynthesis in the differentiating stromal cells.
Endometriosis, a leading cause of infertility in women, leads to impaired stromal cell
decidualization (152). It occurs due to retrograde menstruation during which endometrial
fragments are implanted at ectopic sites on the pelvis and the ovary. As discussed before,
individuals suffering from this pathological condition exhibits increased aromatase activity
and endogenous E biosynthesis in the endometrial stromal cells (153). The heightened E
action in these cells stimulates cell proliferation resulting in altered differentiation (154).
Uterine tumors like leiomyomas, endometrial cancer and endometrial hyperplasia are
additional diseased conditions with enhanced E activity (155). Thus, the significance of local
E biosynthesis during decidualization in healthy diseased free uterine tissue was difficult to
envisage. In our experiments, the concentration of E measured in the condition media varied
from 20-40 pg/ml, which translates to 0.07 nM to 0.15 nM of E, a concentration significantly
lower compared to cells obtained from endometriotic tissue (151). Because E is markedly
elevated in endometriotic cells, we hypothesized that at an optimal concentration of E is
needed to support the differentiation process. We further confirmed our hypothesis by
culturing differentiating stromal cells with an increasing dosage of E. A significant reduction
in the expression of decidual biomarkers in response to high E concentration explained the
differentiation defect in the case of endometriosis.
Nuclear receptor action is the classical signaling pathway through which steroid hormones
mediate their effect in reproductive tissues. Human endometrium displays expression of both
ER and ER in the stromal and glandular epithelial cells (156, 157). While the role of ER
is widely established in stromal cell regeneration and in regulating angiogenic factors in the
proliferative phase, the role of ER remains unknown (158, 159). In the mouse, ER is the
predominant E receptor playing pivotal roles in regulating epithelial receptivity and embryo
attachment in pre-implantation phase. Studies from our laboratory have also revealed that
conditional ablation of ER in endometrial stromal cells using a Cre-loxP technology renders
the mouse absolutely unresponsive to any decidual stimulus, indicating a pivotal role of this
nuclear receptor in decidualization (Laws MJ and Bagchi IC, unpublished observations). In
this study, siRNA mediated ER gene silencing in human endometrial stromal cells also
exhibited severe defect in cell differentiation. Silencing of ER impaired transformation of

122
the fibroblastic stromal cell morphology to the decidual structure, one of the characteristic
features of cellular differentiation. Furthermore, many critical markers of human
decidualization are down regulated in the absence of ER. Conversely, over expression of
ER in endometrial stromal cells led to enhanced expression of the bio-markers, indicating
that de novo synthesis of decidual E functioning via ER plays a critical role in regulating
human endometrial stromal cell differentiation.

123
5.6. FIGURES Figure 5.1: Cytochrome P450 aromatase is induced in differentiating human stromal cells
Figure 5.1: Cytochrome P450 aromatase is induced in differentiating human stromal cells Human endometrial stromal cells were isolated from cycling, healthy individuals during the late proliferative stage of the menstrual cycle and cultured in vitro in complete medium. Differentiation was induced by adding a hormone cocktail containing 1M P and 0.5mM cAMP and cultured for an additional 0-9days (n=3). A: Cell morphology was captured by phase microscopy in the absence (None) or 6 and 9 days after adding the hormone cocktail (middle and right panels). B: Cells were harvested at the 2, 3, 6 and 9 days after adding the hormone cocktail and the isolated total RNA was subjected to q-PCR analysis using gene specific primers for PRL (panel a), IGFBP-1 (panelb), IL-11 (panel c), VEGF (panel d) P450 aromatase (Fig 5.1C).
None 6 days 9 days
Figure 5.1A
a b c
Figure 5.1B
0 day 2 day 3 day 6 day 9 day
0 day 2 day 3 day 6 day 9 day
A
B
0
5
10
15
20
25
0 day 2day 3day 6day 9day
Re
lati
ve
lev
el o
f e
xp
res
sio
n
IL-11
02468
1012
0 day 2day 3day 6day 9day
Rel
ativ
e le
vel
of
exp
ress
ion
VEGF
0 day 2 day 3 day 6 day 9 day
0 day 2 day 3 day 6 day 9 day
C
D
Figure 5.1C
Figure 5.1A
Figure 5.1B
Figure 5.1Cp<0.05
* *
*
*
*
*
*
**
***

124
Figure 5.1 (cont.)
Figure 5.1: Cytochrome P450 aromatase is induced in differentiating human stromal cells D: Cultured stromal cells were fixed before (panel a and b), and after 6days (panel c and d) and 9days (panel e and f) days of hormone cocktail treatment and protein expression of aromatase was monitored. E: Tissue sections obtained from healthy individuals undergoing endometrial biopsies in the late secretory phase was subjected to immunohistochemistry using anti-aromatase antibody. Strong cytoplasmic expression is observed in the stromal cells surrounding the spiral artery (panel a and b). Panel c shows the negative control, immunostained in the absence of a specific primary antibody. Str: stroma, SA: Spiral arteries, LE: Luminal epithelium, GL: Uterine glands.
None 6 days 9 days
IHC: AromataseFigure 5.1D
N/C
Gl
Str
Str
Gl
Figure 5.1E
A
C
B
SA
Str
Figure 5.1D
Figure 5.1E

125
Figure5.2: Functional E biosynthetic machinery in human endometrial stromal cells
Figure 5.2: Functional E biosynthetic machinery in differentiating human endometrial stromal cells Human endometrial stromal cells isolated from cycling and healthy individuals during the late proliferative stage of the menstrual cycle were cultured in the presence of hormone cocktail to induce decidualization. A: Stromal cells were subjected to RNA isolation after 2, 4 and 6 days of culture and subjected to q-PCR analysis using the primers specific to human StAR (steroidogenic acute regulatory protein), P450scc (P450 side chain cleavage), P450c17 (17-lyase) and 3-HSD (3b-hydroxysteroid dehydrogenase). B: Steroid hormone levels were measured by ELISA in the conditioned medium on days 0, 3, 6 and 9 after administration of the hormone cocktail (n=3).
0
1
2
3
4
5
6
7
8
SCC StAR 17alpha 3bHSD
Steroid biosynthetic enzymes
Rel
ativ
e le
vel
of
exp
ress
ion
None
2days
4days
6days
P450 SCC StAR 17lyase 3HSD
Fig 5.2A
0 day 3 day 6 day 9 day
Fig 5.2B
Figure 5.2A
Figure 5.2B

126
Figure 5.3: Endogenous E plays a major role in stromal cell differentiation
Figure 5.3: Endogenous E plays a major role in stromal cell differentiation Primary human endometrial stromal cells were cultured in reduced serum conditions and transfected with aromatase and scramble (NC) siRNA. 24hrs after transfection, differentiation was induced by administering P and cAMP and cells were collected 5days after addition of the hormone cocktail. A: Down regulation of aromatase mRNA expression in cells treated with aromatase siRNA compared to control siRNA transfected cells, monitored by qPCR analysis (upper panel) and reduction of endogenous E concentration in the condition media of aromatase siRNA treated cells, assessed by ELISA (lower panel). B: Harvested cells were subjected to q-PCR analysis to monitor the gene expression of decidualization bio-markers like PRL, IGFBP-1, BMP2, Wnt4 and ER. (n=4, p<0.05)
A
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Arom Prl IGFBP1 Wnt4 Foxo1 VEGF ER-a
Rel
ativ
e le
vel
of
exp
ress
ion
NC siRNA
Ar siRNA
B
Fig 5.3
0
0.2
0.4
0.6
0.8
1
1.2
1.4
NC siRNA Ar siRNA
Re
lati
ve
lev
el o
f e
xp
res
sio
n
Aromatase
Q-PCR
Q-PCR
ELISA
Figure 5.3
****
*
*

127
Figure 5.4: Optimal concentration of E sustains decidualization
Figure 5.4: Optimal concentration of E sustains decidualization Primary human endometrial stromal cells isolated from normal cycling individuals were cultured in vitro. A: Cells were transfected with aromatase siRNA and a control NC siRNA. 24hrs after siRNA transfection cells were treated with a hormone cocktail containing P+cAMP (yellow bars) or E+P+cAMP (green bars). RNA was isolated after 6days of siRNA transfection and the expression of the decidual bio-markers PRL and IGFBP-1 was monitored by qPCR. B: Differentiation was induced in cultured stromal cells by administering increasing concentration of E (10nM, 100nM and 1M) along with P and cAMP. After 6 days of culture, cells were harvested and the RNA isolated was subjected to q-PCR analysis using specific primers for PRL (upper panel) and IGFBP-1 (lower panel). (n=3, p<0.05)
Fig 5.4A
Fig 5.4B
Figure 5.4A
Figure 5.4B
*
*
*
*
*

128
Figure 5.5: ER exhibits predominant stromal cell expression in cultured human endometrial cells
Figure 5.5: ER exhibits predominant stromal cell expression in cultured human endometrial cells Human endometrial stromal cells isolated from cycling, healthy individuals during the late proliferative stage of the menstrual cycle were cultured in vitro. Differentiation was induced by adding a hormone cocktail containing 1M P and 0.5mM cAMP for 0-9days. A: RNA isolated from the differentiating cells after 0, 3, 6 and 9 days of culture were subjected to qPCR analysis using ER specific primers. B: 30g of whole cell protein extracts isolated at the same time-points from the cultured cells were resolved on a 10% SDS-PAGE gel and the protein level of ER was monitored by Western blot analysis. An equal amount of whole cell extract from ovary was loaded as positive control. The expression level of Calnexin, a calcium binding protein, was also monitored to ensure equal loading of the protein extracts. (n=3, p<0.05)
0d 3d 6d 9d OvrERa
Calnexin
Fig 5.5
Q-PCR
Immunoblot
ER
Figure 5.5B
Figure 5.5A
* *

129
Figure 5.6: Endogenous E acts through ER in mediating the stromal decidualization process.
Figure 5.6: Endogenous E acts through ER in mediating the stromal decidualization process. Primary human endometrial stromal cells were transfected with ER and NC specific siRNA. 24hrs after transfection, differentiation was induced by administering P and cAMP and cells were isolated 5days after addition of the hormone cocktail. A: Down regulation of ER mRNA (upper panel) and protein (lower panel) was assessed by qPCR and WB respectively. B: Morphology of cells cultured in the presence of hormone cocktail, transfected with NC siRNA (upper panels) and the ER siRNA (lower panels) at similar concentrations.
NC-20nM ER-20nM
ER
Calnexin
Immunoblot:ERalpha
Fig 5.6A
NC-10nM NC-20nM
ER-10nM ER-20nM
Fig 5.6BFigure 5.6B
Figure 5.6A
*

130
Figure 5.6 (cont.)
Figure 5.6: Endogenous E acts through ER in mediating the stromal decidualization process C: Relative level of expression of the decidualization markers like PRL, IGFBP-1, BMP2, Wnt4 in ER silenced cells compared to the NC siRNA treated cells was assessed by qPCR. D: Fibroblast stromal cells were transduced with an adenovirus encoding ER or a control virus expressing GFP at a multiplicity of infection (MOI) of 50:1. Differentiation was then induced by adding P+cAMP. Cells were harvested 48hrs following viral administration. The RNA isolated was analyzed by qPCR to monitor the expression level of the different decidual bio markers after ER over expression compared to the control virus infected cells. (n=3, p<0.05)
0
0.2
0.4
0.6
0.8
1
1.2
ER PR Prl IGFBP-1 VEGF IL11 Wnt4 FOXO1 GAPDH MMP2
Rel
ativ
e le
vel
of
exp
ress
ion
NC siRNA
ER siRNA
Fig 5.6C
0
5
10
15
20
25
30
35
PR Prl IGFBP-1 Wnt4 VEGF IL-11 GAPDH
Re
lati
ve
lev
el o
f e
xp
res
sio
n
Ad GFP+cAMP
Ad ER+cAMP
Fig 5.6DFigure 5.6D
Figure 5.6C
*
*
* *
**
**
* *
*

131
Summary
Implantation is initiated upon attachment of the embryo to the uterine luminal epithelium,
which in mouse occurs on day 4 of pregnancy. This event is followed by proliferation and
differentiation of uterine stromal cells into morphologically distinct secretory decidual cells,
which control trophoblast invasion and embryonic growth until placentation ensues. As the
blastocyst invades though the maternal wall, the decidualized stromal cells at the immediate
vicinity of the embryo migrate away to accommodate the growing embryo. It is well
established that ovarian steroids, estrogen (E) and progesterone (P), are essential to prepare
the uterus for embryo implantation. While P plays a critical role in decidualization, the role of
E in stromal differentiation and remodeling is less clear. Studies in our labiratory revealed
that the decidual uterus expresses all the enzymatic components of the steroid hormone
biosynthetic machinery, including the key enzyme P450 aromatase, which converts
testosterone to E. The overall finding of this dissertation is focused on identifying the role of
E signaling during decidualization. Importantly, we identified that this pathway is
phyllogenetically conserved in both mouse and human endometrium.
In the preimplantation phase, while the ovarian source of E critically regulates epithelial
receptivity by mediating the action of ER, with the initiation of stromal differentiation,
P450 aromatase, which is induced in the decidualizing stromal cells, along with the other
critical steroid biosynthetic enzymes regulates the de novo synthesis of E. Interestingly, we
have further observed that induction of aromatase in the decidualization phase is regulated by
the transcription factor C/EBPbeta. The intrauterine E plays a major role in modulating the
differentiation of stromal cells by regulating down stream target molecules like PR, Wnt4,
Bmp2. In addition to this PR dependent pathway, it also governs the expression of some
known targets of E signaling constituting the PR independent pathway, including Cx43 and
CYR61. Altered differentiation consequently impairs the release of several cytokines which
through a paracrine mechanism act on the neighboring endothelial cells governing the growth
and maturation of the uterine angiogenic networks. Again, local E regulated the expression of
endothelial cell specific molecules which contributed to the development of angiogenesis.
Finally, we identified the transcription factor FRA-1 as a direct target of E-ER signaling
during decidualization. While FRA-1 is regulated by ER at the transcription level, the

132
activation and nuclear stabilization of FRA-1 is governed by the ERK1/2-MAPK pathway.
FRA-1 has major contributions in governing stromal differentiation. Our studies showed that
inhibition of FRA-1 expression in stromal cells blocked the decidualization process as
indicated by the down regulation of well-known biomarkers of stromal differentiation.
Interestingly, we also observed that down regulation of FRA-1 expression impaired the
migration of stromal cells in an in vitro wound-healing assay. This defect in stromal cell
mobility is mediated through extra cellular matrix modulators like MMP, TIMPs and CD44.
Our study, therefore, established that biosythesis of local E acting through ER is a major
signaling pathway that critically regulates stromal cell differentiation and migration by
regulating the expression of FRA-1 and contributes to the neo-angiogenic networking by
modulating the expression of various paracrine signaling factors. Most importantly, we
observed that this signaling mechanism remains conserved during human decidualization,
playing pivotal roles in stromal differentiation. Fig. 6 summarizes these E mediated
transcriptional networks that critically modulate the success of decidualization during early
pregnancy.
The specific regulation of aromatase in the differentiating uterus still remains unknown.
Aromatase expression is regulated by various tissue specific promoters. Some of the plausible
candidates involved in regulating aromatase expression in the decidual uterus include
members of the monomeric orphan nuclear receptor family, which are key aromatase
transcription regulators in other E-producing tissues. Steroidogenic factor 1 (SF-1) and liver-
receptor homolog-1 (LRH-1) were the two major factors studied. While some of the
preliminary experiments suggest that the expression of aromatase can be regulated by the
orphan nuclear receptor LRH-1, extensive studies require further characterization of the
molecular details.
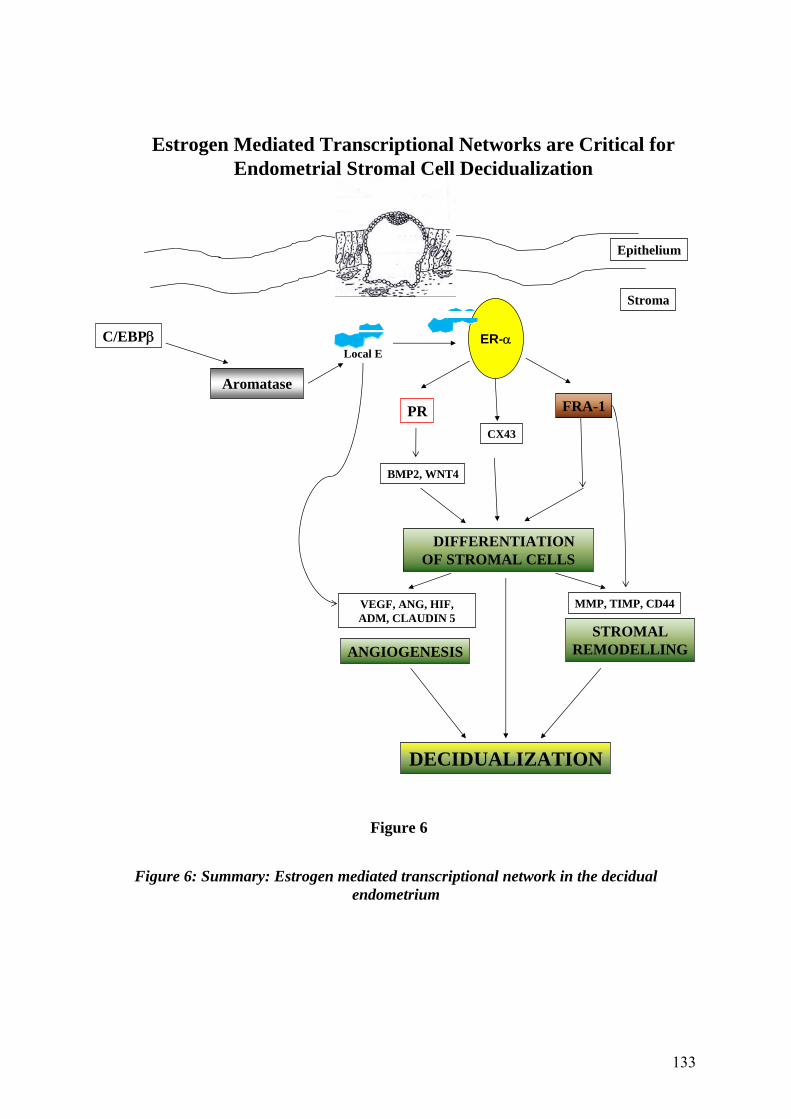
133
Figure 6: Summary: Estrogen mediated transcriptional network in the decidual endometrium
Epithelium
Stroma
CX43
C/EBP
VEGF, ANG, HIF, ADM, CLAUDIN 5
PR
ANGIOGENESIS
DIFFERENTIATION OF STROMAL CELLS
FRA-1
ER-
Aromatase
Local E
MMP, TIMP, CD44
STROMAL REMODELLING
BMP2, WNT4
Estrogen Mediated Transcriptional Networks are Critical for Endometrial Stromal Cell Decidualization
DECIDUALIZATION
Figure 6

134
Bibliography

135
REFERENCES:
1. Yoshinaga K 1988 Uterine receptivity for blastocyst implantation. Ann N Y Acad Sci 541:424-431
2. Carson DD, Bagchi I, Dey SK, Enders AC, Fazleabas AT, Lessey BA, Yoshinaga K 2000 Embryo implantation. Dev Biol 223:217-237
3. Dey SK, Lim H, Das SK, Reese J, Paria BC, Daikoku T, Wang H 2004 Molecular cues to implantation. Endocr Rev 25:341-373
4. Gellersen B, Brosens IA, Brosens JJ 2007 Decidualization of the human endometrium: mechanisms, functions, and clinical perspectives. Semin Reprod Med 25:445-453
5. O'Malley BW, Tsai SY, Bagchi M, Weigel NL, Schrader WT, Tsai MJ 1991 Molecular mechanism of action of a steroid hormone receptor. Recent Prog Horm Res 47:1-24; discussion 24-26
6. Wada-Hiraike O, Hiraike H, Okinaga H, Imamov O, Barros RP, Morani A, Omoto Y, Warner M, Gustafsson JA 2006 Role of estrogen receptor beta in uterine stroma and epithelium: Insights from estrogen receptor beta-/- mice. Proc Natl Acad Sci U S A 103:18350-18355
7. McDevitt MA, Glidewell-Kenney C, Jimenez MA, Ahearn PC, Weiss J, Jameson JL, Levine JE 2008 New insights into the classical and non-classical actions of estrogen: evidence from estrogen receptor knock-out and knock-in mice. Mol Cell Endocrinol 290:24-30
8. Li X, O'Malley BW 2003 Unfolding the action of progesterone receptors. J Biol Chem 278:39261-39264
9. Lydon JP, DeMayo FJ, Funk CR, Mani SK, Hughes AR, Montgomery CA, Jr., Shyamala G, Conneely OM, O'Malley BW 1995 Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities. Genes Dev 9:2266-2278
10. Cheon YP, Li Q, Xu X, DeMayo FJ, Bagchi IC, Bagchi MK 2002 A genomic approach to identify novel progesterone receptor regulated pathways in the uterus during implantation. Mol Endocrinol 16:2853-2871
11. Tranguch S, Cheung-Flynn J, Daikoku T, Prapapanich V, Cox MB, Xie H, Wang H, Das SK, Smith DF, Dey SK 2005 Cochaperone immunophilin FKBP52 is critical to uterine receptivity for embryo implantation. Proc Natl Acad Sci U S A 102:14326-14331
12. Matsumoto H, Zhao X, Das SK, Hogan BL, Dey SK 2002 Indian hedgehog as a progesterone-responsive factor mediating epithelial-mesenchymal interactions in the mouse uterus. Dev Biol 245:280-290
13. Lee K, Jeong J, Kwak I, Yu CT, Lanske B, Soegiarto DW, Toftgard R, Tsai MJ, Tsai S, Lydon JP, DeMayo FJ 2006 Indian hedgehog is a major mediator of progesterone signaling in the mouse uterus. Nat Genet 38:1204-1209
14. Simon L, Spiewak KA, Ekman GC, Kim J, Lydon JP, Bagchi MK, Bagchi IC, DeMayo FJ, Cooke PS 2009 Stromal progesterone receptors mediate induction of Indian Hedgehog (IHH) in uterine epithelium and its downstream targets in uterine stroma. Endocrinology 150:3871-3876
15. Kurihara I, Lee DK, Petit FG, Jeong J, Lee K, Lydon JP, DeMayo FJ, Tsai MJ, Tsai SY 2007 COUP-TFII mediates progesterone regulation of uterine implantation by controlling ER activity. PLoS Genet 3:e102
16. Lee KY, Jeong JW, Wang J, Ma L, Martin JF, Tsai SY, Lydon JP, DeMayo FJ 2007 Bmp2 is critical for the murine uterine decidual response. Mol Cell Biol 27:5468-5478

136
17. Adams RH, Alitalo K 2007 Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol 8:464-478
18. Lim H, Ma L, Ma WG, Maas RL, Dey SK 1999 Hoxa-10 regulates uterine stromal cell responsiveness to progesterone during implantation and decidualization in the mouse. Mol Endocrinol 13:1005-1017
19. Daikoku T, Tranguch S, Friedman DB, Das SK, Smith DF, Dey SK 2005 Proteomic analysis identifies immunophilin FK506 binding protein 4 (FKBP52) as a downstream target of Hoxa10 in the periimplantation mouse uterus. Mol Endocrinol 19:683-697
20. Mantena SR, Kannan A, Cheon YP, Li Q, Johnson PF, Bagchi IC, Bagchi MK 2006 C/EBPbeta is a critical mediator of steroid hormone-regulated cell proliferation and differentiation in the uterine epithelium and stroma. Proc Natl Acad Sci U S A 103:1870-1875
21. Tong W, Pollard JW 1999 Progesterone inhibits estrogen-induced cyclin D1 and cdk4 nuclear translocation, cyclin E- and cyclin A-cdk2 kinase activation, and cell proliferation in uterine epithelial cells in mice. Mol Cell Biol 19:2251-2264
22. Zhu L, Pollard JW 2007 Estradiol-17beta regulates mouse uterine epithelial cell proliferation through insulin-like growth factor 1 signaling. Proc Natl Acad Sci U S A 104:15847-15851
23. Chen B, Pan H, Zhu L, Deng Y, Pollard JW 2005 Progesterone inhibits the estrogen-induced phosphoinositide 3-kinase-->AKT-->GSK-3beta-->cyclin D1-->pRB pathway to block uterine epithelial cell proliferation. Mol Endocrinol 19:1978-1990
24. Gidley-Baird AA 1981 Endocrine control of implantation and delayed implantation in rats and mice. J Reprod Fertil Suppl 29:97-109
25. Yoshinaga K, Adams CE 1966 Delayed implantation in the spayed, progesterone treated adult mouse. J Reprod Fertil 12:593-595
26. Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS, Smithies O 1993 Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc Natl Acad Sci U S A 90:11162-11166
27. Paria BC, Tan J, Lubahn DB, Dey SK, Das SK 1999 Uterine decidual response occurs in estrogen receptor-alpha-deficient mice. Endocrinology 140:2704-2710
28. Couse JF, Curtis SW, Washburn TF, Eddy EM, Schomberg DW, Korach KS 1995 Disruption of the mouse oestrogen receptor gene: resulting phenotypes and experimental findings. Biochem Soc Trans 23:929-935
29. Kos M, Denger S, Reid G, Korach KS, Gannon F 2002 Down but not out? A novel protein isoform of the estrogen receptor alpha is expressed in the estrogen receptor alpha knockout mouse. J Mol Endocrinol 29:281-286
30. Das A, Mantena SR, Kannan A, Evans DB, Bagchi MK, Bagchi IC 2009 De novo synthesis of estrogen in pregnant uterus is critical for stromal decidualization and angiogenesis. Proc Natl Acad Sci U S A 106:12542-12547
31. Ma W, Tan J, Matsumoto H, Robert B, Abrahamson DR, Das SK, Dey SK 2001 Adult tissue angiogenesis: evidence for negative regulation by estrogen in the uterus. Mol Endocrinol 15:1983-1992
32. Grummer R, Hewitt SW, Traub O, Korach KS, Winterhager E 2004 Different regulatory pathways of endometrial connexin expression: preimplantation hormonal-mediated pathway versus embryo implantation-initiated pathway. Biol Reprod 71:273-281

137
33. Laws MJ, Taylor RN, Sidell N, DeMayo FJ, Lydon JP, Gutstein DE, Bagchi MK, Bagchi IC 2008 Gap junction communication between uterine stromal cells plays a critical role in pregnancy-associated neovascularization and embryo survival. Development 135:2659-2668
34. Ferrara N 2000 Vascular endothelial growth factor and the regulation of angiogenesis. Recent Prog Horm Res 55:15-35; discussion 35-16
35. Chakraborty I, Das SK, Dey SK 1995 Differential expression of vascular endothelial growth factor and its receptor mRNAs in the mouse uterus around the time of implantation. J Endocrinol 147:339-352
36. Douglas NC, Tang H, Gomez R, Pytowski B, Hicklin DJ, Sauer CM, Kitajewski J, Sauer MV, Zimmermann RC 2009 Vascular endothelial growth factor receptor 2 (VEGFR-2) functions to promote uterine decidual angiogenesis during early pregnancy in the mouse. Endocrinology
37. Hess AP, Hirchenhain J, Schanz A, Talbi S, Hamilton AE, Giudice LC, Krussel JS 2006 Angiopoietin-1 and -2 mRNA and protein expression in mouse preimplantation embryos and uteri suggests a role in angiogenesis during implantation. Reprod Fertil Dev 18:509-516
38. Ema M, Taya S, Yokotani N, Sogawa K, Matsuda Y, Fujii-Kuriyama Y 1997 A novel bHLH-PAS factor with close sequence similarity to hypoxia-inducible factor 1alpha regulates the VEGF expression and is potentially involved in lung and vascular development. Proc Natl Acad Sci U S A 94:4273-4278
39. Li M, Wu Y, Caron KM 2008 Haploinsufficiency for adrenomedullin reduces pinopodes and diminishes uterine receptivity in mice. Biol Reprod 79:1169-1175
40. Li M, Yee D, Magnuson TR, Smithies O, Caron KM 2006 Reduced maternal expression of adrenomedullin disrupts fertility, placentation, and fetal growth in mice. J Clin Invest 116:2653-2662
41. Bilinski P, Roopenian D, Gossler A 1998 Maternal IL-11Ralpha function is required for normal decidua and fetoplacental development in mice. Genes Dev 12:2234-2243
42. Robb L, Li R, Hartley L, Nandurkar HH, Koentgen F, Begley CG 1998 Infertility in female mice lacking the receptor for interleukin 11 is due to a defective uterine response to implantation. Nat Med 4:303-308
43. Lim H, Paria BC, Das SK, Dinchuk JE, Langenbach R, Trzaskos JM, Dey SK 1997 Multiple female reproductive failures in cyclooxygenase 2-deficient mice. Cell 91:197-208
44. Chakraborty I, Das SK, Wang J, Dey SK 1996 Developmental expression of the cyclo-oxygenase-1 and cyclo-oxygenase-2 genes in the peri-implantation mouse uterus and their differential regulation by the blastocyst and ovarian steroids. J Mol Endocrinol 16:107-122
45. Chakrabarty A, Tranguch S, Daikoku T, Jensen K, Furneaux H, Dey SK 2007 MicroRNA regulation of cyclooxygenase-2 during embryo implantation. Proc Natl Acad Sci U S A 104:15144-15149
46. Collins MK, Tay CS, Erlebacher A 2009 Dendritic cell entrapment within the pregnant uterus inhibits immune surveillance of the maternal/fetal interface in mice. J Clin Invest 119:2062-2073
47. Plaks V, Birnberg T, Berkutzki T, Sela S, Benyashar A, Kalchenko V, Mor G, Keshet E, Dekel N, Neeman M, Jung S 2008 Uterine DCs are crucial for decidua formation during embryo implantation in mice. J Clin Invest
48. Matsumoto H, Ma WG, Daikoku T, Zhao X, Paria BC, Das SK, Trzaskos JM, Dey SK 2002 Cyclooxygenase-2 differentially directs uterine angiogenesis during implantation in mice. J Biol Chem 277:29260-29267

138
49. Bell SC, Jackson JA, Ashmore J, Zhu HH, Tseng L 1991 Regulation of insulin-like growth factor-binding protein-1 synthesis and secretion by progestin and relaxin in long term cultures of human endometrial stromal cells. J Clin Endocrinol Metab 72:1014-1024
50. Kim JJ, Taylor HS, Akbas GE, Foucher I, Trembleau A, Jaffe RC, Fazleabas AT, Unterman TG 2003 Regulation of insulin-like growth factor binding protein-1 promoter activity by FKHR and HOXA10 in primate endometrial cells. Biol Reprod 68:24-30
51. Daly DC, Maslar IA, Riddick DH 1983 Prolactin production during in vitro decidualization of proliferative endometrium. Am J Obstet Gynecol 145:672-678
52. Brosens JJ, Hayashi N, White JO 1999 Progesterone receptor regulates decidual prolactin expression in differentiating human endometrial stromal cells. Endocrinology 140:4809-4820
53. Christian M, Zhang X, Schneider-Merck T, Unterman TG, Gellersen B, White JO, Brosens JJ 2002 Cyclic AMP-induced forkhead transcription factor, FKHR, cooperates with CCAAT/enhancer-binding protein beta in differentiating human endometrial stromal cells. J Biol Chem 277:20825-20832
54. Dimitriadis E, Robb L, Salamonsen LA 2002 Interleukin 11 advances progesterone-induced decidualization of human endometrial stromal cells. Mol Hum Reprod 8:636-643
55. Sharkey AM, Day K, McPherson A, Malik S, Licence D, Smith SK, Charnock-Jones DS 2000 Vascular endothelial growth factor expression in human endometrium is regulated by hypoxia. J Clin Endocrinol Metab 85:402-409
56. McLachlan RI, Cohen NL, Dahl KD, Bremner WJ, Soules MR 1990 Serum inhibin levels during the periovulatory interval in normal women: relationships with sex steroid and gonadotrophin levels. Clin Endocrinol (Oxf) 32:39-48
57. Mote PA, Balleine RL, McGowan EM, Clarke CL 1999 Colocalization of progesterone receptors A and B by dual immunofluorescent histochemistry in human endometrium during the menstrual cycle. J Clin Endocrinol Metab 84:2963-2971
58. Jones RL, Salamonsen LA, Findlay JK 2002 Activin A promotes human endometrial stromal cell decidualization in vitro. J Clin Endocrinol Metab 87:4001-4004
59. Jones RL, Salamonsen LA, Zhao YC, Ethier JF, Drummond AE, Findlay JK 2002 Expression of activin receptors, follistatin and betaglycan by human endometrial stromal cells; consistent with a role for activins during decidualization. Mol Hum Reprod 8:363-374
60. Stoikos CJ, Harrison CA, Salamonsen LA, Dimitriadis E 2008 A distinct cohort of the TGFbeta superfamily members expressed in human endometrium regulate decidualization. Hum Reprod 23:1447-1456
61. Grinius L, Kessler C, Schroeder J, Handwerger S 2006 Forkhead transcription factor FOXO1A is critical for induction of human decidualization. J Endocrinol 189:179-187
62. Buzzio OL, Lu Z, Miller CD, Unterman TG, Kim JJ 2006 FOXO1A differentially regulates genes of decidualization. Endocrinology 147:3870-3876
63. Takano M, Lu Z, Goto T, Fusi L, Higham J, Francis J, Withey A, Hardt J, Cloke B, Stavropoulou AV, Ishihara O, Lam EW, Unterman TG, Brosens JJ, Kim JJ 2007 Transcriptional cross talk between the forkhead transcription factor forkhead box O1A and the progesterone receptor coordinates cell cycle regulation and differentiation in human endometrial stromal cells. Mol Endocrinol 21:2334-2349

139
64. Mak IY, Brosens JJ, Christian M, Hills FA, Chamley L, Regan L, White JO 2002 Regulated expression of signal transducer and activator of transcription, Stat5, and its enhancement of PRL expression in human endometrial stromal cells in vitro. J Clin Endocrinol Metab 87:2581-2588
65. Nagashima T, Maruyama T, Uchida H, Kajitani T, Arase T, Ono M, Oda H, Kagami M, Masuda H, Nishikawa S, Asada H, Yoshimura Y 2008 Activation of SRC kinase and phosphorylation of signal transducer and activator of transcription-5 are required for decidual transformation of human endometrial stromal cells. Endocrinology 149:1227-1234
66. Schneider-Merck T, Pohnke Y, Kempf R, Christian M, Brosens JJ, Gellersen B 2006 Physical interaction and mutual transrepression between CCAAT/enhancer-binding protein beta and the p53 tumor suppressor. J Biol Chem 281:269-278
67. Hu W, Feng Z, Teresky AK, Levine AJ 2007 p53 regulates maternal reproduction through LIF. Nature 450:721-724
68. Kang HJ, Feng Z, Sun Y, Atwal G, Murphy ME, Rebbeck TR, Rosenwaks Z, Levine AJ, Hu W 2009 Single-nucleotide polymorphisms in the p53 pathway regulate fertility in humans. Proc Natl Acad Sci U S A 106:9761-9766
69. Espinoza J, Romero R, Mee Kim Y, Kusanovic JP, Hassan S, Erez O, Gotsch F, Than NG, Papp Z, Jai Kim C 2006 Normal and abnormal transformation of the spiral arteries during pregnancy. J Perinat Med 34:447-458
70. Sugino N, Kashida S, Karube-Harada A, Takiguchi S, Kato H 2002 Expression of vascular endothelial growth factor (VEGF) and its receptors in human endometrium throughout the menstrual cycle and in early pregnancy. Reproduction 123:379-387
71. Fujimoto J, Hori M, Ichigo S, Tamaya T 1997 Ovarian steroids regulate the expression of basic fibroblast growth factor and its mRNA in fibroblasts derived from uterine endometrium. Ann Clin Biochem 34 ( Pt 1):91-96
72. Koga K, Osuga Y, Tsutsumi O, Yano T, Yoshino O, Takai Y, Matsumi H, Hiroi H, Kugu K, Momoeda M, Fujiwara T, Taketani Y 2001 Demonstration of angiogenin in human endometrium and its enhanced expression in endometrial tissues in the secretory phase and the decidua. J Clin Endocrinol Metab 86:5609-5614
73. Psychoyos A 1973 Hormonal control of ovoimplantation. Vitam Horm 31:201-256 74. Psychoyos A 1986 Uterine receptivity for nidation. Ann N Y Acad Sci 476:36-42 75. Curtis SW, Clark J, Myers P, Korach KS 1999 Disruption of estrogen signaling
does not prevent progesterone action in the estrogen receptor alpha knockout mouse uterus. Proc Natl Acad Sci U S A 96:3646-3651
76. Parr MB PE 1989 The implantation reaction. Wynn RM, (eds) Biology of the uterus Plenium Press, New York, 233-277, 1989
77. Lephart ED, Simpson ER 1991 Assay of aromatase activity. Methods Enzymol 206:477-483
78. Gibori G, Rodway R, Rothchild I 1977 The luteotrophic effect of estrogen in the rat: prevention by estradiol of the luteolytic effect of an antiserum to luteinizing hormone in the pregnant rat. Endocrinology 101:1683-1689
79. Gibori G, Sridaran R 1981 Sites of androgen and estradiol production in the second half of pregnancy in the rat. Biol Reprod 24:249-256
80. McCormack JT, Greenwald GS 1974 Progesterone and oestradiol-17beta concentrations in the peripheral plasma during pregnancy in the mouse. J Endocrinol 62:101-107
81. Peng L, Arensburg J, Orly J, Payne AH 2002 The murine 3beta-hydroxysteroid dehydrogenase (3beta-HSD) gene family: a postulated role for 3beta-HSD VI during early pregnancy. Mol Cell Endocrinol 187:213-221

140
82. Abbaszade IG, Arensburg J, Park CH, Kasa-Vubu JZ, Orly J, Payne AH 1997 Isolation of a new mouse 3beta-hydroxysteroid dehydrogenase isoform, 3beta-HSD VI, expressed during early pregnancy. Endocrinology 138:1392-1399
83. Ben-Zimra M, Koler M, Melamed-Book N, Arensburg J, Payne AH, Orly J 2002 Uterine and placental expression of steroidogenic genes during rodent pregnancy. Mol Cell Endocrinol 187:223-231
84. Arensburg J, Payne AH, Orly J 1999 Expression of steroidogenic genes in maternal and extraembryonic cells during early pregnancy in mice. Endocrinology 140:5220-5232
85. Simpson ER, Clyne C, Rubin G, Boon WC, Robertson K, Britt K, Speed C, Jones M 2002 Aromatase--a brief overview. Annu Rev Physiol 64:93-127
86. Fisher CR, Graves KH, Parlow AF, Simpson ER 1998 Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc Natl Acad Sci U S A 95:6965-6970
87. Toda K, Takeda K, Okada T, Akira S, Saibara T, Kaname T, Yamamura K, Onishi S, Shizuta Y 2001 Targeted disruption of the aromatase P450 gene (Cyp19) in mice and their ovarian and uterine responses to 17beta-oestradiol. J Endocrinol 170:99-111
88. Mary J Laws RNT, Neil Sidell, Francesco J DeMayo, John P Lydon, David E Gutstein, Milan K Bagchi, and Indrani C Bagchi 2008 Gap Junctions Between Uterine Stromal Cells Play a Critical Role in Pregnancy-Associated Neovascularization and Embryo Survival. Development (under review)
89. Grummer R, Chwalisz K, Mulholland J, Traub O, Winterhager E 1994 Regulation of connexin26 and connexin43 expression in rat endometrium by ovarian steroid hormones. Biol Reprod 51:1109-1116
90. Hyder SM, Stancel GM 1999 Regulation of angiogenic growth factors in the female reproductive tract by estrogens and progestins. Mol Endocrinol 13:806-811
91. Folkman J 1995 Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1:27-31
92. Tsai MJ, O'Malley BW 1994 Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem 63:451-486
93. Young MR, Colburn NH 2006 Fra-1 a target for cancer prevention or intervention. Gene 379:1-11
94. Weisz A, Rosales R 1990 Identification of an estrogen response element upstream of the human c-fos gene that binds the estrogen receptor and the AP-1 transcription factor. Nucleic Acids Res 18:5097-5106
95. Frost JA, Geppert TD, Cobb MH, Feramisco JR 1994 A requirement for extracellular signal-regulated kinase (ERK) function in the activation of AP-1 by Ha-Ras, phorbol 12-myristate 13-acetate, and serum. Proc Natl Acad Sci U S A 91:3844-3848
96. Chinenov Y, Kerppola TK 2001 Close encounters of many kinds: Fos-Jun interactions that mediate transcription regulatory specificity. Oncogene 20:2438-2452
97. Adiseshaiah P, Lindner DJ, Kalvakolanu DV, Reddy SP 2007 FRA-1 proto-oncogene induces lung epithelial cell invasion and anchorage-independent growth in vitro, but is insufficient to promote tumor growth in vivo. Cancer Res 67:6204-6211
98. Cheon YP, DeMayo FJ, Bagchi MK, Bagchi IC 2004 Induction of cytotoxic T-lymphocyte antigen-2beta, a cysteine protease inhibitor in decidua: a potential regulator of embryo implantation. J Biol Chem 279:10357-10363
99. Li Q, Kannan A, Wang W, Demayo FJ, Taylor RN, Bagchi MK, Bagchi IC 2007 Bone morphogenetic protein 2 functions via a conserved signaling pathway involving

141
Wnt4 to regulate uterine decidualization in the mouse and the human. J Biol Chem 282:31725-31732
100. Li Q, Cheon YP, Kannan A, Shanker S, Bagchi IC, Bagchi MK 2004 A novel pathway involving progesterone receptor, 12/15-lipoxygenase-derived eicosanoids, and peroxisome proliferator-activated receptor gamma regulates implantation in mice. J Biol Chem 279:11570-11581
101. Verde P, Casalino L, Talotta F, Yaniv M, Weitzman JB 2007 Deciphering AP-1 function in tumorigenesis: fra-ternizing on target promoters. Cell Cycle 6:2633-2639
102. Young MR, Nair R, Bucheimer N, Tulsian P, Brown N, Chapp C, Hsu TC, Colburn NH 2002 Transactivation of Fra-1 and consequent activation of AP-1 occur extracellular signal-regulated kinase dependently. Mol Cell Biol 22:587-598
103. Casalino L, De Cesare D, Verde P 2003 Accumulation of Fra-1 in ras-transformed cells depends on both transcriptional autoregulation and MEK-dependent posttranslational stabilization. Mol Cell Biol 23:4401-4415
104. Basbous J, Chalbos D, Hipskind R, Jariel-Encontre I, Piechaczyk M 2007 Ubiquitin-independent proteasomal degradation of Fra-1 is antagonized by Erk1/2 pathway-mediated phosphorylation of a unique C-terminal destabilizer. Mol Cell Biol 27:3936-3950
105. Nelson KG, Takahashi T, Bossert NL, Walmer DK, McLachlan JA 1991 Epidermal growth factor replaces estrogen in the stimulation of female genital-tract growth and differentiation. Proc Natl Acad Sci U S A 88:21-25
106. Thienel T, Chwalisz K, Winterhager E 2002 Expression of MAPkinases (Erk1/2) during decidualization in the rat: regulation by progesterone and nitric oxide. Mol Hum Reprod 8:465-474
107. Fleischmann A, Hafezi F, Elliott C, Reme CE, Ruther U, Wagner EF 2000 Fra-1 replaces c-Fos-dependent functions in mice. Genes Dev 14:2695-2700
108. Jochum W, David JP, Elliott C, Wutz A, Plenk H, Jr., Matsuo K, Wagner EF 2000 Increased bone formation and osteosclerosis in mice overexpressing the transcription factor Fra-1. Nat Med 6:980-984
109. Eferl R, Hoebertz A, Schilling AF, Rath M, Karreth F, Kenner L, Amling M, Wagner EF 2004 The Fos-related antigen Fra-1 is an activator of bone matrix formation. Embo J 23:2789-2799
110. Baker DJ, Nagy F, Nieder GL 1992 Localization of c-fos-like proteins in the mouse endometrium during the peri-implantation period. Biol Reprod 47:492-501
111. Mitchell JA, Lye SJ 2002 Differential expression of activator protein-1 transcription factors in pregnant rat myometrium. Biol Reprod 67:240-246
112. Halder JB, Zhao X, Soker S, Paria BC, Klagsbrun M, Das SK, Dey SK 2000 Differential expression of VEGF isoforms and VEGF(164)-specific receptor neuropilin-1 in the mouse uterus suggests a role for VEGF(164) in vascular permeability and angiogenesis during implantation. Genesis 26:213-224
113. Belguise K, Kersual N, Galtier F, Chalbos D 2005 FRA-1 expression level regulates proliferation and invasiveness of breast cancer cells. Oncogene 24:1434-1444
114. Grewal S, Carver JG, Ridley AJ, Mardon HJ 2008 Implantation of the human embryo requires Rac1-dependent endometrial stromal cell migration. Proc Natl Acad Sci U S A 105:16189-16194
115. Alexander CM, Hansell EJ, Behrendtsen O, Flannery ML, Kishnani NS, Hawkes SP, Werb Z 1996 Expression and function of matrix metalloproteinases and their inhibitors at the maternal-embryonic boundary during mouse embryo implantation. Development 122:1723-1736

142
116. Matsumoto H, Daikoku T, Wang H, Sato E, Dey SK 2004 Differential expression of ezrin/radixin/moesin (ERM) and ERM-associated adhesion molecules in the blastocyst and uterus suggests their functions during implantation. Biol Reprod 70:729-736
117. Ramos-Nino ME, Scapoli L, Martinelli M, Land S, Mossman BT 2003 Microarray analysis and RNA silencing link fra-1 to cd44 and c-met expression in mesothelioma. Cancer Res 63:3539-3545
118. Ramos-Nino ME, Blumen SR, Pass H, Mossman BT 2007 Fra-1 governs cell migration via modulation of CD44 expression in human mesotheliomas. Mol Cancer 6:81
119. Song Y, Qian L, Song S, Chen L, Zhang Y, Yuan G, Zhang H, Xia Q, Hu M, Yu M, Shi M, Jiang Z, Guo N 2008 Fra-1 and Stat3 synergistically regulate activation of human MMP-9 gene. Mol Immunol 45:137-143
120. Weitlauf HM 1988 The physiology of reproduction, eds Knobil, E & Neil, J (Raven, New York), pp231-262, 1988
121. Fraser HM, Wulff C 2003 Angiogenesis in the corpus luteum. Reprod Biol Endocrinol 1:88
122. Rockwell LC, Pillai S, Olson CE, Koos RD 2002 Inhibition of vascular endothelial growth factor/vascular permeability factor action blocks estrogen-induced uterine edema and implantation in rodents. Biol Reprod 67:1804-1810
123. Marelli-Berg FM, Peek E, Lidington EA, Stauss HJ, Lechler RI 2000 Isolation of endothelial cells from murine tissue. J Immunol Methods 244:205-215
124. Hellstrom M, Phng LK, Hofmann JJ, Wallgard E, Coultas L, Lindblom P, Alva J, Nilsson AK, Karlsson L, Gaiano N, Yoon K, Rossant J, Iruela-Arispe ML, Kalen M, Gerhardt H, Betsholtz C 2007 Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature 445:776-780
125. Scharpfenecker M, Fiedler U, Reiss Y, Augustin HG 2005 The Tie-2 ligand angiopoietin-2 destabilizes quiescent endothelium through an internal autocrine loop mechanism. J Cell Sci 118:771-780
126. Bedell VM, Yeo SY, Park KW, Chung J, Seth P, Shivalingappa V, Zhao J, Obara T, Sukhatme VP, Drummond IA, Li DY, Ramchandran R 2005 roundabout4 is essential for angiogenesis in vivo. Proc Natl Acad Sci U S A 102:6373-6378
127. Kruger RP, Aurandt J, Guan KL 2005 Semaphorins command cells to move. Nat Rev Mol Cell Biol 6:789-800
128. Neufeld G, Shraga-Heled N, Lange T, Guttmann-Raviv N, Herzog Y, Kessler O 2005 Semaphorins in cancer. Front Biosci 10:751-760
129. Kuijper S, Turner CJ, Adams RH 2007 Regulation of angiogenesis by Eph-ephrin interactions. Trends Cardiovasc Med 17:145-151
130. Gale NW, Thurston G, Hackett SF, Renard R, Wang Q, McClain J, Martin C, Witte C, Witte MH, Jackson D, Suri C, Campochiaro PA, Wiegand SJ, Yancopoulos GD 2002 Angiopoietin-2 is required for postnatal angiogenesis and lymphatic patterning, and only the latter role is rescued by Angiopoietin-1. Dev Cell 3:411-423
131. Lee CG, Heijn M, di Tomaso E, Griffon-Etienne G, Ancukiewicz M, Koike C, Park KR, Ferrara N, Jain RK, Suit HD, Boucher Y 2000 Anti-Vascular endothelial growth factor treatment augments tumor radiation response under normoxic or hypoxic conditions. Cancer Res 60:5565-5570
132. Laws MJ, Taylor RN, Sidell N, Demayo FJ, Lydon JP, Gutstein DE, Bagchi MK, Bagchi IC 2008 Gap junction communication between uterine stromal cells plays a

143
critical role in pregnancy-associated neovascularization and embryo survival. Development
133. Liebner S, Cavallaro U, Dejana E 2006 The multiple languages of endothelial cell-to-cell communication. Arterioscler Thromb Vasc Biol 26:1431-1438
134. Cavallaro U, Liebner S, Dejana E 2006 Endothelial cadherins and tumor angiogenesis. Exp Cell Res 312:659-667
135. Wallez Y, Huber P 2008 Endothelial adherens and tight junctions in vascular homeostasis, inflammation and angiogenesis. Biochim Biophys Acta 1778:794-809
136. Rodewald M, Herr D, Fraser HM, Hack G, Kreienberg R, Wulff C 2007 Regulation of tight junction proteins occludin and claudin 5 in the primate ovary during the ovulatory cycle and after inhibition of vascular endothelial growth factor. Mol Hum Reprod 13:781-789
137. Brosens JJ, Gellersen B 2006 Death or survival--progesterone-dependent cell fate decisions in the human endometrial stroma. J Mol Endocrinol 36:389-398
138. Irwin JC, Kirk D, King RJ, Quigley MM, Gwatkin RB 1989 Hormonal regulation of human endometrial stromal cells in culture: an in vitro model for decidualization. Fertil Steril 52:761-768
139. Brar AK, Frank GR, Kessler CA, Cedars MI, Handwerger S 1997 Progesterone-dependent decidualization of the human endometrium is mediated by cAMP. Endocrine 6:301-307
140. Neulen J, Hartmann C, Breckwoldt M 1987 Aromatase activity in monolayer cell cultures of human endometrium. Gynecol Endocrinol 1:339-343
141. Tseng L 1984 Effect of estradiol and progesterone on human endometrial aromatase activity in primary cell culture. Endocrinology 115:833-835
142. Tseng L, Malbon CC, Lane B, Kaplan C, Mazella J, Dahler H, Tseng A 1987 Progestin-dependent effect of forskolin on human endometrial aromatase activity. Hum Reprod 2:371-377
143. Tseng L, Mazella J, Chen GA 1987 Effect of relaxin on aromatase activity in human endometrial stromal cells. Endocrinology 120:2220-2226
144. Tseng L, Mazella J, Funt MI, Mann WJ, Stone ML 1984 Preliminary studies of aromatase in human neoplastic endometrium. Obstet Gynecol 63:150-154
145. Tseng L, Mazella J, Sun B 1986 Modulation of aromatase activity in human endometrial stromal cells by steroids, tamoxifen and RU 486. Endocrinology 118:1312-1318
146. Yamamoto T, Shiroshita K, Kitawaki J, Okada H 1989 The inductive effects of progestogens on aromatase activity in stromal cells of human uterine endometrium. J Endocrinol Invest 12:201-204
147. Ryan IP, Schriock ED, Taylor RN 1994 Isolation, characterization, and comparison of human endometrial and endometriosis cells in vitro. J Clin Endocrinol Metab 78:642-649
148. Giudice LC, Telles TL, Lobo S, Kao L 2002 The molecular basis for implantation failure in endometriosis: on the road to discovery. Ann N Y Acad Sci 955:252-264; discussion 293-255, 396-406
149. Aghajanova L, Hamilton A, Kwintkiewicz J, Vo KC, Giudice LC 2009 Steroidogenic enzyme and key decidualization marker dysregulation in endometrial stromal cells from women with versus without endometriosis. Biol Reprod 80:105-114
150. Bulun SE, Mahendroo MS, Simpson ER 1993 Polymerase chain reaction amplification fails to detect aromatase cytochrome P450 transcripts in normal human endometrium or decidua. J Clin Endocrinol Metab 76:1458-1463

144
151. Attar E, Tokunaga H, Imir G, Yilmaz MB, Redwine D, Putman M, Gurates B, Attar R, Yaegashi N, Hales DB, Bulun SE 2009 Prostaglandin E2 via steroidogenic factor-1 coordinately regulates transcription of steroidogenic genes necessary for estrogen synthesis in endometriosis. J Clin Endocrinol Metab 94:623-631
152. Giudice LC, Kao LC 2004 Endometriosis. Lancet 364:1789-1799 153. Bulun SE, Utsunomiya H, Lin Z, Yin P, Cheng YH, Pavone ME, Tokunaga H,
Trukhacheva E, Attar E, Gurates B, Milad MP, Confino E, Su E, Reierstad S, Xue Q 2009 Steroidogenic factor-1 and endometriosis. Mol Cell Endocrinol 300:104-108
154. Bulun SE, Imir G, Utsunomiya H, Thung S, Gurates B, Tamura M, Lin Z 2005 Aromatase in endometriosis and uterine leiomyomata. J Steroid Biochem Mol Biol 95:57-62
155. Bulun SE, Yang S, Fang Z, Gurates B, Tamura M, Zhou J, Sebastian S 2001 Role of aromatase in endometrial disease. J Steroid Biochem Mol Biol 79:19-25
156. Lessey BA, Killam AP, Metzger DA, Haney AF, Greene GL, McCarty KS, Jr. 1988 Immunohistochemical analysis of human uterine estrogen and progesterone receptors throughout the menstrual cycle. J Clin Endocrinol Metab 67:334-340
157. Mylonas I, Jeschke U, Shabani N, Kuhn C, Balle A, Kriegel S, Kupka MS, Friese K 2004 Immunohistochemical analysis of estrogen receptor alpha, estrogen receptor beta and progesterone receptor in normal human endometrium. Acta Histochem 106:245-252
158. Okada H, Tsutsumi A, Imai M, Nakajima T, Yasuda K, Kanzaki H 2009 Estrogen and selective estrogen receptor modulators regulate vascular endothelial growth factor and soluble vascular endothelial growth factor receptor 1 in human endometrial stromal cells. Fertil Steril
159. Critchley HO, Saunders PT 2009 Hormone receptor dynamics in a receptive human endometrium. Reprod Sci 16:191-199
160. Stocco C 2008 Aromatase expression in the ovary: hormonal and molecular regulation. Steroids 73:473-487
161. Zhou J, Gurates B, Yang S, Sebastian S, Bulun SE 2001 Malignant breast epithelial cells stimulate aromatase expression via promoter II in human adipose fibroblasts: an epithelial-stromal interaction in breast tumors mediated by CCAAT/enhancer binding protein beta. Cancer Res 61:2328-2334

145
Appendix

146
A1. ER critically regulates stromal cell decidualization
Stromal cells isolated from day 4 pregnant, pre-implantation mouse uterus were cultured and
differentiation was induced by adding P. The cells were then transfected with mouse ER
specific siRNA (sense sequence GGGAGAAUGUUGAAGCACAtt and anti sense sequence
UGUGCUUCAACAUUCUCCCtc) and the scrambled (NC) siRNA. Cells were cultured for
an additional 48 hrs, and subjected to RNA isolation or fixed with 4 % formaldehyde to
monitor the expression of ER (Fig A-1). Silencing of ER indicated almost 80%
knockdown of the gene both at the transcript and protein levels. Phallodin staining,
demarcating the F-actin filament of the cells revealed a significant alteration of the cellular
architecture when treated with ER siRNA compared to the scrambled siRNA transfected
cells (Fig A-2). RNA isolated from ER siRNA transfected stromal cells was subjected to q-
PCR analysis to monitor the expression of well known targets of stromal differentiation. As
shown in Fig A-3 knockdown of ER expression, significantly down regulated the
expression of Alkp, a known bio-marker of decidualization. In addition, the expression of
several other important regulators of stromal cell differentiation like PR, CEBP, CX-43,
BMP2 and CYR-61, were also significantly inhibited while the expression of GAPDH
remained unaltered. This experiment indicated that ER signaling plays a major role in
mediating stromal differentiation.
Figure A-1: Silencing of ER expression in mouse stromal cells siRNA specific to mouse ER and scrambled (NC) was transfected to primary stromal cells undergoing differentiation in the presence of steroid hormones. RNA isolated after 48hrs of transfection was subjected to qPCR analysis to monitor the gene expression of ER (left panel). The efficient silencing of the target gene was further confirmed by immunostaining siRNA transfected cells with anti ER antibody (right panel).
NC siRNA-20nM ER siRNA-20nM
10x
20x0
0.2
0.4
0.6
0.8
1
1.2
NC siRNA ER siRNA
Re
lati
ve
lev
el o
f e
xp
res
sio
n
ER alpha
Figure A-1
Q-PCR
Immunocytochemistry-ER alphaFigure A-1
*

147
Figure A-2: Cellular architecture of cells transfected with ER siRNA
Primary stromal cell cultures were transfected with ER or scrambled siRNA. Cells were fixed and stained for F-actin expression. Control siRNA treated cells revealed characteristic decidual morphology (left panel) which was significantly impaired after ER silencing (right panel).
Figure A-3: ER critically modulates stromal cell decidualization siRNA treated cells were subjected to RNA isolation and was subjected to qPCR analysis to monitor the expression of several molecules involved in decidualization. Silencing of ER significantly impaired the expression of these genes. (n=3, p<0.05)
NC siRNA-20nM ER siRNA-20nM
Figure A-2
F-actin
0
0.2
0.4
0.6
0.8
1
1.2
1.4
ER alpha PR Alkp CEBPb BMP2 Cnx 43 CYR 61 GAPDH
Re
lati
ve
lev
el o
f e
xp
res
sio
n
NC siRNA
ER siRNA
Figure A-4Q-PCR
Figure A-3
Figure A-2
*
**
**

148
A-2. C/CAAT enhancer binding protein beta (C/EBP regulates P450 aromatase
expression in differentiating stromal cells
The pivotal contribution of aromatase derived E in the decidual phase has been the key focus
of this dissertation research. However, the regulator of aromatase expression in the uterus
during implantation is currently unknown. In the gonads it has been shown that cAMP and
gonadotrophins regulate aromatase expression (160). In human adipose fibroblasts, aromatase
expression is up-regulated by the binding of C/EBP to the promoter (161). The transcription
factor C/EBP is a critical mediator of steroid hormone action in the decidual uterus. It
belongs to the family of basic leucine zipper domain proteins that is important in regulating
cellular processes like proliferation, differentitation, inflammation and apoptosis. C/EBP
null mouse do not undergo implantation and exhibits a severe defect in endometrial cell
proliferation and differentiation of stromal cells to decidual cells. To investigate whether
C/EBP in the mouse stromal cells were involved in regulating P450 aromatase expression,
the transcriptional activity of C/EBP was impaired by the administration of a dominant
negative C/EBP(ACEBP) in primary mouse stromal cell cultures. ACEBP heterodimerizes
with the endogenous C/EBP and prevents its DNA binding, thereby disrupting its
transcriptional activity. As shown in Figure A-3, cells transduced with the adenovirus
expressing ACEBP exhibited almost 80% reduction in aromatase expression compared to the
GFP expressing adenovirus, indicating that aromatase expression in the decidual uterus is
regulated by this C/CAAT box binding transcription factor. The observation was further
confirmed by analyzing the gene expression level in the wildtype and CEBP KO mouse
uterus (data not shown). As anticipated, aromatase expression was significantly inhibited in
the null uterus. Currently, studies are undergoing to address the functional promoter region
and CEBP binding sites in the aromatase promoter during early mouse pregnancy.
Figure A-4
*

149
Figure A-4: Aromatase expression in the differentiating stromal cells is regulated by C/EBP Primary stromal cells isolated from day 4 pre-implantation mouse uterus were cultured in the presence of P to induce differentiation. The functional activity of C/EBP was impaired by the administration of a dominant negative CEBP. P450 aromatase expression was significantly downregulated in the ACEBP transduced cells, while the expression of CEBP remained unaltered. (n=3, p<0.05)

150
A.3.Generation of mouse ERG-1 plasmid
The full length ERG-1 or Itmap-1 and the truncated ERG-1 cDNA excluding the membrane
spanning domain (NCBI accession no: U69699) was PCR amplified from pregnant day 1
mouse uterus mRNA using Pfu DNA polymerase (Stratagene), with the following conditions:
95°C for 5 min, 1 cycle; 94°C for 30 s, 61°C for 120 s, 72°C for 120 s for 30 cycles, followed
by 1 cycle at 72°C for 10 min. The full-length cDNA construct was gel extracted (Qiagen Gel
Extraction Kit), followed by cloning to the pPCR-Script Amp SK+ cloning vector (pPCR-
Script Amp cloning Kit, Promega). Positive colonies were identified by blue-white selection
subjected to plasmid isolation. For the expression of FLAG epitope-tagged mouse ERG-1,
DNA encoding the full-length ERG-1 was cloned into the NotI and BglII sites of pCMV-
FLAG-MAT2 vector (Sigma Algrich) upstream of a FLAG-MAT peptide sequence. The
clones obtained from bacterial transformation were confirmed by restriction enzyme
digestion and DNA sequencing. Confirmed sequences were then further purified using the
Endo Free Maxi Kit (Qiagen) to get transfection-quality cDNA. The fusion protein was
expressed in mammalian HEK-293 cells, denatured in SDS-sample buffer, resolved on an
SDS-PAGE, stained with Coomassie Blue, blotted with anti-FLAG M2 antibody and
visualized by autoradiography.
Figure: A-5: Map of ERG-1 clone
ERG-1
Figure A-4Figure A-5

151
A.4. Primer Sequences
Table A-1. Mouse primer sequences
Mouse Primers Primer Name Sequence 36B4-F 5'- CATCACCACGAAAATCTCCA 36B4-R 5'- TTGTCAAACACCTGCTGGAT Adrmdln Fwd 5'- ACATTCGTGTCAAACGCTACC Adrmdln Rev 5'- ATGCCGTCCTTGTCTTTGTCT Caveolin-F 5'- GAGTACCAACAACCTCAACTGC Caveolin-R 5'- GTTAAGAGTGAGGACAGCAACC CD44 Fwd 5'- GAACAAGGAACCATCAGAGACC CD44 Rev 5'- AATGGCGTAGGGCACTACAC cebpb asense 5'- CTGCTCCACCTTCTTCTG cebpb sense 5'- AGCGACGAGTACAAGATG c-Fos-Fwd 5'- ACAGTTGGACCCAAGACAAAC c Fos-Rev 5'- TGAAACTGGCTCCTACAGCA Claudin5 Fwd 5'- ATGCAGTGCAAGGTGTATGAA Claudin5 Rev 5'- AGGTAACAAAGAGTGCCACCA COUP-TF-Fwd 5'- CGG AGG AAC CTG AGC TAC AC -3' COUP-TF-rev 5'- CAG GTA CGA GTG GCA GTT GA -3' Cx43 #1 5'- CTATCGTGGATCAGCGACCTTC Cx43 #2 5'- CACGGGAACGAAATGAACACC Endothelin-1 Fwd 5'- ACTTCTGCCACCTGGACATC Endothelin-1 Rev 5'- GGTGAGCGCACTGACATCTA erk1-fwd 5'- TAAAAGCCAGCAGCTGAGCAA erk1-rev 5'- AAGCAGATTGGAAGGCTTCA Fra-1F 5'- CAAGCATCGACAGCAGCA Fra-1R 5'- AGAAAGGGAGATGCAAGGCA Fra2- Rev 5'- CACAGGGTTACAGGGCTAGAA Fra2-Fwd 5'- ATCACTCCCGGCACTTCAAA FraKpn1R 5'- CCAGCCGGTACCTCACAAAGCCAGGAGTGT FraXba1F 5'- CCAGCCTCTAGAATGTACCGAGACTACGGG Gcebp as 5'- GCGCAGGGCGAACGGGAAACCG Gcebp sen 5'- AGCCCCTACCTGGAGCCGCTCGCG h-arom PII Fwd 5'- GAGGAGGAGGAGGAAAAAGGA h-arom PII Rev 5'- AAGGCTGGATGGTTTCTGGT ITF-2-Fwd 5'- CTC CAA TCC TTC AAC TCC TGT G -3' ITF-2-Rev 5'- GTC CAG TCT TTC CAA ACG GTC T -3' JunB- Fwd 5'- ACGACCTGCACAAGATGAACC JunB-Rev 5'- TATGAGCTCCCAGTCCCGA LRH-1F 5'- ACAGAAGTCGCGTTCAACAA LRH-1R 5'- GAAGTAGCTGTCCGAATTTCTCA ltmapF 5'- CTGAGGACCCTGCGAAGAT ltmapR 5'- AGGCCTTTCTTGGGGTCCA m-17bHSD1 F 5'- TTGGGACCATTCGGATGCT m-17bHSD1 R 5'- GCCCACCAGCTTTTCATAGAA m-17bHSD2 F 5'- TGGAGTGCTGGACAAAGAGG

152
m-17bHSD2 R 5'- AGTTCACGGCCATGCATTTT Maromatase asen2 5'- AGCTCTGCGCATGACCAAGT Maromatase sen1 5'- TAACATGCTCTTCCTGGGGAT Maromatase asen 5'- CACGCCACCTACTCGATAAAA Maromatase sense 5'- TGAACATCGGAAGAATGCAC MMP-13 fwd 5'- CCCTATCCCTTGATGCCATT MMP-13 rev 5'- TGGGATAACCTTCCAGAATGT MMP-2 fwd 5'- CCCTCAAGAAGATGCAGAAGT MMP-2 rev 5'- TGTGATCTGGTTCTTGTCCC MMP-7 fwd 5'- ATGCAGAAGTTCTTTGGCCTG MMP-7 rev 5'- CTCTCCTTGCGAAGCCAATT MMP-9 fwd 5'- TCTTCCCCTTCGTCTTCCTT MMP-9 rev 5'- TATACAGCGGGTACATGAGCG mStarF 5'- TCACAAAAGGGGACAAAACG mStarR 5'- TGTCACCATTTGCAGTGTTG mus Cyr61 Fwd 5'- ATTAAGGACTCCCTGGACGA mus Cyr61 Rev 5'- ACGATGCATTTCTGGCCATG PDGF-I Reverse 5' AGGTATGATGGCAGAGTCATCC 3' PDGF-II Forward 5' TGCGATCTGAACTCACAGTG 3' SCC-F 5'- AGTCCCAGGAACACAGGAAA SCC-R 5'- CATTCCTTGCGGGTTTTG SF-1F 5'- CTCCAAAAGACTCCTTGCATT SF-1R 5'- TGATGGAGGAAGGAATGGTT erg 570 reverse 5'- GCC TGG ATC CAA CTT GAT GT -3' FraflF 5'- CAGGGCATGTACCGAGACTA FraflR 5'- CCTCACAAAGCCAGGAGTGTA Gel shift Assay oligo AP 2 5'- TTCCGGCTGACTCATCAAGCG AP 1 5'- CGCTTGATGAGTCAGCCGGAA AP 5- Bio 2 TTCCGGCTGACTCATCAAGCG AP 5- Bio 1 CGCTTGATGAGTCAGCCGGAA AP 3- Bio 2 TTCCGGCTGACTCATCAAGCG AP 3- Bio 1 CGCTTGATGAGTCAGCCGGAA
Chip assay primers Pgr ERE Rev 5'-TCCTCGCACCCGTAAATACT Pgr ERE Fwd 5'-GAATTCCAACGCCAGAGATT ADM ERE Fwd 5'-ATCCTCACGTTTATGATGGA ADM ERE rev 5'-CGGATTTCGTAATAAGGGCA RAMP ERE Rev 5'-CTGTGACAGCAGGAGGACAG RAMP ERE Fwd 5'-AGAGTGTACGTGTGGACAGG Pgr distal Rev 5'-ATCACCAGGGAGGTGCTACA Pgr distal Fwd 5'-ACTGTCCAGAATGCCTCCAC Fra 171 Rev 5'-ATTTGTGAACTGGGGGCA Fra 171 Fwd 5'-CCATCTCTGCAGCCTTGATTT Fra 323 Fwd 5'-TGGATTCACAGTGGATGGTCA

153
Fra 323 Rev 5'-GGCTCTAGGGCTCTTTCCTTA Fra 592 Rev 5'-TCTCCTTCCTGAGTCTGGGGT Fra 592 Fwd 5'-GGAAAGAGCCCTAGAGCCCTA Fra 886 Fwd 5'-CAGAGAAAGAGTTTTGGGACC Fra 886 Rev 5'-TGAGGCTGACAGGTATTGAAA Fra 1006 Fwd 5'-TTTCAATACCTGTCAGCCTCA Fra 1006 Rev 5'-ACTATGTGTGTCACCAGCCCA Fra 1842 Fwd 5'-TCACTTCCCAGCTTTCAGACA Fra 1842 Rev 5'-ACGCACCTTGCCGAAGTT
Table A-2: Human Primer sequences
Primer name Sequence 17bHSD1 Fwd AAGGGACTCAAAATCCGTGGC 17bHSD1 Rev AGCATCCGCACAGTCCCTACT 17bHSD2 Fwd CTTGGCCATGCTTTGTGCAA 17bHSD2 Rev TGCTGCAACCTTGCTGTAAGC h-17a lyase F CCAATCTCAGCTCACTGCAA h-17a lyase R GATCGAGACCATCCTGGCTA h36B4 F GTGTTCGACAATGGCAGCAT h36B4 R GACACCCTCCAGGAAGCGA h-3HSD F TATGGGGAAGGAAGCCGATT h-3HSD R CAGGCCACATTGCCAACATA h-Aromatase-F CCTGAATATTGGAAGGATGCA h-Aromatase-R TACTTTCCTGCACAGCCACG hCYCLIN D1 asense AAACCTTCCGGTGTGAAACA hCYCLIN D1 sense TTGCATTCTCACATTGCCAG h-decorin fwd TGGAGCATTTACACCTTTGG h-decorin rev TGATCTCATTCTCATGGGCA hFOXO1-asense TTGGGTCAGGCGGTTCA hFOXO1-sense TGGACATGCTCAGCAGACATC hFra1Fwd AAGGAACTGACCGACTTCCTG hFra1Rev GCTTCCAGCACCAGCTCTA hMMP2-asense AAGACACAGGGGGCACCTT hMMP2-sense AAGGATGGCAAGTACGGCTT h-PR Fwd TGTATTTGTGCGTGTGGGTG h-PR Rev TACAGCCCATTCCCAGGAAG h-scc F GTGATGACCTGTTCCGCTTT h-scc R AACAGGTCTGGGGGAAGGT h-StAR F CCCAATGTCAAGGAGATCA h-StAR R ACAGCGCACGCTCACAAA IGFBP-1 sense GTAGACGCACCAGCAGAG IGFBP-1asense CCAAACTGCAACAAGAATG PRL antisense GGGCTTGCTCCTTGTCTTC PRL sense CTACATCCATAACCTCTCCTCAG















![Jayasree Das arXiv:1507.06733v1 [physics.plasm-ph] 24 Jul 2015 · Jayasree Das Department of Mathematics, Chittaranjan College, 8A Beniatola Lane, Kolkata - 700 009, India. Anup Bandyopadhyay∗](https://static.documents.pub/doc/80x56/5f94c71b87c031600721a5d6/jayasree-das-arxiv150706733v1-24-jul-2015-jayasree-das-department-of-mathematics.jpg)
![MyCoRe - Eine Software für verschiedene Repositorien · Das Konzept von MyCoRe Ein Software-Kern – viele Anwendungen = My Core['maik r]ɔː = My Content Repository](https://static.documents.pub/doc/80x56/5e20b24afea629504c1f0c65/mycore-eine-software-fr-verschiedene-repositorien-das-konzept-von-mycore-ein.jpg)
