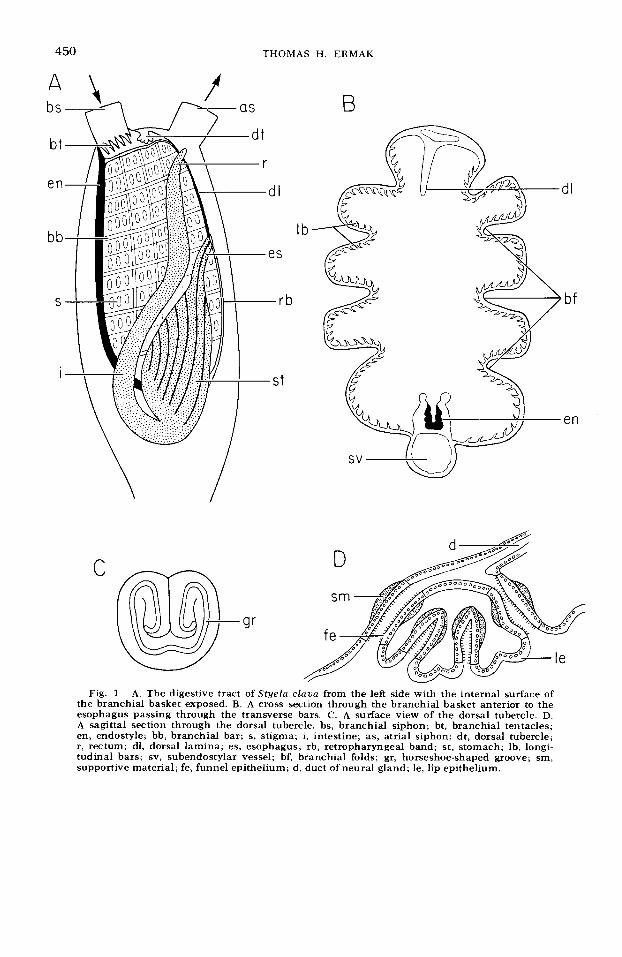
Cell Proliferation in the Digestive Tract of Styela clava (Urochordata: Ascidiacea) as Revealed by Autoradiography with Tritiated Thymidi ne THOMAS H. ERMAK' Scripps lnstitution of Oceanography, La Jolla, Californiu 92037 ABSTRACT The DNA-synthesizing cells in the digestive tract of Styela clava were labeled with tritiated thymidine and detected at increasing time intervals with autoradiography. Most of the epithelia consist of renewing cell populations. These are the dorsal tubercle, dorsal lamina, branchial bars, zone 1 of the endo- style, stigmata, esophagus, stomach, intestine, and rectum. Expanding popula- tions are the branchid tentacles, unspeciafized pharyngeal cells, and pyloric tubules. In the renewing populations, cell proliferation occurs in restricted ger- minal zones of pseudostratified cells. In the postbranchial digestive tract, these germinal zones usually occur at the bases of folds or grooves. Germinal cell nu- clei migrate into the cell apex and undergo mitosis there. With time, germinal cells migrate as a band of labeled cells over the underlying connective tissue into non-proliferating mature zones of ciliated, secretory, or absorptive cells. Aging mature cells are presumably extruded into the gut lumen, maintaining the system in a steady state. Transit times are about ten days for the stigmata; two and a half weeks for the dorsal tubercle, esophageal mucous population, stomach crest population, and stomach groove population; five weeks for the intestine and rectum; and several months for zone 1 of the endostyle and esoph- ageal band population. To date, most of the work on gut cell proliferation employing tritiated thymidine and autoradiography has been confined to the renewing epithelia of mammals (Leblond and Messier, '58; Messier, '60; Leblond et al., '64). Comparable work has been done on relatively few non-mamma- lian vertebrates and invertebrates (Davis and Burnett, '64; Anderson; '65; Holland, '64). The sparsity of such information on ascidians is especially unfortunate because of their vertebrate affinities. Previous work on ascidian gut cell kinetics is limited to a cell formation test (as defined by Leblond et al., '59) for Halocynthia sp. (Mochan et al., '71). However, from anatomical evi- dence alone, the replacement of gut epi- thelial cells has been proposed in several ascidians (Aros and Viragh, '69; Fouque, '53; Relini-Orsi, '68; Thomas, '70). Therefore, the purpose of the present investigation is to elucidate cell prolifera- tion patterns in the digestive tract of the ascidian Styela claua, a large solitary as- cidian found in bays along the California coast. DNA synthesizing cells about to en- ter mitosis were labeled with tritiated thy- J. ExP. ZOOL., 194; 44-66, midine and detected with autoradiography. The fate of the DNA synthesizing cells was followed by taking samples of the digestive tract at increasing time intervals after the administration of tritiated thymidine. General orientation The digestive tract of ascidians (shown schematically in fig. 1A) is composed of a branchial basket, the food collecting and respiratory apparatus, and a postbranchial region, which functions in digestion and absorption of nutrients. The postbranchial region is made of an esophagus, stomach, intestine, and rectum. The pyloric gland, a diverticulum of the gut, is given off the main course of the digestive tract at the junction of the stomach and intestine. Sea water enters the branchial basket through the branchial siphon, passes through the stigmata (gill slits) in the branchial walls, and leaves through the atrial siphon. The major portion of the branchial basket is concerned with the secretion and move- ment of a mucous net across the branchial 1 Present address: Department of Physiology, Univer- sity of California, San Francisco, California 94143. 449
Transcript
Cell Proliferation in the Digestive Tract of Styela clava (Urochordata: Ascidiacea) as Revealed by Au torad iograp hy with Tri tiated Thymi di ne
THOMAS H. ERMAK' Scripps ln s t i t u t ion of Oceanography, La Jolla, Californiu 92037
ABSTRACT The DNA-synthesizing cells i n the digestive tract of Styela clava were labeled with tritiated thymidine and detected at increasing time intervals with autoradiography. Most of the epithelia consist of renewing cell populations. These are the dorsal tubercle, dorsal lamina, branchial bars, zone 1 of the endo- style, stigmata, esophagus, stomach, intestine, and rectum. Expanding popula- tions are the branchid tentacles, unspeciafized pharyngeal cells, and pyloric tubules. In the renewing populations, cell proliferation occurs in restricted ger- minal zones of pseudostratified cells. In the postbranchial digestive tract, these germinal zones usually occur at the bases of folds or grooves. Germinal cell nu- clei migrate into the cell apex and undergo mitosis there. With time, germinal cells migrate as a band of labeled cells over the underlying connective tissue into non-proliferating mature zones of ciliated, secretory, or absorptive cells. Aging mature cells are presumably extruded into the gut lumen, maintaining the system in a steady state. Transit times are about ten days for the stigmata; two and a half weeks for the dorsal tubercle, esophageal mucous population, stomach crest population, and stomach groove population; five weeks for the intestine and rectum; and several months for zone 1 of the endostyle and esoph- ageal band population.
To date, most of the work on gut cell proliferation employing tritiated thymidine and autoradiography has been confined to the renewing epithelia of mammals (Leblond and Messier, '58; Messier, '60; Leblond et al., '64). Comparable work has been done on relatively few non-mamma- lian vertebrates and invertebrates (Davis and Burnett, '64; Anderson; '65; Holland, '64). The sparsity of such information on ascidians is especially unfortunate because of their vertebrate affinities. Previous work on ascidian gut cell kinetics is limited to a cell formation test (as defined by Leblond et al., '59) for Halocynthia sp. (Mochan et al., '71). However, from anatomical evi- dence alone, the replacement of gut epi- thelial cells has been proposed in several ascidians (Aros and Viragh, '69; Fouque, '53; Relini-Orsi, '68; Thomas, '70).
Therefore, the purpose of the present investigation is to elucidate cell prolifera- tion patterns in the digestive tract of the ascidian Styela claua, a large solitary as- cidian found in bays along the California coast. DNA synthesizing cells about to en- ter mitosis were labeled with tritiated thy-
J. ExP. ZOOL., 194; 44-66,
midine and detected with autoradiography. The fate of the DNA synthesizing cells was followed by taking samples of the digestive tract at increasing time intervals after the administration of tritiated thymidine.
General orientation The digestive tract of ascidians (shown
schematically in fig. 1A) is composed of a branchial basket, the food collecting and respiratory apparatus, and a postbranchial region, which functions in digestion and absorption of nutrients. The postbranchial region is made of an esophagus, stomach, intestine, and rectum. The pyloric gland, a diverticulum of the gut, is given off the main course of the digestive tract at the junction of the stomach and intestine. Sea water enters the branchial basket through the branchial siphon, passes through the stigmata (gill slits) in the branchial walls, and leaves through the atrial siphon. The major portion of the branchial basket is concerned with the secretion and move- ment of a mucous net across the branchial
1 Present address: Department of Physiology, Univer- sity of California, San Francisco, California 94143.
449
3
8
XVNXS ‘H SVNOHL OSP
CELL PROLIFERATION I N ASCIDIAN GUT 451
walls and with the creation of water cur- rents to bring in plankton and suspended organic matter. The mucous net is se- creted by the endostyle and moves along the branchial bars by ciliary action. Water currents are produced by numerous cili- ated stigmata in the branchial walls. Par- ticulate matter is trapped by the mucous net and moves toward the anterior part of the basket. Here, the mucous passes down the dorsal lamina to the esophageal opening. Below the branchial siphon, the branchial tentacles exclude larger sus- pended particles, but allow smaller parti- cles to enter the branchial basket. The dorsal tubercle, which projects into the branchial basket, plays a role in chemore- ception (Carlisle, '53).
The postbranchial digestive tract lies on the left side of the branchial basket. The esophagus joins the basket half way down the dorsal side. A t the entrance to the esophagus, the mucous net forms a food cord, passes through the stomach and becomes compacted in the intestine. Mu- cous secretions are added in the esopha- gus; enzymes are secreted mainly in the stomach; and absorption occurs chiefly in the intestine (van Weel, '40; Morton, '60).
The internal lining of the digestive tract of Styela is extensively folded, a feature characteristic of solitary ascidian species. Each region of the digestive tract is cov- ered by the atrial epithelium (an epidermal specialization) on the outside and lined by the digestive epithelium on the inside. In between the epithelia occur blood spaces and connective tissue. In the branchial region, smooth muscles pass below the branchial bars, and in the postbranchial region, the pyloric tubules run below the digestive epithelium.
Much of the digestive tract is lined by simple columnar epithelia, the majority of which are either ciliated, secretory, or absorptive in nature. In several restricted zones throughout the digestive tract, the epithelia are modified into basophilic zones of pseudostratified columnar cells (fig. 2D). These zones are most obvious at the bases of the folds in the esophagus and stomach and in the intestinal groove. These regions have been cited as regions of intense cell proliferation (Fouque, '53; Relini-Orsi, '68), since mitotic figures frequently occur along
the lumenal edges of the pseudostratified regions.
For more complete descriptions of the mechanism of feeding in ascidians and the morphology of the digestive tract in Styela, the reader is asked to refer to McGinitie ('39), Berrill ('50), Morton ('60), Fouque ('53), and Relini-Orsi ('68).
MATERIALS AND METHODS
Styela c l a m is a club-shaped ascidian native to Korea and Japan and lives on docks, pilings, and boats in protected Cali- fornia bays (Abbott and Johnson, '72; Ab- bott, '75). Individuals up to 5 cm in length are relatively clean, showing the brown wrinkled test, but larger ones usually have a coating of bryozoans, algae, and other ascidians. Seventy specimens of S. clava were collected from the docks at Quivira Basin of Mission Bay, San Diego, California, and injected with 5 microcuries of tritiated thymidine (New England Nuclear Corp.) per gram fresh weight. Individuals ranged in weight from one to four grams. The tritiated thymidine (specific activity 6.7 curies per millimore) was diluted 1 : 1 with two times concentrated sea water and in- jected intra-atrially. The animals were maintained at 18"C-2OoC until the time of sacrifice. Ten individuals were sacrificed by fixation in Bouin's fluid at each of the following time intervals: 1 hour, 3 days, 10 days, 15 days, 20 days, 30 days, and 60 days after injection.
The digestive tract was dissected from the body after at least one day's fixation and the various organs dehydrated in alco- hol and embedded in paraffin. Sections were cut at 7 microns and covered with Kodak Nuclear Track Emulsion type NBT-2 by the dipping method. Autoradiograms were exposed for periods from two weeks to two months and developed in Kodak D-19 developer (3 minutes) at 68°F. All slides were stained through the emulsion with hematoxylin.
The following structures or organs were examined: branchial tentacles, dorsal tu- bercle, endostyle, dorsal lamina, branchial bars, stigmata, esophagus, stomach, intes- tine, rectum, and pyloric gland. Tho struc- tures in the branchial basket were not studied: the penpharyngeal band, which is continuous with the dorsal lamina, and
452 THOMAS H. ERMAK
the retropharyngeal band, whose mucous cells and band cells are considered in the esophagus.
RESULTS
Branchial tentacles At one hour after injection, no localized
region of cell proliferation is observed in cross sections of the tentacles, two or three nuclei being labeled per section. No changes are observed after longer time intervals.
Dorsal tubercle The dorsal tubercle is a knob-shaped
protuberance with a deep horseshoe-shaped groove running along its surface (fig. 1C). The exact shape of the groove, however, varies among individuals. The groove is lined by the heavily ciliated funnel epithe- lium while the surface is lined by the non- ciliated lip epithelium (fig. 1 D). Pseudo- stratified regions occur at the base and at the top of the groove. Beneath the funnel epithelium is a band of acidophilic mate- rial which appears to be supportive in function.
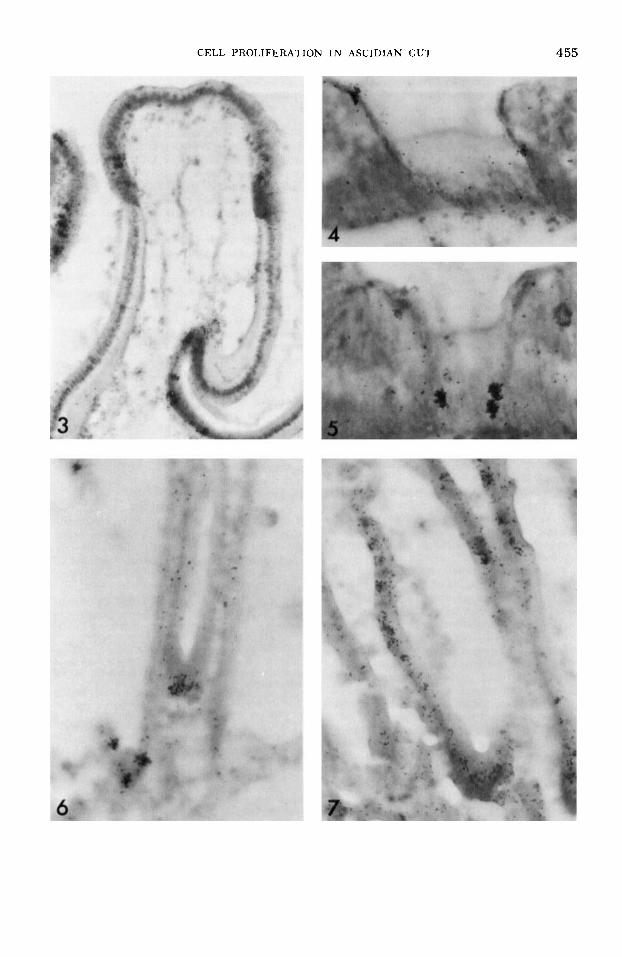
At one hour after injection, many pseu- dostratified cells are labeled, but no cells on the sides of the grooves or on the sur- face of the dorsal tubercle are labeled (fig. 3). By 20 days after injection, cells have migrated onto the sides of the grooves and onto the surface of the dorsal tubercle.
Endostyle The endostyle consists of a deep furrow
running along the ventral surface of the branchial basket (fig. 1B). Most early ac- counts recognize eight cell populations. However, recent ultrastructural examina- tions of zone 5 on the endostyle of several ascidians (Godeaux and Firket, '66, '69) have shown that this zone consists of two separate regions, making nine cell popula- tions in all. By the system of Barrington ('57), zones 6 through 8 are now zones 7 through 9 (fig. 2A). The nine cell popula- tions in the endostyle of Styela clava are roughly arranged in alternating glandu- lar and ciliated regions. Zone 1 has long cilia which reach the top of the endostylar furrow. Zones 3, 5, 6, and 9, on the other hand, have short cilia, while zones 2, 4, and 7 are glandular.
Sokolska ('31) and Levi and Porte ('64) distinguish two compartments in zone 1
consisting of lateral and axial cells. A lat- eral cell compartment lies on each side of the axial cell compartment and partly on top of zone 2 (fig. 2A). At one hour after injection, only lateral cells are labeled (fig. 4). Although only a few cells are la- beled, this is one of the more heavily la- beled zones of the endostyle. By 20 days after injections, no migration of cells is detected. After 60 days, however, several axial cells near the lateral cells are now labeled (fig. 5), indicating that the lateral cells had sunk into the basal portion of zone 1 and differentiate into axial cells.
Although zones 2 through 9 all have la- beled cells at one hour after injection, they show no migration or loss of labeled cells at increasing time intervals. Zones 2 and 4 are the lightest labeled populations while zone 3 is the heaviest labeled population in the endostyle, including zone 1 (see DI sc u s s ION).
Branchial bars and dorsal lamina The dorsal lamina and the ventral (en-
dostylar) surface of each longitudinal bar are covered by ciliated mucous cells while the transverse bars and the dorsal surface of each longitudinal bar are covered by a relatively unspecialized epithelium. The transverse bars are an integral part of the branchial wall. The longitudinal bars, how- ever, are only connected to the rest of the branchial wall at the junction with the transverse bars. As looked at from the in- side of the branchial basket, each longi- tudinal bar bridges over the underlying row of stigmata; either end of the bridge is connected to a transverse bar.
At one hour after injection, many cili- ated cells on the dorsal lamina and on the ventral surface of the longitudinal bars are
Fig. 2 A. A cross section through the endostyle showing the nine cell populations. B. A cross sec- tion through the esophagus. C . The ventral (left) and dorsal (right) internal surfaces of the stomach and descending part of the intestine showing the twisting longitudinal folds on the ventral surface and the straight folds on the dorsal surface of the stomach. D. A cross section through a pseudostrati- fied region. E. A cross section through the stomach. F. A cross section through the intestine in correct position in relation to the stomach in E. cb, band cells; cm, mucous cells; r, raphe; b, bulb; ty, typhosole; f, folds; ig, intestinal groove; cc, simple columnar cells; mf, mitotic figure; tc, transitional cells; pc, pseudostratified columnar cells; gp, groove population; cp, crest population; pt, py- loric tubules.
CELL PROLIFERATION I N ASCIDIAN GUT 453
454 THOMAS H. ERMAK
labeled, but few unspecialized cells are labeled. On the longitudinal bars most la- beled cells occur close to the junction with the transverse bars. On the dorsal lamina, no distinct region of cell proliferation oc- curs, the labeled nuclei being scattered along the entire epithelium.
At increasing time intervals, there is a loss of labeled cells from both the longitud- inal bars and the dorsal lamina. Although the ciliated epithelia appear to be renewed, no definite patterns of cell migration could be determined. There appears to be a gen- eral movement of cells towards the tip of the dorsal lamina and away from the at- tached parts of the longitudinal bars towards the free parts. Cell migration pat- terns are difficult to ascertain because there appears to be no clear separation into germinal and mature compartments. At all times after injection, the unspecial- ized epithelium on the branchial bars has scattered positive reactions.
Stigmatic ep i the l ium Each stigma is lined by heavily ciliated
cells which beat outward when producing water currents. Small pseudostratified re- gions of basophilic cells occur at the an- terior and posterior end of each stigma. At one hour after injection, only pseudo- stratified cells are labeled (fig. 6). Labeled nuclei are not observed in sections of every pseudostratified region, but usually a few nuclei are labeled. No cells on the sides of the stigmatic walls are labeled. At ten days after injection, cells on the side walls are now labeled (fig. 7) indicating that cells from the anterior end of each stigma have migrated posteriorly, and that the cells at the posterior end have migrated anteriorly. The two bands of labeled cells have met each other on the side wall, leaving behind lightly labeled cells in the pseudostratified regions. A t 20 days after injection, the cells lining the stigmata are no longer labeled. Thus, all those cells labeled at the time of injection as well as their immediate prog- eny have been lost from the cell population.
Esophagus The esophagus has five folds and is lined
by two populations of cells. Mucous cells occur on every fold except the base of the ventral fold where band cells occur. The band cells are restricted to one side of the
fold (fig. 2B). Pseudostratified regions oc- cur at the base of each fold and on each side of the band cell population.
At one hour after injection, only those nuclei which occur in a basal position in the pseudostratified region are labeled. Mitotic figures along the lumenal edge of the pseudostratified region and mucous cells and band cells on the sides of the folds are not yet labeled.
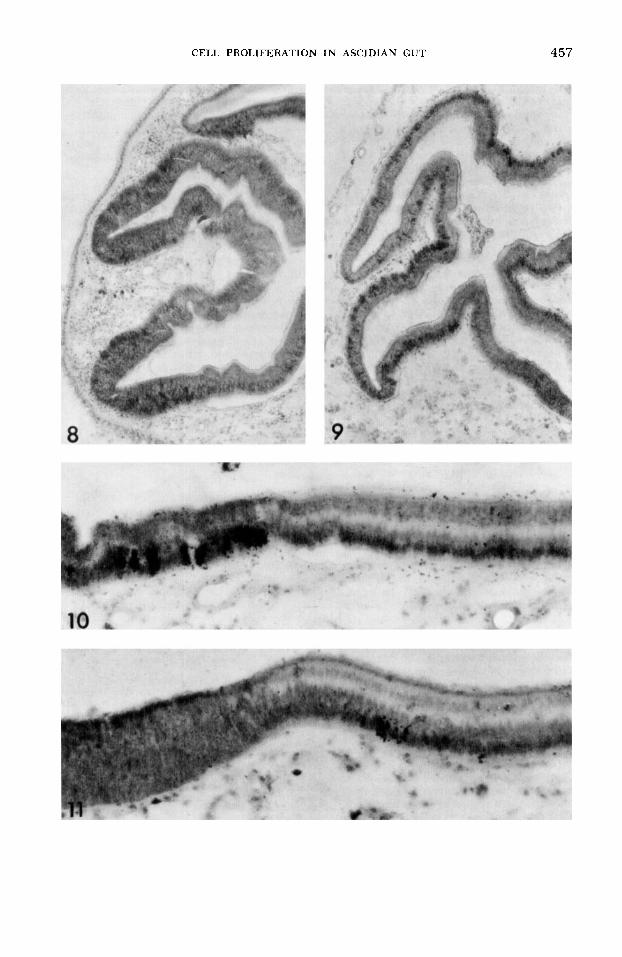
At three days after injection, nuclei are labeled at all levels in the pseudostratified layer (fig. 8). Mitotic figures are also some- times labeled, but no mucous cells or band cells are labeled. The labeling of mitotic figures by three days is a result of the mi- gration of interphase nuclei from the basal to the apical portion of the cell in prepara- tion for mitosis. After mitosis, the daugh- ter nuclei migrate back to the basal por- tion of the cell.
By 10 to 20 days after injection mucous cells but no band cells are now labeled. As shown in figure 9, there is quite a bit of variability in migration rate both be- tween animals and between folds in the same animal.
By 60 days, mucous cells are no longer labeled, presumably because they are ex- truded into the gut lumen; however, the band cells are now labeled (compare figs. 10 and 11). The band cells only originate from the pseudostratified region on the side of the ventral groove. This pseudostra- tified region is unique in that i t produces both mucous cells and band cells; mucous cell production occurs on the opposite side of the pseudostratified region from band cell production.
Stomach The stomach has 25 to 30 longitudinal
folds on its dorsal and lateral surfaces.
Fig. 3 An autoradiogram of the lip and funnel epithelia of the dorsal tubercle one hour after in- jection. X 315.
Fig. 4 An autoradiogram of zone 1 of the en- dostyle 20 days (same as 1 hour) after injection. Only lateral cells are labeled. X 600.
Fig. .5 An autoradiogram of zone 1 of the en- dostyle 60 days after injection. Axial cells are now labeled. X 600.
Fig. 6 An autoradiogram of several stigmata one hour after injection showing labeled pseudo- stratified cells. X 600.
Fig. 7 An autoradiogram of several stigmata ten days after injection showing labeled ciliated cells on the side walls. X 600.
CELL PROLIFERATION IN ASCIDIAN GUT 4 55
456 THOMAS H. ERMAK
The ventral surface is covered by a raphe which expands into a large bulb halfway down the length of the stomach (fig. 2C) and continues as the typhosole in the in- testine. The stomach folds are covered by two cell populations: the groove population which occurs at the base of each fold, and the crest population, which occurs on the top of each fold (fig. 2E). At the anterior and posterior ends of the stomach, the raphe, bulb, and crests, which are all cov- ered by ciliated mucous cells, are continu- ous with each other and with the mucous cells in the esophagus, Chief and zymogen cells (terminology, Burighel and Milanesi, '73) cover the stomach grooves. The chief cells, however, are much more numerous than the zymogen cells, and are absorptive in function. Pseudostratified regions occur at the base of each groove, at the side of each crest, and at the side of the raphe and bulb. Mitotic figures occur along the lumenal edge of the pseudostratified re- gions, but they are most numerous in the groove population.
The folds on the dorsal surface of the stomach are the longest, while the ones directly adjacent to the ventral raphe are the shortest. The dorsal folds are straight (fig. 2C); the ventral folds, however, twist around the raphe and bulb. The anterior ends intersect the raphe while the poste- rior ends intersect the bulb, suggesting that the stomach folds generate hom the ventral surface. As the folds move away from the ventral surface, they lengthen and become higher.
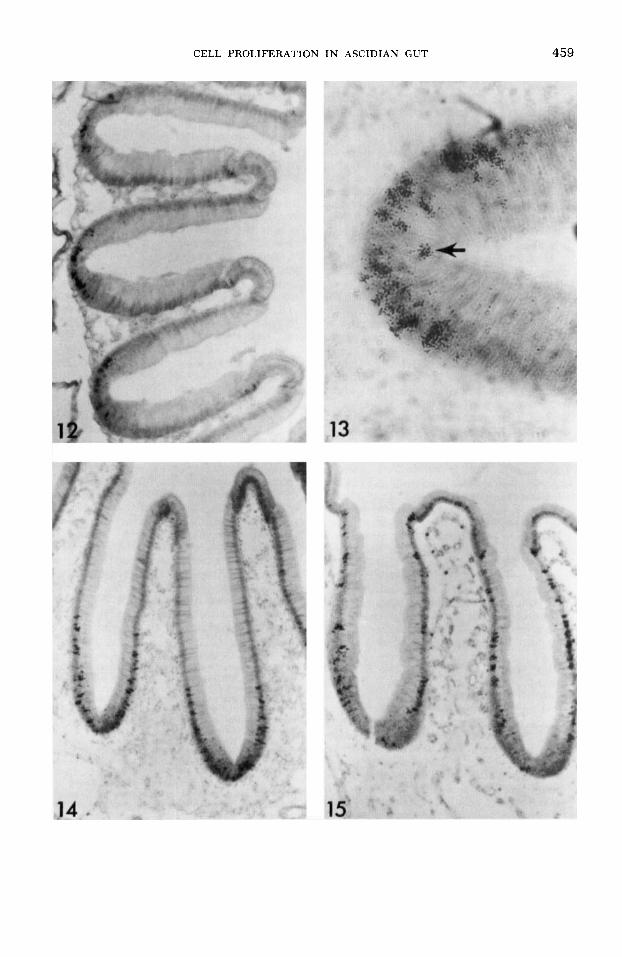
At one hour after injection, only nuclei at the base of each pseudostratified region are labeled (fig. 12). By three days after injection, nuclei at all levels in the pseudostratified region are labeled (fig. 13). No mature cells are yet labeled, but several mitotic figures are la- beled. As in the esophagus, nuclei from the basal portion of each pseudostratified re- gion had migrated toward the lumenal edges of the cells in preparation for mito- sis. Two labeled nuclei immediately adja- cent to each other in the intermediate levels of the pseudostratified region are some- times observed, indicating that they are the two daughter nuclei of a recently di- vided labeled cell. By three days, there has been a slight migration of labeled cells towards the mature zone.
Groove population.
By ten days after injection, mature cells on the sides of the grooves are now labeled. Both chief and zymogen cells are labeled, and several pseudostratified cells are still labeled (fig. 14). Animals killed 15 days after injection have several folds where the entire groove height is labeled (fig. 15). At this time, the originally small band of closely clustered germinal cells has ex- panded into a long band of labeled mature cells intermingled with unlabeled mature cells.
After 20 days, the entire groove height is labeled, but no pseudostratified cells are labeled any longer. By 30 days, few cells in the groove population are still labeled. Thus, most of the DNA synthesizing cells labeled at the time of injection as well as several generations of their progeny have been lost from the epithelium, evidently through cell extrusion at the junction of the groove population with the crest popu- lation on the tops of the folds.
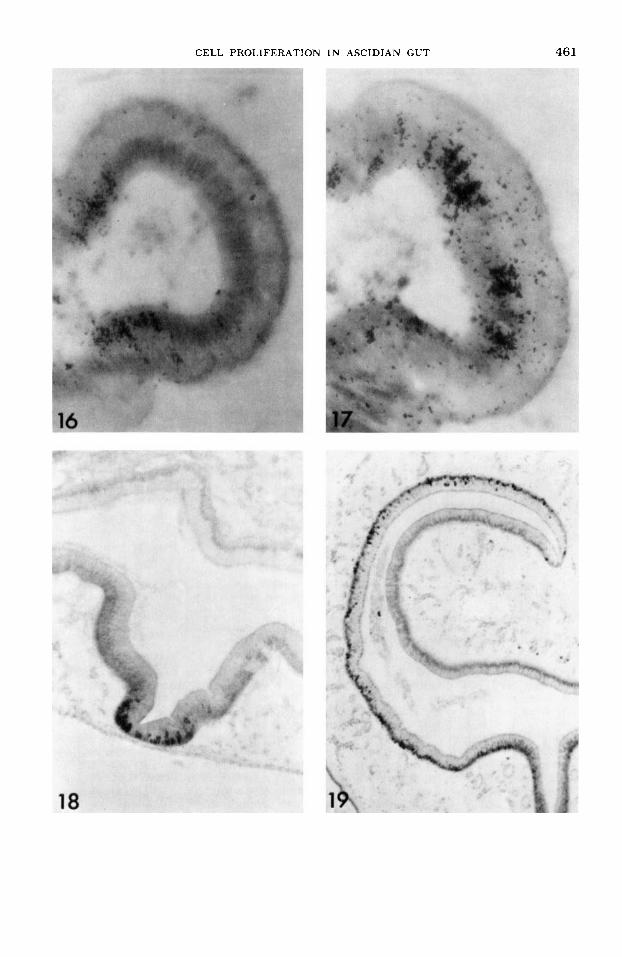
Crest population. At one hour after injection, cells on the sides of the stomach crests are labeled (fig. 16). No mucous cells on the tops of the crests, however, are labeled. Labeled nuclei do not occur in sections of every crest, since the population is small. The crest cells also have fewer grains above their nuclei than the cells in the groove population.
By ten days after injection, the two bands of labeled cells on each side of the crests have migrated toward each other (fig. 17), and by 15 and 20 days, the two bands have met each other on the tops of the crests. By 30 days, the crests are gen- erally unlabeled. Evidently, the mucous cells are extruded into the gut lumen at the tip of each stomach fold. Thus, two separate renewing cell populations occur
Fig. 8 An autoradiogram of two esophageal folds three days after injection. Only cells at the base of the folds are labeled. X 200.
Fig. 9 An autoradiogram of some esophageal folds 20 days after injection. Cells on the tops of the folds are now labeled. X 200.
Fig. 10 An autoradiogram of a pseudostrati- fied region (left) and some adjacent band cells (right) one hour after injection. Only pseudostrati- fied cells are labeled. X 420.
Fig. 11 An autoradiogram of a pseudostrati- fied region (left) and some adjacent band cells 60 days after injection. Pseudostratified cells are no longer labeled but a few band cells are now la- beled. X 420.
CELL PROLIFERATION IN ASCIDIAN GUT 457
458 THOMAS H. ERMAK
on each fold, each renewed by its own ger- minal cells.
The stomach raphe and bulb are basic- ally large crest populations which are re- newed on the same order of time. However, these populations are much larger in size. Many cells are labeled on the bulb at one hour, and migration (in microns per day) is rapid. By 15 to 20 days after injection, the two large bands of mucous cells have met each other on the top of the raphe and bulb, as on the crests.
lntes tine The intestine (fig. 2F) is a long tube
with a typhosole on one side and a deep groove running along the length of the other side. The intestinal groove is lined by pseudostratified cells while the rest of the intestinal wall is lined by absorptive and secretory cells.
At one hour and three days after injec- tion, labeling is much like in the groove population of the stomach with only pseu- dostratified cells in the intestinal groove labeled (fig. 18). Cells migrate at a differ- ent rate, however. After ten days, cells are labeled up to the top of the intestinal groove and frequently onto the side walls of the intestine. Both absorptive and se- cretory cells are labeled. By 15 days, the leading edge of labeled cells has migrated around the side walls almost to the begin- ning of the typhosole.
After 20 days, labeled cells have mi- grated up the sides of the typhosole (fig. 19). The band of labeled cells is clearly defined, unlike in the shorter mature zones in the esophagus and stomach. In front of the band of labeled cells, no cells are la- beled. Behind it, cells are usually only lightly labeled. By 30 days, the leading edges have met each other at the top of the typhosole in a few animals. In others, the leading edges have almost met each other.
Rec turn The rectum is the terminal part of the
digestive tract and is characterized by the lack of a groove of pseudostratified cells and by a typhosole which has transformed into a thin flap. A region of basophilic cells runs along the dorsal surface of the rectum in a position corresponding to the pseudostratified region of the intestine.
Cell proliferation and cell migration pat-
terns in the rectum are similar to those observed in the intestine. The major dif- ference is in the size of the germinal and mature zones, with fewer cells proliferat- ing in the germinal zone of the rectum than in the germinal zone of the intestine.
Pyloric gland The pyloric gland is composed of numer-
ous tubules which are exceedingly numer- ous at the level of the mid-intestine and are solidly packed close to the intestinal epithelium (fig. 2F). Although the tubules are also fairly abundant in the stomach wall, they are not packed close together next to the gut epithelium as in the intes- tine. A few pyloric tubules even occur in the esophagus and intestine. Although the tubules occur associated with the mature cells of the digestive tract, they are not closely associated with the germinal zones, especially in the intestine. The fact that the pyloric tubules occur directly adjacent to the absorptive cells in the intestine but not the germinal cells suggests that this organ might be involved with the process of nutrient absorption and assimilation. At all times after injection, few cells are labeled in the pyloric tubules.
DISCUSSION
The classification of cell populations (as used by Messier and Leblond, '60; Camer- on, '70) in the digestive tract of Styelu c l a m is given in table 1. Static populations, which undergo no cell proliferation, do not occur in the digestive tract nor in the rest of the body (Ermak, '75). Expanding popu- lations have a slow rate of cell prolifera- tion and no detectable cell turnover within the life span of the animal (about 12-18
Fig. 12 An autoradiogram of the stomach grooves one hour after an exposure to tritiated thymidine. Cells are labeled at the base of each groove. x 150.
Fig. 13 An autoradiogram of a germinal zone from a stomach groove three days after injection of tritiated thymidine. Nuclei at various levels within the pseudostratified region and a mitotic figure (arrow) are labeled. X 600.
Fig. 14 An autoradiogram of some stomach grooves ten days after injection of tritiated thymi- dine. Labeled cells now occupy approximately half of the groove height. X 150.
Fig. 15 An autoradiogram of the groove pop- ulation 15 days after an exposure to tritiated thy- midine. Labeled cells now occur along the entire groove height. Few cells at the base of the groove remain labeled. X 150.
CELL PROLIFERATION I N ASCIDIAN GUT 459
460 THOMAS H. ERMAK
months, Johnson, '71). Although zone 3 of the endostyle appears to be an expanding population, longer termed studies would be required to decide whether i t is expand- ing or only slowly renewed.
Renewing populations have a high rate of cell proliferation, usually demonstrate cell migration, and have turnover times within the life span of the animal. Renew- ing populations are divided into two cate- gories based upon their turnover times. Type I populations have turnover times of less than 60 days (usually between 1 and 5 weeks) whereas type I1 populations have turnover times of more than 60 days (prob- ably up to several months). The reason why type I1 cell populations have such long renewal times in comparison to the other renewing populations is not presently un- derstood. The renewal of the two popula- tions in the esophagus is of particular in- terest since both mucous cells (type I) and band cells (type 11) originate from the same stem cell compartment. The patterns of cell migration and loss in the major re- newing populations (dorsal lamina and branchial bars excepted) are shown sche- matically in figure 20.
From the limited work on cell popula- tion kinetics of invertebrates, it seems safe to assume that renewing gut cell pop- ulations are rarely found. Cell renewal occurs in the ascidian branchial wall and also in the gill of the freshwater mussel (Tomasovic and Mix, '74). In addition to the ascidian gut, only part of the crayfish gut (the hepatopancreas) is known to con- tain renewing cell populations (Davis and Burnett, '64). More commonly, the gut cell populations of invertebrates are of the expanding type; such populations occur in sea urchins (Holland, '64) and probably sea stars (Anderson, '65) and oysters (Mix, '71).
My results for Styela clava have con- b e d earlier reports of stem cell popula- tions in the digestive tract of ascidians (Fouque, '53; Relini-Orsi, '68; Thomas, '70; Aros and Viragh, '69). In Styela pli- cata, Fouque ('53) suggested from static light microscope evidence that the cells at the base of the grooves in the stomach migrate up the folds, becoming at first chief and zymogen cells and then differ- entiating into ciliated mucous cells on the crests. In Ciona intestinalis, Thomas ('70)
believed that the undifferentiated cells re- place only the zymogen cells which are then lost from the epithelium. He consid- ered that the chief cells represent a sepa- rate cell-type not derived from the zymogen cell population. My results clarify these points. Two separate cell populations (the groove and crest populations) occur in the stomach of Styela clava, each renewed by its own germinal cells. The chief and zymo- gen cells in the groove population, how- ever, differentiate from the same germinal cells at the groove bases.
In a typical renewing population in the postbranchial digestive tract, DNA is syn- thesized in pseudostratified cells whose nuclei lie close to the basal portion of the cell. As each cell enters mitosis the nucleus migrates toward the lumenal edge of the cell. Labeled mitotic figures appear as soon as five hours after labeling (Ennak, unpub- lished results). After cell division, the two daughter nuclei move back into the region of interphase nuclei. The process of nu- clear migration in preparation for mitosis, interkmetic migration, is responsible for the dispersal of labeled nuclei throughout the various levels of the pseudostratified region by three days after injection. Inter- kinetic migration of nuclei also occurs in many other epithelia such as the epithelium of the neural tube (Watterson, '65), the mouse intestinal epithelium (Kataoka, '70), and the mammalian oviduct epithelium (Wrenn and Wessells, '70). The exact mechanism by which nuclear migration is achieved is not known, but it might in- volve microfilaments (Hinds and Ruffett, '71; Wessells et al., '71).
Cell division is responsible for the in- crease in the number of nuclei labeled with tritiated thymidine at increasing time in-
Fig. 16 An autoradiogram of a stomach crest three days after administration of tritiated thymi- dine. Cells are labeled on the sides of the crest. x 600.
Fig. 17 An autoradiogram of a stomach crest ten days after administration of tritiated thymi- dine. Columnar cells on the top of the crest are now labeled. X 600.
Fig. 18 An autoradiogram of the intestinal groove one hour after injection of tritiated thymi- dine. Nuclei are labeled at the base of the groove. X 160.
Fig. 19 An autoradiogram of one half of the intestine 20 days after administration of tritiated thymidine. Cells are labeled up to the base of the typhosole. X 150.
CELL PROLIFERATION IN ASCIDIAN GUT 461
462
A
THOMAS H. ERMAK
Fig. 20 Schematic diagrams of cell renewal in the digestive tract of Styela cluua showing directions of cell migration (small arrows) and regions of cell loss (large arrows). Black regions indicate germinal zones. Diagrams not to scale. A, lip epithelium (shaded) and funnel epithe- lium of the dorsal tubercle; B, zone I of the endostyle; C, stigmatic cells; D, ciliated band cells (shaded) and ciliated mucuous cells of the esophagus, E, crest population (shaded) and groove population of the stomach; F, intestinal epithelium.
tervals after injection. After a DNA syn- thesizing cell has divided, the daughter cells may differentiate or divide again. Those cells in the germinal zone which lie close to the transitional zone are more likely to differentiate into mature cells while those cells which lie far away from the transitional zone are more likely to di- vide a few more times. In any case, all those cells initially labeled with tritiated thymidine and/or their labeled progeny
eventually move out of the germinal zone, pass through the transitional zone, and then migrate through the mature zone in a linear fashion. Those cells initially la- beled and their immediate progeny move through the epithelium as a band of heav- ily labeled cells. In front of the band, no cells are labeled. Behind it, cells are only lightly labeled or not labeled at all. Those cells which are lightly labeled are descend- ants through many generations from those
CELL PROLIFERATION IN ASCIDIAN GUT 463
TABLE 1
Classification of the cell populutions in the digestive tract of Styela clava according to proliferative behuvior
Type of cell population Example (with transit time)
Expanding populations Branchial tentacles Unciliated pharyngeal cells Zones 2, 3,4, 5 , 6 , 7, 8, and 9 of endostyle Pyloric gland
Funnel epithelium of dorsal tubercle (2.5 weeks) Lip epithelium of dorsal tubercle (2.5 weeks) Dorsal lamina and branchial bars (several weeks) Stigmatic cells (1.5 weeks) Mucous cells of esophagus (2.5 weeks) Crest population of stomach (2.5 weeks) Groove population of stomach (2.5 weeks) Intestinal epithelium (5 weeks) Rectal epithelium (5 weeks) Zone 1 of endostyle (several months) Band cells of esophagus (several months)
Renewing populations A. Type I
(renewal in less than 60 days)
B. Type I1 (renewal in more than 60 days)
cells which were initially labeled with tritiated thymidine and have had their DNA diluted with unlabeled precursors. Assuming a chromosomal number of 32 (Minouchi, '36) for Styela, the progeny of labeled cells would have only one labeled chromosome after six cell divisions (Cron- kite et al., 'Sl), if a labeled cell remained in the germinal zone long enough to divide six times, that is. In any case, the band of labeled cells is finally lost from the cell population, presumably through extrusion of cells into the lumen of the gut.
Although most of the cell proliferation in the germinal compartments is for the replacement of mature cells lost from the cell populations, some proliferation pro- vides for the growth of the digestive tract, especially in small individuals. As animals grow larger, the stigmata multiply, the folds in the esophagus and stomach grow longer, and the intestine grows wider.
There also appears to be some mecha- nism of regulating cell proliferation and maturation in the ascidian digestive tract such as that proposed in the rat intestine (Galjaard et al., '72; Rijke et al., '74). Transit times are the same for the mucous cells in the esophagus and stomach, al- though the sizes of the cell populations (esophageal fold; stomach crest, raphe and bulb) and the actual migration rates (in microns per days) are different. Migration rates are proportional to the sizes of the germinal and mature compartments. Tran- sit times for the chief cells in the stomach
and intestine, however, are dramatically different. Cells in the intestine migrate faster yet are renewed slower than cells in the stomach. It is reasonable to assume that the stomach is under more demands for cell turnover than the intestine. Morton ('60) claims that the stomach is the pri- mary site of digestion in ascidians while Yonge ('25) and van Wee1 ('40) state that the intestine is the primary site of absorp- tion. Presumably the stomach provides a more formidable environment of digestive enzymes.
The renewing populations in the ascidian postbranchial digestive tract are quite sim- ilar to the renewing populations in the stomach and intestine of mammals. The renewal of the mucous cells in the esoph- agus and stomach of Styela is similar to the renewal of the surface epithelium in the mammalian stomach (Messier, '60). The renewal of the band cells in the esoph- agus of Styela is similar to the renewal of the parietal cells in the mammalian stom- ach (Raggins et al., '68; Kataoka, '70) and the Paneth cells in the mammalian intes- tine (Cheng et al., '69; Cheng, '74b). The band cells, parietal cells, and Paneth cells are all renewed at a much slower rate than most of the other cells lining the digestive tract. They also originate from germinal cells which also give rise to the rapidly re- newed cells of the digestive tract.
The renewal of the groove population in the stomach and the renewal of the in- testinal epithelium of Styela is most like
464 THOMAS H. ERMAK
the renewal of the intestinal villi of mam- mals (Leblond and Messier, '58; Messier and Leblond, '60; Cheng and Leblond, '74a,b; Cheng, '74a). In mammals, cell proliferation occurs only in the crypts of Lieberkiihn. Crypt cells migrate up the villi, differentiate into chief and goblet cells, and are finally extruded at the villus tips. Cell migration in mammals, however, is out of pits and up fingerlike projections. Migration in Styela, on the other hand, is linear and up longitudinal folds. Perhaps the most conspicuous difference between ascidian and mammalian gut renewal is in transit time. The time for a labeled cell to reach the villus tip is about one and one-half days in the rat (Messier and Le- blond, 'SO), about three days in the mouse (Leblond and Messier, '58), and about five to six days in man (MacDonald et al., '64). In Styela, transit times are about two and a half weeks in the stomach and about five weeks in the intestine. Cell renewal times in the intestine of Styela, however, are comparable to those in the newt intes- tine (O'Steen and Walker, '60) where re- newal is on the order of a month.
ACKNOWLEDGMENTS
I am indebted to Dr. Nicholas D. Holland for his advice, support, and criticism. I thank Emily Reid for the final drawings of figures 1 and 2.
LITERATURE CITED
Abbott, D. P. 1975 Phylum chordata: Introduc- tion and Urochordata. In: Light's Manual. R. I. Smith and J. T. Carlton, eds. Univ. Calif. Press, Berkeley, pp. 6384355.
Abbott, D. P., and J. V. Johnson 1972 The as- cidians Styela burnharti, s. plicatu, s. clava, and S. montereyensis in California waters. Bull. S. Cal. Acad. Sci., 71 : 95-105.
Anderson, J. M. 1965 Studies on visceral regen- eration in sea stars. 11. Regeneration of pyloric caeca in Asteriidae with notes on the source of cells in regenerating organs. Biol. Bull., 128; 1-23.
Aros, B., and S. Viragh 1969 Fine structure of the pharynx and endostyle of an ascidian (Ciona intestinalis). Acta. Biol. Acad. Sci. Hung., 20: 281-297.
Berrill, N. J. 1950 The Tunicata. Ray Society, London.
Burighel, P., and C. Milanesi 1973 Fine struc- ture of the gastric epithelium of the ascidian Botryllus schlosseri. Vacuolated and zymogen cells. Z. Zellforsch. Microscop. Anat., 145: 541- 555.
Cameron, I. L. 1970 Cell renewal in the organs and tissues of the non-growing adult mouse. Texas Rep. Biol. Med., 28: 203-248.
Cheng, H. 1974 Origin, differentiation and re- newal of the four main epithelial cell types in the mouse small intestine. 11. Mucous cells. Am. J. Anat., 141 : 481-502.
1974 Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine. IV. Paneth cells. Am. J. Anat.,
Cheng, H., and C. P. Leblond 1974 Origin, dif- ferentiation and renewal of the four main epi- thelial cell types in the mouse small intestine. I. Columnar cell. Am. J. Anat., 141 : 461-480.
1974 Origin, differentiation and renew- al of the four main epithelial cell types in the mouse small intestine. V. Unitarian theory of the origin of the four epithelial cell types. Am. J. Anat., 141 ; 537-562.
Cheng, H., J. Merzel and C. P. Leblond 1969 Renewal of Paneth cells in the small intestine of the mouse. Am. J. Anat., 126: 507-524.
Cronkite, E. P., S. W. Greenhouse, G. Brecher, V. P. Bond 1961 Implication of chromosome structure and replication on hazard of tritiated thymidine and the interpretation of data on cell proliferation. Nature, 189: 153-154.
1964 A study of growth and cell differentiation in the hepato- pancreas of the crayfish. Devel. Biol., 10: 122- 153.
Ermak, T. H. 1975 Cell Proliferation in the As- cidian Styela clava: An Autoradiographic and Electron Microscopic Investigation Emphasizing Cell Renewal in the Digestive Tract of This and Fourteen Other Species of Ascidians. Ph.D. dis- sertation, Univ. Calif., San Diego.
Fouque, G. 1953 Contribution ~3 1'Ctude de la glande pylorique des ascidiaces. Ann. Inst. Oceanogr. (Paris), 28: 18CL317.
Galjaard, H., W. Van der Meer-Fieggen and J. Giesen 1972 Feedback control by functional villus cells on cell proliferation and maturation in intestinal epithelium. Exp. Cell Res., 73: 197- 207.
Godeaux, J., and H. Firket 1966 Ultrastructure de l'endostyle de Molgula manhattensis Kay (As- cidie stolidobranche). Compt. Rend. Acad. Sci. (Paris), 262: 48EL490.
1969 Etude au microscope electronique de l'endostyle d'une ascidie stolidobranche M o l - gula manhattensis Kay. Ann. Sci. Nat. Zool. Biol. Animale, 10: 163-187.
Hinds, J. W., and T. L. Ruffett 1971 Cell pro- liferation in the neural tube: an electron micro- scopic and Golgi analysis in the mouse cerebral vesicle. Z. Zellfrsch. Mikr. Anat., 115: 226-264.
Holland, N. D. 1964 Cell Proliferation in Post- embryonic Specimens of the Purple Sea Urchin (Strongylocentrotus purpuratus): an Autoradio- graphic Study Employing Tritiated Thymidine. Ph.D. dissertation, Stanford Univ.
Johnson, J. V. 1971 The annual growth and reproductive cycle of Styela sp. in the Marina del Rey, Venice, California. M. S. thesis, Dept., Zool., Univ. Nebraska.
Kataoka, K. 1970 The fine structure of the pro- liferative cells of the mouse intestine as revealed by electron microscopic autoradiography with H3-thymidine. Z. Zellforsch. Mikr. Anat., 103: 170-178.
1 4 1 : 521-536.
Davis, L. E., and A. L. Burnett
CELL PROLIFERATION I N ASCIDIAN GUT 465
Leblond, C. P., R. C. Greulich and J. P. M. Pereira 1964 Relationship of cell formation and cell migration in the renewal of stratified squamous epithelium. Adv. Biol. Skin, 5: 39-67.
Leblond, C. P., and B. Messier 1958 Renewal of chief cells and goblet cells in the small intes- tine as shown by radioautography after injection of thymidine.Hs into mice. Anat. Rec., 132: 247-259.
1959 Thymidine-Hs as a tool for the investigation of the renewal of cell populations. Lab. Invest., 8: 296308 .
Levi, C., and A. Porte 1964 Ultrastructure de l'endostyle de l'ascidie Microcosmus claudicans Savigny. 2. Zellforsch. Mikr. Anat., 62: 293- 309.
MacDonald, W. C., J. S . Trier and N. B. Everett 1964 Cell proliferation and migration in the stomach, duodenum, and rectum of man: radio- autographic studies. Gastroenterol., 46: 405- 41 7.
McGinitie, G. E. 1939 The method of feeding of tunicates. Biol. Bull., 77: 44-47.
Messier, B. 1960 Radioautographic evidence for the renewal of the mucous cells in the gastric mucosa of the rat. Anat. Rec., 136: 242.
Messier, B., and C. P. Leblond 1960 Cell pro- liferation and migration as revealed by radio- autography after injection of thymidine-Ha into male rats and mice. Am. J. Anat., 106: 247-265.
Minouchi, 0. 1936 Notiz iiber die chromosomen von Tethyum plicatum Les. (Ascidia). Z . Zell- forsch. Microscop. Anat., 23: 790-794.
Mix, M. C. 1971 Cell renewal systems in the gut of the oyster Crassostrea gigus (Mollusca: Bivalva). Veliger, 14: 202-203.
Mochan, 0. T., N. L. Liebson and L. T. Frolova 1971 On localization of proliferative process in the guts of some species of invertebrates (In Russian). Sci. Comm. Inst. Mar. Biol. Far East. Sci. Cent. Akad. Nauk. S. S. S. R., 2: 152-157.
Morton, J. E. 1960 The functions of the gut in ciliary feeders. Biol. Rev., 35: 92-140.
Leblond, C. P., B. Messier and B. Kopriwa
OSteen, W. K., and B. E. Walker 1960 Radio- autographic studies of regeneration. Anat. Rec., 137: 501-509.
Ragins, H., F. Winlze, S. M. Liv and M. Dittbren- ner 1968 The origin and survival of gastric parietal cells in the mouse. Anat. Rec., 162: 99-1 10.
Relini-Orsi, L. 1968 Prime osservazioni morfo- logiche ed istochimiche sull' apparato digerente di Styela plicuta. Les. Boll. Mus. Inst. Biol. Univ. Genova, 36: 157-184.
Rijke, R. P. C., W. Van der Meer-Fieggen and H. Galjaard 1974 Effect of villus length on cell proliferation and migration in small intestinal epithelium. Cell Tiss. Kinet., 7: 577-586.
Sokolska, J. 1931 Contribution l'histologie de l'endostyle des Ascidies. Folia Morfol., 3: 1-34.
Thomas, N. W. 1970 Morphology of cell types from the gastric epithelium of Ciona intestinalis. J. Mar. Biol. Ass. U. K., 50: 737-746.
Tomasovic, S. P., and M. C. Mix 1974 Cell re- newal in the gill of the fresh-water mussel, Mar- garitifera margaritifera: an autoradiographic study using high specific activity tritiated thy- midine. J. Cell Sci., 14: 561-569.
Watterson, R. L. 1965 Structure and mitotic be- havior of the early neural tube. In: Organogene- sis. R. L. Dehaan and H. Ursprung, eds. Holt, Rinehart, and Winston, New York, pp. 129-159.
Weel, P. B. van 1940 Beitrage zur Ernahrungs- biologie der Ascidien. Pub. Sta. Zool. Napoli,
Wessells, N. K., B. S. Spooner, J. F. Ash, M. 0. Bradley, M. A. Luduena, E. L. Taylor, J. T. Wrenn and K. M. Yamada 1971 Microfila- ments in cellular and developmental processes. Science, 171 ; 13S143.
Wrenn, J. T., and N. K. Wessells 1970 Cyto- chalasin B: Effects upon microfllaments involved in morphogenesis of estrogen-induced glands of oviduct. Roc. Nat. Acad. Sci., 66: 904-908.
Yonge, C. M. 1925 Secretion, digestion, and assimilation in the gut of Ciona intestinalis. Brit. J. Exp. Biol., 2: 373-388.