SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
RAD IAT ION IN JURY
1Department of Biostatistics and Translational Medicine, Medical University ofLodz, Lodz 91-738, Poland. 2Department of Radiation Oncology, Harvard MedicalSchool, Dana-Farber Cancer Institute, Boston, MA 02215, USA. 3Medical Gyneco-logic Oncology Program, Dana-Farber Cancer Institute, Harvard Medical School,Boston, MA 02215, USA. 4Department of Radiation Oncology, Albert Einstein Col-lege of Medicine, Montefiore Medical Center, Bronx, NY 10461, USA. 5Departmentof Pathology, Albert Einstein College of Medicine, Montefiore Medical Center,Bronx, NY 10461, USA. 6Department of Pharmacology and Experimental Thera-peutics, F. Edward Hébert School of Medicine, Uniformed Services University ofthe Health Sciences, Bethesda, MD 20814, USA. 7Armed Forces Radiobiology Re-search Institute, Bethesda, MD 20814, USA.*Corresponding author. Email: [email protected] (D.C.); [email protected] (V.K.S.); [email protected] (C.G.)
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
Evolutionarily conserved serum microRNAs predictradiation-induced fatality in nonhuman primatesWojciech Fendler,1 Beata Malachowska,1 Khyati Meghani,2 Panagiotis A. Konstantinopoulos,3
Chandan Guha,4,5* Vijay K. Singh,6,7* Dipanjan Chowdhury2*
Effective planning for themedical response to a radiological or nuclear accident is complex. Because of limited resourcesformedical countermeasures, the key would be to accurately triage and identify victimsmost likely to benefit from treat-ment. We used a mouse model system to provide evidence that serummicroRNAs (miRNAs) may effectively predict theimpact of radiation on the long-term viability of animals. We had previously used nonhuman primates (NHPs) to demon-strate that this concept is conserved and serummiRNA signatures have thepotential to serve as predictionbiomarkers forradiation-induced fatality in ahumanpopulation.We identified a signatureof sevenmiRNAs that are alteredby irradiationin both mice and NHPs. Genomic analysis of these conserved miRNAs revealed that there is a combination of seventranscription factors that are predicted to regulate these miRNAs in human, mice, and NHPs. Moreover, a combinationof threemiRNAs (miR-133b,miR-215, andmiR-375) can identify,withnearly completeaccuracy,NHPsexposed to radiationversus unexposed NHPs. Consistent with historical data, female macaques appeared to be more sensitive to radiation,but the difference was not significant. Sex-based stratification allowed us to identify an interaction between gender andmiR-16-2 expression, which affected the outcome of radiation exposure. Moreover, we developed a classifier based ontwo miRNAs (miR-30a and miR-126) that can reproducibly predict radiation-induced mortality. Together, we have ob-tained a five-miRNA composite signature that can identify irradiated macaques and predict their probability of survival.
ht
by guest on June 30, 2018tp://stm
.sciencemag.org/
INTRODUCTIONThe threat of “dirty bombs” and the possibility of the use of improvisednuclear devices are growing with the changing global sociopoliticalscenario and politically turbulent situations in nuclearwarhead–capablestates.Hence, radiationexposureduring terrorist events, industrial accidents,or natural disasters (such as the nuclear disaster after the tsunami inJapan) is a current and continuing threat for the future. The most im-portant characteristic of such radiation exposure is the heterogeneity ofthe dose delivered to different parts of the body, and the prominence ofsymptomsdepends on themagnitude of damage to the organs, resultingin complex pathophysiological features (1–3). Effective planning for themedical response to a radiologic or nuclear incident is challenging andrequires an in-depth understanding of howmedical triage and treatmentwill occur. The current use of diagnostic screening to estimate the dose ofaccidental radiation exposure is mainly based on three factors: time toonset of radiation sickness, kinetics of lymphocyte depletion, and analysisof chromosomal abnormalities (2, 4). These assays are time-consumingand are often not quantitative enough to draw definitive conclusions. Be-cause acute radiation syndrome (ARS) is a multiorgan syndrome (2), it isunlikely that a single biodosimetry assay can be used for the rapid detec-tion of radiation exposure and monitoring of the clinical symptoms andorgan dysfunction in ARS. Therefore, complementary assays, such as thelymphocyte depletion kinetic assay, can be combined with cytogeneticassays (for example, dicentric chromosome assay andmicronucleus assay)
for dosimetry (2, 5–9). Although cytogenetic assays are the gold standardfor radiation biodosimetry, they may have a limited dose range for detec-tion (up to 5Gy for dicentric chromosome assay andmicronucleus assay).In a radiological mass casualty scenario, resources for biodosimetry andmedical countermeasures would be limited. Therefore, the availabilityof rapid biodosimetry diagnostics that will allow accurate triage andidentify victims most likely to benefit from treatment would be critical.
Serum microRNAs (miRNAs) fall under the emerging “omic” bio-dosimetry assays and can be measured by simple technology that mayeffectively determine whether an individual was exposed to radiation(and if so, whether the dose was sublethal or lethal) and predict long-term survival of exposed individuals (10). With current progress in theminiaturizationofquantitativepolymerase chain reaction (qPCR)ormulti-plexed probe-based assays, an miRNA-based assay has the potential tobecome a point-of-care technology that can be performed outside thelaboratory. It can allow shorter lag time postexposure before the firstsample can be taken compared to lymphocyte depletion kinetics andDNA damage assay using g-H2AX, and it can provide results within12 to 24 hours after exposure. We successfully profiled and identifieda panel of serum miRNAs in mice that were differentially expressed inresponse to total body irradiation (TBI) and correlatedwell with injury atsublethal and lethal doses (11). Akey issuewe addressed is the correlationof serum miRNAs with the impact of radiation, specifically with hema-topoietic injury and animal viability.Working with a narrow dose range,we identified serummiRNAs that distinguish between sublethal (6.5 Gy)and lethal (8 Gy) exposure of C57BL/6J mice (11). This is of paramountimportance because during a radiological emergency, doses sustainedwill almost never be in specific increments, anddistinctionbetween lethaland sublethal doses is the key challenge.Moreover, thismiRNAsignaturealso predicted the impact of radiation on animal survival after pre-treatment with the radioprotective agent amifostine or mitigation usingbone marrow–derived stromal cell transplantation. However, the keyquestion that emerged from this study is whether this concept is evolu-tionarily conserved, that is, whether serummiRNAsignatures can predictradiation-inducedmortality in healthy primates and, ultimately, humans.
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
http://stm.sciencem
ag.orD
ownloaded from
To address this question, we conducted a systematic analysis of serummiRNAs in healthy nonhuman primates (NHPs) exposed to TBI aloneor in combination with a radioprotective agent, g-tocotrienol (GT3)(12). Validation of the initial analysis was conducted in an independentcohort of NHPs. We discovered a signature of seven serum miRNAsthat are altered with TBI in both mice and NHPs. Because of the lack ofexperimental data fromhealthy humans, we adopted an in silico approachto compare the loci coding for thesemiRNAs inmice,NHPs, andhumans.The overall sequence homology in the regulatory elements upstream ofthese miRNAs was moderate across the three species. However, therewas a combination of seven transcription factors that are predicted toregulate these miRNAs in humans, mice, and NHPs. A combination ofthree miRNAs (miR-133b, miR-215, and miR-375) was able to distin-guish NHPs exposed to radiation versus unexposed NHPs with com-plete accuracy, even in the presence of a radioprotector, GT3. ThreemiRNAs (miR-30a, miR-126, and miR-375) correlated with the radio-protective properties of GT3 after TBI. Next, we observed a trend (dif-ference was not statistically significant) in the radiosensitivity of theanimals based on sex, with male NHPs more likely to survive the samedose of radiation relative to their female counterparts. Accounting forthis potential sex bias, we showed that miR-16-2 has a sex-specific as-sociation with radiation-induced mortality, that is, rhesus macaquesthat died showed on average twofold lower expression ofmiR-16-2 thansurvivors of the same sex. This hinted at the necessity of incorporatingsex into predictivemodels for survival after high-dose irradiation. Usingmultivariate logistic regression with sex as a confounding variable, wecreated a statistical model using miR-30a and miR-126, which allowedefficient prediction of radiation-induced mortality. Combining theseresults with our irradiation detection model, we have obtained a five-miRNA signature that allows us to identify irradiated macaques andpredict their survival.
by guest on June 30, 2018g/
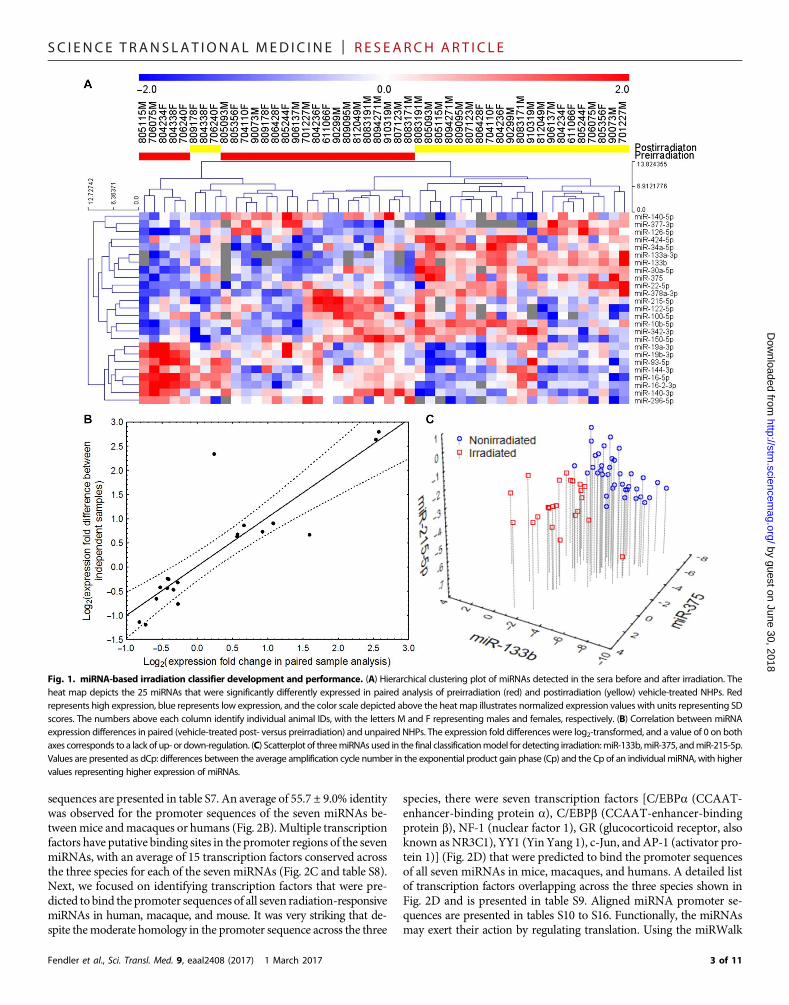
RESULTSSerum miRNA profiling and identification ofradiation-responsive miRNAs in macaquesTo analyze serummiRNAs altered by radiation inmacaques during ini-tial analysis, bloodwas drawn from 48macaques a week before TBI and24 hours after radiation exposure. Pre- and postirradiation samples werepaired for analysis of radiation-responsivemiRNAs. Furthermore, to cor-relate the serummiRNAswith radiation impact, the radioprotective agentGT3 was administered 24 hours before irradiation in 24 macaques (13).BothGT3- and vehicle-treated groupswere divided into three dose levels:5.8 Gy (LD30/60), 6.5 Gy (LD50/60), and 7.2 Gy (LD70/60). The LD repre-sents expected lethality without any supportive care (antibiotics, bloodproducts, and intravenous fluids) to allow for a better demonstration ofthe radioprotective effects of GT3 and to closely simulate amass casualtyscenario where these products are likely to be limited. A total of 651miRNAs were detectable in at least one sample. The average numberof miRNAs detected per sample was 215 ± 44.8, and after filtering formissing data, 174 miRNAs were eligible for analysis. The flowchart ofthe data sets, comparisons, and validation steps (fig. S1) and rawdata fromthe profiling experiment are presented in the Supplementary Material(table S1). Paired pre- and postirradiation samples were compared in thevehicle-treated macaques after removing the batch effect (fig. S2). Thiscomparison identified a list of 25miRNAs that met the significance cri-terion (Fig. 1A and table S2).Among thesemiRNAs, 12were up-regulatedafter irradiation (hsa-miR-133b, hsa-miR-375, hsa-miR-377-3p, hsa-miR-34a-5p, hsa-miR-30a-5p, hsa-miR-424-5p, hsa-miR-342-3p, hsa-
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
miR-10b-5p, hsa-miR-126-5p, hsa-miR-22-5p, hsa-miR-378a-3p, andhsa-miR-133a-3p) and 13 were down-regulated (hsa-miR-215-5p, hsa-miR-122-5p, hsa-miR-150-5p, hsa-miR-144-3p, hsa-miR-140-3p,hsa-miR-16-5p, hsa-miR-19b-3p, hsa-miR-19a-3p, hsa-miR-16-2-3p,hsa-miR-93-5p, hsa-miR-296-5p, hsa-miR-140-5p, and hsa-miR-100-5p). Therewas no significant dose dependence of the radiation-associatedmiRNAs, with variability in expression even within a dose (table S3).This is expected, considering that macaques are not inbred, and each in-dividual animal may have a distinct response to the same dose of radia-tion. Therefore, wemerged the three irradiated groups into one compositeset ofmacaques irradiatedwith a lethal dose of radiation.Having identified25 variable miRNAs in paired analysis, we validated them by comparingthe postirradiation vehicle-treated group with preirradiation samplesallocated to receiveGT3. This result yielded 19miRNAs (table S4) thatshowed significant (P < 0.05) association with radiation in both pairedand independent comparison, convergent in the direction of expressionchange. The expression differences between pre- and postirradiationsamples showed nearly perfect correlation (r = 0.88, P = 0.02; Fig. 1B)between the paired and unpaired comparisons. OnlymiR-133a showeda divergent effect in the unpaired analysis, potentially because it waspresent in very lowquantities andnear the detection threshold innumerouspreirradiation samples.Having established the set of radiation-dependentmiRNAs,weused logistic regressionwith anAkaike information criterion–driven best subset selection to identify an efficient model of separatingall nonirradiated from vehicle-treated irradiatedmacaques. The outcomewas a model based on three miRNAs: miR-133b, miR-215, andmiR-375(Fig. 1C), which showed perfect separation of the groups with an AUC[area under the receiver operating characteristic (ROC) curve] of 1.00[95%confidence interval (CI), 1.00 to 1.00] andmaintained its performancein 10-fold cross-validation (AUC, 1.00; 95%CI, 1.00 to 1.00). Numericalparameters of the model are listed in table S5. Hosmer-Lemeshow’s testshowed a very good fit of themodel (P=1.00; table S6). Because some ofthe animals were missing data for at least one of the miRNAs evaluatedduring model development, 60 samples were eligible for analysis (37preirradiation and 23 postirradiation). A reanalysis with backgroundexpression imputed formissing observations yielded convergent results,with an AUC of 1.00 (95% CI, 1.00 to 1.00), correctly classifying all72 samples.
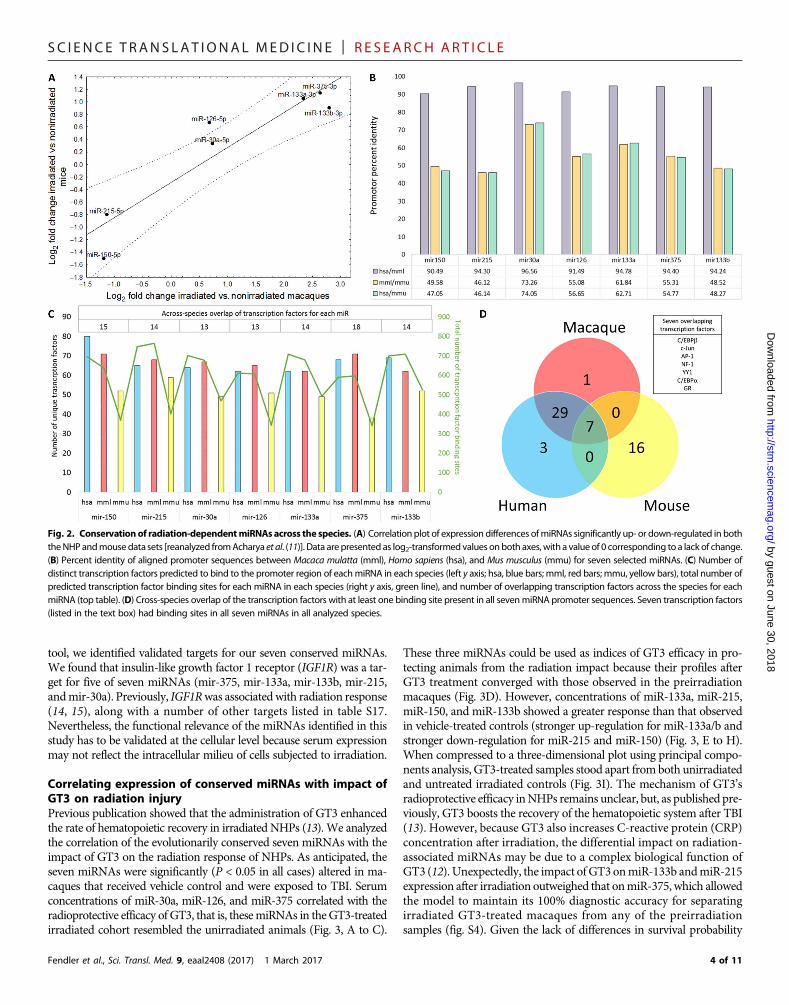
Evolutionary conservation of radiation-responsive miRNAsand their regulationHaving established that irradiated and nonirradiatedNHPsmay be dis-tinguished using a serum miRNA signature, we cross-referenced theradiation-dependent miRNAs with results from our earlier work in amousemodel of irradiation (11). To do so, we reanalyzed themouse dataset by comparing the expression of the 19 miRNAs between 10 controlmice and a pool of 20mice that received 6.5- or 8-Gy radiation exposure,which is comparable to the high, potentially lethal dose used in themacaque experiment. By correlating the fold changes inmiRNAexpres-sion noted in themousemodel and 19miRNAs significantly affected byirradiation in macaques (fig. S3), we identified seven miRNAs thatshowed significant [P < 0.05; false discovery rate (FDR) <0.15] associa-tions with irradiation in both data sets and were convergent in the di-rection of change (Fig. 2A). The expression changes for these sevenmiRNAs showed a strong correlation (r = 0.9303, P = 0.0024) in miceandmacaques. Todeterminewhether thesemiRNAswereunder the con-trol of homologous transcription factors, we interrogated the promotersequences of the miRNAs in mice, macaques, and humans for species-specific transcription factor binding sites. Positions of extracted promoter
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
sequences are presented in table S7. An average of 55.7 ± 9.0% identitywas observed for the promoter sequences of the seven miRNAs be-tweenmice andmacaques or humans (Fig. 2B). Multiple transcriptionfactors have putative binding sites in the promoter regions of the sevenmiRNAs, with an average of 15 transcription factors conserved acrossthe three species for each of the seven miRNAs (Fig. 2C and table S8).Next, we focused on identifying transcription factors that were pre-dicted to bind the promoter sequences of all seven radiation-responsivemiRNAs in human, macaque, and mouse. It was very striking that de-spite themoderate homology in the promoter sequence across the three
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
species, there were seven transcription factors [C/EBPa (CCAAT-enhancer-binding protein a), C/EBPb (CCAAT-enhancer-bindingprotein b), NF-1 (nuclear factor 1), GR (glucocorticoid receptor, alsoknown as NR3C1), YY1 (Yin Yang 1), c-Jun, and AP-1 (activator pro-tein 1)] (Fig. 2D) that were predicted to bind the promoter sequencesof all seven miRNAs in mice, macaques, and humans. A detailed listof transcription factors overlapping across the three species shown inFig. 2D and is presented in table S9. Aligned miRNA promoter se-quences are presented in tables S10 to S16. Functionally, the miRNAsmay exert their action by regulating translation. Using the miRWalk
Fig. 1. miRNA-based irradiation classifier development and performance. (A) Hierarchical clustering plot of miRNAs detected in the sera before and after irradiation. Theheat map depicts the 25 miRNAs that were significantly differently expressed in paired analysis of preirradiation (red) and postirradiation (yellow) vehicle-treated NHPs. Redrepresents high expression, blue represents low expression, and the color scale depicted above the heat map illustrates normalized expression values with units representing SDscores. The numbers above each column identify individual animal IDs, with the letters M and F representing males and females, respectively. (B) Correlation between miRNAexpression differences in paired (vehicle-treated post- versus preirradiation) and unpaired NHPs. The expression fold differences were log2-transformed, and a value of 0 on bothaxes corresponds to a lack of up- or down-regulation. (C) Scatterplot of threemiRNAs used in the final classificationmodel for detecting irradiation:miR-133b,miR-375, andmiR-215-5p.Values are presented as dCp: differences between the average amplification cycle number in the exponential product gain phase (Cp) and the Cp of an individual miRNA, with highervalues representing higher expression of miRNAs.
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
tool, we identified validated targets for our seven conserved miRNAs.We found that insulin-like growth factor 1 receptor (IGF1R) was a tar-get for five of seven miRNAs (mir-375, mir-133a, mir-133b, mir-215,andmir-30a). Previously, IGF1Rwas associatedwith radiation response(14, 15), along with a number of other targets listed in table S17.Nevertheless, the functional relevance of the miRNAs identified in thisstudy has to be validated at the cellular level because serum expressionmay not reflect the intracellular milieu of cells subjected to irradiation.
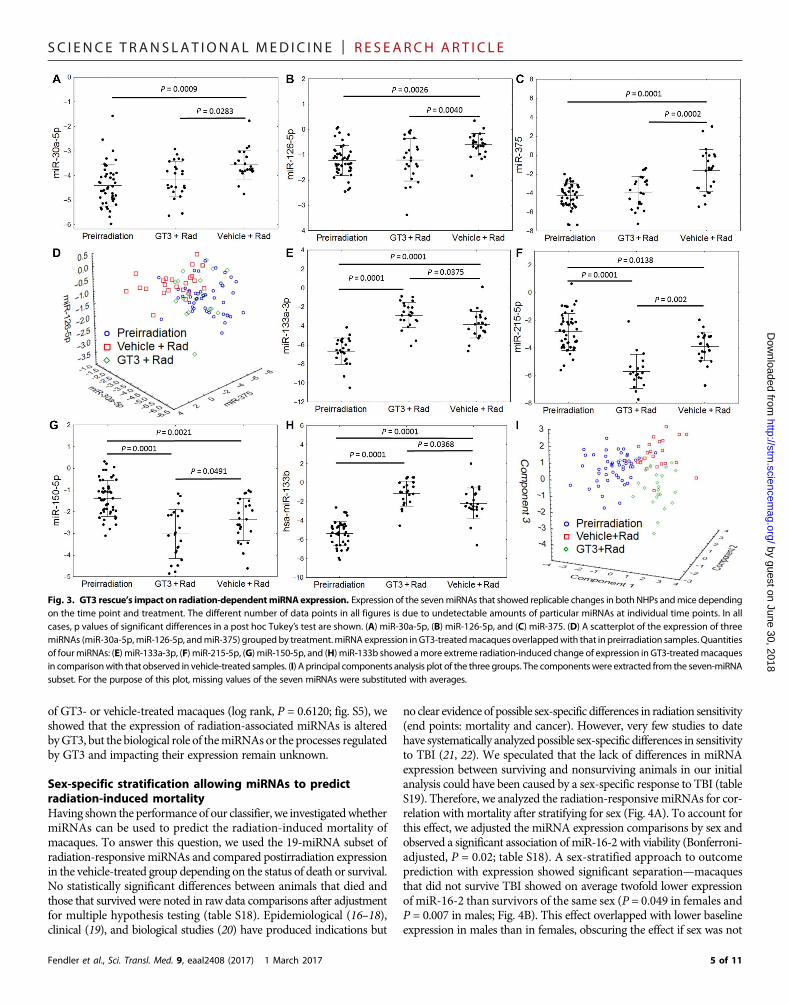
Correlating expression of conserved miRNAs with impact ofGT3 on radiation injuryPrevious publication showed that the administration of GT3 enhancedthe rate of hematopoietic recovery in irradiated NHPs (13). We analyzedthe correlation of the evolutionarily conserved seven miRNAs with theimpact of GT3 on the radiation response of NHPs. As anticipated, theseven miRNAs were significantly (P < 0.05 in all cases) altered in ma-caques that received vehicle control and were exposed to TBI. Serumconcentrations of miR-30a, miR-126, and miR-375 correlated with theradioprotective efficacy of GT3, that is, thesemiRNAs in theGT3-treatedirradiated cohort resembled the unirradiated animals (Fig. 3, A to C).
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
These three miRNAs could be used as indices of GT3 efficacy in pro-tecting animals from the radiation impact because their profiles afterGT3 treatment converged with those observed in the preirradiationmacaques (Fig. 3D). However, concentrations of miR-133a, miR-215,miR-150, and miR-133b showed a greater response than that observedin vehicle-treated controls (stronger up-regulation for miR-133a/b andstronger down-regulation for miR-215 and miR-150) (Fig. 3, E to H).When compressed to a three-dimensional plot using principal compo-nents analysis, GT3-treated samples stood apart fromboth unirradiatedand untreated irradiated controls (Fig. 3I). The mechanism of GT3’sradioprotective efficacy inNHPs remains unclear, but, as published pre-viously, GT3 boosts the recovery of the hematopoietic system after TBI(13). However, because GT3 also increases C-reactive protein (CRP)concentration after irradiation, the differential impact on radiation-associated miRNAs may be due to a complex biological function ofGT3 (12). Unexpectedly, the impact of GT3onmiR-133b andmiR-215expression after irradiation outweighed that onmiR-375, which allowedthe model to maintain its 100% diagnostic accuracy for separatingirradiated GT3-treated macaques from any of the preirradiationsamples (fig. S4). Given the lack of differences in survival probability
Fig. 2. Conservation of radiation-dependentmiRNAs across the species. (A) Correlation plot of expression differences ofmiRNAs significantly up- or down-regulated in boththeNHPandmousedata sets [reanalyzed fromAcharya et al. (11)]. Data arepresented as log2-transformedvalues onboth axes, witha valueof 0 corresponding to a lack of change.(B) Percent identity of aligned promoter sequences between Macaca mulatta (mml), Homo sapiens (hsa), and Mus musculus (mmu) for seven selected miRNAs. (C) Number ofdistinct transcription factors predicted to bind to the promoter region of eachmiRNA in each species (left y axis; hsa, blue bars; mml, red bars; mmu, yellow bars), total number ofpredicted transcription factor binding sites for each miRNA in each species (right y axis, green line), and number of overlapping transcription factors across the species for eachmiRNA (top table). (D) Cross-species overlap of the transcription factors with at least one binding site present in all sevenmiRNA promoter sequences. Seven transcription factors(listed in the text box) had binding sites in all seven miRNAs in all analyzed species.
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
of GT3- or vehicle-treated macaques (log rank, P = 0.6120; fig. S5), weshowed that the expression of radiation-associated miRNAs is alteredbyGT3, but the biological role of themiRNAsor the processes regulatedby GT3 and impacting their expression remain unknown.
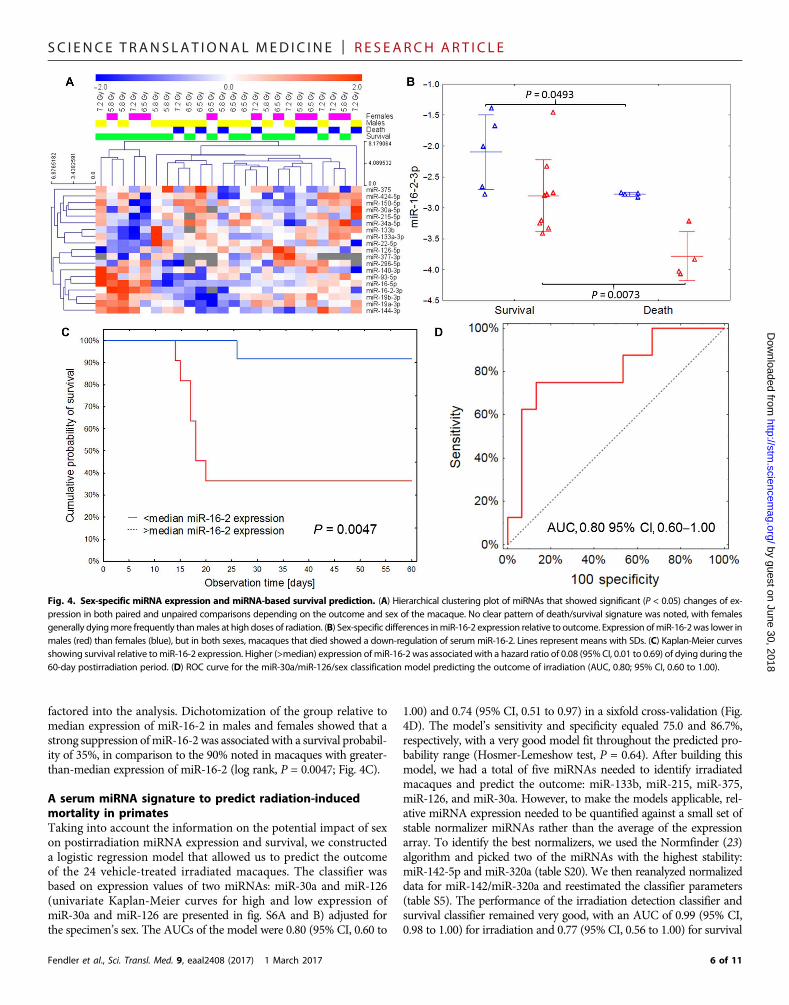
Sex-specific stratification allowing miRNAs to predictradiation-induced mortalityHaving shown the performance of our classifier, we investigatedwhethermiRNAs can be used to predict the radiation-induced mortality ofmacaques. To answer this question, we used the 19-miRNA subset ofradiation-responsive miRNAs and compared postirradiation expressionin the vehicle-treated group depending on the status of death or survival.No statistically significant differences between animals that died andthose that survived were noted in raw data comparisons after adjustmentfor multiple hypothesis testing (table S18). Epidemiological (16–18),clinical (19), and biological studies (20) have produced indications but
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
no clear evidence of possible sex-specific differences in radiation sensitivity(end points: mortality and cancer). However, very few studies to datehave systematically analyzed possible sex-specific differences in sensitivityto TBI (21, 22). We speculated that the lack of differences in miRNAexpression between surviving and nonsurviving animals in our initialanalysis could have been caused by a sex-specific response to TBI (tableS19). Therefore, we analyzed the radiation-responsive miRNAs for cor-relation with mortality after stratifying for sex (Fig. 4A). To account forthis effect, we adjusted the miRNA expression comparisons by sex andobserved a significant association of miR-16-2 with viability (Bonferroni-adjusted, P = 0.02; table S18). A sex-stratified approach to outcomeprediction with expression showed significant separation—macaquesthat did not survive TBI showed on average twofold lower expressionof miR-16-2 than survivors of the same sex (P = 0.049 in females andP = 0.007 in males; Fig. 4B). This effect overlapped with lower baselineexpression in males than in females, obscuring the effect if sex was not
Fig. 3. GT3 rescue’s impact on radiation-dependentmiRNAexpression. Expression of the sevenmiRNAs that showed replicable changes in both NHPs andmice dependingon the time point and treatment. The different number of data points in all figures is due to undetectable amounts of particular miRNAs at individual time points. In allcases, p values of significant differences in a post hoc Tukey’s test are shown. (A) miR-30a-5p, (B) miR-126-5p, and (C) miR-375. (D) A scatterplot of the expression of threemiRNAs (miR-30a-5p,miR-126-5p, andmiR-375) groupedby treatment.miRNA expression inGT3-treatedmacaques overlappedwith that inpreirradiation samples. Quantitiesof fourmiRNAs: (E) miR-133a-3p, (F) miR-215-5p, (G) miR-150-5p, and (H) miR-133b showed amore extreme radiation-induced change of expression in GT3-treatedmacaquesin comparisonwith that observed in vehicle-treated samples. (I) A principal components analysis plot of the three groups. The componentswere extracted from the seven-miRNAsubset. For the purpose of this plot, missing values of the seven miRNAs were substituted with averages.
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
factored into the analysis. Dichotomization of the group relative tomedian expression of miR-16-2 in males and females showed that astrong suppression ofmiR-16-2was associatedwith a survival probabil-ity of 35%, in comparison to the 90% noted in macaques with greater-than-median expression of miR-16-2 (log rank, P = 0.0047; Fig. 4C).
A serum miRNA signature to predict radiation-inducedmortality in primatesTaking into account the information on the potential impact of sexon postirradiation miRNA expression and survival, we constructeda logistic regression model that allowed us to predict the outcomeof the 24 vehicle-treated irradiated macaques. The classifier wasbased on expression values of two miRNAs: miR-30a and miR-126(univariate Kaplan-Meier curves for high and low expression ofmiR-30a and miR-126 are presented in fig. S6A and B) adjusted forthe specimen’s sex. The AUCs of the model were 0.80 (95% CI, 0.60 to
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
1.00) and 0.74 (95% CI, 0.51 to 0.97) in a sixfold cross-validation (Fig.4D). The model’s sensitivity and specificity equaled 75.0 and 86.7%,respectively, with a very good model fit throughout the predicted pro-bability range (Hosmer-Lemeshow test, P = 0.64). After building thismodel, we had a total of five miRNAs needed to identify irradiatedmacaques and predict the outcome: miR-133b, miR-215, miR-375,miR-126, and miR-30a. However, to make the models applicable, rel-ative miRNA expression needed to be quantified against a small set ofstable normalizer miRNAs rather than the average of the expressionarray. To identify the best normalizers, we used the Normfinder (23)algorithm and picked two of the miRNAs with the highest stability:miR-142-5p and miR-320a (table S20). We then reanalyzed normalizeddata for miR-142/miR-320a and reestimated the classifier parameters(table S5). The performance of the irradiation detection classifier andsurvival classifier remained very good, with an AUC of 0.99 (95% CI,0.98 to 1.00) for irradiation and 0.77 (95% CI, 0.56 to 1.00) for survival
Fig. 4. Sex-specific miRNA expression and miRNA-based survival prediction. (A) Hierarchical clustering plot of miRNAs that showed significant (P < 0.05) changes of ex-pression in both paired and unpaired comparisons depending on the outcome and sex of the macaque. No clear pattern of death/survival signature was noted, with femalesgenerally dyingmore frequently thanmales at high doses of radiation. (B) Sex-specific differences inmiR-16-2 expression relative to outcome. Expression ofmiR-16-2was lower inmales (red) than females (blue), but in both sexes, macaques that died showed a down-regulation of serum miR-16-2. Lines represent means with SDs. (C) Kaplan-Meier curvesshowing survival relative tomiR-16-2 expression. Higher (>median) expression ofmiR-16-2 was associatedwith a hazard ratio of 0.08 (95%CI, 0.01 to 0.69) of dying during the60-day postirradiation period. (D) ROC curve for the miR-30a/miR-126/sex classification model predicting the outcome of irradiation (AUC, 0.80; 95% CI, 0.60 to 1.00).
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
bhttp://stm
.sciencemag.org/
Dow
nloaded from
prediction. The association of miR-16-2 persisted for both death (P =0.0116) and sex (P = 0.0003) and was convergent with the direction ofchanges observed in the array average-normalized data set (fig. S7).
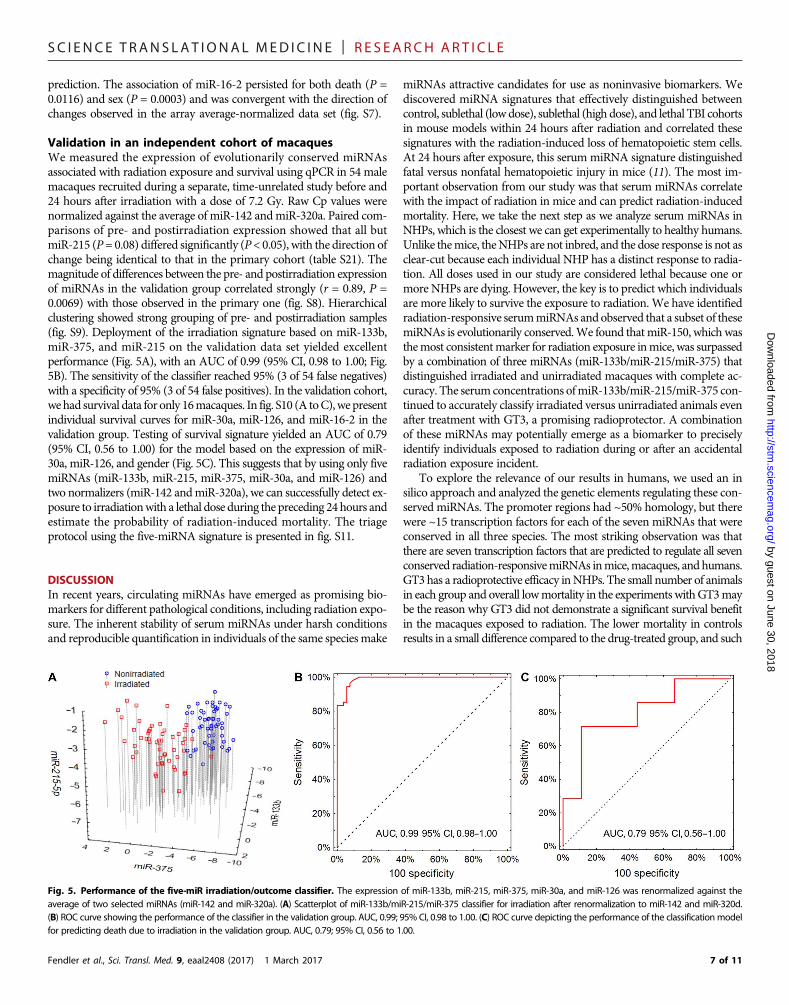
Validation in an independent cohort of macaquesWe measured the expression of evolutionarily conserved miRNAsassociated with radiation exposure and survival using qPCR in 54 malemacaques recruited during a separate, time-unrelated study before and24 hours after irradiation with a dose of 7.2 Gy. Raw Cp values werenormalized against the average of miR-142 andmiR-320a. Paired com-parisons of pre- and postirradiation expression showed that all butmiR-215 (P= 0.08) differed significantly (P< 0.05), with the direction ofchange being identical to that in the primary cohort (table S21). Themagnitude of differences between the pre- and postirradiation expressionof miRNAs in the validation group correlated strongly (r = 0.89, P =0.0069) with those observed in the primary one (fig. S8). Hierarchicalclustering showed strong grouping of pre- and postirradiation samples(fig. S9). Deployment of the irradiation signature based on miR-133b,miR-375, and miR-215 on the validation data set yielded excellentperformance (Fig. 5A), with an AUC of 0.99 (95% CI, 0.98 to 1.00; Fig.5B). The sensitivity of the classifier reached 95% (3 of 54 false negatives)with a specificity of 95% (3 of 54 false positives). In the validation cohort,wehad survival data for only 16macaques. In fig. S10 (A toC),wepresentindividual survival curves for miR-30a, miR-126, and miR-16-2 in thevalidation group. Testing of survival signature yielded an AUC of 0.79(95% CI, 0.56 to 1.00) for the model based on the expression of miR-30a, miR-126, and gender (Fig. 5C). This suggests that by using only fivemiRNAs (miR-133b, miR-215, miR-375, miR-30a, and miR-126) andtwo normalizers (miR-142 andmiR-320a), we can successfully detect ex-posure to irradiationwith a lethal dose during the preceding 24hours andestimate the probability of radiation-induced mortality. The triageprotocol using the five-miRNA signature is presented in fig. S11.
y guest on June 30, 2
DISCUSSIONIn recent years, circulating miRNAs have emerged as promising bio-markers for different pathological conditions, including radiation expo-sure. The inherent stability of serum miRNAs under harsh conditionsand reproducible quantification in individuals of the same speciesmake
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
miRNAs attractive candidates for use as noninvasive biomarkers. Wediscovered miRNA signatures that effectively distinguished betweencontrol, sublethal (lowdose), sublethal (high dose), and lethal TBI cohortsin mouse models within 24 hours after radiation and correlated thesesignatures with the radiation-induced loss of hematopoietic stem cells.At 24 hours after exposure, this serum miRNA signature distinguishedfatal versus nonfatal hematopoietic injury in mice (11). The most im-portant observation from our study was that serum miRNAs correlatewith the impact of radiation in mice and can predict radiation-inducedmortality. Here, we take the next step as we analyze serum miRNAs inNHPs, which is the closest we can get experimentally to healthy humans.Unlike themice, theNHPs are not inbred, and the dose response is not asclear-cut because each individual NHP has a distinct response to radia-tion. All doses used in our study are considered lethal because one ormore NHPs are dying. However, the key is to predict which individualsare more likely to survive the exposure to radiation. We have identifiedradiation-responsive serummiRNAs andobserved that a subset of thesemiRNAs is evolutionarily conserved.We found thatmiR-150, which wasthemost consistentmarker for radiation exposure inmice, was surpassedby a combination of three miRNAs (miR-133b/miR-215/miR-375) thatdistinguished irradiated and unirradiated macaques with complete ac-curacy. The serumconcentrations ofmiR-133b/miR-215/miR-375 con-tinued to accurately classify irradiated versus unirradiated animals evenafter treatment with GT3, a promising radioprotector. A combinationof these miRNAs may potentially emerge as a biomarker to preciselyidentify individuals exposed to radiation during or after an accidentalradiation exposure incident.
To explore the relevance of our results in humans, we used an insilico approach and analyzed the genetic elements regulating these con-served miRNAs. The promoter regions had ~50% homology, but therewere ~15 transcription factors for each of the seven miRNAs that wereconserved in all three species. The most striking observation was thatthere are seven transcription factors that are predicted to regulate all sevenconserved radiation-responsivemiRNAs inmice,macaques, and humans.GT3 has a radioprotective efficacy inNHPs. The small number of animalsin each group and overall lowmortality in the experiments withGT3maybe the reason why GT3 did not demonstrate a significant survival benefitin the macaques exposed to radiation. The lower mortality in controlsresults in a small difference compared to the drug-treated group, and such
018
Fig. 5. Performance of the five-miR irradiation/outcome classifier. The expression of miR-133b, miR-215, miR-375, miR-30a, and miR-126 was renormalized against theaverage of two selected miRNAs (miR-142 and miR-320a). (A) Scatterplot of miR-133b/miR-215/miR-375 classifier for irradiation after renormalization to miR-142 and miR-320d.(B) ROC curve showing the performance of the classifier in the validation group. AUC, 0.99; 95% CI, 0.98 to 1.00. (C) ROC curve depicting the performance of the classificationmodelfor predicting death due to irradiation in the validation group. AUC, 0.79; 95% CI, 0.56 to 1.00.
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
experiments need a large sample size to observe statistically significantdifferences between groups. However, three (miR-30a, miR-126, andmiR-375) of the seven conserved radiation-responsive serum miRNAscorrelated with GT3, that is, the expression of miR-30a, miR-126, andmiR-375 was unchanged by irradiation only when GT3 had been ad-ministered before exposure. Therefore, it is feasible that these miRNAsmay serve as biomarkers for GT3 efficacy, particularly because two ofthem, miR-30a and miR-126, were reproducibly associated with theoutcome of irradiation as well as with GT3 response.
Early studies inmice have indicated that females were, in general, 1.5times more sensitive to x-ray irradiation than males of the same strain,suggesting a constant effect of sex (21). However, for humans, becauseof the limited data availability with regard to both clinical studies andbiological experiments, as well as sometimes conflicting findings of ep-idemiological studies, it has not been possible to make unequivocalstatements about sex-specific differences in radiation sensitivity. Therehave been no systematic studies in NHPs that demonstrate the correla-tion of sex with radiation-induced mortality. Our results are consistentwith the mouse studies showing that TBI-induced mortality is morepronounced in female macaques. However, by taking sex into account,we discovered that the decrease of a single miRNA, miR-16-2, shows asex-specific pattern of expression differing between macaques that expe-rience fatal versus nonfatal radiation injury. Furthermore, by combiningthe radiation biomarker miRNAs (miR-133b, miR-215, and miR-375),the survival indicators (miR-30a and miR-126), and the normalizermiRNAs (miR-142 and miR-320a) into a compound classifier, we cansuccessfully detect exposure to radiation and accurately estimate theprobability of death within 24 hours after exposure (table S5). Consid-ering that these results were generated using a large number of NHPs,we anticipate that the results would extrapolate to a human radiationexposure scenario.
A limitation of our study is the restricted ability to experimentallyvalidate results in NHPs. For example, it is striking that the same seventranscription factors are predicted to regulate the radiation-responsiveserummiRNAs in humans,macaques, andmice.However, these resultsneed to be experimentally validated, and our ability to do so has beenlimited for the following reasons. First, we do not know the source (celltype) of the serum miRNAs, and second, only a limited range of experi-ments are feasible with macaques. Here, we used existing serum samplesfromNHPs collected before and after irradiation.Although the number ofsamples was large enough to obtain the results reported here, these shouldbe further validatedwith a larger number of independentNHP samples toaccount for potential sources of variation in the real world—tissuehandling, varying collection times, and technical factors.
MATERIALS AND METHODSStudy designThe aim of the study was to identify miRNA signatures for efficient de-tection of high-dose irradiation and radiation-induced mortality in pri-mates. The study used 48 animal samples from a study with GT3described in a previous publication (13). That is, the animals weresubjected to randomization and given GT3 or vehicle before irradiationwith three dose levels. No blinding was introduced at that stage. Serumsamples were collected before and after irradiation, and after RNA iso-lation, miRNA expression was quantified by qPCR. After refining thesignatures for irradiation detection and survival prediction, a replicationcohort of additional 54 male macaques from a separate, blinded studywas used to test the performance of the developed models.
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
AnimalsHealthy naïve rhesusmacaques (M.mulatta, Chinese substrain; n = 48)for initial analysis were obtained from Primate Products Inc. and quar-antined for 6 to 7 weeks before the start of the experiment. Clinicallyhealthy male and female rhesus macaques 3 to 5 years of age, weighing3.6 to 8.4 kg, were used in this study.AllNHPswere individually housedin stainless steel cages in environmentally controlled roomsmaintainedat 22° ± 2°C, 30 to 70% relative humidity, 10 to 15 air change cycles perhour, and 12-hour light/12-hour dark cycle. Animals were fed with pri-mate diet (Harlan Teklad T.2050 diet) twice daily and received drinkingwater ad libitum.All the animals received enrichment food (fresh fruits andvegetables, prima treats, peanuts, marshmallows, etc.) once a day,Mondayto Friday. They received mirrors, toys, and challenge balls for enrichment.Televisionswere used for sensory enrichment for 4 to 5 hours, at least threetimes a week. All the animals were able to see, hear, and/or touch theirconspecifics through the cages. All animals were serologically negativeforMacacine herpesvirus 1 (herpes B virus), simian retrovirus, simianT cell leukemia virus, and simian immunodeficiency virus. They werevaccinated formeasles, confirmed to have positive antibody titers, andtested negative for tuberculin. Animals were stratified by sex and bybody weight increases during the quarantine period and then assignedto different treatment groups. This animal study was conducted in afacility accredited by the Association for Assessment and Accredita-tion of Laboratory Animal Care International.
Validation groupThe independent validation cohort consisted of an additional 54 malerhesusmacaques (M.mulatta, Chinese substrain; 3.5 to 5.5 years of age,weighing 4.0 to 8.0 kg) procured from the National Institutes of HealthAnimal Facility. All other details for this cohort were the same as de-scribed above for the initial study analysis. Blood samples were collectedfor serum 1week before and 24 hours after irradiation as in the primarycohort. Of 54 irradiated animals in the validation cohort, 16 were usedfor the survival study,making them eligible for validation of the survivalsignature.
Drug preparation and administrationGT3 (50 or 100 mg/ml) in 5% Tween-80 in saline was purchasedfrom Yasoo Health Inc. The quantity of GT3 for each NHP wasbased on individual NHP weight. Drug and vehicle were administeredat the dorsal scapular area (between the shoulder blades). The areasurrounding the injection site was shaved at least 48 hours beforethe administration of drug, so the site could be easily observed forany adverse skin reactions such as rash/eruption, inflammation, irri-tation, or abscess formation after GT3 or vehicle administration. Im-mediately before drug injection, the site was wiped with 70% isopropylrubbing alcohol and allowed to air dry; the drug was administeredsubcutaneously using a 3-ml disposable luer lock syringe with a 25-gauge5/8-inch needle.
Radiation exposureA maximum of 4 NHPs in the primary cohort and 14 animals in thevalidation cohort were irradiated per day. Food was withheld from eachanimal for about 12 to 18 hours before exposure tominimize the occur-renceof radiation-inducedvomiting.About 30 to45minbefore irradiation,ketaminehydrochloride (10 to15mg/kg)was intramuscularly administeredtoNHPs for sedation, and thenplaced in custom-madePlexiglas irradiationboxes and secured in a seated position. TwoNHPs at a timewere placedon the irradiation platform facing away from each other and exposed
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
with amidline dose of 5.8, 6.5, or 7.2 Gy at a dose rate of 0.6 Gy/min. Todeliver the precise dose, NHPs’ abdominal widths were measured withdigital calipers. Animals were observed throughout the irradiation pro-cedure via in-room cameras. After irradiation, animals were returned tothe transport cart and to their cages in the housing area and monitoredfor recovery from the procedure. Dose rate measurements were basedprimarily on the alanine/electron paramagnetic resonance (EPR) system,currently accepted as one of themost accuratemethods for relatively highradiation doses and used for comparisons between national metrologyinstitutions (24). The calibration curves (EMXmicro spectrometer, BrukerCorp.) used in dose measurements at the Armed Forces RadiobiologyResearch Institute (AFRRI) are based on standard alanine calibration setspurchased from theU.S.National Institute of Standards andTechnology(NIST). The alanine dosimeters obtained fromNISThad been calibratedin terms of absorbed dose to water using the U.S. National StandardRadiation Sources. At AFRRI, identical alanine dosimeters were placedmidline withinNHP phantoms (Plexiglas cylinders 6.9, 10, and 12.5 cmin diameter and 34.5 cm in length) and irradiated to about 100Gy.Mea-surement of their EPR signals using the calibration curve constructedwith alanine dosimeters from NIST provided dose rates to water in thecore bodies ofNHP.A small correction (0.9906)was subsequently appliedto adjust for the difference in mass energy absorption coefficients be-tween water and soft tissue.
To assess the radioprotective efficacy of GT3 as a radiation counter-measure, 48 NHPs from the initial analysis cohort were divided intothree groups of 16 animals, and each groupwas exposed to the followinglethal doses of radiation: 5.8 Gy (LD30/60), 6.5 Gy (LD50/60), and 7.2 Gy(LD70/60) (0.6Gy/min).GT3was administered to theNHPs subcutaneously24 hours before radiation exposure.
ThoseNHPs exposed to 5.8Gywere further divided into two groupsof eightNHPs each, with one group receivingGT3 at 37.5mg/kg (n= 4)or vehicle (n = 4) and the other receiving GT3 at 75 mg/kg (n = 4) orvehicle (n = 4). All animals exposed to 6.5 and 7.2 Gy received GT3 at37.5 mg/kg or vehicle. In all experimental groups, 50% of animals wereinjectedwithGT3, and the remaining 50%were controls (vehicle-treated).
All 54 animals in the validation cohort did not receive any test drugbefore blood collection, which took place at either−7 days or +24 hours.Similarly, the 16 animals in the validation cohort used for the survivalstudy did not receive any test drug during the course of the study.
Blood sample collection from NHPsBloodwas collected from chair-restrainedNHPs via the saphenous veinlocated superficially on the caudal aspect of the lower leg, distal to theknee. At 48 hours before the first blood collection, the hairs surroundingthe collection site were shaved using an electric cordless shaver. Forvenipuncture, a tourniquet was applied to the hind leg below the knee,and the site was cleaned using a 70% isopropyl alcohol wipe and driedwith sterile gauze. The desired volume of blood was collected with a3-ml disposable luer lock syringe with a 25-gauge needle. For serumcollection, the blood samplewas transferred toCapiject serum separatortubes (3T-MG, TerumoMedical Corp.) and allowed to clot for 30 min,then centrifuged at 400g for 10 min. The serum samples were stored at−70°C until use.
Sample selection and study protocolSamples evaluated within this study for initial analysis and validationoriginated from different studies conducted at different times (at least2 years apart). Bloodhadbeendrawnatmultiple time points in the originalprotocol, but for the purpose of identifying early irradiation biomarkers
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
and predictors of survival, we profiledmiRNA expression in the −7-daysample and the one collected 24 hours after irradiation.
miRNA profilingmiRNA profiling was performed as described previously (11, 25) usingqPCR panels from Exiqon with LNA (locked nucleic acid)-modified pri-mers. PCR panels are accurate and convergent with miRNA sequencingdata, provided that the number of copies of anmiRNAexceeds 10 permil-lion (26). Because nomacaque-specificmiRNApanelswere available at thetime of the study, human panels I and II were used instead, and qPCRsignals were decoded using H. sapiens miRNA annotations. Profilingwas performed at Exiqon, and all the reagents for the RNA isolation wereobtained from Exiqon. After blood collection from the macaques, thesampleswere centrifuged at 1000g for 15min to separate the serum. Beforeprofiling, serum samples were thawed on ice and centrifuged at 3000g for5min in a4°Cmicrocentrifuge.Analiquotof 200ml (75ml of serum+125mlofwater)per samplewas transferred toanewmicrocentrifuge tube, and60mlof Lysis Solution BF containing 1 mg of carrier RNA per 60 ml of LysisSolutionBF andRNAspike-in templatemixturewas added to the sample.The total amount of RNA used in the analysis was 65 ng. The tube wasvortexed and incubated for 3min at room temperature, followed by theaddition of 20 ml of Protein Precipitation Solution BF. The tube wasvortexed, incubated for 1 min at room temperature, and centrifuged at11,000g for 3 min. The clear supernatant was transferred to a new collec-tion tube, and 270 ml of isopropanol was added. The solution was vor-texed and transferred to a binding column. The column was incubatedfor 2 min at room temperature and emptied using a vacuummanifold.A 100-mlWash Solution 1 BFwas added to the columns. The liquid wasremoved using a vacuum manifold, 700 ml of Wash Solution 2 BF wasadded, and the columnwas spun at 11,000g to dry the columns entirely.The dry columns were transferred to a new collection tube, and 50 ml ofribonuclease-free H2O was added directly to the membrane of the spincolumn. The column was incubated for 1 min at room temperaturebefore centrifugation at 11,000g. The RNAwas stored in a−80°C freezer.RNA(19ml)was reverse-transcribed in95-ml reactionsusing themiRCURYLNA Universal RT microRNA PCR, Polyadenylation and cDNA syn-thesis kit (Exiqon). Complementary DNAwas diluted 50× and assayedin 10 ml of PCRs according to the protocol formiRCURYLNAUniversalRT microRNA PCR; each miRNA was assayed once by qPCR on themiRNAReady-to-UsePCR, panel I+II usingExiLENTSYBRGreenmastermix. Negative controls excluding template from the reverse transcriptionreaction were analyzed in the same way as the samples. The amplificationwas performed in a LightCycler 480 Real-Time PCR System (Roche) in384-well plates. The amplification curveswere analyzed using theRocheLC software, both for determination of Cp (by the second derivativemethod) and for melting curve analysis.
Ethics statementAll procedures involving animals were approved by the AFRRI Institu-tional Animal Care and Use Committee and the second tier Depart-ment of Defense Animal Care and Use Review Office. This study wascarried out in strict accordance with the recommendations in theGuidefor the Care and Use of Laboratory Animals of the National Institutes ofHealth (27).
Statistical analysisPreprocessing of profiling data.The data set was filtered for miRNAs that were present in 50% of bothpre- and postirradiation samples, with an additional restriction that the
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
miRNA should be detectable in at least two samples from every drug/dose/time point block. Missing data were imputed with 37 cycles as athreshold for miRNA detection. After filtering the data set for miRNAdetection frequency, we evaluated it for potential batch effects resultingfrom the use of different generation Exiqon arrays and removed themusing ComBat—an empirical Bayes method of identifying and removingbatch effect (fig. S2) (28).Statistical methods.Principal components analysis was used for batch effect evaluation.Hierarchical clusteringwith Euclidean distance was used for visualizationof the data set. Comparisons ofmiRNA inpaired pre- and postirradiationsamples were performed using a paired t test. Statistical significance forthis step was stated if a givenmiRNAdiffered with a P value of <0.05 andshowed an FDR (calculated using the Benjamini-Hochberg method) of<0.15. This fairly relaxed significance criterion was established to accountfor pairs dropping out of the analysis if a single pre- or postirradiationsample yielded no miRNA detection. The validation with independentsamples was done using a standard t test under the same threshold forsignificance as in the paired analysis (P < 0.05; FDR, <0.15). The samethreshold was used for identifyingmiRNAs associated with survival. Sex-adjusted comparisons were performed using an analysis of covariancemodel. Logistic regression models for irradiation detection and outcomeprediction were created using an Akaike information criterion–drivenbest subset variable selection. Goodness of fit was evaluated using theHosmer-Lemeshow test (29). Diagnostic accuracy of the model wasevaluated by calculating the AUC. The regression models for radiationeffect identification and outcome prediction were created using 10-foldcross-validation and 6-fold cross-validation, depending on the numberof samples in the two analyses [n = 72 for the irradiation model (all 48preirradiation and 24 vehicle-treated postirradiation) and n = 24 for thesurvival model (only vehicle-treated postirradiation), respectively]. Giventhatweneeded touse all samples for the analysis, adjusting for batch effectand filtering the variables beforemodel development, a full cross-validationas defined in Simon et al. (30) was not performed.Multiple group com-parisonswere performedwith one-way analysis of variance, followed byTukey’s honest significant difference post hoc test for between-groupcomparisons. Kaplan-Meier curves were used to present the outcome,with log-rank test used to confirm statistical significance for differencesin cumulative probabilities of survival between the groupswith differentmiRNA expression. Reanalysis of the Acharya et al. (11) data set wasdone using a standard t test. Correlations were evaluated using Pearson’scorrelation coefficient. For AUCs, 95% CIs were computed, and alltests were two-sided. Threshold for significance for univariate com-parisons was established at P < 0.05. All analyses were done usingStatistica 12.5 (Statsoft) and MultiExperiment Viewer (Dana-FarberCancer Institute).Bioinformatic analyses.Promoter sequences of miRNAs that showed significant changes in thesame direction in bothmacaque andmice data sets were extracted fromGenBank using the University of California, Santa Cruz GenomeBrowser (31) for M. mulatta [November 2015 (BCM Mmul_8.0.1/rheMac8)], M. musculus [December 2011 (GRCm38/mm10)], andH. sapiens [December 2013 (GRCh38/hg38)] (table S7). For intragenicmiRNAs, the analysis of promoter transcription factor binding sites wasperformed for both the miRNA itself and an upstream region of thegene in which the miRNA was located. In each case, the sequences of1500 base pairs upstream to the 5′ end of the gene sequence wereextractedand imported intoPROMO(32), a tool thatuses theTRANSFACdatabase (33), to identify specific transcription factor binding sites in a
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
given sequence and species. ForM.mulatta, weusedprimate transcriptionfactors and binding sites because theM.mulatta datawere not available.ForM.musculus, we usedmouse transcription factors and binding sites,and forH. sapiens, only human ones. The output (transcription factorsbinding to the interrogated genomic sequences from the three species)was cross-referenced to identify common regulatory elements present onan miRNA basis and universal for all miRNAs with similar patterns ofchange after radiation. To check the conservation of promoter sequencesacross the three analyzed species, we used Clustal Omega (34), an onlinemultiple sequence alignment program that can compare three or moresequences by usage of seeded guide trees and hidden Markov modelprofile-profile techniques. Alignment of the three sequences was showninClustalwithout thenumbers format (tables S10 to S16). Percent identityof each pair of promoter sequences between the species was calculated.
SUPPLEMENTARY MATERIALSwww.sciencetranslationalmedicine.org/cgi/content/full/9/379/eaal2408/DC1Fig. S1. Flowchart of miRNA expression comparisons.Fig. S2. Batch effect removal.Fig. S3. Overlap of radiation-induced changes of miRNA expression in the mouse and macaqueexperiments.Fig. S4. Performance of the three-miRNA irradiation classifier on vehicle- and GT3-treatedsamples.Fig. S5. Survival of vehicle- and GT3-treated macaques.Fig. S6. Survival curves for miRNAs retained in the survival prediction model.Fig. S7. Differences of miR-16-2 in data normalized to miR-142/miR-320a average.Fig. S8. Correlation of miRNA expression between the primary and validation groups.Fig. S9. Hierarchical clustering of miRNA expression in the validation group.Fig. S10. Survival curves for miR-30a, miR-126, and miR-16-2 in the validation group.Fig. S11. The five-miRNA diagnostic protocol for irradiation detection and outcome prediction.Table S1. Raw expression data from miRNA qPCR profiling (separate .xls file).Table S2. Paired sample miRNA expression comparisons (separate .xls file).Table S3. miRNA expression in macaques irradiated with different doses.Table S4. Independent sample miRNA expression comparisons (separate .xls file).Table S5. Numerical parameters of logistic regression models.Table S6. Goodness-of-fit evaluation of the irradiation detection model.Table S7. Promoter sequences of miRNAs with similar patterns of radiation-induced expressionchanges.Table S8. Transcription factor binding sites in miRNA promoters of human, macaque, andmouse genomes (separate .xls file).Table S9. Overlap of transcription factor binding sites across the three species.Table S10. Promoter sequences of miR-30a (separate .txt file).Table S11. Promoter sequences of miR-126 (separate .txt file).Table S12. Promoter sequences of miR-375 (separate .txt file).Table S13. Promoter sequences of miR-150 (separate .txt file).Table S14. Promoter sequences of mir-133b (separate .txt file).Table S15. Promoter sequences of miR-133a (separate .txt file).Table S16. Promoter sequences of miR-215 (separate .txt file).Table S17. Validated gene targets of radiation-associated evolutionarily conserved miRNAs(separate .xls file).Table S18. Comparisons of miRNA expression depending on outcome.Table S19. Differences of outcome frequencies depending on sex.Table S20. Selection of best normalizer miRNAs (separate .xls file).Table S21. Differences of radiation-induced miRNA expression in the validation group.
REFERENCES AND NOTES1. F. K. C. Chin, Scenario of a dirty bomb in an urban environment and acute management
of radiation poisoning and injuries. Singapore Med. J. 48, 950–957 (2007).2. J. K. Waselenko, T. J. MacVittie, W. F. Blakely, N. Pesik, A. L. Wiley, W. E. Dickerson,
H. Tsu, D. L. Confer, C. N. Coleman, T. Seed, P. Lowry, J. O. Armitage, N. Dainiak;Strategic National Stockpile Radiation Working Group, Medical management of theacute radiation syndrome: Recommendations of the Strategic National StockpileRadiation Working Group. Ann. Intern. Med. 140, 1037–1051 (2004).
3. C. N. Coleman, H. B. Stone, J. E. Moulder, T. C. Pellmar, Modulation of radiation injury.Science 304, 693–694 (2004).
SC I ENCE TRANS LAT IONAL MED I C I N E | R E S EARCH ART I C L E
by guest on June 30, 2018http://stm
.sciencemag.org/
Dow
nloaded from
4. N. Dainiak, Radiation response: Changing concepts and emerging paradigms. Exp.Hematol. 31, 435–436 (2003).
5. N. Dainiak, J. K. Waselenko, J. O. Armitage, T. J. MacVittie, A. M. Farese, The hematologistand radiation casualties. Hematology Am. Soc. Hematol. Educ. Program 2003,473–496 (2003).
6. D. D. Parker, J. C. Parker, Estimating radiation dose from time to emesis and lymphocytedepletion. Health Phys. 93, 701–704 (2007).
7. R. E. Gowns, E. C. Holloway, M. E. Berger, R. C. Ricks, Early dose assessment followingsevere radiation accidents. Health Phys. 72, 513–518 (1997).
8. J. M. Sullivan, P. G. S. Prasanna, M. B. Grace, L. K. Wathen, R. L. Wallace, J. F. Koerner,C. N. Coleman, Assessment of biodosimetry methods for a mass-casualtyradiological incident: Medical response and management considerations. Health Phys.105, 540–554 (2013).
9. M. M. P. de Lemos Pinto, N. F. G. Santos, A. Amaral, Current status of biodosimetry basedon standard cytogenetic methods. Radiat. Environ. Biophys. 49, 567–581 (2010).
10. V. K. Singh, V. L. Newman, P. L. P. Romaine, M. Hauer-Jensen, H. B. Pollard, Use ofbiomarkers for assessing radiation injury and efficacy of countermeasures. Expert Rev.Mol. Diagn. 16, 65–81 (2016).
11. S. S. Acharya, W. Fendler, J. Watson, A. Hamilton, Y. Pan, E. Gaudiano, P. Moskwa,P. Bhanja, S. Saha, C. Guha, K. Parmar, D. Chowdhury, Serum microRNAs are earlyindicators of survival after radiation-induced hematopoietic injury. Sci. Transl. Med. 7,287ra69 (2015).
12. V. K. Singh, M. Hauer-Jensen, g-Tocotrienol as a promising countermeasure for acuteradiation syndrome: Current status. Int. J. Mol. Sci. 17, E663 (2016).
13. V. K. Singh, S. Kulkarni, O. O. Fatanmi, S. Y. Wise, V. L. Newman, P. L. P. Romaine,H. Hendrickson, J. Gulani, S. P. Ghosh, K. S. Kumar, M. Hauer-Jensen, Radioprotectiveefficacy of gamma-tocotrienol in nonhuman primates. Radiat. Res. 185, 285–298 (2016).
14. U. Raju, D. P. Molkentine, D. R. Valdecanas, A. Deorukhkar, K. A. Mason, T. A. Buchholz,R. E. Meyn, K.-K. Ang, H. Skinner, Inhibition of EGFR or IGF-1R signaling enhancesradiation response in head and neck cancer models but concurrent inhibition has noadded benefit. Cancer Med. 4, 65–74 (2015).
15. M. M. Chitnis, K. A. Lodhia, T. Aleksic, S. Gao, A. S. Protheroe, V. M. Macaulay, IGF-1Rinhibition enhances radiosensitivity and delays double-strand break repair by both non-homologous end-joining and homologous recombination. Oncogene 33, 5262–5273(2014).
16. M. W. Charles, Studies of mortality of atomic bomb survivors. Report 13: Solid cancer andnoncancer disease mortality: 1950–1997. J. Radiol. Prot. 23, 457–459 (2003).
17. D. L. Preston, Y. Shimizu, D. A. Pierce, A. Suyama, K. Mabuchi, Studies of mortality ofatomic bomb survivors. Report 13: Solid cancer and noncancer disease mortality: 1950–1997. Radiat. Res. 160, 381–407 (2003).
18. D. L. Preston, Y. Shimizu, D. A. Pierce, A. Suyama, K. Mabuchi, Studies of mortality ofatomic bomb survivors. Report 13: Solid cancer and noncancer disease mortality: 1950–1997. Radiat. Res. 178, AV146–AV172 (2012).
19. K. Borgmann, E. Dikomey, C. Petersen, P. Feyer, U. Hoeller, Sex-specific aspects of tumortherapy. Radiat. Environ. Biophys. 48, 115–124 (2009).
20. O. Kovalchuk, A. Ponton, J. Filkowski, I. Kovalchuk, Dissimilar genome response to acuteand chronic low-dose radiation in male and female mice. Mutat. Res. 550, 59–72 (2004).
21. J. M. Holland, T. J. Mitchell, The relationship of strain, sex, and body weight to survivalfollowing sublethal whole-body x-irradiation. Radiat. Res. 66, 363–372 (1976).
22. B. A. Carnes, N. Gavrilova, D. Grahn, Pathology effects at radiation doses below thosecausing increased mortality. Radiat. Res. 158, 187–194 (2002).
23. C. L. Andersen, J. L. Jensen, T. F. Ørntoft, Normalization of real-time quantitative reversetranscription-PCR data: A model-based variance estimation approach to identify genessuited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64,5245–5250 (2004).
24. M. Anton, Uncertainties in alanine/ESR dosimetry at the Physikalisch-TechnischeBundesanstalt. Phys. Med. Biol. 51, 5419–5440 (2006).
25. W. Fendler, J. Madzio, K. Kozinski, K. Patel, J. Janikiewicz, M. Szopa, A. Tracz, M. Borowiec,P. Jarosz-Chobot, M. Mysliwiec, A. Szadkowska, A. T. Hattersley, S. Ellard,M. T. Malecki, A. Dobrzyn, W. Mlynarski, Differential regulation of serum microRNA
Fendler et al., Sci. Transl. Med. 9, eaal2408 (2017) 1 March 2017
expression by HNF1b and HNF1a transcription factors. Diabetologia 59,1463–1473 (2016).
26. P. Mestdagh, N. Hartmann, L. Baeriswyl, D. Andreasen, N. Bernard, C. Chen, D. Cheo,P. D’Andrade, M. DeMayo, L. Dennis, S. Derveaux, Y. Feng, S. Fulmer-Smentek,B. Gerstmayer, J. Gouffon, C. Grimley, E. Lader, K. Y. Lee, S. Luo, P. Mouritzen,A. Narayanan, S. Patel, S. Peiffer, S. Rüberg, G. Schroth, D. Schuster, J. M. Shaffer,E. J. Shelton, S. Silveria, U. Ulmanella, V. Veeramachaneni, F. Staedtler, T. Peters,T. Guettouche, L. Wong, J. Vandesompele, Evaluation of quantitative miRNA expressionplatforms in the microRNA quality control (miRQC) study. Nat. Methods 11, 809–815(2014).
27. Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (TheNational Academic Press, ed. 8, 2011).
28. W. E. Johnson, C. Li, A. Rabinovic, Adjusting batch effects in microarray expression datausing empirical Bayes methods. Biostatistics 8, 118–127 (2007).
29. D. W. L. Hosmer, Stanley Applied Logistic Regression (Wiley, 2013).30. R. M. Simon, J. Subramanian, M.-C. Li, S. Menezes, Using cross-validation to evaluate
predictive accuracy of survival risk classifiers based on high-dimensional data. Brief.Bioinform. 12, 203–214 (2011).
31. W. J. Kent, C. W. Sugnet, T. S. Furey, K. M. Roskin, T. H. Pringle, A. M. Zahler, D. Haussler,The human genome browser at UCSC. Genome Res. 12, 996–1006 (2002).
32. X. Messeguer, R. Escudero, D. Farré, O. Núñez, J. Martínez, M. M. Albà, PROMO: Detectionof known transcription regulatory elements using species-tailored searches.Bioinformatics 18, 333–334 (2002).
33. V. Matys, O. V. Kel-Margoulis, E. Fricke, I. Liebich, S. Land, A. Barre-Dirrie, I. Reuter,D. Chekmenev, M. Krull, K. Hornischer, N. Voss, P. Stegmaier, B. Lewicki-Potapov,H. Saxel, A. E. Kel, E. Wingender, TRANSFAC® and its module TRANSCompel®:Transcriptional gene regulation in eukaryotes. Nucleic Acids Res. 34, D108–D110(2006).
34. F. Sievers, A. Wilm, D. Dineen, T. J. Gibson, K. Karplus, W. Li, R. Lopez, H. McWilliam,M. Remmert, J. Söding, J. D. Thompson, D. G. Higgins, Fast, scalable generationof high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol.7, 539 (2011).
Acknowledgments: The views expressed do not necessarily represent the opinions orpolicies of the AFRRI, the F. Edward Hébert School of Medicine, the UniformedServices University of the Health Sciences, the Department of Defense, or the U.S.government. Funding: D.C. is supported by grants R01 AI101897-01 (National Instituteof Allergy and Infectious Diseases) and R01CA142698-07 (National Cancer Institute),the Leukemia and Lymphoma Society Scholar grant, the Claudia Adams Barr Programfor Innovative Cancer Research, the Robert and Deborah First Family Fund, and theTina Brozman Foundation. V.K.S. received grant support from the Defense ThreatReduction Agency (CBM.RAD.01.10.AR.005) and the Congressionally Directed MedicalResearch Program (W81XWH-15-C-0117 and JW140032), Department of Defense.Author contributions: W.F. conducted all the statistical analysis with help from B.M.and K.M. P.A.K. independently validated the analysis. The NHP irradiation and subsequentblood analysis were done under the supervision of V.K.S.; the serum miRNA processingand analysis were conducted in D.C.’s laboratory with help from C.G. The manuscriptwas written by W.F., V.K.S., and D.C. with input from P.K. and C.G. Competing interests:The authors declare that they have no competing interests.
Submitted 11 August 2016Resubmitted 19 December 2016Accepted 10 February 2017Published 1 March 201710.1126/scitranslmed.aal2408
Citation: W. Fendler, B. Malachowska, K. Meghani, P. A. Konstantinopoulos, C. Guha,V. K. Singh, D. Chowdhury, Evolutionarily conserved serum microRNAs predict radiation-induced fatality in nonhuman primates. Sci. Transl. Med. 9, eaal2408 (2017).
nonhuman primatesEvolutionarily conserved serum microRNAs predict radiation-induced fatality in
Singh and Dipanjan ChowdhuryWojciech Fendler, Beata Malachowska, Khyati Meghani, Panagiotis A. Konstantinopoulos, Chandan Guha, Vijay K.
DOI: 10.1126/scitranslmed.aal2408, eaal2408.9Sci Transl Med
well as their chances of survival.miRNAs (adjusted for gender), the authors were able to identify which primates had been exposed to radiation as then analyzed the animals' blood and developed an miRNA-based classifier. By measuring the expression of fivestudied nonhuman primates who had been exposed to radiation and given a radioprotective drug or placebo, and
.et alall and to determine which victims are at risk of death or severe injury and require urgent treatment. Fendler atresponse is vital for saving lives. In particular, it is important to identify those who had been exposed to radiation
When a nuclear accident or attack occurs and people are exposed to radiation, a rapid and accurateRadiation alert