Title Reduced RET expression in gut tissue of individuals carrying risk alleles of Hirschsprung's disease Author(s) Miao, X; Leon, TYY; Ngan, ESW; So, MT; Yuan, ZW; Lui, VCH; Chen, Y; Wong, KKY; Tam, PKH; GarciaBarceló, M Citation Human Molecular Genetics, 2010, v. 19 n. 8, p. 1461-1467 Issued Date 2010 URL http://hdl.handle.net/10722/83609 Rights This is a pre-copy-editing, author-produced PDF of an article accepted for publication in Human Molecular Genetics following peer review. The definitive publisher-authenticated version Human Molecular Genetics, 2010, v. 19 n. 8, p. 1461-1467 is available online at: http://hmg.oxfordjournals.org/content/19/8/1461
Transcript
Title Reduced RET expression in gut tissue of individuals carryingrisk alleles of Hirschsprung's disease
Author(s) Miao, X; Leon, TYY; Ngan, ESW; So, MT; Yuan, ZW; Lui, VCH;Chen, Y; Wong, KKY; Tam, PKH; GarciaBarceló, M
Citation Human Molecular Genetics, 2010, v. 19 n. 8, p. 1461-1467
Issued Date 2010
URL http://hdl.handle.net/10722/83609
Rights
This is a pre-copy-editing, author-produced PDF of an articleaccepted for publication in Human Molecular Genetics followingpeer review. The definitive publisher-authenticated versionHuman Molecular Genetics, 2010, v. 19 n. 8, p. 1461-1467 isavailable online at:http://hmg.oxfordjournals.org/content/19/8/1461
For Peer Review
Reduced RET expression in gut tissue of individuals carrying
risk alleles of Hirschsprung’s Disease
Journal: Human Molecular Genetics
Manuscript ID: HMG-2009-W-01223.R1
Manuscript Type: 1 General Article - US Office
Date Submitted by the Author:
Complete List of Authors: Miao, Xiaoping; School of Public Health, Tongji Medical College, Huazhong University of Science and Technology, Department of Epidemiology and Biostatistics Thomas, Leon; HKU, surgery ELLY, NGAN; HKU, SURGERY Man-ting, so; HKU, surgery yuan, zhengwei; chinese medical college, surgery Vincent, Lui; HKU, surgery Kenneth, Wong; HKU, surgery chen, yan; HKU, surgery Tam, Paul; University of Hong Kong Medical Center,Queen Mary
Hospital, Division of Paediatric Surgery, Department of Surgery Garcia-Barcelo, Maria-Merce; The University of Hong Kong, Surgery and Genome Research Centre
1 Angrist, M., Kauffman, E., Slaugenhaupt, S.A., Matise, T.C., Puffenberger, E.G., Washington, S.S., Lipson, A., Cass, D.T., Reyna, T, Weeks, D.E., et al. (1993) A gene for Hirschsprung disease (megacolon) in the pericentromeric region of human chromosome 10. Nat. Genet., 4, 351-356. 2 Romeo, G., Ronchetto, P., Luo, Y., Barone, V., Seri, M., Ceccherini, I., Pasini, B., Bocciardi, R., Lerone, M., Kaariainen, H., et al. (1994) Point mutations affecting the tyrosine kinase domain of the RET proto-oncogene in Hirschsprung's disease. Nature, 367, 377-378. 3 Plaza-Menacho, I., Burzynski, G.M., de Groot, J.W., Eggen, B.J. and Hofstra, R.M. (2006) Current concepts in RET-related genetics, signaling and therapeutics. Trends Genet., 22, 627-636. 4 Edery, P., Lyonnet, S., Mulligan, L.M., Pelet, A., Dow, E., Abel, L., Holder, S., Nihoul-Fekete, C., Ponder, B.A. and Munnich, A. (1994) Mutations of the RET proto-oncogene in Hirschsprung's disease. Nature, 367, 378-380. 5 Attie, T., Pelet, A., Edery, P., Eng, C., Mulligan, L.M., Amiel, J., Boutrand, L., Beldjord, C., Nihoul-Fekete, C., Munnich, A., et al. (1995) Diversity of RET proto-oncogene mutations in familial and sporadic Hirschsprung disease. Hum. Mol.Genet., 4, 1381-1386. 6 Seri, M., Yin, L., Barone, V., Bolino, A., Celli, I., Bocciardi, R., Pasini, B., Ceccherini, I., Lerone, M., Kristoffersson, U., et al. (1997) Frequency of RET mutations in long- and short-segment Hirschsprung disease. Hum. Mutat., 9, 243-249. 7 Svensson, P.J., Molander, M.L., Eng C., Anvret, M. and Nordenskjold, A. (1998) Low frequency of RET mutations in Hirschsprung disease in Sweden. Clin. Genet., 54, 39-44. 8 Garcia-Barcelo, M., Sham, M.H., Lee, W.S., Lui, V.C., Chen, B.L., Wong, K.K., Wong, J.S. and Tam, P.K. (2004) Highly recurrent RET mutations and novel mutations in genes of the receptor tyrosine kinase and endothelin receptor B pathways in Chinese patients with sporadic Hirschsprung disease. Clin. Chem., 50, 93-100. 9 Tam, P.K. and Garcia-Barcelo, M. (2009) Genetic basis of Hirschsprung's disease. Pediatr. Surg. Int., 25, 543-558. 10 Brooks, A.S., Bertoli-Avella, A.M., Burzynski, G.M., Breedveld, G.J., Osinga, J., Boven, L.G., Hurst, J.A., Mancini, G.M., Lequin, M.H., de Coo, R.F. et al. (2005) Homozygous nonsense mutations in KIAA1279 are associated with malformations of the central and enteric nervous systems. Am. J. Hum. Genet., 77, 120-126. 11 Amiel, J., Sproat-Emison, E., Garcia-Barcelo, M., Lantieri, F., Burzynski, G., Borrego, S., Pelet, A., Arnold, S., Miao, X., Griseri, P., et al. (2008) Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet., 45,1-14. 12 Garcia-Barcelo, M.M., Tang, C.S., Ngan, E.S., Lui, V.C., Chen, Y., So,M.T., Leon, T.Y., Miao, X.P., Shum, C.K., Liu, F.Q. et al. (2009) Genome-wide association study identifies NRG1 as a susceptibility locus for Hirschsprung's disease. Proc. Natl. Acad. Sci. USA., 106, 2694-2699. 13 Borrego, S., Ruiz, A., Saez, M.E., Gimm, O., Gao, X., Lopez-Alonso, M., Hernandez, A., Wright, F.A., Antinolo, G. and Eng, C. (2000) RET genotypes comprising specific haplotypes of polymorphic variants predispose to isolated Hirschsprung disease. J. Med.
Genet., 37, 572-578. 14 Griseri, P., Pesce, B., Patrone, G., Osinga, J., Puppo, F., Sancandi, M., Hofstra, R., Romeo, G., Ravazzolo, R., Devoto, M., et al. (2002) A rare haplotype of the RET proto-oncogene is a risk-modifying allele in hirschsprung disease. Am. J. Hum. Genet., 71, 969-974. 15 Sancandi, M., Griseri, P., Pesce, B., Patrone, G., Puppo, F., Lerone, M., Martucciello, G., Romeo, G., Ravazzolo, R., Devoto, M., et al. (2003) Single nucleotide polymorphic alleles in
the 5' region of the RET proto-oncogene define a risk haplotype in Hirschsprung's disease. J.
Med. Genet., 40, 714-718. 16 Garcia-Barcelo, M.M., Sham, M.H., Lui, V.C., Chen, B.L., Song,Y.Q., Lee, W.S., Yung, S.K., Romeo, G. and Tam, P.K. (2003) Chinese patients with sporadic Hirschsprung's disease are predominantly represented by a single RET haplotype. J. Med. Genet., 40, e122. 17 Fitze, G., Cramer, J., Ziegler, A., Schierz, M., Schreiber, M., Kuhlisch, E., Roesner, D. and Schackert, H.K. (2002) Association between c135G/A genotype and RET proto-oncogene germline mutations and phenotype of Hirschsprung's disease. Lancet, 359, 1200-1205. 18 Griseri, P., Bachetti, T., Puppo, F., Lantieri, F., Ravazzolo, R., Devoto, M. and Ceccherini, I. (2005) A common haplotype at the 5' end of the RET proto-oncogene, overrepresented in Hirschsprung patients, is associated with reduced gene expression. Hum.
Mutat., 25, 189-195. 19 Garcia-Barcelo, M., Ganster, R.W., Lui, V.C., Leon, T.Y., So, M.T., Lau, A.M., Fu, M., Sham, M.H., Knight, J., Zannini, M.S., et al. (2005) TTF-1 and RET promoter SNPs: regulation of RET transcription in Hirschsprung's disease. Hum. Mol. Genet., 14, 191-204. 20 Fitze, G., Appelt, H., Konig, I.R., Gorgens, H., Stein, U., Walther, W., Gossen, M., Schreiber, M., Ziegler, A., Roesner, D., et al. (2003) Functional haplotypes of the RET proto-oncogene promoter are associated with Hirschsprung disease (HSCR). Hum. Mol.
Genet., 12, 3207-3214. 21 Burzynski, G.M., Nolte, IM., Bronda, A., Bos, K.K., Osinga, J., Plaza Menacho, I., Twigt, B., Maas, S., Brooks, A.S., Verheij, J.B., et al. (2005) Identifying candidate Hirschsprung disease-associated RET variants. Am. J. Hum. Genet., 76, 850-858. 22 Burzynski, G.M., Nolte, I.M., Osinga, J., Ceccherini, I., Twigt, B., Maas, S., Brooks, A., Verheij, J., Plaza Menacho, I., Buys, C.H., et al. (2004) Localizing a putative mutation as the major contributor to the development of sporadic Hirschsprung disease to the RET genomic sequence between the promoter region and exon 2. Eur. J. Hum. Genet., 12, 604-612. 23 Pelet, A., de Pontual, L., Clément-Ziza, M., Salomon, R., Mugnier, C., Matsuda, F., Lathrop, M., Munnich, A., Feingold, J., Lyonnet, S., et al. (2005) Homozygosity for a frequent and weakly penetrant predisposing allele at the RET locus in sporadic Hirschsprung disease. J. Med. Genet., 42, e18. 24 Fernandez, R.M., Boru, G, Peciña, A., Jones, K., López-Alonso, M., Antiñolo, G., Borrego, S. and Eng, C. (2005) Ancestral RET haplotype associated with Hirschsprung's disease shows linkage disequilibrium breakpoint at -1249. J. Med. Genet., 42, 322-327. 25 Emison, E.S., McCallion, A.S., Kashuk, C.S., Bush, R.T., Grice, E., Lin, S., Portnoy, M.E., Cutler, D.J., Green, E.D. and Chakravarti, A. (2005) A common sex-dependent mutation in a RET enhancer underlies Hirschsprung disease risk. Nature, 434, 857-863. 26 Heanue, T.A. and Pachnis, V. (2007) Enteric nervous system development and Hirschsprung's disease: advances in genetic and stem cell studies. Nat. Rev. Neurosci., 8, 466-479. 27 Uesaka, T., Nagashimada, M., Yonemura, S. and Enomoto, H. (2008) Diminished Ret expression compromises neuronal survival in the colon and causes intestinal aganglionosis in mice. J. Clin. Invest., 118, 1890-1898. 28 Miao, X., Garcia-Barcelo, M.M., So, M.T., Leon, T.Y., Lau, D.K., Liu, T.T., Chan, E.K., Lan, L.C., Wong, K.K., Lui, V.C., et al. (2007) Role of RET and PHOX2B gene polymorphisms in risk of Hirschsprung's disease in Chinese population. Gut, 56, 736. 29 Veiga-Fernandes, H., Coles, M.C., Foster, K.E., Patel, A., Williams, A., Natarajan, D., Barlow, A., Pachnis, V. and Kioussis, D. (2007) Tyrosine kinase receptor RET is a key regulator of Peyer's patch organogenesis. Nature, 446, 547-551. 30 Stephens, M. and Donnelly, P. (2003) A comparison of bayesian methods for haplotype
reconstruction from population genotype data. Am. J. Hum. Genet., 73, 1162-1169. 31 Stephens, M., Smith, N.J. and Donnelly, P. (2001) A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Genet., 68, 978-989. 32 Barrett, J.C., Fry, B., Maller, J. and Daly, M.J. (2005) Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics, 21, 263-265.
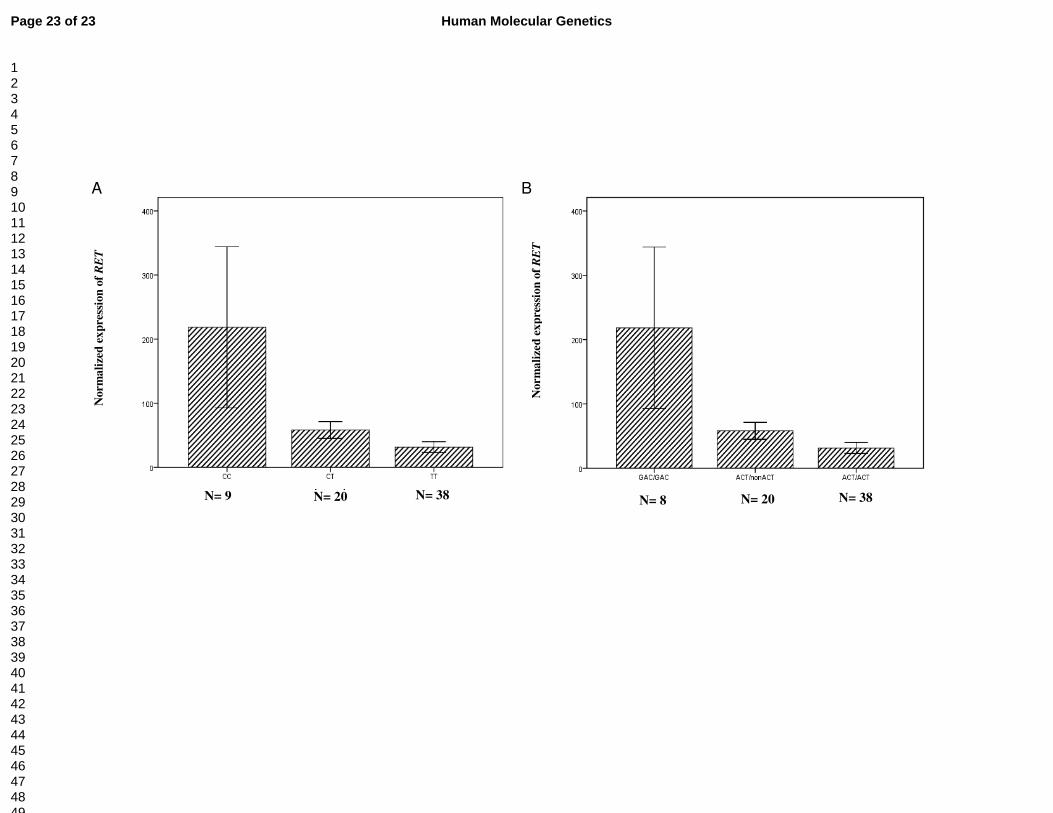
Figure 1. Associations between genetic variants in the regulatory region and normalized RET expression in the colon tissues from the normal portions of HSCR patients and non-HSCR patients. (Y axis represented the RET expression relative to PGP9.5) (A) Significant difference of normalized RET expression were observed among CC, CT and TT genotypes of intron 1 polymorphism (P =0.012, one-way ANOVA). (B) Significant difference of normalized RET expression were observed among different diplotypes ((-5G>A; -1A>C; intron 1C>T) of RET variants (P =0.018, one-way ANOVA). Graphs represent mean ± SE.