ARTICLE Mounir Traı¨kia Æ Dror E. Warschawski Michel Recouvreur Æ Jean Cartaud Æ Philippe F. Devaux Formation of unilamellar vesicles by repetitive freeze-thaw cycles: characterization by electron microscopy and 31 P-nuclear magnetic resonance Received: 2 September 1999 / Revised version: 27 February 2000 / Accepted: 27 February 2000 Abstract It has been reported that repetitive freeze-thaw cycles of aqueous suspensions of dioleoylphosphati- dylcholine form vesicles with a diameter smaller than 200 nm. We have applied the same treatment to a series of phospholipid suspensions with particular emphasis on dioleoylphosphatidylcholine/dioleoylphosphatidic acid (DOPC/DOPA) mixtures. Freeze-fracture electron mi- croscopy revealed that these unsaturated lipids form unilamellar vesicles after 10 cycles of freeze-thawing. Both electron microscopy and broad-band 31 P NMR spectra indicated a disparity of the vesicle sizes with a highest frequency for small unilamellar vesicles (diame- ters £30 nm) and a population of larger vesicles with a frequency decreasing exponentially as the diameter in- creases. From 31 P NMR investigations we inferred that the average diameter of DOPC/DOPA vesicles calcu- lated on the basis of an exponential size distribution was of the order of 100 nm after 10 freeze-thaw cycles and only 60 nm after 50 cycles. Fragmentation by repeated freeze-thawing does not have the same eciency for all lipid mixtures. As found already by others, fragmenta- tion into small vesicles requires the presence of salt and does not take place in pure water. Repetitive freeze- thawing is also ecient to fragment large unilamellar vesicles obtained by filtration. If applied to sonicated DOPC vesicles, freeze-thawing treatment causes fusion of sonicated unilamellar vesicles into larger vesicles only in pure water. These experiments show the usefulness of NMR as a complementary technique to electron microscopy for size determination of lipid vesicles. The applicability of the freeze-thaw technique to dierent lipid mixtures confirms that this procedure is a simple way to obtain unilamellar vesicles. Key words Liposomes Æ Phosphorus-31 NMR Æ Large unilamellar vesicles Æ Freeze-fracture electron microscopy Æ Dioleoylphosphatidylcholine Abbreviations Chol cholesterol Æ DLnPC dilinoleoyl- phosphatidylcholine Æ DMPA dimyristoylphosphatidic acid Æ DMPC dimyristoylphosphatidylcholine Æ DOPA dioleoylphosphatidic acid Æ DOPC dioleoylphospha- tidylcholine Æ DOPG dioleoylphosphatidylglycerol Æ DPPC dipalmitoylphosphatidylcholine Æ EPC egg phosphatidylcholine Æ ESM egg sphingomyelin Æ LPC lysophosphatidylcholine Æ LUV large unilamellar vesicles Æ MLV multilamellar vesicles Æ POPC 1-palmi- toyl-2-oleolylphosphatidylcholine Æ SUV sonicated unil- amellar vesicles Introduction It is well established that mechanical dispersion of dry lipids in aqueous buer forms multilamellar vesicles (MLVs) with typical sizes of the order of a micron. Repetitive freeze-thawing cycles are often applied to MLVs in order to homogenize their lipid composition. This procedure also enhances trapping eciencies due to the breaking of MLVs and the formation of a more homogeneous vesicles population that appears to be largely unilamellar (Mayer et al. 1985). Similarly, freeze- thawing was demonstrated to cause fusion of sonicated unilamellar vesicles (SUVs) (Oku and MacDonald 1983). Fragmentation of dioleoylphosphatidylcholine (DOPC) MLVs into small unilamellar vesicles following freeze-thaw cycles was analyzed in detail by MacDonald et al. (1994). The decrease of the average DOPC vesicle size as a function of number of freeze-thaw cycles was Eur Biophys J (2000) 29: 184–195 Ó Springer-Verlag 2000 M. Traı¨kia Æ D.E. Warschawski Æ P.F. Devaux (&) Physico-Chimie Mole´culaire des Membranes Biologiques, UPR-CNRS 9052, Institut de Biologie Physico-Chimique, 13 Rue Pierre et Marie Curie, 75005 Paris, France e-mail: [email protected]M. Recouvreur Æ J. Cartaud Biologie Cellulaire des Membranes, Institut Jacques Monod, UMR 7592, CNRS, Universite´ Paris 6/Paris 7, 75251 Paris Cedex 05, France
Transcript
ARTICLE
Mounir TraõÈ kia á Dror E. WarschawskiMichel Recouvreur á Jean Cartaud á Philippe F. Devaux
Formation of unilamellar vesicles by repetitive freeze-thaw cycles:characterization by electron microscopyand 31P-nuclear magnetic resonance
Received: 2 September 1999 /Revised version: 27 February 2000 /Accepted: 27 February 2000
Abstract It has been reported that repetitive freeze-thawcycles of aqueous suspensions of dioleoylphosphati-dylcholine form vesicles with a diameter smaller than200 nm. We have applied the same treatment to a seriesof phospholipid suspensions with particular emphasis ondioleoylphosphatidylcholine/dioleoylphosphatidic acid(DOPC/DOPA) mixtures. Freeze-fracture electron mi-croscopy revealed that these unsaturated lipids formunilamellar vesicles after 10 cycles of freeze-thawing.Both electron microscopy and broad-band 31P NMRspectra indicated a disparity of the vesicle sizes with ahighest frequency for small unilamellar vesicles (diame-ters £30 nm) and a population of larger vesicles with afrequency decreasing exponentially as the diameter in-creases. From 31P NMR investigations we inferred thatthe average diameter of DOPC/DOPA vesicles calcu-lated on the basis of an exponential size distribution wasof the order of 100 nm after 10 freeze-thaw cycles andonly 60 nm after 50 cycles. Fragmentation by repeatedfreeze-thawing does not have the same e�ciency for alllipid mixtures. As found already by others, fragmenta-tion into small vesicles requires the presence of salt anddoes not take place in pure water. Repetitive freeze-thawing is also e�cient to fragment large unilamellarvesicles obtained by ®ltration. If applied to sonicatedDOPC vesicles, freeze-thawing treatment causes fusionof sonicated unilamellar vesicles into larger vesicles onlyin pure water. These experiments show the usefulness ofNMR as a complementary technique to electron
microscopy for size determination of lipid vesicles. Theapplicability of the freeze-thaw technique to di�erentlipid mixtures con®rms that this procedure is a simpleway to obtain unilamellar vesicles.
Key words Liposomes á Phosphorus-31 NMR á Largeunilamellar vesicles á Freeze-fracture electronmicroscopy á Dioleoylphosphatidylcholine
Abbreviations Chol cholesterol á DLnPC dilinoleoyl-phosphatidylcholine á DMPA dimyristoylphosphatidicacid á DMPC dimyristoylphosphatidylcholine á DOPAdioleoylphosphatidic acid á DOPC dioleoylphospha-tidylcholine á DOPG dioleoylphosphatidylglycerol áDPPC dipalmitoylphosphatidylcholine á EPC eggphosphatidylcholine á ESM egg sphingomyelin á LPClysophosphatidylcholine á LUV large unilamellarvesicles á MLV multilamellar vesicles á POPC 1-palmi-toyl-2-oleolylphosphatidylcholine á SUV sonicated unil-amellar vesicles
Introduction
It is well established that mechanical dispersion of drylipids in aqueous bu�er forms multilamellar vesicles(MLVs) with typical sizes of the order of a micron.Repetitive freeze-thawing cycles are often applied toMLVs in order to homogenize their lipid composition.This procedure also enhances trapping e�ciencies due tothe breaking of MLVs and the formation of a morehomogeneous vesicles population that appears to belargely unilamellar (Mayer et al. 1985). Similarly, freeze-thawing was demonstrated to cause fusion of sonicatedunilamellar vesicles (SUVs) (Oku and MacDonald1983). Fragmentation of dioleoylphosphatidylcholine(DOPC) MLVs into small unilamellar vesicles followingfreeze-thaw cycles was analyzed in detail by MacDonaldet al. (1994). The decrease of the average DOPC vesiclesize as a function of number of freeze-thaw cycles was
Eur Biophys J (2000) 29: 184±195 Ó Springer-Verlag 2000
M. TraõÈ kia á D.E. Warschawski á P.F. Devaux (&)Physico-Chimie Mole culaire des Membranes Biologiques,UPR-CNRS 9052, Institut de Biologie Physico-Chimique,13 Rue Pierre et Marie Curie, 75005 Paris, Francee-mail: [email protected]
M. Recouvreur á J. CartaudBiologie Cellulaire des Membranes,Institut Jacques Monod,UMR 7592, CNRS, Universite Paris 6/Paris 7,75251 Paris Cedex 05, France
inferred from light absorbance changes and trappingcapacity. A more direct determination of the averagevesicle size was obtained by electron microscopy andphoton correlation spectroscopy and indicated an aver-age diameter of 145 nm after 10 freeze-thawing cycles(MacDonald et al. 1994). However, the latter reportdid not show the size distribution of the freeze-thawedvesicles. The technique of unilamellar vesicle prepara-tion by freeze-thawing is rather simple and can be usedin principle for biotechnical applications (drug encap-sulation) or for biophysical investigations on modelmembranes. It is sometimes complemented by ®ltra-tion, supposedly to obtain homogeneous vesicle size.Indeed, freeze-thawing does not give a population ofvesicles with the same size. Furthermore, fragmenta-tion as well as fusion by repeated freeze-thawing donot have the same e�ciency for all lipid mixtures.A precise knowledge of the size distribution would beuseful in particular for comparison with othertechniques of vesicle preparation such as largeunilamellar vesicles (LUVs) produced with an extruder(Hope et al. 1985) or by a reverse phase procedure(Szoka et al. 1980).
Electron microscopy and light scattering are oftenused to determine vesicle sizes. In this study we dem-onstrate that 31P NMR can be a very powerful techniquefor this purpose. The fragmentation of lipid vesiclesmodi®es NMR lineshapes because of the change inaveraging of the anisotropic chemical shift and relax-ation parameters of the phosphorus nuclei (Dufourcet al. 1992). Thus, one can take advantage of the changein lineshape of 31P NMR spectra to monitor the changein size distribution of di�erent phospholipid mixturesupon repeated freeze-thawing. We show that analysis ofNMR spectra provides an estimate of the size distribu-tion pro®le probably more accurate than statisticalanalysis of freeze-fracture or negative staining electronmicrographs.
Materials and methods
Chemicals
Egg phosphatidylcholine (EPC), egg sphingomyelin(ESM), DOPC, 1-palmitoyl-2-oleolylphosphatidylcho-line (POPC), dimyristoylphosphatidic acid (DMPA),dimyristoylphosphatidylcholine (DMPC), dipalmitoyl-phosphatidylcholine (DPPC), dilinoleoylphosphatidylch-oline (DLnPC), dioleoylphosphatidylethanolamine(DOPE), dioleoylphosphatidylglycerol (DOPG), lys-ophosphatidylcholine (LPC) and cholesterol (Chol) werepurchased from Sigma and were used without furtherpuri®cation. Dioleoylphosphatidic acid (DOPA) wasobtained from DOPC by hydrolysis of the choline headgroup using phospholipase D (Roux et al. 1983). Thepurity of all lipids was veri®ed by thin layerchromatography and by high-resolution NMR in
chloroform. Deuterium oxide (D2O) was obtained fromEurisotop (Saint-Aubin, France), and N-(2-hydroxy-ethyl)piperazine-N¢ (ethanesulfonic acid) (Hepes), ethy-lenediamine- teratraacetic acid (EDTA) and KCl werepurchased from Sigma.
Preparation of freeze-thawed liposomes
MLVs were obtained by mixing phospholipids at thedesired ratio in chloroform. Chloroform was removed®rst at room temperature with a rotary evaporator.Residual organic solvent was removed by pumping forat least 2 h with a mechanical vacuum pump. Either a1:1 (v/v) mixture of H2O and D2O or a 1:1 mixture ofD2O and Hepes bu�er (0.1 M Hepes, 0.1 M KCl,5 mM EDTA, pH 8.0) was added to the lipid ®lm(®nal concentration for lipid mixtures was 100 mg/mL)and the sample vortexed to emulsify the lipid mixture.The lipids in a 5 mL glass tube were then dipped intoliquid nitrogen for rapid cooling. After 3 min, thefrozen lipids were transferred to a bath at 60 °C forthawing. After 3 min in the hot bath, the lipids werefrozen again. This operation was repeated several timesas indicated.
SUVs were prepared by sonication of MLV disper-sion under a stream of argon using a probe-type soni-cator (model VC50, Bioblock Scienti®c, Paris) at 40 Win an ice bath until a clear solution was obtained (whichrequired approximately 30 min). The sample was after-wards centrifuged at 11,000g for 15 min to ensure theremoval of metallic particles.
LUVs were prepared by the reverse phase evapo-ration technique (Szoka et al. 1980). The lipid mixture(25 mg total lipid) in chloroform/methanol solutionwas deposited on the sides of a round-bottom ¯ask byremoval of the organic solvent by rotary evaporation.The lipids were then redissolved in 2 mL diethyl ether.Hepes bu�er (0.5 mL) was added to the organic so-lution of phospholipids and the mixture was sonicatedin a probe-type sonicator for 2 min. The mixture wasplaced on a rotary evaporator and the remainingorganic solvent was removed under vacuum in twostages: evaporation at 400 mmHg until the suspensionbecame a gel, followed by a brief vortex mixing andthen continued evaporation at 730 mmHg until ahomogeneous suspension was obtained. The prepara-tion (50 mg/mL for all lipid mixtures) was extrudedthrough 0.4 lm then 0.2 lm polycarbonate mem-branes.
Electron microscopy
The same vesicles were used for NMR spectroscopy andfor electron microscopy except that vesicles (100 mg/mLfor all lipid mixtures) were mixed with glycerol (50%v/v) as cryoprotectant and frozen rapidly in a propane
185
slush for freeze-fracture investigations. Quenched sam-ples were fractured in a Balzers BAF 300 freeze-fractureapparatus (Balzers, Liechtenstein) equipped with anelectron beam device for platinum and carbon shadow-ing and a quartz crystal monitor for regulating shadowthickness (2 nm for platinum and 20 nm for carbon).The replicas were cleaned in sodium chlorate solutionsand collected on 300-mesh copper grids coated withcollodion ®lm.
The replicas were examined in a Philips CM 12electron microscope operating at 80 keV. The pictureswere taken on Kodak electron microscope ®lms 4489 atnominal initial magni®cations of 15,000´ and 26,000´.The size distributions were measured on 2.5´ magni®edprints. Dimensions of the freeze-fractured vesicles weremeasured as the largest length in the direction perpen-dicular to the shadowing direction. In the case of unil-amellar vesicles, most pro®les of the fractured lipid shellswere circular and it could therefore be assumed that theywere derived from spherical vesicles. As discussed inHeegaard et al. (1990), in the case of an homogeneouspopulation of spherical vesicles with the same diameterD, the e�ect of non-equatorial fractures introduces asmall correcting factor between the weighted mean di-ameter Dm obtained by assuming a random intersectionof identical spheres and the actual diameter D of thesespheres. Namely:
Dm � �p=4�D �1�About 200±1500 vesicles were counted for each sample.Accuracy of size determination was about 10 nm, cor-responding to 0.5 mm on the ruler.
For negative staining, vesicle samples (1±10 mg/mL)were diluted in 0.1 M ammonium acetate bu�er(pH 7.4) containing 10 lg/mL bacitracin. Drops ofvesicles were deposited onto carbon-coated grids, andnegatively stained with 1% aqueous uranyl acetate.Grids were directly observed in the microscope (80 keV).Pictures were taken at a magni®cation of 15,000´ underminimal irradiation conditions.
NMR experiments
NMR measurements were performed on a Bruker 400AMX, wide-bore spectrometer operating at 162 MHzfor 31P using a 10 mm liquid probe. Spectra were ac-quired using a phase cycled Hahn echo with protonbroad band decoupling (WALTZ16) (Rance and Byrd1983). Typical acquisition parameters were: 90° pulselength 10 ls; echo delay 40 ls; recycle delay 3 s or 5 s,after checking that the lineshape is not distorted; spec-tral width 100 ppm (16 kHz). In all experiments, 16 kcomplex points were acquired, zero ®lled to 32 k beforeFourier transformation, exponentially multiplied with50 Hz line broadening, and treated with automaticbaseline correction. Each spectrum was the average of 5±18 k scans.
NMR spectral simulation
The e�ect of vesicle size distribution, polydispersity andmultilamellarity can be accounted for in the simulationsof 31P NMR spectra by considering both the rotationaldi�usion of the vesicles and the lateral di�usion ofphospholipids along the curved surfaces (Burnell et al.1980). The tensor representing the chemical shift an-isotropy of the phosphate moiety is considered to beaxially symmetric due to the rapid motion of thephospholipids around the local normal of the bilayer.Additional averaging is due to phospholipid reorienta-tion caused by vesicle tumbling st or lateral di�usionwithin each vesicle; the latter reorientation is associatedwith a characteristic time, sdi�. One can introduce ane�ective characteristic time s, de®ned by the followingexpression:
1
s� 1
st� 1
sdiff� 6
r2Dt � Ddiff� � �2�
where Dt � kT=8prg is the rotational di�usion coe�cientof the vesicles, g being the viscosity of the medium. Thesimulation program employed in the present study wasinspired from the doctorate thesis of Rance (1981) as de-veloped in Douliez et al. (1994). We have modi®ed theprogram in order to include the isotropic chemical shift ofeach lipid in lipid mixtures and to allow us to test partic-ular size distribution pro®les. The following values wereused in the simulations: T � 298 K; Ddi� � 10)7 cm2/s(Galla et al. 1979) and g � 7.808 ´ 10)4 Poise.
Frequency shifts were referred to the isotropicchemical shift of DOPC. The overall chemical shift an-isotropy (Dr) in the absence of averaging by vesicletumbling and/or lateral di�usion was obtained from thedi�erence between parallel and perpendicular chemicalshifts as measured on the NMR spectrum recorded withMLVs in the ¯uid state before freeze-thawing. The iso-tropic chemical shifts of LPC, DOPE and DOPG weredetermined in SUVs and corresponded to low-®eld shiftsof diso � 0.5 ppm, 0.6 ppm and 1.1 ppm, respectively.For DOPA, diso was in the range 2±3 ppm, towards low®elds. This value is strongly dependent upon pH, tem-perature and ionic strength (Swairjo et al. 1994; TraõÈ kiaet al. 1997). For lipid mixtures, Dr can be evaluated foreach lipid by measuring the distance between the high®eld peak (h � 90°) which is always clearly visible, andthe theoretical position of the isotropic peak, diso of thecorresponding lipid. We found Dr � 45 � 1 ppm forDOPC, DOPG, EPC, POPC, DMPC, DPPC andDLnPC. The same value was obtained for DOPA underour conditions of pH and temperature. When LPC wasincorporated in DOPC vesicles, two powder patternswere observed and revealed that LPC had a signi®cantlysmaller chemical shift anisotropy, with Dr � 35 �1 ppm.
The intrinsic linewidth, which is proportional to 1/T2,is theoretically orientation dependent for 31P NMR.Smith and Ekiel (1984) have reported the followingvalues for phospholipids in MLVs: T2 � 2.4 ms for
186
h � 0 and T2 � 7.4 ms for h � p/2, where h is the anglebetween the normal to the bilayer and the direction ofthe magnetic ®eld. In our simulations the lorentzianlinewidth, Dm, was calculated for each orientation byusing Seelig's formula (Seelig 1978):
Dt � 1
pT2� R1 � R2�3 cos2 hÿ 1�
where R1 and R2 are adjustable parameters; R1 can bedetermined in systems where the anisotropy is averagedout. DOPC has a linewidth in Triton X100 around30 Hz and in SUVs in the range 40±50 Hz. In SUVsmade with DOPC/DOPA, the DOPA linewidth is al-ways found larger than that of DOPC. It is also afunction of the pH and of the packing imposed by thecurvature, which is di�erent for the inner and outerlea¯ets (Swairjo et al. 1994). In practice, R1 values wereadjusted in the range 20±100 Hz for DOPC, DOPG andLPC while R1 was selected in the range 100±500 Hz forDOPA. An optimum value for R2 was 600 Hz. Thisvalue was ®xed in all the simulations.
It can be assumed that all lamellae of a given MLVhave the same tumbling rate, which depends only on theradius of curvature of the most external bilayer. On theother hand, the reorientation of the phospholipids bylateral di�usion depends upon the actual radius of eachindividual vesicle within one MLV. Thus 31P NMRspectra of MLVs can be simulated: (1) by assuming a®xed external radius and a variable number of innerbilayers, or (2) by assuming that the number of innerbilayers is determined by a periodic spacing betweenlamellae, the smaller vesicle possessing an arbitrary di-ameter of 20 or 30 nm, while the external diameter isvariable (Douliez et al. 1994). Obviously, if the numberof lamellae and the external radius are allowed to varyfor the same sample, there are too many parameters toreach unambiguous conclusions. For that reason thelamellarity was determined independently by electronmicroscopy freeze-fracture studies.
For spectra corresponding to a population of unila-mellar vesicles, Douliez et al. (1994) assumed a Gaussiandistribution. However, in our particular case, optimum®ts of the spectra were obtained with a distribution ofvesicle diameters, which decreased exponentially from amaximum for small diameters to zero frequency forlarge diameters.
Results
Freeze-fracture electron microscopy
We have carried out freeze-fracture electron microscopyon various lipid suspensions: DOPC, DOPC/DOPA,DOPC/LPC, DOPC/DOPA/LPC, DOPC/Chol, EPC,DLnPC, DMPC, DPPC, ESM and DMPC/DMPA.Figure 1a±c shows freeze-fracture electron micrographs
of DOPC/DOPA (80:20 mol%) vesicles before and afterfreeze-thawing, with 0, 10 and 50 cycles, respectively. Themultilamellar vesicles become in the majority unilamellarfollowing freeze-thawing, and have a quasi-sphericalshape. Quantitative evaluation of the percentage of un-ilamellar vesicles was achieved by analyzing micrographsobtained with vesicles of di�erent lipid composition ob-tained after 10 freeze-thaw cycles. The percentage ofvesicles remaining multilamellar after fragmentation waspractically zero for DOPC/DOPA or DOPC/LPC, about2% of the vesicles appeared multilamellar in the case ofDOPC alone or DOPC/Chol (70:30 mol%) and about5% in the case of EPC. If less than 10 cycles were used, alarger number of multilamellar vesicles could be counted,in particular with EPC, which is in agreement with theresults of Mayer et al. (1985).
Dimensions of the unilamellar vesicles were deter-mined from the fractured vesicles after 10 freeze-thawcycles (see Materials and methods). Typical histogramsof the percentage of vesicles of various diameters areshown in Fig. 2 for cases where e�cient fragmentationtook place. More than 40% of the counted vesicles had adiameter below 100 nm and the frequency decreased atgreater diameters. The average diameter was reduced bythe presence of DOPA, DOPG or LPC, suggesting a roleof the charged lipids. Increasing the number of freeze-thaw cycles decreased the average vesicle size: after50 cycles, more than 90% of counted vesicles had adiameter below 100 nm.
Note that if instead of histograms of the percentageof vesicles, histograms of the inner volume are repre-sented for various values of D assuming spherical vesi-cles (Fig. 2), the same data (because of the factor D3)reveal that the contribution of large or very large un-ilamellar vesicles (i.e. D ³ 400 nm) is in fact prepon-derant. Thus, in spite of the majority of very smallvesicles, the entrapped volume per lmol of phospholipidis important with such liposome preparations. However,the ¯uctuations in the histograms representing the innervolumes (Fig. 2) emphasize the fact that the actualpopulation of very large vesicles is di�cult to establish ina statistically reliable fashion by electron microscopy,even when a population of several hundreds of unila-mellar vesicles is examined. This is because these verylarge vesicles represent only a small percentage of thetotal population and, hence, counting only a fewhundreds of vesicles is insu�cient for good statistics.
Negative staining electron microscopy
This technique is often considered as more reliable forsize determination. However, owing to the frequentoverlap of vesicles on the grids and since the very smallvesicles are di�cult to visualize, the uncertainty may infact be larger. Figure 1d, for example, shows vesicleswith complicated shapes, which could be a superpositionof several vesicles. Histograms have been made from
187
negative staining data. The overall tendency was similarto that deduced from freeze-fracture experiments with,however, less very small vesicles (data not shown).
31P Broad band NMR and simulated spectra
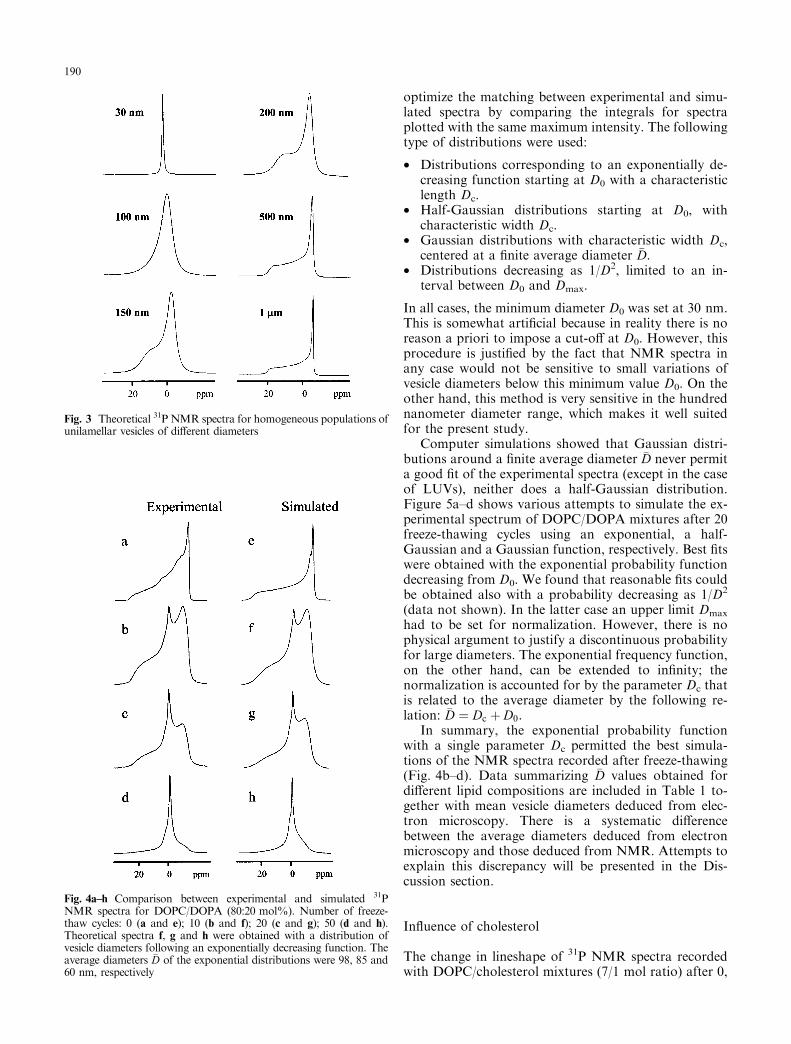
Because of its sensitivity to the time scale of reorienta-tion, 31P broad band NMR allows one to determine thedistribution of vesicle sizes for vesicles with a diameterbelow 1 lm (Burnell et al. 1980). Figure 3 shows a set oftheoretical NMR spectra corresponding to homoge-neous populations of spherical phospholipid vesicleswith diameters varying from 30 nm to 1 lm. Figure4a±d shows experimental spectra obtained with a mix-ture of DOPC/DOPA (80:20 mol%) subjected to 0, 10,20, and 50 freeze-thaw cycles, respectively. Figure 4e±hshows the corresponding simulated spectra.
The theoretical spectrum of Fig. 4e was obtained byassuming MLVs with an onion structure, i.e. assumingthe superposition of vesicles stacked like Russian dollswith a continuous distribution from very large to verysmall vesicles (Douliez et al. 1994). However, the ex-perimental spectrum corresponding to DOPC/DOPAmixtures (Fig. 4a) shows features which probably re¯ectthe existence of a large proportion of vesicles withsub-micron diameters, possibly due to vesicles enrichedin DOPA, hence re¯ecting improper mixing betweenDOPC and DOPA. Another possibility is that theseDOPA containing vesicles have an ellipsoidal shape andhence are partially oriented in the magnetic ®eld (Pottand Dufourc 1995). MLVs made of pure DOPC gave amore classical ``bilayer type'' spectrum (not shown).
When several freeze-thaw cycles are applied to thismixture, the overall NMR lineshapes reveal a decreaseof the average vesicle size. It also demonstrates the
Fig. 1 Freeze-fracture electronmicrographs (a, b, c) and nega-tive staining (d) of aqueousdispersions of DOPC/DOPA(80:20 mol%) after: 0 (a);10 (b and d) and 50 (c) cyclesof freeze-thawing
188
heterogeneity of vesicle sizes obtained by freeze-thawing.The narrow peak around 0 ppm corresponds to thepresence of small vesicles for which the anisotropy of thechemical shift tensor is averaged out. The amplitude ofthis peak increases with the number of cycles (compareFig. 4a and Fig. 4d). The overall lineshape may be
misleading because one sees only the two extreme spec-tral components associated with very large vesicles andvery small vesicles, respectively. In reality, there is acontinuous distribution of vesicle sizes. We have carriedout a series of spectral simulations to determine the mostlikely distribution in the case of: (1) a lipid dispersioncomposed either of DOPC or DOPC/DOPA (80:20mol%) with an increasing number of cycles, and(2) various lipid mixtures after 10 freeze-thaw cycles.
Several types of size distribution were tested andwithin each type of pro®le we varied the parameters to
Fig. 2 Histograms indicating the percentage of vesicles of variousdiameters counted on freeze-fracture electron micrographs after 10freeze-thaw cycles (in gray); the same ®gure shows the correspondinghistograms indicating the percentage of vesicles contributing to thetotal entrapped volume assuming spherical vesicles (hatched symbols)
189
optimize the matching between experimental and simu-lated spectra by comparing the integrals for spectraplotted with the same maximum intensity. The followingtype of distributions were used:
· Distributions corresponding to an exponentially de-creasing function starting at D0 with a characteristiclength Dc.
· Half-Gaussian distributions starting at D0, withcharacteristic width Dc.
· Gaussian distributions with characteristic width Dc,centered at a ®nite average diameter �D.
· Distributions decreasing as 1/D2, limited to an in-terval between D0 and Dmax.
In all cases, the minimum diameter D0 was set at 30 nm.This is somewhat arti®cial because in reality there is noreason a priori to impose a cut-o� at D0. However, thisprocedure is justi®ed by the fact that NMR spectra inany case would not be sensitive to small variations ofvesicle diameters below this minimum value D0. On theother hand, this method is very sensitive in the hundrednanometer diameter range, which makes it well suitedfor the present study.
Computer simulations showed that Gaussian distri-butions around a ®nite average diameter �D never permita good ®t of the experimental spectra (except in the caseof LUVs), neither does a half-Gaussian distribution.Figure 5a±d shows various attempts to simulate the ex-perimental spectrum of DOPC/DOPA mixtures after 20freeze-thawing cycles using an exponential, a half-Gaussian and a Gaussian function, respectively. Best ®tswere obtained with the exponential probability functiondecreasing from D0. We found that reasonable ®ts couldbe obtained also with a probability decreasing as 1/D2
(data not shown). In the latter case an upper limit Dmax
had to be set for normalization. However, there is nophysical argument to justify a discontinuous probabilityfor large diameters. The exponential frequency function,on the other hand, can be extended to in®nity; thenormalization is accounted for by the parameter Dc thatis related to the average diameter by the following re-lation: �D � Dc � D0.
In summary, the exponential probability functionwith a single parameter Dc permitted the best simula-tions of the NMR spectra recorded after freeze-thawing(Fig. 4b±d). Data summarizing �D values obtained fordi�erent lipid compositions are included in Table 1 to-gether with mean vesicle diameters deduced from elec-tron microscopy. There is a systematic di�erencebetween the average diameters deduced from electronmicroscopy and those deduced from NMR. Attempts toexplain this discrepancy will be presented in the Dis-cussion section.
In¯uence of cholesterol
The change in lineshape of 31P NMR spectra recordedwith DOPC/cholesterol mixtures (7/1 mol ratio) after 0,
Fig. 3 Theoretical 31P NMR spectra for homogeneous populations ofunilamellar vesicles of di�erent diameters
Fig. 4a±h Comparison between experimental and simulated 31PNMR spectra for DOPC/DOPA (80:20 mol%). Number of freeze-thaw cycles: 0 (a and e); 10 (b and f); 20 (c and g); 50 (d and h).Theoretical spectra f, g and h were obtained with a distribution ofvesicle diameters following an exponentially decreasing function. Theaverage diameters �D of the exponential distributions were 98, 85 and60 nm, respectively
190
1 or 10 freeze-thaw cycles (Fig. 6) con®rms electronmicroscopy observations (Fig. 2) which showed thefragmentation of DOPC/cholesterol vesicles. NMRcon®rms also that average diameters remain much largerthan in the case of DOPC or DOPC/DOPA vesicles (seeTable 1).
Lipid mixtures that do not fragment
Freeze-fracture electron microscopy was also carried outwith freeze-thawed suspensions of DMPC, DPPC, ESMor DLnPC. In contrast to the previous mixtures, thelatter lipid suspensions remained essentially multila-mellar even after 10 cycles. Electron microscopy as wellas 31P NMR carried out with these lipids revealed atonce that they do not fragment into small vesicles duringthe process of freeze-thawing, at least under our exper-imental conditions. In the case of DMPC/DMPA orDOPC with more than 30% cholesterol, NMR spectraobtained after 10 cycles indicated partial fragmentation.Figure 7 shows micrographs from freeze-fracture elec-tron microscopy and NMR spectra of some of theselipids after 10 cycles of freeze-thawing. The lineshapeobtained with ESM before freeze-thawing suggests par-tial orientation of the MLVs owing to the magnetic ®eldas reported already by several investigators (Brummet al. 1992; 1995; Picard et al. 1999; Pott and Dufourc,Seelig et al. 1985).
Large unilamellar vesicles
Repetitive freeze-thawing was also successful in reducingthe diameter of LUVs with an initial diameter of theorder of 200 nm as obtained by reverse phase evapora-tion. If freeze-thawing was carried out with LUVs inbu�er, NMR spectroscopy indicated a narrowing of the31P NMR spectrum (Fig. 8). Importantly, simulation of
Table 1 Average diameters of unilamellar vesicles obtained after10 freeze-thaw cycles of aqueous dispersions
a Lipids were dispersed in pH 8 bu�erbAverage diameter, �D � D0 � Dc (in nm), deduced from simulationusing an exponential probability function. A variation of �D of theorder of �2 nm su�ces to modify the lineshape, indicating that theprecision obtained for this parameter from the simulations is high.On the other hand, this does not prove the uniqueness of the modelto ®t the datacAverage diameter �D (in nm), obtained directly on the freeze-fracture electron micrographs. In parentheses is the value includingthe correcting factor (see text)d 50 freeze-thaw cycles
Fig. 5 Comparison between an experimental spectrum obtained after20 cycles of freeze-thawing (spectrum a) and simulated spectra withvarious distributions of vesicle sizes: b exponentially decreasingfunction with �D � 85 nm; c, half-Gaussian pro®le, starting atD0 � 30 nm with Dc � 140 nm; d Gaussian pro®le centered at�D � 265nm, with a Gaussian width of 110 nm. The spectra areplotted with the same maximum amplitude. Calculation of theirintegral indicates that if spectrum a corresponds to Ia � 1, then:Ib � 1.023; Ic � 1.115; Id � 1.082. For the simulations, T2 wascalculated with R1 � 20 Hz and 500 Hz for DOPC and DPA,respectively. R2 was ®xed at 600 Hz. Changing arbitrarily R1 and R2
values in order to broaden or to narrow the lines did not allow a better®t in the case of a Gaussian or a semi-Gaussian distribution of vesiclesizes
Fig. 6 Evolution of the 31P NMR spectrum obtained with anaqueous dispersion of DOPC/cholesterol (70:30 mol%) after 0, 1and 10 cycles of freeze-thawing, respectively
191
the experimental spectrum corresponding to LUVs withdiameters around 200 nm before freeze-thawing with aGaussian distribution centered around 200 nm gave agood ®t (Fig. 8a and b). On the other hand, the spectrumobtained after 10 cycles of freeze-thawing (Fig. 8c) ispractically identical to that obtained after 50 cycles whenstarting with MLVs (Fig. 4h), suggesting that fragmen-tation of vesicles by freeze-thawing reaches a limit.
Sonicated vesicles
When sonicated vesicles were subjected to cycles offreeze-thawing, the 31P NMR spectra were also modi®edand indicated the fusion and/or aggregation of at least afraction of the vesicles. The e�ciency of this processdepended on the bu�er: in pure water, after a singlefreeze-thaw cycle, sonicated DOPC SUVs appear to beheterogeneous with the coexistence of small vesicles andvery large vesicles (Fig. 9d); after ®ve cycles the smallvesicles had disappeared, as indicated by the broadNMR spectrum (Fig. 9f). If SUVs of DOPC were madein salt containing bu�er, only a small broadening of thenarrow peak associated with SUVs took place (Fig. 9cand e). Similarly, SUVs made of DOPC/DOPA(80:20 mol%) gave rise to a small line broadening whilewith DOPC/DOPA/LPC (60:13:27) no detectablebroadening was seen after 10 cycles (not shown). Elec-tron microscopy (Fig. 9g and h) demonstrates also thein¯uence of the bu�er and seems to indicate that fusionhappens rather than aggregation, in agreement withPick's measurements of the trapping capacity of soni-cated vesicles submitted to freeze-thawing (Pick 1981).
Discussion
Certain lipid mixtures containing unsaturated chains,when dispersed in bu�er and subjected to cycles offreeze-thawing, form unilamellar vesicles as reported byMac Donald et al. (1994). Here, we used liquid nitrogenfor the cooling and a water bath at 60 °C for heating.We have deliberately tried a single temperature forcooling and limited ourselves to one bu�er compositionsince an extensive study of the nature and concentrationof electrolytes had been done previously by MacDonaldet al. (1994). On the other hand, we have extended theassay to other lipid mixtures for which this protocol canbe applied with success and we have quanti®ed the un-ilamellarity and size distribution of the vesicles.
Electron microscopy as well as 31P NMR indicated arange of heterogeneity of vesicle sizes after freeze-thawing. Both techniques showed that the proportion ofvesicles of a given size can be represented by an expo-nentially decreasing function of the diameter, with acharacteristic averaged diameter �D function of thenumber of cycles. However, we found a systematic dif-ference between the average value determined from
Fig. 7a±d 31P NMR spectra and freeze-fracture electron micro-graphs of aqueous dispersions of lipids that do not fragment oronly partially fragment after 10 cycles of freeze-thawing. a and b:ESM; c and d: DLnPC. a and c show (dotted lines) the NMRspectra before freeze-thawing and (full line) the spectra after 10cycles. The freeze-fracture micrographs were obtained after 10cycles
Fig. 8 Comparison between experimental (a and c) and simulated (band d) 31P NMR spectra of LUVs made of DOPC/DOPA(80:20 mol%). Spectrum a was obtained after extrusion of liposomesthrough 200 nm pores before freeze-thawing; spectrum cwas obtainedafter 10 freeze-thaw cycles applied to the LUV suspension. Thetheoretical spectrum b was obtained by using a Gaussian distributionof vesicle diameters, centered at �D � 225 nm and a Gaussian width of81 nm. Spectrum d was obtained with an exponentially decreasingdistribution and �D � 65 nm. For more details, see text
192
NMR simulations and that deduced from electron mi-croscopy. In the latter case, the average diameter wasmultiplied by a factor between 1.5 and 2. One control ofthe validity of NMR simulation is our ability to simulatethe spectrum of LUVs obtained by phase reversion and®ltration at 200 nm. The average diameter deduced fromthe simulation was indeed very close to 200 nm. Nev-ertheless, one may ask if an erroneous value of the dif-fusion rate constant utilized in other NMR simulationscould explain the discrepancy with electron microscopy.In these calculations, we used Ddi� � 10)7 cm2/s whichis the value proposed by Galla et al. (1979) for DOPC.When examining the data in the literature correspondingto the di�usion of phospholipids in the ¯uid phasearound room temperature, values reported range from
1.3 ´ 10)7 to 10)8 cm2/s (Blume 1993; Devaux andMcConnell 1972). If a lower di�usion constant was used(say 0.5 ´ 10)7 cm2/s), it would be necessary to reducethe average diameter �D in the simulation. Indeed, slowerdi�usion imposes smaller vesicles in order to obtain thesame reorientation rate. Similarly, if the water viscosityis increased arti®cially in the calculation to take intoaccount the high concentration of vesicles, the orienta-tion averaging becomes less e�cient unless the averagediameters are again reduced. Thus, in both instances thediscrepancy between NMR and electron microscopy isaggravated.
In the case of NMR, small vesicles contribute verysigni®cantly to the lineshape, owing to the fact that theygive narrow and therefore intense peaks. Large vesiclesalso contribute to the spectral intensity owing to thelarge number of lipids in each vesicle. The latter con-tribution is in a di�erent region of the spectrum. Moreimportant, owing to the low sensitivity of NMR, a singlespectrum corresponds to the averaging of a very largenumber of vesicles, exceeding by several orders ofmagnitude the number of vesicles that can be reasonablycounted from electron micrographs.
A di�culty with electron microscopy is that vesiclesmay not be counted on a single micrograph if their di-ameters vary considerably, as is frequently the case.Indeed, large vesicles can be counted easily but it isdi�cult, if not impossible, to count very small vesicleson the same micrograph (i.e. with the same magni®ca-tion). Yet micrographs contain vesicles with diametersvarying from 30 nm to almost 1 lm. Thus, an underes-timation of the number of small vesicles that are countedon electron micrographs is highly probable.
Another problem is the actual vesicle shapes. Thecorrecting factor 1.28 which is applied to the size mea-surements in Table 1 (values between parentheses in thethird column) is only valid for perfect spheres (Heegaardet al. 1990). Indeed, freeze-fracture electron microscopyshows essentially spherical objects. However, negativestaining shows vesicles with more complicated shapes.(Fig. 1d). It is possible that glycerol is responsible for thesphericity of vesicles seen by freeze-fracture electronmicroscopy. Recently, we have observed LUVs bycryoelectron microscopy in the absence or presence ofglycerol. It appears that while vesicles are all spherical inthe presence of glycerol, in the absence of glycerol theirshape can be discoid, elongated or invaginated (TraõÈ kiaM, Lambert O, Rigaud J-L, Devaux PF, in preparation).If the vesicles which are used for NMR in the absence ofglycerol have surfaces covered with invaginations andprotrusions, then the orientation averaging will be muchfaster than with perfect spheres. Certain vesicles mayhave an ellipsoidal deformation. The NMR spectra inFig. 7 and perhaps spectrum a in Fig. 4 have featuresindicative of a partial orientation of the vesicles inthe magnetic ®eld. Such magnetic orientation due to theanisotropy of the lipid diamagnetism can happen in thecase of ellipsoidal deformation of the lipid vesicles (Pottand Dufourc 1995). Thus, the non-sphericity of the lipid
Fig. 9 31P NMR spectra and freeze-fracture electron micrographs ofsonicated dispersions of DOPC with bu�er (a, c, e, g) and in water (b,d, f, h). Number of freeze-thawing cycles: a and b, 0; c and d, 1; e, f, gand h, 5
193
vesicles could explain, at least partially, why NMR givessmaller �D values than electron microscopy.
In conclusion, both techniques have their limitationsat a quantitative level. However, they both indicate thesame trend: namely, that the unilamellar vesicle sizes arenot identical, the majority of the vesicles have a smalldiameter and the average diameter is reduced as thenumber of freeze-thaw cycles is increased. Table 1 indi-cates that after 10 freeze-thawing cycles the ``average''vesicle size is comparable to that of LUVs obtained by®ltration with an extruder. However, a large fraction ofthe vesicles have a much smaller diameter. Filtration offreeze-thawed vesicles will eliminate the very large vesi-cles remaining, but ®ltration will not permit one to ob-tain a homogeneous population of vesicles. Thus, inspite of its simplicity, the advantage of this technique forthe production of unilamellar vesicles might be ques-tioned if one needs a population of vesicles perfectlyhomogeneous in size. On the other hand, an advantageof the freeze-thaw technique is that it is not impeded bythe presence of charged lipids, unlike the extrudertechnique (unpublished results from our laboratory).
Is it possible to predict the e�ciency of freeze-thaw-ing for a given lipid composition and one can explain themechanism of fragmentation? As revealed by freeze-fracture electron microscopy, MLVs fragmentation intosmall unilamellar vesicles takes place progressively: ahomogeneous population of unilamellar vesicles requiresbetween ®ve and ten cycles of freeze-thawing. Althoughwe have not attempted a systematic investigation of theminimum number of cycles for each lipid compositionused in these experiments, it is apparent that pure DOPCfragments faster than DOPC/cholesterol mixtures. Forconcentrations of cholesterol above 30%, fragmentationwas not observed. However, there is an upper limit onthe cholesterol concentration that can be accommodatedwithin the bilayer structure and this limit strongly de-pends on the sample preparation procedure (Huanget al. 1999). Excess cholesterol will precipitate as crystalsof pure cholesterol monohydrate and could perturb lightabsorbence measurements. This observation may ex-plain the di�erence between our results and those ofMacDonald et al. concerning the DOPC/Chol mixtures.At any rate, membrane ¯uidity appears to be importantfor this process to take place: DMPC, DPPC and ESM,all lipids that do not fragment, are in a gel phase at thetemperature of ice formation; by contrast DOPC, POPCand EPC, lipids that do fragment, are ¯uid at thattemperature. However, as pointed out already by Mac-Donald et al., the fact that membranes are ¯uid or rigidwhen ice forms is not a de®nitive criteria since DLnPC,which is a lipid with double bonds and a transitiontemperature below 0 °C, does not fragment (see Fig. 7).Because fragmentation takes place only in the presenceof salt, it is likely, as pointed out by previous investi-gators, that osmotic e�ects are involved. Membranebreakage by osmotic shocks will be favored by the in-homogeneous distribution of ions in MLVs (Gruneret al. 1985), the entrapped bu�er exhibiting reduced
solute concentrations. It is noteworthy that very smallvesicles like SUVs are resistant to osmotic shocks; thusvesicles that do fragment seem to reach progressively astate where the surface tension associated with highcurvature is a maximum.
Acknowledgements The authors thank Dr. Erick J. Dufourcfor allowing us to use his simulation program. We also thank Dr.Richard Callaghan for critically reading the manuscript.
References
Blume A (1993) Dynamic properties. In: Cevec G (ed) Phospholi-pids handbook. Dekker, New York, pp 555±509
Brumm T, MoÈ ps A, Dolainsky C, Bayerl TM (1992) Macroscopicorientation e�ects in broad line NMR spectra of model mem-branes at high magnetic ®eld strength. A method for preventingsuch e�ects. Biophys J 61: 1018±1024
Burnell EE, Cullis PR, De Kruij� B (1980) E�ects of tumblingand lateral di�usion on phosphatidylcholine model mem-brane 31P-NMR lineshapes. Biochim Biophys Acta 603:63±69
Devaux P, McConnell HM (1972) Lateral di�usion in spin labeledphosphatidylcholine-multilayers. J Am Chem Soc 94: 4475±4481
Douliez JP, Bellocq AM, Dufourc EJ (1994) E�ect of vesicle size,polydispersity and multilayering on solid state 31P- and 2H-NMR spectra. J Chem Phys 91: 874±880
Dufourc EJ, Mayer C, Strorer J, Altho� G, Kothe G (1992) Dy-namics of phosphate head groups in biomembranes. A com-prehensive analysis using phosphorus-31 nuclear magneticresonance lineshape and relaxation measurements. BiophysJ 61: 42±57
Galla H-J, Hartmann W, Theilen U, Sackmann E (1979) On two-dimensional passive random walk in lipid bilayers and ¯uidpathways in biomembranes J Membr Biol 48: 215±236
Heegaard CW, Le Maire M, Gulik-Krzywicki T, Moller JV (1990)Monomeric states and Ca2+ transport by sarcoplasmic retic-ulum Ca2+ ATPase, reconstituted with an excess of phos-pholipid. J Biol Chem 265: 12020±12028
Hope MJ, Bally MB, Webb G, Cullis PR (1985) Production oflarge unilamellar vesicles by rapid extrusion procedure. Char-acterization of size distribution, trapped volume and ability tomaintain a membrane potential. Biochim Biophys Acta 812:55±65
Huang J, Buboltz JT, Feigenson GW (1999) Maximum solu-bility of cholesterol in phosphatidylcholine and phosphati-dylethanolamine bilayers. Biochim Biophys Acta 1417:89±100
MacDonald RC, Jones FD, Qiu R (1994) Fragmentation into smallvesicles of dioleoylphosphatidylcholine bilayers during freezingand thawing. Biochim Biophys Acta 1191: 362±370
Mayer LD, Hope MJ, Cullis PR, Jano� AS (1985) Solute distri-butions and trapping e�ciencies observed in freeze-thawedmultilamellar vesicles. Biochim Biophys Acta 817: 193±196
Oku N, MacDonald RC (1983) Di�erential e�ects of alkali metalchlorides on formation of giant liposomes by freezing andthawing and dialysis. Biochemistry 22: 855±863
Picard F, Paquet MJ, Levesque J, Be langer A, Auger M (1999) 31PNMR ®rst spectral moment study of the partial magnetic ori-entation of phospholipid membranes. Biophys J 77: 888±902
Pick U (1981) Liposomes with a large trapping capacity preparedby freezing and thawing of sonicated phospholipid mixtures.Arch Biochem Biophys 212: 186±194
194
Pott T, Dufourc EJ (1995) Action of melittin on the DPPC-cholesterol liquid-ordered phase: a solid state 2H- and 31P-NMR study. Biophys J 68: 965±977
Rance M (1981) PhD thesis. University of Guelph, CanadaRance M, Byrd RA (1983) Obtaining high-®delity spin-1/2 powder
spectra in anisotropic media: phase cycled Hahn echo spec-troscopy. J Magn Res 52: 221±240
Roux M, Huynh-Dinh T, Igolen J, Prigent Y (1983) Simple prep-aration of 1,2-dipalmitoyl-sn-glycero-3-phopshoric acid anddeuterated choline derivatives. Chem Phys Lipids 33: 41±45
Seelig J (1978) 31P nuclear magnetic resonance and head groupstructure of phospholipids in membranes. Biochim BiophysActa 515: 105±140
Seelig J, Borle F, Cross TA(1985) Magnetic ordering of phos-pholipid membranes Biochim Biophys Acta 814: 195±198
Smith ICP, Ekiel IH (1984) Phosphorus-31 NMR of phospholipidsin membranes. In: Gorenstern D (ed) Phosphorus-31 NMR.Principles and applications. Academic Press, New York, p 447
Swairjo MA, Seaton BA, Roberts MF (1994) E�ect of vesiclecomposition and curvature on the dissociation of phosphatidicacid in small unilamellar vesicles: a 31P-NMR study. BiochimBiophys Acta 1191: 354±361
Szoka FC, Olson F, Heath T, Vail WJ, Mayhew E, Papahadjo-poulos D (1980) Preparation of unilamellar liposomes of in-termediate size (0.1±0.2 microns) by a combination of reversephase evaporation and extrusion through polycarbonatemembranes. Biochim Biophys Acta 601: 559±571
TraõÈ kia M, Langlais DB, Cannarozzi, GM, Devaux PF (1997) Highresolution spectra of liposomes using MAS NMR. The case ofintermediate size vesicles. J Magn Res 125: 140±144

























![pH gradient loading of anthracyclines into cholesterol ... · sively used in the preparation of liposomes for improving transdermal liposomal drug delivery [25], improving encap-sulation](https://static.documents.pub/doc/80x56/5ec666a52a5b1b797e2a9fef/ph-gradient-loading-of-anthracyclines-into-cholesterol-sively-used-in-the-preparation.jpg)


