Page 1
UNIVERSITY of OULU
OULUN YLIOPISTO
Pro gradu
Isolation and Characterization of Extracellular
Vesicles (EVs) from Renal Carcinoma Cells
Khem Raj Giri
University of Oulu
Faculty of Biochemistry and Molecular Medicine, Biocenter Oulu
2016
Page 2
This work was performed at the Department of Biochemistry, University of Oulu, Biocenter
Oulu.
Oulu, Finland.
Supervisor:
Professor Seppo Vainio
Co-supervisor:
Dr. Anatoliy Samoylenko
Page 3
Contents
Acknowledgements
Abbreviations
1 REVIEW OF THE LITERATURE 1
1.1 Introduction 1
1.2 Extracellular vesicles 2
1.2.1 Apoptotic bodies 5
1.2.2 Microvesicles 5
1.2.3 Exosomes 6
1.3 Exosomes in cancer 7
1.4 Hypoxia and exosomes 8
1.5 Importance of EVs in cancer 10
1.5.1 As biomarkers 10
1.5.2 As therapeutic agents 12
2. AIM OF THE PROJECT 14
3. MATERIALS AND METHODS 15
3.1 Cell culture 15
3.2 Exosome isolation 15
3.3 Exosome labeling 17
3.4 SDS-PAGE 17
3.5 Western blot 18
3.6 Cell migration assay 19
3.7 Cell proliferation assay 19
3.8 Mass spectrometry 20
3.9 Immunohistochemistry 20
3.10 RNA extraction 21
Page 4
3.11 Electron microscopy 22
3.12 Injection of chicken embryos: ex ovo culture 23
4. RESULTS 24
4.1 Characterization of EVs 24
4.1.1 Electron microscope analysis 24
4.1.2 Western Blot 25
4.1.3 Nanoparticle tracking analysis (NTA) 26
4.2 Effects of exosomes 28
4.2.1 Exosome labeling 28
4.2.2 Cell proliferation and motility assay 29
4.2.3 Injection of chicken embryos 30
4.3 Protein analysis by mass spectrometry 31
4.4 RNA analysis 33
5. DISCUSSIONS 34
6. CONCLUSIONS 37
7. REFERENCES 38
Page 5
Acknowledgements
I would like to thank Department of Biochemistry, University of Oulu, Biocenter Oulu, Oulu,
Finland for providing me opportunity to carry out this pro-gradu work. My heartily gratitude goes
to Professor Seppo Vainio for his excellent supervision and continuous support throughout my
work. I am very thankful to my co-supervisor Dr. Anatoliy Samoylenko for his supervision
during my work. His inspiration and guidance were much valuable without which this work
would be incomplete.
I am grateful to my lab technicians Paula Haipus, Hannele Harkman and Johanna Kekolahti-Liias
for technical supports, Dr. Ilya Skovorodkin for his help during in-vivo chicken work. Dr. Timo
Pikkarainen being helpful during the lab works. I am also grateful to my office colleagues
Prateek Singh and Dr. Genevieve Bart for their support throughout my working period.
I am also thankful to Professor Lloyd Ruddock, coordinator of international master´s programme
for the opportunity to be a part of this excellent programme. His help during initial days of the
course is very appreciable.
I would also like to remember my friends Chandan, Siva, Zenglai, Bikram and Histoni for being
there when I needed break from my work. Thank you guys!
Last but not the least, my sincerest appreciation goes to my family for their continuous
encouragement and inspiration throughout my whole life.
Page 6
Abbreviations
RCC Renal cell carcinoma
DC Dendritic cell
EVs Extracellular Vesicles
MSCs Mesenchymal stromal cells
ABs Apoptotic bodies
PS Phosphatidylserine
PBS Phosphate Buffered Saline
MVs Microvesicles
MVBs Multivesicular bodies
HSP Heat shock protein
MHC Major Histocompatibility
HIF Hypoxia inducible factor
MM Metanephric mesenchyme
YFP Yellow fluorescent protein
WT Wild type
DMEM Dulbecco’s Modified Eagle medium
DMSO Dimethyl sulfoxide
HM Homogenization medium
WB working buffer
SDS Sodium dodecyl sulfate
BSA Bovine serum albumin
TEM Transmission electron microscopy
NTA Nanoparticle tracking analysis
LCMS Liquid chromatography mass spectrometry
Rpm Revolutions per minute
Page 7
1
1. LITERATURE SECTION
1.1 Introduction
Kidney cancer is one of the major types of cancer among people, being 7th
most common
cancer among men and 11th
among women. This accounts for around 3% among all cancers
occurring in Europe alone. In 2012, around 115,000 new cases were reported and more than
49,000 deaths were recorded in 40 European countries because of kidney cancer (Steliarova-
Foucher et al. 2014). The mortality rate due to this cancer is higher in Europe and North
America in comparison to Asia and South America. Kidney cancer includes multiple tumors
that originate in kidney, renal pelvis as well as ureter and other urinary organs (Marcos-
Gragera et al. 2015).
Renal cell carcinoma (RCC) is the most common renal tumor, accounting more than 90%
renal malignancies (National Comprehensive Cancer Network 2009). The major factor that is
responsible for increasing malignancy rate is lifestyle. Major etiological factors include sex,
race, age, smoking habit and obesity or overweight. Beside this, genetic factors, nutritional
factors and occupations are also involved. Thus RCC still remains a major challenge for the
researchers and effective therapeutics able to inhibit growth and propagation of tumor is an
urgent need.
Exosomes are the membrane vesicles between size 30-100 nm (Raposo et al. 1996). They are
formed by inward budding from multivesicular endosomes fused with plasma membrane.
Exosomes are secreted by different cells including RBC, B and T lymphocytes, platelets and
dendritic cells (DCs) (Denzer et al. 2000). Exosomes has been purified from different body
fluids such as blood, saliva, urine, serum and nasal secretion.
Role of exosome in kidney cancer is not fully understood yet and is currently deriving much
interest among researchers. The purpose of our study was to find the general effect of kidney
cancer derived exosomes in tumorigenesis. The work includes characterization of these
Page 8
2
exosomes using different techniques, study of effects of these exosomes in tumor formation,
cell proliferation and cell motility. The study also includes identification of cargo proteins
and miRNAs from exosomes.
1.2 Extracellular vesicles
Extracellular Vesicles (EVs) are nano sized vesicles secreted by various cell types including
B and T lymphocytes, mast cells, platelets, dendritic cells and mesenchymal stromal cells
(MSCs) derived from different tissues, and can be isolated from cell culture supernatant and
different biological fluids like urine, blood, semen, amniotic fluid etc. Wide variety of
organisms ranging from simple prokaryotes to higher eukaryotes releases EVs. EVs can be
categorized as exosomes, microvesicles and apoptotic bodies on the basis of size,
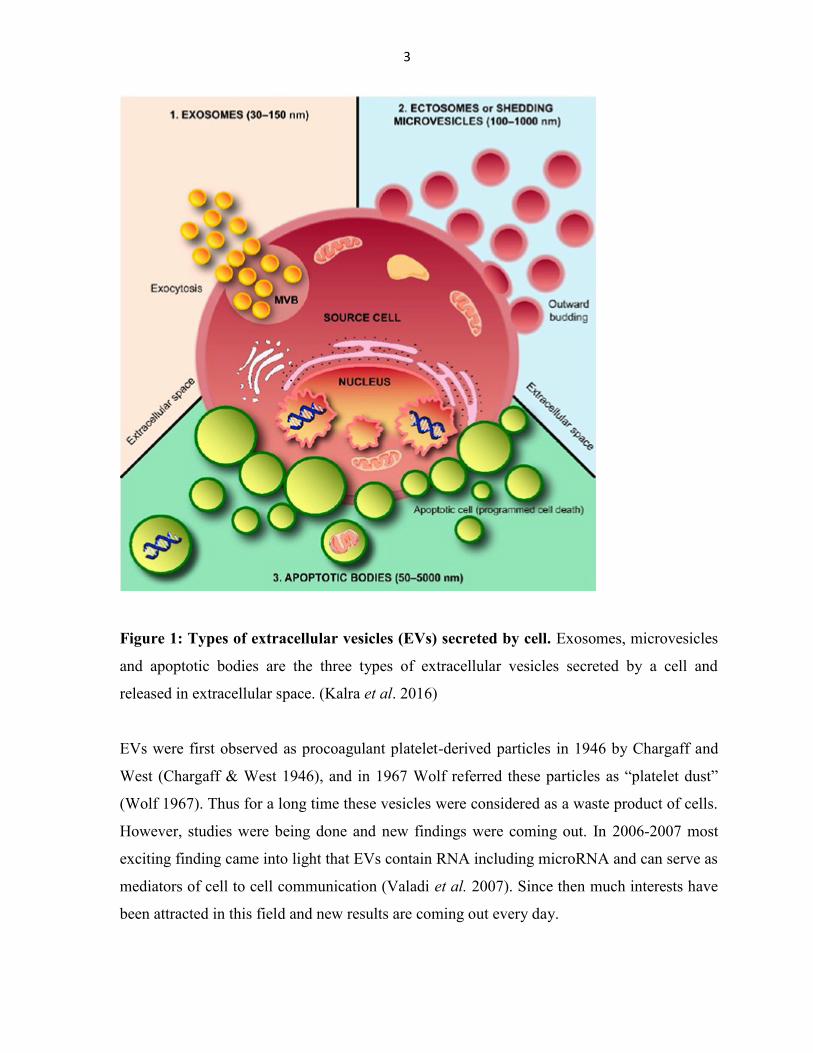
morphology, origin and the way they are released in the microenvironment (fig.1) (Nawaz et
al. 2014). Exosomes originate from multivesicular bodies, microvesicles are derived from the
plasma membrane and apoptotic bodies are produced from cells undergoing apoptosis.
Page 9
3
Figure 1: Types of extracellular vesicles (EVs) secreted by cell. Exosomes, microvesicles
and apoptotic bodies are the three types of extracellular vesicles secreted by a cell and
released in extracellular space. (Kalra et al. 2016)
EVs were first observed as procoagulant platelet-derived particles in 1946 by Chargaff and
West (Chargaff & West 1946), and in 1967 Wolf referred these particles as “platelet dust”
(Wolf 1967). Thus for a long time these vesicles were considered as a waste product of cells.
However, studies were being done and new findings were coming out. In 2006-2007 most
exciting finding came into light that EVs contain RNA including microRNA and can serve as
mediators of cell to cell communication (Valadi et al. 2007). Since then much interests have
been attracted in this field and new results are coming out every day.
Page 10
4
The various EVs from body fluids and cell culture can be isolated and characterized by using
different protocols. The presence of membrane fragments and all types of vesicles in the
starting material can lead to the contamination during specific EVs purification. Therefore,
standardization of protocol for effective purification and analysis of EVs is a major challenge
at the moment. The problem also arises due to similar or overlapping characteristics among
various vesicles and lack of proper biomarkers that can discriminate these vesicles in an
effective way (Gould & Raposo 2013).
The major components of EVs include mRNAs, miRNAs, DNA, proteins and lipids. EVs
have abilities to transfer these materials to the neighboring cells, thus introducing changes in
their phenotype along with the microenvironment (Camussi et al. 2011). So this has been
recognized as new effective means of cell to cell communication in both prokaryotic and
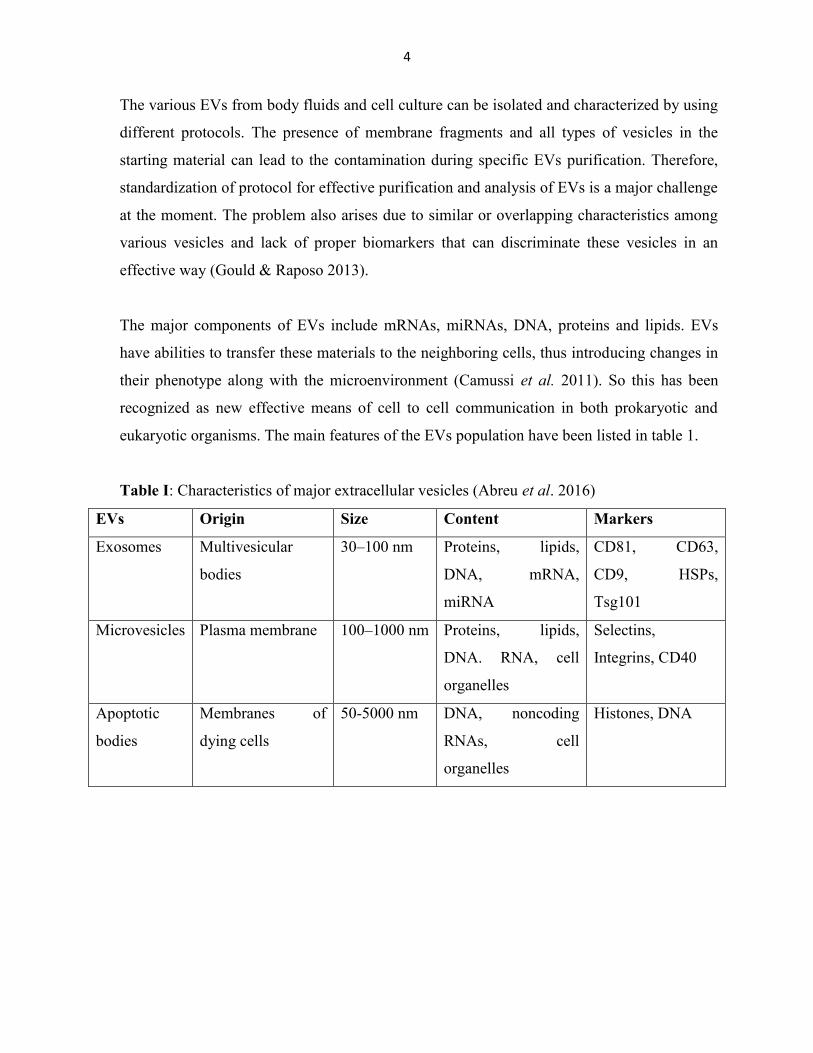
eukaryotic organisms. The main features of the EVs population have been listed in table 1.
Table I: Characteristics of major extracellular vesicles (Abreu et al. 2016)
EVs Origin Size Content Markers
Exosomes Multivesicular
bodies
30–100 nm Proteins, lipids,
DNA, mRNA,
miRNA
CD81, CD63,
CD9, HSPs,
Tsg101
Microvesicles Plasma membrane 100–1000 nm Proteins, lipids,
DNA. RNA, cell
organelles
Selectins,
Integrins, CD40
Apoptotic
bodies
Membranes of
dying cells
50-5000 nm DNA, noncoding
RNAs, cell
organelles
Histones, DNA
Page 11
5
1.2.1 Apoptotic bodies
Apoptotic bodies (ABs) are the least studied members among extracellular vesicles. These
are released from the cells which undergoes apoptosis and the size are measured between 50-
5000nm (Simpson & Mathivanan 2012). Apoptosis is important process and helps in removal
of damaged, infected and old cells from tissues. So, apoptotic bodies are basically packed
structure of cells and cellular debris that needs to be removed. The positive staining for
Annexin V (Hristov et al. 2004) and the presence of phosphatidylserine (PS) on external part
(Martin et al. 1995) makes it similar to microvesicle (MV) and hence difficult to distinguish
from microvesicle.
Since apoptotic bodies were considered as garbage bags for a long time, relatively less is
known about the composition of Abs. However, Mallat et al. studied the ABs of human
monocytic and lymphocytic from atherosclerotic plaques and found these ABs to contain PS,
coagulation factor III and annexin A5 (Mallat et al. 1999). Proteomics studies of ABs from
thrombocyte of mice has identified 142 proteins ranging from heat shock protein, histone
related, cytosolic, oncogenes and few other proteins that are immunologically important
(Turiak et al. 2011).
Further, ABs contains multiple cell organelles that are packed closely along with nuclear
fragments which can be seen by electron microscopy (Elmore 2007). Also, ABs containing
DNA and RNA separately that is rich in either one of these component have been observed
(Halicka et al. 2000). ABs of tumor cells have shown ability to suppress CD8+ cell responses
in vivo (Xie et al. 2009) and growth factor VGEF was found to be expressed higher,
suggesting possible role of apoptotic bodies in tumor propagation.
1.2.2 Microvesicles
Microvesicles (MVs) represent the extracellular vesicles between 100-1000 nm in size. MVs
are released by normal and diseased cells and are often grouped together with exosomes but
these vesicles are unique populations with distinct structures from exosomes.
Page 12
6
MVs are formed by budding and fission of plasma membrane and released into extracellular
space; process which is dependent in calcium signaling and enzyme activity. The changes in
calcium level leads to the activation of calcium dependent enzymes such as scramblase and
floppase along with lipid composition of the plasma membrane (Piccin et al. 2007). One of
the main features of MVs is externalization of phosphatidylserine (PS) which is located on
the outer membrane of microvesicle shedding site. However, MVs without secreted PS has
also been reported (Barteneva et al. 2013).
The function of microvesicles depends on the cargo they carry. Apart from carrying genetic
materials like DNA, mRNA and miRNA, microvesicles also carry transmembrane proteins
and cellular receptors on their surface thus reflecting the characteristics of their parent cells.
Differentiated cells and tumor cells release microvesicles carrying miRNAs or mRNAs in
most of the cases (Mytar et al. 2008).
1.2.3 Exosomes
Exosomes are small vesicles ranging from 30 – 100 nm in size and originate from
multivesicular bodies (MVBs). MVBs fuse with the plasma membrane and their intraluminal
vesicles are released as exosomes in extracellular space (Harding et al. 1983). The term
“exosomes” was given by Trams et al in 1981 (Trams et al. 1981). Exosomes are separated
by multiple number of cells including B and T cells, dendritic cells, mesenchymal stem cells,
epithelial cells, endothelial cells and cancer cells both at normal as well as pathological
condition (Henderson & Azorsa 2012). Apart from this, exosomes are also identified in
different types of body fluids such as urine, blood, serum, saliva and nasal secretion.
Interestingly, it has been shown that the amount of exosomes secreted by cancer cells is
higher than the normal cells (Taylor & Gercel-Taylor 2008; Logozzi et al. 2009).
The major components of exosomes are Proteins, lipids, DNA, miRNA and mRNA. The
study carried out in different cell type shows that exosomes contain 4,563 proteins, 194
lipids, 1,639 mRNA and 764 miRNAs (Mathivanan et al. 2012; Mathivanan & Simpson
2009) making exosomes a complex molecule. The major proteins identified are fusion
proteins, CD63, CD81, CD9, CD82, heat shock proteins (HSP70, HSP90), signal
Page 13
7
transduction proteins, cytoskeletal proteins along with cell type-specific proteins like major
histocompatibility complex (MHC) I and II, A33 antigen (Mathivanan & Simpson 2009) and
differ according to the cells from which they origin.
Lipids in exosomes are less characterized in compared to other components. Exosomes have
their own specific lipid composition differing from parental MVB but still reflect the cells of
origin. Their functions are highly contributed by bioactive lipids such as prostaglandins
(Subra et al. 2010). DNA and RNA including mRNA and miRNA are other major
compositions of exosomes in addition to protein. The presence of RNA makes exosomes as a
significant pathway of cell to cell communication. These mRNAs are found to be functional
and transferable within target cells where they can be translated into proteins (Ratajczak et
al. 2006). Thus mRNA and miRNA both perform specific functions upon their transfer to
other cells indicating that exosomes are potential route for signaling between cells.
1.3 Exosomes in cancer
Intercellular communication is an important process by which cells transmit and receive
signals between nearby or distant cells. Previously, it was thought that intercellular
communication occurs by direct cell-to-cell contact and secreted soluble proteins. Recent
studies have shown a new way of communication among cells mediated by extracellular
vesicles (Thery et al. 2009). These vesicles are effective for signaling among neighbor cells
as well as distant cells.
Metastasis is one of the major causes of deaths in cancer. The mechanism by which the
cancers cells escape the site of primary tumor and migrate to distant cells are not yet fully
understood (Nguyen et al. 2009). For effective metastasis, cells from primary tumor escape
into blood or lymph and grow tumor in a distant cell with favorable microenvironment for
tumor propagation. There is growing evidence that exosomes might play an important role in
this process (Yang C & Robbins PD 2011). Exosomes are capable of passing proteins,
miRNAs as well as anti-cancer drugs from the cells and release regulatory and signaling
molecules (Lee et al. 2011). Some factors released from exosomes can enhance stromal
Page 14
8
remodeling and transformation of hypoxia-mediated mesenchyme which is important for
cancer development (Nieto 2011). By desmoplastic reaction, exosomes can lead to fibroblast
proliferation and can cause immune escape by suppressing antigen specific immune
responses or by upregulating immunosuppressive cell differentiation (Azmi et al. 2013). This
shows that exosomes are critical for tumorigenesis.
Some studies have shown the potential of exosomes for cross tumor propagation. Exosomes
from renal cells have shown to initiate angiogenesis in lung cancer ascites (Nguyen DX &
Massague J 2007) and exosomes from human bone marrow mesenchymal stem cells have
shown to stimulate proliferation of proximal tubular cells by transferring hepatocyte growth
factor-1 receptor mRNA (Tomasoni et al. 2013). These results suggest that exosomes trigger
tumor propagation by horizontal transfer of genetic materials.
There are different mechanisms by which exosomes interact with the recipient cells. One
mechanism involves the binding of exosomal membrane proteins with the ligands of the
recipient cell membrane such as transferrin receptor, integrins, tumor necrosis factor
receptors and tetraspanin proteins like CD9, CD63 and CD81 (Chen et al. 2010). Another
mechanism involves the fusion of exosomes with the membrane of the recipient cell thus
resulting in the transfer of exosome contents such as proteins, miRNAs, mRNAs and other
signaling molecules or endocytosis (Hemler 2003; Tien et al. 2010).
Together with transferring cargo to various cells, exosomes have ability to protect its
contents from degradation due to several degradative enzymes such as RNase, trypsin and
other chemical components due to the presence of lipid bilayer in exosomes (Koga et al.
2011).
Page 15
9
1.4 Hypoxia and exosomes
Adequate oxygen is necessary for normal cellular function and energy metabolism in
organisms. Hypoxia refers to the condition of insufficient oxygen supply and in most of the
time associated with promoting different diseases such as tumor spreading, pre-eclampsia
(PE) and venous thromboembolism (VTE) (Belting & Christianson 2015). Hypoxia inducible
factors (HIFs), are the transcription factors associated with the oxygen homeostasis (Kaelin
& Ratcliffe 2008). HIFs are formed by two subunits, HIF-α and HIF-β and are responsible
for maintenance of different cellular responses (Zhong et al. 1999). HIF dependent hypoxic
response depends on intercellular signals, which then plays active role to provide adaptive
mechanism for cell survival and tissue remodeling in the hypoxic microenvironment. It was
also recently found that hypoxia increased the secretion of tumor enhancing factors which
affects the surrounding tissues in the tumor microenvironment (Chaturvedi et al. 2013).
Many studies have shown that hypoxia enhance the exosomes release in various tumors. In
breast cancer, hypoxia mediated activation of HIF-1 α have been found to increase release of
exosomes which was confirmed by the lower release of exosomes when HIF-1α was
knockdown (King et al. 2012). Extracellular vesicles include a diverse range of vesicles and
hence are conceivable to believe that different mechanisms play a role in hypoxia-regulated
formation and secretion of these vesicles in cell dependent manner.
Hypoxia not only affects during secretion of vesicles but also have great impact on the
messages contained within the vesicles. The messages contained and transferred by the
vesicles are affected by hypoxia resulting in effects in functions. The study showed that
angiogenic pathways and metastatic potential are enhanced by hypoxic microenvironment
through secretion of exosomal proteins transpanins and alix while studying A431 carcinoma
cell line when subjected to hypoxia and then back to oxygen (Park et al. 2010). These studies
show that hypoxia actually promotes the secretion of exosomes inorder to create
microenvironment which favors pro-angiogenic environment which results in the
development of tumors aggressively.
Page 16
10
1.5 Importance of EVs in cancer
1.5.1 As biomarkers
The novel biomarkers play important roles in detection, prognosis and diagnosis of the disease,
particularly in cancer and exosomes have potential to act as significant biomarkers (fig.2).
Exosomes carries various intercellular molecules, hence may act as a source of unidentified
important biomarkers which have potential to predict the disease before it spreads completely
and patient goes under formal diagnosis such as MRI, CT or PET scan (Cho et al. 2012;
Paidpally et al. 2012). Since exosomes display specific expressions which are most of the time
representation of disease state, and also these exosomes can be detected in the body fluids,
making these vesicles ideal candidate for biomarkers of the tumor propagation (Tickner et al.
2014). Both exosomal proteins and RNAs serve as potential biomarkers.
In kidney, proteomics studies of urinary exosomes have analyzed proteins from different parts of
nephron and several biomarkers have been identified. These proteins are effective for
pathological conditions such as acute kidney rejection, acute kidney injury as well as diabetic
nephropathy (Ling et al. 2010; Hongjuan et al. 2009). One study suggested that exosomes
containing aquaporin-1 protein´s excretion decreased in rats under I/R up to 96 hours but after
480 hours, it returned to normal level. The similar result was later observed in patient with renal
allograft transplantation, suggesting that exosomes carrying aquaporin-1 protein is able to detect
I/R induced injury of renal cells. This can be used to predict post transplantation failure (Sonoda
et al. 2009).
More than 1000 proteins have been identified in humans by analyzing urinary exosomes and
among them 34 proteins have been found to be associated with kidney diseases like polycystin-1
protein in autosomal dominant polycystic kidney disease type 1, aquaporin-2 protein in
autosomal dominant and recessive nephrogenic diabetes and Na-K-2Cl symport protein in
antenatal barter syndrome type 1 (Miranda et al. 2010).
Page 17
11
Along with proteins, exosomes also contain mRNA and miRNA which can be used as disease
biomarkers. Studies have shown that miRNA of circulating exosomes are useful in diagnosis of
cancers including lung and ovarian cancer (Taylor & Gercel-Taylor 2008; Rabinowits et al.
2009). In kidney, various miRNAs (miR-200a, miR-200b and miR-429) were found to be
lowered than the normal level in the patient with immunoglobulin A nephropathy and the level
of reduction was associated with the disease severity (Wang et al. 2010). It is also found that
urinary granzyme A mRNA level can distinguish person between diseases cellular rejection and
acute kidney injury (Van Ham et al. 2010). These studies show that the miRNAs and mRNAs are
not only associated with the disease biomarkers but also changes in their level in urinary
exosomes can help to detect the progression of diseases.
Page 18
12
Figure 2: Role of extracellular vesicles as therapeutic agents, biomarkers and in drug
delivery. EVs can activate immune response as well as suppress inflammation, can act in blood
coagulation process, help in communication for neurons and can repair injured tissues, thus
making them ideal for therapeutic agents. Their ability to transfer RNAs to surrounding cells
make them suitable candidate for drug delivery. Also proteins present in these vesicles can act as
biomarkers (Andaloussi et al. 2013).
1.5.2 As therapeutic agents
The ability of exosomes to transfer mRNAs and miRNAs between cells of origin and target cells
opens a new concept of using exosomes as delivery vehicles in therapeutics (fig.2) (Valadi et al.
2007). These transfers can either increase or decrease in the level of protein expression within
the target cells. Angiogenesis in endothelial cells is caused by endothelial progenitor cell-derived
exosomes by possible mechanism of transferring mRNA and miRNA molecules in the target
Page 19
13
cells (Deregibus et al. 2007). Thus exosomes can influence the gene expression of the target
cells.
Exosomes can also serve as important vehicles in the delivery of specific antigens (fig.2). The
idea has been useful in vaccination against acute respiratory syndrome where exosomes loaded
with acute respiratory syndrome S protein (Kuate et al. 2007) against Toxoplasma gondii where
exosomes loaded with specific antigen (Beauvillain et al. 2009) showed positive effects. The
antibodies were neutralized to the most extent and in the case of T. gondii the disease level was
seen to be reduced in mice.
Since exosomes are secreted by almost every type of cells and are able to transfer their cargos
among cells proves their ability to deliver drugs, especially the delivery of therapeutic nucleic
acids, fig. 1(b). Thus exosomes can be considered as ideal nanovectors for drug delivery while
compared to synthetic lipid vesicles or liposomes (Fais et al. 2013). In 2010, the first publication
was done on vesicle mediated transfer of nucleic acid where THP-1 monocyte was transfected
with miR-150 and secreted vesicles were found containing miR-150, which was delivered to the
recessive, cells (Zhang et al. 2010). The capacity of such vesicles in delivery can be an important
success in gene therapy where these vesicles can be engineered to deliver regulatory miRNAs,
siRNAs or even tumor suppressors like PTEN (Gabriel et al. 2013).
The possible role of exosomes in the kidney diseases is being studied. Mostly, the exosomes
have been found to be associated in the paracrine effects in stem cell therapy. During stem cell
therapy of kidney injury, mesenchymal stem cells were able to recover partially through
paracrine factors released from exosomes (Gatti et al. 2011). Similarly, in stem cell therapy of
glomerulonephritis in rats (anti-Thy 1.1 glomerulonephritis), paracrine factors from injected
mesenchymal stem cells were helpful than the factors derived from cells themselves (Kunter et
al. 2006). In these paracrine effects, exosome secretion was found to be involved. Thus
exosomes can have a significant importance in treatment of different diseases including kidney
diseases.
Page 20
14
2. AIM OF THE PROJECT
The general aim of this thesis is to study the role of renal tumor cell derived exosomes in kidney.
The specific aims of this thesis are:
To purify and characterize exosomes derived from mouse kidney cancer cells (Renca).
To study the effect of exosomes in mouse embryonic kidney (MM) cells
To see whether Renca derived exosomes have role in tumor growth
To study proteins and RNA contents of exosomes for biomarker discovery
Page 21
15
3. MATERIALS AND METHODS
3.1 Cell culture
The cell lines used in this study were renal carcinoma (Renca WT) cell line (ATCC® CRL-
2947 TM
, Manassa, VA) and Renca cells stably expressing enhanced yellow fluorescent
protein (Renca YFP), MK3 (early metanephric mesenchyme) and MK4 (later induced
metanephric mesenchyme) (Valerius et al. 2002). The Renca cells were cultured in
Dulbecco’s modified Eagle medium (Life Technologies, UK): nutrient mixture F-12
(DMEM/F12), MK3 and MK4 cells were cultured in normal DMEM, both supplied with
10% FBS and 100 U penicillin/ 0.1 mg/ml streptomycin.
During passage of the cells, the cells were washed with PBS one to two times, trypsin was
added and plates were incubated for 4 minutes at 37°C in order to detach the cells. After
adding media, the cells were splitted in the ratio of 1:3 - 1:5. Cells were cultured in an
incubator at 37°C and 5% CO2 (Panasonic Healthcare, Japan). The cells were frequently
checked for contamination.
To maintain cell lines for a long period of time, cells were trypsinized and centrifuged at
1500 rpm for 4 minutes. The pellets were then dissolved in dimethyl sulfoxide (DMSO) 5-
10%, put in cryogenic tube and stored at -70°C overnight. The tubes were then finally
transferred to liquid nitrogen for long term storage.
3.2 Exosomes isolation
The cells were cultured in 20 big plates until confluency reached up to 80-90%. The previous
media was removed and exosome free media was added. Half number of plates were
incubated in normal condition (37°C and 5% CO2) while the remaining plates were incubated
in hypoxic condition (37°C and 1% O2). After 24 hours of incubation, isolation was carried
out by ultracentrifugation and density-gradient separation method.
Page 22
16
For ultracentrifugation, the cell supernatants were collected and centrifuged at 3000 rpm for
10 minutes to pellet cells and debris. The supernatant was then centrifuged at 2000 rcf and
4°C for 15 minutes inorder to get rid of oncosomes. MVs were pelleted by ultracentrifugation
at 20,000 rcf and 4°C for 45 minutes. The supernatant was filtered using 0.22-micron filter
and finally ultra-centrifuged at 108,800 rcf and 4°C for overnight or 18-22 hours (Sorvall
WX90, Thermo Scientific, US) for exosome isolation. The supernatant was discarded and the
pellet was dissolved in 1x filtered PBS.
For density gradient separation, the cell supernatants were collected in centricon plus-70
filter device (Merck Millipore, Ireland) and concentrated by centrifugation at 3500 rpm and
4°C for 40 minutes. The concentrated sample was recovered by short spin at 1000 rpm for 2
minutes. Homogenization media (HM) and working solution buffer (WB) were prepared.
The working solution (WS) was prepared by adding 1 volume of WB to 5 volumes of
optiprep solution (Sigma). The gradient solution was prepared in a following way:
Percentage (%) WS (ml) HM (ml)
5 1 9
10 2 8
15 4 6
20 8 2
Each gradient solution was sequentially layered on the top of each other in polyallomer
ultracentrifuge tube. The concentrated sample was finally added on the top of the gradient.
The gradients were ultracentrifuged at 24,200 rcf, 4°C for 18 hours (overnight). 1 ml
fractions were collected and diluted in 1x PBS and centrifuged at 24,200 rcf, 4°C for another
3 hours. The supernatant was discarded and pellet was resuspended in 1x PBS.
Page 23
17
Gradient solutions were prepared as follows:
Homogenization medium (HM)
0.25 M sucrose
1 mM EDTA
10 mM Tris-HCl
pH 7.4
Working solution buffer (WB)
0.25 M sucrose
6 mM EDTA
60 mM Tris-HCl
pH 7.4
3.3 Exosomes labeling
10x Exo-Red was added to isolated exosomes in a 1.5 ml eppendorf tube and mixed gently
without vortex. The solution was incubated in 37°C for 10 minutes. The labeling reaction
was stopped by adding ExoQuick-TC reagent (System Biosciences, CA) to the labeled
exosomes samples and mixed by inverting. The tube was incubated on ice for 30 minutes.
The sample was centrifuged at 14,000 rpm for 3 minutes. The supernatant was removed and
the pellet was suspended in 1x PBS. The exosomes were labeled and ready to be monitored
for tracking.
3.4 SDS-PAGE
Before characterization of exosomes by western blot, proteins were first separated by sodium
dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). 10% resolving gel was first
prepared and after its polymerization, 4% stacking gel was layered on the top of resolving
gel. Protein samples were mixed with 1x laemmli buffer and heated at 95°C for 5 minutes.
The 2-mercaptoethanol reduced disulfide bonds and SDS denatures the proteins and provide
Page 24
18
negative charge which will now separate on the basis of molecular weight. The samples were
loaded on each well and run for 90 minutes at 15 mA current. Protein ladder was used
inorder to determine the protein size separated in the gel.
10% resolving gel
H2O 5.22ml
40% acrylamide 2.6ml
1.85M Tris, pH 8.8 2ml
10% SDS 100µl
10% APS 50 µl
TEMED 10 µl
4% stacking gel
H2O 3.92ml
40% acrylamide 500µl
1.85M Tris, pH 6.8 500µl
10% SDS 50µl
10% APS 31µl
TEMED 6.3µl
3.5 Western blot
After completing SDS-PAGE, the proteins were transferred to nitrocellulose membrane. The
nitrocellulose membrane was placed on the top of gel and sandwiched between Whatman
papers on both sides. The blotting was done for 70 minutes at 150V using ice inorder to
avoid excessive heating. Once the transfer was complete, the membrane was blocked in a 5%
blocking buffer for 1 hour. Primary antibody CD81 (Santa Cruz Biotechnology, US) was
added and the membrane was left in a cold room in a shaker overnight. The membrane was
washed with 1xPBST three times 5 minutes each and secondary antibody anti-mouse (Dako,
Denmark) was added and incubated in a shaker for 1 hour. This was followed by another
wash with 1xPBST three times 5 minutes each. Detection reagent was added to the
membrane and incubated for 1 minute and finally imaging was done on the imager.
Page 25
19
Blocking buffer
2.5-gram casein milk was dissolved in 50 ml 1x PBST.
3.6 Cell migration assay
Approximately 30000 cells per well were cultured in a 96 well image lock plate. The plate
was incubated overnight and next day, the wells were more than 80 percent confluent to carry
out experiment. The wells were then treated with mitomycin (1:10 dilution) and incubated for
3 hours at +37°C.
Scratching on the cell plate was done carefully by using wound maker (Essen Bioscience).
The media was removed and the cells were washed with 1xPBS once. After adding new
media, the cells were treated with hypoxia and normoxia exosomes (1µl per well) and
incubated in +37°C incubator. The imaging was done by using incucyte zoom system (Essen
Bioscience, US) for the next 24 hours.
3.7 Cell proliferation assay
Approximately 3000 cells were cultured in a 96 well plate. The cells were 80-90% confluent
after growing for 24 hours. Next day, the old media was removed and new media was added.
The cells were then treated with hypoxia and normoxia exosomes. Sytox Green (1:20
dilution) was added in all wells and incubated at +37°C. The imaging was done by incucyte
zoom (Essen Bioscience US) for next 24 hours.
Page 26
20
3.8 Mass spectrometry
The concentration of protein sample was measured by BCA assay. Specific amount of
protein solution was taken and precipitated by adding 3 aliquots of cold acetone and mixed
by vortexing. The mixture was incubated at -20°C overnight.
The sample was centrifuged at 12,000g for 20 minutes at 4°C. The supernatant was discarded
and the pellet was air dried for 3 minutes. A double volume of Tris buffer (50mM, pH 8.5)
with trypsin at an enzyme: substrate ratio of 1:30 (w/w) was added, vortexed and incubated at
37°C for 3 hours. The sample was then reduced with 25mM DTT at 95°C for 5 minutes and
then alkylated with 100mM iodoacetamide at 37°C for 30 minutes in the dark place. This
was followed by addition of second aliquot of trypsin at an enzyme: substrate ratio of 1:25
(w/w) and incubated at 37°C overnight. This allowed for the complete digestion of the
protein sample.
The tubes were collected from incubator and centrifuged at 10,000g for 5 minutes. The
supernatant was collected in new tube and equal volume of 100% acetonitrile was added. The
mixture was transferred to glass vials avoiding bubbles at the bottom and dried using
speedvac (Thermo Scientific, US). The sample was then processed for mass spectrometry.
3.9 Immunohistochemistry
The section slides were taken out from -70°C freezer and thaw them at room temperature for
15 minutes. The samples were blocked by adding 500µl blocking solution at room
temperature for 1 hour. The samples were then incubated by primary antibody; anti-GFP
(abcam ab 290) at 4°C overnight.
Next day, the samples were taken out from 4°C and allowed to stay at room temperature for 1
hour. Samples were washed with 1x PBS for 6 times 15 minutes each. Secondary antibody;
AF goat-anti-rabbit 546 along with Hoechst was added to the samples and incubated for 1
hour at room temperature. Since secondary antibody is light sensitive, this step was carried
Page 27
21
out in dark place. The samples were then washed with 1x PBS for 3 times for 15 minutes and
finally washed with water.
Blocking solution
5% gat serum 2.5ml
5% FBS 2.5ml
1% BSA 0.5ml
0.1% Triton-X 50µl
Dissolved in 0.3M glycine PBS
Primary antibody (1:500)
3µl anti-GFP dissolved in 1500 µl 1x PBS.
Secondary antibody (1:1000)
2.5µl AF goat-anti-rabbit 546 and 1.25µl Hoechst (1:2000) dissolved in 2.5ml 1x PBS.
3.10 RNA extraction
TRIZOL was added in RNAseA treated exosome and mixed. Chloroform was then added to
the sample, mixed by inverting 30 times. The mixture was left for 5 minutes at room
temperature and span for 15 minutes at 10,000 rpm and 4°C. The aqueous phase was
transferred to the tube containing isopropanol, mixed by vortexing and kept at -80°C for 2
hours. The sample was thaw span for 15 minutes at 13000 rpm 4°C, supernatant was
removed and 1ml 75% ethanol was added to the pellet. After brief vortexing, the sample was
span for 5 minutes at 1000 rpm 4°C, supernatant was discarded and the tube was span for a
few minutes at 12000 rpm at room temperature. The rest of the liquid was removed carefully
and tube was left to dry for few minutes and RNAse free H2O was added and vortexed. After
5 minutes incubation on ice, concentration was measured. DNAseI buffer and DNAseI was
added to the sample and incubated for 10 minutes at room temperature. Trizol and
chloroform were immediately added and mixed by inverting. The mixture was left at room
temperature for 5 minutes and then centrifuged for 15 minutes at 10,000 rpm 4°C. The
Page 28
22
aqueous phase was transferred to the tube with isopropanol, mixed by vortexing and
incubated overnight at -80°C.
Next day, the sample was centrifuged for 15 minutes at 13000 rpm. The supernatant was
removed and 75% ethanol was added, span for 5 minutes at 10000 rpm and supernatant was
removed. The sample was further span for 2 minutes at 12000 rpm at room temperature and
rest of the supernatant was removed. The sample was left to dry for few minutes and RNAse
free water was added, vortexed and put in ice. After 10 minutes concentration of RNA was
measured and precipitated again with RNAse free glycogen, RNAse free sodium acetate and
100% ethanol. The mixture was vortexed, incubated at -80°C for 1 hour. After spinning at
13000 rpm for 15 minutes, the sample was washed with 75% ethanol.
The samples were processed by Next Generation Sequencing (NGS), Ion Torrent in
Biocenter Oulu Sequencing Center and results were analyzed.
3.11 Electron microscopy
Former coated copper grids (400 mesh) were glow-discharged before samples were applied.
Sample solution was mixed gently and 3µl droplet was used for each grid and incubated for
20 minutes. The grid was washed 2 times for 1 minute each with a drop of PBS and for 10
minutes in 1% BSA in PBS. The grid was incubated on a drop of primary antibody for 20
minutes, washed 3 times for 1 minute each in a drop of 1% BSA in PBS and incubated on
another drop of secondary antibody for 20 minutes. The grid was washed 3 times for 1
minute each in a drop of 1% BSA in PBS, incubated on a drop of gold-conjugated protein A
for 20 minutes and washed 3 times for 1 minute in a drop of PBS. Sample was fixed by
incubating grid on a drop of 1% glutaraldehyde in PBS for 5 minutes, washed 8 times 1
minute each in a drop of water and then stained with 2% UA in H2O for 5 minutes inside the
fume hood. Finally, the grid was coated with 2% methylcellulose-UA (0.4%) solution for 10
minutes and dried at room temperature.
Page 29
23
3.12 Injection of chicken embryos: ex ovo culture
Fertilized Hy-line chicken eggs were obtained from Haaviston farm (Finland). The eggs were
incubated in a rotary thermostat at 37.5°C for 2 days. At day 3, egg shell was cut carefully
using sterile scissors and the entire egg content was transferred to petri dish with water
covered with plastic.
Renca YFP cells were trypsinized and centrifuged at low speed for 5 minutes. The pellet was
dissolved in 1x PBS and around 1 million cells were injected into embryo. The injections
were intracardiac and were done in two different conditions; some embryos were injected
with only renca YFP cells and others were injected with renca YFP cells with exosomes. The
embryos were then incubated in incubator at +37°C. They were allowed to grow for 7-10
days and images were taken.
Page 30
24
4. RESULTS
The aim of this thesis was to study the function of kidney cancer derived exosomes. Therefore,
the project includes characterization of these exosomes using multiple methods, their effects on
normal mesenchyme cells and role in tumor enhancing. In the last part of the thesis, we analysed
proteins and RNA contents of the exosomes.
4.1 Characterization of EVs
4.1.1 Electron Microscope analysis
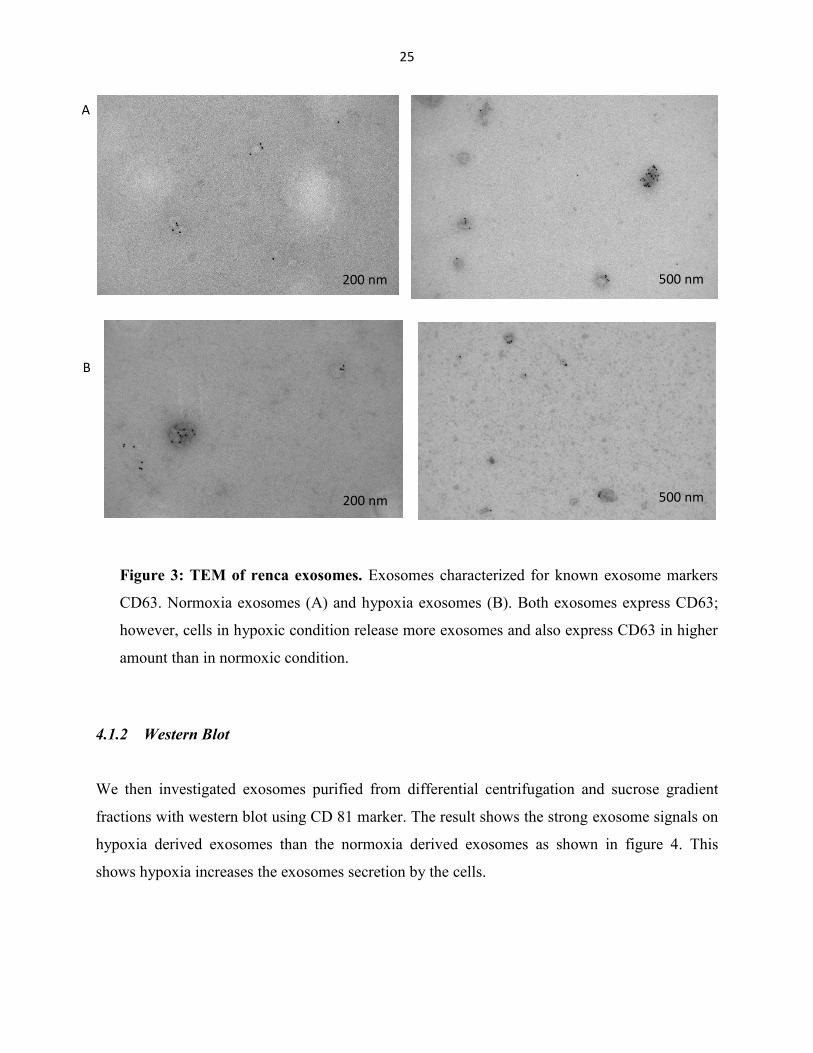
We first investigated the purity of isolated exosomes by transmission electron microscopy
(TEM). The exosomes were immunogold labelled with CD63 antibody and negative stained with
uranyl acetate. Images were taken with Tencai, TEM. We found that majority of vesicles were
labeled with CD63 (fig. 3).
Page 31
25
Figure 3: TEM of renca exosomes. Exosomes characterized for known exosome markers
CD63. Normoxia exosomes (A) and hypoxia exosomes (B). Both exosomes express CD63;
however, cells in hypoxic condition release more exosomes and also express CD63 in higher
amount than in normoxic condition.
4.1.2 Western Blot
We then investigated exosomes purified from differential centrifugation and sucrose gradient
fractions with western blot using CD 81 marker. The result shows the strong exosome signals on
hypoxia derived exosomes than the normoxia derived exosomes as shown in figure 4. This
shows hypoxia increases the exosomes secretion by the cells.
200 nm 500 nm
A
B
200 nm 500 nm
500 nm
500 nm
Page 32
26
Figure 4: Western blot analysis of tumor derived exosomes. A) Vesicles from differential
centrifugation were tested with CD81 marker. The signal was seen in exosome fractions but no
signals were seen in oncosomes and microvesicles fractions. B) Exosomes from hypoxic
condition show strong signal when treated with CD81 marker. This signal is higher than the
normoxia derived exosomes. C) Normoxia derived exosomes have signals when treated with
CD81 marker. The signal shows less secretion of exosomes than hypoxic condition.
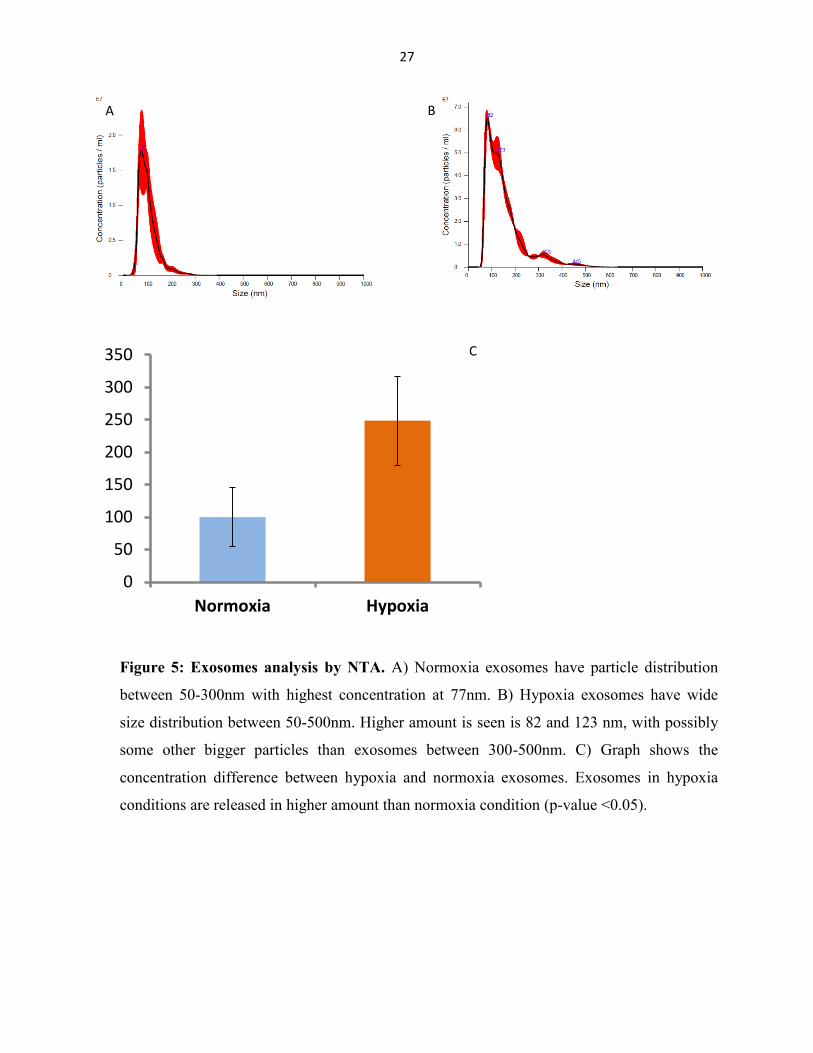
4.1.3 Nanoparticle tracking analysis (NTA)
The size and concentration of exosomes was measured using nanoparticle tracking device
(Malvern Instruments, Malvern, UK). No significant difference in average values was seen
between hypoxia and normoxia exosomes particle sizes. The size distribution in hypoxia sample
is wide that shows the presence of particles with the size above 100 nm. The NTA result also
shows the amount of exosomes is higher in hypoxia than the normoxia condition (fig. 5).
A
B
C
Page 33
27
Figure 5: Exosomes analysis by NTA. A) Normoxia exosomes have particle distribution
between 50-300nm with highest concentration at 77nm. B) Hypoxia exosomes have wide
size distribution between 50-500nm. Higher amount is seen is 82 and 123 nm, with possibly
some other bigger particles than exosomes between 300-500nm. C) Graph shows the
concentration difference between hypoxia and normoxia exosomes. Exosomes in hypoxia
conditions are released in higher amount than normoxia condition (p-value <0.05).
0
50
100
150
200
250
300
350
Normoxia Hypoxia
C
B A
Page 34
28
4.2 Effects of exosomes
4.2.1 Exosome labeling
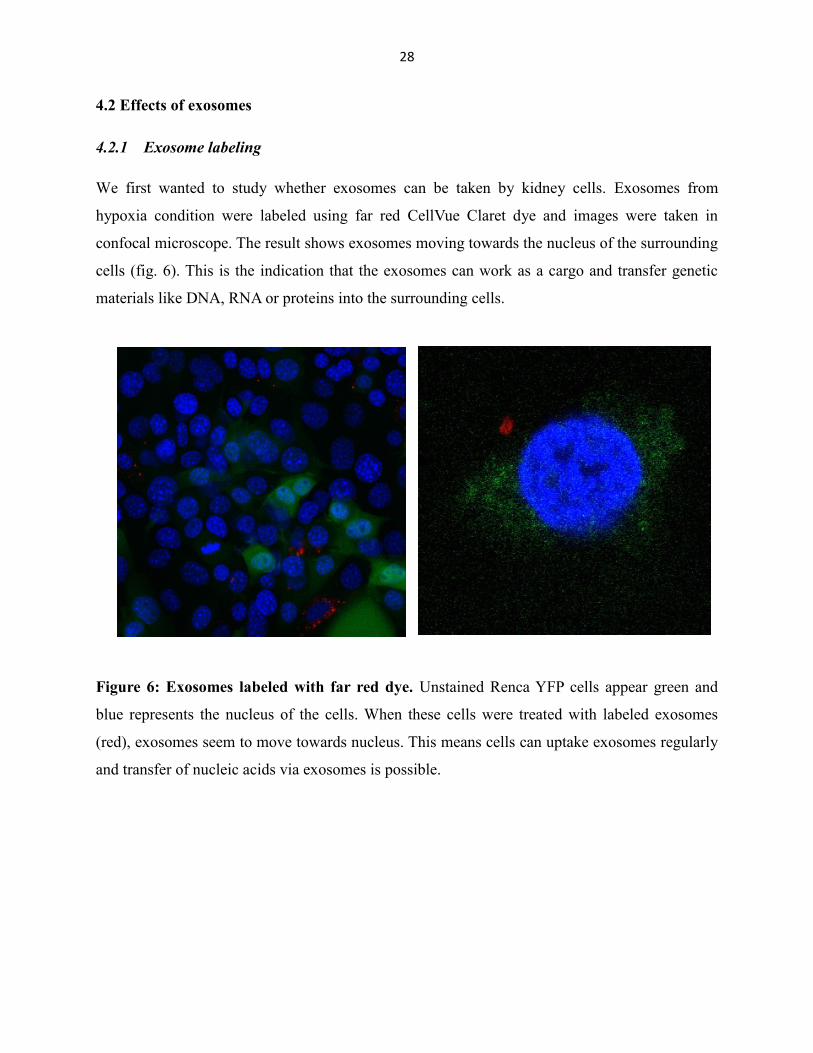
We first wanted to study whether exosomes can be taken by kidney cells. Exosomes from
hypoxia condition were labeled using far red CellVue Claret dye and images were taken in
confocal microscope. The result shows exosomes moving towards the nucleus of the surrounding
cells (fig. 6). This is the indication that the exosomes can work as a cargo and transfer genetic
materials like DNA, RNA or proteins into the surrounding cells.
Figure 6: Exosomes labeled with far red dye. Unstained Renca YFP cells appear green and
blue represents the nucleus of the cells. When these cells were treated with labeled exosomes
(red), exosomes seem to move towards nucleus. This means cells can uptake exosomes regularly
and transfer of nucleic acids via exosomes is possible.
Page 35
29
4.2.2 Cell proliferation and motility assay
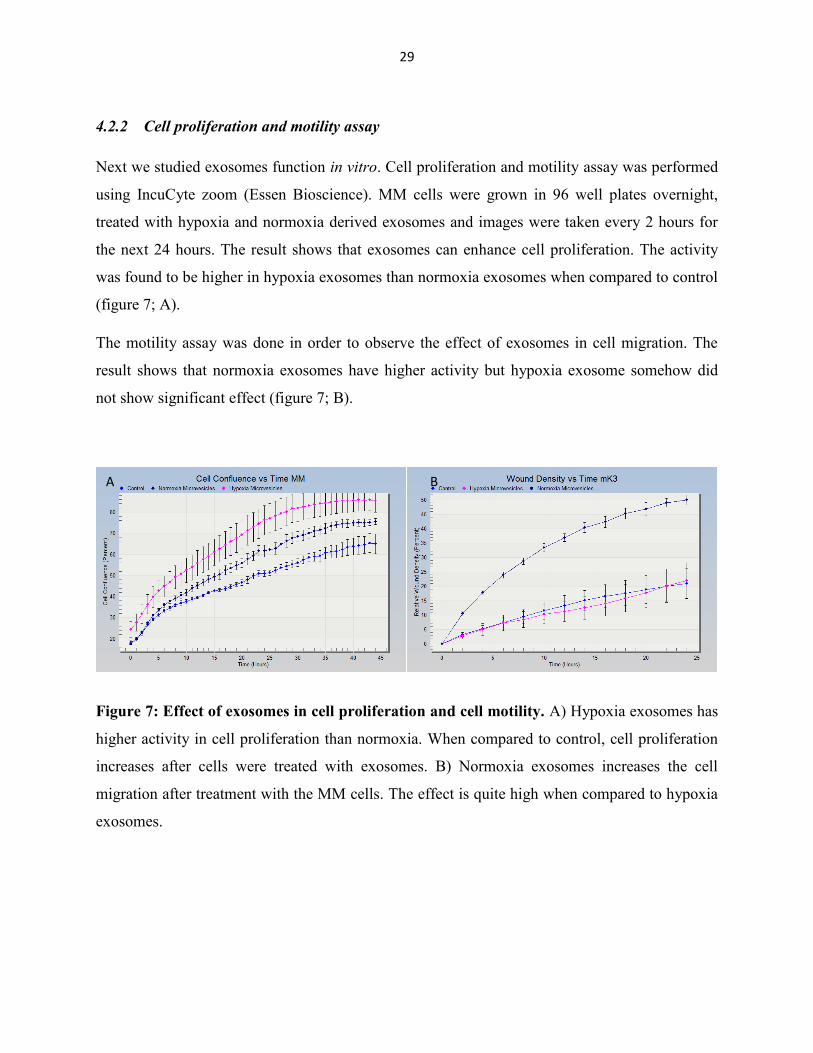
Next we studied exosomes function in vitro. Cell proliferation and motility assay was performed
using IncuCyte zoom (Essen Bioscience). MM cells were grown in 96 well plates overnight,
treated with hypoxia and normoxia derived exosomes and images were taken every 2 hours for
the next 24 hours. The result shows that exosomes can enhance cell proliferation. The activity
was found to be higher in hypoxia exosomes than normoxia exosomes when compared to control
(figure 7; A).
The motility assay was done in order to observe the effect of exosomes in cell migration. The
result shows that normoxia exosomes have higher activity but hypoxia exosome somehow did
not show significant effect (figure 7; B).
Figure 7: Effect of exosomes in cell proliferation and cell motility. A) Hypoxia exosomes has
higher activity in cell proliferation than normoxia. When compared to control, cell proliferation
increases after cells were treated with exosomes. B) Normoxia exosomes increases the cell
migration after treatment with the MM cells. The effect is quite high when compared to hypoxia
exosomes.
A B
Page 36
30
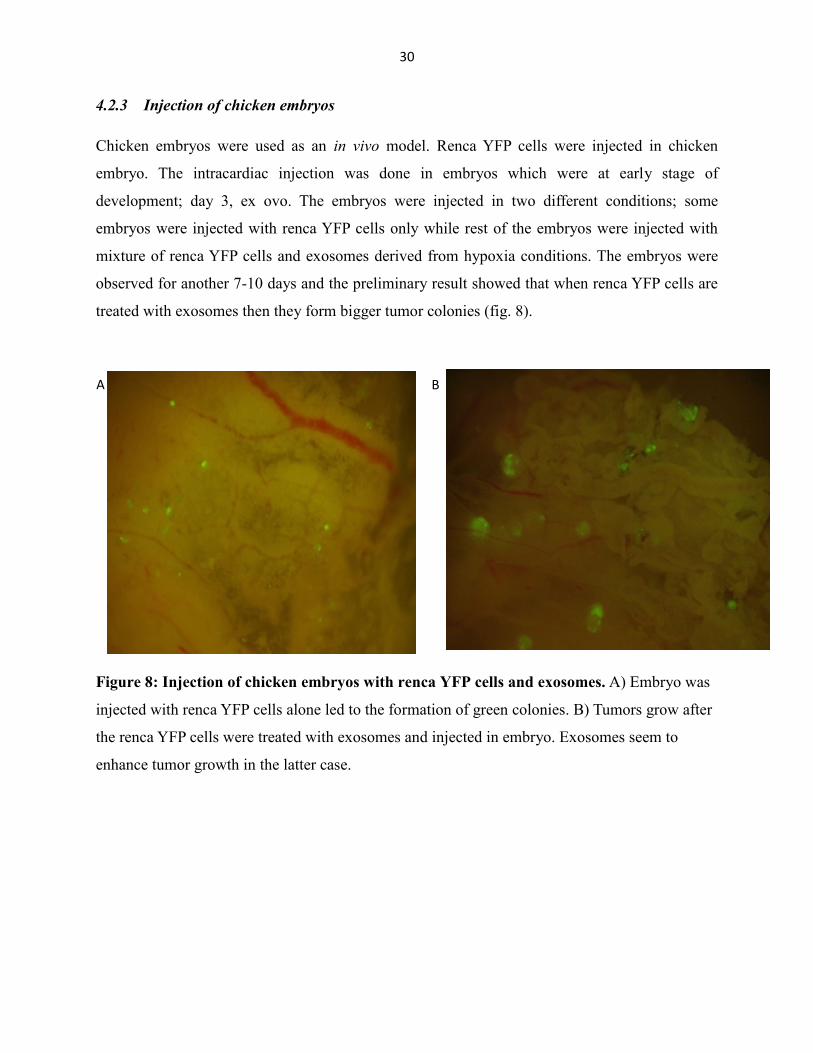
4.2.3 Injection of chicken embryos
Chicken embryos were used as an in vivo model. Renca YFP cells were injected in chicken
embryo. The intracardiac injection was done in embryos which were at early stage of
development; day 3, ex ovo. The embryos were injected in two different conditions; some
embryos were injected with renca YFP cells only while rest of the embryos were injected with
mixture of renca YFP cells and exosomes derived from hypoxia conditions. The embryos were
observed for another 7-10 days and the preliminary result showed that when renca YFP cells are
treated with exosomes then they form bigger tumor colonies (fig. 8).
Figure 8: Injection of chicken embryos with renca YFP cells and exosomes. A) Embryo was
injected with renca YFP cells alone led to the formation of green colonies. B) Tumors grow after
the renca YFP cells were treated with exosomes and injected in embryo. Exosomes seem to
enhance tumor growth in the latter case.
B A
100
Page 37
31
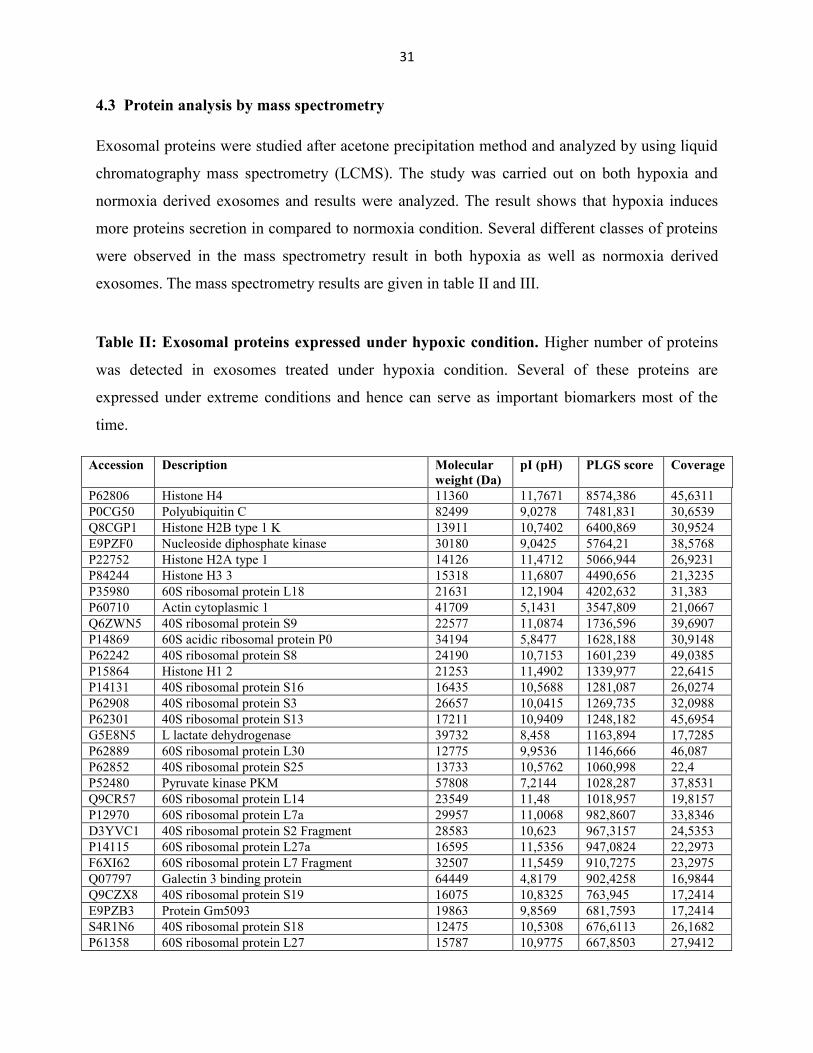
4.3 Protein analysis by mass spectrometry
Exosomal proteins were studied after acetone precipitation method and analyzed by using liquid
chromatography mass spectrometry (LCMS). The study was carried out on both hypoxia and
normoxia derived exosomes and results were analyzed. The result shows that hypoxia induces
more proteins secretion in compared to normoxia condition. Several different classes of proteins
were observed in the mass spectrometry result in both hypoxia as well as normoxia derived
exosomes. The mass spectrometry results are given in table II and III.
Table II: Exosomal proteins expressed under hypoxic condition. Higher number of proteins
was detected in exosomes treated under hypoxia condition. Several of these proteins are
expressed under extreme conditions and hence can serve as important biomarkers most of the
time.
Accession Description Molecular
weight (Da)
pI (pH) PLGS score Coverage
P62806 Histone H4 11360 11,7671 8574,386 45,6311
P0CG50 Polyubiquitin C 82499 9,0278 7481,831 30,6539
Q8CGP1 Histone H2B type 1 K 13911 10,7402 6400,869 30,9524
E9PZF0 Nucleoside diphosphate kinase 30180 9,0425 5764,21 38,5768
P22752 Histone H2A type 1 14126 11,4712 5066,944 26,9231
P84244 Histone H3 3 15318 11,6807 4490,656 21,3235
P35980 60S ribosomal protein L18 21631 12,1904 4202,632 31,383
P60710 Actin cytoplasmic 1 41709 5,1431 3547,809 21,0667
Q6ZWN5 40S ribosomal protein S9 22577 11,0874 1736,596 39,6907
P14869 60S acidic ribosomal protein P0 34194 5,8477 1628,188 30,9148
P62242 40S ribosomal protein S8 24190 10,7153 1601,239 49,0385
P15864 Histone H1 2 21253 11,4902 1339,977 22,6415
P14131 40S ribosomal protein S16 16435 10,5688 1281,087 26,0274
P62908 40S ribosomal protein S3 26657 10,0415 1269,735 32,0988
P62301 40S ribosomal protein S13 17211 10,9409 1248,182 45,6954
G5E8N5 L lactate dehydrogenase 39732 8,458 1163,894 17,7285
P62889 60S ribosomal protein L30 12775 9,9536 1146,666 46,087
P62852 40S ribosomal protein S25 13733 10,5762 1060,998 22,4
P52480 Pyruvate kinase PKM 57808 7,2144 1028,287 37,8531
Q9CR57 60S ribosomal protein L14 23549 11,48 1018,957 19,8157
P12970 60S ribosomal protein L7a 29957 11,0068 982,8607 33,8346
D3YVC1 40S ribosomal protein S2 Fragment 28583 10,623 967,3157 24,5353
P14115 60S ribosomal protein L27a 16595 11,5356 947,0824 22,2973
F6XI62 60S ribosomal protein L7 Fragment 32507 11,5459 910,7275 23,2975
Q07797 Galectin 3 binding protein 64449 4,8179 902,4258 16,9844
Q9CZX8 40S ribosomal protein S19 16075 10,8325 763,945 17,2414
E9PZB3 Protein Gm5093 19863 9,8569 681,7593 17,2414
S4R1N6 40S ribosomal protein S18 12475 10,5308 676,6113 26,1682
P61358 60S ribosomal protein L27 15787 10,9775 667,8503 27,9412
Page 38
32
P62900 60S ribosomal protein L31 14453 10,9688 605,6777 34,4
Q9D8E6 60S ribosomal protein L4 47124 11,4434 593,623 18,6158
I7HLV2 60S ribosomal protein L10 Fragment 23057 10,4004 586,8488 16,9154
P47911 60S ribosomal protein L6 33488 11,1284 575,1203 17,9054
P05213 Tubulin alpha 1B chain 50119 4,7622 558,2223 23,9468
G5E899 Plasminogen activator inhibitor 1 45015 6,0103 469,793 17,9104
P11499 Heat shock protein HSP 90 beta 83229 4,7754 439,9723 15,4696
P62754 40S ribosomal protein S6 28663 11,2896 432,179 15,261
P47963 60S ribosomal protein L13 24290 11,9531 402,5908 18,484
P62702 40S ribosomal protein S4 X isoform 29579 10,5864 332,8276 19,0114
P35979 60S ribosomal protein L12 17793 9,8965 308,0653 33,3333
P84099 60S ribosomal protein L19 23451 11,8901 298,982 18,3673
E9QAZ2 Ribosomal protein L15 24062 11,9707 293,9146 14,7059
P19253 60S ribosomal protein L13a 23449 11,439 251,4231 18,2266
P62267 40S ribosomal protein S23 15797 10,9585 249,2977 15,3846
P14206 40S ribosomal protein SA 32817 4,6025 226,6607 13,5593
P99027 60S acidic ribosomal protein P2 11643 4,1792 201,3828 23,4783
Q148B6 Speriolin 51385 8,3804 164,7531 12,5
P68040 Guanine nucleotide binding protein subunit beta
2 like 1
35054
7,5645
134,4731
20,5047
E9PZF0 Nucleoside diphosphate kinase 30180 9,0425 3022,229 40,4494
P52480 Pyruvate kinase PKM 57808 7,2144 1109,824 37,2881
Q5SX59 Enolase Fragment 20947 9,3311 866,2366 22,449
F6YJW4 60S ribosomal protein L18a 20464 10,5938 166,6728 17,2414
A2BI14 Coiled coil domain containing protein 171 43195 5,1079 156,5664 24,1848
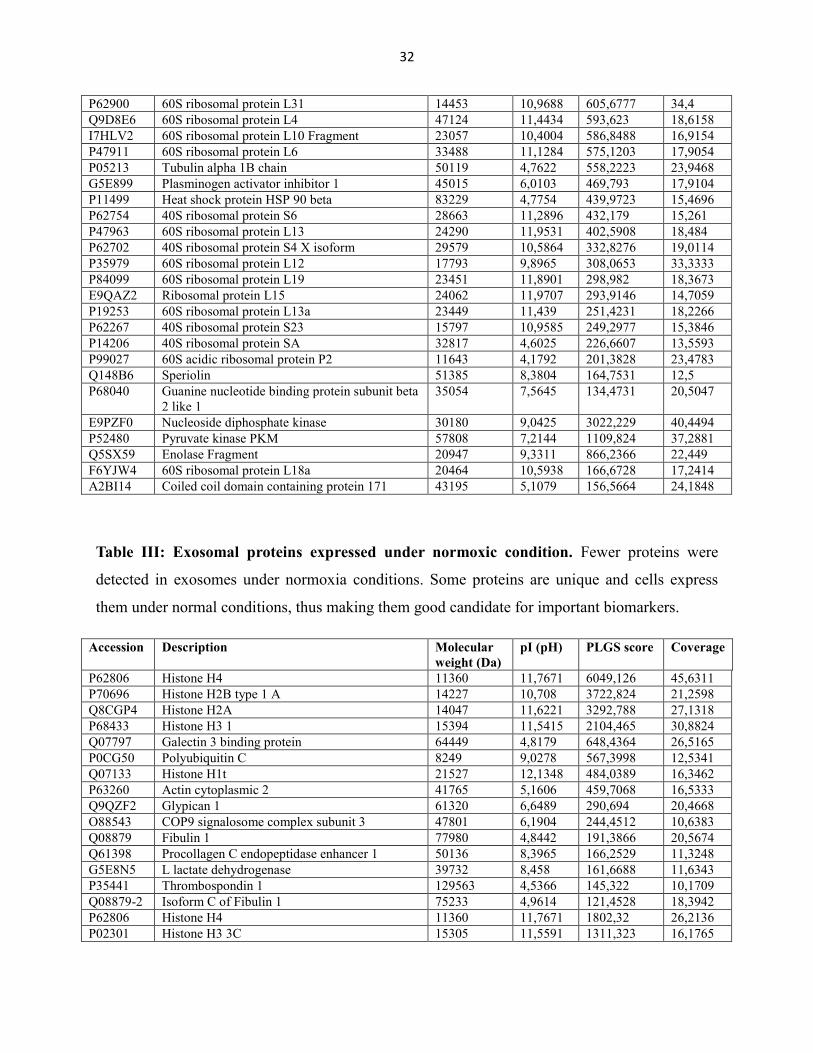
Table III: Exosomal proteins expressed under normoxic condition. Fewer proteins were
detected in exosomes under normoxia conditions. Some proteins are unique and cells express
them under normal conditions, thus making them good candidate for important biomarkers.
Accession Description Molecular
weight (Da)
pI (pH) PLGS score Coverage
P62806 Histone H4 11360 11,7671 6049,126 45,6311
P70696 Histone H2B type 1 A 14227 10,708 3722,824 21,2598
Q8CGP4 Histone H2A 14047 11,6221 3292,788 27,1318
P68433 Histone H3 1 15394 11,5415 2104,465 30,8824
Q07797 Galectin 3 binding protein 64449 4,8179 648,4364 26,5165
P0CG50 Polyubiquitin C 8249 9,0278 567,3998 12,5341
Q07133 Histone H1t 21527 12,1348 484,0389 16,3462
P63260 Actin cytoplasmic 2 41765 5,1606 459,7068 16,5333
Q9QZF2 Glypican 1 61320 6,6489 290,694 20,4668
O88543 COP9 signalosome complex subunit 3 47801 6,1904 244,4512 10,6383
Q08879 Fibulin 1 77980 4,8442 191,3866 20,5674
Q61398 Procollagen C endopeptidase enhancer 1 50136 8,3965 166,2529 11,3248
G5E8N5 L lactate dehydrogenase 39732 8,458 161,6688 11,6343
P35441 Thrombospondin 1 129563 4,5366 145,322 10,1709
Q08879-2 Isoform C of Fibulin 1 75233 4,9614 121,4528 18,3942
P62806 Histone H4 11360 11,7671 1802,32 26,2136
P02301 Histone H3 3C 15305 11,5591 1311,323 16,1765
Page 39
33
P10854 Histone H2B type 1 M 13927 10,7402 652,5275 15,873
P02535-2 Isoform 2 of Keratin type I cytoskeletal 10 57025 4,8164 617,9836 13,369
Q9R0H5 Keratin type II cytoskeletal 71 57346 6,627 470,1037 23,0033
Q7TSY8 Shugoshin like 2 130196 8,9033 61,8569 10,6529
Q07797 Galectin 3 binding protein 64449 4,8179 494,5569 16,5122
D3Z741 Protein 4930444G20Rik 56592 6,2329 266,3577 20,1144
F7CB97 CAP Gly domain containing linker protein 1
Fragment
135913
5,02
196,5467
12,3662
Q06890 Clusterin 51622 5,3247 389,5988 18,9732
P11499 Heat shock protein HSP 90 beta 83229 4,7754 239,0147 14,0884
Q80X71 Transmembrane protein 106B 31152 6,2329 123,0444 13,8182
4.4 RNA analysis
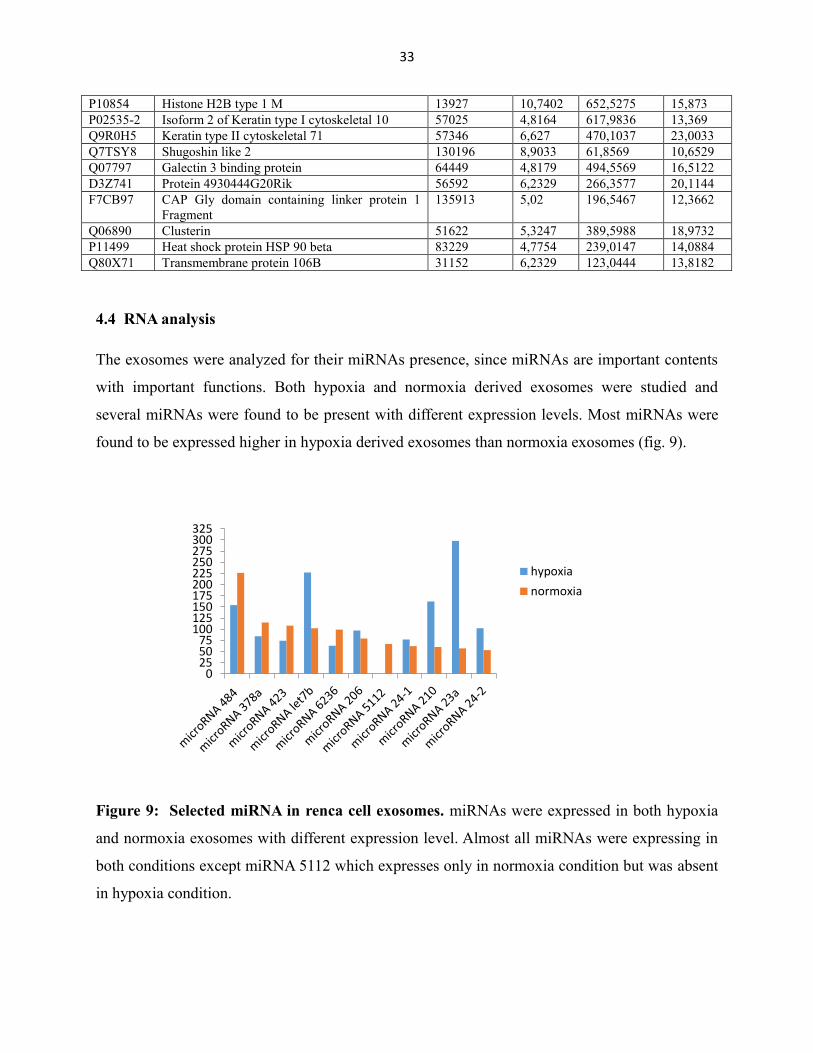
The exosomes were analyzed for their miRNAs presence, since miRNAs are important contents
with important functions. Both hypoxia and normoxia derived exosomes were studied and
several miRNAs were found to be present with different expression levels. Most miRNAs were
found to be expressed higher in hypoxia derived exosomes than normoxia exosomes (fig. 9).
Figure 9: Selected miRNA in renca cell exosomes. miRNAs were expressed in both hypoxia
and normoxia exosomes with different expression level. Almost all miRNAs were expressing in
both conditions except miRNA 5112 which expresses only in normoxia condition but was absent
in hypoxia condition.
0255075
100125150175200225250275300325
hypoxia
normoxia
Page 40
34
5. DISCUSSIONS
Exosomes are extracellular vesicles secreted by almost all type of cells. They are enriched in
biological molecules such as DNA, mRNA, miRNA, proteins and lipids. Hence, the cells use
these vesicles as a means of communication and transfer biological information from parent cells
to the target cells. Such properties make them important players in the field of diseases including
cancer. In this study, tumor derived exosomes from renal carcinoma cells were purified,
characterized and studied. The proteins and RNAs were analyzed, and role of exosomes in tumor
formation was studied.
Currently, there is a lack of standardized purification methods for extracellular vesicles that can
distinguish exosomes from other vesicles. Here, we tried to purify exosomes by two methods:
sequential ultracentrifugation and OptiPrepTM
density gradient ultracentrifugation methods (Van
Deun et al. 2014). Western blot analysis with CD81 marker showed stronger signal in density
gradient purification (fig. 4) than simple ultracentrifugation method (fig. 4 A). However,
nanoparticle tracking analysis (NTA) confirmed the presence of vesicles larger than exosomes
(>100 nm) (fig. 5). Thus getting 100% pure exosomes or any other vesicles with the existing
purification methods is still a big challenge.
To study the effect of exosomes in cell proliferation and migration in recipient cells, MM cells
were either treated with exosomes or with diluent. A significant increase in cell proliferation and
migration was observed after the cells were treated with exosomes (fig. 7). Many studies support
the role of exosomes in tissue repair and positive effects have been seen in cell migration and
proliferation (Zhao et al. 2015). It is assumed that the ability of exosomes to exert such affect is
because of the RNA and protein contents of the exosomes. These biological components are
released into the cells by exosomes resulting in a promotion of cell migration and proliferation.
The in vitro tracking of the labeled exosomes showed that exosomes actually enter into the cells.
The labeled exosomes were uptaken by the cells (fig. 6). This result supports the work of
Ramesh et al. where they treated cells with exosomes of tumor origin and found the exosomes
entering into the cell cytoplasm (Ramesh et al. 2015). Our result shows that cells uptake
exosomes and exosomes release their active biological substances into the cells, promoting
cellular activities such as proliferation, migration and even tumor formation.
Page 41
35
The role of exosomes in intercellular communication has been studied widely. The ability of
exosomes to transfer genetic materials like DNA, RNA and proteins has been found to be
associated with tumor propagation (Minciachhi et al. 2015). As an in vivo experiment, the
chicken embryos were injected with YFP labelled cancer cells either treated with exosomes or
Phosphate Buffered Saline (PBS). Significant tumor colonies growth was observed when the
embryos were injected with tumor derived exosomes (fig. 8). This provides evidence that
exosomes are key player in the tumor propagation. This result supports the work done by Du et
al. who investigated the effect of exosomes in renal carcinoma cells. The human Wharton´s jelly
mesenchyme stem cells derived exosomes stimulated tumorigenesis as well as growth in the
tumor size (Du et al. 2014).
Exosomes are rich in protein contents and in many cellular processes, exosomes can transfer
these protein cargos to target cells thus affecting cellular environment. The transfer of these
protein cargos via exosomes shows the potential role of exosomes in diseases including cancer
(Boukouris & Mathivanan 2015). In this study, we analyzed exosomal proteins from tumor
derived cells treated under hypoxia and normoxia conditions. Diverse range of proteins was
found in exosomes derived from both conditions; however, more proteins were detected in
hypoxia derived exosomes (table II) than in the normal condition (table III). The result is
comparable with the previous findings as many of these proteins have been reported in ExoCarta
protein database. However, many ribosomal proteins are also found in this study. This may be
due to the presence of apoptotic bodies in significant amount. During purification, apoptotic
bodies might have mixed with exosomes.
miRNAs refer to the small non-coding RNAs which play important roles in different biological
processes. One of the aims of our study was to investigate the miRNAs present in Renca
exosomes. Hypoxia and normoxia exosomes were analyzed and result showed the presence of
different miRNAs (fig. 9). Many of these miRNA have been detected in exosomes previously
and our result supports them. One of the miRNA we found in our study is let7b, which has been
described as one of the first miRNAs discovered in mammals and involves in the regulation of
differentiation and proliferation (Peter 2009). MiR-210 is expressed higher in hypoxia exosomes
which is relevant as miR-210 is known to be associated with HIF molecules and hypoxia
(McCormick et al. 2013). We found that miR-5112 is expressed in normoxia condition but is
Page 42
36
completely unexpressed in hypoxia condition. The cells might loss expression of this miRNA
under stressed conditions like hypoxia. This can act as RNA biomarker and can be used in
diagnostic and prognostic process.
Page 43
37
6. CONCLUSIONS
We investigated the role of exosomes in kidney in this study. Exosomes are released constantly
by cells and can be extracted from different body fluids and cell culture supernatants. The lack of
standard purification methods and suitable biomarkers that can separate exosomes from other
extracellular vesicles is still a problem and challenge for the researchers in this field.
The in vitro study showed that exosomes have effects in kidney cells. Metanephric mesenchyme
(MM) cells when treated with exosomes enhanced cell proliferation and motility activity when
compared to control. Hypoxia increases the release of exosomes as compared to the normal
condition.
The in vivo study of chicken model suggested the ability of exosomes to enhance tumor growth.
This suggests the role of exosomes in intercellular communication. The exosomes can transfer
their biological materials into the recipient cells and change microenvironment. This proves
exosomes can enhance tumor growth.
The proteins and RNA analysis of exosomes show the presence of many important proteins and
miRNAs. These exosomal proteins and miRNAs might have capacity to alter biological
functions.
In summary, we were able to purify and characterize exosomes from kidney cancer cell line,
show effect of exosomes in kidney both in vitro and in vivo.
Page 44
38
7. REFERENCES
Abreu SC, Weiss DJ & Rocco PRM (2016) Extracellular vesicles derived from mesenchymal
stromal cells: a therapeutic option in respiratory diseases? Stem Cell Research & Therapy 7: 53.
Andaloussi SEL, Mager I, Breakfield XO & Wood MJA (2013) Extracellular vesicles: biology
and emerging therapeutic opportunities. Nature reviews drug discovery 12: 347-357.
Azmi AS, Bao B & Sarkar FH (2013) Exosomes in cancer development, metastasis, and drug
resistance: a comprehensive review. Cancer Metastasis Rev 32: 623–642.
Barteneva NS, Fasler-Kan E, Bernimoulin M, Stern JN, Ponomarev ED & Duckett L (2013)
Circulating microparticles: square the circle. BMC Cell Biol 14: 23.
Beauvillain C, Juste MO & Dion S (2009) Exosomes are an effective vaccine against congenital
toxoplasmosis in mice. Vaccine 27: 1750–1757.
Belting M & Christianson HC (2015) Role of Exosomes and Microvesicles in Hypoxia-
associated Tumour Development and Cardiovascular Disease. J Intern Med 3: 251-63.
Boukouris S & Mathivanan S (2015) Exosomes in bodily fluids are a highly stable resource of
disease biomarkers. Proteomics Clin Appl 9; 358-367.
Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V & Tetta C (2011)
Exosome/microvesicle-mediated epigenetic reprogramming of cells. American Journal of Cancer
Research 1: 98–110.
Chargaff E & West R (1946) The biological significance of the thromboplastic protein of blood.
J Biol Chem 166: 189-97.
Chaturvedi P, Gilkes DM, Wong CC, Luo W, Zhang H & Wei H (2013) Hypoxia-inducible
factor-dependent breast cancer-mesenchymal stem cell bidirectional signaling promotes
metastasis. The Journal of Clinical Investigation 123: 189–205.
Page 45
39
Chen TS, Lai RC, Lee MM, Choo AB, Lee CN & Lim SK (2010) Mesenchymal stem cell
secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res 38: 215–224.
Cho JA, Park H, Lim EH & Lee KW (2012) Exosomes from breast cancer cells can convert
adipose tissue-derived mesenchymal stem cells into myofibroblast-like cells. Int J Oncol 40:
130–138.
Denzer K, Kleijmeer MJ, Heijnen HF, Stoorvogel W & Geuze HJ (2000) Exosome: from
internal vesicle of the multivesicular body to intercellular signaling device. J. Cell Sci. 113:3365.
Deregibus MC, Cantaluppi V & Calogero R (2007) Endothelial progenitor cell derived
microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of
mRNA. Blood 110: 2440–2448.
Du TJG, Wu S, Cheng Z, Cheng J & Zou X (2014). Microvesicles derived from human
Wharton's jelly mesenchymal stem cells promote human renal cancer cell growth and
aggressiveness through induction of hepatocyte growth factor. PLoS ONE 9: e96836.
Elmore S (2007) Apoptosis: a review of programmed cell death. Toxicologic pathology 35: 495-
516.
Fais S, Logozzi M, Lugini L, Federici C, Azzarito T & Zarovni N (2013) Exosomes: the ideal
nanovectors for biodelivery. Biol Chem 394: 1-15.
Gabriel K, Ingram A, Austin R, Kapoor A, Tang D & Majeed F (2013) Regulation of the tumor
suppressor PTEN through exosomes: a diagnostic potential for prostate cancer. PLoS One 8:
70047.
Gatti S, Bruno S & Deregibus MC (2011) Microvesicles derived from human adult mesenchymal
stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury.
Nephrol Dial Transplant 26:1474–1483.
Gould SJ & Raposo G (2013) As we wait: coping with an imperfect nomenclature for
extracellular vesicles. J Extracell Vesicles 2: 10.
Page 46
40
Halicka HD, Bedner E & Darzynkiewicz Z (2000) Segregation of RNA and separate packaging
of DNA and RNA in apoptotic bodies during apoptosis. Experimental cell research 260: 248-
256.
Harding C, Heuser J & Stahl P (1983) Receptor-mediated endocytosis of transferrin and
recycling of the transferrin receptor in rat reticulocytes. J Cell Biol 97: 329–39.
Hemler ME (2003) Tetraspanin proteins mediate cellular penetration, invasion, and fusion events
and define a novel type of membrane microdomain. Annu Rev Cell Dev Biol 19: 397–422.
Henderson MC & Azorsa DO (2012) The genomic and proteomic content of cancer cell-derived
exosomes. Front Oncol 2: 38.
Hongjuan J, Guan G, Zhang R, Liu G, Cheng J, Hou X & Cui Y (2009) Identification of Urinary
Soluble E-cadherin as a Novel Biomarker for Diabetic Nephropathy. Diabetes/Metabolism
Research and Reviews Diabetes Metab. Res. Rev 25.3: 232-41.
Hristov M, Erl W, Linder S & Weber PC (2004) Apoptotic bodies from endothelial cells enhance
the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood
104: 2761-2766.
Kaelin WG Jr & Ratcliffe PJ (2008) Oxygen sensing by metazoans: the central role of the HIF
hydroxylase pathway. Mol Cell 30: 393–402.
Kalra H, Drummen GPC & Mathivanan S (2016) Focus on extracellular vesicles: introducting
the next small big thing. Int J Mol Sci 17(2): 170.
King HW, Michael MZ & Gleadle JM (2012) Hypoxic enhancement of exosome release by
breast cancer cells. BMC Cancer 12: 421.
Koga Y, Yasunaga M, Moriya Y, Akasu T, Fujita S & Yamamoto S (2011) Exosome can prevent
RNase from degrading microRNA in feces. J Gastrointest Oncol 2: 215–222.
Kuate S, Cinatl J & Doerr HW (2007) Exosomal vaccines containing the S protein of the SARS
coronavirus induce high levels of neutralizing antibodies. Virology 362: 26–37.
Page 47
41
Kunter U, Rong S & Djuric Z (2006) Transplanted mesenchymal stem cells accelerate
glomerular healing in experimental glomerulonephritis. J Am Soc Nephrol 17: 2202–2212.
Lee TH, D’Asti E, Magnus N, Al-Nedawi K, Meehan B & Rak J (2011) Microvesicles as
mediators of intercellular communication in cancer- the emerging science of cellular ‘debris’.
Semin Immunopathol 33: 455–467.
Ling XB, Sigdel TK, Lau K, Ying L, Lau I, Schilling J & Sarwal MM (2010) Integrative Urinary
Peptidomics in Renal Transplantation Identifies Biomarkers for Acute Rejection. Journal of the
American Society of Nephrology 21.4: 646-53.
Logozzi M, De Milito A, Lugini L, Borghi M, Calabrò L, Spada M, Perdicchio M, Marino ML,
Federici C, Iessi E, Brambilla D, Venturi G, Lozupone F, Santinami M, Huber V, Maio M,
Rivoltini L & Fais S (2009) High levels of exosomes expressing CD63 and caveolin-1 in plasma
of melanoma patients. PLoS One 4: 5219.
Mallat Z, Hugel B, Ohan J, Leseche G, Freyssinet JM & Tedgui A (1999) Shed membrane
microparticles with procoagulant potential in human atherosclerotic plaques: A role for apoptosis
in plaque thrombogenicity. Circulation 99:348–353.
Marcos-Gragera R, Mallone S, Kiemeney LA, Vilardell L, Malats N, Allory Y, Sant M &
Eurocare-5 working group (2015) Urinary tract cancer survival in Europe 1999-2007: Results of
the population-based study ERUOCARE-5. European Journal of Cancer 51; 2217-2230.
Martin SJ, Reutelingsperger CP, McGahon AJ, Rader JA, van Schie RC, LaFace DM & Green
DR (1995) Early redistribution of plasma membrane phosphatidylserine is a general feature of
apoptosis regardless of the initiating stimulus: inhibition by overexpression of Bcl-2 and Abl.
The Journal of experimental medicine 182: 1545-1556.
Mathivanan S & Simpson RJ (2009) ExoCarta: A compendium of exosomal proteins and RNA.
Proteomics 9: 4997–5000.
Mathivanan S, Fahner CJ, Reid GE & Simpson RJ (2012) ExoCarta 2012: database of exosomal
proteins, RNA and lipids. Nucleic Acids Res 40: 1241–1244.
Page 48
42
McCormick RI, Blick C, Ragoussis J, Schoedel J, Mole DR & Young AC (2013) miR-210 is a
target of hypoxia-inducible factors 1 and 2 in renal caner, regulates ISCU and correlates with
good prognosis. Br J Cancer 108; 1133-1142.
Miranda KC, Bond DT, McKee M, Skog J, Paunescu TG & Da Slva N (2010) Nucleic acids
within urinary exosomes/microvesicles are potential biomarkers for renal disease. Kidney Int
78:191–199.
Mytar B, Baj-Krzyworzeka M, Majka M., Stankiewicz D & Zembala M (2008) Human
monocytes both enhance and inhibit the growth of human pancreatic cancer in SCID mice.
Anticancer Res 28: 187-192.
National Comprehensive Cancer Network (2009). NCCN Clinical Practice Guidelines in
Oncology: Kidney Cancer. URI:
http://www.nccn.org/professionals/physician_gls/f_guidelines.asp. Citated: 2009/6.
Nawaz M, Camussi G & H. Valadi (2014) The emerging role of extracellular vesicles as
biomarkers for urogenital cancers, Nature Reviews Urology 12: 688–701.
Nguyen DX & Massague J (2007) Genetic determinants of cancer metastasis. Nat Rev Genet 8:
341–352.
Nguyen DX, Bos PD & Massague J (2009) Metastasis: from dissemination to organ-specific
colonization. Nat Rev Cancer 9: 274–284.
Nieto MA (2011) The ins and outs of the epithelial to mesenchymal transition in health and
disease. Annu Rev Cell Dev Biol 27: 347–376.
Paidpally V, Chirindel A, Lam S, Agrawal N, Quon H & Subramaniam RM (2012) FDG-
PET/CT imaging biomarkers in head and neck squamous cell carcinoma. Imaging Med 4: 633–
647.
Park JE, Tan HS, Datta A, Lai RC, Zhang H & Meng W (2010) Hypoxic tumor cell modulates
its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and
exosomes. Molecular & Cellular Proteomics 9: 1085–1099.
Page 49
43
Peter ME (2009) Let-7 and miR-200 microRNAs: guardians against pluripotency and cancer
propagation. Cell Cycle 8; 843-852.
Piccin A, Murphy WG & Smith OP (2007) Circulating microparticles: pathophysiology and
clinical implications. Blood Rev 21: 157–71.
Rabinowits G, Gercel-Taylor C & Day JM (2009) Exosomal microRNA: a diagnostic marker for
lung cancer. Clin Lung Cancer 10: 42–46.
Raposo G, Nijman HW, Stoorvogel W, Leijendekker R, Harding CV & Geuze HJ (1996) B
lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 183:1.
Ratajczak J, Miekus K, Kucia M, Zhang J, Reca R, Dvorak P & Ratajczak MZ (2006)
Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for
horizontal transfer of mRNA and protein delivery. Leukemia 20: 847–56.
Shrivastava A, Filant J, Moxley KM, Sood A, McMeeking S & Ramesh R (2015) Exosomes: a
role for naturally occurring nanovesicles in cancer growth, diagnosis and treatment. Current gene
therapy 15; 182-192.
Simpson R & Mathivanan S (2012) Extracellular microvesicles: The need for internationally
recognised nomenclature and stringent purification criteria. J. Proteom. Bioinform 5:2.
Sonoda H, Yokota-Ikeda N, Oshikawa S, Kanno Y, Yoshinaga K, Uchida K, Ueda Y, Kimiya K,
Uezono S, Ueda A, Ito K & Ikeda M (2009) Decreased Abundance of Urinary Exosomal
Aquaporin-1 in Renal Ischemia-reperfusion Injury. AJP: Renal Physiology 297.4.
Steliarova-Foucher E, O’Callaghan M, Ferlay J, Masuyer E, Forman D & Comber H (2014)
European Cancer Observatory: cancer incidence, mortality, prevalence and survival in Europe.
Version 1.0 European Network of Cancer Registries, International Agency for Research on
Cancer.
Subra C, Grand D & Laulagnier K (2010) Exosomes account for vesicle-mediated transcellular
transport of activatable phospholipases and prostaglandins. J Lipid Res 51: 2105–20.
Page 50
44
Taylor DD & Gercel-Taylor C (2008) MicroRNA signatures of tumor-derived exosomes as
diagnostic biomarkers of ovarian cancer. Gynecol Oncol 110: 13–21.
Thery C, Boussac M, Veron P, Ricciardi-Castagnoli P, Raposo G, Garin J & Amigorena S
(2001) Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular
compartment distinct from apoptotic vesicles. Journal of Immunology 166; 7309-7318.
Thery C, Ostrowski M & Segura E (2009) Membrane vesicles as conveyors of immune
responses. Nat Rev Immunol 9: 581-593.
Tian T, Wang Y, Wang H, Zhu Z & Xiao Z (2010) Visualizing of the cellular uptake and
intracellular trafficking of exosomes by live-cell microscopy. J Cell Biochem 111: 488–496.
Tickner JA, Aaron JU, Sally-Anne S, Derek JR & Kenneth J (2014) Functions and Therapeutic
Roles of Exosomes in Cancer. Front. Oncol. Frontiers in Oncology 4.
Tomasoni S, Longaretti L, Rota C, Morigi M, Conti S & Gotti E (2013) Transfer of growth
factor receptor mRNA via exosomes unravels the regenerative effect of mesenchymal stem cells.
Stem Cells Dev 22: 772–780.
Trams EG, Lauter CJ & Salem N Jr (1981) Exfoliation of membrane ecto-enzymes in the form
of micro-vesicles. Biochim Biophys Acta 645: 63–70.
Turiak L, Misjak P, Szabo TG, Aradi B, Paloczi K, Ozohanics O, Drahos L, Kittel A, Falus A &
Buzas EI (2011) Proteomic characterization of thymocyte-derived microvesicles and apoptotic
bodies in BALB/c mice. J. Proteom 74: 2025–2033.URI: http://eco.iarc.fr.
Valadi H, Ekstrom K & Bossios A (2007) Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 9: 654–659.
Valerius MT, Patterson LT, Witte DP & Potter SS. (2002) Microarray analysis of novel cell lines
representing two stages of metanephric mesenchyme differentiation. Mechanism of Development
112: 219-232.
Page 51
45
Van Deun J, Mestdagh P, Sormunen R, Cocquyt V, Vermaelen k, Vandesompele J, Bracke M,
Wever OD & Hendrix A (2014) The impact of disparate isolation methods for extracellular
vesicles on downstream RNA profiling. Journal of Extracellular vesicles 3; 24858.
Van Ham SM, Heutinck KM & Jorritsma T (2010) Urinary granzyme A mRNA is a biomarker
to diagnose subclinical and acute cellular rejection in kidney transplant recipients. Kidney Int 78:
1033–1040.
Wang G, Kwan BC & Lai FM (2010) Expression of microRNAs in the urinary sediment of
patients with IgA nephropathy. Dis Markers 28: 79–86.
Wolf P (1967) The nature and significance of platelet products in human plasma. Br J Haematol
13; 269-88.
Xie Y, Bai O, Yuan J, Chibbar R, Slattery K, Wei Y, Deng Y & Xiang J (2009) Tumor apoptotic
bodies inhibit CTL responses and antitumor immunity via membrane-bound transforming growth
factor-beta1 inducing CD8+ T-cell energy and CD4+ Tr1 cell responses. Cancer Res 69: 7756-
7766.
Yang C & Robbins PD (2011) The roles of tumor-derived exosomes in cancer pathogenesis. Clin
Dev Immunol 842849.
Zhang Y, Liu D, Chen X, Li J, Li L, Bian Z, Sun F, Lu J, Yin Y, Cai X, Sun Q, Wang Q, Wang
D, Yang J, Liu P, Xu T, Yan Q, Zhang J, Zen K & Zhang CY (2010) Secreted monocytic miR-
150 enhances targeted endothelial cell migration. Mol Cell 39: 133–144.
Zhao Y, Sun X & Cao W (2015) Exosomes Derived from Human Umbilical Cord Mesenchymal
Stem Cells Relieve Acute Myocardial Ischemic Injury. Stem Cells International; 2015:761643.
Zhong H, De Marzo AM & Laughner E (1999) Overexpression of hypoxia-inducible factor
1alpha in common human cancers and their metastases. Cancer Res 59: 5830–5.