Lack of Association between 460 C / T and 936 C / T of the Vascular Endothelial Growth Factor and Angiopoietin-2 Exon 4 G /A Polymorphisms and Ovarian, Cervical, and Endometrial Cancers ECE KONAC, 1 H. ILKE ONEN, 1 JALE METINDIR, 2 EBRU ALP, 1 AYDAN ASYALI BIRI, 3 and ABDULLAH EKMEKCI 1 ABSTRACT Tumor growth, which employs a number of regulators, requires the formation of new blood vessels. The most important regulators are vascular endothelial growth factor (VEGF) and angiopoietin-2 (ANGPT-2). DNA sequence variations in VEGF and ANGPT-2 genes may lead to altered productions and/or activities of these genes. In this study, we aimed to determine the polymorphic effects of the changes in the VEGF 460 C/T, VEGF 936 C/T, and ANGPT-2 exon 4 G/A, which we perceive as risk factors in the progress and metasta- sis of cancer, on the gynecologic cancer patients in the Turkish population. Forty-seven ovarian, 32 cervical, and 21 endometrial cancer patients and 106 healthy controls were studied. The genomic DNA was extracted from the whole blood by using DNA extraction techniques. DNA samples were analyzed by polymerase chain reaction and restriction fragment length polymorphism. There were no significant differences between any of the three types of gynecologic cancer patients and controls in terms of the distribution of VEGF 460, VEGF 936, and ANGPT-2 genotypes and alleles ( p > 0.05). Odds ratios (ORs) were calculated by logistic regression analysis in comparison with the most common homozygote genotype observed in the studied population. No evidence of a relationship that would constitute a risk factor ( p > 0.05) was found between genotype and allele frequencies of patients and controls for VEGF 460, VEGF 936, and ANGPT-2 genes. A multivariable logistic regression analysis with the involvement of covariant factors, such as the history of gynecologic cancer and/or other cancer types in the family, stages of tumor, smoking habits, and existence of other diseases, did not change the results. The present study is the first case-control study of VEGF and ANGPT-2 polymorphisms in relation to ovarian, cervical, and endometrial cancers. INTRODUCTION O VARIAN, CERVICAL, AND ENDOMETRIAL CANCERS are the most common types of gynecologic cancers. Parallel to the increasing female life expectancy, the likelihood of encoun- tering these cancer types, which are often observed in females in the age range of 45–80 and constitute roughly one-fifth of all cancers observed in females, also increases (Repetto et al., 1998). Angiogenesis, which would allow better flow of blood to the ischemic tissues, is the formation of new capillary blood ves- sels (neovascularization) rooting from pre-existing ones. It occurs in normal adult tissues during the healing of a wound and female reproductive cycle, and further activated during the cancer progression. Tumor growth and the process of metas- tasis employ a number of regulators and require the formation of new blood vessels (Saaristo et al., 2000). Various angiogenic growth factors, such as the vascular endothelial growth factor 1 Department of Medical Biology and Genetics, Faculty of Medicine, Gazi University, Besevler, Ankara, Turkey. 2 Department of Obstetrics and Gynecology, Ankara Oncology Education and Research Hospital, Demetevler, Ankara, Turkey. 3 Department of Obstetrics and Gynecology, Faculty of Medicine, Gazi University, Besevler, Ankara, Turkey. DNA AND CELL BIOLOGY Volume 26, Number 7, 2007 # Mary Ann Liebert, Inc. Pp. 453–463 DOI: 10.1089/dna.2007.0585 453
Transcript
Lack of Association between �460 C / T and 936 C / Tof the Vascular Endothelial Growth Factor
and Angiopoietin-2 Exon 4 G /A Polymorphismsand Ovarian, Cervical, and Endometrial Cancers
ECE KONAC,1 H. ILKE ONEN,1 JALE METINDIR,2 EBRU ALP,1
AYDAN ASYALI BIRI,3 and ABDULLAH EKMEKCI1
ABSTRACT
Tumor growth, which employs a number of regulators, requires the formation of new blood vessels. The mostimportant regulators are vascular endothelial growth factor (VEGF) and angiopoietin-2 (ANGPT-2). DNA
sequence variations in VEGF and ANGPT-2 genes may lead to altered productions and/or activities of these
genes. In this study, we aimed to determine the polymorphic effects of the changes in the VEGF �460 C/T,
VEGF 936 C/T, and ANGPT-2 exon 4 G/A, which we perceive as risk factors in the progress and metasta-
sis of cancer, on the gynecologic cancer patients in the Turkish population. Forty-seven ovarian, 32 cervical,
and 21 endometrial cancer patients and 106 healthy controls were studied. The genomic DNA was extracted
from the whole blood by using DNA extraction techniques. DNA samples were analyzed by polymerase chain
reaction and restriction fragment length polymorphism. There were no significant differences between any ofthe three types of gynecologic cancer patients and controls in terms of the distribution of VEGF� 460, VEGF
936, and ANGPT-2 genotypes and alleles ( p> 0.05). Odds ratios (ORs) were calculated by logistic regression
analysis in comparison with the most common homozygote genotype observed in the studied population. No
evidence of a relationship that would constitute a risk factor ( p> 0.05) was found between genotype and allele
frequencies of patients and controls for VEGF� 460, VEGF 936, and ANGPT-2 genes. A multivariable logistic
regression analysis with the involvement of covariant factors, such as the history of gynecologic cancer and/or
other cancer types in the family, stages of tumor, smoking habits, and existence of other diseases, did not
change the results. The present study is the first case-control study of VEGF and ANGPT-2 polymorphisms inrelation to ovarian, cervical, and endometrial cancers.
INTRODUCTION
O VARIAN, CERVICAL, AND ENDOMETRIAL CANCERS are the most
common types of gynecologic cancers. Parallel to the
increasing female life expectancy, the likelihood of encoun-
tering these cancer types, which are often observed in females
in the age range of 45–80 and constitute roughly one-fifth of all
cancers observed in females, also increases (Repetto et al.,
1998).
Angiogenesis, which would allow better flow of blood to the
ischemic tissues, is the formation of new capillary blood ves-
sels (neovascularization) rooting from pre-existing ones. It
occurs in normal adult tissues during the healing of a wound
and female reproductive cycle, and further activated during the
cancer progression. Tumor growth and the process of metas-
tasis employ a number of regulators and require the formation
of new blood vessels (Saaristo et al., 2000). Various angiogenic
growth factors, such as the vascular endothelial growth factor
1Department of Medical Biology and Genetics, Faculty of Medicine, Gazi University, Besevler, Ankara, Turkey.2Department of Obstetrics and Gynecology, Ankara Oncology Education and Research Hospital, Demetevler, Ankara, Turkey.3Department of Obstetrics and Gynecology, Faculty of Medicine, Gazi University, Besevler, Ankara, Turkey.
DNA AND CELL BIOLOGYVolume 26, Number 7, 2007# Mary Ann Liebert, Inc.Pp. 453–463DOI: 10.1089/dna.2007.0585
453
(VEGF) and angiopoietin (ANGPT), as well as cytokines, in-
duce neovascularization in tumors (Cao, 2004; Tait and Jones,
2004; Tammela et al., 2005).
The gene that encodes VEGF is located on chromosome 6
(6p21.3) and comprises a 14-kb-coding region with eight exons
and seven introns (Vincenti et al., 1996). VEGF is a heparin-
binding homodimeric glycoprotein of 45 kDa, and at least six
VEGF isoforms of variable amino acids are produced through
alternative splicing (Ferrara et al., 2003). It has been shown
that elements in both the 50- and 30-flanking regions are sen-
sitive to hypoxia (Minchenko et al., 1994). Additionally,
VEGF strongly stimulates angiogenesis in hypoxic conditions
via hypoxia-inducible factor (HIF), which is a regulated ele-
ment of the VEGF gene (Pugh and Ratcliffe, 2003). A strong
correlation has been found between VEGF expression and in-
creased tumor microvasculature, malignancy, and metastasis in
breast cancer (Saaristo et al., 2000). There is a considerable
variation between individuals in terms of VEGF expression,
and analysis of the 50-flanking region of the gene has shown the
presence of many polymorphisms (Schultz et al., 1999; Watson
et al., 2000). Polymorphisms within the VEGF gene have been
associated with production of VEGF protein and reported to be
susceptible to several diseases whose progress may be critically
affected by angiogenesis (Renner et al., 2000; Watson et al.,
2000; Awata et al., 2002; Cheng-Chieh et al., 2003; Krippl
et al., 2003; Morohashi et al., 2003; Han et al., 2004; Papa-
zoglou et al., 2004a, 2004b; Ray et al., 2004). The most fre-
quently seen polymorphisms are Bsh1236I (C to T) located at
the� 460th (promoter) and Hin1II (C to T) located at the 936th
(30 untranslated) nucleotides of the gene (Cheng-Chieh et al.,
2003; Krippl et al., 2003). Bsh1236I is a suitable genetic
marker of breast cancer (Krippl et al., 2003), whereas Hin1II is
of prostate cancer (Cheng-Chieh et al., 2003). On the basis of
these facts, VEGF may contribute to the development and
progress of gynecologic cancers. However, there have been no
published studies on any populations so far about the associ-
ation of VEGF� 460 C/T and VEGF 936 C/T polymorphisms
with the ovarian, cervical, and endometrial cancers.
The family of human ANGPTs comprises factors with criti-
cally important roles in vascular development and angiogenesis,
especially in the female reproductive tract (Davis et al., 1996;
Maisonpierre et al., 1997; Valenzuela et al., 1999). Due to their
critical roles in adult angiogenesis, the ANGPTs have natu-
rally become the focus of cancer studies (Tait and Jones, 2004).
Three human ANGPTs have been identified: angiopoietin-1
(ANGPT-1), angiopoietin-2 (ANGPT-2), and angiopoietin-4
(ANGPT-4) (Ward et al., 2001). ANGPT-2, comprising 496
amino acids and consisting of nine exons and eight introns, en-
codes the ANGPT-2 protein (Grosios et al., 1999) on chromo-
some 8 (8p23.1) and is highly expressed only at sites of vascular
remodeling in the adult, notably in the ovary, uterus, and placenta
(Maisonpierre et al., 1997; Grosios et al., 1999; Holash et al.,
1999; Valenzuela et al., 1999; Dunk et al., 2000; Ward et al.,
2001). In addition, ANGPT-2 seems to exert vasodilatative ef-
fects in hypoxic conditions, possibly stimulated by the most
prominent mediator of angiogenesis, VEGF (Dunk et al., 2000).
The ANGPT-2 acts through inhibition of the endothelial cell–
specific receptor tyrosine kinase (Tie-2) signaling and leads to a
loosening of cell–matrix and cell–cell contacts, allowing access
to angiogenic inducers, for example, VEGF. Thus, coexpression
of VEGF and that of ANGPT-2 whose mRNA and protein are
detectable in ovarian cancer specimens lead to angiogenesis
(Hirchenhain et al., 2003; Zhang et al., 2003). In ANGPT-2,
three positions were found to be highly polymorphic between
individuals: at position 759 in exon 2, position 1087 in exon 4, and
position 1233 in exon 5 of the cDNA sequence. All detected
polymorphisms are silent mutations and do not result in amino
acid changes in the encoded protein (Ward et al., 2001). The most
common polymorphism of ANGPT-2 is a G/A polymorphism
in exon 4, which is believed to alter protein expression and pos-
sibly inhibit angiogenesis (Pietrowski et al., 2003). ANGPT-2
gene polymorphism was studied, so far, in idiopathic recurrent
miscarriage (IRM), unexplained intrauterine fetal death (IUFD),
and uterine leiomyoma (Pietrowski et al., 2003; Denschlag
et al., 2005; Huber et al., 2005). The biological actions of the
ANGPTs in tumor growth and metastasis have not been fully
ascertained although they have been implicated in the devel-
opment of the vasculature of a wide variety of tumors (Tait and
Jones, 2004). Therefore, the ANGPT-2 polymorphism has only
been linked to obstetric conditions/diseases so far (Huber et al.,
2005). The capability of angiogenic polymorphism to affect the
growth and invasion rates of the tumor led us to investigate the
relationship between them.
In the present study, we aimed to determine whether the
polymorphic effects of the changes in the VEGF� 460 C/T,
VEGF 936 C/T, and ANGPT-2 exon 4 G/A are risk factors
for ovarian, cervical, and endometrial cancer patients in the
Turkish population.
MATERIALS AND METHODS
Study population
A total of 206 subjects, consisting of 106 healthy volunteers
as controls and 100 gynecologic cancer patients who were
selected from patients admitted to the Department of Obstetrics
and Gynecology, Faculty of Medicine, Gazi University, Be-
sevler, Turkey, and to the same department of the Ankara On-
cology Education and Research Hospital, Demetevler, Turkey,
were enrolled in this study. The research was carried out in
accordance with the Declaration of Helsinki (1989) of the
World Medical Association, and the study was approved by
the Committee of Ethics of the Gazi University. All cases and
controls were of Caucasian origin and provided their written
consents for inclusion in the study. Our study conforms to a
great extent the standards presented by Hegele in the form of a
list of the desirable attributes for genetic case–control studies
(Hegele, 2002). In each case, age of menarche and first diag-
nosis; history of gynecologic cancer and/or other cancer types
in the family; stages (I–IV) of tumor; information on smoking
habits; and existence of other diseases, such as psoriasis, dia-
betic retinopathy, rheumatoid arthritis, and cardiovascular dis-
eases (for which angiogenesis may be critical), were obtained
from the patient files. Out of 100 patients, 47 had epithelial
ovarian cancer, 32 women had cervical cancer, and the remain-
ing 21 women had endometrial cancer. Clinical characteristics
of the participating patients were shown in Table 1. The mean
454 KONAC ET AL.
age of menarche of the control group was 13.0� 0.18 years,
and the mean age of the control group was 47� 0.5 years
(range: 25–65 years). None of them had history of gynecologic
cancer or other cancer types in the family, smoking habits, or
other diseases.
Extraction of genomic DNA and amplification of thegenes� 460 VEGF, 936 VEGF, and ANGPT-2
Genomic DNA was isolated from peripheral blood by using a
DNA extraction kit (Heliosis�, Metis Biotechnology, Ankara,
Turkey) according to the manufacturer’s instructions. Amplifica-
tions of the two regions of the VEGF gene and the gene encod-
ing for ANGPT-2 were carried out by placing in a Mastercycler
gradient (Eppendorf, Hamburg, Germany) thermal cycler a total
volume of 50mL PCR mixture containing 50 ng genomic DNA,
2.5 mM magnesium chloride, 100mM deoxyribonucleotide tri-
phosphate, 50 pmol/mL of each primer, and 1.0 U/mL Taq DNA
polymerase. For the� 460 C/T (rs833061), the following pair
of primers produced a PCR product of 175 bp: forward, 50-TGTGCGTGTGGGGTTGAGCG-30; reverse, 50-TACGTGCG
GACAGGGCCTGA-30 (Gene Bank accession no. AF095785;
nucleotides 1883–2057). For the 936 C/T (rs3025039), the
following pair of primers produced a PCR product of 208 bp:
were loaded directly onto 4%, 3%, and 2% agarose gel (con-
taining 0.5% ethidium bromide), respectively. The products were
separated by electrophoresis and visualized by Gel Logic 100 gel
image system (Kodak, Rochester, NY).
Statistical analysis
We used the goodness-of-fit chi-square (w2) test with one
degree of freedom while testing the Hardy–Weinberg equilib-
rium of alleles at individual loci to compare the observed ge-
notype frequencies with the expected genotype frequencies
among the subjects. We then used Pearson’s two-way w2 test to
analyze the distribution of genotype and allele frequencies
of the VEGF� 460 C/T, VEGF 936 C/T, and ANGPT-2 exon 4
G/A polymorphisms in the patients and control group.
When the assumption of the w2 was violated, Fisher’s exact test
was performed. A power calculation was conducted for each
single nucleotide polymorphism (SNP) to indicate which ap-
proximate odds ratio (OR) levels could achieve significant
association between the three types of gynecologic cancer and
the genotypes, given the sample size used and the observed
allele frequencies in controls. We used the logistic regression
analysis to calculate the ORs at 95% confidence interval (CI).
Adjusted ORs for the epidemiological covariant factors, such
as the history of gynecologic cancer and/or other cancer types
in the family, stages of tumor, smoking habits, and existence of
other diseases, were determined by using a multivariable lo-
gistic regression method. We considered results with p< 0.05
as statistically significant. SPSS V.11.5 was employed to carry
out the required statistical analysis.
RESULTS
There were no significant differences between the cases and
controls in mean age and mean age of menarche, suggesting
that the matching based on these two variables was adequate.
The genotype distributions of the three polymorphisms in the
controls and cases were in Hardy–Weinberg equilibrium.
For VEGF� 460 genotypes, frequencies of C alleles of all
three types of gynecologic cancer patients and controls were
TABLE 1. CLINICAL CHARACTERISTICS OF THE
PARTICIPATING PATIENTS
Ovarian
cancer
Cervical
cancer
Endometrial
cancer
Number of cases 47 (47) 32 (32) 21 (21)
Mean ages of
menarche (years)
13� 0.23 13� 0.18 13� 0.14
Mean ages of
first diagnosis (years)
48� 0.51 49� 0.46 50� 0.52
Range (years) 16–73 25–68 39–81
1a 8 (17.0) 7 (21.9) 9 (42.9)
2b
I 1 (2.1) 5 (15.6) 10 (47.6)
II 5 (10.6) 16 (50.0) 5 (23.8)
III 40 (85.1) 10 (31.3) 5 (23.8)
IV 1 (2.1) 1 (3.1) 1 (4.8)
3c 8 (17.0) 4 (12.5) 1 (4.8)
4d 11 (23.4) 5 (15.6) 5 (23.8)
Values in parentheses are percentages.aHistory of gynecologic cancer and/or other cancer types in the
family.bStages of tumor (staging was performed according to the current
International Federation of Gynecology and Obstetrics [FIGO] classi-
fication).cSmoking habits.dExistence of other diseases.
VEGF AND ANGPT-2 POLYMORPHISMS IN GYNECOLOGIC CANCERS 455
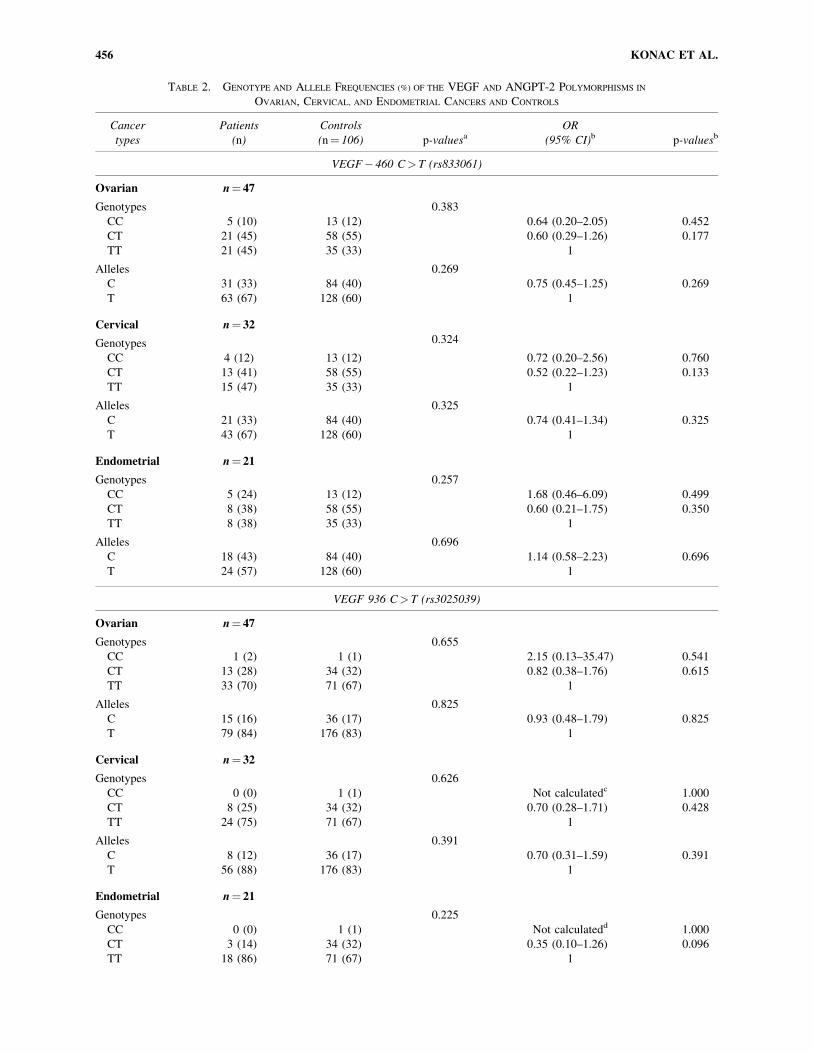
TABLE 2. GENOTYPE AND ALLELE FREQUENCIES (%) OF THE VEGF AND ANGPT-2 POLYMORPHISMS IN
OVARIAN, CERVICAL, AND ENDOMETRIAL CANCERS AND CONTROLS
Cancer
types
Patients
(n)
Controls
(n¼ 106) p-valuesaOR
(95% CI)b p-valuesb
VEGF� 460 C> T (rs833061)
Ovarian n¼ 47
Genotypes 0.383
CC 5 (10) 13 (12) 0.64 (0.20–2.05) 0.452
CT 21 (45) 58 (55) 0.60 (0.29–1.26) 0.177
TT 21 (45) 35 (33) 1
Alleles 0.269
C 31 (33) 84 (40) 0.75 (0.45–1.25) 0.269
T 63 (67) 128 (60) 1
Cervical n¼ 32
Genotypes 0.324
CC 4 (12) 13 (12) 0.72 (0.20–2.56) 0.760
CT 13 (41) 58 (55) 0.52 (0.22–1.23) 0.133
TT 15 (47) 35 (33) 1
Alleles 0.325
C 21 (33) 84 (40) 0.74 (0.41–1.34) 0.325
T 43 (67) 128 (60) 1
Endometrial n¼ 21
Genotypes 0.257
CC 5 (24) 13 (12) 1.68 (0.46–6.09) 0.499
CT 8 (38) 58 (55) 0.60 (0.21–1.75) 0.350
TT 8 (38) 35 (33) 1
Alleles 0.696
C 18 (43) 84 (40) 1.14 (0.58–2.23) 0.696
T 24 (57) 128 (60) 1
VEGF 936 C> T (rs3025039)
Ovarian n¼ 47
Genotypes 0.655
CC 1 (2) 1 (1) 2.15 (0.13–35.47) 0.541
CT 13 (28) 34 (32) 0.82 (0.38–1.76) 0.615
TT 33 (70) 71 (67) 1
Alleles 0.825
C 15 (16) 36 (17) 0.93 (0.48–1.79) 0.825
T 79 (84) 176 (83) 1
Cervical n¼ 32
Genotypes 0.626
CC 0 (0) 1 (1) Not calculatedc 1.000
CT 8 (25) 34 (32) 0.70 (0.28–1.71) 0.428
TT 24 (75) 71 (67) 1
Alleles 0.391
C 8 (12) 36 (17) 0.70 (0.31–1.59) 0.391
T 56 (88) 176 (83) 1
Endometrial n¼ 21
Genotypes 0.225
CC 0 (0) 1 (1) Not calculatedd 1.000
CT 3 (14) 34 (32) 0.35 (0.10–1.26) 0.096
TT 18 (86) 71 (67) 1
456 KONAC ET AL.
smaller than the frequencies of T alleles. We did not find sta-
tistically significant differences in the genotypes and allele
frequencies between the patients and controls of all three types
of gynecologic cancers. The ORs per copy of the C allele were
0.75 (95% CI: 0.45–1.25; p¼ 0.269) for ovarian cancer, 0.74
(95% CI: 0.41–1.34; p¼ 0.325) for cervical cancer, and 1.14
(95% CI: 0.58–2.23; p¼ 0.696) for endometrial cancer (Table
2). In other words, the allelic frequencies of the gene were not
significantly associated with any of the three types of gyne-
cologic cancer patients.
For VEGF 936 genotypes, out of 106 healthy controls, 1 was
type CC, 34 were type CT, and 71 were type TT. In addition, in
the whole group of 100 patients there was only one type CC,
which was encountered in 1 of the ovarian cancer patients. VEGF
936 TT was the most common genotype observed in the Turk-
ish population. Genotype and allele frequencies of VEGF 936
were again not significantly different between the patients and
controls of the three types of gynecologic cancers. The ORs per
copy of the C allele were 0.93 (95% CI: 0.48–1.79; p¼ 0.825) for
ovarian cancer, 0.70 (95% CI: 0.31–1.59; p¼ 0.391) for cervi-
cal cancer, and 0.38 (95% CI: 0.11–1.28; p¼ 0.106) for endo-
metrial cancer (Table 2), again pointing to the fact that the allelic
frequencies of the gene were not significantly associated with
any of the three types of gynecologic cancer patients.
TABLE 2. (Continued)
Cancer
types
Patients
(n)
Controls
(n¼ 106) p-valuesaOR
(95% CI)b p-valuesb
VEGF 936 C> T (rs3025039)
Alleles 0.106
C 3 (7) 36 (17) 0.38 (0.11–1.28) 0.106
T 39 (93) 176 (83) 1
ANGPT-2 exon 4 G>A (rs3020221)
Ovarian n¼ 47
Genotypes 0.989
GG 11 (23) 26 (25) 1.10 (0.41–2.27) 0.892
GA 22 (47) 49 (46) 1.10 (0.45–2.52) 0.893
AA 14 (30) 31 (29) 1
Alleles 0.893
G 44 (47) 101 (48) 1.03 (0.64–1.68) 0.893
A 50 (53) 111 (52) 1
Cervical n¼ 32
Genotypes 0.157
GG 5 (16) 26 (25) 1.01 (0.27–3.68) 0.992
GA 21 (66) 49 (46) 2.23 (0.75–6.60) 0.141
AA 6 (18) 31 (29) 1
Alleles 0.570
G 31 (48) 101 (48) 1.21 (0.62–2.37) 0.570
A 33 (52) 111 (52) 1
Endometrial n¼ 21
Genotypes 0.031
GG 1 (5) 26 (25) 3.35 (0.35–31.91) 0.376
GA 16 (76) 49 (46) 8.49 (1.06–67.60) 0.019
AA 4 (19) 31 (29) 1
Alleles 0.570
G 18 (43) 101 (48) 1.21 (0.62–2.37) 0.570
A 24 (57) 111 (52) 1
Values in parentheses are percentages.aResults of ovarian, cervical, and endometrial cancer groups compared with control group.bCalculations were performed TT vs. CT and CC for VEGF� 460 and 936; AA vs. GA and GG for ANGPT-2.cThere are no CC-genotype patients.dThere are no CC-genotype patients.
OR, odds ratio; CI, confidence interval.
VEGF AND ANGPT-2 POLYMORPHISMS IN GYNECOLOGIC CANCERS 457
Finally, regarding the ANGPT-2 exon 4 G/A genotypes,
forty-eight percent of the control group and 43–48% of the
cancer patients were found to have G allele frequencies. There
were no significant differences in the genotype and allele fre-
quencies of ANGPT-2 between the controls and patients. The
ORs per copy of the G allele were 1.03 (95% CI: 0.64–1.68;
p¼ 0.893) for ovarian cancer, 1.21 (95% CI: 0.62–2.37;
p¼ 0.570) for cervical cancer, and 1.21 (95% CI: 0.62–2.37;
p¼ 0.570) for endometrial cancer (Table 2). In other words, the
allelic frequencies of the gene were again not significantly
associated with the three types of gynecologic cancer patients.
Power calculation was conducted for each SNP (VEGF�460 C/T, VEGF 936 C/T, and ANGPT-2 exon 4 G/A) to
find out approximate OR levels, which would achieve signifi-
cant association between the three types of gynecologic cancer
and the genotypes, given the sample sizes used and the ob-
served allele frequencies in controls. OR levels calculated by
the power analysis were from 2.7 (1.3–5.8) to 15.2 (1.7–132.1)
for ovarian cancer, from 3.05 (1.35–6.89) to 8.7 (1.9–38.8) for
cervical cancer, and from 3.4 (1.3–8.96) to 9.4 (1.9– 47.2) for
endometrial cancer with respect to reference genotypes (power
of 0.80, significance level of 0.05).
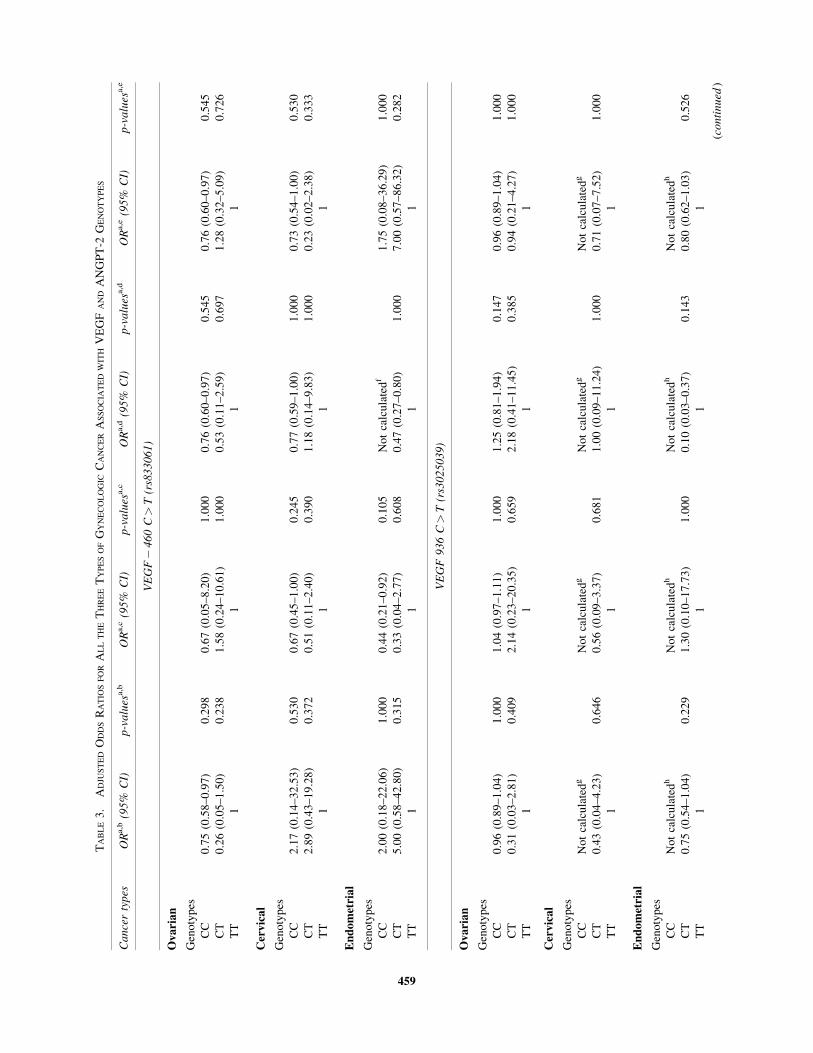
Multivariable logistic regression analysis demonstrated that
as far as VEGF� 460 C/T, VEGF 936 C/T, and ANGPT-2
exon 4 G/A polymorphisms were concerned, the occurrences of
ovarian, cervical, and endometrial cancers were not associated
with the history of gynecologic cancer or other cancer types in
the family, smoking habits, stages of tumor, or existence of
other diseases (Table 3).
DISCUSSION
Humans are believed to carry over a million distinct SNPs
(Li et al., 2001). Determination of SNPs is a new means to
study the etiology of polygenetic disorders with complex in-
heritance patterns (Denschlag et al., 2005). For instance, they
affect the sensibility of cancer development and progression
via actions on the pathways of tumor angiogenesis. This may
be achieved by enhanced or reduced transcription, altered
posttranscriptional or posttranslational activity, or changes in
the tertiary structure of the gene product (Liu et al., 1995).
Polymorphisms of VEGF and ANGPT-2 genes may lead to
differences in their expression levels among individuals and
could potentially contribute to a variety of pathological pro-
cesses. Since the implications of VEGF and ANGPT-2 poly-
morphisms for gynecologic cancers have not been studied so
far, we, in this study, investigate the polymorphic effects of the
changes in the VEGF� 460 C/T, VEGF 936 C/T, and
ANGPT-2 exon 4 G/A on the ovarian, cervical, and endometrial
cancer patients in the Turkish population.
VEGF is a major angiogenic factor and is a prime regulator
of endothelial cell proliferation (Ferrara et al., 2003). A 28-bp
element within the human VEGF promoter believed to contain
a consensus binding site for hypoxia-inducible factor-1 (HIF-1)
and is necessary to increase transcription of a chloramphenicol
acetyltransferase (CAT) reporter gene in endothelial cells
cultured under hypoxic conditions (Liu et al., 1995). Many
reports have indicated that VEGF might combine with up-
stream signals or other growth factors to enhance prostate
cancer growth and progression (Ravindranath et al., 2001).
VEGF� 460 C/T polymorphism is associated with prostate
cancer. While the TT homozygotes, the most common geno-
type found in patients with prostate cancer in Taiwan, seemed
to impose a significant risk for the emergence of prostate
cancer, it did not play a role in the cancer progression (Cheng-
Chieh et al., 2003). Another study showed that TT homozy-
gotes and the T allele of the VEGF� 460 gene were associated
with a higher risk of endometriosis. In other words, VEGF
polymorphism is likely to contribute to the pathogenesis of en-
dometriosis and may become a useful marker for predicting
endometriosis susceptibility (Hsieh et al., 2004). However, the
distribution of genotype and allele frequencies of the VEGF�460 C/T polymorphism did not differ significantly between en-
dometriosis patients and controls in South Indian and Korean
populations (Bhanoori et al., 2005; Kim et al., 2005). In another
study in which we had investigated VEGF� 634 G>C poly-
morphism, we reported that VEGF� 634 genetic variants did
not play a relevant role in the pathogenesis of ovarian, cervical,
and endometrial cancers (Konac et al., 2006). However, in our
recent study (Konac et al., 2007), in which we had investigated
HIF-1a gene polymorphisms, results suggested that the C1772T
polymorphism of the HIF-1a might be associated with cervical
and endometrial cancers. In this study, we found that the least
common genotype of VEGF� 460 C/T polymorphism was
CC homozygote in both the gynecologic cancer patients and the
control group. The distribution of genotypes and allele frequen-
cies of the� 460 C/T polymorphism in controls in South
Indian population (Bhanoori et al., 2005) is consistent with the
distribution of genotypes and allele frequencies presented in the
current study. Studies on Korean population have revealed that
the most common genotype was TT in endometriosis and lung
cancer patients (Kim et al., 2005; Lee et al., 2005). We found
that� 460 T allele carriers (CTþTT genotypes) were most
frequent among the cancer patients. However, there was no sta-
tistically significant difference between the patients and the con-
trols in terms of genotypes ( p> 0.05). Multivariable logistic
regression analysis for the VEGF� 460 demonstrated that the
occurrences of ovarian, cervical, and endometrial cancers were
not associated with the history of gynecologic cancer or other
cancer types in the family, stages of tumor, smoking habits, and
existence of other diseases ( p> 0.05) (Table 3).
A C/T polymorphism at position 936 in the 30 untranslated
region of the VEGF gene has been associated with VEGF
plasma levels (Renner et al., 2000; Krippl et al., 2003). Al-
though it is unclear how the polymorphism in the untranslated
region of the gene influences its protein production, several
reports have demonstrated that SNPs of the VEGF gene are
associated with VEGF synthesis (Watson et al., 2000; Awata
et al., 2002). The T variant, which is linked to lower VEGF
levels, has been associated with breast cancer, sarcoidosis, and
spontaneous preterm delivery (Krippl et al., 2003; Morohashi
et al., 2003; Papazoglou et al., 2004a). Individuals with VEGF
936 TT or CT genotype demonstrated a statistically significant
association with preterm delivery compared with those sharing
936 CC genotype in the Greek population (Papazoglou et al.,
2004a). The 936 C allele carries a potential binding site for
activator protein 4 (AP-4), a helix-loop-helix transcription
factor enhancing expression of several viral and cellular genes
by binding to specific enhancer sites, which are abolished in the
458 KONAC ET AL.
TABLE
3.
AD
JUS
TE
DO
DD
SR
AT
IOS
FO
RA
LL
TH
ET
HR
EE
TY
PE
SO
FG
YN
EC
OL
OG
ICC
AN
CE
RA
SS
OC
IAT
ED
WIT
HV
EG
FA
ND
AN
GP
T-2
GE
NO
TY
PE
S
Cancertypes
OR
a,b(95%
CI)
p-values
a,b
OR
a,c(95%
CI)
p-values
a,c
OR
a,d(95%
CI)
p-values
a,d
OR
a,e(95%
CI)
p-values
a,e
VEGF�460C>T(rs833061)
Ovarian
Gen
oty
pes
CC
0.7
5(0
.58
–0
.97
)0
.29
80
.67
(0.0
5–
8.2
0)
1.0
00
0.7
6(0
.60
–0
.97
)0
.54
50
.76
(0.6
0–
0.9
7)
0.5
45
CT
0.2
6(0
.05
–1
.50
)0
.23
81
.58
(0.2
4–
10
.61
)1
.00
00
.53
(0.1
1–
2.5
9)
0.6
97
1.2
8(0
.32
–5
.09
)0
.72
6
TT
11
11
Cervical
Gen
oty
pes
CC
2.1
7(0
.14
–3
2.5
3)
0.5
30
0.6
7(0
.45
–1
.00
)0
.24
50
.77
(0.5
9–
1.0
0)
1.0
00
0.7
3(0
.54
–1
.00
)0
.53
0
CT
2.8
9(0
.43
–1
9.2
8)
0.3
72
0.5
1(0
.11
–2
.40
)0
.39
01
.18
(0.1
4–
9.8
3)
1.0
00
0.2
3(0
.02
–2
.38
)0
.33
3
TT
11
11
Endometrial
Gen
oty
pes
CC
2.0
0(0
.18
–2
2.0
6)
1.0
00
0.4
4(0
.21
–0
.92
)0
.10
5N
ot
calc
ula
ted
f1
.75
(0.0
8–
36
.29
)1
.00
0
CT
5.0
0(0
.58
–4
2.8
0)
0.3
15
0.3
3(0
.04
–2
.77
)0
.60
80
.47
(0.2
7–
0.8
0)
1.0
00
7.0
0(0
.57
–8
6.3
2)
0.2
82
TT
11
11
VEGF936C>T(rs3025039)
Ovarian
Gen
oty
pes
CC
0.9
6(0
.89
–1
.04
)1
.00
01
.04
(0.9
7–
1.1
1)
1.0
00
1.2
5(0
.81
–1
.94
)0
.14
70
.96
(0.8
9–
1.0
4)
1.0
00
CT
0.3
1(0
.03
–2
.81
)0
.40
92
.14
(0.2
3–
20
.35
)0
.65
92
.18
(0.4
1–
11
.45
)0
.38
50
.94
(0.2
1–
4.2
7)
1.0
00
TT
11
11
Cervical
Gen
oty
pes
CC
No
tca
lcu
late
dg
No
tca
lcu
late
dg
No
tca
lcu
late
dg
No
tca
lcu
late
dg
CT
0.4
3(0
.04
–4
.23
)0
.64
60
.56
(0.0
9–
3.3
7)
0.6
81
1.0
0(0
.09
–1
1.2
4)
1.0
00
0.7
1(0
.07
–7
.52
)1
.00
0
TT
11
11
Endometrial
Gen
oty
pes
CC
No
tca
lcu
late
dh
No
tca
lcu
late
dh
No
tca
lcu
late
dh
No
tca
lcu
late
dh
CT
0.7
5(0
.54
–1
.04
)0
.22
91
.30
(0.1
0–
17
.73
)1
.00
00
.10
(0.0
3–
0.3
7)
0.1
43
0.8
0(0
.62
–1
.03
)0
.52
6
TT
11
11
(continued
)
459
TABLE
3.
(Continued
)
Cancertypes
OR
a,b(95%
CI)
p-values
a,b
OR
a,c(95%
CI)
p-values
a,c
OR
a,d(95%
CI)
p-values
a,d
OR
a,e(95%
CI)
p-values
a,e
ANGPT-2
exon4G>A(rs3020221)
Ovarian
Gen
oty
pes
GG
2.7
3(0
.24
–3
0.6
6)
0.6
04
1.3
0(0
.07
–2
3.4
3)
1.0
00
1.8
0(0
.26
–1
2.3
0)
0.6
61
1.0
7(0
.18
–6
.21
)1
.00
0
GA
2.2
2(0
.22
–2
2.7
)0
.64
30
.45
(0.0
4–
4.6
0)
0.6
43
0.4
5(0
.05
–3
.72
)0
.58
60
.59
(0.1
1–
3.2
9)
0.6
61
AA
11
11
Cervical
Gen
oty
pes
GG
0.8
0(0
.04
–1
7.2
0)
1.0
00
0.3
0(0
.02
–4
.91
)0
.54
52
.00
(0.1
3–
31
.98
)1
.00
00
.44
(0.2
1–
0.9
2)
0.4
55
GA
1.2
5(0
.11
–1
3.9
2)
1.0
00
0.9
2(0
.13
–6
.78
)1
.00
00
.20
(0.0
1–
3.9
1)
0.3
54
0.7
8(0
.63
–0
.97
)1
.00
0
AA
11
11
Endometrial
Gen
oty
pes
GG
1.3
3(0
.76
–2
.35
)1
.00
01
.33
(0.7
6–
2.3
5)
1.0
00
No
tca
lcu
late
di
0.6
7(0
.30
–1
.48
)1
.00
0
GA
1.2
0(0
.84
–1
.72
)0
.35
31
.50
(0.6
7–
3.3
4)
0.1
76
0.9
4(0
.83
–1
.06
)1
.00
00
.92
(0.7
9–
1.0
8)
1.0
00
AA
11
11
OR
,o
dd
sra
tio
;C
I,co
nfi
den
cein
terv
al.
aC
alcu
lati
ons
wer
eper
form
edT
Tvs.
CT
and
CC
for
VE
GF�
46
0an
d9
36
;A
Av
s.G
Aan
dG
Gfo
rA
NG
PT
-2.
bO
Rad
just
edfo
rh
isto
ryo
fg
yn
ecolo
gic
can
cer
and
/or
oth
erca
nce
rty
pes
inth
efa
mil
y.
cO
Rad
just
edfo
rst
ages
of
tum
or.
dO
Rad
just
edfo
rsm
ok
ing
hab
its.
eO
Rad
just
edfo
rex
iste
nce
of
oth
erd
isea
ses.
f Th
ere
are
no
CC
-gen
oty
pe
pat
ien
tsw
ho
smo
ke.
gT
her
ear
en
oC
C-g
enoty
pe
pat
ien
ts.
hT
her
ear
en
oC
C-g
enoty
pe
pat
ien
ts.
i Th
ere
are
no
GG
-gen
oty
pe
pat
ien
tsw
ho
smo
ke.
460
936 T allele (Renner et al., 2000). The loss of this potential
binding site could be responsible for decreased VEGF expres-
sion by the T allele (Hu et al., 1990). In our study, for VEGF 936
C/T polymorphism, there were no statistically significant dif-
ferences between the genotype and allele frequencies of the
patients and the control group ( p> 0.05). The reason for this
result is that genotype TT was the most common in both the
control group and the patients (Table 2). Interestingly, we found
only one CC genotype (ovarian) among the whole patient group
and another in the control group. There was no CC genotype in
cervical and endometrial patients. In contrast, a low number of
TT homozygotes was found in patients and controls in Austrian
(Krippl et al., 2003) and Tunisian population (Sfar et al., 2006).
In another study, allelic frequency of 936 T variant in pre-
eclampsia, in which VEGF plays a crucial role, was found to be
20.2% (Papazoglou et al., 2004a). Furthermore, carriers of the
VEGF 936 T allele were less frequent in breast cancer patients
than in healthy subjects, indicating that this genetic variant may
be protective against breast cancer (Krippl et al., 2003). The
936 CT genotype and the combined 936 CT and TT genotype
were also associated with a significantly decreased risk of small-
cell lung cancer compared with the 936 CC genotype (Lee et al.,
2005). The discrepancy might be due to ethnic composition
differences between the studies, sizes of study population, or the
fact that the mechanism by which the 936 C/T polymorphism
affects serum levels of VEGF might differ during the progres-
sion of breast, lung, and gynecologic cancers. No evidence of a
relationship between clinicopathological characteristics, in-
cluding family history, stages and grades of tumor, smoking
habits, and existence of other diseases, and 936 C/T poly-
morphism in ovarian, cervical, and endometrial cancer patients
was observed.
ANGPT-2 appears to be highly polymorphic in a few indi-
viduals. The polymorphism in exon 2 suggests that this position
has been the site of two independent mutations and that the
surrounding structure is conducive to mutations (Ward et al.,
2001). On the other hand, the polymorphism in exon 4 of
ANGPT-2 is a silent mutation and does not result in amino acid
changes in the encoded protein (Ward et al., 2001). However,
silent mutations are often associated with expression differences
in various genes on the protein level (Stanford et al., 2000). It
should be noted that there are just a few studies on ANGPT-2
exon 4 G/A polymorphism, such as those on IUFD and IRM
(Pietrowski et al., 2003; Huber et al., 2005). The G/A poly-
morphism in exon 4 of ANGPT-2 is not associated with IRM in
a Middle European white population (Pietrowski et al., 2003).
Furthermore, the polymorphism does not seem to be a candidate
gene for unexplained IUFD in Caucasian women (Huber et al.,
2005). Denschlag et al. (2005) reported that there was no asso-
ciation between the ANGPT-2 genotype and its allele frequen-
cies and leiomyoma in Caucasian population. Nevertheless, there
are no data published to date with respect to this polymorphism
in gynecologic cancers. In our study, we found that the most
common genotype in all three types of gynecologic cancer pa-
tients and controls was GA heterozygote (Table 2). Therefore,
there was no statistically significant difference between the
patients and the controls in terms of genotypes and allele fre-
quencies ( p> 0.05). In addition, ORs per copy of the G allele
showed that carriers of an ANGPT-2 G allele did not seem to
increase the risk of gynecologic cancers in the Turkish popula-
tion. In addition, there was no statistically significant associa-
tion between the genotypes of patients and family history, stages
of tumor, smoking habits, and existence of other diseases (Table
3). Since the association between the three polymorphisms we
studied and gynecologic cancers has not yet been investigated,
we were unable to compare our results with those of similar
studies covering other ethnic populations.
In conclusion, the results of this study suggest that VEGF�460 C/T, VEGF 936 C/T, and ANGPT-2 exon 4 G/A poly-
morphisms are not associated with any of the three types of
gynecologic cancers. It is unlikely that these three types of
polymorphisms account for a substantial proportion of ovarian,
cervical, and endometrial cancers. The main limitation of this
study is the small sample size due to our criteria for the se-
lection of the cases and controls. Therefore, larger prospective
and multiethnic studies are needed to investigate the functional
relevance of the three polymorphic genes and their receptors to
gynecologic cancer development and progression. Further-
more, the functionality of the polymorphic regions needs to be
analyzed by expression studies. Certainly, further in vivo and in
vitro experimentations are required to demonstrate the specific
role of VEGF and ANGPT-2 in angiogenesis of gynecologic
cancer types and elucidate the mechanisms involved. Our re-
sults, which should be bolstered by future complementary
in vivo, in vitro, and gene expression studies, are expected to
help better understand the bond between the SNPs and the
ovarian, cervical, and endometrial cancers, as well as the pro-
spective diagnosis and treatment phases.
ACKNOWLEDGMENT
This study has been supported by Gazi University Research
Fund (No. 11/2004-13).
REFERENCES
AWATA, T., INOUE, K., KURIHARA, S., OHKUBO, T., WATA-NABE, M., INUKAI, K., INOUE, I., and KATAYAMA, S. (2002).A common polymorphism in the 50-untranslated region of the VEGFgene is associated with diabetic retinopathy in type 2 diabetes. Di-abetes 51, 1635–1639.
BHANOORI, M., ARVIND BABU, K., PAVANKUMAR REDDY,N.G., LAKSHMI RAO, K., ZONDERVAN, K., DEENADAYAL,M., KENNEDY, S., and SHIVAJI, S. (2005). The vascular endo-thelial growth factor (VEGF)þ 405 G>C 50-untranslated regionpolymorphism and increased risk of endometriosis in South Indianwomen: a case control study. Hum. Reprod. 20, 1844–1849.
CAO, Y. (2004). Antiangiogenic cancer therapy. Semin. Cancer Biol.14, 139–145.
CHENG-CHIEH, L., HSI-CHIN, W., FUU-JEN, T., HUEY-YI, C., andWEN-CHI, C. (2003). Vascular endothelial growth factor gene�460C/T polymorphism is a biomarker for prostate cancer. Urology62, 374–377.
DAVIS, S., ALDRICH, T.H., JONES, P.F., ACHESON, A., COMP-TON, D.L., JAIN, V., RYAN, T.E., BRUNO, J., RADZIEJEWSKI,C., MAISONPIERRE, P.C., and YANCOPOULOS, G.D. (1996).Isolation of angiopoietin-1, a ligand for the TIE2 receptor, bysecretion-trap expression cloning. Cell 87, 1161–1169.
DENSCHLAG, D., BETTENDORF, H., WATERMANN, D., KECK,C., TEMPFER, C., and PIETROWSKI, D. (2005). Polymorphism ofthe p53 tumor suppressor gene is associated with susceptibility touterine leiomyoma. Fertil. Steril. 84, 162–166.
VEGF AND ANGPT-2 POLYMORPHISMS IN GYNECOLOGIC CANCERS 461
DUNK, C., SHAMS, M., NIJJAR, S., RHAMAN, M., QIU, Y.,BUSSOLATI, B., and AHMED, A. (2000). Angiopoietin-1 and an-giopoietin-2 activate trophoblast Tie-2 to promote growth and mi-gration during placental development. Am. J. Pathol. 156, 2185–2199.
FERRARA, N., GERBER, H.P., and LECOUTER, J. (2003). The bi-ology of VEGF and its receptors. Nat. Med. 9, 669–676.
GROSIOS, K., LEEK, J.P., MARKHAM, A.F., YANCOPOULOS,G.D., and JONES, P.F. (1999). Assignment of angiopoietin 4, 1 and2 to human chromosome band(s) 20p13.3, 8q22.3-23 and 8p23.1,respectively by in situ hybridization and radiation hybrid mapping.Cytogenet. Cell Genet. 84, 118–120.
HAN, S.W., KIM, G.W., SEO, J.S., KIM, S.J., SA, K.H., PARK, J.Y.,LEE, J., KIM, S.Y., GORONZY, J.J., WEYAND, C.M., and KANG,Y.M. (2004). VEGF gene polymorphisms and susceptibility torheumatoid arthritis. Rheumatology 43, 1173–1177.
HEGELE, R.A. (2002). SNP judgments and freedom of associaiton.Arterioscler. Thromb. Vasc. Biol. 22, 1058–1061.
HIRCHENHAIN, J., HUSE, I., HESS, A., BIELFELD, P., DEBRUYNE, F., and KRUSSEL, J.S. (2003). Differential expression ofangiopoietins 1 and 2 and their receptor Tie-2 in human endome-trium. Mol. Hum. Reprod. 9, 663–669.
HOLASH, J., MAISONPIERRE, P.C., COMPTON, D., BOLAND, P.,ALEXANDER, C.R., ZAGZAG, D., YANCOPOULOS, G.D., andWIEGAND, S.J. (1999). Vessel cooption, regression and growth intumors mediated by angiopoietins and VEGF. Science 284, 1994–1998.
HSIEH, Y.Y., CHANG, C.C., TSAI, F.J., YEH, L.S., LIN, C.C., andPENG, C.T. (2004). T allele for VEGF gene� 460 polymorphism atthe 50-untranslated region: association with a higher susceptibility toendometriosis. J. Reprod. Med. 49, 468–472.
HU, Y.F., LUSCHER, B., ADMON, A., MERMOD, N., and TIJAN, R.(1990). Transcription factor AP-4 contains multiple dimeriza-tion domains that regulate dimer specificity. Genes Dev. 4, 1741–1752.
HUBER, A., GRIMM, C., PIETROWSKI, D., ZEILLINGER, R.,BETTENDORF, H., HUSSLEIN, P., and HEFLER, L. (2005).An angiopoietin-2 gene polymorphism in unexplained intrauterinfetal death: a multi-center study. J. Reprod. Immunol. 65, 47–53.
KIM, S.H., CHOI, Y.M., CHOUNG, S.H., JUN, J.K., KIM, J.G., andMOON, S.Y. (2005). Vascular endothelial growth factor geneþ 405C/G polymorphism is associated with susceptibility to advancedstage endometriosis. Hum. Reprod. 20, 2904–2908.
KONAC, E., ONEN, H.I., METINDIR, J., ALMAYANLAR, E.,ASYALI BIRI, A., and EKMEKCI, A. (2006). The relationshipbetween vascular endothelial growth factor� 634 G>C polymor-phism and ovarian, cervical and endometrial cancers. Korean J.Genet. 28, 99–107.
KONAC, E., ONEN, H.I., METINDIR, J., ALMAYANLAR, E.,ASYALI BIRI, A., and EKMEKCI, A. (2007). An investigation ofrelationships between hypoxia-inducible factor-1� gene polymor-phisms and ovarian, cervical and endometrial cancers. Cancer De-tect. Prev. 31, 102–109.
KRIPPL, P., LANGSENLEHNER, U., RENNER, W., YAZDANI-BIUKI, B., WOLF, G., WASCHER, T.C., PAULWEBER, B.,HAAS, J., and SAMONIGG, H. (2003). A common 936 C/Tgene polymorphism of vascular endothelial growth factor is as-sociated with decreased breast cancer risk. Int. J. Cancer 106, 468–471.
LEE, S.J., LEE, S.Y., JEON, H.S., PARK, S.H., JANG, J.S., LEE,G.Y., SON, J.W., KIM, C.H., LEE, W.K., KAM, S., PARK, R.W.,PARK, T.I., KANG, Y.M., KIM, I.S., JUNG, T.H., and PARK, J.Y.(2005). Vascular endothelial growth factor gene polymorphisms andrisk of primary lung cancer. Cancer Epidemiol. Biomarkers Prev. 14,571–575.
LI, W.H., GU, Z., WANG, H., and NEKRUTENKO, A. (2001). Evo-lutionary analyses of the human genome. Nature 409, 847–849.
LIU, Y., COX, S.R., MORITA, T., and KOUREMBANAS, S. (1995).Hypoxia regulates vascular endothelial growth factor gene expres-sion in endothelial cells: identification of a 50 enhancer. Circ. Res.77, 638–643.
MAISONPIERRE, P.C., SURI, C., JONES, P.F., BARTUNKOVA, S.,WIEGAND, S.J., RADZIEJEWSKI, C., COMPTON, D.,MCCLAIN, J., ALDRICH, T.H., PAPADOPOULOS, N., DALY,T.J., DAVIS, S., SATO, T.N., and YANCOPOULOS, G.D. (1997).Angiopoietin-2, a natural antagonist for Tie2 that disrupts in vivoangiogenesis. Science 277, 55–60.
MINCHENKO, A., SALCEDA, S., BAUER, T., and CARO, J. (1994).Hypoxia regulatory elements of the human vascular endothelialgrowth factor gene. Cell. Mol. Biol. Res. 40, 35–39.
MOROHASHI, K., TAKADA, T., OMORI, K., SUZUKI, E., andGEJYO, F. (2003). Vascular endothelial growth factor gene poly-morphisms in Japanese patients with Sarcoidosis. Chest 123, 1520–1526.
PAPAZOGLOU, D., GALAZIOS, G., KOUKOURAKIS, M.I., KON-TOMANOLIS, E.N., and MALTEZOS, E. (2004a). Associationof� 634 G/C and 936 C/T polymorphisms of the vascular endo-thelial growth factor with spontaneous preterm delivery. Acta Ob-stet. Gynecol. Scand. 83, 461–465.
PAPAZOGLOU, D., GALAZIOS, G., KOUKOURAKIS, M.I., PA-NAGOPOULOS, I., KONTOMANOLIS, E.N., PAPATHEODOR-OU, K., and MALTEZOS, E. (2004b). Vascular endothelial growthfactor gene polymorphisms and pre-eclampsia. Mol. Hum. Reprod.10, 321–324.
PIETROWSKI, D., TEMPFER, C., BETTENDORF, H., BURKLE, B.,NAGELE, F., UNFRIED, G., and KECK, C. (2003). Angiopoietin-2polymorphism in women with idiopathic recurrent miscarriage.Fertil. Steril. 80, 1026–1029.
PUGH, C.W., and RATCLIFFE, P.J. (2003). Regulation of angiogen-esis by hypoxia: role of the HIF System. Nat. Med. 9, 677–684.
RAVINDRANATH, N., WION, D., BRACHET, P., and DJAKIEW, D.(2001). Epidermal growth factor modulates the expression of vas-cular endothelial growth factor in the human prostate. J. Androl. 22,432–443.
RAY, D., MISHRA, M., RALPH, S., READ, I., DAVIES, R., andBRENCHLEY, P. (2004). Association of the VEGF gene withproliferative diabetic retinopathy but not proteinuria in diabetes.Diabetes 53, 861–864.
RENNER, W., KOTSCHAN, S., HOFFMANN, C., OBERMAYER-PIETSCH, B., and PILGER, E. (2000). A common 936 C/T mutationin the gene for vascular endothelial growth factor is associated withvascular endothelial growth factor plasma levels. J. Vasc. Res. 37,443–448.
REPETTO, L., GRANETTO, C., AAPRO, M., and PECORELLI,S. (1998). Gynecologic cancers. Crit. Rev. Oncol. Hematol. 27,147–150.
SAARISTO, A., KARPANEN, T., and ALITALO, K. (2000). Mech-anisms of angiogenesis and their use in the inhibition of tumorgrowth and metastasis. Oncogene 19, 6122–6129.
SCHULTZ, A., LAVIE, L., HOCHBERG, I., BEYAR, R., STONE, T.,SKORECKI, K., LAVIE, P., ROGUIN, A., and LEVY, A.P. (1999).Interindividual heterogeneity in the hypoxic regulation of VEGF:significance for the development of coronary artery collateral cir-culation. Circulation 100, 547–552.
SFAR, S., HASSEN, E., SAAD, H., MOSBAH, F., and CHOU-CHANE, L. (2006). Association of VEGF genetic polymorphismswith prostate carcinoma risk and clinical outcome. Cytokine 35, 21–28.
STANFORD, P.M., HALLIDAY, G.M., BROOKS, W.S., KWOK,J.B., STOREY, C.E., CREASEY, H., MORRIS, J.G., FULHAM,M.J., and SCHOFIELD, P.R. (2000). Progressive supranuclear palsypathology caused by a novel silent mutation in exon 10 of the taugene: expansion of the disease phenotype caused by tau gene mua-tions. Brain 123, 880–893.
TAIT, C.R., and JONES, P.F. (2004). Angiopoietins in tumours: theangiogenic switch. J. Pathol. 204, 1–10.
TAMMELA, T., ENHOLM, B., ALITALO, K., and PAAVONEN, K.(2005). The biology of vascular endothelial growth factors. Cardi-ovasc. Res. 65, 550–563.
N., MAISONPIERRE, P.C., DAVIS, S., and YANCOPOULOS,G.D. (1999). Angiopoietins 3 and 4: diverging gene counterparts inmice and humans. Proc. Natl. Acad. Sci. USA 96, 1904–1909.
VINCENTI, V., CASSANO, C., ROCCHI, M., and PERSICO, G.(1996). Assignment of the vascular endothelial growth factor gene tohuman chromosome 6p21.3. Circulation 93, 1493–1495.
WARD, E.G., GROSIOS, K., MARKHAM, A.F., and JONES, P.F.(2001). Genomic structures of the human angiopoietins showpolymorphism in angiopoietin-2. Cytogenet. Cell Genet. 94, 147–154.
WATSON, C.J., WEBB, N.J., BOTTOMLEY, M.J., and BRENCH-LEY, P.E. (2000). Identification of polymorphisms within the vas-cular endothelial growth factor (VEGF) gene: correlation withvariation in VEGF protein production. Cytokine 12, 1232–1235.
ZHANG, L., YANG, N., PARK, J.W., KATSAROS, D., FRAC-CHIOLI, S., CAO, G., O’BRIEN-JENKINS, A., RANDALL, T.C.,RUBIN, S.C., and COUKOS, G. (2003). Tumor-derived vascularendothelial growth factor up-regulates angiopoietin-2 in host endo-
thelium and destabilizes host vasculature, supporting angiogenesis inovarian cancer. Cancer Res. 63, 3403–3412.