of July 2, 2018.This information is current as Functional Arrest of Immature NK Cells
T Cell Maintenance and a+Impaired CD8 Mice ShownullγDiabetic/SCID/IL-2R
Cell-Engrafted Nonobese Stem+Long-Term Human CD34
Lang, Rupert Handgretinger and Udo F. HartwigMang, Ayline Wilhelm, Ingo Mueller, Wolfgang Herr, PeterSchmauke, Barbara Goecke, Alexander Hohberger, Philippa Maya C. André, Annika Erbacher, Christian Gille, Vanessa
Long-Term Human CD34+ Stem Cell-Engrafted NonobeseDiabetic/SCID/IL-2Rgnull Mice Show Impaired CD8+ T CellMaintenance and a Functional Arrest of Immature NK Cells
Maya C. Andre,* Annika Erbacher,* Christian Gille,† Vanessa Schmauke,*
Barbara Goecke,* Alexander Hohberger,‡ Philippa Mang,* Ayline Wilhelm,*
Ingo Mueller,* Wolfgang Herr,‡ Peter Lang,* Rupert Handgretinger,*,1 and
Udo F. Hartwig‡,1
Allogeneic hematopoietic stem cell transplantation represents the most effective form of immunotherapy for chemorefractory dis-
eases. However, animal models have been missing that allow evaluation of donor-patient–specific graft-versus-leukemia effects.
Thus, we sought to establish a patient-tailored humanized mouse model that would result in long-term engraftment of various
lymphocytic lineages and would serve as a donor-specific surrogate. Following transfer of donor-derived peripheral blood stem-
cells into NOD/SCID/IL-2Rgnull (NSG) mice with supplementation of human IL-7, we could demonstrate robust engraftment and
multilineage differentiation comparable to earlier studies using cord blood stem cells. Phenotypical and functional analyses of
lymphoid lineages revealed that >20 wk posthematopoietic stem cell transplantation, the majority of T lymphocytes consisted of
memory-type CD4+ T cells capable of inducing specific immune functions, whereas CD8+ T cells were only present in low
numbers. Analysis of NSG-derived NK cells revealed the expression of constitutively activated CD56brightCD162 killer Ig-like
receptornegative NK cells that exhibited functional impairments. Thus, the data presented in this study demonstrate that humanized
NSG mice can be successfully used to develop a xenotransplantation model that might allow patient-tailored treatment strategies
in the future, but also highlight the need to improve this model, for example, by coadministration of differentiation-promoting
cytokines and induction of human MHC molecules to complement existing deficiencies in NK and CD8+ T cell development. The
Journal of Immunology, 2010, 185: 2710–2720.
Allogeneic hematopoietic stem cell transplantation (HSCT)using bone marrow (BM) or mobilized peripheral bloodstem cells (PBSCs) represents an effective immunotherapy
for the treatment of high-risk leukemia. Because ,30% of patientshave a HLA-identical donor, transplantation of HLA-incompatiblestem cells remains the therapy of choice (1). However, the extentof HLA mismatches that is beneficial remains a matter of debate.Whereas certain HLA mismatches have been shown to exert po-tent graft-versus-leukemia (GvL) reactivity mediated by alloreac-tive NK cells (2, 3), disparities at single or multiple HLA alleles
increase the risk of severe graft-versus-host disease (4, 5) mediatedby alloreactive donor T cells. T cell depletion is an effective meansfor reducing alloreactivity (6) and is mandatory for donors with onefully mismatched HLA haplotype (7). However, in this haploident-ical setting, rigorous T cell depletion strongly reduces adoptiveimmunity to pathogens and abrogates donor T cell-mediated GvLimmunity, resulting in higher mortality and relapse (8, 9). Therefore,we were interested in developing a donor-patient–specific HSCTxenotransplantation model to study new immunotherapeutic strate-gies for improving donor T lymphocyte and NK cell-mediated GvLimmunity (2, 10, 11) in a HLA-defined situation using long-term(.20 wk) humanized mice.Different immunodeficient mice strains have been increasingly
used to generate surrogate models for engraftment of human he-matopoietic stem cells (HSCs) (12, 13). Recently, NOD.Cg-Prkdcscid IL2rgtmWjl/Sz (NOD/SCID/IL-2Rgnull [NSG]) mice havebeen reported that lack T, B, and NK cells due to a deficientcommon cytokine receptor g-chain, resulting in defective cytokinesignaling of IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21 (14–16). NSGmice demonstrate superior engraftment of human cord blood andadult CD34+ HSCs and multilineage differentiation when com-pared with previously described immunodeficient mouse strains.Transplantation of adult human CD34+ PBSCs into NSG micesupplemented with exogenous human IL-7 has been demonstratedto result in high levels of human CD45+ cells with development ofCD3+ T cells and NK cells, Ig-secreting B cells, myeloid cells,and plasmacytoid dendritic cells (DC) (16). However, there is littleinformation available on the engraftment of adult PBSCs intoNSG mice and the functional properties of human lymphocytesemerging from these HSCTs in long-term engrafted mice. In
*Department of Pediatric Hematology/Oncology and †Department of Neonatology,University Children’s Hospital, Eberhard Karls University, Tuebingen; and ‡Univer-sity Medical Center, 3rd Department of Medicine, Hematology and Oncology,Johannes Gutenberg University, Mainz, Germany
1R.H. and U.F.H. share senior authorship.
Received for publication February 22, 2010. Accepted for publication June 17, 2010.
This work was supported by Grants KFO 183, TP 4, and TP 6 from the DeutscheForschungsgemeinschaft (to R.H., P.L., W.H., and U.F.H.).
Address correspondence and reprint requests to Dr. Rupert Handgretinger or UdoF. Hartwig, Children’s University Hospital, Department of Pediatric Hematology/Oncol-ogy, Hoppe-Seyler-Str.1, D-72076 Tuebingen, Germany (R.H.) and University MedicalCenter, 3rd Department of Medicine, Johannes Gutenberg University, Building for R&D,Obere Zahlbacherstrasse 63, D-55131Mainz, Germany (U.F.H.). E-mail addresses: [email protected] (R.H.) and [email protected] (U.F.H.)
The online version of this article contains supplemental material.
addition, most of the previous studies have been performed withumbilical cord blood (UCB)-derived HSCs, an approach that doesnot allow patient-related studies in a haploidentical setting.We report a comprehensive study to examine the use of adult
PBSCsforestablishingaxenotransplantationmodelthatwouldallowstudies on human GvL reactivity in long-term engrafted recipientsfollowing transfer of leukemia in a donor-patient–specific situa-tion. As the treatment of CD34+ PBSC-engrafted NSG mice withhuman IL-7 was shown to improve the generation and long-termsurvival of human CD3+ T lymphocytes in humanized mice (16)(U.F.H., unpublished results), we included human IL-7 in all ourinvivo studies. Following transplantation ofCD34+-selectedPBSCsinto adult NSG recipients and additional weekly supplementationwith Fc–IL-7, we could demonstrate a robust engraftment efficiencyand multilineage development, although faster engraftment atweek 12 post-HSCT was observed in comparative studies usingCD34+ UCB-HSC–transplanted mice. Phenotypic analyses per-formed at 12–30 wk posttransplantation demonstrated the presenceof early developing monocytes and naive B cells, followed byT cells and NK cells. Surprisingly, the majority of T lymphocytesconsisted of memory-type CD4+ T cells, whereas only smallnumbers of CD8+ T cells were detected in mice.20 wk after trans-plantation, suggesting that IL-7 alone might be insufficient to main-tain peripheral CD8+ T cell homeostasis. Whereas NSG-derivedT cells showed Ag-specific immune responses to human alloanti-gens, NK cells displayed an aberrant phenotype and were function-ally inert.Thus, the data presented in this study demonstrate that long-term
PBSC-engrafted NSG mice can be successfully used to generatea small animal xenograft model for testing feasibility and potentialof individualized patient-tailored treatment strategies, but alsohighlight the need for improvement of this xenotransplantationmodel, for example, by supporting differentiation-promoting cyto-kines and implementing the expression of human MHC moleculesto complement currently existing deficiencies on NK and CD8+
T cell developmental and peripheral homeostatic requirements.
Materials and MethodsMice
NOD.Cg-Prkdcscid IL2rgtmWjl/Sz (also termed NSG), C57BL/6, andBALB/c mice were purchased at The Jackson Laboratory (Bar Harbor,ME) and maintained under specific pathogen-free conditions in the re-search animal facilities of the Universities Tuebingen and Mainz, Ger-many. All approved experimental animal procedures were conductedaccording to German federal and state regulations.
Mobilization, harvest, and isolation of PBSCs
PBSCs were mobilized from parents of children diagnosed with variousmalignancies prior to planned HSCT. Five days prior to PBSC collection,the donor received 10 mg/kg/d G-CSF (Neupogen; Amgen, ThousandOaks, CA) s.c., and leukaphereses were performed on days 5 and 6. Fol-lowing approval by the local ethics committee, parents gave informedconsent to donate ,5% of the PBSCs to be transplanted into NSG miceand to use PBSCs for research purposes in case the patient deceased priorto HSCT. UCB-HSCs from full-term deliveries were collected with in-formed consent and approval by the local ethics committee. CD34+ andCD133+ cells were positively selected from leukapheresis products byMACS with the respective cell isolation kits from Miltenyi Biotec (Ber-gisch Gladbach, Germany). Purity of the cell fractions was evaluated usinga FACSCalibur flow cytometer (BD Biosciences, Heidelberg, Germany).The purity of CD34+ or CD133+ HSC preparations was documented toconsistently be higher than 90% with a contaminating CD3+ T cell contentof 0.14 6 0.1% and a CD56+ NK cell content of 5.3 6 7.3% (see alsoSupplemental Data).
Transplantation of HSCs into NSG mice
Transplantation of adult PBSCs into adult NSG recipients. Eight- to 12-wk-old recipients were sublethally irradiated using a 137Cs irradiator (Gammacell
1000 Elite; MDS Nordion, Fleurus, Belgium) (female NSG mice, 200 cGy;malemice, 250 cGy). Four hours later, recipientswere i.v. injected into the tailvain with 1 3 106 donor-derived CD34+ or CD133+ PBSCs. Reconstitutedmice received weekly i.v. injections of 20 mg Fc–IL-7 fusion protein permouse generously provided byMerck (Darmstadt, Germany) until sacrificed.
Transplantation of neonatal cord blood-derived PBSCs into adult NSGrecipients. Cord blood was collected into heparinized tubes. CD34+
HSCs were isolated using CD34 MicroBeads and MACS technology(Miltenyi Biotec) and stored at 280˚C until use. For transplantation, 1 3106 HSCs were injected i.v. into irradiated mice, as described above,followed by weekly i.v. injections of Fc–IL-7 until mice were sacrificed.
Flow cytometry
After transplantation, peripheral blood, spleen, and BM were collected atdifferent time points, as outlined in results for flow cytometric analyses ofhuman lymphocytes after staining using the following, nonmurine cross-reactive, mAbs: CD62L (clone Dreg56) FITC, CD25 (2A3) allophycocya-nin, CD8 (SU1) PerCP, CD4 (SU3) PerCP, CCR7 (3D12) PECy7, CD28(CD28.2) FITC, CD4 (SK3) FITC, CD3 (SK7) PerCP, CD38 (HIT2)AF700, CD45RO (UCHL1) PE, CD33 (P67.6) PE, CD38 (HB-7) PE,CD34 (8G12) allophycocyanin, IgD (IA6-2) FITC, IgM (G20-127) PE,CD56 (NCAM16.2) FITC, CD56 (My31) PE, CD56 (B159) allophycocya-nin, CD158a (HP-3E4) FITC, CD94 (HP-3D9) FITC, CD45 (HI30)PECy7, and the corresponding isotype controls were all purchased fromBD Pharmingen (Heidelberg, Germany). CD19 (HIB19) allophycocyanin,CD20 (2H7) PerCP, CD45RA (HI100) allophycocyanin, CD45 (HI30) PE,CD4 (RPA-T4) allophycocyanin, and matching isotype controls were fromBiolegend (Munich, Germany). NKp30 (Z25) PE, NKp44 (Z231) PE,NKp46 (BAB281) PE, CD158b (GL183) PE, and NKG2A (Z199) PE,including corresponding isotype control, were from Beckman Coulter(Krefeld, Germany); CD16 (LNK16) AF700 and the corresponding isotypecontrol were from ExBio Praha (Vestec, Czech Republic). CD14 (MEM15)FITC and the corresponding isotype control were purchased from Immu-notools (Friesoythe, Germany); CD158e (DX9) allophycocyanin and iso-type control were purchased from R&D Systems (Wiesbaden, Germany).Flow cytometric analyses were conducted on an LSR II (BD Biosciences)using Diva software following gating of human CD45+ cells.
Isolation of NSG-derived NK cells, CD3+ T cells, and humanPBMCs
CD56+ NK cells and CD3+ T cells were isolated from freshly extractedspleens and BM of engrafted NSG mice by two-step immunomagnetic cellsorting using whole blood CD56 MicroBeads and CD3 MicroBeads, re-spectively (Miltenyi Biotec). Cell purity was determined by FACS analysisto be 90–98%. NKT cell isolation was excluded by flow cytometric anal-ysis of CD56+CD3+ cells. In the majority of cases, the content of CD56+
CD3+ NKT cells was ,1%. In selected cases, in which the NKT contentwas higher than 5%, an additional round of CD3+ positive selection wasperformed. The remaining CD56+CD32 cell fraction was used to isolatethe RNA for the subsequent killer Ig-like receptor (KIR) typing procedure.Human PBMCs were isolated from whole blood using Ficoll-Paque (Bio-chrom, Berlin, Germany) density gradient centrifugation to be used aspositive control in various assays. Washed cells were resuspended inVLE RPMI 1640 (Biochrom), containing 10% FCS (Biochrom), and cul-tured overnight for next day use.
KIR typing
Total RNAwas extracted from NK cells isolated as described above usingthe RNeasy Mini Kit from Qiagen (Hilden, Germany). KIR typing wasperformed by primer-specific detection of KIR gene-transcribed mRNA us-ing the KIR typing kit from Miltenyi Biotec and PCR technology. An am-plified product of a housekeeping gene served as an internal control.
Intracellular IFN-g staining
For in vivo detection of intracellular IFN-g synthesis, splenocytes orhuman PBMCs were cultured for 4 h in the presence of Golgi Plug (BDBiosciences), according to the manufacturer’s instructions. During these4 h, a leukocyte activation mixture (BD Pharmingen) containing PMA andionomycin was used either without or with addition of 100 U/ml IL-2(Proleukin; Novartis, Basel, Switzerland). PBMCs and NSG-derived sple-nocytes were included as positive and negative controls, respectively. Cellswere stained for 20 min with CD56 FITC and CD3 PerCP Ab, permeabi-lized (Fix & Perm Kit; ADG Bio Research, Hamburg, Germany), and incu-bated for 45 min with PE anti–IFN-g (BD Pharmingen) to finally be ana-lyzed by flow cytometry.
CD3+ T cells were isolated from spleens and BM of NSG mice transplantedwith HSCs from one donor or from human PBMCs, as described above.NSG- or C57BL/6 3 BALB/c F1 mice (H-2b3d)-derived DCs were gener-ated from murine BM, as described previously (17). Briefly, femur-derivedBM cells were maintained in culture using RPMI 1640 supplemented with10% FCS, 1% penicillin/streptomycin, and 10% of supernatant obtainedfrom murine GM-CSF producing X6310 cell transfectants for 8 d with50% (v/v) culture medium change every second day. Accordingly, humanHLA-matched and mismatched DCs were generated from PBMCs afterenrichment of monocytes by adherence to plastic and culture in serum-free X-vivo 15 medium (Cambrex Bioscience, Brussels, Belgium) contain-ing 1000 U/ml GM-CSF (Leukomax; Sandoz, Munich, Germany) and 1000U/ml IL-4 (R&D Systems), as described previously (18). MLC of CD3+
responder cells (1 3 105/well) and 1 3 104 irradiated (30 Gy) DCs weremaintained in RPMI 1640 containing 10%FCS, 1% penicillin/streptomycin,and 1% nutrition additives (NCTC-135 medium; Invitrogen, Karlsruhe,Germany) in 96-well triplicates for 5 d. PHA (Sigma-Aldrich, Holzkirchen,Germany) was added to some cultures at 10 and 2.5 mg/ml, respectively, tomeasure alloantigen-independent stimulation. Proliferation of CD3+ T cellswas determined by thymidine uptake (0.5 mCi/well) for the last 15 h ofculture and quantified using a b-plate liquid scintillation counter (Wallac,Turku, Finland). Results represent means of triplicate wells 6 SD.
ELISPOT analysis
IFN-g ELISPOT assays were performed, as recently described (18). Briefly,NSG-derived CD3+ T cells at 5 3 104/well and DCs at 1 3 104/well wereseeded in ELISPOT plates in serum-free X-vivo 20 medium. T cells withoutAPCs or stimulated in the presence of PHA (2.5 mg/ml) served as controls.After 20h incubation at 37˚C, IFN-g spots were visualized and counted usingan Axioplan 2 microscope combined with the computer-assisted image anal-ysis system KS ELISPOT 4.1 (Carl Zeiss Vision, Hallbergmoos, Germany).Results represent means of triplicate wells 6 SD.
Chromium release assay
Splenocytes and BM of up to three humanized NSG mice engrafted withPBSCs of one individual donor were pooled and cultivated overnight. Theerythroleukemia cell line K562 was used as target cells and labeled with50mCi 51Cr (AmershamBiosciences, Freiburg,Germany) for 1.5 h. E:Tratiosvaried between 5:1 and 1:1 and were calculated based on the flow cytometricdetermination of the percentage of CD56+ humanNK cells (varying between1.4 and 3%). The presence of relevant numbers of NKT cells was excluded bystaining forCD56+ andCD3+ cells. LabeledK562 cells and humanPBMCsorpooled murine splenocytes/BM cells were cocultivated for 4 h, in someexperiments with the addition of IL-2 (Proleukin) at a concentration of 100U/ml. Spontaneous release of chromium was ,10% of the maximum releasetaken from the target cells lysed in 1% Triton X-100 (Roche, Mannheim,Germany). Percentage of lysis was calculated as follows: 1003 (experimen-tal release 2 spontaneous release)/(maximum release 2 spontaneous re-lease). Data represent means of quadruplicates.
Pneumococcal immunization (Supplemental Data)
For determination of B cell functionality, humanized NSG mice (week 12–16) were immunized with Pneumovax (Merck) with a dosage adjusted tothe murine body weight (10 ml vaccine diluted in 200 ml PBS s.c.) cor-responding to 0.5 mg each of the 23 different polysaccharide types andboosted with the same dosage 4 wk later. As analysis of pneumococcalserum IgM often results in unreliable data not adequately reflecting thesuccess of a pneumococcal immunization, we determined serum levels ofwhole IgM in NSG mice before immunization, prior to the boost and 2 wkafter the boost using the ELISA for human IgM (Mabtech, Hamburg,Germany), including a human serum probe as internal control.
Statistics
Nonparametric methods were used to compare groups when the distributionof the dependent variable was not normal (Mann-Whitney U test). Resultsare shown as the means 6 SD.
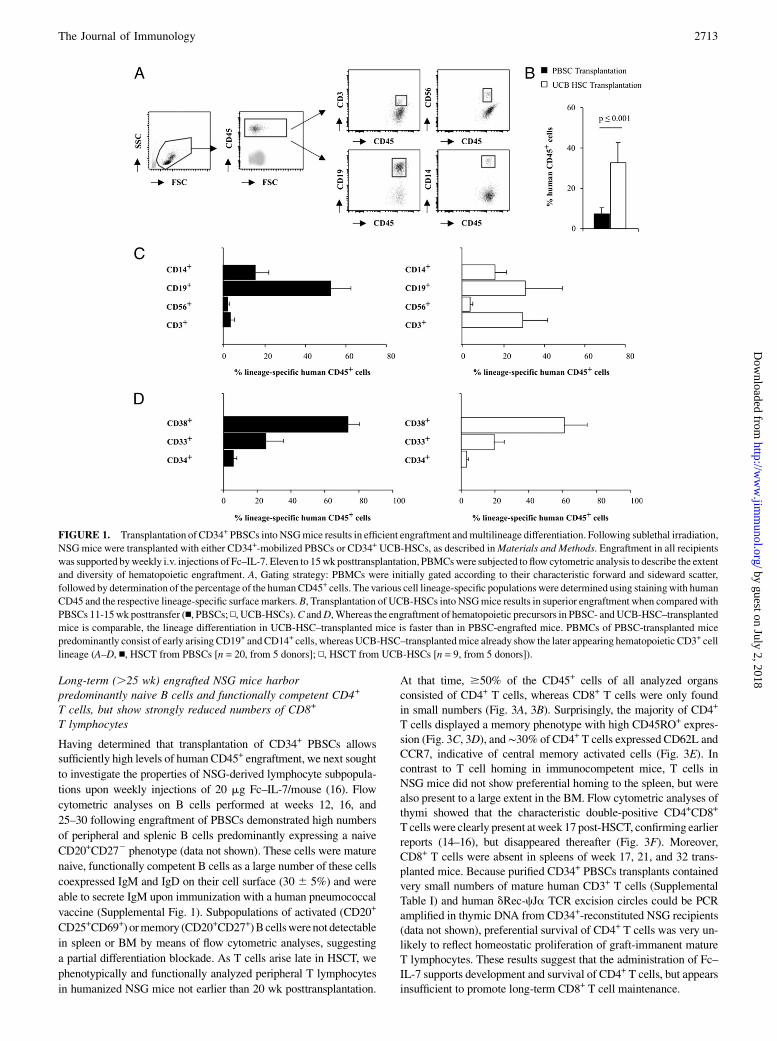
ResultsTransplantation of donor-derived CD34+ PBSCs into NSGmice results in efficient engraftment and multilineagedifferentiation
Because previous experiments were mainly performed with CD34+
UCB-HSCs, we first compared engraftment efficacy and the extent
of lineage differentiation following transplantation of mobilizedCD34+ PBSCs and CD34+ UCB-HSCs into sublethally irradiatedadult NSG recipients. As previous reports (16) and our early stud-ies (data not shown) demonstrated a beneficial effect of humanIL-7 particularly on the generation and peripheral survival of hu-man T lymphocytes in CD34+ PBSC-engrafted mice, we chose toadminister Fc–IL-7 to all experimental groups. Our gating strategywas based on the initial gating on all mononuclear cells (includingmonocytes and granulocytes) according to their characteristic for-ward and sideward scatter and their expression of human CD45+
cells, followed by staining with lineage-specific surface markers(Fig. 1A). This strategy underestimates engraftment rates whencompared with published data often exclusively gating on individ-ual lymphocytic cell populations. However, because HSCT intoNSG also resulted in the generation of a substantial proportion ofmonocytes, this gating strategy appeared to be more precise inestimating the percentage of chimerism. Comparison of engraft-ment rates as determined by the percentage of CD45+ cells atweek 11-15 posttransplantation demonstrated significantly highernumbers of CD45+ cells in mice engrafted with UCB-HSCs thanin mice that had received PBSCs, although the overall absoluteleukocyte count was comparable in both experimental groups(32.7 6 10% versus 7.2 6 3.2%; p # 0.001) (Fig. 1B). In addi-tion, we found a lower amount of early developing CD19+ B cellsand higher numbers of late appearing CD3+ T cells in UCB-HSC–transplanted mice (Fig. 1C), suggesting that transfer of UCB-HSCs results in faster engraftment and lineage differentiation.However, analyses performedat later timepoints (week16–24) showedcomparable levels of CD45+ cells and differentiated cells in bothPBSC- and UCB-HSC–engrafted mice, indicating that the quality ofHSC engraftment ultimately was the same (data not shown). Fre-quencies of CD14+ and CD56+ expressing monocytes or NK cells,respectively, as well as CD33+ and CD38+ cell lineage progeni-tors were comparable in PBSCs and UCB-HSC–transplanted NSGmice (Fig. 1C, 1D).We conclude from these data that transplantation of PBSCs
from adult donors into NSG mice with additional administrationof Fc–IL-7 results in sufficient engraftment and lineage-specificcell differentiation to allow further studies on lymphocyte sub-populations.
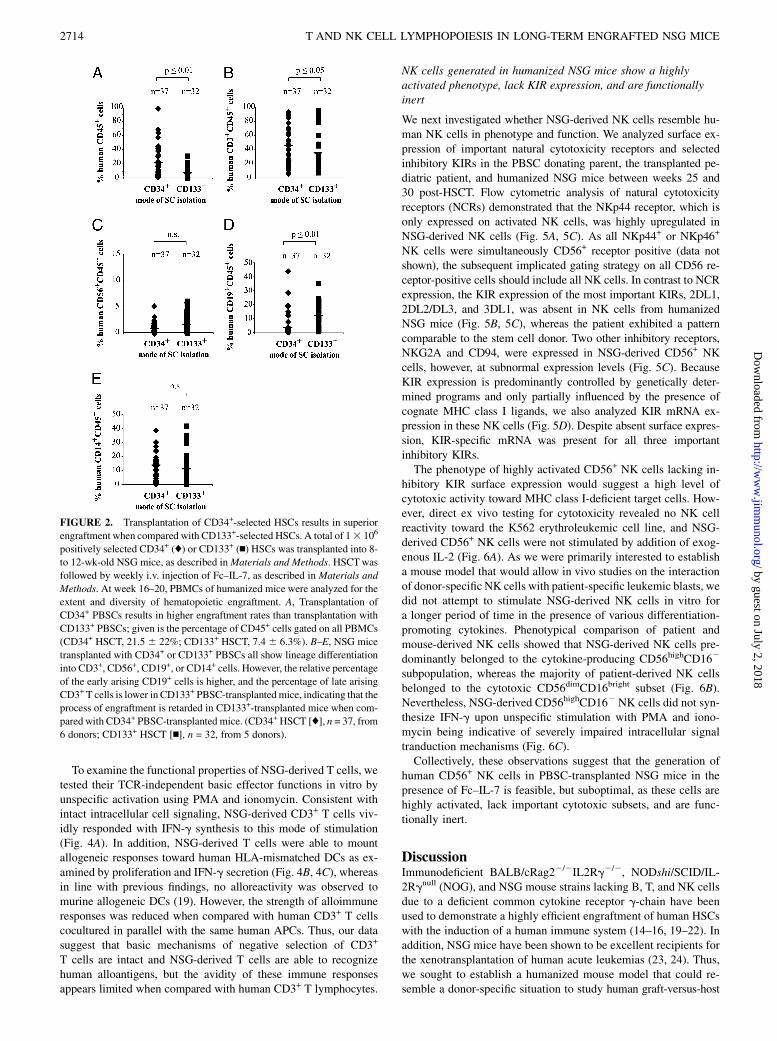
Transplantation of CD34+-selected HSCs results in superiorengraftment when compared with CD133+-selected PBSCs
Because we sought to use this xenotransplantation model asa surrogate for haploidentical HSCT and our patients are routinelytransplanted both with CD34+- and CD133+-selected PBSCs, wealso evaluated the engraftment efficiencies of CD34+ and CD133+
positively selected PBSCs. Engraftment rates obtained in NSGmice transplanted with CD34+ PBSCs were significantly higherthan in mice that received CD133+ PBSCs (CD34+ HSCT, 21.5 622% (n = 37); CD133+ HSCT, 7.4 6 6.3% (n = 32); p # 0.01)(Fig. 2A). Analysis on development of lineage-specific cells dem-onstrated that mice transplanted with CD133+ PBSCs had signif-icantly higher proportions of the early arising CD19+ B cells, butreduced numbers of later evolving CD3+ T cells (Fig. 2B, 2D).Whereas CD14+ cells were detected at comparable percentages inboth groups, CD56+ NK cells were only present in low numbers,indicating that the microenvironment in humanized NSG micemight be suboptimal to support growth and differentiation of NKcells (Fig. 2C, 2E).Thus, HSCT with both CD34+- and CD133+-selected PBSCs is
feasible; however, transplantation with CD34+-selected PBSCsresults in higher engraftment rates and faster lymphoid–lineagedifferentiation than HSCT of CD133+-selected PBSCs.
2712 T AND NK CELL LYMPHOPOIESIS IN LONG-TERM ENGRAFTED NSG MICE
in spleen or BM by means of flow cytometric analyses, suggesting
a partial differentiation blockade. As T cells arise late in HSCT, we
phenotypically and functionally analyzed peripheral T lymphocytes
in humanized NSG mice not earlier than 20 wk posttransplantation.
At that time, $50% of the CD45+ cells of all analyzed organsconsisted of CD4+ T cells, whereas CD8+ T cells were only foundin small numbers (Fig. 3A, 3B). Surprisingly, the majority of CD4+
T cells displayed a memory phenotype with high CD45RO+ expres-sion (Fig. 3C, 3D), and∼30% of CD4+ T cells expressed CD62L andCCR7, indicative of central memory activated cells (Fig. 3E). Incontrast to T cell homing in immunocompetent mice, T cells inNSG mice did not show preferential homing to the spleen, but werealso present to a large extent in the BM. Flow cytometric analyses ofthymi showed that the characteristic double-positive CD4+CD8+
T cells were clearly present atweek 17 post-HSCT, confirming earlierreports (14–16), but disappeared thereafter (Fig. 3F). Moreover,CD8+ T cells were absent in spleens of week 17, 21, and 32 trans-planted mice. Because purified CD34+ PBSCs transplants containedvery small numbers of mature human CD3+ T cells (SupplementalTable I) and human dRec-cJa TCR excision circles could be PCRamplified in thymic DNA from CD34+-reconstituted NSG recipients(data not shown), preferential survival of CD4+ T cells was very un-likely to reflect homeostatic proliferation of graft-immanent matureT lymphocytes. These results suggest that the administration of Fc–IL-7 supports development and survival of CD4+ T cells, but appearsinsufficient to promote long-term CD8+ T cell maintenance.
FIGURE 1. Transplantation of CD34+ PBSCs into NSGmice results in efficient engraftment andmultilineage differentiation. Following sublethal irradiation,
NSGmice were transplanted with either CD34+-mobilized PBSCs or CD34+ UCB-HSCs, as described inMaterials and Methods. Engraftment in all recipients
was supported byweekly i.v. injections of Fc–IL-7. Eleven to 15wk posttransplantation, PBMCswere subjected to flow cytometric analysis to describe the extent
and diversity of hematopoietic engraftment. A, Gating strategy: PBMCs were initially gated according to their characteristic forward and sideward scatter,
followed by determination of the percentage of the humanCD45+ cells. Thevarious cell lineage-specific populations were determined using stainingwith human
CD45 and the respective lineage-specific surfacemarkers.B, Transplantation of UCB-HSCs into NSGmice results in superior engraftment when compared with
PBSCs 11-15wk posttransfer (n, PBSCs; N, UCB-HSCs).C andD,Whereas the engraftment of hematopoietic precursors in PBSC- andUCB-HSC–transplanted
mice is comparable, the lineage differentiation in UCB-HSC–transplanted mice is faster than in PBSC-engrafted mice. PBMCs of PBSC-transplanted mice
predominantly consist of early arising CD19+ andCD14+ cells, whereas UCB-HSC–transplantedmice already show the later appearing hematopoietic CD3+ cell
lineage (A–D, n, HSCT from PBSCs [n = 20, from 5 donors]; N, HSCT from UCB-HSCs [n = 9, from 5 donors]).
To examine the functional properties of NSG-derived T cells, wetested their TCR-independent basic effector functions in vitro byunspecific activation using PMA and ionomycin. Consistent withintact intracellular cell signaling, NSG-derived CD3+ T cells viv-idly responded with IFN-g synthesis to this mode of stimulation(Fig. 4A). In addition, NSG-derived T cells were able to mountallogeneic responses toward human HLA-mismatched DCs as ex-amined by proliferation and IFN-g secretion (Fig. 4B, 4C), whereasin line with previous findings, no alloreactivity was observed tomurine allogeneic DCs (19). However, the strength of alloimmuneresponses was reduced when compared with human CD3+ T cellscocultured in parallel with the same human APCs. Thus, our datasuggest that basic mechanisms of negative selection of CD3+
T cells are intact and NSG-derived T cells are able to recognizehuman alloantigens, but the avidity of these immune responsesappears limited when compared with human CD3+ T lymphocytes.
NK cells generated in humanized NSG mice show a highlyactivated phenotype, lack KIR expression, and are functionallyinert
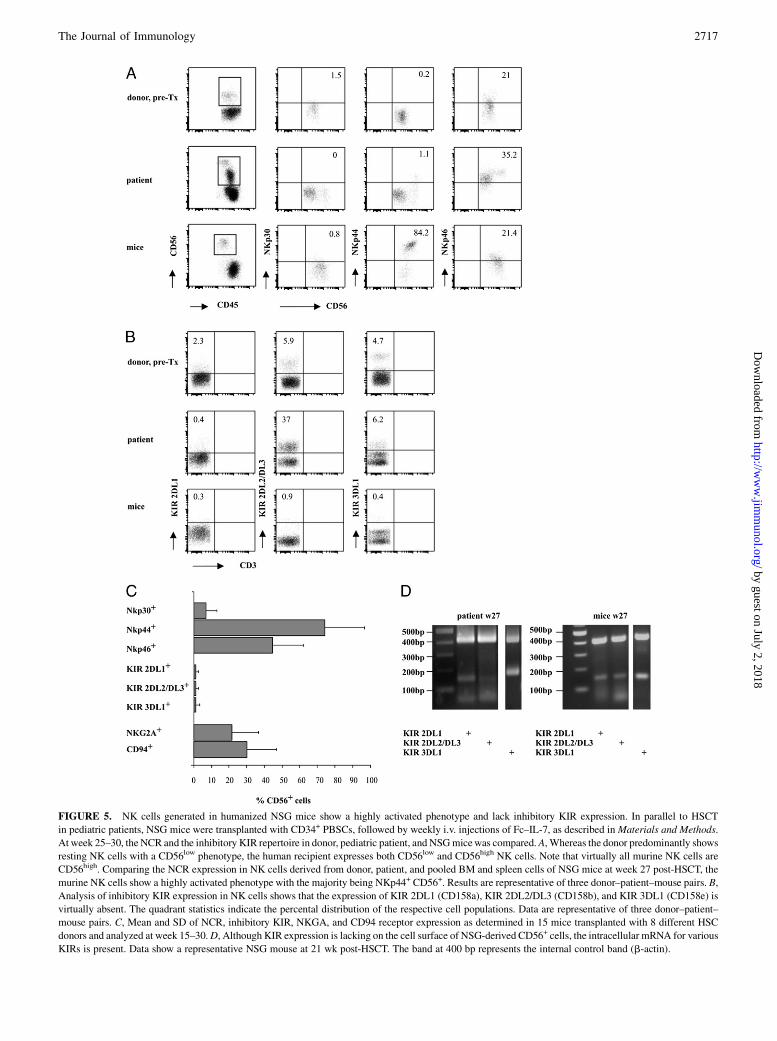
We next investigated whether NSG-derived NK cells resemble hu-man NK cells in phenotype and function. We analyzed surface ex-pression of important natural cytotoxicity receptors and selectedinhibitory KIRs in the PBSC donating parent, the transplanted pe-diatric patient, and humanized NSG mice between weeks 25 and30 post-HSCT. Flow cytometric analysis of natural cytotoxicityreceptors (NCRs) demonstrated that the NKp44 receptor, which isonly expressed on activated NK cells, was highly upregulated inNSG-derived NK cells (Fig. 5A, 5C). As all NKp44+ or NKp46+
NK cells were simultaneously CD56+ receptor positive (data notshown), the subsequent implicated gating strategy on all CD56 re-ceptor-positive cells should include all NK cells. In contrast to NCRexpression, the KIR expression of the most important KIRs, 2DL1,2DL2/DL3, and 3DL1, was absent in NK cells from humanizedNSG mice (Fig. 5B, 5C), whereas the patient exhibited a patterncomparable to the stem cell donor. Two other inhibitory receptors,NKG2A and CD94, were expressed in NSG-derived CD56+ NKcells, however, at subnormal expression levels (Fig. 5C). BecauseKIR expression is predominantly controlled by genetically deter-mined programs and only partially influenced by the presence ofcognate MHC class I ligands, we also analyzed KIR mRNA ex-pression in these NK cells (Fig. 5D). Despite absent surface expres-sion, KIR-specific mRNA was present for all three importantinhibitory KIRs.The phenotype of highly activated CD56+ NK cells lacking in-
hibitory KIR surface expression would suggest a high level ofcytotoxic activity toward MHC class I-deficient target cells. How-ever, direct ex vivo testing for cytotoxicity revealed no NK cellreactivity toward the K562 erythroleukemic cell line, and NSG-derived CD56+ NK cells were not stimulated by addition of exog-enous IL-2 (Fig. 6A). As we were primarily interested to establisha mouse model that would allow in vivo studies on the interactionof donor-specific NK cells with patient-specific leukemic blasts, wedid not attempt to stimulate NSG-derived NK cells in vitro fora longer period of time in the presence of various differentiation-promoting cytokines. Phenotypical comparison of patient andmouse-derived NK cells showed that NSG-derived NK cells pre-dominantly belonged to the cytokine-producing CD56highCD162
subpopulation, whereas the majority of patient-derived NK cellsbelonged to the cytotoxic CD56dimCD16bright subset (Fig. 6B).Nevertheless, NSG-derived CD56highCD162 NK cells did not syn-thesize IFN-g upon unspecific stimulation with PMA and iono-mycin being indicative of severely impaired intracellular signaltranduction mechanisms (Fig. 6C).Collectively, these observations suggest that the generation of
human CD56+ NK cells in PBSC-transplanted NSG mice in thepresence of Fc–IL-7 is feasible, but suboptimal, as these cells arehighly activated, lack important cytotoxic subsets, and are func-tionally inert.
DiscussionImmunodeficient BALB/cRag22/2IL2Rg2/2, NODshi/SCID/IL-2Rgnull (NOG), and NSG mouse strains lacking B, T, and NK cellsdue to a deficient common cytokine receptor g-chain have beenused to demonstrate a highly efficient engraftment of human HSCswith the induction of a human immune system (14–16, 19–22). Inaddition, NSG mice have been shown to be excellent recipients forthe xenotransplantation of human acute leukemias (23, 24). Thus,we sought to establish a humanized mouse model that could re-semble a donor-specific situation to study human graft-versus-host
FIGURE 2. Transplantation of CD34+-selected HSCs results in superior
engraftment when compared with CD133+-selected HSCs. A total of 13 106
positively selected CD34+ (♦) or CD133+ (n) HSCs was transplanted into 8-
to 12-wk-old NSG mice, as described inMaterials and Methods. HSCTwas
followed by weekly i.v. injection of Fc–IL-7, as described in Materials and
Methods. At week 16–20, PBMCs of humanized mice were analyzed for the
extent and diversity of hematopoietic engraftment. A, Transplantation of
CD34+ PBSCs results in higher engraftment rates than transplantation with
CD133+ PBSCs; given is the percentage of CD45+ cells gated on all PBMCs
and GvL immunobiology in vivo and potentially serve as a preclin-ical model to test new individualized concepts of immunotherapeu-tic strategies upon cotransfer of patient-specific leukemia in thefuture.Most humanized mouse models reported to date have been
engrafted with UCB-HSCs, an approach that is not well suitable forHLA-defined in vivo studies in a donor-patient–specific xenotrans-plantation model. In addition, because adoptive transfer of humanPBMCs results in xenogeneic graft-versus-host disease in thesemice (25), studying GvL-specific responses using this approachwill be difficult. We, therefore, transplanted donor-derived CD34+
PBSCs into NSG mice to develop a xenograft model that wouldallow studies on long-term T and NK cell biology as a surrogate tohaploidentical HSCTs in pediatric patients performed with CD34+
HSC from the same donor.Previous work on human lymphocytes generated in NSG or
NOG mice has largely been conducted without supplementinghuman cytokines (15, 20, 26–29). However, addition of exogenoushuman IL-7 to CD34+ PBSC-engrafted NSG mice was shown toimprove development of human T lymphocytes (16). Moreover,because IL-7 is instrumental in T, B, and DC development (30–32)and plays a pivotal role in homeostatic proliferation of T cells(33), we did not compare engraftment efficiency and lineage dif-ferentiation on human HSC-engrafted mice in the presence or
absence of human IL-7, but administered Fc–IL-7 at weekly inter-vals following HSCT and examined the phenotype and function-ality of lymphocytes from NSG mice over 20–30 wk.Comparative analyses on engraftment rates and efficiency upon
transplantation of PBSCs or UCB-HSCs under these conditionsrevealed that transfer of UCB-HSCs results in faster engraftmentand generation of human CD45+ cells when compared with PBSC-engrafted NSG mice 12 wk after HSCT. Because early developingCD19+ B cells were found to be decreased, but late appearing CD3+
T cells increased, this might indicate a faster differentiation potentialof UCB-HSCs. These differences were not due to differential leuko-cyte counts that were found to be comparable in both experimentalgroups.Moreover, lineage-specific CD33+ and CD38+ progenitors aswell as monocytes and NK cells were present in comparable num-bers, and the amount of lymphocytes in PBSC- or UCB-HSC–engrafted mice reached comparable levels at week 16 posttransfer.Thus, transplantation of donor-derivedCD34+ PBSCs intoNSGmiceproved to be sufficient to allow detailed phenotypical and functionalanalyses of human lymphocytes in long-term transplanted mice.Because we perform pediatric HSCTs using both CD34+- and
CD133+-enriched HSCs, we next examined engraftment efficien-cies of these stem cell subsets. Interestingly, transplantation ofCD34+ HSCs in NSG mice resulted in significantly higher engraft-ment rates and faster lineage differentiation than transplantation of
FIGURE 3. Long-term (.25 wk) engrafted NSG mice harbor phenotypically normal CD4+ T cells, whereas the number of CD8+ T cells is strongly
decreased. NSG mice were transplanted, as described in Materials and Methods, followed by weekly injections of Fc–IL-7. PBMCs, BM, and spleen cells
of long-term engrafted mice were subjected to flow cytometric analysis to describe the extent and diversity of T cell lineage differentiation. A and B, More
than 50% of the CD45+ cells of all analyzed organs consist of CD4+ T cells, whereas CD8+ T cells are only found in small numbers. Note that NSG-derived
T cells do not show preferential homing to the spleen, but are also present to a large extent in the BM (n, PBMCs; gray bar, BM; N, spleen). C and D, Ten to
15% of the CD4+ T cells are naive CD45RA+ T cells, whereas the majority of CD4+ T cells display a memory T cell phenotype with high CD45RO+
expression. E, Thirty percent of the CD4+ T cells show a central memory phenotype as determined by CD62L and CCR7 expression. Data are representative
of five mice transplanted with four different HSC donors analyzed at week 27–32 post-HSCT. F, Double-positive CD4+CD8+ T cells are present in the
thymus of a week 17 transplanted NSG mouse, whereas thymi of NSG mice at week 21 and 32 post-HSCT lack characteristic double-positive CD4+CD8+
T cells. Splenocytes of the identical mice at weeks 17, 21, and 32 do not display any CD8+ T cells. Thymi and splenocytes were stained for CD3+, CD4+,
and CD8+ expression in NSG mice at week 17, 21, and 32 post-HSCT. CD4+ and CD8+ expression was determined following gating on all CD3+ T cells.
The quadrant statistics indicate the percental distribution of the respective cell populations.
CD133+ HSCs. Gene expression profiling studies have attributeddiffering roles to CD34+ and CD133+ HSCs (34) in the processof cell cycle, maintenance of chromatin architecture, and DNAmetabolism, and in line with this, our results are compatible witha rather undifferentiated nature of CD133+ HSCs.
Collectively, our data demonstrate that transplantation of CD34+
PBSCs into adult NSG mice with additional Fc–IL-7 administra-tion can be efficiently used to generate myeloid and lymphoidlineages.Surprisingly, phenotypic analyses of these lymphoid populations
25–30 wk following HSCT revealed that the majority of T cellsfound in PBSC-engrafted NSG mice consisted of CD4+ T cells,whereas only small numbers of CD8+ T cells were found. More-over, most CD4+ T cells expressed a CD45RO+ memory pheno-type. Although we cannot completely exclude any engraftmentof mature T cells persisting in the graft in such highly permis-sive mice, preferential expansion and survival of mature CD4+
T lymphocytes reflecting this phenotype appear very unlikely giventhe small numbers of CD3+ T lymphocytes cotransplanted and thepresence of dRec-cJa TCR excision circles detected in the thymusof CD34+ HSC-engrafted NSG recipients, as shown before (16). AsIL-7 mediates its effect primarily on naive and memory T cells(35), the repetitive Fc–IL-7 injections may at least partially accountfor the high percentage of memory T cells in our mice. BecauseCD4+CD8+ double-positive thymocytes and CD8+ T cells weredetected in thymi of humanized NSG mice up to 17 wk post-HSCT, but were absent at later stages of HSCT, we assume thatadministration of IL-7 alone is in fact sufficient to maintain a poolof polyclonal CD4+, but not of CD8+ T cells. Along this line, resultsfrom our group on better engraftment of purified donor CD8+
T cells or leukemia-reactive donor CD8+ T cell lines in the pres-ence of IL-15 support these data (M. Nonn, J. Knapstein, S.A.Khan, M. Horz, E. Distler, A. Brunk, M.C. Ande, R. Handgretinger,M. Theobald, W. Herr, and U.F. Hartwig, submitted for publica-tion). In addition to IL-7, survival and homeostatic proliferation ofCD8+ T cells are promoted by IL-15 (33), and recent results showimproved CD8+ T cell development in Rag22/2IL2Rg2/2 micewhen IL-15 is presented in trans (36). Moreover, a lack of T cellmaintenance (37) and increased CD95 (Fas)-mediated susceptibil-ity of CD8+ T cells to undergo apoptosis have been described inhumanized mice (20), possibly reflecting anti-apoptotic effects ofIL-7 and IL-15 (33). Because TCR-driven signaling provided byhuman MHC class I-restricted recognition of Ags might furtherpromote survival and homeostasis of CD8+ T cells in humanizedNSG mice (21), we thus assume that the suboptimal cytokine mi-lieu and weak TCR–MHC interactions due to insufficient humanMHC class I expression on a limited number of human APCs inlymphoid compartments might account for the reduced life span ofCD8+ T cells in our long-term engrafted NSG mice (38). Becauseour T cell analyses in humanized mice were performed at later timepoints than previously described results (16, 20), it is difficult tocompare our findings with these data.In contrast to CD8+ T cells, mature naive B cells were found in
large numbers in humanized NSG mice and were able to secreteIgM upon immunization, confirming previous findings (20). Thus,despite the fact that humanized NSG mice lack activated (CD20+
CD25+CD69+) or memory (CD20+CD27+) B cells, the suggestedpartial differentiation blockade (20) of B cell-lineage cells appearsto be limited to later stages of B cell differentiation.Despite low numbers of CD8+ T lymphocytes, functional analyses
of NSG-derived CD3+ T cells demonstrated increased IFN-g syn-thesis upon unspecific stimulation, indicating intact basic signaltransduction mechanisms. Moreover, T cells proliferated vigorouslyand exhibited effector functions when stimulated with allogeneichuman DCs, whereas no alloimmunity was observed following co-culture with MHC-matched murine (NSG) or allogeneic (F1) DCs,as reported earlier (19). Hence, our results support emerging data inhumanized mice, which demonstrate positive selection of humanHSC-derived T cells by murine thymic epithelial cells, followed by
FIGURE 4. CD3+ T cells generated in long-term (.25 wk) transplanted
NSG mice display effector functions to human, but not to murine alloanti-
gens. A, NSG-derived CD3+ T cells are able to respond with IFN-g synthesis
to unspecificT cell stimulation. For these invitro experiments, splenocytes of
humanizedNSGmicewere cultivatedwith PMA/Iono. or PMA/Iono. + IL-2,
followed by intracellular IFN-g staining, as described in Materials and
Methods. A, Shows the flow cytometric data of one mouse representative
of three independent experiments with a total of six mice. Included is the
internal IFN-g control staining of PBMCs of a healthy donor. The quadrant
statistics indicate the percental distribution of the respective cell populations.
B, CD3+ T cells retain their capability to recognize Ag in the context of
human HLA molecules. Pooled CD3+ T cells from PBMCs, spleen, and
BM were stimulated with either irradiated MHC-matched murine NSG-
derived or MHC-mismatched human or murine F1 DCs, as described in
Materials and Methods, and tested for their proliferative alloresponses de-
termined by [3H]thymidine incorporation. NSG-derived T cells (n) show
alloimmune reactions to human DCs, but not to MHC-matched murine
NSG-derived DCs or allogeneic (F1) murine DCs. Included are proliferative
responses of human CD3+ T cells (n) toward murine or human DCs as an
internal standard. Note that the extent of alloreactive T cell proliferation is
reduced when compared with human control PBMCs. PHA-induced T cell
activation was used as a positive control, as described in Materials and
Methods. C, NSG-derived T cells display alloreactive effector functions
against human MHC-mismatched DCs. Alloimmune responses were deter-
mined by IFN-g ELISPOT assay, as described in Materials and Methods.
Data are representative of three experiments with a total of nine mice at 25–
30 wk post-HSCT. PMA/Iono., PMA/ionomycin.
2716 T AND NK CELL LYMPHOPOIESIS IN LONG-TERM ENGRAFTED NSG MICE
FIGURE 5. NK cells generated in humanized NSG mice show a highly activated phenotype and lack inhibitory KIR expression. In parallel to HSCT
in pediatric patients, NSG mice were transplanted with CD34+ PBSCs, followed by weekly i.v. injections of Fc–IL-7, as described inMaterials and Methods.
At week 25–30, the NCR and the inhibitory KIR repertoire in donor, pediatric patient, and NSGmicewas compared. A, Whereas the donor predominantly shows
resting NK cells with a CD56low phenotype, the human recipient expresses both CD56low and CD56high NK cells. Note that virtually all murine NK cells are
CD56high. Comparing the NCR expression in NK cells derived from donor, patient, and pooled BM and spleen cells of NSG mice at week 27 post-HSCT, the
murine NK cells show a highly activated phenotype with the majority being NKp44+ CD56+. Results are representative of three donor–patient–mouse pairs. B,
Analysis of inhibitory KIR expression in NK cells shows that the expression of KIR 2DL1 (CD158a), KIR 2DL2/DL3 (CD158b), and KIR 3DL1 (CD158e) is
virtually absent. The quadrant statistics indicate the percental distribution of the respective cell populations. Data are representative of three donor–patient–
mouse pairs. C, Mean and SD of NCR, inhibitory KIR, NKGA, and CD94 receptor expression as determined in 15 mice transplanted with 8 different HSC
donors and analyzed at week 15–30.D, Although KIR expression is lacking on the cell surface of NSG-derived CD56+ cells, the intracellular mRNA for various
KIRs is present. Data show a representative NSG mouse at 21 wk post-HSCT. The band at 400 bp represents the internal control band (b-actin).
negative selection on both thymic epithelial cells andDCs of murineand human origin (13, 39) to enable recognition of human MHCmolecules as alloantigens. Because the MHC haplotypes of DCsfrom F1 and NSG mice are quite similar, this may have contributedto the suboptimal immune responses to murine alloantigens. Finally,consistent with previous reports obtained in NOG mice (20), theextent of immune responses elicited by NSG-derived T cellsappeared less avid when compared with human CD3+ T cells stim-ulated with the same allostimulator cells. This result further indi-cates that human T cell immunity in NSG mice might be improvedupon additional cytokine support and expression of human MHCmolecules.
In summary, our data suggest that Fc–IL-7 administration de-cidedly promotes B cell precursor and both CD4+ T cell develop-ment and differentiation toward a memory phenotype; however,Ag-specific T cell immune responses occur at a reduced efficiencywhen compared with human T lymphocytes.In addition to T cells, we examined the phenotype and function
of NSG-derived NK cells. To our knowledge, this is the first studyinvestigating human NK cell immunity in humanized mice by com-paring the nature of NK cells from transplanted patients with re-constituted NSG recipients. Flow cytometric analyses of patient-derived NK cells demonstrated the presence of NK cells withlow levels of activation and high inhibitory KIR 2DL2/DL3 expres-sion as frequently observed during the early posttransplantation pe-riod in patients (R.H., unpublished observation). In contrast, NSG-derived NK cells demonstrated signs of constitutive activation withhigh levels of NKp44 expression together with subnormal levels ofinhibitory KIRs and selected inhibitory lectin-like receptors, suchas CD94 and NKG2A. Moreover, the important cytotoxic NK cellsubset of CD56dimCD16+ NK cells that was abundantly expressedin transplanted patients was virtually absent in NSG mice. Be-cause adoptive transfer of donor NK cells into irradiated NSGmice supplemented with IL-7 did neither result in engraftmentnor expansion of NK cells up to 20 wk postinfusion (our unpub-lished data) and NSG-derived NK cells did not display a donor-specific phenotype, we conclude that the observed immature,functionally inert CD56+CD162KIR2 NK subpopulation did notresult from cotransfer of graft-immanent NK cells, but was indeedHSC derived. Moreover, our results are supported by Huntingtonet al. (36), who recently reported that NK cells generated in hu-manized Rag22/2IL2Rg2/2 mice with additional administrationof IL-15 linked to its IL-15R a-chain developmentally progressedfrom a CD56highCD162KIR2 phenotype to a more matureCD56lowCD16+KIR+ phenotype. Thus, the phenotypical aberra-tions observed in our NSG mice may in fact result from a matu-rational arrest of NK cells due to lack of human IL-15.Functional analyses of NSG-derived NK cells demonstrated that
NK cells isolated from Fc–IL-7–supplemented NSG mice are func-tionally inert and do neither display signs of intracellular IFN-gcytokine synthesis upon unspecific cell stimulation nor showin vitro cytotoxic activity against the K562 erythroleukemia cellline. Thus, our observations thatNSG-derivedNKcells lack essentialinhibitory receptors, but are at the same time unable to exert NK cell-specific cytotoxicity, are consistent with reports that NK cells fromMHC class I-deficient animals or humans may be tolerant as a resultof induced hyporesponsiveness (40, 41). As described by the theoryof licensing, KIRs on NK cells are able to exhibit differential effectsduring endogenousmaturation uponMHCclass I interaction (42, 43).In line with this, studies on fully MHC-mismatched BALB/c→B6chimeras have demonstrated that both donor and host NK cells be-come specifically tolerant of donor and host Ags, whereas alloreac-tivity against b2-microglobulin2/2 BM cells is preserved (44). Incontrast, host NK cells in a rat→mouse xenotransplantation modelare rendered functionally anergic and lack important NK cell cyto-toxic functions similar to NK cell-deficient Ly49A transgenic mice(45). Thus, it is conceivable that in addition to a lack of supportivecytokines, an inadequate MHC class I-KIR interaction affectedNK cell ontogeny and resulted in a maturational arrest in our xeno-transplantation model.In summary, our phenotypical and functional data are consis-
tent with a model suggesting that cytotoxic CD56lowCD16+KIR+
NK cells represent the terminal stage of NK cell differentiationnot yet achieved in our xenotransplantation model.In conclusion, our results suggest that the establishment of a
patient-specific xenotransplantation model following engraftment
FIGURE 6. NSG-derived CD56+ NK cells are functionally inert. A, NK
cells derived from NSG mice (week 16–25) lack cytotoxic activity against
the NK-sensitive erythroleukemic K562 cell line. Cytotoxicity was deter-
mined using a 4-h 51Cr release assay, as described in Materials and Meth-
ods. Data are representative of three independent experiments with pooled
cells of one to three mice (when mice had been transplanted with the same
SC donor). B, CD16 expression in NSG-derived CD56+ NK cells is low.
Note that the transplanted patient shows two NK cell subsets, with the
majority of NK cells being CD56dimCD16+. In contrast, murine NK cells
exclusively belong to the CD56high subpopulation and are CD162. C,
NSG-derived CD56+ NK cells are unable to respond with IFN-g synthe-
sis to unspecific NK cell stimulation. For these in vitro experiments,
splenocytes of humanized NSG mice were cultivated with PMA/Iono. or
PMA/Iono. + IL-2, followed by intracellular IFN-g staining, as described in
Materials and Methods. C, Shows the flow cytometric data of one mouse
representative of three independent experiments with a total of four mice.
Included is the internal IFN-g control staining of PBMCs of a healthy
donor. The quadrant statistics indicate the percental distribution of the
of donor-derived CD34+ PBSCs with the generation of a multi-lineage human immune system in NSG mice is feasible, but hasseveral limitations that need to be optimized. We, therefore, pro-pose that introduction of MHC molecules possibly expressed ona murine MHC-deficient background in addition to the continuoussupply of growth, homeostasis, and differentiation-promoting cyto-kines may refine the generation of long-term humanized mice notonly to evaluate new forms of patient-tailored immunotherapy inHSCT, but also to improve, for example, HIV- and Ab-based can-cer therapy studies in these mice.
AcknowledgmentsWe thank the stem cell laboratory of the University Children’s Hospital
(Tuebingen, Germany) for providing the donor-derived PBSCs. We also
thank Dr. W. Strittmatter, Merck (Darmstadt, Germany), for generously
providing the human Fc–IL-7.
DisclosuresThe authors have no financial conflicts of interest.
References1. Petersdorf, E. W. 2004. HLA matching in allogeneic stem cell transplantation.
Curr. Opin. Hematol. 11: 386–391.2. Ruggeri, L., M. Capanni, E. Urbani, K. Perruccio, W. D. Shlomchik, A. Tosti,
S. Posati, D. Rogaia, F. Frassoni, F. Aversa, et al. 2002. Effectiveness of donornatural killer cell alloreactivity in mismatched hematopoietic transplants. Sci-ence 295: 2097–2100.
3. Bethge, W. A., C. Faul, M. Bornhauser, G. Stuhler, D. W. Beelen, P. Lang,M. Stelljes, W. Vogel, M. Hagele, R. Handgretinger, and L. Kanz. 2008. Hap-loidentical allogeneic hematopoietic cell transplantation in adults using CD3/CD19 depletion and reduced intensity conditioning: an update. Blood Cells Mol.Dis. 40: 13–19.
4. Sasazuki, T., T. Juji, Y. Morishima, N. Kinukawa, H. Kashiwabara, H. Inoko,T. Yoshida, A. Kimura, T. Akaza, N. Kamikawaji, et al. 1998. Effect of matchingof class I HLA alleles on clinical outcome after transplantation of hematopoieticstem cells from an unrelated donor: Japan Marrow Donor Program. N. Engl. J.Med. 339: 1177–1185.
5. Petersdorf, E. W., J. A. Hansen, P. J. Martin, A. Woolfrey, M. Malkki, T. Gooley,B. Storer, E. Mickelson, A. Smith, and C. Anasetti. 2001. Major-histocompatibility-complex class I alleles and antigens in hematopoietic-celltransplantation. N. Engl. J. Med. 345: 1794–1800.
6. Ho, V. T., and R. J. Soiffer. 2001. The history and future of T-cell depletion asgraft-versus-host disease prophylaxis for allogeneic hematopoietic stem celltransplantation. Blood 98: 3192–3204.
7. Aversa, F., A. Tabilio, A. Velardi, I. Cunningham, A. Terenzi, F. Falzetti,L. Ruggeri, G. Barbabietola, C. Aristei, P. Latini, et al. 1998. Treatment of high-risk acute leukemia with T-cell-depleted stem cells from related donors with onefully mismatched HLA haplotype. N. Engl. J. Med. 339: 1186–1193.
8. Handgretinger, R., M. Schumm, P. Lang, J. Greil, A. Reiter, P. Bader,D. Niethammer, and T. Klingebiel. 1999. Transplantation of megadoses of purifiedhaploidentical stem cells. Ann. NY Acad. Sci. 872: 351–362.
9. Kato, S., H. Yabe, M. Yasui, K. Kawa, T. Yoshida, A. Watanabe, Y. Osugi,K. Horibe, and Y. Kodera. 2000. Allogeneic hematopoietic transplantation ofCD34+ selected cells from an HLA haplo-identical related donor: a long-termfollow-up of 135 patients and a comparison of stem cell source between the bonemarrow and the peripheral blood. Bone Marrow Transplant. 26: 1281–1290.
10. Wehler, T. C., M. Nonn, B. Brandt, C. M. Britten, M. Grone, M. Todorova,I. Link, S. A. Khan, R. G. Meyer, C. Huber, et al. 2007. Targeting the activation-induced antigen CD137 can selectively deplete alloreactive T cells from anti-leukemic and antitumor donor T-cell lines. Blood 109: 365–373.
11. Kabelitz, D., W. R. Herzog, B. Zanker, and H. Wagner. 1985. Human cytotoxicT lymphocytes. I. Limiting-dilution analysis of alloreactive cytotoxic T-lymphocyte precursor frequencies. Scand. J. Immunol. 22: 329–335.
12. Shultz, L. D., F. Ishikawa, and D. L. Greiner. 2007. Humanized mice in trans-lational biomedical research. Nat. Rev. Immunol. 7: 118–130.
13. Manz, M. G. 2007. Human-hemato-lymphoid-system mice: opportunities andchallenges. Immunity 26: 537–541.
14. Ito, M., H. Hiramatsu, K. Kobayashi, K. Suzue, M. Kawahata, K. Hioki,Y. Ueyama, Y. Koyanagi, K. Sugamura, K. Tsuji, et al. 2002. NOD/SCID/gamma(c)(null) mouse: an excellent recipient mouse model for engraftment ofhuman cells. Blood 100: 3175–3182.
15. Ishikawa, F., M. Yasukawa, B. Lyons, S. Yoshida, T. Miyamoto, G. Yoshimoto,T. Watanabe, K. Akashi, L. D. Shultz, and M. Harada. 2005. Development offunctional human blood and immune systems in NOD/SCID/IL2 receptorgamma chain(null) mice. Blood 106: 1565–1573.
16. Shultz, L. D., B. L. Lyons, L. M. Burzenski, B. Gott, X. Chen, S. Chaleff,M. Kotb, S. D. Gillies, M. King, J. Mangada, et al. 2005. Human lymphoid andmyeloid cell development in NOD/LtSz-scid IL2R gamma null mice engraftedwith mobilized human hemopoietic stem cells. J. Immunol. 174: 6477–6489.
17. Lutz, M. B., N. Kukutsch, A. L. Ogilvie, S. Rossner, F. Koch, N. Romani, andG. Schuler. 1999. An advanced culture method for generating large quantities ofhighly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 223:77–92.
18. Hartwig, U. F., M. Nonn, S. Khan, I. Link, C. Huber, and W. Herr. 2008. De-pletion of alloreactive donor T lymphocytes by CD95-mediated activation-induced cell death retains antileukemic, antiviral, and immunoregulatoryT cell immunity. Biol. Blood Marrow Transplant. 14: 99–109.
19. Traggiai, E., L. Chicha, L. Mazzucchelli, L. Bronz, J. C. Piffaretti,A. Lanzavecchia, and M. G. Manz. 2004. Development of a human adaptiveimmune system in cord blood cell-transplanted mice. Science 304: 104–107.
20. Watanabe, Y., T. Takahashi, A. Okajima, M. Shiokawa, N. Ishii, I. Katano, R. Ito,M. Ito, M. Minegishi, N. Minegishi, et al. 2009. The analysis of the functions ofhuman B and T cells in humanized NOD/shi-scid/gammac(null) (NOG) mice(hu-HSC NOG mice). Int. Immunol. 21: 843–858.
21. Strowig, T., C. Gurer, A. Ploss, Y. F. Liu, F. Arrey, J. Sashihara, G. Koo,C. M. Rice, J. W. Young, A. Chadburn, et al. 2009. Priming of protective T cellresponses against virus-induced tumors in mice with human immune systemcomponents. J. Exp. Med. 206: 1423–1434.
22. Hiramatsu, H., R. Nishikomori, T. Heike, M. Ito, K. Kobayashi, K. Katamura,and T. Nakahata. 2003. Complete reconstitution of human lymphocytes fromcord blood CD34+ cells using the NOD/SCID/gammacnull mice model. Blood102: 873–880.
23. Agliano, A., I. Martin-Padura, P. Mancuso, P. Marighetti, C. Rabascio,G. Pruneri, L. D. Shultz, and F. Bertolini. 2008. Human acute leukemia cellsinjected in NOD/LtSz-scid/IL-2Rgamma null mice generate a faster and moreefficient disease compared to other NOD/scid-related strains. Int. J. Cancer 123:2222–2227.
24. Sanchez, P. V., R. L. Perry, J. E. Sarry, A. E. Perl, K. Murphy, C. R. Swider,A. Bagg, J. K. Choi, J. A. Biegel, G. Danet-Desnoyers, and M. Carroll. 2009. Arobust xenotransplantation model for acute myeloid leukemia. Leukemia 23:2109–2117.
25. King, M. A., L. Covassin, M. A. Brehm, W. Racki, T. Pearson, J. Leif, J. Laning,W. Fodor, O. Foreman, L. Burzenski, et al. 2009. Human peripheral bloodleucocyte non-obese diabetic-severe combined immunodeficiency interleukin-2receptor gamma chain gene mouse model of xenogeneic graft-versus-host-likedisease and the role of host major histocompatibility complex. Clin. Exp. Immu-nol. 157: 104–118.
26. Noort, W. A., J. Wilpshaar, C. D. Hertogh, M. Rad, E. G. Lurvink, S. A. vanLuxemburg-Heijs,K.Zwinderman,R.A.Verwey,R.Willemze, and J.H.Falkenburg.2001. Similar myeloid recovery despite superior overall engraftment in NOD/SCIDmice after transplantation of human CD34+ cells from umbilical cord blood ascompared to adult sources. Bone Marrow Transplant. 28: 163–171.
27. Lepus, C. M., T. F. Gibson, S. A. Gerber, I. Kawikova, M. Szczepanik,J. Hossain, V. Ablamunits, N. Kirkiles-Smith, K. C. Herold, R. O. Donis, et al.2009. Comparison of human fetal liver, umbilical cord blood, and adult bloodhematopoietic stem cell engraftment in NOD-scid/gammac2/2, Balb/c-Rag12/2
gammac2/2, and C.B-17-scid/bg immunodeficient mice. Hum. Immunol. 70:790–802.
28. Yahata, T., K. Ando, Y. Nakamura, Y. Ueyama, K. Shimamura, N. Tamaoki,S. Kato, and T. Hotta. 2002. Functional human T lymphocyte development fromcord blood CD34+ cells in nonobese diabetic/Shi-scid, IL-2 receptor gamma nullmice. J. Immunol. 169: 204–209.
29. Giassi, L. J., T. Pearson, L. D. Shultz, J. Laning, K. Biber, M. Kraus, B. A. Woda,M. R. Schmidt, R. T. Woodland, A. A. Rossini, and D. L. Greiner. 2008. Ex-panded CD34+ human umbilical cord blood cells generate multiple lymphohe-matopoietic lineages in NOD-scid IL2rgamma(null) mice. Exp. Biol. Med. 233:997–1012.
30. Di Santo, J. P., and H. R. Rodewald. 1998. In vivo roles of receptor tyrosinekinases and cytokine receptors in early thymocyte development. Curr. Opin.Immunol. 10: 196–207.
31. Dittel, B. N., and T. W. LeBien. 1995. The growth response to IL-7 duringnormal human B cell ontogeny is restricted to B-lineage cells expressingCD34. J. Immunol. 154: 58–67.
32. Vogt, T. K., A. Link, J. Perrin, D. Finke, and S. A. Luther. 2009. Novel functionfor interleukin-7 in dendritic cell development. Blood 113: 3961–3968.
33. Surh, C. D., and J. Sprent. 2008. Homeostasis of naive and memory T cells.Immunity 29: 848–862.
34. Hemmoranta, H., S. Hautaniemi, J. Niemi, D. Nicorici, J. Laine, O. Yli-Harja,J. Partanen, and T. Jaatinen. 2006. Transcriptional profiling reflects shared andunique characters for CD34+ and CD133+ cells. Stem Cells Dev. 15: 839–851.
35. Rochman, Y., R. Spolski, and W. J. Leonard. 2009. New insights into the reg-ulation of T cells by gamma(c) family cytokines. Nat. Rev. Immunol. 9: 480–490.
36. Huntington, N. D., N. Legrand, N. L. Alves, B. Jaron, K. Weijer, A. Plet,E. Corcuff, E. Mortier, Y. Jacques, H. Spits, and J. P. Di Santo. 2009. IL-15 trans-presentation promotes human NK cell development and differentiation in vivo.J. Exp. Med. 206: 25–34.
37. Legrand, N., K. Weijer, and H. Spits. 2006. Experimental models to study de-velopment and function of the human immune system in vivo. J. Immunol. 176:2053–2058.
38. Tanchot, C., F. A. Lemonnier, B. Perarnau, A. A. Freitas, and B. Rocha. 1997.Differential requirements for survival and proliferation of CD8 naive or memoryT cells. Science 276: 2057–2062.
39. Zinkernagel, R. M., and A. Althage. 1999. On the role of thymic epithelium vs.bone marrow-derived cells in repertoire selection of T cells. Proc. Natl. Acad.Sci. USA 96: 8092–8097.
40. Hoglund, P., C. Ohlen, E. Carbone, L. Franksson, H. G. Ljunggren, A. Latour,
B. Koller, and K. Karre. 1991. Recognition of beta 2-microglobulin-negative
(beta 2m2) T-cell blasts by natural killer cells from normal but not from beta
2m2 mice: nonresponsiveness controlled by beta 2m2 bone marrow in chimeric
mice. Proc. Natl. Acad. Sci. USA 88: 10332–10336.41. Liao, N. S., M. Bix, M. Zijlstra, R. Jaenisch, and D. Raulet. 1991. MHC class I
deficiency: susceptibility to natural killer (NK) cells and impaired NK activity.
Science 253: 199–202.
42. Yokoyama, W. M., and S. Kim. 2006. How do natural killer cells find self toachieve tolerance? Immunity 24: 249–257.
43. Yokoyama, W. M., and S. Kim. 2006. Licensing of natural killer cells by self-major histocompatibility complex class I. Immunol. Rev. 214: 143–154.
44. Zhao, Y., H. Ohdan, J. O. Manilay, and M. Sykes. 2003. NK cell tolerance inmixed allogeneic chimeras. J. Immunol. 170: 5398–5405.
45. Kawahara, T., J. I. Rodriguez-Barbosa, Y. Zhao, G. Zhao, and M. Sykes. 2007.Global unresponsiveness as a mechanism of natural killer cell tolerance in mixedxenogeneic chimeras. Am. J. Transplant. 7: 2090–2097.
2720 T AND NK CELL LYMPHOPOIESIS IN LONG-TERM ENGRAFTED NSG MICE