55 Gene, 34 (1985) 55-61 Elsevier GENE 1211 Molecular cloning of the RADIO gene of Succharomyces cereuisiue (Recombinant DNA; yeast; DNA repair; radiation sensitivity; Ml3 phage vector; pyrimidine dimers; hypersensitivity) Louise Prakash, Deunis Dumais, Renata Polakowska, Giuditta Perozzi* and Satya Prakash* Department of Radiation Biology and Biophysics, University of Rochester School of Medicine, Rochester, NY 14642 Tel. (716) 275-2656, and *Department of Biology, University of Rochester, Rochester, NY 14627 (U.S.A.) Tel. (716) 275-3847 (Received August 27th, 1984) (Revision received November 13th, 1984) (Accepted November 14th, 1984) SUMMARY We have cloned the RADIO gene of Saccharomyces cerevisiae and physically mapped it to a l.O-kb DNA fragment. Strains containing disruptions of the RADIO gene were found to show enhanced UV sensitivity compared with the previously characterized radlO-1 or radlO-2 mutants. The UV sensitivity of the disruption mutant is comparable to the highly UV sensitive radl-19, ra&-A, and radl-2 mutants. INTRODUCTION In the eukaryote S. cerevisiae, ten genes, RADI, RAD2, RAD3, RAD4, RAD7, RADIO. RAD14, R4D16, RAD23 and MMS19, are involved in exci- sion of UV-induced pyrimidine dimers (Unrau et al., 1971; Resnick and Setlow, 1972; Waters and Moustacchi, 1974; P&ash, 1975; 1977a, b; Reynolds, 1978; Prakash and Prakash, 1979; Miller et al., 1982a). Mutants in some of these genes are highly defective in the first step of excision repair, that of incision, while the others show varying degrees of reduced incision capacity. The radl, raa!2, rad3, rad4, radl0 and mmsl9 mutants are defective in incision of DNA containing pyrimidine dimers (Wilcox and Prakash, 1981; Reynolds and Abbreviations: A, deletion; kb, kilobases or kilobase pair; “(superscript), resistance; tet, tetracycline; UV, ultraviolet light; [ 1. indicates plasmid-carrier state. 0378-l 119/85/%03.30 0 1985 Elsevier Science Publishers Friedberg, 1981) or interstrand crosslinks (Miller et al., 1982b; Jachymczyk et al., 1981; Magafla- Schwencke et al., 1982). The radll mutant, which is also excision defective, exhibits a lowered incision proficiency than the Rad’ strain (Wilcox and Prakash, 1981; Miller et al., 1982b). The rad7 and rad23 mutants are partially defective in the removal of pyrimidine dimers and crosslinks (Miller et al., 1982a). The rad16 mutant shows a partial defect in pyrimidine dimer removal (P&ash, 1977b; Wilcox and P&ash, 1981) but appears to be normal in nicking of crosslinked DNA (Miller et al., 1982b). Our goal is to characterize the protein products of the genes affecting the incision process and to deter- mine the incision mechanism. We have cloned and characterized the R4DI (Higgins et al., 1983a), RAD2 (Higgins et al., 1984) and RAD3 (Higgins et al., 1983b) genes. In this paper, we describe the isolation of the RADlO gene and its localization to about 1 kb of DNA. In addition, a radl0 mutant
Transcript
55 Gene, 34 (1985) 55-61 Elsevier
GENE 1211
Molecular cloning of the RADIO gene of Succharomyces cereuisiue
Louise Prakash, Deunis Dumais, Renata Polakowska, Giuditta Perozzi* and Satya Prakash*
Department of Radiation Biology and Biophysics, University of Rochester School of Medicine, Rochester, NY 14642 Tel. (716) 275-2656, and *Department of Biology, University of Rochester, Rochester, NY 14627 (U.S.A.) Tel. (716) 275-3847
(Received August 27th, 1984) (Revision received November 13th, 1984) (Accepted November 14th, 1984)
SUMMARY
We have cloned the RADIO gene of Saccharomyces cerevisiae and physically mapped it to a l.O-kb DNA fragment. Strains containing disruptions of the RADIO gene were found to show enhanced UV sensitivity compared with the previously characterized radlO-1 or radlO-2 mutants. The UV sensitivity of the disruption mutant is comparable to the highly UV sensitive radl-19, ra&-A, and radl-2 mutants.
INTRODUCTION
In the eukaryote S. cerevisiae, ten genes, RADI,
RAD2, RAD3, RAD4, RAD7, RADIO. RAD14,
R4D16, RAD23 and MMS19, are involved in exci- sion of UV-induced pyrimidine dimers (Unrau et al., 1971; Resnick and Setlow, 1972; Waters and Moustacchi, 1974; P&ash, 1975; 1977a, b; Reynolds, 1978; Prakash and Prakash, 1979; Miller et al., 1982a). Mutants in some of these genes are highly defective in the first step of excision repair, that of incision, while the others show varying degrees of reduced incision capacity. The radl, raa!2, rad3, rad4, radl0 and mmsl9 mutants are defective in incision of DNA containing pyrimidine dimers (Wilcox and Prakash, 1981; Reynolds and
Abbreviations: A, deletion; kb, kilobases or kilobase pair; “(superscript), resistance; tet, tetracycline; UV, ultraviolet light; [ 1. indicates plasmid-carrier state.
Friedberg, 1981) or interstrand crosslinks (Miller et al., 1982b; Jachymczyk et al., 1981; Magafla- Schwencke et al., 1982). The radll mutant, which is also excision defective, exhibits a lowered incision proficiency than the Rad’ strain (Wilcox and Prakash, 1981; Miller et al., 1982b). The rad7 and rad23 mutants are partially defective in the removal of pyrimidine dimers and crosslinks (Miller et al., 1982a). The rad16 mutant shows a partial defect in pyrimidine dimer removal (P&ash, 1977b; Wilcox and P&ash, 1981) but appears to be normal in nicking of crosslinked DNA (Miller et al., 1982b).
Our goal is to characterize the protein products of the genes affecting the incision process and to deter- mine the incision mechanism. We have cloned and characterized the R4DI (Higgins et al., 1983a), RAD2 (Higgins et al., 1984) and RAD3 (Higgins et al., 1983b) genes. In this paper, we describe the isolation of the RADlO gene and its localization to about 1 kb of DNA. In addition, a radl0 mutant
56
obtained by disrupting the RADIO gene in the yeast genome, is far more UV-sensitive than the m&O-1 or rudlO-2 mutants.
MATERIALS AND METHODS
(a) Yeast and bacterial strains
The following yeast strains were employed: 962-3b, MATor adez leul rudlU-1 (obtained from Richard Snow); GP7-32, MATa canR his3-dl tipl-289 RAD’ ; GP7-123, MATa canR his3-Al ~1-289 ~~3-52 rudlO-2; LP2817-8D, MATa
RAD’ (obtained from Bill Weiss); DBY746, MATa his3-A 1 Ieu2-3 leu2-112 trpl-289 ura3-52 RAD+ (obtained from David Botstein); and 7799-5C, MATa h&4-17 uru3-52 RAD + (obtained from Gerry Fink).
Synthetic complete medium (P&ash et al., 1980) lacking uracil was used to maintain and grow plasmid-bearing yeast strains. Escherichia coli strain
JM103 was used to propagate Ml3 derivative phages, while HBlOl was used to propagate all plasmids.
(b) Plasmids
We employed a yeast r~ornb~~t bank, obtained from David Botstein, consisting of partial Suu3A digests of yeast genomic DNA which had been ligated into the BumHI site of the 7.6kb hybrid plasmid YEp24 (Botstein et al., 1979; Carlson and Botstein, 1982). YEp24 contains pBR322, and a 2.2-kb EcoRI fragment of yeast 2 p circle DNA, conferring the ability to replicate autonomously in high copy number in yeast, and a l.l-kb Hind111 fragment containing the yeast U&A3 gene, for selec- tion in yeast.
YIPS, another derivative of pBR322, is a 5.5-kb plasmid containing the yeast URA3 gene but no sequences conferring the ability to replicate autono- mously in yeast. Therefore, it can transform yeast only by homologous integration into the yeast genome (Botstein et al., 1979; Scherer and Davis, 1979).
pTB220, derived from YIp5, is a 7.7-kb plasmid containing the yeast 2 p circle origin of replication obtained from YEp24 (Higgins et al., 1984). Plasmid pTB220 contains unique BumHI, EcoRI, Hind111 and Sal1 sites, replicates autonomously in yeast and is maintained in high copy number.
(c) Transformation procedures and UV irradiation
Tr~sfo~ations were carried out by the method of Ito et al, (1983) in which lithium acetate is used to treat intact yeast cells, which promotes DNA uptake. The UV survival of different yeast strains was measured as described by Prakash and Prakash (1977).
(d) Other methods
Procedures used for puriIication of DNA were as described previously by Higgins et al. (1983b).
RESULTS
(a) Isolation of the RADlO gene
We used the yeast genomic bank constructed in the vector YEp24 (Carlson and Botstein, 1982) to isolate the RADfO gene by complementation of the radlO-2 ura3-52 strain GP7-123 for UV resistance. One Rad + tr~sform~t was obtained from approx. 9000 Ura+ ~~sfo~~ts, This strain harbored a plasmid, designated pTB 185, which complemented both the radlO-2 strain GP7-123 (Fig. 1) as well as the radlO-1 strain LP2817-8D. A restriction map of the 11.3-kb yeast DNA insert in plasmid pTB185 is given in Fig. 2.
(b) Genetic mapping of the cloned DNA segment
Since complementation of radlO strains for UV resistance could result from suppression of the Rad - phenotype by some other gene when present on a multicopy plasmid, the 3.15-kb BgflI fragment, located at 0.40 to 3.55 kb of pTB185 (Fig. 2), was genetically mapped to the rudl0 site in the yeast genome. The 3.15-kb BgZII fragment was ligated into the BamHI-restricted integrating vector YIPS which
0.01
0.001
0.0001
UV FLUENCE (J/m*)
contains the lJRA3 gene. The plasmid generated, pTB208, can transform yeast by integration at homologous regions in the yeast genome (Scherer and Davis, 1979), which in this case, would occur at either the uru3 or radl0 sites, if the cloned DNA segment contains the RADIO gene. However, inte- gration at a particular locus can be directed by a double-strand cleavage in the DNA homologous to the desired site of integration (OK-Weaver et al., 1983). Therefore, plasmid pTB208 that had been linearized by cutting at the BumHI site at position 0.7 kb (Fig. 2) in the yeast DNA insert was used to transform the ura3-52 rudlO-2 strain GP7-123 to
Fig. 1. UV survival of yeast strains with and without various R4D10-insert-containing plasmids. Cells were grown in minimal
medium supplemented with the appropriate nutrients except that plasmid containing strains were grown in selective medium lacking uracil. Irradiation was as described by Prakash and P&ash (1977). Only the relevant genotype of strains is given here. 0, Strain GP7-32, MD+; * , LP2817-8D, ura3-52 r&O-1; v, GP7-123, uru3-52 rudlO-2; 0, GP7-123[pTB185]; A, GP7-123[pTB215]; A, GP7-123[pTB240]; 0, GP7-123-
[pTB233]; n , GP7-123[pTB208]; V, GP7-123[pDD16]; 0,
GP7_123[YEp24]; +, GP7-123[pTB218]; open hexagon, GP7-123[pDD17]; SX46A, ura3-52 RAD +, transformed to Ura+ and Rad- to generate rudl0 disruptions with the 2.6-kb SalI-BumHI fragment of pDD37, which contains the UR43 gene inserted within the RADIO gene, x , disruption 1; filled hexagon, disruption 2.
I 1 I I I I 1 I I I 1 I f
kb 0 I 2 3 4 5 6 7 8 9 IO II 12
*GSB EXbHSPv Xb V G Ii E HCXGPX E l-i SXGG* Plasmid
Ps Pvti vector Gnnplementation
UlM’(// I III I of fad10
____ ____ pTSlt35 YEp24 +
I pTS201 YEp24 +
I I PTS203 YW
I I PT’-J~ YEp24 +
I I PT= YIPS +
1 I pTB212 YW +
pTS215 YEp24 +
Xbt
I I pTS214 YIP5
Fig. 2. Restriction map of the yeast DNA insert in plasmid pTB185 and complementation pattern of pTB185 subclones. Insert of the yeast DNA segment in pTB185, given by the open bar, is in the BumHI site of the ref gene of YEp24. The thin line represents YEp24 DNA and asterisks denote the fusion of insert with plasmid. Symbols for restriction enzymes are as follows: A, AvaI; B, BarnHI; C, SacI; E, EcoRI; G, BglII; H, HindIII; Hc, ZfincII; P, SphI; Ps, MI; Pv, PvuII; S, SalI; V, EcoRV; X, XhoI; Xb, XbaI. Xbt denotes the XbaI site destroyed by blunt ending of XbaI digested plasmid pTB212, followed by ligation to yield the plasmid pTB214.
58
Ura + . A total of 56 transformants were obtained, of which 45 were Rad + Ura+ and 11 were Rad- Ura+ . The Rad- Ura’ transformants most likely arose as a result of gene conversion. Three of the Rad + Ura+ transformants were crossed to the Rad’ ~~3-52 strain LP2682-4B to generate the diploids RP-3 I, LP-28 19 and LP-2820, respectively. Analysis of 3520 and 16 tetrads, respectively, from these three spot-mated diploids yielded only Rad’ spores, indicating that integration of pTB208 had occurred at the radl0 site in the yeast genome. Furthermore, genetic analysis of 20 tetrads from the sporulated diploid RP-33, obtained by crossing one of the Rad- Ura+ transformants to the Rad’ ~~~3-52 strain LP2682-4B, gave only parental ditype tetrads, i.e., 2 Rad - Ura + : 2 Rad + Ura- spores. All these results provide genetic evidence that the 3.15-kb BglII fragment contains the RADfO gene.
(c) Location of the RADIO gene in the cloned DNA insert
Subcloning of the 11.3-kb DNA insert in plasmid pTB 185 (Fig. 2) was achieved by constructing plas- mids with various deletions of ML)10 region DNA, or inserting DNA restriction fragments in suitable plasmid vectors and testing for complementation ability. The subcloning and complementation results are summarized in Figs. 2 and 3. The plasmid pTB201, which is deleted for the 3.0-kb XhoI frag- ment, and the plasmid pTB209, which is deleted for the X/z01 fragment and the two leftwards Sal1 frag- ments, complemented the rudlO-2 mutation (Fig. 2).
t I
kb 0 0.5 1.0 1.5 2.0
Plasmld Vf!CtOr Camplementation
SA Pv Xb HcVHcG of radl0
t-1 pTB215 YEp24 +
i I pTEiZl8 YEp24
r I pTB233 YEp24 +
I I pTB240 pTB220 +
I I pODI6 pTf3220 +
1 pODI7 pTB220
Fig. 3. Restriction map of the 1.85-kb &WI-BgZII yeast DNA insert in plasmid pTB215 and complementation pattern of pTB215 subclones. Symbols for restriction sites are as in the legend to Fig. 2.
The 3.85-kb &g&I-XhoI fragment in plasmid pTB203 did not complement the rudlO-2 mutation (Fig. 2), whereas the 3.15-kb Bg111 fragment in plasmid pTB208 and the 1.85-kb SalI-BgEII fragment in plasmids pTB212 and pTB215 did complement the rudlO-2 mutation (Figs, 1 and 2). The&z1 site in the 1.85-kb SalI-BglII fragment is internal to the RADIO
gene, since disruption at this site led to loss of complementing activity in both the integrating plas- mid pTB214 (Fig. 2) and the autonomously replicat- ing plasmid pTB218 (Figs. 1 and 3). For further subcloning of the 1.8%kb &II-BgZII fragment, the 0.22-kb SalI-PvuII segment situated at O-O.22 kb (Fig. 3) of pTB215 was removed, generating the plasmid pTB233 which complements the radlO-2
mutation (Figs. 1 and 3). To determine whether the far right region of pTB2 15 is required for RADlO
function, the region between EcoRV and BglII was deleted, generating the plasmid pTB240, which complements the rudlO-2 mutation (Figs. 1 and 3). Since plasmid pTB233, which lacks DNA sequences to the left of PvuII, and plasmid pTB240, which lacks DNA sequences to the right of EcoRV, both restore Rad + levels of UV resistance to the rudlO-2 strain GP7-123, the RADIO gene must lie within the 1.3-kb PvuII-EcoRV fragment. To delimit the RADIO gene even further, BAL 3 1 deletions were generated from the EcoRV site and BarnHI linkers attached. DNA segments of varying lengths were cloned as SalI- BarnHI fragments. The smallest complementing SalI-BumHI fragment, found in plasmid pDDl6, restored normal levels of UV resistance to the rudlO-2 strain GP7-123 (Figs. 1 and 31, while the largest non-complementing SalI-BumHI fragment, located in plasmid pDDl7, gave a UV response similar to the r~dlO-2 mut~t strain (Figs. 1 and 3). Our subcloning results indicate that the ML)10 gene is contained within approx. 1 kb of DNA between the PvuII site on the left at position 0.22 kb and the right end of the insert in plasmid pDDl6 at 1.25 kb (Fig. 3).
(d) Disruption of the RADIO gene
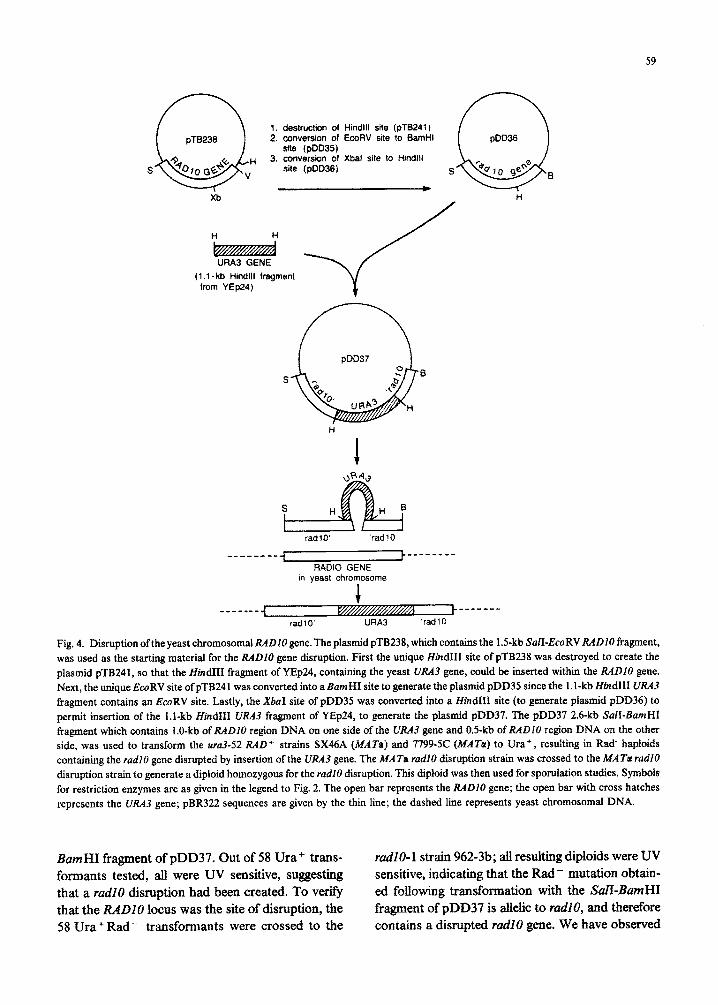
The RADIO gene was disrupted by the method of Rothstein (1983) by insertion of the UEA3 gene at the XbuI site which is internal to the RADlO gene, as shown in Fig. 4. The ~~3-52 Rad+ strain SX46A was transformed to Ura+ with the 2.6-kb SalI-
59
1, destruction of Hindlll site (pTB241) 2. conversion of EcoRV site to EarnHI
(1 .l - kb Hindlll fragment from YEp24)
Y
/\ ~0037
RADIO GENE m yeast chromosome
4
-------i-------
Fig. 4. Disruption of the yeast chromosomal RADlO gene. The plasmid pTB238, which contains the 1.5kb SalI-EcoRV R4DlO fragment, was used as the starting material for the RADZO gene disruption. First the unique Hind111 site of pTB238 was destroyed to create the plasmid pTB241, so that the Hind111 fragment of YEp24, containing the yeast UR43 gene, could be inserted within the RADZO gene. Next, the unique EcoRV site of pTB241 was converted into aBamH1 site to generate the plasmid pDD35 since the l.l-kb Hind111 WA3 fragment contains an EcoRV site. Lastly, the J&z1 site of pDD35 was converted into a EIindIII site (to generate plasmid pDD36) to permit insertion of the l.l-kb Hind111 U&t3 fragment of YEp24, to generate the plasmid pDD37. The pDD37 2.6kb MI-BumHI fragment which contains I.O-kb of MD10 region DNA on one side of the URA3 gene and OS-kb of RADIO region DNA on the other side, was used to transform the ~~3-52 RAD + strains SX46A (MATa) and 779%5C (MATa) to Ura+, resulting in Rad- haploids containing the radI0 gene disrupted by insertion of the UR43 gene. The MATa rudi0 disruption strain was crossed to the MATa tad10
disruption strain to generate a diploid homozygous for the radl0 disruption. This diploid was then used for sporulation studies. Symbols for restriction enzymes are as given in the legend to Fig. 2. The open bar represents the RADIO gene; the open bar with cross hatches represents the URA3 gene; pBR322 sequences are given by the thin line; the dashed line represents yeast chromosomal DNA.
BamHI fragment of pDD37. Out of 58 Ura+ trans- ru&O-1 strain 962-3b; all resulting diploids were UV formants tested, all were UV sensitive, suggesting sensitive, indicating that the Rad- mutation obtain- that a ru&O disruption had been created. To verify ed following transformation with the MI-BarnHI that the RADIO locus was the site of dis~ption, the fragment of pDD37 is allelic to rudl0, and therefore 58 Ura+ Rad - ~~sfo~~ts were crossed to the contains a disrupted radI0 gene. We have observed
60
that these r&U disruption mutants show much greater UV sensitivity than the rudlO-2 mutant or the rudlO-1 mutant (Fig. 1). The rudl0 disruption does not a&et viabi~ty or growth rate of vegetative cells or sporulation and spore viability (not shown).
DISCUSSION
We have isolated the RADIO gene and mapped it to 1-kb of DNA between thePvuI1 site on the left and the end of the insert on the right in plasmid pDD 16 (Fig. 3). The RADIO gene is much smaller in size than the RADl, RAD2 and RAD3 genes, which encode 3.1-, 3.3- and 2%kb transcripts, respectiveIy (Higgins et al., 1983a, b; 1984).
We have also created a disruption in the RADIO gene by ~te~a~g the UR43 gene within it. The radlO disruption mutant shows much greater UV sensitivity than either the rudlO-1 or rudlO-2 mu- tants, its UV sensitivity being comparable to the highly UV sensitive radl-19 (Higgins et al., 1983a) rad2-A (Higgins et al., 1984) and rud3-2 (Higgins et al., 1983b) mutants. Since it has been previously shown that the rudlO-2 mutant is defective in incising DNA at pyrimidine dimers (Wilcox and Prakash, 1981), the enhanced sensitivity of the rudl0 disrup- tion mutant could be due to a complete defect in pyrimidine dimer incision, while the rudl O-2 mutant could have a reduced rate of incision relative to the Rad + strain. Alternatively, in addition to the incision defect, the radl0 disruption mutant could be defec- tive in some other aspect of DNA repair, thereby accounting for its greater UV sensitivity compared with the rudlO-1 or radlO-2 mutants. The radl0 dis~ption does not affect viability of vegetative cells or spores, as has also been observed with the radl and rud2 deletions (Higgins et al., 1983a; 1984; Naumovski and Friedberg, 1984). The rud3 deletion mutant, on the other hand, is a recessive lethal (Higgins et al., 1983b; Naumovski and Friedberg, 1983) indicating an essential role of this incision gene.
ACKNOWLEDGEMENTS
This investigation was supported by grants CA35035 to S.P. and GM19261 to L.P. from NIH,
and contract no. DE-ACO2-74EVO3490 with the U.S. Department of Energy at the University of Rochester Department of Radiation Biology and Biophysics and has been assigned report No. DOE/EV/03490-2429.
REFERENCES
Botstein, D., Falco, SC., Stewart, SE., Brennan, M., Scherer, S., Stinchcomb, D.T., Struhl, K. and Davis, R.W.: Sterile host yeasts (SHY): a eukaryotic system of biological containment for recombinant DNA experiments. Gene 8 (1979) 17-24.
Carlson, M. and Botstein, D.: Two ~fferenti~ly regulated mRNAs with different 5’ ends encode secreted and intra- cellular forms of yeast invertase. Cell 28 (1982) 145-154.
Higgins, D.R., Prakash, S., Reynolds, P. and P&ash, L.: Molecular cloning and characterization of the RADI gene of Saccharomyces eerevisiae. Gene 26 (1983a) 119-126.
Higgins, D.R., P&ash, S., Reynolds, P., Polakowska, R., Weber, S. and Prakash, L.: Isolation and characterization of the RAD3 gene of Saccharomyces cerevisiae and inviability of rad3 deletion mutants. Proc. Natl. Acad. Sci. USA 80 (1983b) 5680-5684.
Higgins, D.R., Prakash, L., Reynolds, P. and Prakash, S.: Isolation and characterization of the RAD2 gene of Saccharo- myces cerevisiae. Gene 30 (1984) 121-128.
Ito, H., Fukuda, Y., Murata, K. and Kimura, A.: Transformation of intact yeast cells treated with alkali cations. J. Bacterial. 153 (1983) 163-168.
Jach~czyk, W.J., von Borstel, R.C., Mowat, M.R.A. and Hastings, P.J.: Repair of interstrand cross-links in DNA of Saccharomyces cerevisiae requires two systems for DNA repair: the RAD3 system and the RAD51 system. Mol. Gen. Genet. 182 (1981) 196-205.
Johnston, L.H. and Nasmyth, K.A.: Saceharomyces cerevisiue cell cycle mutant cdc9 is defective in DNA ligase. Nature 274 (1978) 891-893.
Maga~a-Schwencke, N., Henriques, J.A.P., Chanet, R. and Moustacchi, E.: The fate of 8-methoxypsoralen photoinduced cross-links in nuclear and mitochondrial yeast DNA: com- parison of wild type and repair-deficient strains. Proc. Natl. Acad. Sci. USA 79 (1982) 1722-1726.
Miller, R.D., P&ash, L. and Prakash, S.: Defective excision of pyrimidine dimers and interstrand DNA crosslinks in rad7
and rad23 mutants of Saccharomyces cerevisiae. Mol. Gen. Genet. 188 (1982a) 235-239.
Miller, R.D., Prakash, L. and Prakash, S.: Genetic control of excision of Saccharomyces cerevisiae interstrand DNA cross- links induced by psoralen plus near-UV light. Mol. Cell. Biol. 2 (198213) 939-948. l
Naumovski, L. and Friedberg, E.C.: A DNA repair gene required for the incision of damaged DNA is essential for viability in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 80 (1983) 4818-482’
61
Naumovski, L. and Friedberg, E.C.: Saccharomyces cerevisiae
Orr-Weaver, T.L., Szostak, J.W. and Rothstein, R.J.: Genetic applications of yeast transformation with linear and gapped plasmids. Methods Enzymol. 101 (1983) 228-245.
P&ash, L.: Repair of pyrimidine dimers in nuclear and mito- chondrial DNA of yeast irradiated with low doses of ultra-
violet light. J. Mol. Biol. 98 (1975) 781-795. Prakash, L.: Repair of pyrimidine dimers in radiation-sensitive
mutants rad3, rad4, rad6. and rod9 of Saccharomyces cere-
sensitive mutants radl0 and radld of Saccharomyces cere-
visae. Mol. Gen. Genet. 152 (1977b) 125-128. Prakash, L. and Prakash, S.: Isolation and characterization of
MMS-sensitive mutants of Saccharomyces cerevisiae. Genet- ics 86 (1977) 33-55.
Prakash, L. and Prakash, S.: Three additional genes involved in pyrimidine dimer removal in Saccharomyces cerevisiae: RAD7,
RAD14 and MMSI9. Mol. Gen. Genet. 176 (1979) 351-359. Prakash, S., Prakash, L., Burke, W. and Montelone, B.A.: Effects
of the RADSZ gene on recombination in Saccharomyces
cerevisiae. Genetics 94 (1980) 3 l-50. Resnick, M.A. and Setlow, J.K.: Repair of pyrimidine dimer
damage induced in yeast by ultraviolet light. J. Bacterial. 109 (1972) 979-986.
Reynolds, R.J.: Removal of pyrimidine dimers from Saccharo-
myces cerevisiae nuclear DNA under nongrowth conditions as detected by a sensitive enzymatic assay. Mutation Res. 50
(1978) 43-56. Reynolds, R.J. and Friedberg, EC.: Molecular mechanisms of
pyrimidine dimer excision in Saccharomyces cerevzkiae: inci- sion of ultraviolet-irradiated deoxyribonucleic acid in vivo. J. Bacterial. 146 (1981) 692-704.
Rothstein, R.J.: One-step gene disruption in yeast. Methods
Enzymol. 101 (1983) 202-211. Scherer, S. and Davis, R.W.: Replacement of chromosome
segments with altered DNA sequences constructed in vitro.
Proc. Natl. Acad. Sci. USA 76 (1979) 4951-4955. Unrau, P., Wheatcroft, R. and Cox, B.S.: The excision of
pyrimidine dimers from DNA of ultraviolet irradiated yeast. Mol. Gen. Genet. 113 (1971) 359-362.
Waters, R. and Moustacchi, E.: The disappearance ofultraviolet- induced pyrimidine dimers from the nuclear DNA of expo- nential and stationary phase cells of Saccharomyces ceretiiae
following various post-irradiation treatments. Biochim. Biophys. Acta 353 (1974) 407-419.
Wilcox, D.R. and Prakash, L.: Incision and postincision steps of
pyrimidine dimer removal in excision-defective mutants of Saccharomyces cerevisiae. J. Bacterial. 148 (1981) 618-623.