ORIGINAL ARTICLE Novel Spiroplasma sp. Isolated From CWD Is an Extreme Bacterial Thermoacidophile That Survives Autoclaving, Boiling, Formalin Treatment, and Significant Gamma Irradiation Frank O. Bastian, MD, James Lynch, BSc, and Wei-Hsung Wang, PhD Abstract Rapid spreading of chronic wasting disease (CWD) in wildlife and captive cervid populations has exposed lack of progress in deal- ing with the transmissible spongiform encephalopathies (TSE) of man and animals. Since the TSE transmissible agent was resistant to extremes in environmental and chemical treatments, focus was on an unconventional agent including the prion theory. Recent break- through research has revealed consistent isolation of a novel Spiro- plasma sp. from TSE-affected tissues that propagates in cell-free media and on agar. Here, we developed a live culture assay to test whether the CWD spiroplasma isolate possessed unconventional bi- ologic properties akin to those of the transmissible agent of TSE. The CWD spiroplasma isolate survived boiling for 1 hour, standard liquid autoclaving, 10% formalin treatment overnight, and gamma irradiation of 20 kGy. The CWD spiroplasma isolate is an acido- phile, growing best at pH 2. The biologic resistance of the CWD spi- roplasma isolate may be due to unusual phage-like viruses found in the bacterial pellet or to DNA-protein binding. Because the CWD spiroplasma isolate has biologic properties consistent with the causal agent of the TSEs, TSE research focus should be redirected to devel- opment of diagnostic tests and preventive vaccines for control of CWD based upon the bacterium. Key Words: Chronic wasting disease, Creutzfeldt Jacob disease, Drop assay, Gamma irradiation, Prion, Scrapie, Thermoacidophile. INTRODUCTION Molecular and morphological studies have shown that a filterable wall-less bacterium, spiroplasma, is consistently as- sociated with transmissible spongiform encephalopathies (TSE) (1, 2). Recent breakthrough research has shown that this novel microbe can be isolated from tissues affected with TSE including scrapie in sheep, chronic wasting disease (CWD) in deer, and Creutzfeldt-Jakob disease (CJD) in humans (3). This spiroplasma TSE isolate has previously been recovered from CWD and scrapie-affected brain and lymphoid tissues by passage in embryonated eggs (4). When inoculated intracranially into sheep and goats, the spiroplasma TSE iso- lates caused development of spongiform encephalopathy. More specifically, the presence of vacuolated neurons in the brainstem of these experimentally inoculated animals was identical to characteristic lesions seen in the obices of natu- rally occurring TSE in deer, sheep, and goats (4). We have used our new culture method to re-isolate the novel spiro- plasma from brain and lymphoid tissues from these experi- mentally inoculated ruminants into cell-free culture (3). The spiroplasma TSE isolate was easily identified in the broth cul- ture by dark-field microscopy and by formation of typical sub- surface spiroplasma colonies on Bacto Schaedler agar plates (3). This process fulfills the scientific criteria for determina- tion of causality, otherwise known as Koch’s postulates. These observations brought up the question of whether this novel TSE spiroplasma isolate possessed the unconven- tional biologic properties determined for the transmissible agent of scrapie in the 1960–1970s (5, 6). The consistent in vitro isolation of the novel TSE-related spiroplasma in cell- free broth culture and as subsurface colonies on agar plates has provided the mechanism to fully characterize the biologic properties of this bacterial pathogen (3). The properties of the TSE-spiroplasma isolate can then be compared with those of the transmissible scrapie agent. We exposed the cultures of TSE-spiroplasma isolates in Brucella media to test chemical and environmental extremes to determine survival. The re- markable similarities of these resistant studies of the spiroplasma TSE isolate to the early studies of the scrapie agent (5, 6) strongly support the role of this novel Spiroplasma sp. as a candidate causal agent of the TSEs. From the Bastian Laboratory for Neurological Disease Research, New Or- leans, Louisiana (FOB, JL); Radiation Safety Office/Center for Energy Studies, Louisiana State University, Louisiana (W-HW); Texas Tech University, Department of Environmental Toxicology, Lubbock, Texas (FOB); and Tulane Medical School Department of Pathology, New Or- leans, Louisiana (FOB). Send correspondence to: Frank O. Bastian, MD, Bastian Laboratory for Neu- rological Disease Research, New Orleans, LA 70115; E-mail: [email protected]“LIFT GRANT,” by LSU Research and Technology Foundation. LSU; ACRES (Audubon Center for Research of Endangered Species); the Uni- fied Sportsmen of Pennsylvania; the Deer Breeder’s Corporation and the Sardo foundation for financial support. The Bastian Laboratory for Neu- rological Disease Research is sponsored by “The Terra Cor Institute” A nonprofit organization. The authors have no duality or conflicts of interest to declare. Supplementary Data can be found at academic.oup.com/jnen. 993 V C 2019 American Association of Neuropathologists, Inc. All rights reserved. J Neuropathol Exp Neurol Vol. 78, No. 11, November 2019, pp. 993–1001 doi: 10.1093/jnen/nlz081 Downloaded from https://academic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 October 2019
Transcript
ORIGINAL ARTICLE
Novel Spiroplasma sp. Isolated From CWD Is an ExtremeBacterial Thermoacidophile That Survives Autoclaving,Boiling, Formalin Treatment, and Significant Gamma
Irradiation
Frank O. Bastian, MD, James Lynch, BSc, and Wei-Hsung Wang, PhD
AbstractRapid spreading of chronic wasting disease (CWD) in wildlife
and captive cervid populations has exposed lack of progress in deal-
ing with the transmissible spongiform encephalopathies (TSE) of
man and animals. Since the TSE transmissible agent was resistant to
extremes in environmental and chemical treatments, focus was on an
unconventional agent including the prion theory. Recent break-
through research has revealed consistent isolation of a novel Spiro-plasma sp. from TSE-affected tissues that propagates in cell-free
media and on agar. Here, we developed a live culture assay to test
whether the CWD spiroplasma isolate possessed unconventional bi-
ologic properties akin to those of the transmissible agent of TSE.
The CWD spiroplasma isolate survived boiling for 1 hour, standard
liquid autoclaving, 10% formalin treatment overnight, and gamma
irradiation of 20 kGy. The CWD spiroplasma isolate is an acido-
phile, growing best at pH 2. The biologic resistance of the CWD spi-
roplasma isolate may be due to unusual phage-like viruses found in
the bacterial pellet or to DNA-protein binding. Because the CWD
spiroplasma isolate has biologic properties consistent with the causal
agent of the TSEs, TSE research focus should be redirected to devel-
opment of diagnostic tests and preventive vaccines for control of
CWD based upon the bacterium.
Key Words: Chronic wasting disease, Creutzfeldt Jacob disease,
Drop assay, Gamma irradiation, Prion, Scrapie, Thermoacidophile.
INTRODUCTIONMolecular and morphological studies have shown that a
filterable wall-less bacterium, spiroplasma, is consistently as-sociated with transmissible spongiform encephalopathies(TSE) (1, 2). Recent breakthrough research has shown thatthis novel microbe can be isolated from tissues affected withTSE including scrapie in sheep, chronic wasting disease(CWD) in deer, and Creutzfeldt-Jakob disease (CJD) inhumans (3). This spiroplasma TSE isolate has previously beenrecovered from CWD and scrapie-affected brain and lymphoidtissues by passage in embryonated eggs (4). When inoculatedintracranially into sheep and goats, the spiroplasma TSE iso-lates caused development of spongiform encephalopathy.More specifically, the presence of vacuolated neurons in thebrainstem of these experimentally inoculated animals wasidentical to characteristic lesions seen in the obices of natu-rally occurring TSE in deer, sheep, and goats (4). We haveused our new culture method to re-isolate the novel spiro-plasma from brain and lymphoid tissues from these experi-mentally inoculated ruminants into cell-free culture (3). Thespiroplasma TSE isolate was easily identified in the broth cul-ture by dark-field microscopy and by formation of typical sub-surface spiroplasma colonies on Bacto Schaedler agar plates(3). This process fulfills the scientific criteria for determina-tion of causality, otherwise known as Koch’s postulates.
These observations brought up the question of whetherthis novel TSE spiroplasma isolate possessed the unconven-tional biologic properties determined for the transmissibleagent of scrapie in the 1960–1970s (5, 6). The consistentin vitro isolation of the novel TSE-related spiroplasma in cell-free broth culture and as subsurface colonies on agar plateshas provided the mechanism to fully characterize the biologicproperties of this bacterial pathogen (3). The properties of theTSE-spiroplasma isolate can then be compared with those ofthe transmissible scrapie agent. We exposed the cultures ofTSE-spiroplasma isolates in Brucella media to test chemicaland environmental extremes to determine survival. The re-markable similarities of these resistant studies of thespiroplasma TSE isolate to the early studies of the scrapieagent (5, 6) strongly support the role of this novel Spiroplasmasp. as a candidate causal agent of the TSEs.
From the Bastian Laboratory for Neurological Disease Research, New Or-leans, Louisiana (FOB, JL); Radiation Safety Office/Center for EnergyStudies, Louisiana State University, Louisiana (W-HW); Texas TechUniversity, Department of Environmental Toxicology, Lubbock, Texas(FOB); and Tulane Medical School Department of Pathology, New Or-leans, Louisiana (FOB).
Send correspondence to: Frank O. Bastian, MD, Bastian Laboratory for Neu-rological Disease Research, New Orleans, LA 70115; E-mail:[email protected]
“LIFT GRANT,” by LSU Research and Technology Foundation. LSU;ACRES (Audubon Center for Research of Endangered Species); the Uni-fied Sportsmen of Pennsylvania; the Deer Breeder’s Corporation and theSardo foundation for financial support. The Bastian Laboratory for Neu-rological Disease Research is sponsored by “The Terra Cor Institute” Anonprofit organization.
The authors have no duality or conflicts of interest to declare.Supplementary Data can be found at academic.oup.com/jnen.
993VC 2019 American Association of Neuropathologists, Inc. All rights reserved.
J Neuropathol Exp NeurolVol. 78, No. 11, November 2019, pp. 993–1001doi: 10.1093/jnen/nlz081
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
IsolationThe spiroplasma TSE isolate was prepared in the follow-
ing manner (3). A small portion of TSE-affected brain or lym-phoid tissues (5 mg) from 5 individual CWD-affectedwhite-tailed deer (Odocoileus virginianus). Each sample wasplaced onto a sterile plastic petri dish and minced with sterilescalpels. The tissue sample was transferred to a sterile grindertube and 5 mL Brucella media (10% fetal bovine serum [FBS])added. Following homogenization, the tube was capped andcentrifuged at 3000g at room temperature for 10 minutes. Thesupernatant was collected in a 5-mL syringe and the pellet dis-carded. The supernatant was filtered through a 0.45-lm sy-ringe filter and 10 mL Brucella media (10% FBS) was added.The preparation was incubated at 35�C for 12–14 days incapped tubes.
Preparation of 1% Brucella (10% FBS) AgarPlates
Fourteen g Brucella broth powder was added to 450 mLtype 2 H2O, then 5.0 g Agar powder was added and the prepa-ration heated until boiling, making sure all agar was dissolved.The mixture was autoclaved then cooled in water bath to<65�C. Fifty milliliter of FBS was added to the media, whichwas then poured into 100� 15 mm plates (20 mL into each)and left uncovered for 30–60 minutes to cool. The agar plateswere repackaged in parafilm and kept refrigerated at 4�C untiluse.
Inoculation of Bacto Schaedler Agar PlatesThe broth culture was pelleted at 20 000g for 10 minutes
and the pellet resuspended in 500 mL phosphate buffered sa-line (PBS) and briefly vortexed to disperse organisms. Fivemicroliter drops were placed onto the agar plates and allowedto air dry for �5 minutes. The inoculated plates were sealedwith parafilm, inverted and incubated for 14 days at 35�C.
Staining of PlatesThe inoculated agar plates were stained with Oil Red O
according to the method of Cluss and Somerson (7). An OilRed O stock solution was prepared and filtered through What-man No. 1 filter paper. A Coomassie Blue R-250 counter stainwas prepared and similarly filtered. The Oil Red O dye was di-luted (6 parts dye to 4 parts water) just prior to staining. Theplates were flooded with 10 mL of the diluted dye for 10–15 minutes, washed with water and allowed to dry. The platewas then flooded with 2 mL Coomassie Blue R-250 stain for30–40 seconds, washed with water and left to dry upsidedown. The lid was placed on plate that was then sealed withparafilm.
Examination of PlatesThe plates were examined and photographed using a
Leica DMIL LED phase-contrast inverted microscope
equipped with a DMC 4500 color camera and long focallength objective lenses.
Agar Plate Drop Assay for Testing SurvivorshipThe broth culture samples were prepared as follows.
First, �20 mL of the test culture was spun down and collectedinto 1 pellet. That pellet was then resuspended in 20 mL 1�PBS and vortexed until the pellet was completely broken.Twelve milliliter were divided equally into 12 centrifuge tubesof 1 mL each for heat and chemical treatments. Four milliliterswere divided into glass jars of 2 mL each for autoclave treat-ment. Five microliter from each culture sample was platedprior to treatment as controls. All spins were 20 000g 4�C30 minutes.
Autoclaved SamplesTwo milliliter sample was autoclaved in a glass jar for 1
liquid cycle. After autoclaving, the sample was collected andspun, the pellet was resuspended in 5 mL media. For auto-claved control, 2 mL sample was refrigerated while the culturesample was being autoclaved. The sample was then spundown and resuspended in 5 mL media.
Heat-Treated SamplesThree of the 1-mL samples were placed in a heating
block at 95�C. The samples were removed and moved to therefrigerator at 4�C after the predetermined heating time: 1sample after 2 minutes, 1 after 5 minutes, and 1 after60 minutes. As a control, a fourth tube was refrigerated duringthe whole duration. After treatment, the samples were spundown and resuspended in 5 mL media.
Samples Under Acid ConditionsTest cultures in Brucella media were prepared at pH 2,
5, and 7. To prepare pH 5, �200mL 1 N HCl was added to20 mL Brucella media. To prepare pH 2 media,�1400mL 1 NHCl was added to 20 mL Brucella media. Three 1-mL sampleswere spun down and resuspended: 1 in 5 mL of standard me-dia, 1 in pH 5 media, and 1 in pH 2 media.
Formalin-Treated SamplesFormalin dilutions were prepared by dilution of standard
37% formaldehyde in PBS. To make 4% formaldehyde,1.08 mL 37% formaldehyde was added to 8.92 mL PBS for atotal of 10 mL. To make 10% formaldehyde, 2.70 mL 37%formaldehyde was added to 7.3 mL PBS for a total of 10 mLsolution. Two 1-mL samples were spun down and resuspendedin formaldehyde solution, 1 in 4% and 1 in 10%. A third 1-mLsample was spun down and resuspended in PBS to serve as acontrol. All 3 tubes were kept in a heat block at 37�C over-night. Samples were then spun down and resuspended in 5 mLof media.
Bastian et al J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019
994
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
Triton X-100-Treated SamplesTriton X-100 was diluted to 1% in PBS. One 1-mL sam-
ple was spun down and resuspended in 1% triton X-100. Asecond 1-mL was spun down and resuspended in PBS to be acontrol. Both samples were kept in a heat block at 37�C over-night. Samples were then spun down and resuspended in 5 mLmedia.
Sodium Dodecyl Sulfate-Treated SamplesA small pellet of spiroplasma TSE isolate was sus-
pended in 567mL of TE buffer, 30mL of 10% sodium dodecylsulfate (SDS), and 3mL of proteinase K (20 mg/mL).
Guanidine Hydrochloride-Treated SamplesA small pellet of spiroplasma TSE isolate was resus-
pended in 600mL of 4 M guanidine hydrochloride and100 mM Tris-HCl pH 9 buffer (61.14 g of GuHCl, 0.8 g N-lau-ryl sarcosine, 1702 g of NaCl, 0.707 g Tris-HCl, 1.395 Tris-base in 160 mL water). Both SDS- and GuHCl-treated sampleswere heated at 55�C for 24 hours. Once cooled to room tem-perature, 5-mL drops were plated directly on Brucella agarplates, air dried for 10 minutes then the plates were sealedwith parafilm and incubated for 2 weeks. The plates were ex-amined using an inverted microscope with phase microscopyand no stain.
Cobalt-60-Treated SamplesA pellet of TSE spiroplasma isolate was collected and
resuspended in PBS in 400mL screw top freezer tubes, thenplaced in a secondary larger plastic tube before being sent forradiation. Sample tubes and controls were sent for radiationtreatment and positive and negative controls were refrigeratedas well. Before being sent, 5mL was plated on agar from eachtube to confirm colony presence before radiation. Once sent,sample tubes and PBS control were irradiated up to 20 kGy.Therefore, at the end, we would have 2 tubes for each treat-ment (1 tube positive and 1 tube PBS-negative control) atwhatever kGy treatments we were trying. The treatments werealways done the same way. A drop assay was done with 5mLplaced on the agar plates and incubated at 35�C for 1 week.The samples (irradiated and control) were also regrown bytaking 100mL of the treated sample and adding 300mL of me-dia to it in a 1.5-mL centrifuge tube. After incubation at 35�Cfor 1.5 weeks, the samples were spun, combined, and platedon agar (drop assay) (8).
Radiation ProcedureThe gamma irradiation was conducted using a JL Shep-
herd Associates Irradiator Model 484 (JL Shepard & Associ-ates, San Fernando, CA). The source was cobalt-60. Theexposure rate was 1,743 R/minute at the center of the verticalline (i.e. 5-in) above the turntable at position #2 (9). Exposureto dose conversion (in air): 1 R o 0.87 rad (10) (SupplementaryData S1).
Dot Blot ProcedureThe reagents included TBS (20 mM Tris-HCl 150 mM
NaCl pH 7.5), TBS-T (0.05% Tween 20 in TBS), BSA/TBS-T(0.1% BSA in TBS-T), and nitrocellulose membrane (Bio-Rad, Hercules, CA). The detailed procedure is as follows: Agrid was drawn on a nitrocellulose membrane and 2 lL ofeach of the samples was pipetted onto the nitrocellulose mem-brane. The membrane was dried and nonspecific sites wereblocked by soaking in 5% BSA in TBS-T (0.5–1 hour at roomtemperature). The membrane was incubated with primarymonoclonal prion antibody (0.1–10 lg/mL for purified anti-body) (Cayman Chemical Company Clone SAF 83 [CaymanChemical Company, Ann Arbor, MI]) and washed 3 timeswith TBS-T (3� 5 minutes). The membrane was incubatedwith secondary antibody conjugated with horseradish peroxi-dase, washed 3 times with TBS-T (15 minutes� 1,5 minutes� 2), then once with TBS (5 minutes).
Negative Stain Electron MicroscopyA drop of log phase culture (as determined by dark-field
examination of a 20-day-old culture) was placed on aformvar-coated copper grid and stained with phosphotungsticacid hematoxylin (PTAH). The grid was examined when dryusing a JEOL transmission electron microscope (JEM1011)(JEOL USA, Inc., Peabody, MA).
Extraction of Nucleic Acids From a PelletCell pellets were washed in PBS and suspended in
0.5 mL of the buffer. SDS was added to bring the final concen-tration of SDS at 0.1%. An equal volume of phenol was addedto the lysate. After mixing, a brief spin in a microfuge sepa-rated the phases (phenol on bottom, aqueous layer on top).The aqueous layer (leaving the interphase behind) was re-moved using a pipetman and placed into a new tube. This pro-cedure was repeated twice. When the interphase looked clean,an extraction was done with a 50/50 mix of phenol-chloroform instead of straight phenol. An extraction was doneonce with straight chloroform (chloroform phase on top, aque-ous on the bottom). One-tenth volume of 3 M sodium acetate(pH 8.0) was added and 3 volumes of ethanol. The mixturewas placed in a –20�C freezer for 1 hour, and then centrifugedat high speed for 5–10 minutes. The solution was drawn off,and a small DNA pellet visible in the bottom of the tube wassuspended in TE buffer (protocol according to Dr. Kevin Dyb-vig, Professor of Medicine, Program in Immunology, Univer-sity of Alabama Medical School [UAB]). The nucleic acidextraction shown was also successful using the QiagenDNeasy Power Soil Kit (Qiagen, Hilden, Germany) accordingto kit instructions.
RESULTSOur success in consistently isolating a novel Spiro-
plasma sp. from CWD-affected lymph nodes has allowed usto begin characterizing this organism that we believe is a can-didate causal agent of TSE. One question is whether the prionamyloid is associated with the bacterium. We tested this withimmunoblot using monoclonal antibody against the prion
J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019 CWD Isolate Is an Extreme Bacterial Thermoacidophile
995
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
amyloid. These studies showed that the pellets of organismsfrom the cultured spiroplasma CWD isolate showed no associ-ated prion proteins (Fig. 1). This information will be importantin future studies when these CWD spiroplasma isolates areused to develop animal models, which are needed not only tostudy the infection in either rodent or ruminant models, but tovalidate diagnostic tests and preventive vaccines based uponthe bacterial isolate.
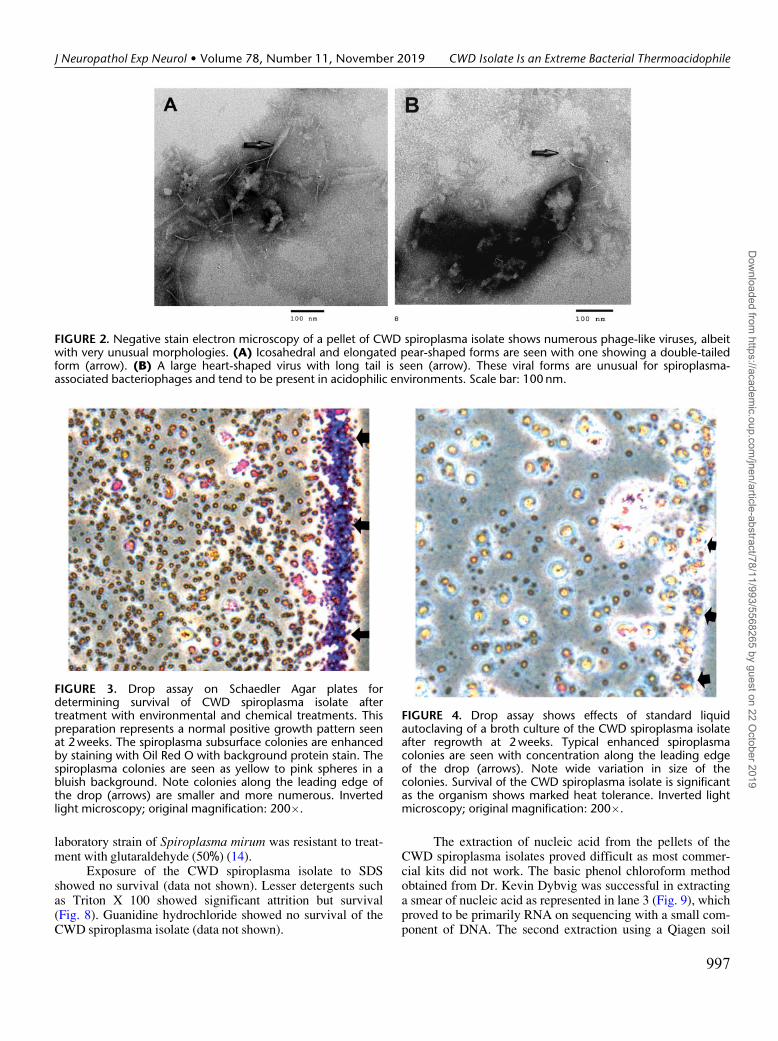
In further studies of the Spiroplasma CWD isolates, weexamined pellets of the organisms growing in the Brucellabroth at peak infection at 2 weeks postinoculation by negativestain electron microscopy. As shown in Figure 2, unusual vari-eties of phage-like viruses were found. These were unlike thetypical icosahedral or rod shaped spiroplasma bacteriophages(11). Instead, there were pear-shaped forms and long-tailedicosahedral viruses. An unusual double-tailed virus was seen.The viruses with these unusual forms are usually from an aci-dophilic environment and (12) may involve both bacteria andarchaea. Acidophiles are usually thermophiles, as in this case.These viruses may be important in occurrence of these ex-treme biologic properties (13) of the CWD spiroplasmaisolate.
Oil Red O staining procedure for detection of the tinyspiroplasma subsurface colonies on the agar showed the spiro-plasma growth as distinct yellow to red stained spiroplasmacircular subsurface colonies that are easily separated from blu-ish background; stain enhancing identification and enumera-tion of the spiroplasma colonies derived from TSE samples(7). The use of this modification of our method is superior tosimple phase microscopy since this Oil Red O stains
cholesterol and other membrane lipids in the membranes ofspiroplasma organisms embedded in the individual colonies.
The live drop assay that we developed clearly showsgrowth by the formation of the spiroplasma-specific roundsubsurface colonies that are enhanced by the Oil Red O stain-ing of membranes lipids of the organism (Fig. 3). The coloniesalong the leading edge of the drop are smaller and more nu-merous. Since it would be difficult to enumerate these colo-nies, this study is presented as a qualitative test whereinestimates of the effect of each environmental treatment aremade. Quantitative studies will be done in future studies usingother means.
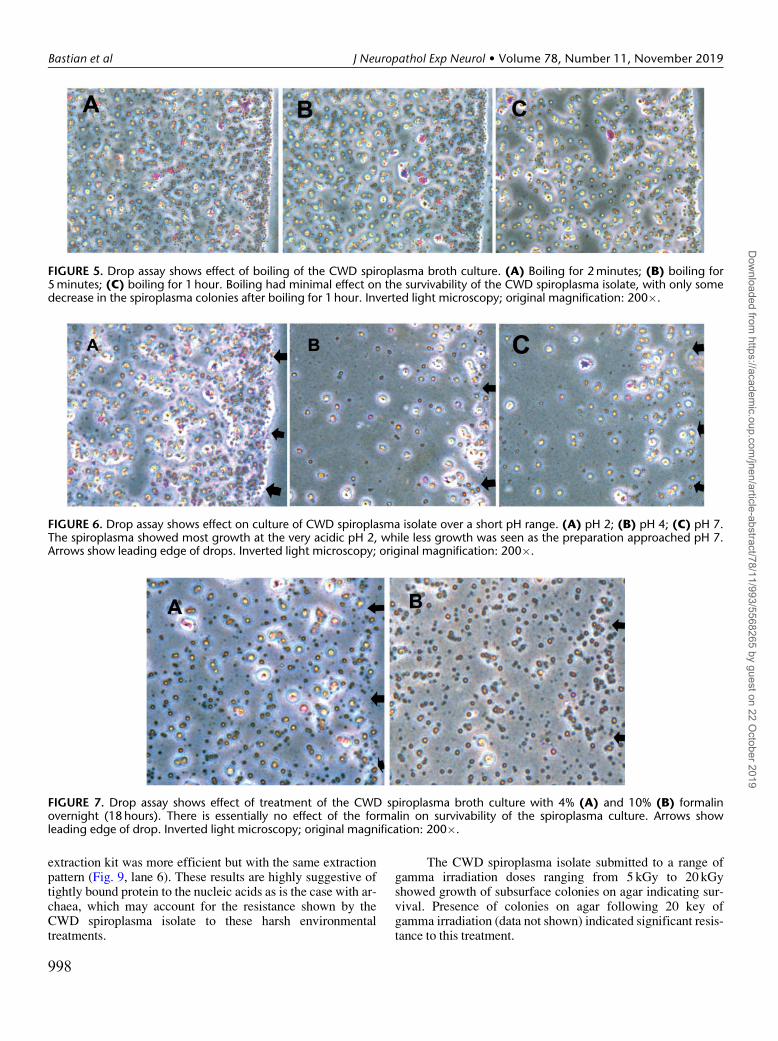
The assay shows the CWD spiroplasma isolate to be athermophile in that it survives extreme heat treatments. Theorganism survives standard liquid autoclaving (Fig. 4). Collec-tions are seen along the leading edge of the drop and there ismore variability in the size of the colonies compared with thecontrol in Figure 3. The survivability of the CWD spiroplasmaisolate in this case is significant in that the protein (prion) onlyhypothesis was solely based upon the supposition that a con-ventional microbe could not survive autoclaving. Similarly,the CWD spiroplasma isolate was not affected by boiling(Fig. 5A–C), confirming that it is an extreme thermophile.
Testing of the CWD spiroplasma isolate to acidityshows most growth in extreme acidic conditions (pH 2)(Fig. 6A–C), indicating that it is an extreme acidophile (i.e.survives highly acidic pH 3 and below) (13).
Formalin treatment had minimal effect on the surviv-ability of the CWD spiroplasma isolate, at either 4% or 10%exposure (Fig. 7A, B). We have previously reported that the
FIGURE 1. Dot blot with monoclonal prion antibody shows no prion amyloid in a lysate of the CWD spiroplasma isolate.Columns 1, 2, and 3 are the CWD-affected deer lymph node, control sheep brain, and culture lysate of the CWD spiroplasmaisolate, respectively, treated with proteinase K for 1 hour at 37�C. Columns 4, 5, and 6 are the same samples in the same orderbut not treated with proteinase K. Row A: Undiluted samples. Row B: Samples diluted 1:10. Row C: Samples diluted 1:1000. RowD: The lysis buffer by itself; this is a negative control to check for background. The deer CWD-infected lymph node and thenormal sheep brain stain positive for prion without proteinase K treatment; however, with proteinase K treatment, the CWD-affected lymph node is the only positive sample.
Bastian et al J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019
996
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
laboratory strain of Spiroplasma mirum was resistant to treat-ment with glutaraldehyde (50%) (14).
Exposure of the CWD spiroplasma isolate to SDSshowed no survival (data not shown). Lesser detergents suchas Triton X 100 showed significant attrition but survival(Fig. 8). Guanidine hydrochloride showed no survival of theCWD spiroplasma isolate (data not shown).
The extraction of nucleic acid from the pellets of theCWD spiroplasma isolates proved difficult as most commer-cial kits did not work. The basic phenol chloroform methodobtained from Dr. Kevin Dybvig was successful in extractinga smear of nucleic acid as represented in lane 3 (Fig. 9), whichproved to be primarily RNA on sequencing with a small com-ponent of DNA. The second extraction using a Qiagen soil
FIGURE 2. Negative stain electron microscopy of a pellet of CWD spiroplasma isolate shows numerous phage-like viruses, albeitwith very unusual morphologies. (A) Icosahedral and elongated pear-shaped forms are seen with one showing a double-tailedform (arrow). (B) A large heart-shaped virus with long tail is seen (arrow). These viral forms are unusual for spiroplasma-associated bacteriophages and tend to be present in acidophilic environments. Scale bar: 100 nm.
FIGURE 3. Drop assay on Schaedler Agar plates fordetermining survival of CWD spiroplasma isolate aftertreatment with environmental and chemical treatments. Thispreparation represents a normal positive growth pattern seenat 2 weeks. The spiroplasma subsurface colonies are enhancedby staining with Oil Red O with background protein stain. Thespiroplasma colonies are seen as yellow to pink spheres in abluish background. Note colonies along the leading edge ofthe drop (arrows) are smaller and more numerous. Invertedlight microscopy; original magnification: 200�.
FIGURE 4. Drop assay shows effects of standard liquidautoclaving of a broth culture of the CWD spiroplasma isolateafter regrowth at 2 weeks. Typical enhanced spiroplasmacolonies are seen with concentration along the leading edgeof the drop (arrows). Note wide variation in size of thecolonies. Survival of the CWD spiroplasma isolate is significantas the organism shows marked heat tolerance. Inverted lightmicroscopy; original magnification: 200�.
J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019 CWD Isolate Is an Extreme Bacterial Thermoacidophile
997
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
ctober 2019
extraction kit was more efficient but with the same extractionpattern (Fig. 9, lane 6). These results are highly suggestive oftightly bound protein to the nucleic acids as is the case with ar-chaea, which may account for the resistance shown by theCWD spiroplasma isolate to these harsh environmentaltreatments.
The CWD spiroplasma isolate submitted to a range ofgamma irradiation doses ranging from 5 kGy to 20 kGyshowed growth of subsurface colonies on agar indicating sur-vival. Presence of colonies on agar following 20 key ofgamma irradiation (data not shown) indicated significant resis-tance to this treatment.
FIGURE 5. Drop assay shows effect of boiling of the CWD spiroplasma broth culture. (A) Boiling for 2 minutes; (B) boiling for5 minutes; (C) boiling for 1 hour. Boiling had minimal effect on the survivability of the CWD spiroplasma isolate, with only somedecrease in the spiroplasma colonies after boiling for 1 hour. Inverted light microscopy; original magnification: 200�.
FIGURE 6. Drop assay shows effect on culture of CWD spiroplasma isolate over a short pH range. (A) pH 2; (B) pH 4; (C) pH 7.The spiroplasma showed most growth at the very acidic pH 2, while less growth was seen as the preparation approached pH 7.Arrows show leading edge of drops. Inverted light microscopy; original magnification: 200�.
FIGURE 7. Drop assay shows effect of treatment of the CWD spiroplasma broth culture with 4% (A) and 10% (B) formalinovernight (18 hours). There is essentially no effect of the formalin on survivability of the spiroplasma culture. Arrows showleading edge of drop. Inverted light microscopy; original magnification: 200�.
Bastian et al J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019
998
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
No spiroplasma colonies grew on agar following treat-ment of the CWD spiroplasma isolate with guanidine hydro-chloride (data not shown) or following exposure to the anionicdetergent SDS (data not shown) indicating that these chemi-cals are effective in killing the CWD spiroplasma isolate.
A summary of the susceptibility of the CWD spiro-plasma isolates to various chemical and physical treatmentextremes is shown in the Table.
DISCUSSIONA novel Spiroplasma sp. isolated from CWD, CJD and
scrapie-affected tissues has been identified as a candidatecausal agent/s for TSEs based upon fulfillment of Koch’s pos-tulates of causality (3). The assessment of the spiroplasma rolein the pathogenesis of TSE also fits the Bradford Hill criteria(Supplementary Data S2). Critical to this conclusion is that thespiroplasma CWD isolates studied in this report showed re-markable similarities of biologic properties ascribed to thetransmissible scrapie agent as reported in the 1960–1970s (5,6). We used a newly developed live assay wherein the viabilityof the bacterium was determined by its ability to grow in Bru-cella media and to form typical spiroplasma subsurface colo-nies on special agar plates (3). Using this methodology, wedemonstrated the spiroplasma CWD isolates to be resistant toextremes in chemical and environmental treatments. Thenovel spiroplasma CWD isolate was resistant to heatingextremes, surviving boiling for 1 hour. It is noteworthy thatthere appeared to be minimal effect of heating on survivabilityof the spiroplasma CWD cultures as determined by numbersof colonies. In this context it was not surprising that the spiro-plasma CWD isolate survived a standard liquid autoclavingcycle. These studies were repeated at least 3 times. Testing ofvarious pH levels revealed that the more vigorous growth ofthe CWD isolate was seen at pH 2, indicating that this novelspiroplasma isolate is a distinctly acidophilic bacterium. Ex-posure of the CWD isolate to 10% formalin treatment for18 hours caused a slight decrease in numbers of colonies. Pre-liminary studies also showed a remarkable resistance of thebacterium to gamma irradiation (20 000 gy), which is in therange of the radioresistant Deinococcus radiodurans (15, 16).Note that this degree of radio resistance compares with theearly TSE studies (5) since irradiation of tissue homogenatesis greater by a factor of 8 (17), whereas our studies were doneusing planktonic microbes. Indeed, future studies are neces-sary to fully characterize the resistance of this novel CWD spi-roplasma isolate using quantitative methods.
The TSE isolate is phenotypically a spiroplasma (3) butpossesses biologic properties unlike any known spiroplasmaspecies. The remarkable biologic resistance of the spiroplasmaCWD isolate could be due the association of unusualbacteriophage-like viruses seen in the pellets by negative stainelectron microscopy. These particular viruses have not beenseen in other spiroplasma preparations and bear close morpho-logical resemblance on negative stain electron microscopy toknown archaea viruses (18, 19), suggesting a relationship ofthis novel spiroplasma CWD isolate to this group of environ-mental resistant organisms. Also repeated nucleic acid extrac-tions from the pellets of the CWD spiroplasma isolate using
FIGURE 8. Drop assay shows effect of treatment of the CWDspiroplasma broth culture with Triton X100 overnight(18 hours). This detergent treatment showed moderatenegative effect on spiroplasma survivability with wide variationin colony size. Arrows show leading edge of drop. Invertedlight microscopy; original magnification: 200�.
FIGURE 9. Gel electrophoresis of nucleic acid extraction ofpellets of CWD spiroplasma isolate. Lane 1 are DNA 100 bpmarkers. The nucleic extractions of interest are seen inseparate samples as shown in lane 3 and lane 6 (Qiagen soilkit). The nucleic acid smear in both represents primarilyRNA pattern. Subsequent sequencing revealed mostly RNAwith a small component of DNA. These results werereproducible using the extraction procedure provided byDr. Dybvig.
J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019 CWD Isolate Is an Extreme Bacterial Thermoacidophile
999
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
several diverse methods were unsuccessful with recovery ofmostly a large RNA component. Sequencing revealed primar-ily RNA and a minimal DNA component. Preliminary bioin-formatic studies of the RNA sequence has revealed that theCWD isolate is a novel spiroplasma species related to but notidentical to the S. mirum clade. These data corroborate studiesthat showed no immune crossreactivity of the CWD or scrapieSpiroplasma isolates with S. mirum (14). The repeated failedattempts to extract sufficient DNA from the CWD isolate pel-lets for sequencing suggest that there may be an unusual com-plex binding of protein to the DNA similar to that describedfor archaea. Further molecular studies will be necessary to ad-dress the taxonomic classification of these novel spiroplasmaTSE isolates (16).
Although at this time we cannot definitively categorizethis organism, we are able, since it is growing in vitro, to pre-pare antibodies to surface epitopes and potentially make a se-rological diagnostic test (i.e. enzyme-linked immunosorbentassay [ELISA]) and potentially a hunter handheld instant testto identify CWD infection. Monoclonal antibodies to specificimmune reactive proteins on the membrane surface of the or-ganism could be used to make these tests very specific andperhaps be used to identify specific strains of the organism.Widespread use of such a test would produce accurate surveil-lance data regarding the epidemiology of CWD infection innature. Such information could be used to trace sources ofCWD infection. Pellets from multiple in vitro cultures of theSpiroplasma CWD isolate could be used to develop a preven-tive nasal or injectable vaccine to apply to captive cervidssuch as those housed by deer farmers or exotic ruminants pre-sent in zoo populations. These vaccines could be used to pre-vent the spread of CWD to cattle, to which CWD has beenexperimentally passaged (20). Protecting animals in the wildis another issue that may require development of an oral vac-cine or a darting program. The live assay for CWD infectionusing culture in Brucella media and as colonies on Bacto
Schaedler agar plates has application to detect the spiroplasmain grasses and dirt since these 2 entities may be involved in thespread of CWD. The diagnostic tests under development (de-scribed above) and the live test used in this study could be ap-plied for early diagnosis CJD patients, whereas at presentthere is no workable diagnostic test other than prion detectionin a brain biopsy or autopsy tissues (1). Hopefully, we will beable to control and possibly eliminate the CWD threat (21)and prevent destruction of the deer farming and hunting indus-tries. This research showing that CJD is a bacterial diseasewill allow aggressive antimicrobial therapy in early diagnosedcases of CJD.
ACKNOWLEDGMENTSMost of the experiments were done in Dr. Bastian’s lab-
oratory in the Department of Animal Sciences, LouisianaState University Agricultural Center during his tenure at thatinstitution. We thank Greg McCormick (Hansen Center) forassisting in electron microscopic studies.
REFERENCES1. Bastian FO. Creutzfeldt-Jakob Disease and Other Transmissible Spongi-
form Encephalopathies. New York, NY: Mosby/Year Book 19912. Bastian FO. The case for involvement of spiroplasma in the pathogenesis
of transmissible spongiform encephalopathies. J Neuropathol Exp Neurol2014;73:104–14
3. Bastian FO, Lynch J, Hagius S, et al. Novel Spiroplasma sp. culturedfrom brains and lymph nodes of ruminants affected with transmissiblespongiform encephalopathy. J Neuropathol Exp Neurol 2018;77:64–73
4. Bastian FO, Sanders DE, Forbes WA, et al. Spiroplasma spp. from trans-missible spongiform encephalopathy brains or ticks induce spongiformencephalopathy in ruminants. J Med Microbiol 2007;56:1235–42.
5. Gibbs C, Jr, Gajdusek DCLatarjet R. Unusual resistance to ionizing radi-ation of the viruses of kuru, Creutzfeldt-Jakob disease and scrapie (un-conventional viruses). Proc Natl Acad Sci USA 1978;75:6268–70
6. Zlotnik I, Rennie JC. The effect of heat on the scrapie mouse brain. Brit JExp Pathol 1967;48:171–9
8. Cember H. Introduction to Health Physics, 3rd Edition. New York, NY:The McGraw-Hill, Inc. 1996
9. Golashvili TV. Chart of the Nuclides, 15th Edition, GE Nuclear Energy.New York: KAPI., Inc. 1996
10. Johnson TA, Cember H. Introduction to Health Physics, 4th Edition.New York, NY: McGraw-Hill, Inc. 2009
11. Cole RM, Tully JG, Popkin TJ, et al. Morphology, ultrastructure, andbacteriophage infection of the helical mycoplasma-like organism (Spiro-plasmas citri gen. nov., sp. nov.) cultured from “stubborn” disease of cit-rus. J Bacteriol 1973;115:367–86
12. Cardenas JP, Valdes J, Quatrini R, et al. Lessons from the genomes of ex-tremely acidophilic bacteria and archaea with special emphasis on biol-eaching microorganisms. Appl Microbiol Biotechnol 2010;88:605–20
13. Mirete S, Morgante V, Gonz�alez-Pastor JE. Acidophiles: Diversity andmechanisms of adaptation to acidic enviroments. In: Stan-Lotter H, Fen-drihan S, eds. Adaption of Microbial Life to Environmental Extremes.Madrid, Spain: Springer International Publ 2017
14. Bastian FO, Boudreaux CM, Hagius SD, et al. Spiroplasma found in theeyes of scrapie affected sheep. Vet Ophthalmol 2011;14:10–7
15. Cox MM, Keck JL, Battista JR. Rising from the ashes: DNA repair inDeinococcus radiodurans. PLoS Gen 2010;6:1–2
16. Rainey FA, Ray K, Ferreira M, et al. Extensive diversity of ionizing-radiation-resistant bacteria recovered from Sonoran desert soil and de-scription of nine new species of the genus Deinococcus obtained from asingle soil sample. Appl Environ Microbiol 2005;71:5225–35
17. Da Silva Aquino KA. Sterilization by gamma irradiation. In: Adrovic F,ed. Gamma Radiation, Chapter 9. InTech Open, 2012:171–206. DOI:
TABLE. Susceptibility of the CWD Spiroplasma Isolates toChemical and Physical Treatment Extremes
Susceptibility of TSE Spiroplasma Isolate
Sample Exposure Survival Nonviable
Autoclaving (standard liquid) þþ 30 min (Fig. 4)
Gamma irradiation þ* 20 kgy
95�C þþþþ 2 min (Fig. 5A)
“ þþþþ 5 min (Fig. 5B)
“ þþþ 1 hour (Fig. 5C)
ACID pH 2 þþþþ 1 week (Fig. 6A)
pH 4 þþ 1 week (Fig. 6B)
pH 7 þþ 1 week (Fig. 6C)
Formalin (4%) 37�C þþþþ Overnight (Fig. 7A)
Formalin (10%) 37�C þþþþ Overnight (Fig. 7B)
Guanidine hydrochloride 1 hour –*
Sodium dodecyl sulphate (SDS) 1 hour –*
Triton X detergent þþ Overnight (Fig. 8)
*Data not shown.
Bastian et al J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019
1000
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O
10.5772/34901. Available at: https://www.intechopen.com/books/gamma-radiation/sterilization-by-gamma-irradiation
18. Hochstein RA, Amenabar MJ, Munson-McGee JH, et al. Acidianus tailedspindle virus: A new archaeal large tailed spindle virus discovered byculture-dependent methods. J Virol 2016;90:3458–68
19. Pietila MK, Demina TA, Atanaova NS, et al. Archael viruses and bac-teriophages: Comparisons and contrasts. Trends Microbiol 2014;334–44
20. Hamir AN, Kunkle RA, Cutlip RC, et al. Experimental transmission ofchronic wasting disease agent from mule deer to cattle by the intracere-bral route. J Vet Diagn Invest 2005;17:276–81
21. Needham MD, Vaske JJ, Petit JD. Risk sensitivity and hunter perceptionsof chronic wasting disease risk and other hunting, wildlife, and healthrisks. Hum Dimens Wildlife 2017;22:197–216
J Neuropathol Exp Neurol • Volume 78, Number 11, November 2019 CWD Isolate Is an Extreme Bacterial Thermoacidophile
1001
Dow
nloaded from https://academ
ic.oup.com/jnen/article-abstract/78/11/993/5568265 by guest on 22 O