Plzf regulates limb and axial skeletalpatterningMaria Barna1, Nicola Hawe1, Lee Niswander2 & Pier Paolo Pandolfi1
The promyelocytic leukaemia zinc finger (Plzf) protein (encoded by the gene Zfp145) belongs to the POZ/zinc-fin-ger family of transcription factors. Here we generate Zfp145–/– mice and show that Plzf is essential for patterningof the limb and axial skeleton. Plzf inactivation results in patterning defects affecting all skeletal structures of thelimb, including homeotic transformations of anterior skeletal elements into posterior structures. We demonstratethat Plzf acts as a growth-inhibitory and pro-apoptotic factor in the limb bud. The expression of members of theabdominal b (Abdb) Hox gene complex, as well as genes encoding bone morphogenetic proteins (Bmps), is alteredin the developing limb of Zfp145–/– mice. Plzf regulates the expression of these genes in the absence of aberrantpolarizing activity and independently of known patterning genes. Zfp145–/– mice also exhibit anterior-directedhomeotic transformation throughout the axial skeleton with associated alterations in Hox gene expression. Plzf istherefore a mediator of anterior-to-posterior (AP) patterning in both the axial and appendicular skeleton and actsas a regulator of Hox gene expression.
1Department of Human Genetics and Molecular Biology Program, 2Howard Hughes Medical Institute and Molecular Biology Program, Memorial Sloan-Kettering Cancer Center, Sloan-Kettering Division, Graduate School of Medical Sciences, Cornell University, New York, New York, USA. Correspondenceshould be addressed to P.P.P. (e-mail: [email protected]).
IntroductionThe vertebrate limb develops from small buds that arise from thelateral plate mesoderm. Cells at the tip of the limb bud rapidlyproliferate, whereas mesenchyme cells at the base of the budbegin to terminally differentiate. A functionally defined zone ofpolarizing activity (ZPA) is located in the posterior limb budmesenchyme and is marked by the presence of Shh (ref. 1). Thisregion is important in mediating AP patterning of the developinglimb bud, but the signals acting downstream or independently ofShh signalling that are responsible for directly regulating thegrowth and patterning of the limb are not well characterized,although the Hox genes have been implicated.
In the axial structures, the Hox genes are expressed along theAP axis and are responsible for the specification of segmentalidentity of vertebral structures2. Gain-of-function and loss-of-function experiments have demonstrated that misexpression ofa given Hox gene results in a morphological transformation ofthe tissue into a structure that would have normally expressedthis gene2. In addition, ectopic expression of Hox genes causesalterations in both limb and axial skeletal patterning.
The gene ZFP145 (encoding PLZF) was identified by virtue ofchromosomal translocations involving RARA (encoding retinoicacid receptor-α) leading to acute promyelocytic leukaemia3
(APL). PLZF is a nuclear phosphoprotein, containing at the car-boxy terminus nine Krüppel-type zinc-finger domains, whichbind directly to DNA (refs 3–5). At the amino terminus, PLZFcontains a BTB/POZ domain that mediates self-association andtranscriptional repression through recruitment of nuclear co-repressors6–8. Here we show that Plzf is essential for patterning ofthe limb and axial skeleton and acts as a main upstream regulatorof Hox gene expression.
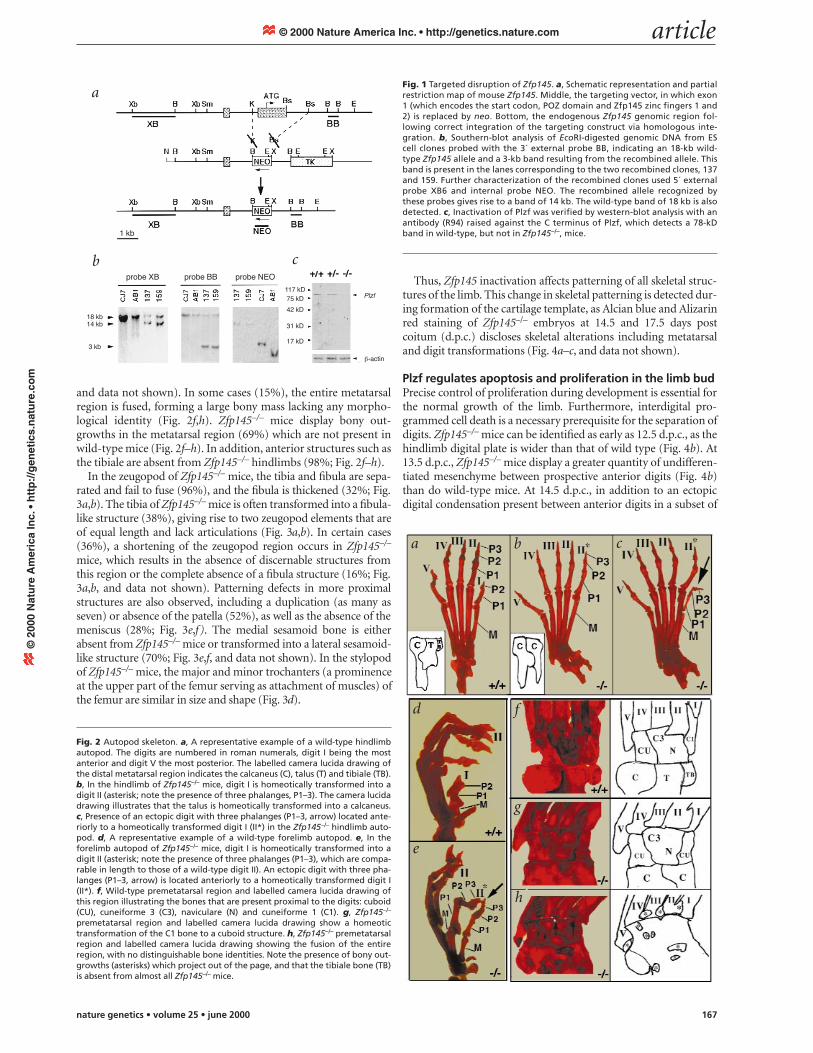
ResultsTargeted disruption of Zfp145 in embryonic stem cellsBy homologous recombination, we substituted exon 2 of Zfp145,which contains the 5´ region spanning the region encoding the POZdomain and Plzf zinc fingers 1 and 2, with a neomycin-resistancegene cassette (Fig. 1a,b). We demonstrated inactivation of Plzf bythe absence of any detectable protein from cells from Zfp145–/– miceby western-blot analysis (Fig. 1c, and data not shown).
Limb homeosis and patterning defects in Zfp145–/– miceThe vertebrate hindlimb is divided into three regions: stylopod(femur), zeugopod (tibia and fibula) and autopod (tarsals,metatarsals and phalanges). Zfp145–/– mice display morphologicaldefects affecting all the skeletal structures of the hindlimb, whereasskeletal defects in the forelimb occur at a lower frequency.
In the hindlimb autopod of these animals, digit I is homeoticallytransformed into a digit II (92%), as indicated by the presence of anadditional phalanx (Fig. 2a,b). In a subset of cases (40%), Zfp145–/–
mice possess an extra digit preaxially, which morphologicallyresembles a digit V in the hindlimb, or showed a complete absenceof digit I (Fig. 2c, and data not shown). The talus, which normallylies proximal to digits I, II and III, is also homeotically transformedinto a calcaneus (90%; Fig. 2a,b). Morphological defects in the fore-limb of Zfp145–/– mice occur at a lower frequency (5%) and arelargely restricted to the autopod. As with their hindlimb, Zfp145–/–
mice display a homeotic transformation of digit I into a digit II inthe forelimb and an additional digit is present preaxially, whichmorphologically resembles a digit V (Fig. 2d,e).
In the metatarsal region, homeotic transformations change theanterior C1 bone into a neighbouring naviculare bone (38%) orinto the most posterior structure, the cuboid bone (42%; Fig. 2f,g,
and data not shown). In some cases (15%), the entire metatarsalregion is fused, forming a large bony mass lacking any morpho-logical identity (Fig. 2f,h). Zfp145–/– mice display bony out-growths in the metatarsal region (69%) which are not present inwild-type mice (Fig. 2f–h). In addition, anterior structures such asthe tibiale are absent from Zfp145–/– hindlimbs (98%; Fig. 2f–h).
In the zeugopod of Zfp145–/– mice, the tibia and fibula are sepa-rated and fail to fuse (96%), and the fibula is thickened (32%; Fig.3a,b). The tibia of Zfp145–/– mice is often transformed into a fibula-like structure (38%), giving rise to two zeugopod elements that areof equal length and lack articulations (Fig. 3a,b). In certain cases(36%), a shortening of the zeugopod region occurs in Zfp145–/–
mice, which results in the absence of discernable structures fromthis region or the complete absence of a fibula structure (16%; Fig.3a,b, and data not shown). Patterning defects in more proximalstructures are also observed, including a duplication (as many asseven) or absence of the patella (52%), as well as the absence of themeniscus (28%; Fig. 3e,f ). The medial sesamoid bone is eitherabsent from Zfp145–/– mice or transformed into a lateral sesamoid-like structure (70%; Fig. 3e,f, and data not shown). In the stylopodof Zfp145–/– mice, the major and minor trochanters (a prominenceat the upper part of the femur serving as attachment of muscles) ofthe femur are similar in size and shape (Fig. 3d).
Thus, Zfp145 inactivation affects patterning of all skeletal struc-tures of the limb. This change in skeletal patterning is detected dur-ing formation of the cartilage template, as Alcian blue and Alizarinred staining of Zfp145–/– embryos at 14.5 and 17.5 days postcoitum (d.p.c.) discloses skeletal alterations including metatarsaland digit transformations (Fig. 4a–c, and data not shown).
Plzf regulates apoptosis and proliferation in the limb budPrecise control of proliferation during development is essential forthe normal growth of the limb. Furthermore, interdigital pro-grammed cell death is a necessary prerequisite for the separation ofdigits. Zfp145–/– mice can be identified as early as 12.5 d.p.c., as thehindlimb digital plate is wider than that of wild type (Fig. 4b). At13.5 d.p.c., Zfp145–/– mice display a greater quantity of undifferen-tiated mesenchyme between prospective anterior digits (Fig. 4b)than do wild-type mice. At 14.5 d.p.c., in addition to an ectopicdigital condensation present between anterior digits in a subset of
Fig. 1 Targeted disruption of Zfp145. a, Schematic representation and partialrestriction map of mouse Zfp145. Middle, the targeting vector, in which exon1 (which encodes the start codon, POZ domain and Zfp145 zinc fingers 1 and2) is replaced by neo. Bottom, the endogenous Zfp145 genomic region fol-lowing correct integration of the targeting construct via homologous inte-gration. b, Southern-blot analysis of EcoRI-digested genomic DNA from EScell clones probed with the 3´ external probe BB, indicating an 18-kb wild-type Zfp145 allele and a 3-kb band resulting from the recombined allele. Thisband is present in the lanes corresponding to the two recombined clones, 137and 159. Further characterization of the recombined clones used 5´ externalprobe XB6 and internal probe NEO. The recombined allele recognized bythese probes gives rise to a band of 14 kb. The wild-type band of 18 kb is alsodetected. c, Inactivation of Plzf was verified by western-blot analysis with anantibody (R94) raised against the C terminus of Plzf, which detects a 78-kDband in wild-type, but not in Zfp145–/–, mice.
Fig. 2 Autopod skeleton. a, A representative example of a wild-type hindlimbautopod. The digits are numbered in roman numerals, digit I being the mostanterior and digit V the most posterior. The labelled camera lucida drawing ofthe distal metatarsal region indicates the calcaneus (C), talus (T) and tibiale (TB).b, In the hindlimb of Zfp145–/– mice, digit I is homeotically transformed into adigit II (asterisk; note the presence of three phalanges, P1–3). The camera lucidadrawing illustrates that the talus is homeotically transformed into a calcaneus.c, Presence of an ectopic digit with three phalanges (P1–3, arrow) located ante-riorly to a homeotically transformed digit I (II*) in the Zfp145–/– hindlimb auto-pod. d, A representative example of a wild-type forelimb autopod. e, In theforelimb autopod of Zfp145–/– mice, digit I is homeotically transformed into adigit II (asterisk; note the presence of three phalanges (P1–3), which are compa-rable in length to those of a wild-type digit II). An ectopic digit with three pha-langes (P1–3, arrow) is located anteriorly to a homeotically transformed digit I(II*). f, Wild-type premetatarsal region and labelled camera lucida drawing ofthis region illustrating the bones that are present proximal to the digits: cuboid(CU), cuneiforme 3 (C3), naviculare (N) and cuneiforme 1 (C1). g, Zfp145–/–
premetatarsal region and labelled camera lucida drawing show a homeotictransformation of the C1 bone to a cuboid structure. h, Zfp145–/– premetatarsalregion and labelled camera lucida drawing showing the fusion of the entireregion, with no distinguishable bone identities. Note the presence of bony out-growths (asterisks) which project out of the page, and that the tibiale bone (TB)is absent from almost all Zfp145–/– mice.
Zfp145–/– mice (30%), all Zfp145-mutant hindlimbs show a delayedregression of interdigital mes-enchyme (Fig. 4b). We confirmedthat apoptosis of interdigital regionswas reduced in Zfp145–/– mice byTUNEL and Nile blue staining. Celldeath is not detected between digits Iand II of Zfp145–/– hindlimbs at 13.5d.p.c. and 14.5 d.p.c. (Fig. 4c, anddata not shown). Furthermore, adultZfp145–/– mice display interdigitalwebbing in hindlimb (60%) andforelimb (5%), largely between digitsI and II (Fig. 4c).
Zfp145–/– mice also displaychanges in cell proliferation. Flowcytometric analysis revealed anincrease in the percentage of cellsthat incorporate BrdU, in utero, in the hindlimb autopod ofZfp145–/– mice at 13.5 d.p.c. and 14.5 d.p.c. (Fig. 4d). No differ-ences in proliferation are apparent in Zfp145–/– mice at 15.5 d.p.c.(data not shown), when patterning of the digit primordia is com-plete. Immunohistochemistry of hindlimb tissue sections with theKi-67 antibody revealed an increased percentage of proliferatingcells in the interdigital regions of Zfp145–/– mice at 13.5 d.p.c.(data not shown). Thus, Plzf inhibits proliferation and functionsas a pro-apoptotic factor in the limb.
Expression of Zfp145 in developing limbMorphological alterations in Zfp145–/– hindlimb correlate withthe regions of Zfp145 expression in the limb. Onset of Zfp145
expression is concomitant with limb-bud formation, andZfp145 is expressed at 10.5 d.p.c. and 11.5 d.p.c. throughout thelimb mesenchyme. At 12.5 and 13.5 d.p.c., Zfp145 expression ishighest in the interdigital regions surrounding the perichon-drium of prospective digit anlages (Fig. 5).
Homeotic transformations are not caused by ectopicpolarizing activityThe gene Shh is involved in pattern formation along the AP limbaxis9,10. Shh and components of the Shh signalling pathway, suchas Ptch and Gli, are expressed normally in Zfp145–/– limbs (Fig. 6b,and data not shown). In addition, Fgf4 and Fgf8 transcripts arenot anteriorized in Zfp145–/– limbs (Fig. 6b, and data not shown).
Transplantation of anterior meso-derm from mouse mutants that pos-sess an ectopic polarizing region to theanterior margin of a host chick limbbud results in digit duplications9.Inactivation of Plzf does not result inectopic polarizing activity, as a normaldigit pattern is observed following agraft of anterior mesenchyme fromZfp145–/– mice (n=7) or wild-type lit-termates (n=38) to a chick limb bud(data not shown). These results sug-gest that Plzf acts downstream or inde-pendently of Shh to regulate limbpatterning.
Fig. 3 Zeugopod skeleton. a, Wild-type hindlimb zeugopod region with a normally fused tibia (T) and fibula (F). b, AZfp145–/– hindlimb zeugopod in which the fibula is thickened and the tibia has been homeotically transformed into afibula-like structure. c, Wild-type hindlimb. d, Zfp145–/– hindlimb. The brackets and arrows highlight the zeugopod.Only rudimentary tibia and fibula structures are present in the Zfp145–/– zeugopod. Note the equal size and shape ofthe major and minor trochanter of the femur (Fe) as indicated by an asterisk. e,f, Knee region of the hindlimb of awild-type and a Zfp145–/– mouse, respectively. Note in Zfp145–/– limbs the presence of two patellas (P), which areshifted proximally, and the absence of the meniscus (me) and medial sesamoid (ms) bone (arrows).
Fig. 4 Increased proliferation and decreasedapoptosis in the hindlimb of Zfp145–/– mice. a, Alcian blue and Alizarin red staining of 16.5-d.p.c. Zfp145–/– embryos revealed skeletal alter-ations including the transformation of digit Iinto a digit II identity and ectopic skeletal con-densations (arrow). b, Scanning electronmicroscopy of the hindlimb of wild-type andZfp145–/– embryos. Roman numerals indicatedigit condensations. c, Nile blue staining in thehindlimb of Zfp145–/– and control mice at 13.5d.p.c. The arrow indicates the lack of Nile bluestaining between digits I and II of the Zfp145–/–
hindlimb. Bottom, an adult Zfp145–/– hindlimbwith interdigital webbing between digits I andII (arrow). d, Flow cytometric analysis of limb tis-sues, as assessed by BrdU uptake. Zfp145–/– micehad a statistically significant increase (P<0.002)in the number of BrdU-positive cells in the auto-pod of the hindlimb at 13.5 d.p.c. and 14.5d.p.c.
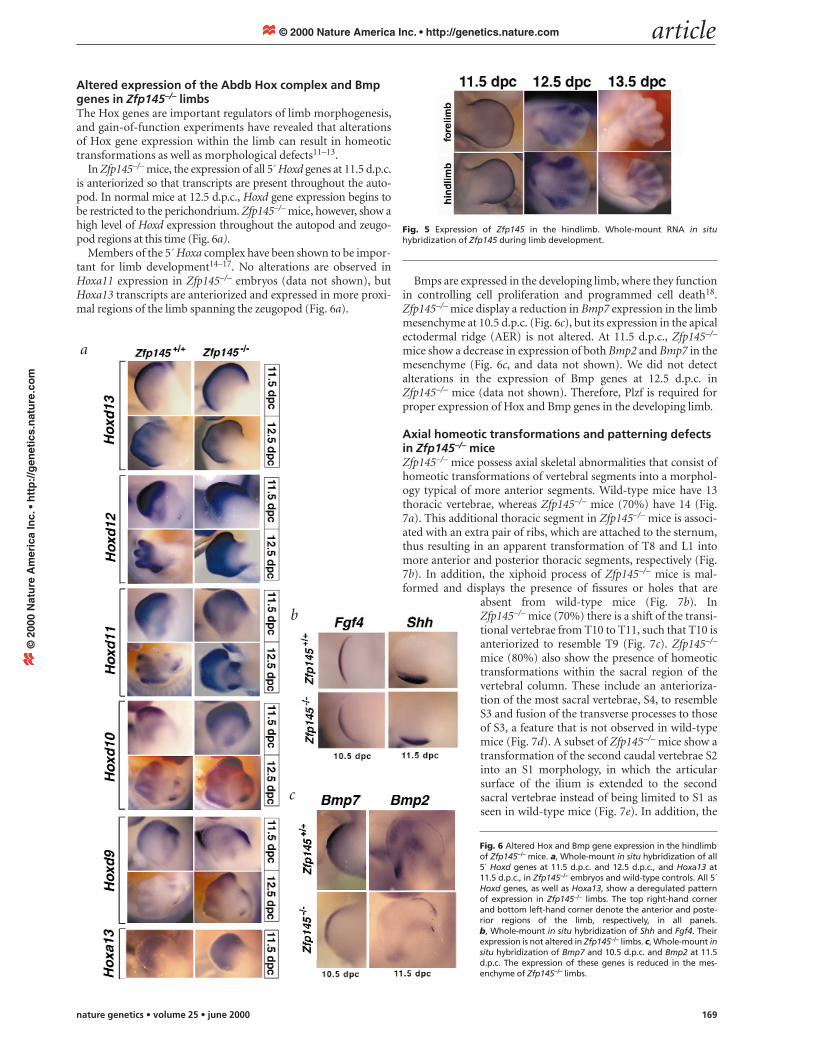
Altered expression of the Abdb Hox complex and Bmpgenes in Zfp145–/– limbsThe Hox genes are important regulators of limb morphogenesis,and gain-of-function experiments have revealed that alterationsof Hox gene expression within the limb can result in homeotictransformations as well as morphological defects11–13.
In Zfp145–/– mice, the expression of all 5´ Hoxd genes at 11.5 d.p.c.is anteriorized so that transcripts are present throughout the auto-pod. In normal mice at 12.5 d.p.c., Hoxd gene expression begins tobe restricted to the perichondrium. Zfp145–/– mice, however, show ahigh level of Hoxd expression throughout the autopod and zeugo-pod regions at this time (Fig. 6a).
Members of the 5´ Hoxa complex have been shown to be impor-tant for limb development14–17. No alterations are observed inHoxa11 expression in Zfp145–/– embryos (data not shown), butHoxa13 transcripts are anteriorized and expressed in more proxi-mal regions of the limb spanning the zeugopod (Fig. 6a).
Bmps are expressed in the developing limb, where they functionin controlling cell proliferation and programmed cell death18.Zfp145–/– mice display a reduction in Bmp7 expression in the limbmesenchyme at 10.5 d.p.c. (Fig. 6c), but its expression in the apicalectodermal ridge (AER) is not altered. At 11.5 d.p.c., Zfp145–/–
mice show a decrease in expression of both Bmp2 and Bmp7 in themesenchyme (Fig. 6c, and data not shown). We did not detectalterations in the expression of Bmp genes at 12.5 d.p.c. inZfp145–/– mice (data not shown). Therefore, Plzf is required forproper expression of Hox and Bmp genes in the developing limb.
Axial homeotic transformations and patterning defectsin Zfp145–/– miceZfp145–/– mice possess axial skeletal abnormalities that consist ofhomeotic transformations of vertebral segments into a morphol-ogy typical of more anterior segments. Wild-type mice have 13thoracic vertebrae, whereas Zfp145–/– mice (70%) have 14 (Fig.7a). This additional thoracic segment in Zfp145–/– mice is associ-ated with an extra pair of ribs, which are attached to the sternum,thus resulting in an apparent transformation of T8 and L1 intomore anterior and posterior thoracic segments, respectively (Fig.7b). In addition, the xiphoid process of Zfp145–/– mice is mal-formed and displays the presence of fissures or holes that are
absent from wild-type mice (Fig. 7b). InZfp145–/– mice (70%) there is a shift of the transi-tional vertebrae from T10 to T11, such that T10 isanteriorized to resemble T9 (Fig. 7c). Zfp145–/–
mice (80%) also show the presence of homeotictransformations within the sacral region of thevertebral column. These include an anterioriza-tion of the most sacral vertebrae, S4, to resembleS3 and fusion of the transverse processes to thoseof S3, a feature that is not observed in wild-typemice (Fig. 7d). A subset of Zfp145–/– mice show atransformation of the second caudal vertebrae S2into an S1 morphology, in which the articularsurface of the ilium is extended to the secondsacral vertebrae instead of being limited to S1 asseen in wild-type mice (Fig. 7e). In addition, the
Fig. 6 Altered Hox and Bmp gene expression in the hindlimbof Zfp145–/– mice. a, Whole-mount in situ hybridization of all5´ Hoxd genes at 11.5 d.p.c. and 12.5 d.p.c., and Hoxa13 at11.5 d.p.c., in Zfp145–/– embryos and wild-type controls. All 5´Hoxd genes, as well as Hoxa13, show a deregulated patternof expression in Zfp145–/– limbs. The top right-hand cornerand bottom left-hand corner denote the anterior and poste-rior regions of the limb, respectively, in all panels. b, Whole-mount in situ hybridization of Shh and Fgf4. Theirexpression is not altered in Zfp145–/– limbs. c, Whole-mount insitu hybridization of Bmp7 and 10.5 d.p.c. and Bmp2 at 11.5d.p.c. The expression of these genes is reduced in the mes-enchyme of Zfp145–/– limbs.
Fig. 5 Expression of Zfp145 in the hindlimb. Whole-mount RNA in situhybridization of Zfp145 during limb development.
transverse process of the second sacral vertebrae of Zfp145–/– miceresembles that of an S1 vertebrae in wild-type mice (Fig. 7e). Fur-thermore, Zfp145–/– mice show a loss in the number of caudal ver-tebrae, resulting in a shortening of the tail, and fusion of cartilagebetween vertebrae, causing a kinked tail (Fig. 7f). Plzf inactivationtherefore affects patterning of the axial skeleton.
Alterations in Hox gene expression in the AP axisLoss-of-function and overexpression experiments of Hox genes inthe mouse result in transformations of axial skeletal structures19,20.The paraxial expression patterns of both Hoxc6 and Hoxc8 areexpanded posteriorly in Zfp145–/– embryos, as the posterior bound-aries of expression of both genes are shifted by at least two preverte-brae (Fig. 8). Other Hox genes (such as Hoxb2) are expressednormally in the axial skeleton and limbs of Zfp145–/– embryos (Fig.8, and data not shown). Alterations in the expression patterns ofHox genes are associated with regions of Zfp145 expression, asZfp145 transcripts are present in both spinal cord and preverte-brae21. Zfp145 activity therefore regulates the expression of selectiveHox genes in both axial and appendicular skeletons.
DiscussionRole of Zfp145 in control of limb and axial skeletal patterning.Zfp145 controls AP patterning in the limb in both distal andproximal structures. This may be mediated, at least in part,through regulation of Hox genes, which are anteriorized andectopically expressed in Zfp145–/– mice. It has been previouslysuggested that Hox genes may mediate patterning of the limbalong the AP axis; however, Hox loss-of-function mutants largelyresult in truncation of specific skeletal elements and not AP pat-terning defects. In contrast, gain-of-function studies of individ-ual Hox genes result in AP patterning changes, largely restrictedto one or a few elements. Zfp145–/– mice display defects in limbpatterning, affecting skeletal elements along the entire proxi-modistal axis. The more extensive AP phenotype observed in
Zfp145–/– mice may be due to the concomitant alterations inexpression of the 5´ Hoxa and Hoxd genes. In the axial skeleton,Zfp145–/– mice also exhibit anterior-directed homeotic transfor-mation. The thoracic transformations are similar to those associ-ated with misexpression of Hoxc6 and Hoxc8, and in Zfp145–/–
embryos the posterior boundaries of expression of both genes aremisexpressed. Plzf can therefore regulate Hox gene expression inboth the axial and appendicular skeleton. In this regard, we haveidentified Zfp145 binding sites within the Abdb Hox gene locus(unpublished data) and it may be that Plzf acts as a transcrip-tional repressor to directly regulate Hox gene expression.
Plzf regulates apoptosis and proliferation in the developinglimb. Plzf also influences the expression of Bmp genes, which is
Fig. 7 Homeotic transformations and patterning defects in the axial skeleton ofZfp145–/– mice. a, Vertebral column of wild-type and Zfp145–/– mice. The totalnumber of thoracic vertebrae and ribs are increased in Zfp145–/– mice. b, Left,sternum with associated attached ribs in wild-type and Zfp145–/– mice. Note thepresence of an additional pair of attached ribs in Zfp145–/– mice. Right, close-upof the xiphoid process in wild-type and Zfp145–/– mice. The arrowhead indicatesthe presence of a hole or fissure in the xiphoid process of Zfp145–/– mice. c, Tran-sitional articular processes (arrow) of thoracic vertebrae in wild-type andZfp145–/– mice. Note the shift of the transitional vertebrae from T10 in wild typeto T11 in Zfp145–/– mice (asterisk). d, Alizarin red staining of adult sacral verte-brae in wild-type and Zfp145–/– mice. In Zfp145–/– mice, the morphology of thefourth sacral vertebrae (S3*) is altered as its transverse processes are completelyfused to those of S3 (filled arrowhead). e, Left, Alizarin red staining of adultsacral vertebrae in Zfp145–/– mice in which the second sacral vertebrae exhibits amorphology characteristic of a wild-type S1. Right, transitional processes ofsacral vertebrae seen in the left panel. Note the transformation of the transi-tional process (arrow) of the second sacral vertebrae (S1*) in Zfp145–/– mice tothat of a wild-type S1. f, Tail morphology of wild-type and Zfp145–/– mice. Notethat Zfp145–/– mice show a truncated and kinked tail.
Fig. 8 Expression of Hox genes within prevertebrae (Pv). Whole-mount in situhybridization is shown of Hoxc6, Hoxc8 and Hoxb2 in wild-type and Zfp145–/–
embryos at 12.5 d.p.c. Note the posterior expansion in Hoxc6 and Hoxc8expression domains in Zfp145–/– embryos. No differences in Hoxb2 expressionwere observed.
reduced in Zfp145–/– mice. Bmp7-null mice exhibit anterior limbpatterning defects, including the presence of an extra preaxialdigit22,23. The diminished expression of Bmp genes in Zfp145–/–
embryos may therefore also contribute to the limb skeletal pat-terning defects observed in these mice. In addition, Bmps maymediate the pro-apoptotic and growth inhibitory function ofZfp145. The misexpression of Hoxd genes within the interdigits ofZfp145–/– embryos at 12.5 d.p.c. may also result in decreased pro-grammed cell death, consistent with the retroviral overexpressionof Hoxd11 in the chick, which results in webbing of the digits11.
Morphological alterations in the forelimb of Zfp145–/– mice arepresent at a lower frequency (5%) than in the hindlimb (100%). Inthis respect, it is notable that two homologues of Zfp145 haverecently been identified24, one of which is highly expressed in fore-limb25. Therefore, the loss of Zfp145 function in the forelimb maybe compensated by the presence of Zfp145 homologues.
The ability of Zfp145 to regulate Hox and Bmp expressionappears to be downstream or independent of Shh. In contrast,other mouse mutants that exhibit posterior transformations ofanterior skeletal elements display ectopic polarizing activity, dueto either direct activation of Shh signalling or loss of a repressorof Shh activity. Plzf can therefore mediate patterning of the limbindependently or downstream of a polarizing signal.
Implications for leukaemogenesis. APL is characterized by theclonal expansion of malignant myeloid cells blocked at thepromyelocytic stage of haemopoietic development. As a conse-quence of translocations between chromosomes 11 and 17, tworeciprocal fusion genes are generated that encode PLZF–RARAand RARA–PLZF fusion proteins, which are co-expressed in theleukaemic blast26. Both proteins can act as dominant-negativeproducts on PLZF function26. Our findings therefore have impli-cations for the pathogenesis of APL, in which aberrant PLZFfunction might lend a selective growth and survival advantage tothe leukaemic cells through deregulated expression of PLZF tar-get genes. In this regard, HOX genes and BMPs have been impli-cated in the control of proliferation and differentiation ofprimitive haemopoietic cells27,28. Aberrant regulation of HOXgenes in APL may be exacerbated by the fact that PLZF–RARAcan concomitantly interfere with the transcriptional function ofRARA, which has also been implicated in the regulation of HOXgene expression.
MethodsGeneration and identification of Zfp145–/– mice. We isolated Zfp145genomic clones by screening a 129 Sv mouse genomic library (Stratagene)with a human ZFP145 probe. To generate the targeting construct, we sub-cloned a 10-kb BamHI fragment into pBluescript vector, in which the KpnIand XhoI sites had been deleted from the multiple cloning site. This facili-tated removal of a 3.5-kb KpnI/BspEI fragment and replacement with theneomycin cassette, thereby deleting the 5´ Plzf coding region. This was thensubcloned into the PYTK vector, which contains the TK gene29. The target-ing construct was electroporated into CJ7 ES cells. We selected transfec-tants in G418 (350 µg/ml) and gancyclovir (2 µM) and expanded them forSouthern-blot analysis. Chimaeric mice were produced by microinjectionof two independently generated, targeted ES cell clones with normal kary-otypes into 3.5-d.p.c. C57BL6/J blastocysts (Jackson Laboratories). We ver-ified germline transmission of the mutated allele by Southern-blot analysis.
Generation of Plzf antibody and western-blot analysis. We carried outimmunoblot analysis using the ECL western-blot kit (Amersham) follow-ing the manufacturer’s protocol. Whole-cell extracts from 10.5-d.p.c.Zfp145–/– and wild-type embryos were used for western-blot analysis. Thefilters were blocked with 10% nonfat dry milk in TBST for 1 h, incubatedwith anti-Plzf antibodies for 1–2 h at RT, washed 3 times in TBST and incu-bated with appropriate secondary antibodies conjugated to horseradishperoxidase. The western-blot signals were developed using reagents sup-
plied by the manufacturer. The filters were stripped with Tris-HCl (62.5mM, pH 6.8), 2% SDS and β-mercaptoethanol (100 mM) at 50 °C for 30min, and re-hybridized with an anti-β-actin antibody (1:3,000). We usedtwo antibodies for western-blot analysis: NPGP recognized the Plzf POZdomain30; and R94 recognized a peptide (KPEEIPPDWRIEKTY) located atthe very C terminus of the Plzf protein (aa 653–667).
Skeletal analysis of Zfp145–/– mice. For skeletal analysis, skin and internalviscera of 26 Zfp145–/– animals were removed. We then fixed the animalsovernight in ethanol and carried out Alcian blue and Alizarin red stainingprocedures as described31. The clearing step was modified to 2% potassiumhydroxide treatment overnight for adult mice.
Scanning electron microscopy. Embryos at different developmental stageswere dissected in PBS and fixed overnight in 4% paraformaldehyde and 1%glutaraldehyde in cacodylic buffer (0.1 M). After postfixation with 1%OsO4 in cacodylate buffer (0.1 M; 1 h at 4 °C), the embryos were dehydrat-ed by an ascending ethanol series and dried by the critical point method,sputtered-coated with palladium-gold and examined with a scanning elec-tron microscope.
Nile blue staining. We dissected 13.5- and 14.5-d.p.c. embryos in PBS,placed them into a solution of 1.5% Nile blue (Sigma) in DMEM (Gibco)for 45 min at RT, then placed them in PBS. The limbs were removed andphotographed from a dorsal perspective.
Flow cytometry. We injected BrdU labelling solution (Amersham)intraperitoneally into pregnant female mice at 13.5, 14.5 and 15.5 days ofgestation (1 ml/100 g body weight). One hour after injection, the mice werekilled and embryos dissected in PBS. A single-cell suspension of limb tissuewas obtained by tissue grinding. Cells were washed twice with PBS andfixed in 70% ethanol for 30 min, and washed again with PBS. A solution of2N HCl/0.5% Triton X-100 was added slowly while cells were gently vor-texed and incubated for 30 min. We added a solution of sodium borate (0.1M) to neutralize the acid and resuspended cells in PBS/1% BSA/0.5%Tween-20 for 30 min before staining. Cells were then incubated with FITC-conjugated anti-BrdU antibody for 30 min, vortexed, resuspended in PBSand labelled cells sorted on a FACS machine.
Polarizing activity assay. A piece of anterior or posterior mesenchyme(∼150 µm cube) from Zfp145–/– or wild-type littermate embryos(11.5–12.5 d.p.c.) was implanted under the AER on the anterior side of astage 19–20 chick limb bud. We fixed host embryos at day 8–10 of incuba-tion and stained the skeleton with Alcian blue. Digit duplications (patternranged from 2234 to 432234) were observed following posterior mes-enchyme graft from Zfp145–/– or wild-type limbs (n=15; genotype did notaffect the extent of duplication).
Whole-mount in situ hybridization. We carried out whole-mount in situhybridization using digoxygenin-labelled antisense RNA probes asdescribed32. We detected the label using an alkaline phosphatase-conjugat-ed anti-digoxygenin antibody (Boehringer).
AcknowledgementsWe thank D. Ruggero and S. Pizette for discussions; M. Branford and C.Kaduro for help with experiments; J.H. Dong and V. Soares for help with thegeneration and the management of the Zfp145 mutants; N. Lampin and B.Gumbiner for help and advice with the electron scanning microscopyanalysis; D. Duboule and P. Gruss for Hoxd and Hoxa probes, respectively;M. Scott and A. Joyner for Ptch and Gli probes, respectively; M. Capecchi andA. Boulet for Hoxc6 and Hoxc8 probes; and P.G. Pelicci and M. Ruthardt fora PLZF antibody and probe. P.P.P. is a Scholar of the Leukemia andLymphoma Society (formerly known as the Leukemia Society of America).L.N. is an Assistant Investigator of the Howard Hughes Medical Institute.This work was supported by the NIH (CA-08748 and CA-71692 awarded toSloan-Kettering Institute and P.P.P.).
1. Pearse, R.V. & Tabin, C.J. The molecular ZPA. J. Exp. Zoology 282, 677–690 (1998).2. Krumlauf, R. Hox genes in vertebrate development. Cell 78, 191–201 (1994).3. Chen, Z. et al. Fusion between a novel Krüppel-like zinc finger gene and the
retinoic acid receptor-α locus due to a variant t(11;17) translocation associatedwith acute promyelocytic leukaemia. EMBO J. 12, 1161–1167 (1995).
4. Li, J.Y. et al. Sequence-specific DNA binding and transcriptional regulation by thepromyelocytic leukemia zinc finger protein. J. Biol. Chem. 272, 22447–22455(1997).
5. Reid, A. et al. Leukemia translocation gene, PLZF, is expressed with a specklednuclear pattern in early hematopoietic progenitors. Blood 86, 4544–4552 (1995).
6. Chang, C.C., Ye, B.H., Chaganti, R.S. & Dalla-Favera, R. BCL-6, a POZ/zinc-fingerprotein, is a sequence-specific transcriptional repressor. Proc. Natl Acad. Sci. USA93, 6947–6952 (1996).
7. Hong, S.H., David, G., Wong, C.W., Dejean, A. & Privalsky, M.L. SMRT corepressorinteracts with PLZF and with the PML-retinoic acid receptor-α (RARα) and PLZF-RARα oncoproteins associated with acute promyelocytic leukemia. Proc. NatlAcad. Sci. USA 94, 9028–9033 (1997).
8. He, L.-Z. et al. Distinct interactions of PML-RARα and PLZF-RARα withtranscriptional co-repressors determine differential responses to retinoic acid inAPL. Nature Genet. 18, 126–135 (1998).
9. Tickle, C. The number of polarizing region cells required to specify additionaldigits in the developing chick wing. Nature 289, 295–298 (1981).
10. Riddle, R.D., Johnson, E.L. & Tabin, C. Sonic Hedgehog mediates the polarizingactivity of the ZPA. Cell 75, 1401–1416 (1993).
11. Morgan, B.A., Izpisua-Belmonte, J.C., Duboule, D. & Tabin, C.J. Targetedmisexpression of Hox-4.6 in the avian limb bud causes apparent homeotictransformations. Nature 358, 236–239 (1992).
12. Goff, D.J. & Tabin, C.J. Analysis of Hoxd-13 and Hoxd-11 misexpression in chicklimb buds reveals that Hox genes affect both bone condensation and growth.Development 124, 627–636 (1997).
13. Knezevic, V. et al. Hoxd-12 differentially affects preaxial and postaxialchondrogenic branches in the limb and regulates Sonic hedgehog in a positivefeedback loop. Development 124, 4523–4536 (1997).
14. Small, K.M. & Potter, S.S. Homeotic transformations and limb defects in HoxA11mutant mice. Genes Dev. 7, 2318–2328 (1993).
15. Yokouchi, Y. et al. Misexpression of Hoxa-13 induces cartilage homeotictransformation and changes cell adhesiveness in chick limb buds. Genes Dev. 9,2509–2522 (1995).
16. Fromental-Ramin, C. et al. Hoxa-13 and Hoxd-13 play a crucial role in thepatterning of the limb autopod. Development 122, 2997–3011 (1996).
17. Davis, A.P., Witte, D.P., Hsieh-Li, H.M., Potter, S.S. & Capecchi, M.R. Absence ofradius and ulna in mice lacking hoxa-11 and hoxd-11. Nature 375, 791–795(1995).
18. Zou, H. & Niswander, L. Requirement for BMP signaling in interdigital apoptosisand scale formation. Science 272, 738–741 (1996).
19. Pollack, R.A., Jay, G. & Bieberich, C.J. Altering the boundaries of Hox3.1expression: evidence for antipodal gene expression. Cell 71, 911–923 (1992).
20. Jegalian, B.G. & Robertis, E.M. Homeotic transformations in the mouse inducedby overexpression of a human Hox3.3 transgene. Cell 71, 901–910 (1992).
21. Avantaggiato, V. et al. Developmental analysis of murine promyelocyte leukemiazinc finger (PLZF) gene expression: implications for the neuromeric model of theforebrain organization. J. Neurosci.15, 4927–4942 (1995).
22. Luo, G. et al. BMP-7 is an inducer of nephrogenesis, and is also required for eyedevelopment and skeletal patterning. Genes Dev. 9, 2808–2820 (1995).
23. Dudley, A.T., Lyons, K.M. & Robertson, E.J.A. Requirement for bonemorphogenetic protein-7 during development of the mammalian kidney andeye. Genes Dev. 9, 2795–2807 (1995).
24. Hoatlin, M.E. et al. A novel BTB/POZ transcriptional repressor protein interactswith the Fanconi anemia group C protein and PLZF. Blood 94, 3737–3747 (1999).
25. Davis, J.M. et al. Novel BTB/POZ domain zinc-finger protein, LRF, is a potentialtarget of the LAZ-3/BCL-6 oncogene. Oncogene 18, 365–375 (1999).
26. He, L.Z., Merghoub, T. & Pandolfi, P.P. In vivo analysis of the molecularpathogenesis of acute promyelocytic leukemia in the mouse and its therapeuticimplications. Oncogene 18, 5278–5292 (1999).
27. Bhatia, M. et al. Bone morphogenetic proteins regulate the developmentalprogram of human hematopoietic stem cells. J. Exp. Med. 189, 1139–1148 (1999).
28. Van Oostveen, J., Bijl, J., Raaphorst, F., Walboomers, J. & Meijer, C. The role ofhomeobox genes in normal hematopoiesis and hematological malignancies.Leukemia 13, 1675–1690 (1999).
29. Tybulewicz, V.L., Crawford, C.E., Jackson, P.K., Bronson, R.T. & Mulligan, R.C.Neonatal lethality and lymphopenia in mice with a homozygous disruption of thec-abl proto-oncogene. Cell 65, 1153–1163 (1991).
30. Ruthardt, M. et al. Opposite effects of the acute promyelocytic leukemia PML-retinoic acid receptor α (RAR α) and PLZF-RAR α fusion proteins on retinoic acidsignalling. Mol. Cell. Biol. 17, 4859–4869 (1997).
31. Kessel, M. & Gruss, P. Homeotic transformations of murine vertebrae andconcomitant alteration of Hox codes by retinoic acid. Cell 67, 89–104 (1991).
32. Wilkinson, D.G. Whole-mount in situ hybridization of vertebrate embryo. in InSitu hybridization: A Practical Approach (ed. Wilkinson, D.G.) 75–83 (IRL, Oxford,1992).