PESTICIDE BIOCHEMISTRY AND PHYSIOLOGY 13, 121-128 (1980) Properties of Tribolium Gut Chitin Synthetasel EPHRAIM COHEN’ AND JOHN E. CASIDA Received November 21, 1979: accepted January 3, 1980 An insect chitin synthetase (CS) is readily assayed using the microsomal fraction (-0.5 mg protein) from an homogenate of Tribdium custuneum larvae. This enzyme preparation is incubated at 22°C with uridine 5’-diphospho-N-acetyl[~H]glucosamine in 355 ~1 of 25 m&f Tris-HCI buffer containing 10 mM MgCl,, 17 mM N-acetylglucosamine, and 1 n&I dithiothreitol. Other divalent cations and amino sugars are less effective activators or are inhibitory. T. cas~ane~rnr CS is strongly inhibited by polyoxin D and uridine 5’-diphosphate. These activation and inhibition properties of Triholiom custunrrrm gut CS are similar to those of fungal CS. The polymerization product formed by the Triholium enzyme is stable in alkali but hydrolyzed by chitinase. Enzymes of Tribohrm cor~firsrrm. Triholium hrevicornis. Tenrhrio molitor, and Gallrria mrllonel/a are also active under the same conditions. These enzymes are from the gut and probably from the peritrophic membrane. Integumental CS activity is not detected under the indicated assay conditions. INTRODUCTION Chitin is the major skeletal component of all arthropods and many fungi (l-4). Chitin synthetase (CQ3 (EC 2.4.1.16) carries out the polymerization step in chitin synthesis. This enzyme is readily assayed in many fungal systems (4) but there is only one series of studies reporting an insect CS (5, 6). Other workers failed with many species in attempts to assay insect CS (7-lo), a likely target site for benzoyl- phenyl ureas and a few other insecticides (7- 12). This report describes the assay and prop- erties of a cell-free CS system from tene- brionid larvae. ’ Study supported in part by National Institute of Environmental Health Sciences Grant PO1 ES00049. Presented in part at the Pesticide Chemistry Division, American Chemical Society, National Meeting, Honolulu, Hawaii, April 2-6, 1979. ” Permanent address: Department of Entomology, Faculty of Agriculture. The Hebrew University of Jerusalem. Rehovot. Israel. :( Abbreviations used: CS, chitin synthetase; dpm, disintegrations per minute; DTT, dithiothreitol; GlcNAc, N-acetylglucosamine: LSC, liquid scintilla- tion counting; UDP, uridine 5’-diphosphate; UDP- GlcNAc. uridine 5’.diphospho-N-acetylglucosamine; TCA. trichloroacetic acid: tic, thin-layer chromatog- raphy. MATERIALS AND METHODS Chemiculs UDP-[glucosamine-6-3H]GlcNAc (7 Ci/ mmol), D-[glucosamine-l-l”C]GlcNAc hy- drochloride, and [I-‘“Clglucosamine were from New England Nuclear (Boston, Mass.), and UDP-[glucosamine-1-‘“Cl- GlcNAc (31 mCi/mmol) was from ICN Chemical and Radioisotope Division (Ir- vine, Calif.). Unlabeled chemicals were from Sigma Chemical Co. (St. Louis, MO.) or Calbiochem-Behring Corp. (San Diego, Calif.). Insects Tribofium castaneum (Herbst) (CTC-12), T. brevicornis LeC. (Riverside +/+), and T. confusum duVa1 (Chicago +/+) were reared at 28°C on wheat flour supplemented with 7.5% casein. To maintain relatively homogeneous populations, eggs collected at 2- to 3-day intervals were transferred to fresh diets. Other insects used were from cultures in the Department of Entomologi- cal Sciences at Berkeley. Imaginal disks of Drosophila melanogaster Meigen were provided by J. W. Fristrom (Department of Genetics, University of California, Berke- ley). 121 0048.3575/80/020121-08$02.00/O Copyright 0 1980 by Academic Press. Inc. AU rights of reproduction in any form reserved.
Transcript
PESTICIDE BIOCHEMISTRY AND PHYSIOLOGY 13, 121-128 (1980)
Properties of Tribolium Gut Chitin Synthetasel
EPHRAIM COHEN’ AND JOHN E. CASIDA
Received November 21, 1979: accepted January 3, 1980
An insect chitin synthetase (CS) is readily assayed using the microsomal fraction (-0.5 mg protein) from an homogenate of Tribdium custuneum larvae. This enzyme preparation is incubated at 22°C with uridine 5’-diphospho-N-acetyl[~H]glucosamine in 355 ~1 of 25 m&f Tris-HCI buffer containing 10 mM MgCl,, 17 mM N-acetylglucosamine, and 1 n&I dithiothreitol. Other divalent cations and amino sugars are less effective activators or are inhibitory. T. cas~ane~rnr CS is strongly inhibited by polyoxin D and uridine 5’-diphosphate. These activation and inhibition properties of Triholiom custunrrrm gut CS are similar to those of fungal CS. The polymerization product formed by the Triholium enzyme is stable in alkali but hydrolyzed by chitinase. Enzymes of Tribohrm cor~firsrrm. Triholium hrevicornis. Tenrhrio molitor, and Gallrria mrllonel/a are also active under the same conditions. These enzymes are from the gut and probably from the peritrophic membrane. Integumental CS activity is not detected under the indicated assay conditions.
INTRODUCTION
Chitin is the major skeletal component of all arthropods and many fungi (l-4). Chitin synthetase (CQ3 (EC 2.4.1.16) carries out the polymerization step in chitin synthesis. This enzyme is readily assayed in many fungal systems (4) but there is only one series of studies reporting an insect CS (5, 6). Other workers failed with many species in attempts to assay insect CS (7-lo), a likely target site for benzoyl- phenyl ureas and a few other insecticides (7- 12).
This report describes the assay and prop- erties of a cell-free CS system from tene- brionid larvae.
’ Study supported in part by National Institute of Environmental Health Sciences Grant PO1 ES00049. Presented in part at the Pesticide Chemistry Division, American Chemical Society, National Meeting, Honolulu, Hawaii, April 2-6, 1979.
” Permanent address: Department of Entomology, Faculty of Agriculture. The Hebrew University of Jerusalem. Rehovot. Israel.
UDP-[glucosamine-6-3H]GlcNAc (7 Ci/ mmol), D-[glucosamine-l-l”C]GlcNAc hy- drochloride, and [I-‘“Clglucosamine were from New England Nuclear (Boston, Mass.), and UDP-[glucosamine-1-‘“Cl- GlcNAc (31 mCi/mmol) was from ICN Chemical and Radioisotope Division (Ir- vine, Calif.). Unlabeled chemicals were from Sigma Chemical Co. (St. Louis, MO.) or Calbiochem-Behring Corp. (San Diego, Calif.).
Insects
Tribofium castaneum (Herbst) (CTC-12), T. brevicornis LeC. (Riverside +/+), and T. confusum duVa1 (Chicago +/+) were reared at 28°C on wheat flour supplemented with 7.5% casein. To maintain relatively homogeneous populations, eggs collected at 2- to 3-day intervals were transferred to fresh diets. Other insects used were from cultures in the Department of Entomologi- cal Sciences at Berkeley. Imaginal disks of Drosophila melanogaster Meigen were provided by J. W. Fristrom (Department of Genetics, University of California, Berke- ley).
121 0048.3575/80/020121-08$02.00/O Copyright 0 1980 by Academic Press. Inc. AU rights of reproduction in any form reserved.
122 COHEN AND CASIDA
insect Chirin Synthetase Assay Preparation of microsomes. Last-instar
larvae were homogenized (mortar and pes- tle) at 10% (w/v) in 100 mM, pH 7.4, K phosphate buffer containing 0.25 M sucrose and 1.15% KCl. The homogenate was fil- tered through two layers of gauze and cen- trifuged at 12,OOOg for 30 min at 2°C. The supernatant was filtered through glass wool and centrifuged at 105,OOOg for 60 min. The microsomal pellet was washed with and suspended in an appropriate buffer for CS assay.
Etzzyme assay. Unless otherwise specified, the reaction mixture consisted of 50 ~1 of microsomal suspension as the en- zyme source (-0.5 mg protein) plus 300 all of 25 mZt4, pH 7.2, Tris-HCl or 50 mM, pH 6.5, Na phosphate buffer containing 10 n-&J MgCl,, 17 mM GlcNAc, and 1 mM DTT (final concentrations). The substrate UDP- [3H]GlcNAc (0.5 &i, l,lOO,OOO dpm; 46 ng, 71 pmol) was added last in a 5+1 vol- ume. Following incubation at 22”C, cold 5% TCA (2.5 ml) was added and the mixture cooled on ice for at least 1 hr. The precipi- tate was collected on glass-fiber filters (2.4 cm, GF/C Whatman) and washed with cold 5% TCA (6 x 2.5 ml). The filters were placed in scintillation vials and washed twice with cold 5% TCA (5 ml). The filters were air-dried overnight and further dried in an oven for 30 min at 60°C. They were then treated with 0.5 ml Soluene-100 (Packard Instrument Co., Downers Grove, Ill.) for 4 hr, and then 10 ml of a toluene- based scintillation mixture was used for LSC. Protein was determined by the method of Lowry et al. (13).
Verification of chitin biosynthesis. These studies used the T. castaneum product. To determine the resistance to alkaline deg- radation, the residue from filtration and drying of the TCA-precipitate fraction was fortified with 50 mg chitin powder and sub- jected to a series of digestions with recov- ery of the 12,OOOg (lo-min) sediment in each case. The total “H or 14C was determined by combustion and LSC. The treatments and
sequence were: 20% NaOH, loo”C, 1 hr, which disintegrates the filters; water wash; 2% HCl, 25°C 1 hr; 20% NaOH, lOo”C, 1 hr; washes of water (2x) and acetone (2x).
For determination of chitinase-degrad- able product, the TCA-insoluble precipi- tate on the filter was washed with water (9~) and then treated with 50 r&f Na phosphate, pH 6.6, buffer (0.5 ml) contain- ing 2 mg chitinase (EC 3.2.1.14) (Cal- biochem-Behring) or with 0.5 ml chitinase-free buffer. The soluble radioac- tivity after 20 hr at 30°C was determined by LSC on a 0.2-ml aliquot using a toluene- methyl Cellosolve-based scintillation mix- ture. The insoluble portion after three water washes was digested in Soluene for LSC as above.
Thin-layer chromatography was used to differentiate intermediates from the poly- mer formed (14). The reaction products were compared at 0 and 30 min after intro- ducing the substrate by first boiling (2 mitt) or adding glacial acetic acid (20 ~1) and then spotting the mixture on silica gel plates for development with n-propanol-water (7:3) (14) or on ammonium acetate-treated MN- 300 cellulose plates for development with ethanol-ammonium acetate (0.5 M, pH 3.8) (5:2, v/v) (10). Radioautography and LSC served to quantitate the products at the origin (polymer assumed to be chitin) and at higher Rf values (the substrate and nonpolymeric metabolites).
To determine the nature of the polymeric material, the TCA precipitate as before but with 14C substrate was treated with 6 N HCI at 100°C for 24 hr and an aliquot was di- rectly spotted for tic (three developments with n-propanol-water, 7:3) (R, 0.79 for glucosamine) and product detection by radioautography.
A special type of experiment with incu- bations directly on the filter paper was used to examine the possibility that chitin label- ing did not involve a polymerase but instead an exchange reaction between UDP- GlcNAc and the terminal group of a chitin chain. The extent of chitin labeling was de-
Triholrrm CHITIN SYNTHETASE PROPERTIES 123
termined after 10 min of reaction. Similar reactions at 10 min were washed with buffer (9x), and while the enzyme was still on the filter paper new incubation medium was added for an additional 20-min incubation with UDP-[“H]GlcNAc in the usual manner or with unlabeled UDP-GlcNAc (2 mM); the former reaction verified the enzyme ac- tivity and the latter tested whether or not chitin labeling was decreased by exchange.
RESULTS
Nature and Properties of T. castaneam Chitin Synthetase Evidence for designating the product as
chitin. The product is predominantly an alkali-stable and insoluble polymer since more than 50% of the TCA-insoluble radioactivity is retained in insoluble form on treatment twice with hot 20% NaOH and multiple washing steps. The losses that occur may result from the presence of short polymer chains, degradation to oligomers which do not sediment on centrifugation, and the many washing steps involved. TIC experiments also indicate polymer forma- tion since a portion of the ‘“C material after incubation with UDP-[14C]Gl~NA~ was re- tained at the origin in tic solvent systems that move other known metabolites and the substrate free of the origin; a control ex- periment in which the reaction was stopped immediately after UDP-[‘“C]GlcNAc addi- tion gave no 14C at the origin. The product is hydrolyzed under two conditions nor- mally used in characterizing chitin. First, at least 70% is degraded by chitinase as re- vealed by comparisons with incubations without chitinase. Second, the only product from HCl hydrolysis chromatographs in the region of glucosamine.
Evidence for designating the enzyme as chitin synthetase. UDP-[“H]GlcNAc is readily utilized, but [‘“C]GlcNAc and [“‘Clglucosamine do not label the insoluble material. Incubation with UDP-GlcNAc after UDP-[“H]GlcNAc does not alter the [“Hlchitin yield, whereas further incubation with labeled substrate gives higher [3H]-
chitin yields as expected; thus, there is no exchange of terminal GlcNAc groups in preexisting chitin with radioactive substrate or monomer. As detailed later, chitin syn- thesis is increased by Mg”+ and GlcNAc as generally expected for CS. The system undergoes strong product inhibition, by 50% with 11 ~IV UDP on IO-min preincu- bation prior to substrate addition (12). Of great importance is the finding that this en- zyme is sensitive to inhibition by polyoxin D, i.e., 50% inhibition at 4 ~IV with IO-min preincubation (12).
Localization. This is a membrane-bound enzyme cosedimenting with the mitochon- dria (12,OOOg pellet, -70% of the total ac- tivity) and microsomes (105,OOOg pellet, -30% of the total activity); less than 1% of the activity appears in the 105,OOOg super- natant fraction (or in an initial 1OOg pellet). The specific activity of the CS is similar for the mitochondrial and microsomal fractions and varies from 700 to 2500 dpm [“H]chitin/min/mg protein with different preparations over a period of 1 year. Further studies with T. castaneum used only the microsomal fraction since it was more homogeneous. Electron micrographs of ultrathin sections of the microsomal pel- let revealed membranes, vesicles, and ribo- somes but no bacteria, viruses, or other foreign organisms.
The specific activity of pupal CS was about one-eighth of that for larval enzyme. With the larvae, the activity drops to -50% of normal on starvation for 3 to 5 days. The larval enzyme activity is present in the gut as noted in more detail with T. brevicornis as discussed later.
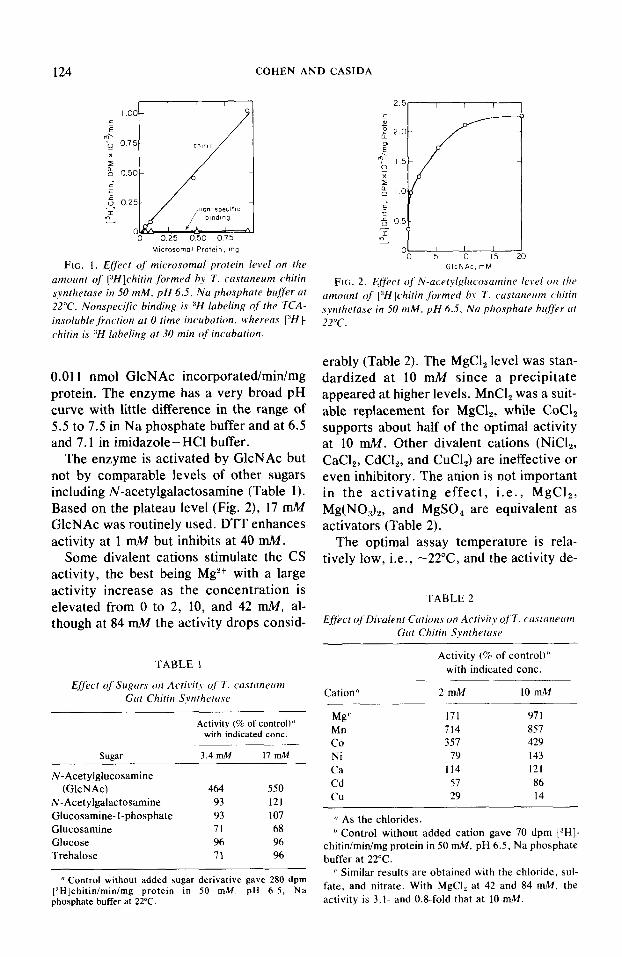
Optimizing assay conditions. Chitin labeling is proportional to enzyme concen- tration up to at least 1 mg microsomal pro- tein, with - 1% of the total labeling attribut- able to nonspecific binding (Fig. 1). The UDP-[3H]Gl~NA~ level normally used is 46 ng or 71 pmol, of which 2-4% is incorpo- rated into chitin within 30 min. The specific activity at higher substrate levels (200 to 2000 pLM) involving enzyme saturation was
124 COHEN AND CASIDA
25 I .oo
E g
m’ ; 2.0
so 075 ;
F n’
5 b 15
0 0.50 ;
E z 0 10
5 025
F g 05 0
0 0.25 0 50 0.75 Mtcrosomol Profan. mg :t;l
2
0 0 5 IO I5 20
FIG. 1. Effect of microsomal protein lervl on the GlcNAc. mM
amoant of [3Hi]chitin formed by T. custaneam chitin synthetase in 50 mM, pH 6.5. Na phosphate buffer at
FIG. 2. Ejfect of N-acetylglltco.samine le\,e/ on the
22°C. Nonspecijk binding is “H labeling of the TCA- amount of [:‘HJchitin formed by T. castaneam chitin
insoluble fraction at 0 time incubation, whereas rJHf synthetase in 50 mM. pH 6.5. Na phosphate buffer at 22°C.
chitin is “H labeling at 30 min of incubation.
0.011 nmol GlcNAc incorporated/min/mg protein. The enzyme has a very broad pH curve with little difference in the range of 5.5 to 7.5 in Na phosphate buffer and at 6.5 and 7.1 in imidazole-HCl buffer.
The enzyme is activated by GlcNAc but not by comparable levels of other sugars including N-acetylgalactosamine (Table 1). Based on the plateau level (Fig. 2), 17 mM GlcNAc was routinely used. DTT enhances activity at 1 mM but inhibits at 40 mM.
Some divalent cations stimulate the CS activity, the best being Mg2+ with a large activity increase as the concentration is elevated from 0 to 2, 10, and 42 n&I, al- though at 84 mM the activity drops consid-
TABLE 1
Effect of Sagars on Activity of 7‘. castaneam Gut Chitin Synthetase
(’ Control without added sugar derivative gave 280 dpm [“Hlchitinlminimg protein in 50 mM. pH 6.5, Na phosphate buffer at 22°C.
erably (Table 2). The MgCl, level was stan- dardized at 10 mkf since a precipitate appeared at higher levels. MnCl, was a suit- able replacement for MgCl,, while CoCl, supports about half of the optimal activity at 10 rnk!. Other divalent cations (NiCl,, CaCl,, Cd& and CuCl,) are ineffective or even inhibitory. The anion is not important in the activating effect, i.e., MgC12, Mg(NO,),, and MgSO, are equivalent as activators (Table 2).
The optimal assay temperature is rela- tively low, i.e., -22”C, and the activity de-
TABLE 2
Ejjtct oj’Divaient Cations on Actitaity of T. castaneum Gut Chitin Synthetase
Activity (% of control)” with indicated cont.
Cation” 2mM 10 m.M
Mg’ 171 971
Mn 714 857
CO 357 429
Ni 79 143
Ca 114 121 Cd 57 86
CU 29 14
I’ As the chlorides. b Control without added cation gave 70 dpm [JHl-
chitinlminimg protein in 50 mM. pH 6.5, Na phosphate buffer at 22°C.
c Similar results are obtained with the chloride, sul- fate, and nitrate. With MgCl, at 42 and 84 mM. the activity is 3.1. and 0.8-fold that at 10 mM.
Tribolum CHITIN SYNTHETASE PROPERTIES 125
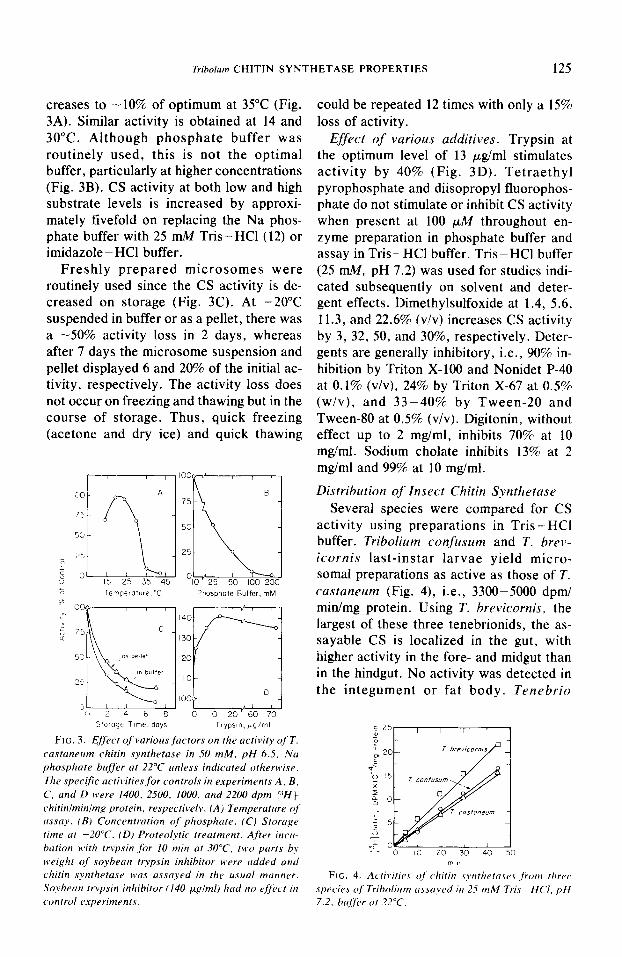
creases to -10% of optimum at 35°C (Fig. 3A). Similar activity is obtained at 14 and 30°C. Although phosphate buffer was routinely used, this is not the optimal buffer, particularly at higher concentrations (Fig. 3B). CS activity at both low and high substrate levels is increased by approxi- mately fivefold on replacing the Na phos- phate buffer with 25 n&I Tris-HCl (12) or imidazole - HCl buffer.
Freshly prepared microsomes were routinely used since the CS activity is de- creased on storage (Fig. 3C). At -20°C suspended in buffer or as a pellet, there was a -50% activity loss in 2 days, whereas after 7 days the microsome suspension and pellet displayed 6 and 20% of the initial ac- tivity, respectively. The activity loss does not occur on freezing and thawing but in the course of storage. Thus, quick freezing (acetone and dry ice) and quick thawing
lo Temperature.“C Phosphate Buffer, mM 0
“I IOOQ , , , , , “4 , , ,
Storage itme. days Trypsln,pq/ml
FIG. 3. Effect of varioas factors on the acfivify of T. castaneam chitin synthetase in 50 mM, pH 6.5, Na phosphate buffer at 22°C anlrss indicated otherwise. The specific actiL.ities for controls in experiments A. B. C, and D were 1400. 2500. 1000. and 2200 dpm [YHf chitinlminimg protein, respectively. (A) Temperature of assay. (BI Concentration of phosphate. (C) Szorage time at -20°C. (D) Proteolytic treatment. After incw bation with trypsin for 10 min at 30°C. tbcv parts b> weight of soybean trypsin inhibitor H’ere added and chitin s>~ntheta.se was assayed in the asaal manner. Soybean trypsin inhibitor (140 pglml) had no c~fcvt in control cj.xperiments.
could be repeated 12 times with only a 15% loss of activity.
Effect of various additives. Trypsin at the optimum level of 13 pg/ml stimulates activity by 40% (Fig. 3D). Tetraethyl pyrophosphate and diisopropyl fluorophos- phate do not stimulate or inhibit CS activity when present at 100 pM throughout en- zyme preparation in phosphate buffer and assay in Tris - HCl buffer. Tris - HCl buffer (25 mM, pH 7.2) was used for studies indi- cated subsequently on solvent and deter- gent effects. Dimethylsulfoxide at 1.4, 5.6, 11.3, and 22.6% (v/v) increases CS activity by 3, 32, 50, and 30%, respectively. Deter- gents are generally inhibitory, i.e., 90% in- hibition by Triton X-100 and Nonidet P-40 at 0.1% (v/v), 24% by Triton X-67 at 0.5% (w/v), and 33-40% by Tween-20 and Tween-80 at 0.5% (v/v). Digitonin, without effect up to 2 mg/ml, inhibits 70% at 10 mgiml. Sodium cholate inhibits 13% at 2 mg/ml and 99% at 10 mg/ml.
Distribution of Insect Chitin Synthetase Several species were compared for CS
activity using preparations in Tris - HCl buffer. Tribolium confczsum and T. brev- icornis last-instar larvae yield micro- somal preparations as active as those of T. castaneum (Fig. 4), i.e., 3300-5000 dpm/ min/mg protein. Using T. brevicornis, the largest of these three tenebrionids, the as- sayable CS is localized in the gut, with higher activity in the fore- and midgut than in the hindgut. No activity was detected in the integument or fat body. Tenebrio
d 25-,
FIG. 4. Ac,til,itie.$ oj chitin syrrthetase.r jkm three spec,ie> of Triboliam assayed itr 25 mM Tris -HC’I. pH 7.2. bujirer at 22°C.
126 COHEN AND CASIDA
molitor L. gives significant CS activity, mostly residing in the gut, with a specific activity of 6300 dpm/min/mg protein for the gut homogenate and its 900g and 12,000g sediment fractions; the 12,000g supernatant was inactive.
A survey of insects other than the tene- brionids revealed only one, Gulleria melonella L., that gave active CS prepara- tions (3800 dpmlminlmg protein on assaying guts of last-instar larvae). Species or organs giving inactive homogenates (~75 dpm/ min/mg protein) were Myzlrs persicue (Sulzer) (whole homogenates of nymphs and adults), Oncopeltus fusciatus (Dallas) (whole homogenates of embryos, last-instar nymphs, and newly emerged adults), Tele- ogryllus commodus Walker (guts from eighth-instar female nymphs), Byrsotricz fumigatu (Guerin) (guts from females), Spodopteru exigua (Hubner) (guts from last instar larvae), and D. melanoguster (imagi- nal disks treated with P-ecdysone to acti- vate chitin synthesis?.
DISCUSSION
An active CS system was prepared from Tribolium larvae. Its activity is localized in the fore- and midgut and is reduced in starved larvae and pupae. It is therefore likely that the polymerizing enzyme is as- sociated with the formation of the peri- trophic membrane, a chitoprotein (1, 15). It is important to establish that this CS is of insect origin and not due to a fungal con- taminant, since CS occurs in many fungi. In this respect, it is an advantage to work with Tribohm since their alimentary canal is free of microbial flora (16) and they are maintained on a very dry diet, minimizing microbial contamination. Whereas Tribolium are normally reared on a diet of brewer’s yeast and flour, in the present ex- periments casein was used in place of yeast. Several types of evidence indicate that only insect CS is assayed in the current investi- gation. The CS activity was the same for Tribolium larvae raised on diet treated with
’ J. W. Fristrom, personal communication.
fungistats (sorbic acid and methyl p- hydroxybenzoate) as on normal diet. Elec- tron microscopy studies did not reveal any microbial contamination of the 105,OOOg pellet. Finally, the CS activity of Tribolium is demonstrable without the elaborate pro- cess of enzyme treatments necessary to detect CS activity in yeast and lilamentous fungi.
Chitin synthetase from T. custuneum is similar in many respects to that from fungi, although the fungal enzymes are much more active (17-20). They both require GlcNAc and MgZ+. GlcNAc is the only ac- tive sugar with the Tribolium enzyme, whereas with some fungal enzymes other sugars are also effective (17). It is proposed that with fungal CS, GlcNAc stimulates ac- tivity by associating with an allosteric site (17, 18), a speculation also applicable to the insect enzyme. In both T. custaneum and fungal systems, Mg2+ can be replaced by Mn”+ and Co”+ (18, 19). Both the insect and the fungal enzymes are strongly inhibited by UDP and polyoxin D, the product of CS action and an antibiotic similar in structure to UDP-GlcNAc, respectively (12, 17, 18, 20). There are additional similarities be- tween the insect and fungal enzymes in re- sponse to other inhibitors (12) and higher activity in Tris than in phosphate buffer (19, 21). Fungal enzyme is released only on treatment with trypsin or other proteases (18,20), while an exogenous protease is not essential for activation of the gut enzyme. Inhibition of organophosphate-sensitive proteases during CS preparation and assay does not alter the activity of T. custunrum CS, providing some evidence for the ab- sence of an endogenous protease activator. Trypsin, in stimulating T. castuneum CS activity up to 40%, may expose blocked or masked active sites of the enzyme. An ex- ogenous primer is not necessary for either T. custuneum or fungal (14, 20) CS activity. Thiols are useful in preparing fungal CS (22), but with T. custanesrm enzyme DTT is of only moderate effect, although it pre- vents inhibition by -SH reagents (12). Fun-
Tribohm CHITIN SYNTHETASE PROPERTIES 127
gal CS can be solubilized with digitonin (20), using levels inhibitory to the insect CS. Many other surfactants and emulsifiers and dimethylsulfoxide inhibit or stimulate T. castaneum CS activity, indicating that they alter protein and membrane confor- mations.
Chitin synthetase activity is demon- strable not only in three Tribolium species but also in the guts of T. molitor and G. mellonella. Attention was focused on op- timizing the system for T. castaneum CS assays. Accordingly, the conditions are not necessarily optimal for other species. A previously reported CS system from whole Spodoptera eridania (Cramer) larvae (5, 6) utilized completely different methodology than the present experiments so it is not possible to compare the findings directly. The Tribolium gut enzyme is relatively sta- ble and easy to assay. This is not the case for integumental CS (8). The gut enzyme is probably involved in continuous secretion of the peritrophic membrane. Integumental CS is likely to be fundamentally similar to its counterpart in the gut, except for hor- monal regulation and functional organiza- tion of the enzyme complex associated with polymerization and fibrillogenesis. The authors have made some progress in ob- taining CS activity in cell-free systems from Trichoplusia ni (Htibner) integument, i.e., the system is sensitive to inhibition by polyoxin D and the product is resistant to alkaline degradation. This preliminary finding suggests that further optimization may yield an integumental CS as well as a gut CS from insects, and that they will probably be similar in their basic prop- erties.
ACKNOWLEDGMENTS
We thank A. Sokoloff (California State College, San Bernardino, Calif. for supplying T. hrfipicornis and T. c~mfirsfrrn and C. E. Dyte (Pest Infestation Labora- tory. Slough, Buckinghamshire, England) for providing T. ~~I.S/(IIICIII~I.
REFERENCES
1. K. M. Rudall and W. Kenchington, The chitin system. Biol. Rev. 49, 597 (1973).
2. A. C. Neville, “Biology of the Arthropod Cuti- cle,” Springer-Verlag. New York, 1975.
3. H. R. Hepburn (Ed.), “The Insect Integument.” Elsevier, New York, 1976.
4. R. A. A. Muzzarelli. “Chitin.” Pergamon Press. Oxford, 1977.
5. E. Jaworski. L. Wang, and Cl. Marco, Synthesis of chitin in cell-free extracts of Prodenitr eridrrnia. Nature (LondonJ 198, 790 (1963).
6. C. A. Porter and E. G. Jaworski, Biosynthesis of chitin during various stages in the metamorpho- sis of Prodeniu c,ridania. .I. Insect Pizysiol. 11,
1151 (1965). 7. B. Surholt, Studies in l,ivo and in vitro on chitin
synthesis during the larval-adult moulting cycle of the migratory locust, Locusta migratoricl L.,
J. camp. Physiol. 102, 135 (19751. 8. A. Vardanis, An in vitro assay system for chitin
synthesis in insect tissue, Lifr Sci. 19, 1949 (19761.
9. A. Verloop and C. D. Ferrell, Benzoylphenyl ureas-a new group of larvicides interfering with chitin deposition, in “Pesticide Chemistry in the 20th Century” (J. R. Plimmer, Ed.). ACS Symposium Series 37, pp. 237-270. 1977.
10. N. P. Hajjar and J. E. Casida, Structure-activity relationships of benzoylphenyl ureas as toxi- cants and chitin synthesis inhibitors in On- copeltrrs ,firsc.itrtn.\, Prstk. Biochem. Pirysiol. 11, 33 (1979).
II. W. H. van Eck. Mode of action of two benzoyl- phenyl ureas as inhibitors of chitin synthesis in insects. Insecr Biochem. 9, 295 (1979).
12. E. Cohen and J. E. Casida. Inhibition of Triholicrm gut chitin synthetase, Pestic. Biochrm. Physiol.. 13, 129 (1980).
13. 0. H. Lowry, N. J. Rosebrough, A. L. Farr, and R. J. Randall, Protein measurement with the Folin phenol reagent, J. Bid/. Chem. 193, 265 (1951).
14. G. W. Gooday and A. de Rousset-Hall, Properties of chitin synthetase from Coprinus cinrrrus. J. Gen. Microhiol. 89, 137 (1975).
15. W. Peters, Investigations on the peritrophic mem- branes of Diptera, in “The Insect Integument” (H. R. Hepburn. Ed.), pp. 5155543. Elsevier. New York, 1976.
16. A. Sokoloff, “The Biology of Triholium,” Vol. 2.
p. 258, Oxford University Press, Oxford, 1974. 17. A. de Rousset-Hall and G. W. Gooday, A kinetic
study of a solubilized chitin synthetase prepa- ration from Coprinus cinrwrrs, J. Gen.. Mic- robial. 89, 146 (1975).
18. J. Ruiz-Herrera, E. Lopez-Romero. and S. Bartnicki-Garcia, Properties of chitin syn- thetase in isolated chitosomes from yeast cells of Mlrcor rorrsii. J. Biol. Chrm. 252, 3338 (1977).
19. J. F. Peberdy and P. M. Moore. Chitin synthetase
128 COHEN AND CASIDA
in Mortiurd/u vinrrcrut Properties. cellular lo- 21. Y. N. Jan, Properties and cellular localization of calization and synthesis in growing cultures, J. chitin synthetase in Phycomyrs h/akes/rran~~.s. Gen. Microhid. 90, 228 ( 1975). J. Bid. Chem. 249, 1973 (1974).
20. A. Duran and E. Cabib, Solubilization and partial 22. R. Schekman and V. Brawley, Localized deposi- purification of yeast chitin synthetase. Confir- tion of chitin on the yeast cell surface in re- mation of the zymogenic nature of the enzyme, sponse to mating pheromone. Proc. Nat. Acad. J. Bid. Chem. 253, 4419 (1978). Sci. f USA) 76, 645 (1979).