Regulation of expression of 1,25D3-MARRS/ERp57/PDIA3 inrat IEC-6 cells by TGF� and 1,25(OH)2D3
Benjamin Rohea, Susan E. Saffordb, Ilka Nemerec, Mary C. Farach-Carsona,∗
a Department of Biological Sciences, University of Delaware, Newark, DE 19716, United Statesb Department of Biology, Lincoln University, Lincoln University, PA, United Statesc Department of Nutrition and Food Sciences, Utah State University, Logan, UT, United States
a r t i c l e i n f o
Article history:
Published on line 26 December 2006
Keywords:
Vitamin D3
Intestine
Calcium absorption
1,25D3-MARRS
Rapid response
a b s t r a c t
We examined the transcriptional regulation of expression of the redox-sensitive Membrane-
Associated-Rapid Response, Steroid-binding (1,25D3-MARRS) protein specific for 1,25(OH)2D3
in a rat small intestinal cell line, IEC-6, that demonstrates rapid responses to 1,25(OH)2D3.
1,25D3-MARRS binds and is activated by 1,25(OH)2D3, but is not itself up-regulated by treat-
ment with 1,25(OH)2D3, nor is there a Vitamin D response element (VDRE) in its proximal
promoter. We previously reported that transforming growth factor � (TGF�) increased steady
state levels of 1,25D3-MARRS transcript and protein approximately two-fold [Rohe B, Safford
SE, Nemere I, Farach-Carson, MC. Identification and characterization of 1,25D3-membrane-
associated rapid response, steroid (1,25D3-MARRS)-binding protein in rat IEC-6 cells. Steroids
2005;70:458–63]. To determine if this up-regulation could be attributed to the function of a
highly conserved consensus smad 3 binding element present in the proximal promoter of
the 1,25D3-MARRS gene, we created a promoter-reporter [SEAP] construct that was respon-
sive to TGF� (200 pM). Deletion or mutation of the smad 3 element greatly reduced the
response of the 1,25D3-MARRS promoter to TGF�. Subsequent studies found that the smad
3 response element is bound by a protein found in the IEC-6 nuclear extract, most likely
smad 3. Interestingly, although 1,25(OH)2D3 alone did not increase expression of the 1,25D3-
MARRS promoter-reporter, co-treatment of transfected IEC-6 cells with 1,25(OH)2D3 and
TGF� shifted the dose–response curve to a lower effective concentration (100 pM peptide).
We conclude that TGF� is a transcriptional regulator of 1,25D3-MARRS expression via a
functional smad 3 element and that cross-talk with non-classical 1,25(OH)2D3-stimulated
pathways occurs. The findings have broad implications for redox-sensitive signaling phe-
nomena including those that regulate phosphate transport in the intestine.
include protein disulfide isomerase (PDI) and ERp72 [2]. We
1. Introduction
The recently identified 1,25D -MARRS (membrane-associated-
3
rapid response, steroid-binding) protein, identical toERp57/PDIA3, is a key regulator of 1,25(OH)2D3-stimulatedintestinal phosphate absorption in chickens [1]. The protein
ith diets enriched in the antioxidants Vitamins C and E dou-led net phosphate absorption in vivo as compared to chicksed a control diet [3]. Finally, antioxidant diets increasedinding of [3H]1,25(OH)2D3 to 1,25D3-MARRS, leading us touggest that 1,25D3-stimulated phosphate transport in thentestine is sensitive to redox-sensitive transcription- and
embrane-initiated signaling pathways.Very little is known of the mechanisms that regulate
xpression of the 1,25D3-MARRS receptor itself, although itsevels are associated with the amount of phosphate thatan be transported in response to 1,25(OH)2D3 [1,4]. To thisnd, we analyzed the immediate proximal promoter [within000 bp] of the transcription start site of the 1,25D3-MARRSeceptor gene which lies at chromosome 3q35 in the rat.xamination of this region revealed a smad 3 consensus ele-ent [GTCTGGCCC] but no consensus VDRE. Consistent with
his, we found that treatment of IEC-6 cells withTGF�1, butot 1,25(OH)2D3, doubled steady-state transcript and protein
evels of the 1,25D3-MARRS receptor [5] Stimulation was dose-ependent with a maximum increase occurring at 200 pMGF�1.
Rapid actions of 1,25(OH)2D3 have been shown to alter generanscription via mechanisms independent of a VDRE, and
ay involve activation of Ca2+ signaling [6], a phenomenonecently named “excitation–transcription coupling” [7]. Inhese cases, the seco-steroid hormone also alters membraneon permeability, characteristically shifting the threshold ofctivation for voltage sensitive ion channels toward the restingembrane potential, a so called “left shift” [8,9]. Such action is
ostulated to cooperatively sensitize peptide hormone recep-ors to their ligands, typically reducing the concentration ofeptide hormone needed for cell activation. This type of mech-nism defines a form of receptor “cross-talk” that creates aunctional synergy between steroid and peptide hormones10].
In this study, we used a promoter-reporter construct tonalyze the response of the smad 3 element in the rat 1,25D3-ARRS promoter to TGF�1. We used a deletion/mutation
pproach to demonstrate the functionality of the proximal ele-ent, and surveyed cellular extracts for the presence of TGF�1
athway proteins and nuclear extracts for smad 3 binding pro-eins. Finally, we investigated the ability of 1,25(OH)2D3, whendded with TGF�1, to alter the dose response for increase ofteady state levels of 1,25D3-MARRS receptor expression.
. Experimental
.1. Cell culture
at IEC-6 cells were purchased from ATCC (Manassas, VA)nd maintained in DMEM with 5% (v/v) heat inactivated fetalovine serum (FBS) (Gibco Invitrogen Corporation Carlsbad,A) at 37 ◦C with 5% CO2. Cells were grown to 80% confluencynd passaged with trypsin/EDTA.
.2. Hormone treatment
ells were withdrawn from serum 24 h prior to treatment.ells then were treated with TGF�1 alone at the indicated con-
7 ) 144–150 145
centrations or co-treated with TGF�1 and 1 nm 1,25(OH)2D3 for18 h.
2.3. ELISA
IEC-6 cells were withdrawn from serum in the medium for48 h. After serum withdrawal, media was collected and ana-lyzed by ELISA for TGF�1 levels. All treatments were performedin serum free DMEM at the indicated concentrations.
2.4. Transfection
IEC-6 cells were allowed to attach and proliferate in six wellculture plates for 24 h in media containing 5% (v/v) FBS,then withdrawn from serum for 24 h. One half hour prior totransfection, the cells were placed in Opti-MEM media (GibcoInvitrogen Corporation) and incubated under normal incu-bation conditions. Cells then were transfected using 4 �L ofLipofectamine 2000 kit and protocol (Gibco Invitrogen), anda total amount of 3 �g of DNA (1.5 �g of pEGFP-N3 for trans-fection efficiency detection and 1.5 �g of empty, control orpromoter/reporter construct plasmid) per well.
2.5. RT-PCR
Cells were harvested at 80% confluency and total RNA wasisolated by use of RNAeasy Mini-prep® kits and a protocol sup-plied by the manufacturer (Qiagen Valencia, CA). Five hundredng of RNA were used in conjunction with OmniscriptTM kitand a protocol provided by the manufacturer (Qiagen). Poly-merase chain reaction (PCR) was performed with the cDNAtemplate using Hot Star TaqTM polymerase kit and protocol(Qiagen). Samples then were identified by electrophoresison 1.2% (w/v) agarose (Gibco Invitrogen) in TAE at 90 V for45–60 min. Quantitative (Q, or real time)-PCR was performedas above using the QuantitectTM SYBR® Green kit and protocol(Qiagen). Primers used in this study include rat smad 2: 5′-TCTAACAGAACTGCCGCCT-3′, 5′-GGAAGGTCTCTCCAACCC-TC-3′; rat smad 3: 5′-GCAGTGGCAATCACAGAGAA-3′, 5′-AA-CAGCCTGGGAGAGACTCA-3′; rat smad 4: 5′-GTTGCAGATA-GCTTCAGGGC-3′, 5′-GGATCCACGTATCCATCCAC-3′; rat smad7: 5′-GCACAAAGTGTTCCCTGGTT-3′, 5′-ACAACAAAACCGACC-GAAAG-3′; rat SARA: 5′-CACGGGTGTGAAAGGAGATT-3′,5′-AGTGCATCCCGGTAAAGTTG-3′; and Q-PCR primers,1,25D3-MARRS-RT: 5′-AAACTCAACTTTGCTGTAGCTAGCC-3′,5′-TGCATGACAAACTTCTCTCCTTTAGC-3′.
2.6. Promoter/reporter constructs
Genomic DNA from IEC-6 cells grown to 80% confluencewas extracted using the Genomic Tip® 20/G kit and pro-tocol (Qiagen) for cultured cells using proteinase K at a20 �g/�L final concentration as per kit instructions. Twoprimer sets were produced to generate a PCR productof 943 bp upstream of the 1,25D3-MARRS transcriptionalstart site and a 3′ truncated PCR product of 867 bp remov-
ing the smad 3 response element. These products thenwere cloned individually into the pSEAP2Basic vector fromBDBiosciences Clontech (San Jose, CA). The smad 3 mutantconstruct was produced by use of the Quikchange II XL
( 2 0
smad 2, smad 3, smad 4, smad 7 and smad anchor for receptoractivation (SARA) (Fig. 1), all of which are transducers of theintracellular signaling pathway stimulated by TGF�1. Controlslacking reverse transcriptase were blank (data not shown).
Fig. 1 – IEC-6 cells produce TGF�1 signal transductionpathway transcripts. RT-PCR analysis using specific primers
146 s t e r o i d s 7 2
mutagenesis kit and protocol from Stratagene (La Jolla,CA) with a mismatch primer set on the full length 1000 bppromoter in pSEAP2Basic. Primers used in this study includerMARRS promoter: 5′-AAGCTGAGCCTGTGCTTGAT-3′, 5′-CAGAGAACCGAGATGGGAGA-3′; rMARRSdel: 5′-AACCCCCTA-GGCCTGCTC-3′; and rMPSmut: 5′-GGTCTGCCCGGGCCTACTG-CGCGAGCGCAGG-3′, 5′-CTGCCAGCGCTCGCGCAGTAGGCCCG-GGCAG-3′.
2.7. Secreted alkaline phosphatase detection
Cell media samples (200 �L) were collected, centrifuged toremove cell debris and stored at −20 ◦C. Prior to analysis,samples were thawed on ice and secreted alkaline phos-phatase activity was measured using the BD Great EscAPeSEAP assay and protocol (BD Biosciences Clontech, San Jose,CA). Chemilumenescence was measured by using a plate lumi-nometer and data is expressed in relative luminescence units(RLU).
2.8. Nuclear extracts
IEC6 cells were cultured to 70% confluence, then the mediawas removed and the cells were washed with warm PBS. Thecells were lifted by addition of cold PBS/5 mM EDTA and incu-bated for 1 h at 4 ◦C. The cells were pelleted by centrifugationat 2000 rpm (350 × g) on a tabletop centrifuge for 5 min at 4 ◦C.The pellet was resuspended in PBS containing 5 mM EDTA andthen centrifuged again for 5 min at 2000 rpm at 4 ◦C. The pel-let was resuspended in 2 cell volumes of cold hypotonic buffer(10 mM HEPES pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA,0.75 mM spermidine, 0.15 mM spermine, 1 mM DTT, 10 �L/mLPICS II and 1 �L/mL PICS III). The cells were allowed to swellfor 30 min on ice. The solution was adjusted to 0.1% (v/v)NP40 to lyse the cells. The lysed cells were centrifuged at13,000 × g for 5 min. The supernatant then was dialysed in amini Slide-A-Lyzer® unit (Pierce, Rockford, IL) using a solutionof 20 mM, HEPES pH 7.9, 0.1 M KCl, 0.3 mM EDTA, 20% glycerol(v/v), 0.5 mM PMSF and 1 mM DTT for 4 h at 4 ◦C with constantstirring and a solution change at 2 h [11].
2.9. Generation of biotinylated DNA probe
Sense and antisense oligos of the region containing the smad3 response element were created and a working concentra-tion was produced by diluting the oligonucleotides to 1 �M inwater. The probes then were biotinylated using the Biotin 3′
End DNA Labeling kit and protocol (Pierce). The biotinylatedoligos were removed from the reaction by chloroform:isoamylalcohol extraction. Equal volumes of the sense and antisenseoligos were annealed by heating to 95 ◦C then cooling slowlyto room temperature.
2.10. Electromobility shift assays (EMSA)
The binding assay was carried out using the LightShift®
Chemiluminescent EMSA kit and protocol (Pierce). The reac-tion consisted of 2 �L of reaction buffer supplemented with1 �L 50% glycerol, 1 �L 100 mM MgCl2, 5 �L 1 �g/�L poly-dAdT,1 �L 1% NP40, and 0.5 �L of 50 nM biotinylated probe with or
0 7 ) 144–150
without 4 �L of 500 nM cold probe and IEC6 nuclear extract asnecessary. The reaction was incubated for 20 min at room tem-perature before running through a 4% (w/v) polyacrylamidenon-denaturing gel in 0.5× TBE for 3 h. The contents of thegel then were transferred to membrane using a mini-blot wettransfer apparatus in 0.5× TBE at an amperage of 0.35 for3 h in the cold with stirring. The membrane then was cross-linked with UV light, and exposed for approximately 30 s.The membrane was blocked, conjugated, washed and treatedwith peroxidase conjugate as described by the manufacturer’s(Pierce) protocol. Light sensitive film then was exposed to blotfor several hours and developed.
2.11. Statistical analysis
All values are expressed as means ±standard error. Exper-iments were repeated a minimum of two times. Statisticalvalues were generated using the Instat statistical programfrom GraphPad using one way analysis of variance (ANOVA)followed by either the Tukey-Kramer or Student T post-test asappropriate.
3. Results
3.1. IEC-6 cell responses to TGFˇ1
Consistent with our previous data [5], IEC-6 cells respondedto exogenous addition of TGF�1. We consistently found anincrease in the steady state levels of 1,25(OH)2D3-MARRSreceptor transcript and protein in cells treated with 200 pMTGF�1 compared to vehicle treated control cells. Using Q-PCR threshold as the assay, we found that treatment of IEC-6cells with 200 pM TGF�1 produced a two-fold increase in1,25(OH)2D3-MARRS receptor transcript versus untreated cells(not shown). To determine if IEC-6 cells contain the neces-sary TGF�1 pathway transduction machinery, RT-PCR analysiswas performed using cDNA derived from IEC-6 cell transcripts.Analysis revealed that IEC-6 cells contain the mRNA encoding
recognizing members of the machinery of the TGF�1signaling pathway reveals IEC-6 cells actively producetranscripts of the expected size. Negative controls wereblank (not shown).
s t e r o i d s 7 2 ( 2 0 0 7 ) 144–150 147
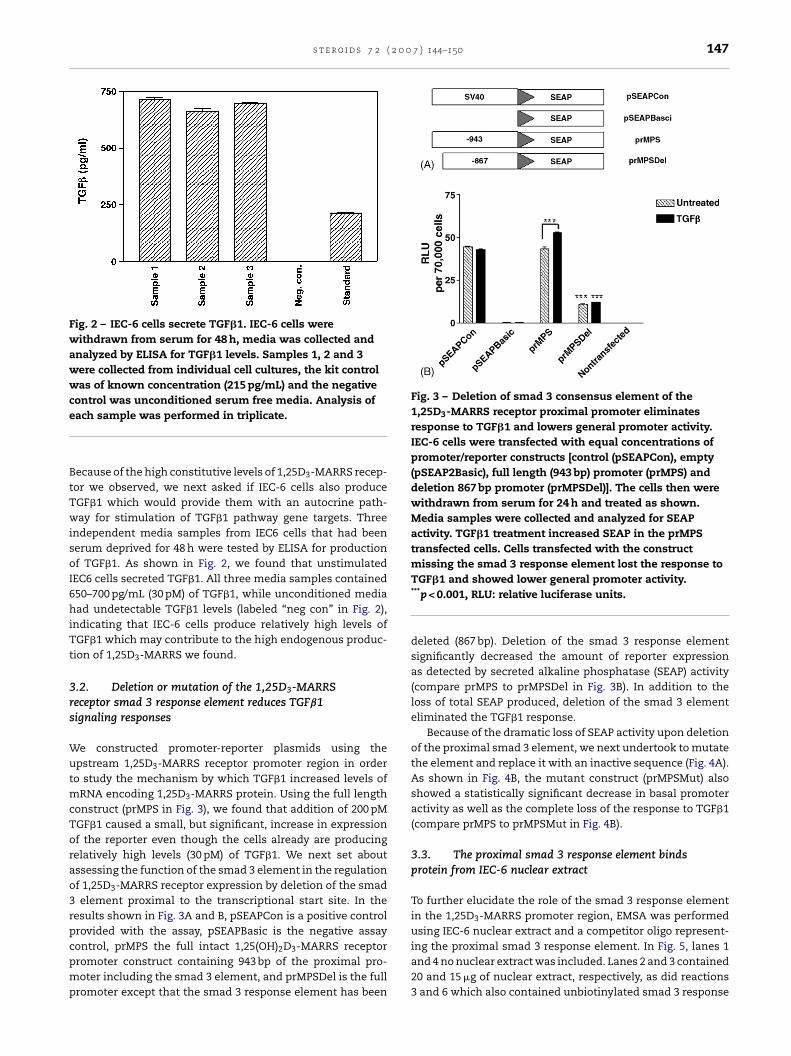
Fig. 2 – IEC-6 cells secrete TGF�1. IEC-6 cells werewithdrawn from serum for 48 h, media was collected andanalyzed by ELISA for TGF�1 levels. Samples 1, 2 and 3were collected from individual cell cultures, the kit controlwas of known concentration (215 pg/mL) and the negativecontrol was unconditioned serum free media. Analysis ofe
BtTwisoI6hiTt
3rs
WutmcTorao3rpcpmp
Fig. 3 – Deletion of smad 3 consensus element of the1,25D3-MARRS receptor proximal promoter eliminatesresponse to TGF�1 and lowers general promoter activity.IEC-6 cells were transfected with equal concentrations ofpromoter/reporter constructs [control (pSEAPCon), empty(pSEAP2Basic), full length (943 bp) promoter (prMPS) anddeletion 867 bp promoter (prMPSDel)]. The cells then werewithdrawn from serum for 24 h and treated as shown.Media samples were collected and analyzed for SEAPactivity. TGF�1 treatment increased SEAP in the prMPStransfected cells. Cells transfected with the constructmissing the smad 3 response element lost the response to
ach sample was performed in triplicate.
ecause of the high constitutive levels of 1,25D3-MARRS recep-or we observed, we next asked if IEC-6 cells also produceGF�1 which would provide them with an autocrine path-ay for stimulation of TGF�1 pathway gene targets. Three
ndependent media samples from IEC6 cells that had beenerum deprived for 48 h were tested by ELISA for productionf TGF�1. As shown in Fig. 2, we found that unstimulatedEC6 cells secreted TGF�1. All three media samples contained50–700 pg/mL (30 pM) of TGF�1, while unconditioned mediaad undetectable TGF�1 levels (labeled “neg con” in Fig. 2),
ndicating that IEC-6 cells produce relatively high levels ofGF�1 which may contribute to the high endogenous produc-
ion of 1,25D3-MARRS we found.
.2. Deletion or mutation of the 1,25D3-MARRSeceptor smad 3 response element reduces TGFˇ1ignaling responses
e constructed promoter-reporter plasmids using thepstream 1,25D3-MARRS receptor promoter region in ordero study the mechanism by which TGF�1 increased levels of
RNA encoding 1,25D3-MARRS protein. Using the full lengthonstruct (prMPS in Fig. 3), we found that addition of 200 pMGF�1 caused a small, but significant, increase in expressionf the reporter even though the cells already are producingelatively high levels (30 pM) of TGF�1. We next set aboutssessing the function of the smad 3 element in the regulationf 1,25D3-MARRS receptor expression by deletion of the smadelement proximal to the transcriptional start site. In the
esults shown in Fig. 3A and B, pSEAPCon is a positive controlrovided with the assay, pSEAPBasic is the negative assay
ontrol, prMPS the full intact 1,25(OH)2D3-MARRS receptorromoter construct containing 943 bp of the proximal pro-oter including the smad 3 element, and prMPSDel is the full
romoter except that the smad 3 response element has been
TGF�1 and showed lower general promoter activity.***p < 0.001, RLU: relative luciferase units.
deleted (867 bp). Deletion of the smad 3 response elementsignificantly decreased the amount of reporter expressionas detected by secreted alkaline phosphatase (SEAP) activity(compare prMPS to prMPSDel in Fig. 3B). In addition to theloss of total SEAP produced, deletion of the smad 3 elementeliminated the TGF�1 response.
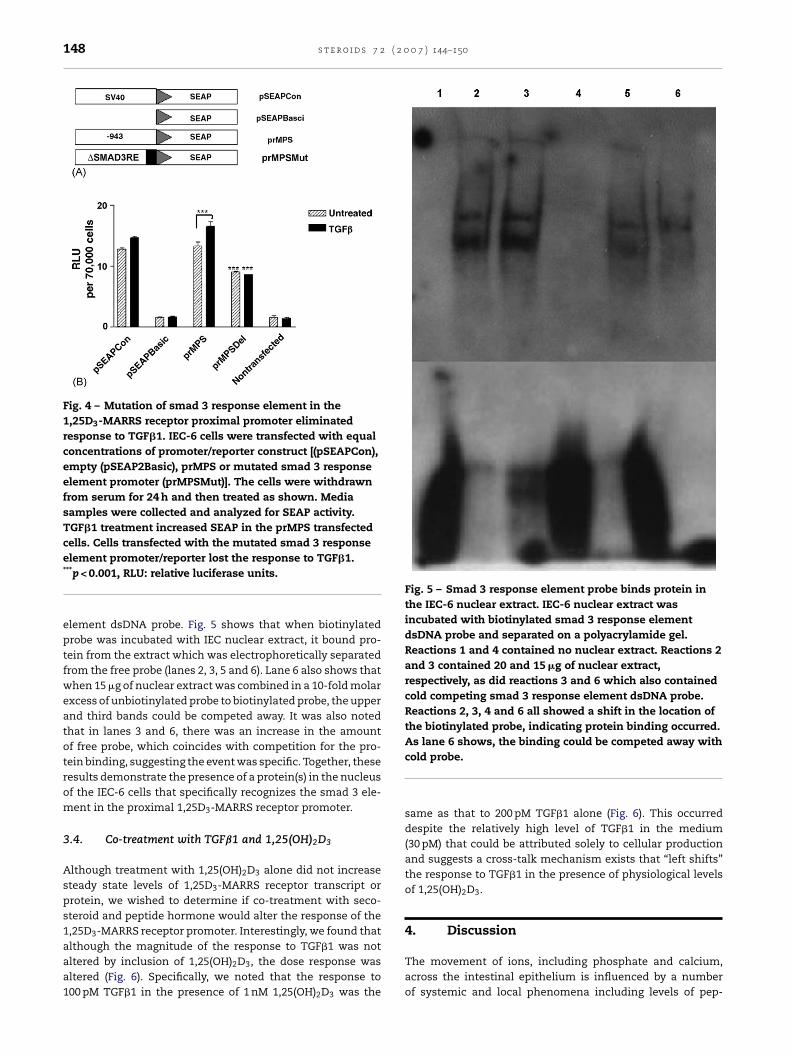
Because of the dramatic loss of SEAP activity upon deletionof the proximal smad 3 element, we next undertook to mutatethe element and replace it with an inactive sequence (Fig. 4A).As shown in Fig. 4B, the mutant construct (prMPSMut) alsoshowed a statistically significant decrease in basal promoteractivity as well as the complete loss of the response to TGF�1(compare prMPS to prMPSMut in Fig. 4B).
3.3. The proximal smad 3 response element bindsprotein from IEC-6 nuclear extract
To further elucidate the role of the smad 3 response elementin the 1,25D3-MARRS promoter region, EMSA was performedusing IEC-6 nuclear extract and a competitor oligo represent-
ing the proximal smad 3 response element. In Fig. 5, lanes 1and 4 no nuclear extract was included. Lanes 2 and 3 contained20 and 15 �g of nuclear extract, respectively, as did reactions3 and 6 which also contained unbiotinylated smad 3 response
148 s t e r o i d s 7 2 ( 2 0 0 7 ) 144–150
Fig. 4 – Mutation of smad 3 response element in the1,25D3-MARRS receptor proximal promoter eliminatedresponse to TGF�1. IEC-6 cells were transfected with equalconcentrations of promoter/reporter construct [(pSEAPCon),empty (pSEAP2Basic), prMPS or mutated smad 3 responseelement promoter (prMPSMut)]. The cells were withdrawnfrom serum for 24 h and then treated as shown. Mediasamples were collected and analyzed for SEAP activity.TGF�1 treatment increased SEAP in the prMPS transfectedcells. Cells transfected with the mutated smad 3 response
Fig. 5 – Smad 3 response element probe binds protein inthe IEC-6 nuclear extract. IEC-6 nuclear extract wasincubated with biotinylated smad 3 response elementdsDNA probe and separated on a polyacrylamide gel.Reactions 1 and 4 contained no nuclear extract. Reactions 2and 3 contained 20 and 15 �g of nuclear extract,respectively, as did reactions 3 and 6 which also containedcold competing smad 3 response element dsDNA probe.Reactions 2, 3, 4 and 6 all showed a shift in the location ofthe biotinylated probe, indicating protein binding occurred.As lane 6 shows, the binding could be competed away with
element promoter/reporter lost the response to TGF�1.***p < 0.001, RLU: relative luciferase units.
element dsDNA probe. Fig. 5 shows that when biotinylatedprobe was incubated with IEC nuclear extract, it bound pro-tein from the extract which was electrophoretically separatedfrom the free probe (lanes 2, 3, 5 and 6). Lane 6 also shows thatwhen 15 �g of nuclear extract was combined in a 10-fold molarexcess of unbiotinylated probe to biotinylated probe, the upperand third bands could be competed away. It was also notedthat in lanes 3 and 6, there was an increase in the amountof free probe, which coincides with competition for the pro-tein binding, suggesting the event was specific. Together, theseresults demonstrate the presence of a protein(s) in the nucleusof the IEC-6 cells that specifically recognizes the smad 3 ele-ment in the proximal 1,25D3-MARRS receptor promoter.
3.4. Co-treatment with TGFˇ1 and 1,25(OH)2D3
Although treatment with 1,25(OH)2D3 alone did not increasesteady state levels of 1,25D3-MARRS receptor transcript orprotein, we wished to determine if co-treatment with seco-steroid and peptide hormone would alter the response of the1,25D3-MARRS receptor promoter. Interestingly, we found that
although the magnitude of the response to TGF�1 was notaltered by inclusion of 1,25(OH)2D3, the dose response wasaltered (Fig. 6). Specifically, we noted that the response to100 pM TGF�1 in the presence of 1 nM 1,25(OH)2D3 was the
cold probe.
same as that to 200 pM TGF�1 alone (Fig. 6). This occurreddespite the relatively high level of TGF�1 in the medium(30 pM) that could be attributed solely to cellular productionand suggests a cross-talk mechanism exists that “left shifts”the response to TGF�1 in the presence of physiological levelsof 1,25(OH)2D3.
4. Discussion
The movement of ions, including phosphate and calcium,across the intestinal epithelium is influenced by a numberof systemic and local phenomena including levels of pep-
s t e r o i d s 7 2 ( 2 0 0
Fig. 6 – Co-treatment of IEC-6 cells with TGF�1 and1,25(OH)2D3 decreased the effective dose of TGF�1. IEC-6cells were serum withdrawn for 24 h then co-treated withTGF�1 and 1,25(OH)2D3 or TGF�1 alone for 18 h. Q-PCRshowed that the level of 1,25D3-MARRS receptor transcriptafter 200 pM TGF�1 treatment was similar to that after100 pM TGF�1 and 1 nM 1,25(OH)2D3 co-treatment. Barsl
tofirneauscemp
nmwsspiopAistIo
pac
abeled (−) are the negative controls.
ide and steroid hormones, diet, oxidative status, and degreef inflammation [3,12,13]. We believe that the newly identi-ed binding protein for 1,25(OH)2D3, namely 1,25D3-MARRSeceptor (ERp57/PDIA3) is a key player that functions as aew receptor for control of intestinal transport under a vari-ty of conditions. To determine how levels of the proteinre influenced by two hormones, TGF�1 and 1,25(OH)2D3, wendertook a study using a proximal promoter-reporter con-truct, specifically an examination of the function of a highlyonserved smad 3 element near the start of transcription. Thislement is present in three species that we examined (rat,ouse, and human) suggesting it might be functional under
hysiological conditions in a variety of species.In the first part of this study, we determined if the intesti-
al cell line that we chose for our studies, IEC-6, contained theachinery associated with TGF�1 responsiveness. Althoughe only examined the presence of transcripts encoding
everal smad proteins and a co-regulator (SARA) of TGF�1ignaling, we believe that the proteins also are likely to beresent in IEC-6 cells as they have been reported to be present
n a variety of intestinal epithelial cells [14,15]. The presencef this pathway may be part of the response pathway forrotection from inflammation, as has been reported [15,16].lternatively, it may function in growth or differentiation of
ntestinal epithelial cells as occurs in tissues undergoing con-tant regeneration and remodeling as do fast renewing cells ofhe colonic crypt [17]. In either case, this pathway is present inEC-6 cells and we propose that it contributes to the regulationf expression of the 1,25D3-MARRS receptor.
The 1,25D3-MARRS receptor transcript and protein areresent at high levels in IEC-6 cells [5], similar to those of �
ctin. We thus considered that there must be relatively highonstitutive levels of expression and wondered if smad 3 sig-
7 ) 144–150 149
naling contributed to this ongoing expression. We examinedthe levels of TGF�1 secreted into the medium of IEC-6 cells cul-tured in serum free media and found that cells made between650 and 700 pg/mL (30 nM) of TGF�1. Despite this, they main-tained the ability to respond to further addition of exogenousTGF�1, with a threshold concentration of 200 pM needed (Ref.[5] and this work). Deletion of the proximal smad 3 elementgreatly reduced basal reporter expression and completelyabolished the ability of added TGF�1 to increase expres-sion. Thus, this element clearly represents a critical regionof the 1,25D3-MARRS receptor promoter important in bothconstitutive and stimulated transcript expression. Becausedeletion of a nucleotide string can alter the spacing and three-dimensional presentation of the cis elements in a proximalpromoter, we next elected to perform a mutational transfor-mation that would leave spacing intact and merely eliminatethe consensus smad 3 element. When this was performed,the effects on basal reporter expression were less severe, butthe elimination of the TGF�1 responsiveness still was evi-dent. This clearly demonstrated that the smad 3 element inthe proximal 1,25D3-MARRS receptor promoter is functionaland is responsible for the increased transcription when theTGF�1 concentration is increased from 30 pM (endogenous) to230 pM (endogenous plus exogenously added). We next wishedto determine if the active cis element was bound by proteinspresent in IEC-6 nuclear extracts. Electromobility shift assaysusing nuclear extracts from IEC-6 cells clearly showed thepresence of protein which stably bound to the smad 3 element.The association of the protein with the smad 3 consensuselement was readily competed by a specific oligonucleotide,indicating that the association was specific. We had hopedto unequivocally demonstrate that the active binding proteinwas, in fact, smad 3, but a dearth of anti-smad 3 antibodiescapable of “supershifting” smad 3 containing complexes inEMSA assays limited this aspect of the study. We attempted touse a commercially available anti-smad 3 antibody, but wereconsistently unable to obtain supershifts of the complex evenwith the positive controls. This problem was not overcomedespite exhaustive troubleshooting with technical support.Interestingly, addition of the smad 3 antibody did reduce theamount of signal from the shifted third band in two separateexperiments, hinting that the presence of the antibody mightdisrupt formation of a smad 3-containing complex (data notshown). Thus, we believe that the active protein in the nuclearextract is most likely smad 3 based on the competition withthe specific competitor oligonucleotide and reduction of com-plex formation. Definitive identification will have to await theavailability of a smad 3 antibody capable of supershifting smad3 containing nuclear complexes. Despite this limitation, thepresence of a smad 3 element binding protein(s) in nuclearextracts from untreated cells suggests that IEC6 cells main-tain a state that represents a constitutive autocrine responseto TGF�1, but that can be up-regulated by exogenous additionof higher levels of TGF�1 (>200 pM).
Because of the likelihood of cross-talk between steroidand peptide hormone pathways, we decided to determine if
1,25(OH)2D3 could affect the dose response to TGF�1, per-haps shifting it to lower levels. Our laboratory previouslyreported a similar synergy between 1,25(OH)2D3 and parathy-roid hormone [18]. In the presence of physiological levels of
( 2 0
r
Birth Defects Res C Embryo Today 2005;75(1):58–71.
150 s t e r o i d s 7 2
1,25(OH)2D3 (1 nM), we found that the dose of TGF�1 neededto increase smad 3 element mediated up-regulation of thereporter was reduced by half, to 100 pM. This occurs despitethe fact that the 943 bp of the proximal promoter we used doesnot contain a consensus VDRE, thus we believe that this mostlikely involves a non-classical action of 1,25(OH)2D3, perhapsone linked to calcium signaling as reported previously [6].
In summary, based on the findings reported in this study,we conclude that TGF�1 is a transcriptional regulator of1,25D3-MARRS expression via a functional smad 3 element inthe proximal promoter and that cross-talk with non-classical1,25(OH)2D3-stimulated pathways occurs. The findings havebroad implications for redox-sensitive signaling phenomenonincluding those that regulate phosphate transport in the intes-tine, and for processes involving chronic inflammation of theintestinal epithelium, such as occur during colitis which arereported to involve smad 3 [14].
Acknowledgments
This work was supported in part by a grant from the NIDCRDE12641 (MCF-C), in part by USDA/CSREES 2004-35206-14134(IN) and in part by NIAMS AR48554 (SES).
e f e r e n c e s
[1] Nemere I, Farach-Carson MC, Rohe B, Sterling TM, NormanAW, Boyan BD, et al. Ribozyme knockdown functionally linksa 1,25(OH)2D3 membrane binding protein (1,25D3-MARRS)and phosphate uptake in intestinal cells. Proc Natl Acad Sci(USA) 2004;101(19):7392–7.
[2] Ellgaard L, Ruddock LW. The human protein disulphideisomerase family: substrate interactions and functionalproperties. EMBO Rep 2005;6(1):28–32.
[3] Nemere I, Wilson C, Jensen W, Steinbeck M, Rohe B,Farach-Carson MC. Mechanism of 24,25-dihydroxyvitaminD3-mediated inhibition of rapid, 1,25-dihydroxyvitaminD3-induced responses: role of reactive oxygen species. J CellBiochem 2006;99(6):1572–81.
[4] Sterling TM, Nemere I. 1,25-dihydroxyvitamin D3 stimulatesvesicular transport within 5 s in polarized intestinalepithelial cells. J Endocrinol 2005;185(1):81–91.
[5] Rohe B, Safford SE, Nemere I, Farach-Carson MC.Identification and characterization of
0 7 ) 144–150
1,25D3-membrane-associated rapid response, steroid(1,25D3-MARRS)-binding protein in rat IEC-6 cells. Steroids2005;70:458–63.
[6] Farach-Carson MC, Bergh JJ, Xu Y. Integrating rapidresponses to 1,25-dihydroxyvitamin D3 with transcriptionalchanges in osteoblasts: Ca2+ regulated pathways to thenucleus. Steroids 2004;69:543–7.
[7] Wamhoff BR, Bowles DK, Owens GK.Excitation-transcription coupling in arterial smooth muscle.Circ Res 2006;98(7):868–78.
[12] Laverty G, Elbrond VS, Arnason SS, Skadhauge E. Endocrineregulation of ion transport in the avian lower intestine. GenComp Endocrinol 2006;147(1):70–7.
[13] McCole DF, Rogler G, Varki N, Barrett KE. Epidermal growthfactor partially restores colonic ion transport responses inmouse models of chronic colitis. Gastroenterology2005;129(2):591–608.
[14] Tokumasa A, Katsuno T, Tanaga TS, Yokote K, Saito Y, SuzukiY. Reduction of Smad3 accelerates re-epithelialization in amurine model of colitis. Biochem Biophys Res Commun2004;317(2):377–83.
[15] Walia B, Wang L, Merlin D, Sitaraman SV. TGF�
down-regulates IL-6 signaling in intestinal epithelial cells:critical role of SMAD-2. FASEB J 2003;17(14):2130–2.
[16] Fuss IJ, Boirivant M, Lacy B, Strober W. The interrelated rolesof TGF�and IL-10 in the regulation of experimental colitis. JImmunol 2002;168(2):900–8.
[17] Hauck AL, Swanson KS, Kenis PJ, Leckband DE, Gaskins HR,Schook LB. Twists and turns in the development andmaintenance of the mammalian small intestine epithelium.
[18] Li W, Farach-Carson MC. Parathyroid hormone-stimulatedresorption in calvaria cultured in serum-free medium isenhanced by the calcium-mobilizing activity of1,25-dihydroxyvitamin D3. Bone 2001;29:231–5.