Role of Vanadium (V) in the Differentiation of C3H10t1/2 Cells Towards Osteoblast Lineage: A Comparative Analysis with Other Trace Elements Swati Srivastava & Narender Kumar & Rajani Salunke Thakur & Partha Roy Received: 10 October 2012 / Accepted: 3 January 2013 / Published online: 18 January 2013 # Springer Science+Business Media New York 2013 Abstract In recent time, vanadium compounds are being used as antidiabetic drug and in orthopedic implants. How- ever, the exact role of this incorporated vanadium in im- proving the quality of bone structure and morphology is not known. The impact of vanadium ion was studied and com- pared to other trace metal ions with respect to the prolifer- ation and osteoblast differentiation of C3H10t1/2 cells. Toxicity profile of these trace metal ions revealed a descend- ing toxicity trend of Fe 2+ > Zn 2+ > Cu 2+ > Co 2+ > Mn 2+ > V 5+ > Cr 2+ . The effect of vanadium and other trace metal ions on osteoblast differentiation was evaluated by culturing the cells for 10 days in osteoblastic medium supplemented with different trace ions at concentrations lower than their cytotoxic doses. The results indicated that vanadium has maximum impact on the induction of osteoblast differentia- tion by upregulating alkaline phosphatase activity and min- eralization by up to 145 and 150 %, respectively (p <0.05), over control. Cu 2+ and Zn 2+ had a mild inhibitory effect, while Mn 2+ , Fe 2+ , and Co 2+ demonstrated a clear decrease in osteoblast differentiation when compared to the control. The data as presented here demonstrate that orthopedic implants, if supplemented with trace metals like vanadium, may provide a source of better model for bone formation and its turnover. Keywords Trace element . Vanadium ion . Osteoblast . Differentiation . Toxicity Introduction All trace elements (TEs) belong to the category of micro- nutrients, which are needed by human body in very small quantities (generally less than 100 mg/day) for several bio- logical structures and functions [1]. Among all TE, vanadi- um is the most studied element because of its medicinal value. Various investigations have confirmed and extended the antidiabetic effect of vanadium compound both in vitro and in vivo [2] which include their ability to improve glucose homeostasis and insulin resistance in animal models of type 1 and type 2 diabetes mellitus. Therefore, based on these reports, vanadium compounds have been proposed to exert insulin-mimicking effects on biological systems [3]. Since insulin is already known to be a potent bone anabolic agent [4], using vanadium compound as osteoblast differen- tiating agent may open up another new area of research on bone science for development of bone implants. TE and their alloys have been used extensively for a wide range of applications as prosthetic materials for bone and tooth tissue reconstruction [5]. The suitable mechanical and physical properties of these materials have made them at- tractive as implant materials. However, in vivo corrosion of these metals is a major potential disadvantage of these implants. TE analysis has shown that metal ions can be localized in blood or in bone tissues adjacent to implants [6]; for example, vanadium drugs are reported to accumulate in bones in the form of vanadate and finally replacing phosphate in the apatite lattice of bone mineral [7]. There- fore, the functional activity of cells is clearly influenced by the physicochemical properties of the implants with which they are in contact. Inorganic salts, such as sodium vanadate (Na 3 VO 4 ), sodium metavanadate (NaVO 3 ) or vanadium pentaoxide (V 2 O 5 ), and organo-vanadium complexes, were S. Srivastava : N. Kumar : R. S. Thakur : P. Roy (*) Molecular Endocrinology Laboratory, Department of Biotechnology, Indian Institute of Technology Roorkee, Roorkee 247667 Uttarakhand, India e-mail: [email protected]Biol Trace Elem Res (2013) 152:135–142 DOI 10.1007/s12011-013-9602-2
Transcript
Role of Vanadium (V) in the Differentiationof C3H10t1/2 Cells Towards Osteoblast Lineage:A Comparative Analysis with Other Trace Elements
Swati Srivastava & Narender Kumar &
Rajani Salunke Thakur & Partha Roy
Received: 10 October 2012 /Accepted: 3 January 2013 /Published online: 18 January 2013# Springer Science+Business Media New York 2013
Abstract In recent time, vanadium compounds are beingused as antidiabetic drug and in orthopedic implants. How-ever, the exact role of this incorporated vanadium in im-proving the quality of bone structure and morphology is notknown. The impact of vanadium ion was studied and com-pared to other trace metal ions with respect to the prolifer-ation and osteoblast differentiation of C3H10t1/2 cells.Toxicity profile of these trace metal ions revealed a descend-ing toxicity trend of Fe2+ > Zn2+ > Cu2+ > Co2+ > Mn2+ >V5+ > Cr2+. The effect of vanadium and other trace metalions on osteoblast differentiation was evaluated by culturingthe cells for 10 days in osteoblastic medium supplementedwith different trace ions at concentrations lower than theircytotoxic doses. The results indicated that vanadium hasmaximum impact on the induction of osteoblast differentia-tion by upregulating alkaline phosphatase activity and min-eralization by up to 145 and 150 %, respectively (p<0.05),over control. Cu2+ and Zn2+ had a mild inhibitory effect,while Mn2+, Fe2+, and Co2+ demonstrated a clear decreasein osteoblast differentiation when compared to the control.The data as presented here demonstrate that orthopedicimplants, if supplemented with trace metals like vanadium,may provide a source of better model for bone formationand its turnover.
Keywords Trace element . Vanadium ion . Osteoblast .
Differentiation . Toxicity
Introduction
All trace elements (TEs) belong to the category of micro-nutrients, which are needed by human body in very smallquantities (generally less than 100 mg/day) for several bio-logical structures and functions [1]. Among all TE, vanadi-um is the most studied element because of its medicinalvalue. Various investigations have confirmed and extendedthe antidiabetic effect of vanadium compound both in vitroand in vivo [2] which include their ability to improveglucose homeostasis and insulin resistance in animal modelsof type 1 and type 2 diabetes mellitus. Therefore, based onthese reports, vanadium compounds have been proposed toexert insulin-mimicking effects on biological systems [3].Since insulin is already known to be a potent bone anabolicagent [4], using vanadium compound as osteoblast differen-tiating agent may open up another new area of research onbone science for development of bone implants.
TE and their alloys have been used extensively for a widerange of applications as prosthetic materials for bone andtooth tissue reconstruction [5]. The suitable mechanical andphysical properties of these materials have made them at-tractive as implant materials. However, in vivo corrosion ofthese metals is a major potential disadvantage of theseimplants. TE analysis has shown that metal ions can belocalized in blood or in bone tissues adjacent to implants[6]; for example, vanadium drugs are reported to accumulatein bones in the form of vanadate and finally replacingphosphate in the apatite lattice of bone mineral [7]. There-fore, the functional activity of cells is clearly influenced bythe physicochemical properties of the implants with whichthey are in contact. Inorganic salts, such as sodium vanadate(Na3VO4), sodium metavanadate (NaVO3) or vanadiumpentaoxide (V2O5), and organo-vanadium complexes, were
S. Srivastava :N. Kumar : R. S. Thakur : P. Roy (*)Molecular Endocrinology Laboratory, Departmentof Biotechnology, Indian Institute of TechnologyRoorkee, Roorkee 247667 Uttarakhand, Indiae-mail: [email protected]
Biol Trace Elem Res (2013) 152:135–142DOI 10.1007/s12011-013-9602-2
tested in most of these studies [8]. Out of these, V2O5 is themost commonly available compound as a source of vanadi-um, but till date, not much has been studied in vitro with thiscompound to understand its role in cellular differentiationprocesses. The aim of the present study was to determine ifthere is any direct effect of V2O5 (as a source of vanadiumion) on osteoblast differentiation and to compare its effectwith other TEs which are already known to regulate bonemetabolism and are being used in orthopedic implants.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM), fetal bovineserum (FBS), and streptomycin–penicillin antibiotic–anti-mycotic solutions were procured from Gibco (BRL,Inchinnan, UK). All trace metal ions used in this study weredivalent ions in nitrate solution except vanadium pentaox-ide, where vanadium is in oxidation state (V). These sol-utions were purchased from SRL (Mumbai, India) and wereof 99 % purity. All other chemicals used were of cell culturegrade from HiMedia (Mumbai, India) and Sigma-Aldrich(St. Louis, MO, USA).
Cell Culture
Mouse embryonic fibroblast cell line C3H10t1/2, a multipo-tent stem cell line of mouse origin, was purchased fromNational Center for Cell Sciences, Pune, India. The cells weremaintained in an undifferentiated state on tissue culture platein DMEM supplemented with 10 % FBS, 1× antibiotic–anti-mycotic solution, and 2 mM glutamine. During all the experi-ments, the cells were within passage numbers 23–25.
Cell Viability Test
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazoliumbromide (MTT) assay was carried out as described previously[9]. Briefly, C3H10t1/2 cells were seeded in 96-well plates atlow density (0.1×102 cells/cm2) and grown in its culturemedium along with the TEs at various concentrations asindicated. After 24 h of incubation, the medium was replacedwith fresh medium without any test chemicals. MTT (HiMe-dia, Mumbai, India) reagent was added to each well at a finalconcentration of 0.5 mg/ml, followed by incubation for 4 h toform formazan. The MTT-containing medium was then aspi-rated and 200 μL of DMSO (HiMedia, Mumbai, India) and25 μL of Sorensen's glycine buffer (0.1 M glycine and 0.1 MNaCl, pH10.5) were added to it to solubilize water-insolubleformazon. The absorbance was then determined in an ELISAplate reader (Oasys, Austria) at 570 nm.
Osteoblast Differentiation
For evaluating the impact of TEs on osteoblast differ-entiation, the cells were plated on six-well tissue cultureplates and cultured in osteoblast differentiation media(DMEM containing 10 % FBS, 50 μM ascorbic acid,10 nM dexamethasone, and 10 μM β-glycerophosphate)with different concentrations of trace ions. The cellswere then maintained in this differentiation media forfurther 10 days.
Alkaline Phosphatase Staining and Activity
For the alkaline phosphatase staining, the cells were fixed in4 % paraformaldehyde at room temperature (RT) for 2 minthen washed in phosphate-buffered saline (PBS) three timesfollowed by incubation with NBT/BCIP solution (Genie,Bangalore, India) for 30 min at RT. Finally, the cells wererinsed with water and observed under an inverted micro-scope (Axiovert 25, Carl Zeiss, Germany).
To analyze alkaline phosphatase (ALP) activity, the cellsin 24-well plates were rinsed three times with PBS and thensonicated in the presence of PBS followed by the estimationof protein using BCA reagent (Sigma, St. Louis, MO, USA).ALP activity was estimated based on the earlier method [10]with minor modifications. In brief, the assay mixturescontained 0.1 M 2-amino-2-methyl-1-propanol, 1 mMMgCl2, 8 mM p-nitrophenyl phosphate disodium, and cellhomogenates. After 4 min of incubation at 37 °C, thereaction was terminated with 0.1 N NaOH, and the absor-bance was measured at 405 nm. A standard curve wasprepared using p-nitrophenol as standard substrate. Eachvalue as obtained spectrophotometrically was normalizedto the protein concentration.
Alizarin Red Staining
For staining the cells, the media were drained off and thecell monolayers were washed with PBS. The cells were thenfixed with 4 % formaldehyde solution and incubated at 4 °Cfor 30 min. The cells were then washed thoroughly withdistilled water to remove excess formaldehyde and Alizarinred staining solution was added and incubated at roomtemperature for an additional 20 min, followed by removalof stain and washing with PBS. The cells were then ob-served under the inverted microscope (Axiovert 25, CarlZeiss, Germany).
von Kossa Staining
This staining was performed to observe the extent of phos-phate deposition in the cells after mineralization and it wasaccording to the method described earlier [11]. Briefly, the
136 Srivastava et al.
media were removed from the monolayer culture of cells.Then, about 1 ml of PBS was added to the culture forwashing of the cells. The cells were first fixed in 4 %formaldehyde, and then, a 5 % solution of silver nitratewas added to each well and the plate was kept in sunlightfor 30 min or until calcium turned black. Excess stain wasthen removed and washed with distilled water and finallyincubated in 5 % sodium thiosulfate for 2 min. Cells werethen washed and observed under microscope.
RT-PCR Analysis of Osteoblast Marker Genes
Total RNAwas extracted from the treated cells according tothe method described earlier [12]. RNAwas then quantifiedand an equal amount of the RNAs were transcribed with thehelp of the RT-PCR kit from Bangalore Genei (Bangalore,India) according to the manufacturer's instructions followedby PCR. The sequences of the primer used in this study andthe product sizes are listed in Table 1. PCR was performedby denaturing at 94 °C for 60 s and annealing for 30 s atvarious temperatures (Table 1) followed by extension at 72 °Cfor 60 s. The PCR products were then separated on 1.5 %agarose gel and visualized in a gel documentation system(Bio-Rad, USA).
Propidium Iodide Staining
For propidium iodide staining, the cells after 10 days ofdifferentiation were washed twice with PBS. They werethen incubated in propidium iodide solution (1 μg/ml)and RNase (10 μg/ml), for 30 min at RT in dark.Finally, after 30 min, the cells were washed with PBSand observed under fluorescence microscope (Axiovert 25,Carl Zeiss, Germany).
Statistical Analysis
Values are expressed as the mean±SD. The statistical sig-nificance was evaluated by one-way ANOVA at 5 % level ofsignificance. The statistical package used was Origin 6.1(OriginLab Corporation. USA).
Results and Discussion
Some of the bone-related disorders appear to be related totype1 diabetics since it has been reported that patients withtype1 diabetes have decreased bone mineral density [13]. Inthis regard, insulin has also been reported to cause enhancedbone development [4]. Due to the insulin mimetic action ofvanadium, various compounds of this element are also sup-posed to affect bone metabolism. Apart from its insulin-mimicking action, vanadium has wide role in the manufactureof hard tissue implants especially in bone tissue engineering.Hence, analyzing the role of vanadium in bone metabolismappears to be a critical issue since this may lead to bettermodification of implants by using this element. The main goalof this study was to analyze the role of vanadium (V5+) inosteoblast differentiation. This action of vanadiumwas furthercompared with other trace ions that are regularly been used inbone implants. In the present study, the viability of C3H10t1/2cells was first tested using varying doses of the trace elementsranging from 10 to 900 μM concentrations. The rationale forselecting this wide range of doses was to make it maximallyphysiologically relevant as possible since it is known that thedaily intake and accumulation of all these trace ions withinhuman body range from 10 μg to 1 mg (1). Effects of variousTEs on the cytotoxicity of cells were determined after 24 h ofincubations. The viability of cells increased to 137 and 142 %,respectively, (p<0.05) in the presence of Cr2+ and V5+ at aconcentration of 10 μM (Fig. 1). This increase in viability ofcells might be due to high metabolic activity or increasedproliferation of cells in presence of Cr2+ and V5+ which couldbe attributed to the growth hormone-mimicking nature ofvanadium and its compounds [14], since growth factors havelong been recognized as the proteins that can stimulate orinhibit cell proliferation [15]. On the other hand, the increasedproliferation in presence of Cr may be attributed to its antiox-idant property [16]. Fe2+, Co2+, and Mn2+ did not show anytoxic effect at lower concentrations. However, at 100 μM,although V5+ showed almost 95 % of cell viability, for Cu2+,Cr2+, Fe2+, Co2+, Mn2+, and Zn2+, the viability of cells re-duced to 77, 93, 78, 97, 92, and 86 %, respectively, indicatinga marginal toxicity at this dose. These data for vanadium were
Table 1 List of primers andproduct size (in base pair) usedfor RT-PCR
Role of Vanadium (V) in Differentiation of C3H10t1/2 Cells 137
in agreement with a recent report where it has been shown thatV5+ did not exert any toxic effect on L6 myotube cell line till200 μM concentrations [17]. Since after 200 μM concentra-tion treatment most of the TEs caused significant toxicity tocells, we chose three lower concentrations (25, 50, and
100 μM) for differentiation study. We did not test any con-centrations below that since some of the ions (V5+ and Cr2+)showed enhanced proliferative effect at those concentrationswhich might interfere in the induction of differentiation bythese ions.
Fig. 1 Effect of various TEs onthe proliferation and viability ofC3H10t1/2 cells after 24 h oftreatment. Vehicle-treated cellswere given a value of 100. Thedata represent mean±SD, n=3.Letters a, b, and c indicate sig-nificant levels of differences atp<0.05 with respect to “con-trol,” “a,” and “b,” respectively,within each ions
Fig. 2 Effects of various TEs onthe formation of early osteoblasticcells from undifferentiatedC3H10 t1/2 cells after 10 days ofdifferentiation. a Alkalinephosphatase assay ofdifferentiated cells. b Opticalimages of ALP-stained cells. Thedata are mean±SD, n=3. Lettersa and b indicate significant levelsof differences at p<0.05 with re-spect to “control” and “a,” re-spectively, within each ions. ALPactivity of control cells (vehicletreated) was considered as 100 %
138 Srivastava et al.
To evaluate the impact of TEs on the osteoblast differenti-ation of C3H10t1/2 cells, the ALP activity, which is an early-stage osteoblast marker, was examined after 10 days of differ-entiation period. Cellular ALP activity of control cells wasconsidered to be 100 %, which significantly increased in thepresence of V5+ at 25 μM (145±2.3 %) and 50 μM (110±1.9 %) concentrations, respectively (Fig. 2a; p<0.05). Asexpected, ALP activity was markedly inhibited at 100 μMconcentration in case of all the trace ions with the leastreduction however noticed in case of V5+-stimulated cells.The influence of TE treatment on cellular ALP activity in
the extracellular matrix was measured by ALP staining whichis depicted in Fig. 2b. As observed with the analysis of ALPactivity, ALP staining as well indicated a dose-dependentresponse in the presence of all ions. V5+-stimulated ALPactivity was more remarkable over other ions, when comparedto vehicle-treated cells. It has already been reported thatvanadium compounds have marked impact on alkaline phos-phatase activity [18].
After differentiation period, the deposited calcium phos-phate nodules, characteristics of osteoblastic mineralization,were detected by Alizarin red and von Kossa staining, where
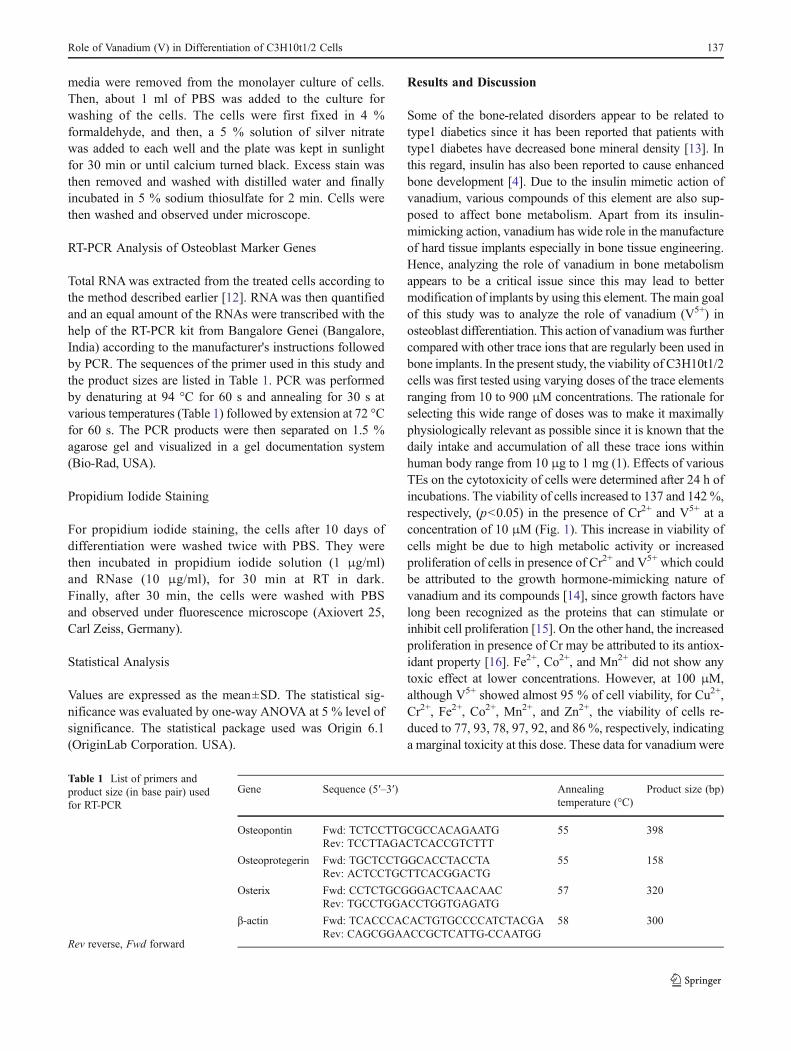
Fig. 3 Effects of various TEs onthe mineralization patterns ofC3H10t1/2 cells after 10 days ofdifferentiation. a Optical imagesof Alizarin red stained cells. bQuantification of Alizarin red instained cells. The data are mean±SD, n=3. Letters a, b, and cindicate significant levels ofdifferences at p<0.05 withrespect to “control,” 25 μMV5+,and 50 μMV5+-treated cells,respectively. Staining as shownby vehicle-treated cells (control)was considered as 100 %
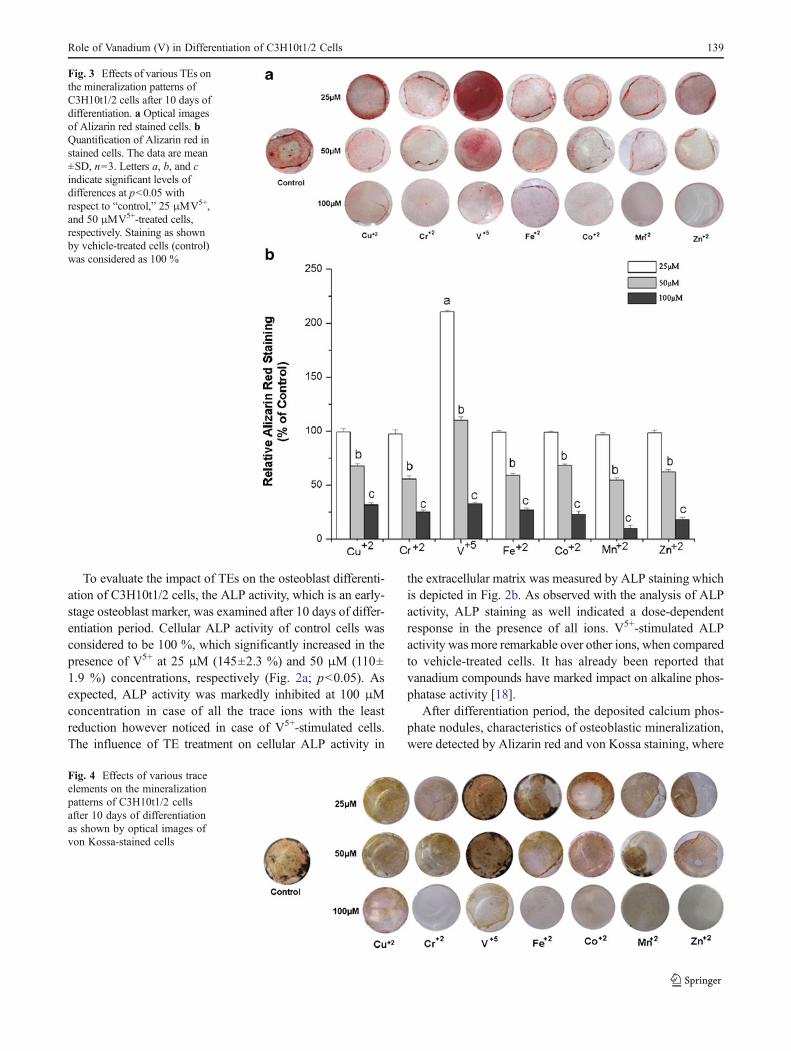
Fig. 4 Effects of various traceelements on the mineralizationpatterns of C3H10t1/2 cellsafter 10 days of differentiationas shown by optical images ofvon Kossa-stained cells
Role of Vanadium (V) in Differentiation of C3H10t1/2 Cells 139
Alizarin red and von Kossa combines with calcium andphosphate ions, respectively, thus indicating mineralizationof developing bones. As shown in Fig. 3a, b, V5+ showed asignificant increase in mineralization over control cells at25 μM (210 %), which however decreased almost to thelevel of control cells at 50 μM (p<0.05). As expected, at100 μM, all ions showed a remarkable reduction in themineralization. A similar pattern of inhibition of minerali-zation was shown by von Kossa staining in C3H10t1/2 cellsas depicted in Fig. 4. Facchini et al. [19] investigated theeffect of vanadium on bone metabolism in both diabetic andnondiabetic rats. They showed that vanadium compoundimproves diabetes-related bone dysfunction primarily byimproving the diabetic state and also appeared to increasebone formation in nondiabetic rats. In our study, we showedthat vanadium ion caused the direct and maximum upregu-lation of osteoblast differentiation, which was in accordancewith a recent report where it was shown that the deprivationof this ion induces skeletal deformation in the forelegs ofgoats [20]. Vanadium-induced enhanced mineralization mayalso be linked to its effect on collagen biosynthesis in cells.It has been reported that vanadium compounds upregulatethe synthesis of collagen in cells which play a major role inthe formation and function of bones [21]. Besides that,vanadium is also known to accumulate in the apatite latticeof bone tissue [22], which in turn results in enhancedmineralization of bones.
To further confirm the above findings, expressions of fewpotent osteoblast marker genes were checked and are shownin Fig. 5. At 25 μM concentration, V5+ exerted a significantupregulation in the expression of osteopontin gene as com-pared to control, while Cr2+, Fe2+, and Co2+ showed aremarkable decrease in its expression. A similar trend wasobserved while analyzing the expression patterns of anothermarker of osteogenesis, i.e., osterix. Except V5+ which
significantly upregulated its expression, other elementsexerted either nominal or negative impacts on the expressionof osterix. The other osteogenic gene as tested here, osteo-protegerin, was found to be upregulated only in the presenceof V5+ and was absent in the presence of all other ions. Insummary, V5+ positively stimulated the expression of all theosteogenic marker genes, as tested here, at least at 25 μMconcentration which was almost negligible in case of othermetal ions. Among all the TEs that we tested here, only V5+
salts are known to inhibit protein tyrosine phosphatases,which in turn enhance the activation of extracellular recep-tor kinase (ERK) within cells [23]. Since it is already knownthat active ERKs induce the expression of osteogenic genes,indirect upregulation of ERKs by V5+ may be responsiblefor the overexpression of these osteogenic genes like osteo-pontin, osterix, and osteoprotegerin.
Differences in the cellular responses to trace metal ions are acomplex process with several mechanisms operating simulta-neously. In the present study, the toxicity trend followed bytrace elements was Fe2+ > Zn2+ > Cu2+ > Co2+ >Mn2+ > V5+ >Cr2+, while the osteoblast differentiation potential of ions fol-lowed V5+ > Cu2+ > Fe2+ > Zn2+ > Co2+ > Mn2+ > Cr2+.Mahealani et al. [24] reported that permeability of ions in the
which is more permeable to cell membrane can affect cellmetabolism more effectively than non-permeable ions. Thishypothesis was further supported by a previous report wherehighly soluble Co complexes were found to readily cross thecell membranes via the anion transporter, whilst the cell mem-brane permeability was reported to be low for Cr complexeswhich were less soluble at physiological pH [25], and hence, Coshowed more toxicity than Cr [26]. As shown in the presentstudy, Zn is more permeable to cells than Co and Cr and, hence,causes more toxicity at higher concentration than Co and Cr and
Fig. 5 Representativetranscriptional analysis for theexpression of various osteogenicmarker genes in response to TEsas determined by RT-PCR
Fig. 6 Effect of various TEs onthe apoptosis of C3H10t1/2 cellsover the whole osteoblastdifferentiation process asdetermined by propidium iodidestaining following stimulation bydifferent test chemicals. Arrowsindicate apoptotic bodies withinthe nucleus (×20 objectives). Ccontrol
140 Srivastava et al.
cells decreases in the order of Zn2+ > Co2+ > Mn2+ > Cd2+ >Cr2+. Based on the above report, we hypothesized that the ion
also affects osteoblast differentiation more efficiently. However,further detailed analysis is needed to provide any conclusiveevidence for this complex crosstalk between bioavailability,toxicity, and metabolic effects of these ions on cells.
In order to check if long-term exposure of ions duringdifferentiation was responsible for apoptosis of cells, differ-entiated cells were stained with propidium iodide. By stain-ing these cells with propidium iodide, the apoptoticcondition of the cells could be analyzed where the healthycells can be identified by intact bright red nucleus, whereasthe apoptotic cells can be recognized by the fragmentedDNA in the nucleus. As shown in Fig. 6, the exposure ofcells for 10 days to all the TEs at 25 μM concentrationsrendered them healthy with no signs of apoptosis. While at50 μM concentration, long exposure of Cr2+ and Co2+
resulted in degraded DNA pattern with almost no effectsin response to other elements. However, at 100 μM concen-tration, all the ions induced a profound toxicity thus con-firming the earlier data where no responses were seen bythese elements at this concentration. Further, vanadium hasalso been reported to induce toxic effects in several tissuesat pharmacological doses [27], and this could be correlatedto our findings where V5+ was found to show decreasedosteoblast differentiation potency at higher concentrations.
In conclusion, the above findings together illustrate thatlow dose of vanadium could be an effective inducer in thedifferentiation of osteoblasts without causing any significanttoxicity to the cells even after prolonged exposure. To the bestof our knowledge, no other studies addressed this aspect ofvanadium-induced osteogenesis and its comparatively higherefficacy over other trace elements. Hence, the data as pre-sented here could provide some basic concepts about themodification of the choice of appropriate trace element addi-tives as well as their concentrations in orthopedic and dentalimplants so as to restore the normal function of impaired hardtissues with minimal adverse effects like toxicity. Further,based on these data, detailed analysis of the mechanism ofaction of vanadium both at in vitro and in vivo level iswarranted to elucidate their exact mode of actions.
Acknowledgments This work was funded by the Department of Bio-technology, Government of India (reference no. DBT-JRF/08-09/459),Ministry of Human Resource and Development (MHR03-412-Fig ‘B’),and Uttarakhand State Council for Science and Technology (reference no.UCS&T/R&D/LS-10/11-12/4224/1) as funded project to PR.
References
1. Fraga CG (2005) Relevance, essentiality and toxicity of traceelements in human health. Mol Aspects Med 26:235–244
2. Ming L, Jason JS, Wenjun D, Debbie CC (2009) Anti-diabeticeffects of sodium 4-amino-2,6-dipicolinatodioxovanadium (V)
dihydrate in streptozotocin-induced diabetic rats. J Inorg Biochem103:585–589
3. Thompson KH, Orvig C (2006) Vanadium in diabetes: 100 yearsfrom phase 0 to phase I. J Inorg Biochem 100:1925–1935
4. Thrailkill KM, Lumpkin CK Jr, Bunn RC, Kemp SF, Fowlkes JL(2005) Is insulin an anabolic agent in bone? Dissecting the diabeticbone for clues. Am J Physiol Endocrinol Metab 289:735–745
5. Navarro M, Michiardi A, Castano O, Planell JA (2008) Biomate-rials in orthopaedics. J R Soc Interface 5:1137–1158
6. Zhi LS, John CW, Hanks CT (1997) Effects of metal ions onosteoblast-like cell metabolism and differentiation. J Biomed Ma-ter Res 34:29–37
7. Fukui K, Fujisawa Y, Ohya Nishiguchi H, Kamada H, Sakurai H(1999) In vivo coordination structural changes of a potent insulin-mimetic agent, bis(picolinato)oxovanadium(IV), studied by elec-tron spin-echo envelope modulation spectroscopy. J Inorg Bio-chem 77:215–24
8. Srivastava AK, Mehdi MZ (2005) Insulino-mimetic and anti-diabetic effects of vanadium compounds. Diabet Med 22(1):2–13
9. Mosmann T (1983) Rapid colorimetric assay for cellular growthand survival: application to proliferation and cytotoxicity assays. JImmunol Methods 65:55–63
10. Bessey OA, Lowry OH, Brock MJ (1946) A method for the rapiddetermination of alkaline phosphatase with five cubic millilitres ofserum. J Biol Chem 164:321–329
11. Meloan SN, Puchtler H (1985) Chemical mechanisms of stainingmethods: von Kossa's technique. What von Kossa really wrote anda modified reaction for selective demonstration of inorganic phos-phate. J Histotechol 8:11–13
12. Chomczynski P, Sacchi N (1987) Single step method of RNAisolation by acid guanidinium thiocyanate–phenol–chloroform ex-traction. Anal Biochem 162:156–159
13. Muñoz-Torres M, Jódar E, Escobar-Jiménez F, López-Ibarra PJ,Luna JD (1996) Bone mineral density measured by dual X-rayabsorptiometry in Spanish patients with insulin-dependent diabetesmellitus. Calcif Tissue Int 58(5):316–319
14. Tsiani E, Fantus IG (1997) Vanadium compound: biologicalactions and potential as pharmacological agents. Trends Endocri-nol Metab 8(2):51–58
16. Hsing-Hsien C, Ming-Hoang L, Wen-Chi H, Chen-Ling H (2004)Antioxidant effects of chromium supplementation with type 2diabetes mellitus and euglycemic subjects. J Agric Food Chem52(5):1385–1389
17. Hwang SL, Chang HW (2012) Natural vanadium-containing Jejuground water stimulates glucose uptake through the activation ofAMP-activated protein kinase in L6 myotubes. Mol Cell Biochem360(1–2):401–9
18. Cortizo AM, Salice VC, Etcheverry SB (1994) Vanadium com-pounds: their action on alkaline phosphatase activity. Biol TraceElem Res 41:331–339
19. Facchini DM, Yuen VG, Battell ML, McNeill JH, Grynpas MD(2006) The effects of vanadium treatment on bone in diabetic andnon-diabetic rats. Bone 38(3):368–77
20. Nielsen FH, Uthus EC (1990) Vanadium in biological systems:physiology and biochemistry. Kluwer, London
21. Schwartz AV (2003) Diabetes mellitus: does it affect bone? CalcifTissue Int 73:515–519
22. Stankiewicz PJ, Tracey AS, Crans DC (1995) Inhibition ofphosphate-metabolizing enzyme by oxovanadium (V) complexes.Met Ions Biol Syst 31:287–324
23. Hila TK, Judith K, Tal S, Shira GA, Galit S, Hava GH, Yuval YE(2003) Protein tyrosine phosphatase ε inhibits signaling bymitogen-activated protein kinases. Mol Cancer Res 1:541–550
Role of Vanadium (V) in Differentiation of C3H10t1/2 Cells 141
24. Mahealani K, Monteilh Z, Meredith C, Monica JSN, Andrew MS,Reinhold P, Andrea F (2003) TRPM7 provides an ion channelmechanism for cellular entry of trace metal ions. J Gen Physiol121:49–60
25. Ramirez-Diaz MI, Diaz-Perez C, Vargas E, Riveros-Rosas H,Campos-Garcia J, Cervantes C (2008) Mechanisms of bacte-rial resistance to chromium compounds. Biometals 21:321–332
26. Cyrille F, Alain P, Fackson M, John A, David JZ, MaryamT OLH(2006) Effect of cobalt and chromium ions on human MG-63osteoblasts in vitro: morphology, cytotoxicity, and oxidative stress.Biomaterials 27:3351–3360
27. Preet ABL, Gupta MR, Siddiqui PK, Yadava NZ (2005) Restorationof ultrastructural and biochemical changes in alloxan induced diabet-ic rat sciatic nerve on treatment with Na3VO4 and Trigonella—apromising antidiabetic agent. Mol Cell Biochem 278:21–31











![Perles Vanadium [Compatibility Mode]](https://static.documents.pub/doc/80x56/5571fe6349795991699b4865/perles-vanadium-compatibility-mode.jpg)











