Light and Electron Microscopic Observations of the Life Cycle of Sarcocystis orientalis sp. n.
in the Rat (Rattus norvegicus) and the Malaysian Retieulated Python (Python reticulatus)*
V. Z a m a n and F rede r i ek C. Colley
Department of Parasitology, Faculty of Medicine, University of Singapore and Institute for l~edical Researeh, Kuala Lumpur, Malaysia and
the G. W. Hooper Foundation, University of California, San Franciseo
Received April 1, 1975
Summary. A light and electron microscopic study of Sarcocystis orientalis sp. n. was made. The life cycle of this parasite is in two hosts. Gametogony is in the intestinal epithelial cells of ~ predator, Python reticulatus. I~osl~ora-like oocysts developed. Sporocysts average 9.1 by 7.7 ~m. Rats (Rattus norvegicus) were infected with sporocysts and asexual stages devel- oped. Ten days after infection large zoites (average 7.85 by 2.48 ~m) were observed free in peripheral blood and within white blood cells. Small schizonts producing merozoites 2-3 ~m long were seen in lung tissue. Tissue cysts developed in skeletal musele and produced numerous cystozoites (average 5.53 by 1.38 tzm). Firm structure was similar to previously described Sarcocysti8 spp.
In t roduct ion
Rzepczyk (1974) recen t ly r epor ted the deve lopmen t of Sarcocystis-like infect ion in the muscles of l abo ra to ry b red Rattus norvegicus which h a d been fed oocysts of an Isospora species f rom the Aus t r a l i an ca rpe t py thon , Morelia spilotes variegata.
W e recovered s imilar Isospora-like oocysts f rom the Malays ian re t i cu la ted py~hon, Python reticulatus Schneider , 1801. Oocysts were fed to l a b o r a t o r y ra t s and Sarcocystis-like infectiorr of s t r i a t ed muscles and other t issue was observed.
The l ight and e lect ron microscopic morpho logy of the deve lopmen ta l s tages of this paras i te and other deta i l s of i ts life cycle are given in th is paper .
Materials and Methods The pythons used in this study were purchased ffom a Chinese butcher shop in Singapore,
specializing in snake and other exotic meat. :Nine Python reticulatus shedding oocysts were killed. The entire alimentary tract was removed and examined for developmental stages of the parasite. Impression smears were made lrom the stomach, duodenum, ileum, jejunum, caecum, striated and cardiac muscles, liver, lungs, kidney and spleen. Smears were fixed in absolute methanol and stained with Giemsa. Additional tissue from the alimentary tract was fixed in 10% formal-saline, embedded in paraffin and sectioned. Slides were stained with hematoxylin-eosin and }?AS.
* Supported in part by the University of California International Center for Medical Research (UC ICMR, Hooper Foundation, University of CaliIornia, San Francisco 94143) with research grant AZ 10051 to the Department of international Health, Sehool of Medicine, University of Califoraia, San :Francisco, from the :National Institute of Allergy and Infectious Diseases, NationM Institutes of Health, U.S. Public Health Service.
1 Z. Parasitenk.
170 V. Zaman and F. C. Colley
Intestinal epithelium was fixed in 5 % glutaraldehyde and processed for electron microscopy according to standard technique.
Thirty female, two months old white rats (Rattus norvegicus) (Wistar strain) and 20 white mice (Mus musculus) were infeeted per os with oocyst suspension via a polythene tube and syringe. The total number of oocysts administered to each rat was not determined but in each ease the dose was heavy, at least 50 sporoeysts per 40×40 field. From 0.1 to 0.5 ml of oocyst suspension was given to each rat and mouse.
Daily thin blood films from eaeh infeeted rat and mouse were fixed in absolute methanol and stained with Giemsa.
All but two rats died after infection. When possible rats were autopsied at the time of death and brain, striated and heart muscle, lungs, spleen, liver and kidneys were proeessed for light microscopy. The striated museles were from the oesophagus, diaphragm and the tongue.
In addition 20 uninfeeted two months old female rats (Wistar strain) and 20 female labora- tory bred white miee were killed and the striated musele of the oesophagus, diaphragm and the tongue were examined histologieally for the presence of Sarcocystis, so as to exelude the possibility of natural infeetion.
Results
Sarcocystis orientalis sp. n.
Light Microscope S tudy
1. Development in Python
No schizonts were observed in any organs of the python. Gametocytes and ooeyts were observed th roughout the small intestine but were most numerous in the duodenum.
Microgametocytes. Microgametoeßes were uneommon. They were usually seen within parasi tophorous vaeuoles and were only observed above the host nucleus. Mierogametocytes were round, f rom 6-8 ~ in diameter with approx- imately 18 nuelei (Fig. 1). Each free microgamete had aa elongate rmeleus, about 5 ~z long and 0.5 ~ wide with two flagellae, eaeh about 7 ~ long (Fig. 2).
Macrogametocytes. Maerogametocytes artd ooeysts were extremely numerous in the intestinal epithelium. Young maerogametoeytes were elongate, about 8 9 long by 3 ~ wide, with a large dense nueleus (Fig. 3). Older maerogametocytes were round or ovoid, f rom 5-10 ~xm in diameter and contained numerous dense stainirtg granules (Fig. 4). Dense granules congregated around the periphery of mature maerogametes. Refraetile vacuoles were also seen in the parasite eyto- plasm at this stage (Fig. 5). Maerogametoeytes were usually observed within parasi tophorous vaeuoles either beneath or above the host nucleus.
Oocysts. Ooeysts were observed in the intestinal epithelium in all stages of sporu- lation. The major i ty of ooeysts reeovered in the feees were sporulated. Ooeysts were subspherical or ovoid. The oocyst wall was eomposed of a single eolorless membrane t ight ly applied to the two sporoeysts so tha t the oocyst sometimes had an "hour -g lass" shape. This membrane ruptured easily and m a n y free sporocysts were observed in feces. Twenty live ooeysts measured 14-17 ~x in length by 10-12 ~z in width (mean 15.4 by 11,2 ~z). Lerlgth-width ratios 1.3 to 1.5 (mean 1.38). No oocyst residuum or polar grarmle. Sporoeysts were ellipsoidM, no Stieda body.
Twenty five sporoeysts measured 8-11 ~z in length by 7-10 ~x in width (mean 9.1 by 7.7 ~x). Length-width ratios 1.1 to 1.3 (mean 1.24). The sporocyst residuum
Life Cycle of Sarcocystis orientalis sp. n. 171
Figs. 1--5. Impression smears from ileum of python. Giemsa. 1 Microgametocyte with numerous nuclei. 2 Mierogametes. Note elongate flagellae. 8 Elongate maerogametoeyte. 4 Macrogamete in host eell. 50 lde r maerogamete. Note dense granules and reIractile vacuoles.
All Figs. × 1500
Fig. 6. Two sporulated ooeysts and a sporocyst. Note the elongate sloorozoite and granular residuum within the slooroeyst. Unstained, interferenee-eontrast photograloh. × 1500
Fig. 7. Zoite in blood film from rat. Giemsa. × 1500
Figs. 8--10. Impression smears from lung of rat. Giemsa. 8 Zoite (arrow) in white blood cell. 9 Schizont with 8 nuelei. 10 Cluster of small merozoites. All Figs. × 1500
172 V. Zaman and F. C. Colley
Fig. 11. :Numerous cysts (arrows) in muscle surrounding the esophagus of a rat × 250
Fig. 12. Cross section of cysts in muscle. PAS. × 1500
Fig. 13. Longitudinal section of cyst in muscle. Note striated cyst wall (arrow). Hematoxylin eosin. × 1500
Life Cycle of Sarcocystis orientalis sp. n. 173
consisted of compaet coarse granules, orten concentrated at one side of the sporocyst. Sporozoites were sansage shaped, about 8 ~ long (Fig. 6).
2. Development in Rat
Tweaty-eight of 30 rats died 5-12 days following infection with oocysts. All of these showed achte respiratory distress in the form of rapid and shallow breathing 1-2 days before death. Six of the 28 rats also developed partiM paralysis of their hi•d limbs. On autopsy all of the rats had massive pleural effusion; however, no parasites were found in the pleural fluid.
Zoites. Zoites appeared in the blood 4-9 days following infeetion. These zoites were banana shaped and had a large nueleus» approximately 2.5 ~ in diameter. Twenty-five zoites measured 6.8-9.1 by 2.0-3.3 iz (mean 7.85 by 2.48 ~t) (Fig. 7).
Most zoites were free in the plasma but were also occasionally observed within white blood eells (polymorphs) (Fig. 8). Zoites wirb two nuclei were sometimes seen ia white blood cells. Zoites were observed in impression smears of all organs examined, with the exeeption of the brain.
Schizonts. Small sehizonts 5-6 ~ in diameter with a maximum of 8 nuclei were observed in the lungs (Fig. 9). Small merozoites measuring approximately 2-3 ~ long developed within these sehizonts (Fig. 10). Sehizonts were observed only in lung tissue.
Tissue cysts. The two rats that survived infection had reeeived on]y a smM1 dose of ooeyst suspension (0.1 tal) and did not show any visible illness. Two months after infection muscles of the thorax, diaphragm, tongue, esophagns and heart were examined. Heavy infection with typical Sarcocystis eysts was observed in striated musele only (Fig. 11). Cysts were e]ongate. The longest cyst observed was 4.4 mm in length. Average cyst diameter was abont 2.0 mm (Fig. 12). The cyst wall averaged 4.5 ~ in width and appeared striated with hematoxylin-eosin stMn (Fig. 13). I~o trabeeulae were observed within the eysts. Twenty formol- saline fixed eystozoites were 4.5-6.0 ~ long and 1.0-1.5 ~ wide (mean 5.53 by 1.38 ~). Host tissue surrounding cysts appeared normal.
Electron Microscope Study
1. Devdopment in Python
Microgametocytes. ¥oung microgametocytes were within parasitophorous vacuoles in the host cell. ~olysaccharide granules were distributed peripherally, beneath the pel]iele of the microg~metocyte. Mitochondria and small vesictes were scattered throughout the cytoplasm (Figs. 14, 15). ~uclei had peripherM eoneentratiort of chromatin. Some rmclei were dividing. Nuelei near the pelliele were orten assoeiated with elongate mitoehondria. A basal body was sometimes observed between the nueleus and the pellicle (Fig. 14). No mieropores were observed.
During maturation of the microgametocyte the nuclei protl~uded into the parasitophorous vacuole. There was thickening of the pellicle on euch side of the nueleus. Flagellae with the typieal 9 + 2 configuration of tubules were seen within the parasitophorous vacuole. Po]ysaceharide granules and large vacuoles were scattered throughout the eytoplasm (Fig. 15).
Fig. 14. Mierogametocyte in intestin~l epithelium of python. Polysaccharide granules are distributed peripherally within the parasite. Two nuclei are dividing. A basel body and
several elongate mitochondria may be seen adjacent to nuclei. ×9860
Fig. 15. OIder microgametocyte showing 4 nuclei protruding into the parasitophorous vacuole. Note thickening of pellicle adjacent to nuclei aad flagellae in the parasitophorous vacuole.
× 16000
Life Cycle of Sarcocystis orientalis sp. n. 175
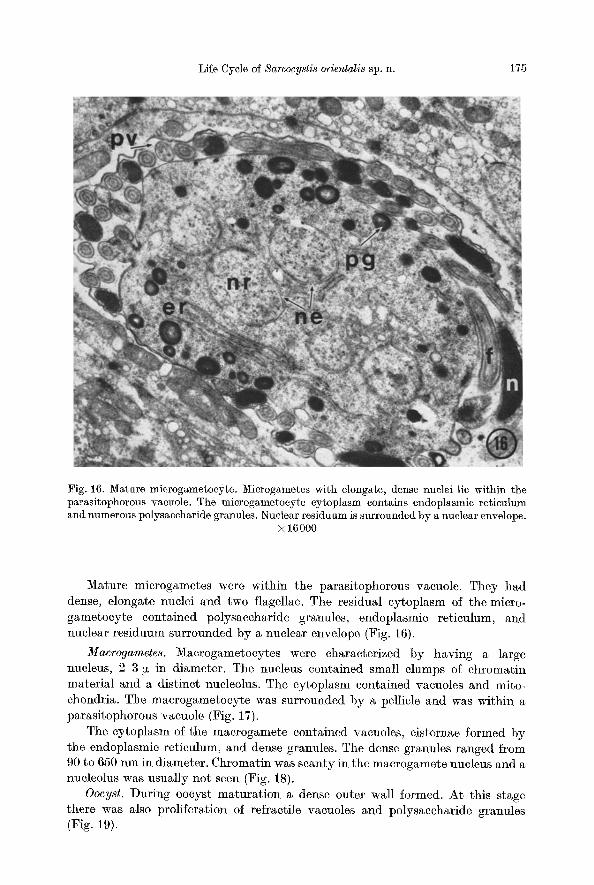
Fig. 16. Mature microgametocyte. Microgametes with elongate, dense nuclei lie within the parasitophorous vacuole. The microgametocyte cytoplasm contains endoplasmic reticulum und numerous polysaccharide granules. Nuclear residuum is surrounded by a nuclear envelope.
× 16000
Mature microgametes were within the parasitophorous vacuole. They had dense, elongate nuclei and two flagellae. The residual eytoplasm of the micro- gametocyte eontained po]ysaccharide granules, endoplasmic reticulum, and nuclear residuum surrounded by a nuclear envelope (Fig. 16).
Macrogametes. Macrogametocytes were charaeterized by having a large nueleus, 2-3 ~ in diameter. The nucleus contained small clumps of chromatin material and a distinet nueleolus. The cytoplasm eontained vacuoles and mito- chondria. The macrogametocyte was surrounded by a pellicle and was within a par~sitophorous vaeuole (Fig. 17).
The cytoplasm of the macrogamete eontained vaeuoles, cisternae formed by the endoplasmie reticulum, and dense granules. The dense grann]es ranged from 90 to 650 nm in diameter. Chromatin was scanty in the maerogamete nucleus and a nueleolus was usually not seea (Fig. 18).
Oocyst. During ooeyst maturat ion a dense outer wa]l formed. At this stage there was also proliferation of refraetile vaeuoles and polysaceharide granules (Fig. 19).
176 V. Zaman and F. C. Colley
Fig. 17. Marcogametocyte lying in parasitophorous vacuole. Note large nucleus with nucleolus. X 20000
}'ig. 18. Young macrogamete. Cytoplasm contains dense granules, cisternae and vacuoles. x 10080
Life Cycle of Sarcocystis orientalis sp. n. 177
The oocyst wall was apparently formed by the breakdown of the dense granules of the macrogamete into smaller partieles which fused to form a compaet dense layer (Fig. 20).
The ooeyst wall eonsisted of five concentrie layers. The outer layer was formed by the dense granules. Four underlying membranes formed the inner portion of the eyst wall. The remnants of the macrogamete pellicle were seen above the outer layer. Beneath the inner membranes was a clear space, 5-6 nm. Beneath this space two inner membranes formed the pellic]e surrounding the cytoplasmic mass (Fig. 21). The mature eyst wall was compact, approximately 0.8 ~ thiek, and eompletely enelosed the eytoplasmie mass of the ooeyst (Fig. 22).
2. Development in Rat
Tissue cysts. Only tissue cysts were observed with the eleetron microscope. The eyst was surrounded by a complex eyst wall. The cyst vacuole contained many elongate cystozoites. Metroeytes were observed around the periphery of the eyst vaeuole (Fig. 23).
Each eystozoite was eovered by a pellicle and contained a nuc]eus approx- imately 1.3 [z in diameter. The nucleus eontained clumps of dense chromatin. Some cells contained 2 nuelei. Micronemes and rhoptries were loeated anterior to the nueleus. A eonoid approximately 20 nm in diameter was seen at the anterior end of many cystozoites (Fig. 24). Micropores were oceasionally observed in the pellicle of eystozoites.
Eaeh metrocyte had a ]arge nueleus approximately 2.5 ~ in diameter. The nucleus eontained dense clumps of chromatin and also aggregates of dense granules which may represent chromosome material. The eytoplasm of the metroeyte contained numerous vesicles and mitoehondria. Invaginations were oceasionally seen in the metroeyte pelliele (Fig. 26).
Cyst wall. The outer layer of the cyst wall was composed of nnmerous villi about 4 ~ long and 1 tz wide. The vi]li were elosely paeked, separated by an inter- villus space about 0.4 ~ wide. Each villus was surrounded by a dense wall. Numerous mieropores, approximate]y 60 nm in diameter were seen in the walls of the villi (Figs. 25, 26). The end of the villus was rounded where it contacted the surrounding muscle cells. A concentration of dense particles was usually seen within the villus at this point. The basa] end of the villus narrowed to a diameter of about 0.3 ~z. The walls of the villus were continuous with the inner layer of the cyst wall (Fig. 25). Numerous mierotubules approximately 80 nm in diameter and 150 nm long projected from the base of the villi into a reservoir-like spaee above the irreer layer of the cyst wall. This reservoir contained numerous granules, vesicles and host eell mitoehondria. Direetly beneath the inner layer was a moder- ately dense granular band.
Diseussion
The coccidian nature of the life cycle of Sarcocystis was first revealed whea Fayer (1970, 1972) observed transformation of eystozoites from musele cysts of graekles into gametocytes and oocysts in cell eultures.
Fig. 19. Young oocyst containing numerous vacuoles and polysaceharide granules. Note developing oocyst wall. X 8400
Fig. 20. Seetion oB developing eyst wall. Dense partieles (arrows) fuse to form a dense layer. × 25200
Fig. 21. Section of ooeyst w~ll showing layers. A remnant of the macrog~mete pe]licle (p) lies above the dense layer (dl). Four membranes (arrows) form the immer portion of the oocyst wall. Two additional membranes (m) form the pellicle of the cytoplasmic mass. × 100000
Fig. 22. Section of an oocyst showing the final appearance of the wall. × 36000
Liße Cycle oB Sarcocystis orientalis sp. n. 179
Fig. 23. Tissue cyst. The cyst is surrounded by a thiek wall. The cyst vaculoe contains numer- ous cystozoites. Metroeytes are seen at the periphery of the vaeuole. × 8900
Fig. 24. Cystozoites. Note rhoptries, micronemes and conoid. One cell (lower right) contains two nuclei. × 16000
180 V. Zaman and F. C. Colley
Fig. 25. Sec~ion of cyst wall. Closely packed villi are separated by an intervillus space which opens into a subvillus reservoir. Dense granular material is coneentrated within the distal end of some villi. The basal end of eaeh villus is continuous with the inner layer oB the cyst
wall. A granular band lies beneath the inner layer. X 12000
Life Cyele of Sarcocystis orientalis sp. n. 181
Sexual stages of Sarcocystis were subsequently deseribed experimentally in the intestinal tissues of dogs and cats (Heydorn and Rommel, 1972a, b) and in tissue culture (Fayër, 1970, 1972 ; Vetterling et al., 1973).
The life eycle of Sarcocystis is now known to oeeur in two hosts. The sexual eyele normally takes place in a predator and the asexual eycle in its prey. Sporo- eysts or ooeysts are shed in the feces of the predator and are ingested by the prey. Sehizonts multiply asexually in the prey and zoites develop into musele eysts.
When musele eysts are eaten by the predator they grow into sexual stages in the intestine. Oocysts sporulate in the intestine and are passed out in the feces (Markus et al., 1974). The life eycle of S. orientalis follows the above pattern. The predator in the retieulated python and the prey the rat.
In cats infected with S. ]usi[ormis macrogametocytes and oocysts were found in a subepithelial location (Heydorn and t~ommel, 1972b). However, in the reticulated python gametocytes and oocysts of S. orientalis were observed ex- clusively in intestinal epithelial cells.
Fine structure of microgametogenesis of S. orientalis is similar to tha t reported for Eimeria intestinalis, E. per]orans, E. tenella and Toxoplasma gondii (Scholty- seck et al., 1972). Microgametes are small and resemble those of Sarcocystis sp. (Vetterling et al., 1973).
Two kinds of wall forming bodies (WB I and I I ) have been described from various eimerians (Scholtyseck et al., 1969, 1971) and T. gondii (Colley and Zaman, 1970; Pelster and Piekarski, 1972). The dense granules observed in oocysts oB S. orientalis and in Sarcocystis sp. (Vetter]ing et al., 1973) resemble wall forming bodies I. The granules in macrogametes of S. orientalis differ slightly from those of Sarcocystis sp. (Vetterling et al., 1973) in being denser and smaller. In both of these species the granules appear to break into smaller pieces and coalesce to form the oocyst wall. The ooeyst wall of S. orientalis is similar to that of Eimeria spp. (Scholtyseck, 1969, 1971) and Sarcocystis sp. (Vetterling et al., 1973) in being composed of 5 layers.
The oocyst of S. orientalis closely resembles those of S. tenella artd S. ]usi/ormis in dogs and cats (Markus et al., 1974). This ooeyst is also similar to those of several lsospora species reported from snakes (Pellérdy, 1965, 1969). Further studies may show tha t many Isospora-like inIections actually represent the predator phase of a Sarcocystis life cycle.
When sporocysts of S./usi]ormis, reeovered from dogs, were fed to calves asexual multiplication was observed after 20 days (Fayer and Johnson, 1973). Markus et al. (1974) observed two types of schizonts in sections oB ealf kidney, one having large and one having small merozoites.
Two types of zoites were also seen in S. orientalis. The small zoites were seen in the impression smears o~ the lung and the large zoites in the peripheral blood and the white cells.
Fig. 26. Numerous microtubules extend from the basal portion of the villus into the subvillar reservoir. Numerous micropores may be seen on the surface oB the villi. A metrocyte lies ag~inst the granular band. The metroeyte eontains a large nueleus with dense chromatin clumps and aggregates of granules (asterisk). An invagination of the pellicle may be seen.
× 16000
182 V. Zaman and F. C. Colley
Species diagnosis of Sarcocystis orientalis n. sp.
Predator Prey
Type host
Geographie area
Sexual development
Asexual development
Python reticulatus (Malaysian reticulated python)
Lowland forests, Malaysia and Singapore
Gametocytes and oocysts in epithelial cells of small intestine. Sporoeysts average 9.1 ~zm long by 7.7 tzm wide with sporocyst residuum. Oocyst covered by thin membrane
Rattus norvegicus, and possibly other rodents
s a m e
Large zoites (average 7.85 ~m long by 2.48 fzm wide) free in peripheral blood. Dividing forms in white blood cells. Small schizonts 5-6 ~m in diameter in lungs produce merozoites about 2-3 ~zm long
Cysts in skeletal muscle only, up to 4.4 mm long, 2.0 mm wide. Thick (4.2 ~m) striated wall. No trabeculae seen. Cystozoites banana shaped. (Average 5.53 ~m long by 1.38 ~m wide) in seetions
Syntypes and paratypes. Deposited with the Department of Parasitology, Faculty of Medicine, University of Singapore, and the ZooIogy Institute of the University of Bonn, West Germany.
Markus et al. (1974) have suggested that asexual multiplication occurs prima- rily in the reticulo-endothelial tissue of the host.
Taxonomy of Sarcocystis is usually based on the host (prey) in whieh the parasite is found and the morphology of the cyst wall and the cystozoites (Man- dour, 1965; Shaw and Lainson, 1969). Since the predator host is unknown for many species of Sarcocystis the characteristics of tissue eysts in the prey host are still important in distinguishing species.
Three Sarcocystis species have been described from Brazilian rodents by Shaw and Lainson (1969). The muscle cysts of S. orientalis differ from these as follows. From S. proechimyos in the spiny rat (Proechimyos guyannensis) in having roun4ed rather than spine-like villi and in lackiag trabeculae; from S. oryzomyos in the rice rat (Oryzomyos capito) in lacking invaginations in the cyst wall and in lackiug trabeeulae; from S. azevedoi, also in the riee rat, in having a mueh thicker cyst wall and in lacking invaginations in the wall. In addition, muscle cysts of S. orientalis differ from those of S. muris in miee (Mus musculus) in having a thick striated eyst wall rather than a thin one with tooth-like projections.
In Ma]aysia Dissanaike (1975) has described S. booliati from the moonrat Echinosorex gymnuris (Erinaceidae). S. orientalis differs from S. bootiati in having a thick (4.5 ~) rather than a thin (0.5 ~) eyst wall. I t also differs from Sarco-
Life Cyele oB Sarcocystis orientalis sp. n. 183
cystis spp. f rom the slow loris (Zaman, 1970), Ind ian water buffalo (Zamart and Colley, 1972) and long-tailed macque mortkey (Prathap, 1973) in Malaysia in lacking trabeculae in the cyst.
Sarcocystis has been reported from the muscles of several species of rats in the Philippines (Carlos and Schaeffer, 1972) ar~d Indormsia (Browrt et al., 1974) bu t the morphology of the parasites was not described in detail. I t is probable tha t wild rats, other thart R. norvegicus, ar t hosts of the asexual phase of the life cyele of S. orientalis in Malaysia.
Fine structure of tissue cysts of Sarcocystis also reveals considerable morpho- logical variat ion between species. I towever, all species studied to date are similar in having micropores in the surface of the cyst wall (Ludvik, 1960; Zeve et al., 1966; SimpsorL artd Forrester, 1973; Mehlhorn and Scholtyseck, 1973). These micropores apparent ly function in absorbing nutr ient material f rom the host eell. I n S. orientalis the surface area of the wall is greatly increased by the presence of closely packed villi and by microtubules at the base of each villus. Similar villi have beeia described in S. miescheriana (Ludvik, 1960).
Metrocytes in the cyst of S. orientalis are similar to those described in S. tenella by Mehlhorn and Scholtyseck (1973). Cystozoites of S. orientalis closely resemble those of other Sarcocystis species (Ludvik, 1960; Zeve et al., 1966; Simpson, 1966; Simpson and Forrester, 1973; Mehlhorn and Scholtyseck, 1973) and also T. gondii, Besnoitia jellisoni, a~d Frenlcelia spp. (Scholtyseck et al., 1973).
Or~ the basis of out observatiorls on the life cycle of S. orientalis wc feel t ha t this parasite should be considered a separate species.
Abbreviations for Electron Micrographs b basal portion of villus rac metrocyte
in wall of tissue cyst mi mitoehondrion bb basal body mp mieropore c chromatin mt microtubules cs eis~ernae mu muscle cells cv cyst vacuole n nucleus cw eyst wall ne nuclear envelope cz cystozoite au nucleolus dc concentra~ed dense particles nr nuclear residuum dg dense granule ow oocyst wall dl dense layer p pellicle er endoplasmic reticulum pg polysaceharide graaule g granular layer pv parasitophorous vaeuole he host cell r reservoir in tissue cyst wall hn host nucleus ro rhoptry il irreer layer t thiek area in pelliele in invagination v vaeuole iv intervillus space vl villus m membranes of pellicle vs vesicle
References Blanchard, 1%. : Note sur les Sarcosporidies et sur un essai de classification de ces Sporozoaires.
Bull. Soc. Zool. t~r. 10, 244 (1885) Browa, I%. J., Carney, W. P., Peerten, P. F. D. ran, Sudomo, M.: Sarcocystis from rats in
Sulawesi, Indonesia. Southeast Asian J. Trop. Med. Pub. tIlth. 5, 451452 (1974)
184 V. Zaman and F. C. Colley
Carlos, E. R., Scharfer, B. T. : Sarcocystis in the Philippines: A histologic review of 202 cases in rats. Southeast Asian J. Trop. Med. Pub. Hlth. 8, 371-375 (1972)
Colley, F., Zaman, V. : Observations on the endogenous stages of Toxoplasma gondii in the cat ileum. II . Electron microscope study. Southeast Asian J. Trop. Med. Pub. Hlth. 1, 465-480 (1970)
Dissanaike, A. S., Poopalachelvam, M.: Sarcocystis booliati n. sp. and a parasite of un- determined taxonomic position, Octoplasma garnhami n. gen. n. sp., flora the moonrat, Echinosorex gymnurus. Southeast Asian J. Trop. Med. Pub. Hlth. (in press, 1975)
Fayer, R.: Sarcocystis: development in cultured avian and mammalian cells. Science 168, 1104-1105 (1970)
Fayer, R. : Gametogony of Sarcocystis sp. in cell culture. Science 17ä, 65-67 (1972) Fayer, R., Johnson, A. J. : Sarcocystis /usi]ormis: Development of cysts in calves infected
with sporocysts from dogs. Proc. helminth. Soc. Wash. 41(1), 105-108 (1974) Heydorn, A. O., Rommel, M. : Beiträ~ge zum Lebenszyklus der Sarkosporidien. II . Hund und
Katze als Übertr/~ger der Sarkosporidien des Rindes. Ibid. 85, 121-123 (19723) Heydorn, A. O., Rommel, M.: Beitr/~ge zum Lebenszyklus der Sarkosporidien. IV. Ent-
wicklungsstadien von S./usi/ormis in der Dünndarmschleimhaut der Katze. Ibid. 85, 333-336 (1972b)
Ludvik, J. : The electron-microscopy of Sarcocystis meischeriana Kuhn 1865. J. Protozool. 7, 128-135 (1960)
Mandour, A. M. : Sarcocystis garnhami n. sp., in the skeletal muscle of an opossum, Didelphis marsußialis. J. Protozool. 12, 606-609 (1965)
Markus, M. B., Killick-Kendrick, R., Garnham, P. C. C. : The coecidial natural and life-eycle of Sarcocystis. J. trop. Med. Hyg. 77, 248-259 (1974)
Mehlhorn, H., Scholtyseek, E. : Elektronenmikroskopische Untersuchungen an Cystenstadien von Sarcocystis tenella aus der Oesophagus-Muskulatur des Schafes. Z. Parasitenk. 41, 291-310 (1973)
Pellérdy, L. P. : Coeeidia and Coecidiosis. Akadémiai Kiadé, Budapest, 657 p. 1965 Pellérdy, L. P. : Catalogue. Eimeriidea (Protozoa, Sporozoa). Supplementum 1. Akademiai
Kiado, Budapest, Hungary, 80 p. 1969 Pelster, B., Piekarski, G. : Elektronenmikroskopische Analyse der Mikrogametenentwicklung
bei ToxopIasma gondii. Z. Parasitenk. 87, 267-277 (1971) Pelster, B., Piekarski, G.: Untersuchungen zur Feinstruktur des Makrogameten von Toxo-
Blasma gondii. Z. Parasitenk. 89, 225-232 (1972) Prathap, K. : Sarcocystis in the Malaysian long-tailed monkey, Macaca irus. Trans. roy. Soc.
trop. Med. Hyg. 67, 615 (1973) Bzepczyk, C. M. : Evidenee of a rat-snake life cycle for Sarcocystis. 1nr. J. Parasit. 4, 447-449
(1974) Scholtyseck, E., Mehlhorn, H., Hammond, D. M. : Electron mieroseope studies of mieIo-
gametogenesis in coceidia and related groups. Z. Parasitenk. 88, 95-131 (1972) Scholtyseck, E., Mehlhorn, H., Müller, E. G.B. : Identifikation von Merozoiten der vier
cystenbildenden Coceidien (Sarcocystis, Toxoplama, Besnoitia, Frenkelia) auf Grund feinstruktureller Kriterien. Z. Parasitenk. 42, 185-206 (1973)
Scholtyseek, E., Mehlhorn, I-I., Hammond, D. M., Müller, E. G. B., Rammel, A., Heller, G. : Licht- und elektronenmikroskopische Untersuchungen zur Bildung der Oocystenhülle bei Eimerien (Eimeria per]orans, E. stiedae und E. tenella). Z. Parasitenk. 81, 289-298 (1969)
Shaw, J. J., Lainson, R. : Sarcocystis of rodents and marsupials in Brazil. Parasitology 59, 233-244 (1969)
Simpson, C. F. : Electron microscopy oB Sarcocystis/usi/ormis. J. Parasit. 52, 607-613 (1966) Simpson, C. F., Forrester, D. J. : Eleetron microscopy of Sarcocystis sp. : Cyst wall, micropore,
rhoptries, and an unidentified body. Int. J. Parasit. 3, 467-470 (1973) Vetterling, J .M. , Pacheco, N.D. , Fayer, R.: Fine structure of gametogony and oocyst
formation in Sarcocystis sp. in cell culture. J. Protozool. 20, 613-621 (1973) Zaman, V. : Sarcocystis sp. in the slow loris, 2Vycticebus coucang. Trans. roy. Soc. trop. Med.
Hyg. 64, 195-196 (1970)
Life Cycle of Sarcocystis orientalis sp. n. 185
Zaman, V., Colley, F. : Fine structure of Sarcocystis ]usi/ormis from the Indian water buffalo (Bubalas bubalas) in Singapore. Southeast Asian J. Trop. Med. Pub. Hlth. 8, 489-495 (1972)
Zeve, V. H., Price, D. L., Herman, C. M[. : Electron microscope study of Sarcocysgs sp. Exp. Parasit. 18, 338-346 (1966)
V. Zaman Dept. of Parasitology Faculty of Medicine University of Singapore Sepoy Lines Singapore 3